ri. fettp diet of powdered milk, sugar and dried meat powder was sufficient for maintenance of adult...
TRANSCRIPT

LABORATORY REARING, TOXICITY OF CYROMAZINE AND THE
EFFECT OF TEMPERATURE AND MANURE MOISTURE ON OPHYRA
AENESCENS (WIEDEMANN) (DIPTERA: MUSCIDAE)
by
Lorraine Marie Koller
Thesis submitted to the Faculty of the Virginia Polytechnic Institute and State
University in partial fulfillment of the requirements for the degree of
APPROVED:
L. T. Kok
MASTER OF SCIENCE
Ill
Entomology
E. C. Turner, Jr., Chairman
May, 1989
Blacksburg, Virginia
I . fettp ri. a. Pfei~r

LABORATORY REARING, TOXICITY OF CYROMAZINE AND THE
EFFECT OF TEMPERATURE AND MANURE MOISTURE ON OPHYRA
AENESCENS (WIEDEMANN) (DIPTERA: MUSCIDAE)
by,
Lorraine Marie Koller
E. Craig Turner, Jr., Chairman
Entomology
(ABSTRACT)
The effect of protein in adult and larval diets for Ophyra aenescens
(Wiedemann) (Diptera: Muscidae ), a facultative predator of the house fly, Musca
domestica L., was examined. A larval diet of coarse wheat bran, vermiculite and
dried meat powder was found adequate for rearing 0. aenescens larvae. Protein
content was important for maximum larval emergence and for Fl adult longevity
and fecundity. Adult diet of powdered milk, sugar and dried meat powder was
sufficient for maintenance of adult 0. aenescens flies. Protein was needed in adult
diet for optimal fecundity.
Cyromazine was toxic to 0. aenescens at high levels (1.0 ppm), but at 0.75
ppm 0. aenescens tolerated cyromazine better than a susceptible strain of house
flies. At these cyromazine levels, mass release of 0. aenescens into poultry houses
is possible one to two days after cyromazine has been removed from the chicken
feed.
The effect of temperature and manure moisture on 0. aenescens was
studied. At temperatures of 18°C, emergence of Ophyra aenescens was
significantly lower than at temperatures of 21 and 27°C. Predation by 0.

aenescens at 27°C on house fly larvae was significant at ratios of 3:1, 2:1and1:1
(house fly to 0. aenescens). A constant manure moisture was important in the
development of 0. aenescens larvae and its ability to prey on house fly larvae. At
constant levels of 50, 60, and 70% manure n;ioisture, 0. aenescens substantially
reduced house fly larval numbers at ratios of 3:1, 2:1and1:1.

ACKNOWLEDGEMENTS
I wish to thank Dr. E. C. Turner, Dr.,L. T. Kok, and Dr. D. G. Pfeiffer for
being members on my committee.
I am grateful to and for their technical
assistance, logical thinking and advice; Dr. Kok for use of an environmental
chamber; for building the fly cages used in the experiments;
of the Poultry Science Department for his never ending supply of chicken
manure; Dr. Ruszler of the Poultry Science Department for use of his chickens in
the cyromazine experiment; Dr. Ray Noblet of Clemson University for analyzing
the different larval media types; and Dr. Birch, Dr. Krutchkoff and Dr. Pirie of the
Statistics· Department for their advice.
Financial support was made possible by the Virginia Agriculture Council,
grant number 4-37043 and by Dr. Turner.
I wish to thank the following people: and
who were a constant source for inspiration. Special thanks goes to
for his advice, encouragement, friendship and his enthusiasm for science.
Finally, I would like to thank my Mom and Dad for calling me every Sunday
night to listen to me and to cheer me onand for sending care packages when I
needed it.
iv

! '
TABLE OF CONTENTS
ABS'ffi.ACT : ........... ~ .................. · ...... ~ ......................................................................... ii
ACKNOWLEDGEMENTS ...............................................•................................. iv
INTRODUCTION:· ..........•............• : .•.. : .••... ~ .. ~.~ ••..•.. ~ •............................................... 1
I. UTERATURE REVIEW ..•.•. ~ .... ·~ ..................................................................... 3
II. THE EFFECT OF DIFFERENT PROTEIN LEVELS IN LARY AL
AND ADULT DIETS ON THE DEVELOPMENT AND
REPRODUCTION·OF•OPHYRAAENESCENS (WIEDEMANN)
(DIPTERA: 'MUSCIDAE) ...........• ~: ..... ; ... ~~ ............... ;; ........................................ 13
ill. THE TOXICITY OF CYROMAZINE TO OPHYRA AENESCENS
(WIEDEMANN}(DIPTERA: MUSCIDAE} ..........•....................................... 28
IV. THE EFFECT OF TEMPERATURE AND MANURE MOISTURE
ON THE PREDATORY HABITS OF OPHYRA AENESCENS
(WIEDEMANN} ON MUSCA DOMESTICA L. ........................................... 37
LITERATURE CITED ...................................................................................... 5 0
VITA ............................................................. .' ............... ~ .......................................... 57
v

1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
LIST OF TABLES
Mean number of flies emerged and standard error of Ophyra aenescens reared on different larval diets placed into the rearing room.. . ............................................................... 23
Mean number and standard error of flies emerged from 150 first instar Ophyra aenescens seeded into larval diets with varying protein levels. . ................................................ 24
Fecundity and longevity with standard error for Fl male and female Ophyra aenescens reared from different protein levels. . ................................................................................... 25
Fecundity.and longevity with standard error for males and female Ophyra aenescens fed adult diets with varying protein levels. . .................................................................... 26
Percent mortality of Musca domestica and Ophyra aenescens from cyromazine in rearing media. . ............................................ .32
Slope, LC90 and fiducial limits from probit analysis or cyromazine in rearing media. . .................................................... .32
Percent mortality of Musca domestica and Ophyra aenescens from cyromazine in chicken manure. . ......................................... .33
Slope, LC and fiducial limits from probit analysis ofOcyromazine in chicken manure. . ................................................... 34
Percent emergence of Musca domestica and Ophyra aenescens from chicken manure one day after and one week after cyromazine was removed from food. . ................................. 35
Effect of manure moisture and competition with Musca domestica on emergence of Ophyra aenescens. . ............................... .43
Effect of manure .moisture and competition with Ophyra aenescens on emergence of Musca domestica. . ............................... .44
Effect of temperature and competition with Musca domestica on emergence of Ophyra aenescens. . ........................................... .45
Effect of temperature and competition with Ophyra aenescens at different ratios on the emergence of Musca domestica ............... : .......... ~ ..................................................................... 46
vi

LIST OF FIGURES
Figure Page
1. House fly emergence at 70, 60 and 50% manure moisture at different house fly to Ophyra aenescens ratios ..................................... : ............................................................ 47
2. House fly emergence at 27, 21and18°C at different house fly to Ophyra aenescens ratios. ····••••••n••·····································································································48
vii

INTRODUCTION
Poultry systems today are providing greater efficiency in animal production.
At the same time these systems also increase problems in manure disposal and fly
control. Manure pits in caged layer poultry houses provide an ideal environment
for the development of the house fly, M-usca domestica L. (Diptera: Muscidae ).
The house fly, primarily a nuisance pest, can be a severe problem when
populations reach high levels. This fly disturbs farm workers and frequently
disperses to nearby residential areas. Large numbers of house flies in residential
areas can prompt complaints from neighbors and may be a potential health threat
(Greenberg 1971).
Traditionally, pesticides are used in house fly control, but with insect
resistance becoming a problem in many poultry houses, an Integrated Pest
Management system (IPM) is becoming more popular. This system involves
cultural and biological control methods with minimal chemical input. Within this
system an alternative approach in biological control of the house fly is through the
use of a facultative predator, Ophyra aenescens (Wiedemann) (Diptera: Muscidae ).
The larvae of 0. aenescens are predators of house fly larvae and mass releases of
0. aenescens into poultry houses can reduce house fly populations (unpublished
data). Mass releasing an insect requires knowledge about rearing the insect. Little
information exists about rearing 0. aenescens for mass release.
After using the proper cultural and biological control practices there is still
often a need to use insecticides to control house flies. One of the best ways to
control flies is through the use of feed-through larvicides. This is an ideal method
to control house flies because it eliminates labor costs and uses no expensive spray
equipment. Cyromazine (Larvadex™) (Ciba-Geigy Corp., Greensboro, NC), a
1

2
feed-through insect growth regulator, is widely used for larval control in caged
layer poultry houses. Although cyromazine is known to affect only dipterous
insects, its toxicity to 0. aenescens is unknown.
Because of the predatory habits of th~ larva, 0. aenescens has been
proposed as a biological control agent for use in an IPM program (Nolan &
Kissam 1985). Thus, egg producers would benefit financially because 0. aenescens
would reduce the amount of chemicals required to· obtain satisfactory house fly
control. The implementation of an IPM program using 0. aenescens may also
retard the occurrence of insecticide resistance. .
The information from this research will be used to.integrate 0. aenescens
into an IPM program for house fly control. The specific objectives of this study are
to determine the:
1. effect of varying protein levels in both larval and adult diet on fly
emergence, fecundity and longevity.
2. level of toxicity of cyromazine to 0. aenescens larvae.
3. effects of different moistllre levels and temperature on 0. aenescens
ability to prey on house fly larvae.

LITERATURE REVIEW
Ophyra aenescens. Ophyra aenescens was first described by Wiedemann in
1830 (Johnson & Venard 1957) and placed~ the genusAnthomyia. According to
Johnson & Venard (1957), Stein in 1897 transferred it toDphyra Robineau-
Desvoidy. The genus Ophyra was first placed in the family Anthomyiidae but
Crampton (1944) after examiningthe terminalia statedthatOphyra was a typical
muscid and placed it in the family Muscidae where it remains today.
Where is 0; aenescens found?. According to Greenberg (1971), Ophyra
aenescens occurs in the United States from Oregon to Arizona, Illinois to the East
Coast. This fly is also neotropical and oceanic. Ophyra aenescens has spread to
Europe (Adams 1984, Skidmore 1985). Ophyra aenescens has been found in Texas
around animal.confinement areas (Robertson & Sanders 1979) and in poultry
houses in South Carolina (Nolan & Kissam 1985) and Florida (Hogsette 1979).
Mail & Schoof{1954) found 0. aenescens moving around actively in refuse
dumping areas in West Virginia. Hill & Williams (1980) found 0. aenescens to be
the most numerous fly in high rise poultry houses in Indiana.
In lab experiments performed by Johnson & Venard (1957), the eggs of 0.
aenescens hatched in 12 to 16.hours·at 28°C. The development period for the
three larval instars averaged 9 days while the pupal stage averaged about 4 days at
27°C. The adult is shiny black and slightly smaller than the house fly. The male
fly has contiguous eyes while the female's eyes are separated. The females hold
theirwings parallel to each other and the male~s wings are crossed half way
between the fifth and sixth veins (Johnson & Venard 1957). Adult males lived an
average of 15 days while females lived an average of 20 days (Johnson & Venard
3

