review of literature 20 - inflibnetshodhganga.inflibnet.ac.in/bitstream/10603/89379/8/08_chapter...
TRANSCRIPT

Review of Literature 20
2.1 Textile Industry in Rajasthan The industrial development of Rajasthan began between 1950 and 1960.
Large and small scale industries sprung up in the Kota, Jaipur, Udaipur, Bhilwara and
other industrial estates of Rajasthan. The key industries of Rajasthan include textile,
rugs, woolen goods, vegetable oil and dyes. Heavy industries include copper and zinc
smelting and the manufacture of railway rolling stock. The private sector industries
include steel, cement, ceramics and glass wares, electronic, leather and footwear,
stone and chemical industries. Textile sector holds for about 20% accountability of
investment made in the state. Rajasthan contributes over 7.5 percent of India's
production of cotton and blended yarn (235,000 tonnes in 2002-03) and over 5
percent of fabrics (60 million sq meters). The state holds a leading position in
spinning of polyester viscose yarn & synthetic suiting (at Bhilwara) and processing,
printing & dyeing of low cost, low weight fabric (at Pali, Balotra, Sanganer and
Bagru). In total the production of textiles in Rajasthan accounts for 21.96 % of the
total national production scenario. Large quantities of spun yarn and hence is the
fourth largest producer in India (http://www.mapsofindia.com/maps/rajasthan/
rajasthanindustry.htm). Besides, Jaipur is also a well-known center for manufacturing
garments primarily for exports. Some knitting units are in process of setting up their
ventures at Neemrana. The economic growth, the state of Rajasthan has witnessed
with the textile sector is depicted in Figure 9.
Figure 9: Industrial Export from Rajasthan (Rs. In crore) with special emphasis to textile industry (Source: Commissioner, Department of Industry; Rajasthan) (State of Environment report for Rajasthan: 2007)
0
200
400
600
800
1000
1200
1400
1600
1999-20002000-20012001-20022002-20032003-2004
Rs.
(in
Cro
res)
Annual industrial Export of diffrent industries
Textile
Gems and Jewellery
Engineering
Marble and Granite
Electrical and Electronics
Wool and Woolens
Chemical and Allied
Drugs and Pharmaceuticals
Plastics and Linoleum
Handicrafts
Handloom

Review of Literature 21
The figure above clearly indicates the significance of textile industry in
Rajasthan. Keeping the fact in mind, that a coin always has two facets; likewise,
textile industry is has proven to be a boon and a bane for societal development. On a
positive front, it is a growing sector with respect to economic growth of Rajasthan
and on the negative side; the industrial growth is always been associated with
augmentation in environmental pollution particularly related to inland water, air,
noise and soil; if not stringently monitored. The industries in the state have been
categorized into red and orange depending upon the environmental pollution:-
•••• Highly polluting industries have been classified as red category which
includes man made fiber manufacturing (Rayon & Others). Yarn and textile
processing industries which involve scouring, bleaching, dyeing, printing or
any effluent / and process that relate to generation of emission.
•••• The moderately polluting industries are classified as orange category and
include yarn and textile manufacturing /processing not involving scouring,
bleaching, dyeing, printing or any effluent /emission generating process
including spinning/weaving unit.
As per the categorization of Rajasthan State Pollution Control Board
(RSPCB), textile units have been included both in red and orange categories, so as to
cover these entire units under the pollution control system (RSPCB office order No.
F.14 (57) Policy / RSPCB/ Plg/ 9219 – 9259 dated 21.12.2010).
Worth mentioning is the fact that textile industry occupies the top notched
position in contributing to the hazardous wastes amongst all industries (State of
Environment Report for Rajasthan, 2007). Table 5 represents the sector specific
generation of hazardous waste by industrial units.

Review of Literature 22
Table 5: Sector – Specific Number of Hazardous Waste Generating Industries
S.No Product Number of Units SS/Med* LS** Total
1 Asbestos and it’s products - 16 16
2 Benefiaciation of ore 1 - 1
3 Cement 14 - 14
4 Ceramic 2 - 2
5 Chemical 23 36 59
6 CETP’s 3 1 4
7 Drug and Pharmaceuticals 8 23 31
8 Dye and it’s intermediates 3 33 36
9 Electronics 2 1 3
10 Engineering 41 41 82
11 Fertilizer 14 1 15
12 Mining 6 2 8
13 Paints, Varnish and ink 2 4 6
14 Pesticides 2 36 38
15 Petroleum Drilling/storage 19 - 19
16 Power plants 3 - 3
17 Primary production of copper/zinc /lead 3 - 3
18 Secondary production of copper/zinc 1 13 14
19 Secondary production of zinc /lead - 32 32
20 Steel rolling mills 2 21 23
21 Tanneries - 9 9
22 Textile 70 19 89
23 Thermal power 2 - 2
24 Waste oil refineries 3 30 33
25 Workshop 2 - 2
26 Miscellaneous 1 1 2 *Small/ Medium Scale **Large Scale (Source - RSPCB Hazardous Waste Report, 2007) Evidently, textile industries are major sources of environmental pollution. As
the textile industries consume large quantities of water and generate waste water in
proportionate order. Pali has always been into limelight as one of the major textile
hubs of Rajasthan which proportionately generates huge volumes of waste water.