4
1957). Hogsette (1979) reported similar results in that females lived for an
average of 20 days and outlived males by an average of 3 to 4 days. One pair of
adults can produce 300:-400 offspring (Johnson & Venard 1957).
Rearing. Little information on Ophy~a spp. rearing exists. Roddy (1955)
used a larval media consisting of whole milk, Brewer's yeast, bacto-agar and water
for rearing 0. aenescens. Johnson & Venard (1957) used the Chemical Specialties
Manufacturing Association ( CSMA) preparation medium, fish meal diamalt, cake
yeast, and brewer's yeast. Hogsette (1979) used a similar media, consisting of
CSMA, wheat flour, fish meal, N31C03, alfalfa meal and water. Olckers & Hulley
(1984) reared colonies of Ophyra capensis (Wiedemann) on lucerne meal, wheat
bran, molasses, yeast, powdered milk and a fungus. suppressor. Nolan (1987) used
a diet consisting of CSMA medium, soybean meal, two whole chicken eggs and
water.
Johnson & Venard (1957) used fish meal for adult food of 0. aenescens.
Hogsette (1979) gave adult 0. aenescens .flies cane sugar, dry fish meal and
nasogastric mix. Nolan (1987) used a mixture of powdered milk, table sugar and
egg for the adult food. Olckers & Hulley (1984) used sugar and whole milk
powder for adult food of 0. capensis.
In basic biological studies on 0. aenescens by Hogsette (1979), a fortified
larval diet (CSMA medium plus alfalfa meal, N31C03, fish meal and wheat flour)
did not shorten larval development time as compared·with an unfortified diet
(CSMA media). The fortified diet did increase the number of pupae that formed
and the number of adults thatemerged. Size of emerging adults was the same for
both fortified and unfortified diets. Sugar in adult food increased the adult life
span 9 times. The life span for flies without sugar in the adult food was 2 days

5
while the life·span was 18 days for flies feeding on sugar.
Predaceous behavior. The larvae of Ophyra spp. are predaceous on other
dipterous larvae (Anderson & Poorbaugh 1964a, Peck 1969, Peck & Anderson
1969) but not cannibalistic (Anderson & Poqrbaugh 1964b, Hogsette 1979).
Hogsette (1979) examined the cephaloskelton of the third-instar larvae of 0.
aenescerzs and concluded that the larvae can live either in a saphrophagous manner
or they can be carnivorous. Similar findings were reported by Schumann(1982).
Anderson & Poorbaugh (1964a, 1964b) reportedthat Ophyra leucostoma
(Wiedemann) larvae may kill 2 to 20 Musca domestica L. maggots per day. In
laboratory studies by Olckers & Hutley (1984), 0. caperzsis consumed 4-5 house fly
larvae per day. Nolan & Kissam (1985) observed 0. aenescerzs feeding onM.
domestica, Phormia regina (Meigen) and Sarcophaga sp. larvae in laboratory
colonies. Hogsette (1979) found thatO. aenescerzs could destroy more than 20
first-instar house fly larvae per day during larval development. Muller (1982)
discovered that joint breeding of 0. aenescerzs and M. domestica led to a high
mortality rate of M. domestica. 0. aenescerzs was a facultative predator and
developed normally without consuming fly larvae.
Because of the predaceous habit of the larvae of the genus Ophyra, it has
been suggested. they could be used as biological control agents against the house
fly (Derbeneva-Ukhova 1943, Conway 1973, Muller et al. 1981, Olckers & Hulley
1984, Nolan & Kissam 1985, Ribbeck et al. 1987). Many researchers feel the
adults of 0. aenescerzs are pestiferous and excessive numbers of this fly are just as
annoying as the house fly (Hogsette 1979, Axtell 1986a, Axtell & Rutz 1986). The
nuisance potential of adult Ophyra flies was examined by Nolan & Kissam (1987).
They found that dispersal from poultry houses with high populations of 0.

l
.6
aenescens was minimal. This agrees with other workers (Anderson & Poorbaugh
1964a, 1964b, Legner & Dietrick 1974)in thatOphyra spp: adults tend to hide in
secluded places.to mate -and.congregate.
House Fly. Linnaeus described the h<;mse fly .in 1758 (West 1951). It is an
ubiquitous insect (West 1951 ), is cosmopolitan in· distribution and meets all
conditions requfred of a disease vector (Greenberg 1971). Jt is important because
it coexists with m~ consuming·both contaminated~an:d;non~cdntaminated·food,
has great flight activity .and dispersal and alternates between feces and Jood
(Greenberg 1971); ·
The development of this fly is highly.dependent upon temperature which
influences both survival and time required for development(Harwood & James·
1979). The egg stage takes·8~12 hours'to hatch, larval.stage takes 5 days, pupal
stage.takes 4-5.days, with a total of 10 days from egg tc>' adultat3o0c. Under
temperate·conditio:risit woUldtake3weeks•(Harwood.&JamesJ979) .. During the
srinnller in the southeastern United States; generation times are between ·six and·
ten days. (Axtell 1986b )>The average life span• of the adult can range-from· 14· to 70
days (Harwood&James1979); House flies can overwinterin•any.stage
(Greenberg1971). ·
The house fly is vecy prolific (Harwood & James 1979); A· female mates
and beginslaying .. eggs 3-4 days afteremerge11ce· (Axtelt1986a). Some housefly
females can lay over 1000 eggs (LaBrecque: etaL 1972) ·in• the laboratory but
usually females will .lay 4-6 batches of 100-150 eggs in her lifetime• (Axtell l986a ).
Greenberg {1973)reported a dispersion. of 2.3 km to lL8 km per day. They
mostly stay within lto 3 km-of: their breeding place butcan: fly 5 or 6 km in large

7
numbers (Harwood & James 1979). Mark recapture traps have indicated that the
house fly can go as far as 20-32 km from their source (Linquist et al. 1951) using
wind drift to increase the distance (Oldroyd 1965).
Resting spots of house flies were also, investigated. Traps held overnight
caught 85% ofhouse flies.within houses (Anderson & Poorbaugh 1964b).
Larvae of house flies can be found in refuse areas (Mail & Schoof 1954)
and inhorse, cow and chicken manure (Harwood & James.1979). In chicken
manure, a temperature of 27°C and a moisture of 60-75 % was determined to be
optimum for house fly larval development (Miller et al. 1974).
Fly Control. Manure management and fly control are closely related
problems for poultry facilities (Barth 1986). The moisture content of fresh manure
that accumulates beneath cages ranges from 70-80% (Hart 1963, Card & Nesheim
1975) and has a pH of 6butincreases due to bacterial growth after a few hours
(Beard & Sands 1973). This is optimal for house fly larval development(Miller et
al. 1974). Keeping the manure dry (below 60%) reduces.the habitat for fly larval
development (Axtell 1986a, 1986b ). Maintaining watering systems and preventing
moisture from entering into the fly breeding areas is critical in fly control (Axtell
1986b, Barth 1986). If this cannot be done, increasing the moisture level higher
than 80% where manure becomes anaerobic will render it unsuitable for house fly
development (Miller et al. 1974) but this may create conditions that favor other
noxious flies (Axtell 1986b ). Keeping the manure dry will also encourage natural
fly predator and parasite populations to develop (Axtell 1986b ). The activities of
predators and parasites in accumulated manure in Florida resulted in a 97%
reduction in the number of house flies recovered from artificially infested

8
containers (Propp & Morgan 1985).
Three families of mites are known to prey upon house fly eggs or larvae:
Macrochelidae, Uropodidae and Parasitidae (Axtell 1963, .1986b ). Macrocheles
muscaedomesticae (Scopoli), a phoretic mite,, can kill house fly eggs and first instar
larvae by piercing the chorion or integument and sucking out the contents (Axtell
1963, 1964 ) .. Other important predatory mites in poultry manure are the uropodid
mite Fuscruopoda vegetans (DeGeer), and. the parasitid mites, Poecilochirus spp.
(Geden et al. 1988). Many chemicals used to control house flies are harmful to
mites (Axtell 1966). Larvicides were reported to destroy mites but adulticides left
them unharmed (Axtell 1970) ..
Many beetles especially Histeridae and Staphylinidae are common in
poultry manure (Peck1969, Pfeiffer & Axtell 1980). Carcinops pumilio (Erichson)
and Alphitobius diaperinus (Panzer) were consistently the most abundant species in
three regions of North Carolina (Pfeiffer & Axtell 1980). Carcinops pumilio adults
are known predators of house fly eggs (Peck1969, Geden et al. 1988) while the
larvae wiHfeed on house fly instars (Geden et al. 1988). The role of A. diaperinus
as a predator of house fly larvae was examined by Despins et al. (1988). Both
adults ~d late instar A. diaperinus larvae had a significant impact on house fly
emergence in the laboratory. Not enough research has been conducted to reveal
the importance of other species of beetles found in poultry manure (Pfeiffer &
Axtell 1980).
The major parasites of house flies in confined animal systems are small
wasps (Hymenoptera) which belong to the family Pteromalidae. These parasites
are common biological control. agents of flies in poultry houses (Legner & Brydon
1966, Morgan & Patterson 1975, Rutz & Axtell 1980, Rueda & Axtell 1985). Mass

I
releases of these parasites -can result in a decrease 'in .house fly populations (Ables
& Shepard 1974,,0lton&Legner 1975, Rutz &.Axtell 1981, Shepard & Kissam
1981) although some.species of parasites were not active.throughout the year
(Olton & Legner 1975, Rutz & Axtell 19~0) and~perfotmed better in dry manure
than wet manure {Rutz & Axtell 1980). Parasites were also required to be
climatically adapted to the ateawhere they are released (Axtell & Rutz 1986).
Manure removal schedules. can affect abundance of house fly and predator
populations. Monthly or biweekly removal of manure favored fly populations
while the. predators studied were most abundant in ;Unremoved manure (Peck &
Anderson 1970)~ Lower numbers .offlies emerging from: poultry manure was
correlated with increasing masses of breeding habitat due to increased populations
of scavengers, predators and parasites,(Legner et al~ 1973). If manure must be
removed, ·leaving a base of old,manure .. behind in the pit would favor beneficial
arthropod predators and also absorb excess.moisture from new droppings (Legner
& Brydon.1966, Peck &Anderson1970, Axtell 1986a).
Environmental conditions in manure pits were measured to see what role
theymight playonthe developmentof flies and beetles:(Stafford & Collison 1987).
Temperatures in the pits had weekly means of 18.l.to 20.o0 c which is adequate for
fly development all year round. ·Mean temperatures in upper manure layers were
near optimum (29 .5~35.8°C) for beetle development. Relative humidity was found
to be around.65% .and no seasonal trends were found. In British poultry houses,
moisture content of manurewas in.dependent of temperature and relative humidity
(Bills 1973)~
· Chemical Larvicides •. Larvicides are an effective way to control livestock
pests,. either applied directly to manure or as a feed additive• (Miller 1970).

10
Larvicides should only be used after biological and cultural controLtactics have
been applied and a high level of fly control is desired (Axtell 1986b ). Insecticides
are effective if they complement biological and cultural controls (Axtell 1986a).
A chemical should not be 'routinely used if it is known to be toxic to
parasites and predators (Axtell 1986a). When spraying manure directly, the
chemical should only be applied· to those areas in which abundance of flyJarvae is
observed (Axtell 1986b). Bailey et al. (1968) stated that sprayinglarge quantities
of manure is ineffective due to larval movement mixing the chemical with the
mediumand quickly making the concentrationtoolmy. Major insecticides
currently used as larvicides are: stirophos (Rabon™), dimethoate (Cygon ™) and
cyromazine (Larvadex™) (Axtell 1986b)~.
Pathogens. The fungus Entomophthora muscae (Cohn) Fresenius is a
widely known natural enemy of adult muscoid flies. ·This pathogen can be
transmitted within house fly populations during periods of comparatively dry and .
cool weather and when the relative humidity within the boundary layer
surrounding the fly's body is at or near the saturation point (Kramer 1980).
Kramer & Steinkraus (1987) achieved 100% mortality of house flies in a small
poultry house by placing infected flies into the house.
Hall & Arakawa (1959)successfullyusedB. thuringiensis vaL thuringiensis.
Berliner to kill house fly maggots in laboratory tests~ The pathogen Bacillus
thuringiensis Berliner was fed to caged layers for fly control buta reduction in feed
consumption, body weight and egg production was noted (Burns et al. 1961).
Insect Growth Regulators. Insect growth regulators (IGR) have been
tested for control of the housefly. ·Several chemicals.have.been effective
experimentally for feed-through fly control buthave not been registered for use

11
due to residues in animals or products (Axtell 1986b )~ Laboratory bioassays using
encapsulated methoprene, anIGR, resulted in good control of house fly
emergence at a rate of 5.0 ppm after the chickens were fed treated feed for eight
days (Breeden et al. 1975). Similar results were found by Morgan et al. (1975) and
birds did not experience a weight loss. However, methoprene was not effective as
atopical spray on poultry droppings under field conditions (Morgan et al. 1975)
and in field studies, satisfactory fly control was not achieved· due to existing cross
resistance to methoprene (Breeden et al. 1980).
Diflubenzuron (Dimilin ™) has also been tested as a feed'-through larvicide
for control of the face fly and the house fly (Miller 1974). In field and laboratory
studies, Diflubenzuron affected intermediate to late stage larvae but had no effect
on house fly eggs or fly pupae (Ables et al. 1975). Diflubenzuron also had no
effect on the parasitoid Muscidifurax raptor Girault and Sanders or its ability to
parasitize house fly pupae (Ables et al. 1975). When Diflubenzuron was applied to
fly resting surfaces in conjunction with releases of the parasitoid, M. raptor, a
significant reduction in the number of house flies on the poultry farm resulted
(Shepard & Kissam 1981). When Diflubenzuron was fed to chickens, fly control
was obtained but residues were found in all egg samples (Miller et al. 1975).
Cyromazine is the only registered feed-through insect growth regulator for
caged poultry to control the house fly and other Diptera. Numerous studies on its
effect on dipterous insects have been performed (Hall & Foehse 1980, Williams &
Berry 1980, Mulla & Axelrod 1983a, 1983b, Meyer et al. 1987). Cyromazine had
no adverse effects on populations of manure-inhabiting mites which prey on fly
eggs and larvae (Axtell & Edwards 1983). Populations of beneficial Staphylinidae
and Histeridae were also unaffected by treatments of

12
cyromazine (Meyer et al. 1984 ). Cyromazine was found to be the most cost
effective treatment and provided the highest level of control in small commercial
caged layer poultry houses (Quisenberry & Foster 1984).
In laboratory studies, a 10% soluble concentrate of cyromazine in rearing
media caused 100% mortality of 0. aenescens at levels between 0.025 - 0.10%
(Hogsette 1979). No pupae were found; all flies died in the larval stage. A 1.5
ppm level of feed-through cyromazine had no effect on the 0. aenescens ' ' '
population in a poultry house in South Carolina (Nolan & Kissam 1985).
In laboratory studies, tolerance to cyromazine was detected in a wild,
organophosphate-resistant house fly strain (Bloomcamp et al. 1987). In field
studies at poultry farms in southern New England, cyromazine was ineffective after .
one and a half years of intermittent use at one farm and at a second farm,
adequate fly control was never achieved {Anderson et al. 1986). Reasons for this
were not understood but resistance was suggested.

1 . I
1
I ,I
/.
THE EFFECT OF DIFFERENT PROTEIN LEVELS IN LARVAL AND ADULT
DIETS ON THE DEVELOPMENT AND REPRODUCTION OF OPHYRA
AENESCENS (WIEDEMANN) (DIPTERA: MUSCIDAE)
Ophyra aenescerzs (Wiedemann) (Diptera: Muscidae ), a facultative
predator, has beeh.proposed as a biological control agent for house flies in caged
layer poultry houses (Nolan & Kissam 1985). When introducing predators for use
in an IPM program, it is important to produce sufficient numbers of the predator
efficiently and economically. Various diets have been used to rear the 0.
aenescerzs larvae using the Chemical Specialties Manufacturing Association
preparation (CSMA). Johnson & Venard (1957) usedCSMAwith diamalt, fish
meal, yeast and brewer's yeast while Hogsette (1979) used a similar diet containing
CSMA, wheat flour, fish meal, N<liC03, and alfalfa meal. Nolan (1987) simplified
the ingredients by using CSMA with soybean meal and chicken eggs.
Ingredients for the adult food have also varied; the adult food used by
Johnson & Venard (1957) consisted of dry fish meal, but Hogsette (1979) used a
nasogastric mix with fish meal. Nolan (1987) used a different mixture consisting of
powdered milk, sugar and· a chicken egg.
Because the diets were either costly or time consuming to produce, a less
expensive diet with simple ingredients was needed to rear 0. aenescerzs. Following
a suggestion by Robert D. Hall (personal comm.), a larval diet of coarse wheat
bran, vermiculite, dried meat powder (50% protein) and a chicken egg was used.
Preliminary tests indicated that this diet worked well. The ingredients for the adult
diet consisted of powdered.milk, sugar and dried meat powder .
Since the dried meat powder is an important part of both diets, the
13

14
objective was to. determine the optimum amount of protein needed in the diet to
rear healthy flies without reducing the number offlies emerging. Experiments
conducted were todetermine : (1) the number of flies emerging from media types
with varying levels of protein thathad been a) placed in the 0. aenescens rearing '
room, and b) seeded with a. known number of first instar 0. aenescens larvae,· (2)
the .effect of varying levels of protein in larval media on the Fl adult 0. aenescens
longevity and fecundity, and (3) the effectof different levels ofprotein in adult
food on longevity andfecundity of adultO. aenescens.
Materials and Methods
Effect of different protein levels in larvaldiet on the number of flies
emerging from. the media. The standard. diet for rearing larvae consists of, by
volume,. eight parts vermiculite, twelve parts wheat bran, one part dried meat
powder (protein supplement), and one chicken egg. The proteinleveland the
presence of the chicken egg were varied in this test. The proteinJevelswere: (1)
dietwith no protein supplement (OX); (2) dietwith half the protein supplement
(1/2X); (3} the standard diet (lX) and (4) dietwith twice the protein supplement
(2X). Because the egg might supply additional protein to the larvae, the presence
of eggs was also .examined using these four diet types (EOx, El/2x, Elx and E2x).
Individual diets (237ml) andwater (enoughto moisten diet)were
combined in small containers (474 ml) and replicated three times. Inthe egg
treatments, 10 ml of raw beaten egg was mixed into the top of the medium. Cups
were placed randomly in a walk-in rearing room, which had a controlled
temperature (27°C) with continuous light and constant air flow. Only one diet
type was.placed into the rearing room at a time. Females were allowed free access

15
to oviposit in the cups for 24 h. To allow for larval growth, the medium was
transferred to a larger container which held· an additional 4 7 4 ml of the wet
corresponding medium. A layer of vermiculite was added on top to prevent mold
growth (Born 1954). The containers were coyered with a screen top and placed
into an environmental chamber (27°C) and the emerging flies counted. Data were
analyzed by the Kruskal-Wallis test (SAS Institute· 1985); ·Amino acid analysis was
determined for larval diets 2x, 1x, and Ox from the Food Science Department,
Clemson University; Clemson, South Carolina.
Effect of different protein levels in larval diet on the adult fly·emergence
from a known number of 0. aenescens larvae. In a similar experiment, diets
containing different protein levels were mixed as before and one hundred and fifty
first-instar 0. aenesceris were seeded into each diet. A layer of vermiculite was
added on top. Containers were covered with a screen top and placed into an
environmental chamber (27°C). Emerging adults were counted. Data were
analyzed using two factor ANOV A and the LSD test for means separation (SAS
Institute 1985).
Effect of different protein levels in the adult diet on longevity and fecundity
of .Q. aenescens adults. The standard adult diet consisted of two parts dried milk,
two parts sugar and one.part dried meat powder. The amount of dried meat
powder in the food was varied in this test as follows: (1) A2 with 480 ml of dried
meat powder; (2) Al with 240 ml of dried meat powder; (3) Al/2 with 120 ml of
dried meat powder; and ( 4) AO with dried milk and sugar and no dried meat
powder; The final treatment used no adult food, just oviposition medium and
water (AH20). Qviposition medium was coarse wheat bran and vermiculite.

16
When the medium was taken out of the adult cages and transferred to bigger
containers, a protein supplement was added to provide a source of nutrition for the
developing larvae.
ln·adult experiments, ten newly·emer~ed male.and female flies were
anesthetized with C02 and placed into rearing cages (30x30x35 ml). Each adult
diet was replicated three times. Cages were kept in a cabinet with a constant
temperature of 30°C. Adult food was provided (120 ml) in 240 ml cups and
changed when needed. Water was alsoprovided in paper cups with Styrofoam™
shavings to reduce mortality due to drowning. Dead flies were recorded and
removed from the cage each day. Oviposition medium was provided on day 2 and
transferred the following day to a bigger container. It was then placed into a
cabinet with a constant temperature of 30°C. The medium was changed everyday
until all females were dead. After five days,. larvae were counted and recorded.
Larvae were counted because it was difficult to count accurately the eggs deposited
in the medium. It was realized that some egg and larval mortality could occur but
since all treatments were similarly evaluated, the results should be comparable
(Turner & Hair 1967).
The· number of larvae per female fly-days was based on the total number of
larvae collected, divided by the total.number of days the females lived. Longevity
for. each replicate was determined by using the median age of the flies in each
cage. Data were analyzed using two factor ANOV A and the LSD test for means
separation (SAS Institute 1985).
Fecundity and longevity of.Fl O. aenescens flies reared from different levels
of protein. In a final experiment, the flies that emerged from the larval media. test
(rearing room test) were collected to determine if they produce viable offspring.

17
Ten newly emerged male and female flies were anesthetized with co2 and placed
into rearing cages (30x30x35 ml); These first generation flies were treated in a
similar manner as the flies in the previous adult food experiment. Adult food was
a 2-2-1 ·ratio of the sugar, powdered milk and.dried meat powder respectively.
Results
Based on amino acid analysis, the percent of crude protein in the diets,
using.amino acid analysis, was 12% for Ox, 17.1% for lx, and 245% for2x.
Although 1/2x medium was not analyzed, extrapolating from the results of the
other diets, 1/2xwould contain approximately 14.Sto 15% crude protein.
Effect of different protein levels in larval diet on the number of emerged
flies. Flies oviposited in all media types (Table 1). Significant differences were
found between diets (Chi-square =0.0056). Due to the small number of replicates,
a mean separation test was not possible. There are trends that can be observed
from the· data. Diets E2xand El/2x had large numbers offlies·emerging from the
diet while diets Ox and 1/2x had the smallest number emerging. ·The addition of 10
ml of raw beaten chicken egg to the diets increased the number of flies emerging
except for dietElx. ·Reasons for. this poor performance of Elx are· unclear.
Effect of different protein levels in the larval diet on adult fly emergence
from a known number of 0. aenescens larvae. Two factor analysis on larval diets
indicate a significantinteraction between larval media with protein and the
presence of eggs (Table 2). For media with an egg on top, EOx was significantly
different from diets El/2x, Elx and E2x. There were similar findings with the
diets with no egg on top (diet Ox was significantly different from diets 1/2x, 1x and
2x). Comparing the media with egg to the media without egg, only EOx and Ox ·

18
were significantly different. Lack of protein in the larval medium can affect the
emergence of flies. A protein source is thus needed because a lack of protein will
reduce the number of flies emerging. The addition 10 ml of raw beaten chicken
egg to the medium increased the number ·of t;lies emerging when no other protein
source was present (EOx vs. Ox).
Fecundity and longevity of Fl adults 0. aenescens reared·from different
larval protein levels. Statistically, .there were no differences in fecundity
(P=0.0836) for the Fl generation 0. aenescens flies reared from the different
larval diets (Table 3). Numerically, the larvae per female fly-days were similar in
diets 2x, 1x and l/2x (21, 20 and 2llarvae per female fly-days respectively).
Although the Ox diet was lower ( 4 larvae per female fly-days) it was not
significantly different.
Two factor analysis indicates a significant difference in longevity of Fl
generation flies reared on different larval diets (P = 0.0016). Diets 2x and lx were
significantly different from l/2x and Ox (Table 3). Fl male flies lived significantly
longer than Fl females (P=0.0001) (Table 3). Longevity of Fl males due to the
larval diets (P=0.0399) was statistically different (Table 3). Fl males reared in lx
media lived the longest (32 days). Males reared in diets 2x, l/2x, and Ox were not
significantly different, having longevities of 24, 16, and 20 days respectively.
Differences were also found in female longevity (P=0.0230) (Table 3). Female
longevity was longest on lx media (17.8 days). Results from diets2x and l/2x were
similar and Ox diet yielded the shortest lived flies (9 days).
Effect of different protein levels in the adult diet on longevity and fecundity
of Q. aenescens adults. No significant difference in the number of larvae produced
due to different levels of protein in adult diets was.found (P=0.0545) (Table 4).