Review of Literature 23
It houses around 800 textile units, of these, 200 fall under orange category
and the remaining 600 under the red category, of which 75-80 are non- operational
(Centre for Science and Excellence (CSE), September, 2014). In Rajasthan, the semi
arid and arid belts rely on groundwater resources due to paucity of surface waters
(Bhadra et al. 2013). The deterioration of surface and groundwaters due to
industrial and urban waste has long been recognized (Olayinka, 2004). The
rivers and stream are common sites for disposal of industrial effluents. The textile
dyeing and printing units situated at Pali have been discharging effluents in the
tributary Bandi which owes it’s origin to Luni basin (Khandelwal and Chauhan,
2005). The untreated industrial effluent flows regularly in the dry bed of river about
45 km downstream Pali city (Rathore, 2011). Particularly, the release of effluents
influxed with dye into the water environment is unacceptable because of their colour,
direct release and end products are toxic and may be carcinogenic or mutagenic to life
forms because of benzidine, naphthalene and other aromatic compounds (Suteu et al.
2009; Zaharia et al. 2011).
The groundwater pollution caused by the discharge of untreated and partially
treated textile effluents into river Bandi has been an issue of utmost concern which
awaits immediate and justified addressal. Recent study conducted by The Centre for
Science and Environment (CSE) on pollution caused by by textile mills in Pali was
of the opinion that none of the samples like surface waters, effluents from different
mills assessed for physico-chemical properties were in compliance with the standards
prescribed by Central Pollution Control Board (CPCB) (Times of India, dated
19.09.2014) The depletion in quality of water has an undue effect on human beings
as well as aquatic ecosystem directly or indirectly (Chinda et al. 2004; Ugochukwu,
2004; Emongor et al. 2005).
National Green Tribunal (NGT) had strictly ordered 550 industrial units to
shut down on account of water pollution caused by textile industries in Pali in March,
2014 (Rajasthan Patrika, dated 03/ 06/ 2014).
A study conducted by Geological Survey India (GSI) in January, 2014 was
aimed to assess the nature of discharging effluents from textile industries and it’s

Review of Literature 24
effect on natural resources in their vicinity. This study remarkably revealed that the
effluents had caused irreparable damage to agriculture and groundwater significantly
(Times of India, dated 2.1.2014).
Nevertheless, an initiative of Times of India (TOI) and RSPCB led to the
closure of all textile units in Pali town on account of excessive effluent generation
and it being drained directly into river Bandi (November, 2012). Previously, a similar
initiative taken by Central Groundwater Board (CGB) which was taglined as “In
Pali, a river being killed” was of opinion that due to excessive release of effluent; 50
Million litres per Day (MLD) against the permissible which is 36 MLD, resulted into
alkalinity of groundwater and hence the wells were rendered unfit for drinking
purposes. (September, 2012). A brief account of textile units in Pali city is given in
Table 6.
Table 6: Distribution of Textile Industries in diff erent localities of Pali city
(Rathore, 2012)
S.No Location Number of Units
1 Mandia Road Industrial Area Phase III 473
2 Industrial Area I and II 121
3 Main Mandia Road, Ramdev Nagar, Residential Area, Gandhi Nagar
173
4 Sumerpur Road, Bajrang Bari, Ramleela Maidan, Sojat Road, Village Mandia
95
5 Composite Unit- Maharaja Shri Umaid Mills Ltd., Pali, Marwar
1
6 Total 860
The tools like Geographic Information System (GIS) and Remote Sensing had
been used to assess groundwater pollution due to industrial effluents from textile
units around Pali city. Indiscriminate discharge of these effluents in the past two
decades caused severe damage to the agricultural land and groundwater resources
within 3 km buffer zone of the river. Remote Sensing and Geographic Information
System (GIS) techniques are found to be better tools in assessing the damage in

Review of Literature 25
agricultural crops during 1997 and 2006 with the help of high resolution satellite
images (Bhadra et al. 2013).
2.2 Characteristics of textile effluents and CETP an approach to combat
water pollution
Effluent discharged from the textile industries has variable characteristics in
terms of pH, dissolved oxygen, organic and inorganic chemical content. Together
with industrialization, awareness towards the environmental problems arising due to
effluent discharge is of critical importance. Pollution caused by dye effluent is
mainly due to durability of the dyes in wastewater (Jadhav et al. 2007).
Textile industries are large industrial consumers of waters as well as
producers of wastewaters and contribute to a total of 14% of industrial pollution in
India (business.mapsofindia.com).
This industry involves processing or conversion of raw material/fabric into
finished cloth involving various processes/ operations and consumes large quantities
of water and produces extremely polluted waste effluents (Asia et al. 2006; Andre et
al. 2007). The effluent is primarily composed of extremely diversified inorganic
compounds, polymers and organic substances (Mishra and Tripathy, 1993) in
dissolved, colloidal or suspended forms and is typically coloured due to the presence
of residual dyestuffs (Patel and Pandey, 2008). The effluent is characterized by a
high BOD (Wang et al. 2007; Annuar et al. 2009) which depletes the dissolved
oxygen; both of which are aesthetically and environmentally unacceptable. The
release of these highly coloured effluents can damage directly the receiving waters
(Chen et al. 2003). Because of the stable structure of synthetic dyes used in textile
industries, the retention of colour remains a permanent feature and is not removed by
municipal waste water systems (Shaul et al. 1991; Robinson et al. 2002; Forgacs et
al. 2004; Couto, 2009). It is estimated that globally 280 000 tons of textile dyes are
discharged in textile industrial effluent every year (Jin et al. 2007).
The risk factors for pollution are primarily associated with the different
production steps in which wet processes include scouring, desizing, mercerizing,