19
Fecundity ranged from. 27 larvae per female fly-days to 11 larvae per female fly-
days for diet AO; even in the treatment with no adult food (AH20), the flies still
laid eggs (13 larvae per female fly-days).
Two factor analysis on adult diets indi~ate a significant difference in
longevity between diets for 0. aenescens.flies (P=0.0001) (Table 4). Diets A2, Al,
Al/2 and AO were significantly different from the AH20treatment (Table 4).
Male flies lived significantly longer than did female flies (P=0.0001). There was
also a significant interaction between adult dietand sex (P=0.0016). For the
treatment of no adult food (AH20), the longevity for male flies was lower than the
female longevity for that treatment.
Longevity was not significantly different between females feeding on the
different adult diets (P=0.0631} (Table 4). Male longevity differed between adult
diets (P=0.0002) (Table 4). The male 0. aenescens lived the longest time on diet
Al/2 (33.6 days) but was not significantly different from diets Al .and A2 (27.8 and
25.3 days respectively) .. Longevity for males significantly decreasedtothree days
when given no adult food (AH20).
DISCUSSION
Larval diet. Female 0. aenescens show a preference for media with high
protein levels with a chicken egg on top. Media containing diets 2x, E2x, lx and
El/2x all produced similar numbers of emerging flies. Diet Elx had enough
protein, .even more than El/2x diet, but it did not attract as many ovipositing flies
as the others. It is uncertain why this occurred. Conditions within the rearing
room such as humidity and temperature, may change from day to day. These

20
conditions may affect the fly's abilitytoJay eggs or they may· not be favorable for
the fly to.lay eggs atthattime;· The number of ovipositingfeniales in the rearing
room also may vary from day to day to cause the difference in the.number of Fl
flies emerged~ The amount ofprotein in the <!iet and the presence of eggs appears
to affect female 0.: aenescens choice of;oviposition site but :because there was
considerable variation. in the·.·experiment .this ·is not conclusive .. ·
... • Proteili levels.in:the. larval media:is important in the development of .the
instars and emergence of the fly. The 10 ml of raw beaten chicken egg placed on
top of the media was not enough to, make a significant difference .in the number
emerged. The one exception was EOx medium (10R67.) which had significantly
more flies emerging thariOx (30.17)~
The.longevity of Fl generation adults was significantly greater when the
larvaewerereared on diets 2x andlx~ Flies lived20.:25 days:onthose diets while
adults livedorily 14 days oll'diets 1/2x and.Ox.·.The Fl males lived significantly
longer (23 days) than the. female flies• (13 days) .. Adult males emerging from the
different larval media lived the longest on the lx media{33 days) and was .
significantly differentfromthose exposed: to diets l/2x (16 days).and Ox (20 days).
Females lived the longestwhen reared.from diets J.X,(17.8 days) and:2x (163 days).
These diets were significantlydifferentfrom Ox diet(9days). < ·.
Fecundity was similar for diets 2x/lx and l/2x (21.2, 20.021.0.larvae per ·
female fly-days, respectively);, Females reared on Ox. had.a lower fecundity ( 4.4
larvae per female fly-days), but was not significantly different fromthe other diets.
A minimum amount of protein· in the larval dietis needed forlongevity and
fecundityofadultO~ aeriescens flies~ . ·
·. Adult Diet· 'J'herewas no difference inJoilgevity offlies.reated on different

l I
21
adult diets but these diets were significantly differentfrom the treatment with no
food; Adult male longevity was similar for diets Al/2, A2 and A1(34, 28, 25 days
respectively) and was notdifferentfromdietAO (20 days).; However; longevity was
significantly reduced when flies were not give:µ adult ·food at all (A.HiO, 3 days). ··
Varying thelevels.ofprotefu in theadultdietmadelittle difference in the female
fly longevity. Female longevity decreased with protein levels but when not given
any adult food, the female flies stilllived an average of8 days~ Diets were not
significantly different for female fongevity at all levels. Varying the protein
amount (ie. dried meat powder) in adult diets has little effect on adult longevity. It
may be that dried milk and sugar are adequate for maximum longevity.
Fecundity of the flies for diet A2 was 27 larvae per female fly-days but was
not significantly different from diets AO and AH20 (11, 13 larvae per female fly-
days respectively). It seems that fecundity of adult flies is not influenced by the
protein in adult diets. In the AHzO treatment, female 0. aenescens still laid viable
eggs even when no protein source was given. The oviposition media contained
only wheat bran and vermiculite. ·In contrast, Johnson and Venard ( 1957) reported
that no eggs hatched until flies fedonanimal flesh. Glaser (1924) stated that sugar
or starch together with protein are necessary oviposition factors for house flies. A
diet of protein was needed for the maturation of ova for blowflies (Mackerras
1933). My results indicate that protein content in the vermiculite and wheat bran
must be adequate for maturation of the ova in the 0. aenescens female.
Both Johnson and Venard (1957) and Hogsette (1979) reported that
females lived 3-4 days longerthan the males (reared at26.6°C). This opposes the
results presented here. In all my experiments the males lived longer than the
females. The rearing temperature (30°C) might be the reason for this. Similar

22
results occurred with the adult house fly when reared at high temperatures (35°C)
(M. G. Fletcher, personal communication). The higher temperature did affect the
fecundity of the house flies but 50% of the eggs were laid sooner at this
temperature than at 30°C. 0. aenescens fem~es would probably outlive males at
temperatures lower than 30°C. Research is needed to confirm this.

23
Table 1. Mean number of flies emerged and standard error (SE) of Ophyra aenescens reared on different larval diets placed into the rearing room.a
Diet Mean SE(±)
E2x 1908.7 236.6
El/2x 1818.7 126.1
2x 1123.7 340.8
lx 923.7 231.9
Elx 170.3 79.4
EOx 141.0 101.4
Ox 54.3 33.8
1/2x 27.7 11.3
aDue to the small number of replicates (n=3), a means separation test was not carried out. ·

24
Table 2. Mean number and standard error (SE) of flies emerged from 150 first instar Ophyra aenescens seeded into larval diets with varying protein levels.
Diet Type
Ox 1/2x 1x 2x
Mean Emergence0 ±SE
Egg 108.7 ±7.87b * 125.7±4.53a 130.2±4.61a 135. 7 + 2.57 a
No egg 30.2±8.16c 113.3±15.06a 121.3±8.77a 128.7±3.47a
0 Means with the same letterin rows are not significantly different.
*Means in this column were found to be significantly differenL There were no differences in the other columns.
LSD test used to separate means (P = 0.05).

Table 3. Fecundity and longevity with standard error (SE) for Fl male and female Ophyra aenescens reared from different larval protein levels.
Fecundity" Diet Meanb
Mean Longevity (days)+ SE Males Females
Mean Longevity+ SE of adult flies
2Jf 21.24+3.84a 24.16± 4.68ab 16.33±0.33ab 20.25±3.87a
lx 20;04±5.99a 32.83±4.04a 17.83±1.92a 25.33 ±5.53a
1/2x 21.03±5.32a 16.33±0.92b 11.16±2.52bc 13.75 +2.36b
Ox 4.44±2.14a 19.83±2.45b 9.00±l.52c 14.42±3.89b
, Meand 23.29a 13.58b
acalculationmade by dividing the total number of larvae produced by the total number of days the female Ophyra aenescens lived (Larvae/female fly-days).
bMeans with the same letter in each column are not significantly different. LSD test used to separate means (P = 0.05).
c2x, lx and 1/2x represent decreasing amounts of protein in larval diets. Diet Ox contains no protein, only vermiculite and wheat bran.
d Average longevity of male and of female 0. aenescens for all diets. Means with different letters are significantly different.
N V1

Table 4. Fecundity and longevity with standard error (SE) for male and female Ophyra aenescens fed adult diets with varying protein levels.
Fecundity" Mean Longevity (days )±SE Mean Longevity±SE Media .Mean±SEb Males Females of adult flies
A2c 26.87±1.94a 27.83±4.26ab 19.33±2.03a 23.59 +4.0la
Al 20.75±5.05a 25.33±3.38ab 15.16±1.59a 23.58±3.99a
Al/2 17.84±1.35a 33.66±1.74a 13.50±2.93a 20.25±6.73ab
AO ll.13±4.36a 19.50±3.0lb 13.50±2.93a 16.51 +3.27b
AHzO 13.32±2.75a 2.66±0.33c 8.33±0.88a 5.50+ 1.89c
Meand 21.80a 13.97b
°Calculation made by dividing the total number of larvae produced by the total number of days the female 0. aenescens lived (Larvae/female fly-days). ·
bMeans with the same letter in each column are not significantly different. LSD test used to separate means (P = 0.05). ·.
c A2, Al and Al/2 represent decreasing amounts of protein in adult diets, AO diet contains only sugar and dried milk, and AH20 represents treatment with no adult food given.
d Average longevity of male and of female 0. aenescens for all diets. Means with different letters are significantly different.
N
°'

27
LITERATURE CITED
Born, D. E. 1954. Mold control in fly rearing media. J. Econ. Entomol. 47: 367.
Glaser, R. W. 1924. The effect of food on longevity and reproduction in flies. J. Exp. Zool. 38: 383-412. '
Hogsette, J. A., Jr. 1979. The evaluation of poultry pest management techniques in Florida poultry houses. Ph.D dissertation U. of Fla. Gainesville, Fl 307 pp.
Johnson, W. T. & C. E. Venard~ 1957. Observations on the biology and morphology of Ophyra aenescens (Diptera: Muscidae). Ohio J. Sci. 57: 21-26.
Mackerras, M. J. 1933. Observations on the life-histories, nutritional requirements and fecundity of blowflies. Bull. Entonml. Res. 24: 353-362.
Nolan III, M. P. 1987. Development of an integrated pest management program for the control of the house fly Musca domestica L., in caged layer poultry houses in South Carolina. Ph.D dissertation Clemson Univ. Clemson, S. C. 179 pp.
Nolan III, M. P. & J.B. Kissam. 1985. Ophyra aenescens: A potential bio-control alternative for house fly control in poultry houses. J. Agric. Entomol. 2: 192-195.
SAS Institute. 1985. SAS user's guide: statistics, 5th ed. SAS Institute, Cary, N. C.
Turner Jr., E. C. & J. A. Hair. 1967. Effect of diet on longevity and fecundity of laboratory-reared face flies. J. Econ. Entomol. 60: 857-860.