Review of Literature 26
bleaching, dyeing and finishing. Desizing, scouring and bleaching processes produce
large quantities of wastewater (Yusuff et al. 2004) and is characterized by high pH,
temperature, detergents, oil, suspended and dissolved solids, dispersants, leveling
agents, toxic and non-biodegradable matter, colour and alkalinity (Nese et al. 2007;
Govindwar et al. 2012; Sharma et al. 2013). Important pollutants in textile effluent
are mainly recalcitrant organics, colour, toxicants and surfactants, chlorinated
compounds. Extreme fluctuations in COD, BOD, pH, colour and salinity of textile
effluents are reported which may be attributed to 8000 different types of chemicals
like salts, metals, surfactants, organic processing assistants, sulphide and
formaldehyde (Zollinger, 1987; Talarposhti et al. 2001; Kanu and Achi, 2011;
Jamaluddin and Nizamuddin, 2012; Sharma et al. 2014). In cases most of the
BOD/COD ratios are found to be around 1:4, indicating the presence of non-
biodegradable substances (Arya and Kohli, 2009). The process of adding colour to
the fibres is known as dyeing which normally requires large volumes of water not
only in the dye bath, but also during the rinsing step. The water let out after the
production of textiles contains a large amount of dyes and other chemicals which are
harmful to the environment. The level of toxicity or harmfulness of the textile
effluents varies among industries (Kiriakidou, 1999). The effluent generated during
the process of rinsing and in dye bath (Dos Santos, 2005) always contains a portion
of unfixed dye (1mg/l) (Gonclaves, 2000) which imparts strong colour to the effluent
(Soares et al. 2006). Rathore et al. 2011 was of opinion that, the colour of textile
effluents is usually subject to daily and seasonal variations. The routine of production
and the other characteristics vary according to the nature of textile manufactured.
The concentration of these unfixed dyes and trace metals like Cr, As, Cu and Zn was
found to be high in textile wastewater (Eswaramoorthi et al. 2008). The presence of
these heavy metals affects the aesthetic value, transparency of water and solubility of
gases in lakes, rivers and other water bodies (Agarval, 2001). Dyes in water give out
a bad colour and can cause diseases like haemorrhage, ulceration of skin, nausea,
severe of skin irritation and dermatitis (Nese et al. 2007). The penetration of sunlight
can be blocked from water surface preventing photosynthesis and increased BOD
which reduces the process of re oxygenation and growth of photoautotrophic

Review of Literature 27
organisms is hampered by and large (Laxman, 2009). Figure 10 represents schematic
representation of unit processes in a textile industry.
Figure 10: Schematic representation of unit processes in a textile industry
Sharma et al. in 2014 explored one of the famous textile hotspots of Jaipur
region, Bagru. This area is internationally famous for bagru prints. The study was
conducted in months of July, August and September 2013 to assess the quality of
water resources affected by textile effluents in terms of pH, electrical conductivity,
total dissolved solids, total suspended solids, dissolved oxygen, chemical oxygen
demand and chloride. The study was found to have non adherence of effluent
standards for discharge of textile effluent prescribed by RSPCB. Thoker et al. in
2012; Jamaluddin and Nizamuddin in 2012, had conducted similar studies to assess
the quality of textile effluents in Chittagong, Bangladesh.
Gray cloth
Settling
Bleaching and washing
Dyeing and printing
Steaming
Washing
Finishing
Folding

Review of Literature 28
Rathore in 2011 and 2012 conducted a study to assess the quality of river
Bandi with special emphasis to parameters like COD, suspended solids, chloride,
sulphate, and chromium and compared with standards laid down by Bureau of Indian
Standards (BIS). A significant reduction in pollution load of river Bandi during
monsoon season was observed which may be attributed to flow of freshwater
upstream of Pali city.
Joshi and Santani in 2013 examined the physicochemical properties like
temperature, pH, colour, odour, COD, BOD, EC, TSS, TDS and inorganic elements
like sodium, potassium and phosphorus levels of effluent discharged by Sumukh
textile mill, Vapi, Gujarat, India.
Textile industry is not only confined to northern and western parts of India,
Tamil Nadu has also emerged as a potential state for the same. The textile mills are
concentrated in Coimbatore, Tirupur, Salem, Palladam, Karur and Erode. Tamil
Nadu has around 3, 50,000 power looms manufacturing cotton fabrics and accounts
for about 30 per cent of India's export of textile products. The Erode district in Tamil
Nadu is well known for marketing of textile products of handloom, powerloom and
readymade garments (Chellasamy and Karuppiah, 2005).
Textile effluents generated by textile dyeing industries of Tirupur and
Coimbatore, Tamil Nadu were thoroughly studied for their physicochemical
parameters. The values of each parameter were found to be well above the laid
standards (Rao and Prasad, 2011; Rajeswari et al. 2013a).
Continuous discharge of untreated textile effluent into Noyyal river basin has
deteriorated the quality of water and has rendered it unfit for irrigational purposes
(Mahalakshmi and Saranathan, 2014). Sundar and Saseedharan in 2006 and 2008
investigated the groundwater quality in Noyyal river basin. The result demonstrated
that due to industrialization, high level of dyeing effluents in and around the river is
responsible for deterioration of groundwater quality (Palanysamy et al. 2008). The
hydro-geochemistry analysis for groundwater of the river basin has been examined to
characterize the groundwater (Sunder and Saseedharan, 2008). Usharani et al. 2010