THE EFFECT OF CYROMAZINE ON OPHYRA AENESCENS (WIEDEMANN)
(DIPTERA: MUSCIDAE)
Accumulated manure of deep pit poultry houses is an ideal environment for '
the development of the house fly, Musca domestica L. Dispersal of this fly into
nearby residential areas has become both a nuisance and a public health threat.
This has resulted in new regulations governing the suppression of these insects
(Anderson et al. 1986).
Feed-through insect growth regulators are an ideal way to control house flies
because this method eliminates most labor and equipment costs of house fly control
(Mulla & Axelrod 1983b, Miller 1970); Cyromazine (Larvadex™) (Ciba-Geigy
Corp., Greensboro, NC), an insect growth regulator, is incorporated into chicken
feed and prevents development of house flies (Hall & Foehse 1980, Williams &
Berry 1980, Mulla & Axelrod 1983, Anderson et al. 1986).
Cyromazine activity is thought to be limited mainly to dipterous insects and
has been shown not to affect manure-inhabiting predaceous mites and beetles
adversely (Axtell & Edwards 1983). Ophyra aenescens (Wiedemann) (Diptera:
Muscidae ), a facultative predator, has been proposed as a biological control agent
against house flies in caged layer poultry houses (Nolan & Kissam 1985). They also
reported that use of cyromazine at a high rise poultry farm appeared to have no
effect on 0. aenescens.
It is important to determine if 0. aenescens and cyromazine can be used
together in an IPM program to control house flie.s. The objective of this study was
to determine the toxicity ofcyromazine to 0. aenescens and compare it to the
toxicity of a susceptible house fly strain.
28 ! I '

29
Materials·and Methods
Cyromazine toxicity in artificial media. Cyromazine was incorporated into
artificial larval media. The larval media consi~ted of eight parts vermiculite, twelve
parts wheat bran and one part dried meat powder. Weighed amounts of
cyromazine from a 1 % premix were added and mixed to 100 g of artificial rearing
media to obtain concentrations of 0.0, 0.75,1.0, 1.5, 2.0, 25 and 3.0 ppm. Each
concentration was replicated three times. About 250 ml of water was mixed into
each container to provide a suitable substrate for larvae. One hundred first instar
0. aenescens (from laboratory colony) were seeded into each container (800 ml),
covered with a screen top and placed into temperature controlled cabinets
maintained at 27°C. Emerging adults were counted and removed. This experiment
was also conducted using susceptible house fly.larvae .. The laboratory colony of
house flies consisted of flies from a poultry farm that did not use cyromazine. Data
were analyzed using probit analysis on the Statistical Analysis System (SAS Institute
1985).
Cyromazine toxicity in chicken manure. Cyromazine was tested as a feed-
through larvicide in chicken feed. Various dosages of a 1 % premix of cyromazine
were mixed into.22.5 kg (50 lbs) of standard layer chicken feed for 10 min. using a
H600 Hobart feed mixer. Dosages were: 0.0, 0.5, 0.65, 0.85, 1.0 and 1.5 ppm. The
experiment was conducted at the Virginia Tech Poultry Research Center using 61
week-old White Leghorn chickens. Five.birds represented a replicate and four
replicates were used for each treatment rate. The feedwas given to the birds for
seven days, after which the food.trough and water system were cleaned and refilled
with untreated food and water. Samples of manure were collected one day before I I l' ,, I
11

30
treatment, days 1, 3, and 7 during treatment and days 8 and 14 post treatment. The
manure samples were collected separately from each replicate (total manure of five
birds), placed in labeled plastic bags and frozen. Prior to bioassay, samples were
thawed for 24 h to room temperature. An eqqal amount of manure (237 ml) from
each sample was placed into two plastic containers (474 ml)and seeded with either
100 first instar 0. aenescens or with 100 first instar.house·flies. Containers were
held at ambient temperature (21-27°C) and relative humidity. All flies that
emerged were allowed to die before counting. Data were analyzed using probit
analysis on the Statistical -Analysis System (SAS Institute 1985).
Results and Discussion
There were too many data points above the 60% mortality range in the
lower treatments. This caused the pro bit analysis to estimate the LC50 value with
large fiducial limits. Thus, the LC90 value was used to compare 0. aenescens and
M. domestica.
Test using artificial media. Ophyra aenescens tolerated higher
concentrations of cyromazine thanM. domestica(Table 5). Fewer 0. aenescens
died at 0.75 ppm (27%) compared with the house fly (65%). As the dosage of
cyromazine increased, mortality increased for both species. The LC90 value for 0.
aenescens was 1.27 and 1.11 for M. domestica (Table 6). Fiducial limits did overlap
for the LC90 values, indicating that they are_ not significantly different. Percent
mortality was high (Table 5) in the 0.75 ppm and 1.0 ppm treatment. 0. aenescens
may be more tolerant of lower doses of cyromazine (below 0.75 ppm) compared
with the house fly but mortality increases with cyromazine dosages above 0.75 ppm.
------
I. 1:

31
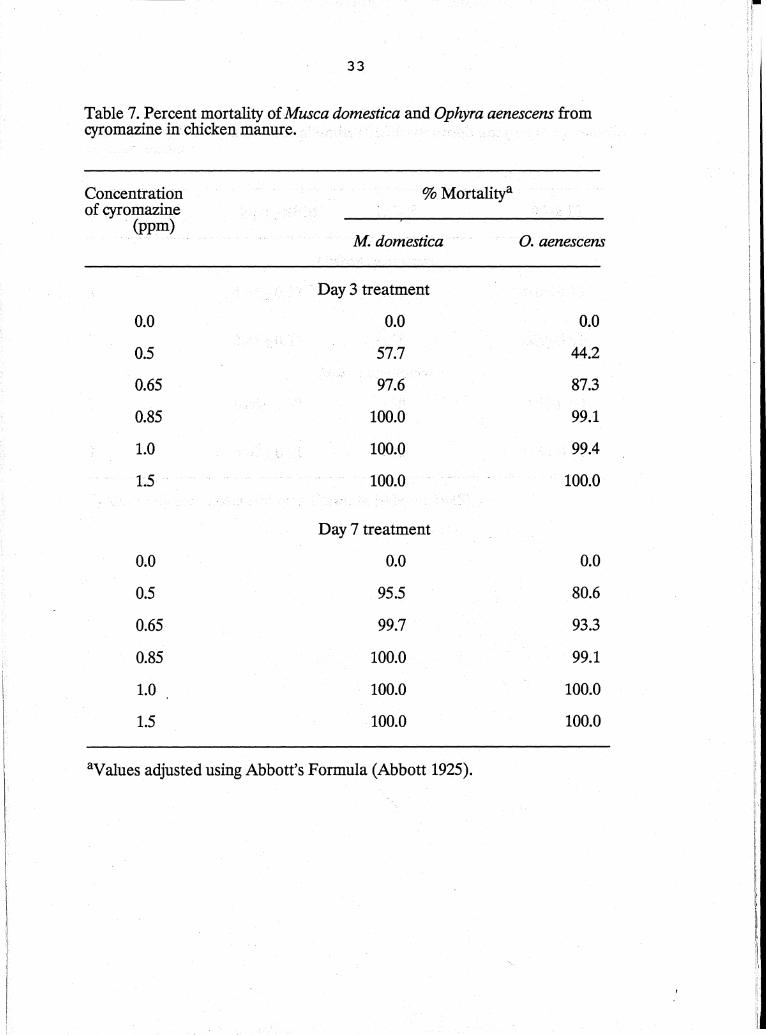
Test using chicken manure. Ophyra aenescens and house fly mortality
increased from day 3 to day 7 (Table 7). The LC90 values for 0. aenescellS tested on
days 3 and 7 were 0.68 and 0.58, respectively (Table 8). Musca domestica bioassay
resulted in LC90 values of 0.58 and 0.45 for days 3 and 7. The fiducial limit values
for 0. aenescellS and the house fly did not overlap.indicating they are significantly
different. The house fly percent emergence was lower in the post treatment (day 8)
compared with the control suggesting that some cyromazine was present in the
manure (Table 9). The percent emergence in 0. aenescellS (day 8) remained the
same in all treatments. One week after cyromazine was removed (day 14 ), no
differences were found in percent emergence between the control and treatments
for either the house fly or 0. aenescellS.
The data indicate that 0. aenescellScan tolerate higher dosages of
cyromazine thanthe house fly. This would agree with the findings Nolan & Kissam
(1985) where 0. aenescellS became the dominant fly in poultry houses after
cyromazine was used as a control for house flies. In an IPM program using 0.
aenescellS and cyromazine at doses of 1.0 ppm and lower, releases of 0. aenescellS
might be possible one day after cyromazine use has stopped. More detailed
research is needed to investigate the effects of cyromazine on 0. aenescellS at lower
doses. Also field studies need to be performed to understand how cyromazine
affects the dynamics of the 0. aenescellS population over time in poultry houses.

32
Table 5. Percent mortality of M-usca domestica and Ophyra aenescerzs from cyromaz~e in rearing media.
Concentration % Mortalitya of~omazine
pm) M. domestica 0. aenescerzs
0.0 0.0 0.0
0.75 65.1 26.7
1.0 81.3 74.4
1.5 98.1 94.7
2.0 100.0 99.4
2.5 100.0 100.0
3.0 100.0 100.0
avalues adjusted using Abbott's Formula (Abbott1925).
Tab~e 6~ SloJ?e,. LC90 and fiducial limits (FL) from pro bit analysis of cyromazine in rearing media. · •· ·
Slope±.SEM
M. doinestica 2.45+0.31
0. aenescerzs 3.34_±_0.26
LC a 90
.. • 1.11
1.27
avalues adjusted with Abbott's Formula (Abbott 1925).
95%FL
1.06-i.17
1.17-1.49
33
Table 7. Percent mortality of Musca domestica and Ophyra aenescens from cyromazine in chicken manure .... ·

34
Table 8. Slope, LC90 and fiducial limits (FL) from probit.analysis of cyromazine in chicken manure.
Day Slope±SEM LC90 a 95%FL
Ophyra aenescerzs
3 4.44±0.52 0.68 0.64-0.75
7 2.85±0.21 0.58 0.56-0.61
Musca domestica
3 6.81±1.37 0.58 0.57-0.60
7 4.07+0.91 0.45 0.38-0.48
avalues adjusted using Abbott's Formula (Abbott 1925).

35
Table 9. Percent emergence of Musca domestica and Ophyra aenescens from chicken manure one day after and one week after· cyromazine was removed from food.
Dosage
0.0 0.5 0.65 0.85 l.O l.5
Musca domestica Day8
93.0 100.0 89.0 29.0 48.0 8.5
Day 14
85.0 87.0 85.0 97.0 86.0 77.0
Ophyra aenescens Day8
60.0 60.0 63.0 62.0 67.0 41.0
Day 14
75.0 63.0 61.0 63.0 56.0 94.0
'I i 1.

36
LITERATURE CITED
Abbott, W. S. 1925. A method for computing .the effectiveness of an insecticide. J. Econ. Entomol. 18: 265-267.
Anderson, J. F., A. Spandorf, L.A. Magnarelli and W. Glowa. 1986. Control of house flies in commercial poultry houses in Connecticut. Poultry Sci. 65: 837-844.
Axtell, R. C. & T. D. Edwards. 1983. Efficacy and nontarget effects of Larvadex™ as a feed additive for controlling house flies in caged-layer poultry manure. Poultry Sci. 62: 2371-2377.
Hall, R. D. & M. Foehse. 1980. Laboratory and field tests of CGA-72662 for control of the house fly and face. fly in poultry, bovine,. or swine· manure. J. Econ. Entomol. 73: 564-569.
Miller, R. W. 1970. Larvicides for fly control-A review. Bull. Entomol. Soc. Am. 16: 154-158.
Mulla, M. S. & H. Axelrod. 1983b~ Evaluation of the IGR Larvadex™ as afeed-through treatment for the control of pestiferous flies on poultry ranches. J. Econ. EntomoL 76: 515-519. ·
Nolan III, M. P~ & J.B. Kissam. 1985. Ophyra aenescens: A potentialbio-control alternative for house fly control in poultry houses. J. Agric. Entomol. 2: 192-WS. .
SAS Institute. 1985. SAS user's guide: statistics, 5th ed. SAS Institute, Cary, N. C.
Williams, R. E. & J. G. Berry. 1980. Evaluation of CGA 72662 as a topical spray and feed additive for controlling house flies breeding in chicken manure. Poultry Sci. 59: 2207-2212.

THE EFFECT OF TEMPERATURE AND MANURE MOISTURE ON THE
PREDATORYliABITS OF OPHYRA AENESCENS (WIEDEMANN) ON MUSCA
DOMESTICA L.
Temperature and manure moisture are important factors regulating the
development of arthropod pest populations in caged layer poultry houses.
Temperature and accumulated manure in high.rise poultry houses provide
favorable conditions for development offlies year round (Stafford & Collison
1987). Optimum conditions for house fly, Musca domestica L., development are
21°c (Miller et al. 1974) and a manure moisture content of 7.0% (Peck and
And~rson 1969). Optimum development of Ophyra aenescens {W,iedemann), a
facultative predator ofthe house fly, was 26.6°C(Johnson & Venard 1957). Ophyra
leucostoma (Wiedemann), a i:elated species, was most abundant in areas with a 63% c
· manure moisture content in poultry houses in California (Peck & Anderson 1969). . . .
Temperatures in pits of poultry houses in Bent Mt., Virginia ranged from a
maximum of 21°c to a minimum of 14.8°C, and an average temperature of 21.8°C
which will support a house fly population (unpublished data).
The larvae of Ophyra spp. are predaceous on.other dipterous larvae
(Anderson & Poorbaugh 1964a, Peck 1969, Peck & Anc.lerson 1969),but not
cannibalistic (Anderson & .Poorbaugh 1964b, Hogsette 1979). In laboratory studies,
Nolan &Kissam (1985) observed.O. aenescens feeding on Musca domestica,
Phormia regina (Meigen) and Sarcophaga sp.larvae. Hogsette (1979) found that D.
aenescens could destroy more than20 first-instat house fly larvae per day during
larval development.
Ambient temperature and moisture content. of the manure may affect. 0.
37

38
aenescens' ability to control house fly larvae. The purpose of this study was to
determine the effect of temperature and manure moisture levels on the ability of 0.
aenescens larvae to control house flies.
Materials and Methods
Varying house fly to 0. aenescens ratios and manure moisture. Fresh
poultry manure was collected from the Virgiriia Tech Poultry Research Center.
The manure was taken to Price's Fork Research Facility, frozen for 24 h and then
dried in an oven at 90°C for two days. One hundred grams of oven-dried manure
was added to a plastic container. Based on the weight of the manure, moisture
levels were prepared by adding a measured amount of water to the cup to condition
the manure to obtain three percentage moisture levels, 50, 60, and 70%. The
experiment was replicated three times for each moisture level. At these levels,
house fly and 0. aenescens larvae were added at different ratios, 3:1, 2:1and1:1,
(house fly to 0. aenescens) and controls were set up, 1:0, 2:0, 3:0 and 0:1. A top
layer of vermiculite was added to prevent mold growth. All containers were
weighed and put into a cabinet which maintained a temperature of 27°C and 50%
RH. The containers were weighed each day for five days (when pupae were
noticed) and water was added to maintain the proper percent moisture level. The
number of emerging flies was recorded;
Varying house fly to 0. aenescens ratios and ambient temperature. Fresh
poultry manure was collected from the Virginia Tech Poultry Research Center and
then frozen for 24 h. The manure was thawed and a 237 ml sample was put into
containers ( 474 ml). Percent moisture of fresh manure was between 60 and 70%.

39
House fly and 0. aenescens larvae were seeded into the containers at different
ratios, 3: 1, 2: 1 and J: 1 {house fly to 0. aenescens) and controls were used, 1 :0, 2:0,
3:0 and 0:1. Ratios were replicated three times. A layer of vermiculite was added
to the surface. The containers were put into ~o cabinets which had constant
temperatures of 21and27°C respectively. For a third temperature, 18°C, a cabinet
witha constant, low temperature was not available, therefore, containers were
placed into a cool roomwhere the temperature was recorded each day. The
temperature ranged from 17-20°C with an average of 18°C;·adequately
representing the effect of low temperature onO. aenescens; .Emerged flies were
collected and the numbers recorded. Data were analyzed using two factor
ANOV A, ttestand LSD means separation (SAS Institute 1985).
Results
Moisture test: Effect of. manure moisture and competition with M.
domestica on emergence of 0. aenescens. No differences in percent emergence
were found for 0. aenescens when combined with varying ratios of M. domestica
(3:1, 2:1, 1:1) as compared with the control (0:1) (P=0.8047). There was a
significant manure moisture effect. Emergence of Ophyra aenescens at 70 and 60%
moisture level was 84 and 77%, respectively (Table 10) which was significantly
differentfrom 46% emergence at the 50% moisture level (P = 0.0001 ).
Effect of manure moisture on percent emergence of M •. domestica in
controls. House fly emergence in the controls was 81.3, 68.8 and 74.1 % at moisture
levels 50,. 60 and 70% respectively (Table 11 ). Two factor analysis indicated a
significant interaction(P=0.0363) between moisture and controls for house fly

40
emergence .. Atthe 2:0control for 60% moisture, emergence was 51%(Table11).
This percent emergence was lower than any control at any moisture level. Reasons
. for this are unclear. At the other moisture levels percent emergence between
controls did not differ more than 8%.
Effect .of manure• moisture and competition with 0. aenescens on emergence
ofM. domestica. Eniergence forthe house flieswhen combined withO. aenescens
was significantly lower from the house fly controls. At the 70% moisture level,
differences in house fly emergence were found at all three ratios (3:1, 2:1, 1:1)
(P=0.0011, P=0.0004, P=0.0001) (Fig. 1). At the 60%1evel, house fly emergence
was significantly different from the controls (P=0.0001, P=0.0139, P=0.0002 for
ratios 3:1, 2:1, 1:1, respectively) (Fig. l). Significant differences from the controls
were also found for house fly emergence at the 50% moisture level (P=0.0017,
P=0.0332, P=0.0003 for ratios 3:1,2:1, 1:1, respectively) (Fig. 1). The 0. aenescens
predation rate was significantly lower at the 50% moistureJevel (P = 0.0341) than at
other levels (Table 11 ). I tis assumed house flies died due to 0. aenescens
predation rather than other causes because the house fly emergence in controls was
at its highest in the 50% moisture level (Table 11).
Temperature Test. Effect of Temperature and competition with Musca
domestica on emergence ofOphyra aenescens. There were no differences between
0. aenescens emergence at the different ratios (3:1, 2:1, l:l)compared withthe
control (0:1). The developing house fly larvae had no detrimental effects on 0.
aenescens. There was however, a significant temperature effect(P=0.0001). The
percent emergence ofO. aenescens inthe 18°Ccontainers was 14.6% while at 27°
and 21°c the rate of emergence was 87 and 83.3% respectively (Table 12).
Effect of temperature on M. domestica inthe controls. For the house fly

41
controls, there was a decrease in the number of flies emerging at the 18°C level
(P = 0.0002). The percent emergence at 18°C was 67% as compared with 87 and
86% at 27° and 21°c, respectively (Table 13).
Effect of temperature· and competition, with 0. aenescens at different ratios
on the emergence ofM. domestica. House.fly emergence vary at the.three
temperatures (Table 13). At 18°C, there were no differences in house fly
emergence betweenthe controls (3:0, 2:0, 1:0) and the ratios (3:1, 2:1, 1:1),
(P=0.2858, P=0.0927, P=0.1802, respectively)•(Fig.2). At 21°c, there was no
difference in emergence at the· 3: 1. ratio (P = 0.1093), but there. were. differences at
the 2:1and1:1 ratiosas compared with the controls (P=0.0041, P=0.0018) (Fig. 2).
At the 27°C, differences in house fly emergence were found at all three ratios (3:1,
2:1, 1:1) (P=0.0005, P=0.0080, P=0.0145) (Fig. 2).
Ophyra aenesceru develops and emerges at 27° and 21°c but not as well at
18°C. This in turn affects its ability to prey on house fly larvae. The house fly also
had a decrease in emergence at 18°C but it is not clear whether this reduction was
due to 0. aenesceru predation or due to the effects of temperature.
The data indicate that 0. aenesceru emergence is influenced by temperature
and also by a constant moisture level. Emergence of 0. aenesceru was highest at
27°C and at constant moisture levels of 70% in chicken manure. Predation on
house fly larvae by 0. aenesceru seems to follow these conditions as the highest
predation of house fly maggots was found at 27°C and at 70% moisture level.
House fly emergence was better than 0. aenesceru in 50% moisture; in fact
M. domestica emergence was highest at 50% moisture while 0. aenesceru
emergence was lowest at the 50% moisture level. It seems M. domestica has a
greater moisture range for development than 0. aenesceru.

42
Although there was significant difference for house fly emergence between
controls and treated containers in the temperature test, there wasn't a dramatic
reduction in house fly emergence as was the case in the moisture test. Temperature
might be a factor for this in the 21° and 18°C ~est, but a lack of constant moisture
in the 27°C could be the reason for the increase in the house fly emergence. The
drying of manure in the temperature test may have hampered 0. aenescens's ability
to seek out house fly larvae. Since they are facultative predators, this would not
hurt 0. aenescens emergence. It would seem constant moisture (as in the moisture
test) helps with 0. aenescens predation on house fly larvae while the lack of
moisture (as in the 27°C temperature test) seems to.hinder predation. Thus, 0.
aenescens would be a more effective predator in areas of wet manure.

43
Table 10. Effect of manure moisture and competition with Musca domestica on emergence of Ophyra aenescens.
Manure Moisture(%)
Ratioa 70 60 50
Percent Emergence
3:1 '77.3 77.3 46.4
2:1 85.3 82.6 34.4
1:1 85.3 62.6 60.0
0:1 90.4 85.3 42.4
Meanb 84.6a 77.0a 46.0b
aRatio of M. domestica to 0. aenescens.
bMeans with the same letter are not significantly different. LSD test used to separate means (P = 0.05).

l ~··
44
Table 11. Effect of manure moisture and competition with Ophyra aenescens on emergence 'Of Musca dome~tica.
Ratioa
3:1
3:0
2:1
2:0
1:1
1:0.
Meanb
Ratio
Controls
70
2.6
73.0
1.3
74~6
2.6
74.6
2.2a
74.lab
Manure Moisture ( % )
60'
Percent·Emergence
io 74.0
5.3
51.3
4.0
81.3
3.7a
68.8b
aRatio of M. domestica to 0. aenescens.
50
10.1
79.3
18.6
86.0
9.3
78.6
12.7b
81.3a
bMeans with the same letter in rows are not significantly different. LSD test used to separate means (P = 0.05).
,,

45
Table 12. Effect of temperature and competition with Musca domestica on emergence of Ophyra aenescens.
Temperature (°C)
Ratioa 27 21 18
Percent Emergence
3:1 88.0 76.0 21.2
2:1 85.2 94.4 16.0
1:1 86.4 73.2 5.2
0:1 88.0 89.2 16.0
Meanb 86.8a 83.2a 14.4b
aRatio of M .. domestic a to 0. aenescerzs.
bMeans with the same letter are not significantly different. LSD test used to separate means (P = 0.05).