Review of Literature 29
has also analyzed the physico-chemical and bacteriological characteristics of Noyyal
river and it’s groundwater quality. Santhosh and Devi in 2010 have integrated land
use and land cover detections using remote sensing and GIS for Coimbatore district.
Zahiruddin and Kandasamy, 2013 studied the germination profile of Cajanus cajan
(Red gram), Phaseolus aureus, (Green gram) and Vigna mungo (Black gram) from
the family Fabaceae. When irrigated by textile effluent taken in varying
concentration from three different textile mills of Kanchipuram, Tamil Nadu, India,
significant differences in germination profile were observed.
Studies pertaining to toxicity of textile effluents on soil microflora have been
reported (Kaur et al. 2010). The parameters like germination, delay index,
physiological growth and plant pigments are affected because utilization of textile
effluent for irrigation purposes is rendered unfit (Garg and Kaushik, 2008). In India
the untreated sewage water and wastewater from textile industries containing variety
of chemicals such as aniline, caustic soda, acids, bleaching powder including heavy
metals, are used in irrigating agricultural fields, for growing vegetables and other
crop plants owing to paucity of water resources (Chandra et al. 2009). The effluent
water takes the dissolved toxicants to crop plants and it’s consumers (Kala and Khan,
2009). Vegetables grown in the agricultural fields using untreated textile wastewater
for irrigation are adversely affected. Use of wastewater alters the nutritional value of
the vegetables grown there and in long run consumption of such vegetables may
impose health hazards in human beings, which is a matter of concern. Irrigating
crops with wastewater containing high nutrient load often leads to prolific weed
growth and undesirable growth of microorganisms (Modasiya et al. 2013). Debnath
et al. 2014 studied the effect of untreated textile effluent generated by textile
industries of Sanganer, Jaipur on irrigation of crops. A similar study conducted on
untreated textile effluent in Sanganer was aimed to assess the bioaccumulation of
different heavy metals by Lycopersicon esculentum (wild tomato) when a varying
concentration of effluent was used for irrigation purposes (Khan and Marwari, 2010).
On the contrary, it has also been reported that waste water reuse for
irrigational purposes is a strategic approach for water pollution abatement ( Bhabindra,
2003) and certain components like nitrogen and phosphorus (Zahiruddin et al. 2013)

Review of Literature 30
in textile effluent when present in lower concentration are beneficial for the plant
growth (Kannan and Upreti, 2008) .
Considering, water pollution caused by textile industries as a global concern,
the physico-chemical properties of textile effluents have been envisaged in different
parts of the world.
Imtiazuddin et al. in 2012 conducted an extensive study for a period of four
months i.e. November, December, January and February in 2009-2010. The samples
were collected from different textile mills in Pakistan and analyzed for various
parameters such as TDS, COD, BOD, pH, EC and heavy metals like cadmium,
chromium, copper, iron, manganese, nickel, potassium, phosphorous, sodium,
sulphur, zinc.
The wastewater treatment plants of 11 textile mills (Turkey) for woven fabric
and knit fabric finishing industry were investigated (Nese et al.2007) and treatment
plants were evaluated for their performance by in situ inspections and analyses of
influent and effluent samples. The analysis of effluent indicated non-adherance with
the IWSA (Istanbul Water and Sewerage Administration) discharge standards.
Keeping the problem of water pollution caused by textile industries as a
major target, CETP had been proved to be a remedial alternative to minimize the
excessive problem caused.
Forty percent of the industrial wastewater generated in India comes from
small size industries. With the adoption of the Water Act (1990), small size industries
should treat their effluent in order to limit pollution concentration at the minimum
acceptable standards laid down by the State Pollution Control Boards (SPCB).
Nevertheless, the size of these facilities makes the installation of a standard Effluent
Treatment Plant (ETP) unaffordable (Singh et al. 2011). The onset of CETP’s was
the pioneering effort of Ministry of Environment and Forests (MoEF) aimed at
treating effluents generated by different small scale industries; to devise remedial
measures to which may result into generation of an eco friendly effluent. The first
CETP came into existence in 1985 in Jedtimetla near Hyderabad, Andhra Pradesh to