46
Table 13. Effect of temperature and competition with Ophyra aenescens at different ratios on the emergence of Musca domestica.
Ratioa
3:1
3:0
2:1
2:0
1:1
1:0
Ratio
Controls
27
60.4
85.6
60.6
88.0
57.3
88.0
59.4a
87.2a
Temperature (°C)
2r
Percent Emergence
53.3
84.0
42.6
89.3
29.3
85.3
41.Tu
86.2a
aRatio of M. domestica to 0. aenescens.
18
54.6
61.3
54.0
68.6
56.0
70.1
54.8a
66.8b
bMeans with the same letter in rows.,are not significantly different. LSD test used to separate mean~ (~~-0.05).\

100
75 w u z w 0 50 °' w :::; w 00!
25
0
100
I 60% MOISTURE
80
w u
60 z w '-' £1: w :::; 40 w ~
20
0 3-0 3-1 2-0 2-1 1-0 1-1
RATIO OF HOUSE FLIES TO 0. AENESCENS
70% MOISTURE
3-0 3-1 2-0 2-1 1-0 1-1 RATIO OF HOUSE FLIES TO Q., ~~N!~CJ:~~-
UJ u z w
- --------100..--~~~~~~~~~~~~~~~~~~~---,
50% MOISTURE
75
~ 50 w :::; w 00!
25
0 +----'---'-r 3-0 3-1 2-0 2-1 1-0 1-1 RATIO OF HOUSE FLIES TO Q., ~q.l_E;~~E!l~-
Fig. 1 House fly emergence at 70, 60 and 50% manure moisture at different house fly to Ophyra aenescens ratios.
+:--....::..

w (J z w 0 0:: w :::!! w <>11
100 27 <t
75
50
25
0
3-0 3-1 2-0 2-1 1-0 1-1 RATIO OF HOUSE FLIES TO Q. ~~ti_E.§!'.2~N_?_
~
I
'
,,
3-0 3-1 2-0 2-1 1-0 1-1 RATIO OF HOUSE FLIES TO 0. AENESCENS - --------
' 100
75 w (J z ~ 0:: 50 w :::< w <>11
25
0
21 "C
I I I I I I I
3-0 3-1 2-0 2-1 1-b 1-1
RATIO OF HOUSE FLIES TO 0....: ~E_!Jf~CJ:.!::I~-
Fig. 2 House fly emergence at 27, 21 and 18° C at different house fly to Ophyra aenescens ratios.
.!;--00
I

49
LITERATURE CITED
Anderson, J. R. & J. H. Poorbaugh. 1964a. Biological control possibility for house flies. Calif. Agric. 18: 2-4.
, Anderson, J. R. & J. H. Poorl>augh. 1964b. Observations on the ethology and
ecology of various diptera associated with northern California poultry ranches. J. Med. Entomol. 1: 131-147.
Hogsette, J. A., Jr. 1979. The evaluation of poultry pest management techniques . in Florida poultry houses. Ph.D dissertation U. of Fla. Gainesville, Fl 307
pp.
Johnson, W. T. & C. E. Venard. 1957. Observations on the biology and morphology. of Ophyra aenescens (Diptera: Muscidae). Ohio J. Sci. 57: 21-26. .
Miller, B. F., J. S. Teotia & T. 0. Thatcher. 1974. Digestion of poultry manure by Musca domestica. Br. Poultry Sci. 15: 231-234.
Nolan III, M. P. & J.B. Kissam. 1985. Ophyra aenescens: A potential bio-control alternative for house fly control in poultry houses. J. Agric. Entomol. 2: 192-195.
Peck, J. H. 1969. Arthropod predators of immature diptera developing in poultry droppings in northern California. Part II. Laboratory studies on feeding behavior and predation potential of selected species. J. Med. Entomol. 6: 168-171.
Peck, J. H. & J. R. Anderson. 1969. Arthropod predators of immature diptera developing in poultry droppings in northern California. Part I. Determination, seasonal abundance and natural cohabitation with prey. J. Med. Entomol. 6: 163-167.
SAS Institute. 1985. SAS user's guide: statistics, 5th ed. SAS Institute, Cary, N. C.
Stafford HI, K. C. & C. H.Collison .. 1987. Manure pit temperatures and relative humidity of Pennsylvania high rise poultry houses and their relationship to arthropod population development. Poultry Sci. 66: 1603-1611.

COMPLETE LIST OF REFERENCES
Abbott, W. S. 1925. A method for computing the effectiveness of an insecticide. J. Econ. Entomol. 18: 265-267.
Ables, J. R. &·M. Shepard. 1974. Responses and competition of the parasitoids Spalangi.a endius and Muscidifurax rap~or (Hymenoptera: Pteromalidae) at different densities of house fly pupae. Can. Entomol. 106: 825•830.
Ables, J. R., R. P. West & M. Shepard. 1975 .. Response of the house fly and its parasitoids to dimilin (TH•6040)~ J. Econ Entomol. 68:.622-624.
Adams, R. G. 1984. Ophyra species as predators in animal houses, with a key to species occurring in Europe (Diptera: Muscidae). Entomol. Gaz; 35: 243-246.
Allderson, J. F., A. Spandorf, L.A. Magnarelli & W. Glowa. 1986. Control of house flies in commercial poultry houses in Connecticut.~. Poultry Sci. 65: 837-844.
Anderson, J. R. & J.·H· Poorbaugh ... 1964a. Biological control possibility for house flies. Calif. Agric. 18: 2-4. ·
Anderson,J. R. & J. H. Poorbaugh. 1964b. Observations on the ethology and ecology of various diptera associated with northern California poultry ranches. J. Med. Entomol. 1: 131-147.
Axtell, R. C. 1963. Acarina occurring in domestic animal manure. Ann. Entomol. Soc. Am. 56: 628-633.
Axtell. R. C. 1964. Phoretic relationship of some common manure;. inhabiting Macrochelidae (Acarina: Mesostigmata)to the house fly. Ann. Entomol. Soc. Am. 57: 584-587.
Axtell, R. C. 1966. Comparative toxicities of insecticides to house fly larvae and Macrocheles muscaedomesticae, a mite predator of the house fly. J. Econ. Entomol. 59: 1128-1130.
Axtell, R. C. 1970. Fly control in caged-poultry houses: Comparison of larviciding and integrated control programs. J. Econ. Entomol. 63: 1734-1737.
Axtell, R. C. 1986a. Fly control in confined livestock and poultry production. Technical Monogragh CIBA-GEIGY Corp. 59 pp.
'·
Axtell, R. C. 1986b. Fly management in poultry production: cultural, biological, and chemical. Poultry Sci. 65: 657-667.
Axtell, R. C. & T. D. Edwards. 1983. Efficacy and nontarget effects of Larvadex™ as a feed additive for controlling house flies in caged;.layer poultry manure. Poultry Sci. 62: 2371-2377.
50
I i

I I
51
Axtell, R. C. & D. A. ~utz~ 1986. Role ~f par~i!~S and predators as biological fly control .agents m pou1!_IY pr~du,ction facilities, . SR-JOO I?P· In~- S Patterson and.D. A Rutz, eds. Biological Control of Muscmd Flies. Miscellaneous Publications 61, ED.tomologicalSociety of Ariierica, College Park, Md. ·
Bailey, D. L., D. W. Meifert,-& P. M. Bishop., 19QJ. Controlof l'louse flies in poultry houses with larvicides. Fla. E1~.tomol.. 51: 107-111.
Barth, c •• L~ · 1986.·· Fly control through manure management.. Poultry Sci. 65: 668-674. . . ·.·. . . . . . .
Beard,.R. L •. & D. C. Sands.1973 •. }?actors affecting degradation of poultry manure · byJlies. Environ. Entomol. 2: 801-806. · · · ·· · · · ·
Bills, G~ T. 1973. Biologicalflycontrol in.qeep-pit.poultryhouses .. Br~ Poultry Sci. 14: 209-212. .. . . . . .. .
Bloomcamp, C. L.,·R. S.Patterson.& P. G. Koehler. 1987. Cyromazine resistance in the house fly (piptera: Muscidae) .. ~· E~<m~Entomol. 80: 352~357.
Born, D. E .. 1954. Mold control fu fly rearing media. J. Econ .. Entomol. 47: 367.
BJ;eeden, G. C.,E. C.TurnerJr.·8'.W. L. Beane~.1975 •. fylethoprene.~a feed additive for control of the house fly breeding in chickeu manure. J. Econ E;ntomol. .68: 451-452. ·. · .··. . :.. . .. • ·•··.·. . : .· .
Breeden, G. c., E. c. Turner Jr~, w. :L. Beane, R. w. Miller & L. c. Pickens. 1980. The effect ofmethoprene asafeed.additiye on house fly emergence in pmtltry houses. Poultry.Sci. .. 60: ~56~562. · ·· . . · · . .. ·. ·
Burns, E. C., ~. H. Wilson lt B. A. Tower. 19(;1~ Effect of feeding Bacillus . .·. .· ... thuringi.ensisto caged layersfor f1y coD:trol. .J. ~con. Entomol. 54: 913-.915.
Card, L. E.& M.C. Neshei111. ~975.···· Poultry. Production. ·.Lea and Febiger,. · Philadelphia •. 392pp; . · · · · ·
Conway,~·.~ ··1973~. Thenli~ro-faunaof California-system poultry houses in · Bntam. Br. Poult. Sci .. 14: 213-216.. ·. .. .
Crampton; G~ C. 1944. A conip~atl~e morphological study of the terminalia of . . male calypterate cyclorrhaphous diptera ~dtheir acalypterate relatives.
Bull. Brooklyn Entomol. Soc;:. 39: l-31.
Derbeneva-Ukhova, V. P. 194i Surl'ecologie des mouclles de fumier a Kabarda . .• ··. ~ev. Appl. Entomol. Ser. B~ 31:124•12~. (~nglis;h,summary). ·.. .
Despins, J: L.,J. A. V~uglian & E. c. ·Tum~r, Jr. 1988.'iiole of the lesser · mealworm,Alphitobius diaperi.nus (Panzer) (Coleoptera: Tenebrionidae), as
a predator of the house fly, Musca domestica L. (D1ptera: Muscidae ), in poultry houses. Coleopts. Bull. 42: 211-216.