Review of Literature 31
treat waste waters generated from pharmaceutical and chemical industries
(Maheswari and Dubey, 2000).
The concept of CETP has four underlying objectives given in Figure 11.
Figure 11: Objectives determining the viability of a CETP
It has been clearly shown, that compared to individual ETP’S, CETP are
more cost effective in reaching the effluent concentration standards (Pandey and
Deb, 1998). Taking this into consideration, the massive problem of water pollution
caused by textile industries in Pali, CETP’s have been installed with the aim to
pollution abatement (Dhariwal and Ahmed, 2009). A brief overview of different
CETP’s in Pali has been reported earlier (RSPCB, 2007) (Table 7).
Table 7: Different CETP’s installed for treating textile effluents in Pali city
(RSPCB, 2007)
S.No Year Capacity Purpose Present
Status Cost Performance Problems Units
PWPCTRF
Unit 1 1983 5MLD Textile Operational 38
Lacs Non-compliance
with standards
Needs up gradation
435 in all
Unit II 1997 8.4MLD Textile Operational
Unit III 1999 9MLD Textile Operational 7.5
Crores
To achieve economics of scale in waste to minimise pollution cost
To circumvent the problem of space as CETP is a centralised facility
To reduce the problems of monitoring for the pollution control boards.
To organize the disposal of treated wastes and sludge and to improve the recycling and reuse possibilities.

Review of Literature 32
Assessment of CETP at regular intervals for inlet and outlet parameters has
been an important aspect to be explicitly considered. This justifies the functioning of
CETP but also correlates the significant reduction in pollution load after treatment
(Kore and Kore, 2012). The CETP located in Kagal Five Star MIDC, Kagal Nagpur
was studied for both inlet and outlet parameters like pH, TSS, TDS, oil and grease,
COD and BOD.
Desai and Kore in 2011 conducted a study to evaluate the performance
efficiency of an ETP of a textile industry located in Kagal-Hatkanangale
(Maharashtra Industrial Development Corporation) MIDC area, Kolhapur
(Maharashtra). Both inlet and outlet parameters like pH, BOD, COD and TDS were
taken into account.
Patel et al in 2013 undertook a study to evaluate efficiency of an ETP of a
textile industry located in Narol, Ahmedabad (Gujarat). The parameters such as pH,
BOD, COD, TDS, TSS and ammonical nitrogen (NH3-N) were considered for both
inlet and outlet samples.
Rajeswari et al. in 2013b analysed and compared the efficiency of three
textile focused CETP’s located at Telugupalayam, Tamil Nadu for it’s outlet
parameters like temperature, pH, colour, odour , EC, TDS , TDS, COD, BOD, total
hardness, total alkalinity, chlorides and sulphates.
Singh et al. in 2011 attempted to evaluate the performance of CETP’s in Pali
based on criteria like design, operation, maintenance and administration and
contrasted the outcomes with the standards prescribed by Rajasthan State Pollution
Control Board (RSPCB).
Performance of CETP, Mandia Road, Pali was evaluated not only for it’s
inlet and outlet parameters but all stages of treatment like primary clariflocculator,
aeration tank and secondary clarifier were analysed for physicochemical parameters
like pH, BOD, COD, TSS for a period of 4 months on a regular basis (Dhariwal and
Ahmad, 2009).

Review of Literature 33
Along with the liquid effluent, sludge, a byproduct of effluent treatment can
create a menace for environment, because it contains many toxic chemicals released
by textile industries. Therefore, it requires proper and safe disposal (Patel and
Pandey, 2008).
Patel and Pandey in 2008, identified 4 CETP’s catering to textile industries
Mandia Road, Pali; Balotra, Barmer; Manikapuram and Mannarai in Tirupur, Tamil
Nadu and thoroughly investigated physico-chemical properties and heavy metals
present in sludge generated by textile industries.
A continuous growth in the demand of water in agricultural, industrial,
municipal and recreational sectors had been witnessed throughout the country. Due
to paucity of surface water, in semi arid and arid regions, there is a dependency on
groundwater resources to a great extent. In many areas in Rajasthan, alarming trends
of groundwater contamination have been reported due to discharge of untreated
industrial effluents (Rathore, 2012; Khan et al. 2014). Since 1980s, Pali city area is
facing severe problem of pollution due to discharge from wide spread dyeing
industry into the Bandi river and industries here discharge a variety of chemicals,
dyes, acids and alkalis besides heavy metals and other toxic compounds. Due to the
non-biodegradable nature and longtime persistence in the environment, the toxic
waste often accumulates through trophic levels causing a deleterious biological effect
(Bhadra et al. 2013). It has been mentioned in literature that synthetic dyes are the
major culprit’s which not only pollute the environment but are difficult to treat owing
to their recalcitrant nature (Pereira and Alves, 2012).
2.3 Synthetic Dyes: The main culprits
Colourants (dyes and pigments) are important industrial chemicals.
According to the technological nomenclature, pigments fall into the category of
colourants which are insoluble in the medium to which they are added, whereas dyes
are soluble in the medium. The world’s first commercially successful synthetic dye,
named mauveine, was discovered accidently in 1856 by William Henry Perkin. Such
substances known as colourants with considerable colouring capacity are widely
employed in the textile, pharmaceutical, food, cosmetics, plastics, photographic and

Review of Literature 34
paper industries (Carneiro et al. 2007). Dyes are classified according to their
application and chemical structure and are composed of a group of atoms known as
chromophores, responsible for imparting colour to the dye. This fact is attributed to
(1) absorption of light in the visible spectrum (400–700 nm), (2) presence of atleast
one chromophore (colour-bearing group) or the colour enhancer (Pereira and Alves,
2012), (3) have a conjugated structure of alternating double and single bonds, and (4)
electrons, which possess resonance and stabilize organic compounds (Abrahart,
1977). Exclusion of any of these factors leads to loss of colour. These chromophore-
containing centers are based on diverse functional groups, such as azo,
anthraquinone, methine, nitro, arylmethane, carbonyl and others. In addition,
electrons withdrawing or donating substituents so as to generate or intensify the
colour of the chromophores are denominated as auxochromes. The most common
auxochromes are amine, carboxyl, sulfonate and hydroxyl (Christie, 2001; Dos
Santos et al. 2007; Prasad and Rao, 2010).
The colour imparted to fibers is permanent, such that the colour is not lost
(wash fast) when exposed to sweat and light (light fast), water and many chemical
substances including oxidizing agents and also to microbial attack (Rai et al. 2005;
Saratele et al. 2011). Textile effluents primarily consist of synthetic dyes as
colourants (Pereira and Alves, 2012) that are amongst the major priority pollutants
that accumulate in environment as recalcitrant xenobiotic compounds due to
continuous industrial inputs that have created a serious impact on environment.
Synthetic dyes in particular are stable, toxic organic compounds (Hao et al.
2001; Hu et al. 2009) which are mostly carcinogenic like benzidine and other
aromatic compounds (Przytas et al. 2012) mutagenic (Weisburger, 2002; Mathur et
al. 2007) and pose serious health hazards which may remain persistent for longer
durations (Forgacs et al. 2004; Shedbalkar et al. 2008; Sinha et al. 2009; Kumar and
Praveen, 2011). The end users are the textile industries and account for about two-
thirds of the total dye market (Elisangela et al. 2009) and to reciprocate, generate
enormous volume of aqueous wastes (Sen and Demirer, 2003; Dos Santos, 2005;
Ben Mansour et al. 2012). The dye effluents are discharged from the dyeing process
as some of dye remains unfixed to the fibres (Wong and Yu, 1999; Fang et al. 2004;

Review of Literature 35
Lavanya et al. 2014) and is therefore visually identifiable (Kilic et al. 2007) and in
some cases concentration as low as 1 mg/ml is also detectable (Pandey et al. 2007;
Wijetunga et al. 2010).
A need to establish an economic and effective way of dealing with the textile
dyeing waste in response to the production costs of different unit operations is
necessitated (Park et al. 2007). The Ecological and Toxicological Association of the
Dyestuffs Manufacturing Industry (ETAD) was inaugurated in 1974 (Chequer et al.
2011) with the goals of minimizing environmental damage, protecting users and
consumers and cooperating with government and public concerns in relation to the
toxicological impact of their products (Anliker, 1979). Government legislation is
becoming more and more stringent, especially in more developed countries,
regarding the removal of dyes from industrial effluents (Robinson et al. 2001; Kuhad
et al. 2004; Ogubue and Sawidis, 2011) and hence the removal of bioaccumulated
xenobiotic dyes from textile effluents is a matter of international concern (Gienfrada
and Rao, 2008).
Dyes are usually classified by their Colour Index (CI), developed by the
Society of Dyes and Colourist (1984). Accordingly, dyes are firstly listed by a
generic name based on it’s application to fibre and it’s colour, followed by assigning
a 5-digit CI number based on it’s chemical structure, if known (O'Neill et al.1999).
Examples include Acid Blue 120 (26400), Reactive Red 4 (18105), and Mordant
Yellow 10 (1401). They can be grouped in different classes: acid, basic, direct,
disperse, metallic, mordant, pigment, reactive, solvent, sulphur and vat dyes, which
reflects their macroscopic behaviour and also their prevailing functionalities. They
are used in accordance to their compatibility with the type of textile substrate being
processed Acid, direct and reactive dyes are water-soluble anionic dyes; basic dyes
are cationic, whereas disperse, pigment and solvent dyes are non-ionic (Hao et al.
2000). Disperse dyes are sparingly soluble in water for application in hydrophobic
fibres from an aqueous dispersion. They are often of anthraquinone and sulfide
structure, with many -C =O, -NH- and aromatic groups (Fu and Viraraghavan, 2001).
Based upon the chemical structure or chromophore, they may be catergorized like
azo (monoazo, disazo, triazo, polyazo), anthraquinone, phthalocyanine, triarylmethane,