52
Geden, C. J., R. E. Stinner, & R. C. Axtell. 1988. Predation by predators of the house fly in poultry manure: Effects of predator density, feeding history, interspecific interference, and field conditions. Environ. Entomol. 17: 320-329.
Glaser, R. W. 1924. The effect of food on longevity and reproduction in flies. J. Exp. Zool. 38: 383-412. ,
Greenberg, B. 1971. Flies and disease. Vol. 1. Princeton Univ. Press, Princeton. 856pp.
Greenberg, B. 1973. Flies and disease. Vol. 2. Princeton Univ. Press,. Princeton. 447pp.
Hall, I. M. & K. Y. Arakawa. 1959. The susceptibility of the house fly, Musca domestica Linnaeus, to Bacillus thuringi.ensis var. thuringi.ensis Berliner. J. Insect Pathol. 1: 351-355.
Hall, R. D. & M. Foehse. 1980. Laboratory and field tests of CGA-72662 for control of the house fly and face fly in poultry, bovine, or swine manure. J. Econ. Entomol. 73: 564-569.
Hart, S. A. 1963;. Fowl Fecal Facts. World's Poultry Sci. J. 19: 262-272.
Harwood, R. F. & M. T. James. 1979. Entomology in human and animal health. Macmillan Pub. Co., Inc. New York. 548 pp.
Hill, G. D. & R. E. Williams. 1980. Seasonal abundance of flies and associated predators in egg-layer houses in Indiana. Poultry Sci. 59: 1620-1621.
Hogsette, J. A., Jr. 1979. The evaluation of poultry pest management techniques in Florida poultry houses. Ph.D dissertation U. of Fla. Gainesville, Fl 307 pp.
Johnson, W. T. & C. E. Venard. 1957. Observations on the biology and morphology of Ophyra aenescens (Diptera: Muscidae). Ohio J. Sci. 57: 21-26.
Kramer, J.P. 1980. The house fly mycosis caused by Entomophthora muscae: Influence of.relative humidity on infectivity and conidial germination. J. N. Y. Entomol. Soc. 88: 236-240.
Kramer, J. P & D. C. Steinkraus. 1987. Experimental induction of the mycosis caused by Entomophthora muscae in a population of house flies (Musca domestica) within a poultry building. J. N. Y. Entomol. Soc. 95: 114-117.
LaBrecque, G. C., D~ W. Meifert & D. E. Wiedhass. 1972. Dynamics of house fly and stable fly populations. Fla. Entomol. 55: 101-106.

I I.
53
Legner,E. F.,w~ R. Bowen;W~D. McKeeU:;W.·F. Rooney&R. F. Hobza.1973. Inverse relationships between mass ·of breeding.habitat and•.synanthropic fly emergence. and the measuremeht of'population densities with sticky tapes in California inland valleys. Environ. Entomol. 2: 199-205. ·
Legiler, E. F. & H. W;. Brydon. 1966.· Suppression of dung-inhabiting fly · . populations by pupal parasites~ .. Ann: ~ntomol. Soc; Am ... 59: 638-651.
Legner, E. F. & E. I. Dietrick. 1974. Effectiveness of supervised contl'ol practices in .·. ' ·. , lowering populati?n de11sities'of synanthropic fiies'on poultry' ranches.
Entomophaga 19; 479-482. . .. · · . ·. ,, .·· . .
Lindquist, A. W., W.W. Yates & R. A. Hoffman. 1951. Studies. of the flight habits · .·of three species of.flies tagged with radioactive phosphorus .. J. ·Econ; ..
EntomoL 44: 397-400~ · · · · ·. · · ·
Mackertas; M. J.1933 •.. 0bsetvations onthelife~histories;.nhtrional requirements and fecundity of blowflies. Bull. Entomol. ~es. 24: 353-362.
Mail, G; A. & H. F. Schoof •. 1954. Overwintering habits.of domestic flies at Charleston, West Virginia. Ann: Entomol. Soc.Amer. 47: 668.,.676.
Meyer; J. A., W. ri. McKeell. & B. A. Mullell.s. 1987. Fact~~s affecting control of Fannia SP.P· (Diptera: Muscidae) with cyromazine feed,.through on caged-
. layer facilities in southern California.: ·J~ Econ.EntomoL.80: 817•821. . . . •·. . -·· '
Meyer, J~ A., W. F~ Rooney & B. A. Mrillens •. · 1984~·.· Effect of l~~dex,feed-through on cool-season development of filth flies and beneficial coleoptera in poultry manure in southern California. Southwest. Entomol. 9: 52-56.
Miller, B~ R,J.S~·.Teotia &·T;O.Thatcher.1974. Digestion.ofpoultry manure by Musca domestica. Br. Poultry Sci. 15: 231-234.
Miller, R. W. 1970 •. XLli.rvieides for fly'.control•A review~· :BulLEntomoLSoc. Am. 16: 154•158. ' .. ' ..
Miller, R. W. 1974. TH 6040 as a feed additive for control of the face fly and house . fly; .J. Econ Entcnnot·67: 697 .. ·. .; · .. · .· .' •. • ·· · ·· . ·
,' - ' ' . Miller, R. W., C. Corley & K. R. Hill. 1975. Feeding TH 6040 to chickens: Effect
on larval house flies in manure and determination of residues in eggs. J. EcOn. ED.tomOl:~'.68:.181~182·. " .. ,~ · ·· ! • •• • •
Morgan, P~ B., G. C. LaBrecque, D •. E. Weidhaas & A. Benton. 1975. The effect of . ··'••·. methoprene, an insect growth'regulator; on Musca domestica.(Diptera:
Muscidae). Can.Entomol.)07:413-417. ·.. . . > ...
. . ~.
r r
.{

54
Morgan, P. B. & R. S. Patterson. 197 5. Field parasitization of house flies by ·. . natural populations. of Pathycrepoideus vindemiae (Rondani), Muscidifurax
raptor Girault·andSanders,.3.ndSpalangi,anigroaeneaCurtis .. Fla. Entomol. 58:202.
Mulla, M. S. & H. Axelrod. 1983a. Evaluation of Larvadex, a new IGR for the controLofpestiferousflies on poriltryranches~ J.•EconEntomol. · 76:520• 524. ·.. . ' .. · . . ' .·. . ' .· - ·-.
Mulla, M. S. & H. AXelrod. 1983b. Evaluation of the IGR Larvadex™ as a feed-- · · through treatment for the control of pestiferous.flies on: poultry ranches; J.
Econ. Entomol. 76: 515-519; ,. . - .. ·· ··
Mull~r,P. 1982. The importance of the Musca domestica natural enemyOphyra· aenescens (Diptera: Muscidae):: III~ ;Laboratory experiments on the interaction between the larvae of Musca domestica and Ophyra aenescens. Angew. Parasitol. 23: 143-154. (in German with English abstract).
Muller, P.; H. Schumann, P. Betke,· H. Schultka; R. Ribbeck, & T. Hiepe. 1981. The importance of the Musca domestica antagonist Ophyra aenescens (Diptera: Muscidae ). I. The occurrence of Ophyra aenescens in animal production fustallations; Angew; Parasitol. 22: 212-216. >(in German with· English abstract) .. ~·· · ·· · · .. · . ·•· •· •· · . . _ . • · ·· · .
. . Nolan III, M. P. 1987. ·Development of an integrated pest management program · · for the control of the house fly Musca domesticaL~, in caged layer poultry
housesinSouth Carolina;· Ph.D disserta.tionCle:i:nson Univ. Clemson;S. C. l79pp·· . . . ,
Nolan III, M. P~ & J. B. Kissam. 1985. Ophyra aenescens: A potential bio•control alternative for house fly control in poultry houses; LAgric. Entomol. 2: 192-195. . . .
NolanIII; M~ P~.&J. B. Kissam~·1987. Nuisance potential.of a dump fly, Ophyra aenescens (Diptera: Muscidae ), breeding at poultry farms. Environ. ·. EntomoL 16: 828-831. · · · · · ·
Olckers, T .• ·.& P. E: Hulley •. 1984. Facultative predation.of house fly larvae by larv:ae of Ophyra capensiS. (Wiedemann) (Diptera: Muscidae ); J; Entomol. Sth. Afr. 47: 231-237. · · . .
Oldroyd; H. 1965. The nati:rral history offlies. W. W; Norton and Co;, Inc: New York ... :3z4·pp. ·. , .. , · · ·
Olton, G. S.-& .E. F~ Legner. 1975. Winter inoculative releases of parasitoids to reduce house flies in poultry manure. J. Econ. Entomol. 68: 35-38.
- > ' • -. ,- • ·- ~ -~¥· • -~ .~' -
Peck, J. H.' 1969. kthropod predators of immature diptera developing in poultry droppings in northern California. Part II .. Laboratory studies on feeding behavior and predation potential of selected species. J. Med. EntomoL 6: 168~171.

55
Peck, J. H. & J. R. Anderson. 1969. Arthropod predators of immature diptera developing in poultry droppings in northern California. Part I. Determination, seasonal abundance and natural cohabitation with prey. J. Med. Entomol. 6: .163-167.
Peck, J. H. & J. R. Anderson. 1970. Influence of poultry manure removal schedules on various diptera larvae and selected arthropod predators. J. Econ. Entomol. 63: 82-90.
Pfeiffer, D. G. & R. C. Axtell. 1980. Coleoptera of poultry inanure in caged-layer houses in North Carolina. Environ. Entomol. 9: 21-28.
Propp, G. D. & P. B. Morgan. 1985. Mortality of eggs and first-stage larvae of the house fly, Musca domestica L. (Diptera: Muscidae ), in poultry manure. J. Kansas Entomol. Soc. 58: 442-447.
Quisenberry, S.S. & D. E. Foster. 1984. Cost-benefit evaluation of house fly (Diptera: Muscidae) control in caged layer poultry houses. Poultry Sci. 63: 2132-2139.
Ribbeck, R., P. Betke, P. Muller, H. Schumann & T. Hiepe. 1987. Damage by and co~trol of flies in intensive livestock production. Monatshefte fur Veterinarmedizin. 42: 517-521. (in German with English abstract).
Robertson, S. H. & D. P. Sanders. 1979. Species composition and seasonal distribution of the dipterous fauna inhabiting swine confinement housing in west Texas. Southwest. Entomol. 4: 89-95.
Roddy, L. R. 1955. A morphological study of the respiratory horns associated with the puparia of some diptera, especially Ophyra aenescens (Wied.). Ann. Entomol. Soc. Am. 48: 407-415.
Rueda, L. M. & R. C. Axtell. 1985. Comparison of hymenopterous parasites of house fly, Musca domestica (Diptera: Muscidae ), pupae in different livestock and poultry production systems. Environ. Entomol. 14: 217-222.
Rutz, D. A. & R. C. Axtell. 1980. House fly (Musca domestica) parasites (Hymenoptera: Pteromalidae) associated with poultry manure in North Carolina. Environ. Entomol. 9: 175-180.
Rutz, D. A. & R. C. Axtell. 1981. House fly (Musca domestica) control in broiler-breeder poultry houses by pupal parasites (Hymenoptera: Pteromalidae ): Indigenous parasites species and releases of Muscidifurax raptor. Environ. Entomol. 10: 343-345.
SAS Institute. 1985. SAS user's guide: statistics, 5th ed. SAS Institute, Cary, N. C.-,
I s

56
Schumann, H. 1982. The importance of the Musca domestica natural enemy Ophyra aenescens (Di_ptera: Muscidae ). II. Morphology of the development stages. Angew. Paras1tol. 23: 86-92. (in German with English abstract).
Shepard, M. & J.B. Kissam. 1981. Integrated control of house flies on poultry farms: Treatment of house fly resting surfaces with diflubenzuron plus releases of the parasitoid, Muscid~rax raptor. J. Ga Entmol. Soc. 16: 222-227. ,
Skidmore, P. 1985. The biology of the Muscidae of the world. Klower Academic Publ., Wingham, MA. 550 pp.
Stafford III, K. C. & C.H. Collison. 1987. Manure pit temperatures and relative humidity of Pennsylvania high-rise poultry houses and their relationship to arthropod population development. Poultry Sci. 66: 1603-1611.
Turner Jr., E. C. & J. A. Hair. 1967. Effect of diet on longevity and fecundity of laboratory-reared face flies. J. Econ. Entomol. 60: 857-860.
West, L. S. 1951. The housefly. Comstock Publ. Co., Ithaca. 584 pp.
Williams, R. E. & J. G. Berry_. 1980. Evaluation of CGA 72662 as a topical spray and feed additive for controlling house flies breeding in chicken manure. Poultry Sci. 59: 2207-2212.

The vita has been removed from the scanned document