Review of Literature 36
diarylmethane, indigoid, azine, oxazine, thiazine, xanthene, nitro, nitroso, methine,
thiazole, indamine, indophenol, lactone, aminoketone, hydroxyketone stilbene and
sulphur dyes. Most of the mordant dyes are anionic in nature with few exceptional
cationic forms. In aqueous solution, a net charge on anionic dyes is attributed to the
presence of sulphonate (S03-) groups, while the presence of net positive charge on
cationic dyes is due to protonated amine or sulfur containing groups. Solubility in a
dye molecule is a trait of sulphonic group. Disperse vat dyes (indigo) are insoluble in
water; however in presence of reducing conditions, they are convertible into a 'leuco'
form (soluble in alkaline aqueous solutions) which are penetrated into the fibres
during dyeing process. The dyes with metal-complex exhibit higher light and wash
fastness because of presence of transition metals, such as chromium, copper, nickel
or cobalt that modify the surface chemistry between the dye molecule and the fabric
(Hao et al. 2000).
Based on the application to fibre, different classes of dyes are used Reactive
dyes are most commonly used as they can be applied to both in natural (wool, cotton,
silk) and synthetic (modified acrylics) fibres (O'Neill et al. 1999). The characteristic
feature of Reactive dyes which makes them different from other class of dyes is that
their molecules contain one or more reactive groups capable of forming a covalent
bond with a compatible fibre group. They possess the properties of high wet-fastness,
brilliance and range of hues (Hao et al. 2000). Their usage as synthetic fibres has
enhanced. Acid and basic dyes are used for dyeing all natural fibres (wool, cotton,
silk) and some synthetics (polyesters, acrylic and rayon). Direct dyes are applied
directly to cellulose fibres. Furthermore, they are used for colouring rayon, paper,
and leather and to certain extent nylon. The application of mordant dyes is limited to
the colouring of wool, leather, furs and anodised aluminium. Solvent dyes are used
for colouring inks, plastics, and wax, fat and mineral oil products. The direct dyes are
the second largest dye class in the colour index with respect to the amount of the
dyes. Direct dyes are relatively large molecules with high affinity for cellulose fibres
which is attributed to Van der Waals forces that make them bind to the fibre. About
1600 direct dyes are listed but only 30% of them are in current production. Society of

Review of Literature 37
Dyers and Colourists classifies direct dyes by their dyeing characteristics (Preston,
1986).
Azo dye are the largest group of dyes (Saranraj et al. 2013) with -N=N- as a
chromophore in an aromatic system. There are monazo, disazo, trisazo, and polyazo
dyes depending upon the number of azo-groups present. Diazotisation of a primary
amine, in presence of HCl + NaNO2 at freezing temperature, produces a diazonium
salt which in turn coupled with aromatic compounds, produces an azo-dye (Lavanya
et al, 2014) .The phenomena of diazotization was discovered by a german scientist
Gries in 1858 (Saranraj et al.2013). Of all known dyestuffs in the world, azo dyes
make up about a half, making them the largest group of synthetic colourants and the
most common synthetic dyes released into the environment (Zhao and Hardin, 2007).
Almost one million tons of dyes are annually produced in the world, of which azo
dyes, characterized by an azo-bond (R1–N=N–R2), represent about 70% by weight
(Hao et al.2000).
Azo dyes are the most common synthetic colourants mostly used for yellow,
orange and red hues. They are released to the environment via textile, pharmaceutical
and chemical industries. Structurally, they are characterised by reactive groups that
form covalent bonds with HO-, HN-, or HS- groups in fibres (cotton, wool, silk,
nylon). Approximately, 10–15 % of the dyes are released into the environment
during dyeing of different substrates, such as synthetic and natural textile fibres,
plastics, leather, paper, mineral oils, waxes, and even foodstuffs and cosmetics
(Sponza and Isik, 2005).
Azo dyes are second to polymers in terms of the number of new compounds
submitted for registration in the United States (US) under Toxic Substance Control
Act (TSCA) (Brown and DeVito, 1993). Chemically, the textile azo dyes are
characterized by relatively high polarity and high recalcitrance. Recalcitrance is
difficult to evaluate because of the dependence of degradation on highly variable
boundary conditions (e.g., redox milieu or pH). The azo dyes can accept protons
because of the free electron pair of the nitrogen, and the free electron pair of nitrogen
interacts with the delocalized π-orbital system. Acceptor substituents at the aromatic
ring such as –Cl or –NO2 cause an additional decrease in the basic character of

Review of Literature 38
aminic groups. Donor groups such as –CH3 or –OR (in meta and para position) lead
to an increase in the basicity of aromatic aminic groups. However, donor
substituients in the ortho position can sterically impede the protonation and
consequently decrease the basicity of aminic groups. The azo dyes are characterized
by amphoteric properties when molecules contain additional acidic groups such as
hydroxyl, carboxyl or sulfoxyl substituents. Depending on pH value, the azo dyes
can be anionic (deprotonation at the acidic group), cationic (protonated at the amino
group) or non-ionic. Environmental partitioning is influenced by substituents as well
as the number of carbon atoms and aromatic structure of the carbon skeleton
influence environmental partitioning. Amino group causes a higher boiling point, a
higher water solubility, a lower Henry’s law constant, and a higher mobility in
comparison with hydrocarbons (the amino group can also reduce the mobility by
specific interactions with solids via covalent bonding to carbonyl moieties or cation
exchange) (Börnick and Schmidt, 2006).
Azo dyes are structurally diverse in nature thus they are not uniformly
susceptible to microbial attack (O’ Neill et al. 1999; Couto, 2009) especially in case
of cotton fibres. Azo dyes cannot be degraded under aerobic conditions; under
anaerobic conditions, the azo linkage can be reduced to form aromatic amines which
are colourless but can be toxic, carcinogenic and mutagenic (Cripps et al. 1990;
Pinheiro et al. 2004). Several amino substituted azo dyes including 4-
phenylazoaniline and N-methyl- and N,N-dimethyl-4-phenylazo anilines are
mutagenic and carcinogenic (Yaneva and Georgieva, 2012). Direct dyes constitute
the major part of azo dyes which are primarily used for dyeing cotton fabrics. One
such dye which founds a common use in textile sector with special emphasis to Pali’s
scenario is Direct Red 28 commonly known as CR.
CR is a benzidine-based (Shinde and Thorat, 2013) direct, anionic diazo dye
(Jaladoni-Buan et al. 2010) chemically prepared by coupling tetrazotised benzidine
with two molecules of napthionic acid. Chemically, it is a sodium salt of
benzidinediazo-bis-1- naphtylamine-4 sulfonic acid (Jaladoni-Buan et al. 2010;
Perumal et al. 2012; Shinde and Thorat, 2013). Primarily, it has been reported to be a
carcinogenic direct diazo dye used for colouration of paper products (Cripps et. al.

Review of Literature 39
1990; Jaladoni-Buan et al. 2010). Besides having found it’s immense usage in textile,
it is also used in medicine (as a biological stain) and as an indicator (Perumal et al.
2012). The effluents containing CR are generated from a number of industrial
activities: textiles, printing and dyeing, paper, rubber, plastics industries (Purkait et
al. 2007; Vimonses et al. 2009).
This dye has been known to cause allergic reactions and it’s cytotoxicity for
bacteria and algae are well documented. Besides this, these dyes also exhibit
cytotoxicity; genotoxicity; hematotoxicity; neurotoxicity, as well as carcinogenicity
(Rajamohan and Kartikeyan, 2006; Kumar and Sawhney, 2011). The study
conducted by Puvaneswari in 2006 suggests dye to be carcinogenic for urinary
bladder of humans and tumorigenic in animals. The probable cause for
carcinogenicity underlies the fact that it forms carcinogenic amines such as benzidine
through cleavage of one or more azo groups is expected to metabolize to benzidine
(Mathur et al. 2005). This reason validates it to be under category of banned dyes
(Pielesz, 1999). It has also found to induce mutagenicity (Sharma et al. 2009;
Vimonses et al. 2009; Han et al. 2009; Sabnis, 2010; Zvezdelina et al. 2012). Kumar
and Sawhney in 2011 studied phytotoxicity of CR and the impact of the dye on
parameters of plant growth. The recalcitrance of CR has been attributed to the
presence of aminobiphenyl group and azo bonds, two features generally considered
as xenobiotic (Pinheiro, 2004; Sponza and Isik, 2005). Thus, the treatment of CR
contaminated wastewater is a complex process due to it’s aromatic structure,
imparting the dye it’s physicochemical, thermal and optical stability and resistance to
biodegradation and photodegradation (Purkait et al. 2007; Vimonses et al. 2009;
Smaranda et al. 2011). Consequently, due to the harmful effects of this organic
compound, the wastewaters containing CR must be treated properly before being
discharged to receiving water bodies.
2.4 Microbial Diversity: Exploring the unexplored
Microorganisms comprise of a huge and almost untapped reservoir of myriad
forms of molecular and chemical diversity in nature, as they constitute the most
diverse forms of life and are resourceful in terms of extrapolating some innovative
applications for the mankind. They have been evolving for nearly 4 billion years and
occupy almost every ecological niche on this planet and exhibit a tremendous

Review of Literature 40
potential in exploiting a vast range of energy sources. The literature suggests that it is
the microbial protoplasm which constitutes for about 50% of the living protoplasm
on this planet. Being highly adaptive in nature, micro-organisms have shown their
presence even in extremely diverse environments pertaining to which they have
developed an extensive range of metabolic pathways (Jain et al. 2005). This
exemplifies an excellent example of sustained presence in an environment. In natural
environments, microorganisms are present in mixed populations (Flemming and
Wingender, 2010). Different species with different characteristics live together, often
in very complex communities. Microbial community can be described by separating
the individual components. This segregation forms the basis for most techniques to
study microbial communities in natural environments. The presence of
microorganisms in extreme conditions of stress or contaminated environments
facilitates their use in different biotechnological interventions as their enzymatic
systems are coded by genes which could be exploited. Additionally, the use of such
highly tolerant autochthonous strains in wastewater bioremediation is a focal point of
research (Gomaa and Momtaz, 2007). Only a fraction of the culturable bacteria
present in an environmental sample can be successfully isolated and cultivated
(Hugenholtz, 2002). To circumvent the problem of isolating the different species by
growth condition preferences, DNA-based molecular techniques over the last few
decades have been developed and had revealed an enormous reservoir of unexplored
or unculturable but viable microbes. Un-culturability is a condition that
encompasses: (i) Lack of specific growth requirements (nutritional, temperature and
aeration); (ii) slow-growing strains, out-competed in the presence of fast- growers
and (iii) injured organisms, which are incapable of overcoming the stressful
conditions imposed by cultivation. These categories may not represent specific
taxonomic positions but account for about 99% of the environmental bacterial
diversity. This large genetic diversity can potentially be used as a bioresource,
leading to development of novel biotransformation, bioremediation processes and
bioenergy generation (Kalia et al. 2003 a, b; Lee et al. 2006). Culturing these
unexplored microbes in controlled laboratory environment requires extensive
knowledge of their fastidious growth requirements. This has in fact been the driving
force for development of new methods to access this vast microbial wealth (Wanger
and Loy, 2002; Kalia et al. 2003 a, b; Handelsman, 2004; Green and Keller, 2006).
However, the development of culture-independent methods and the

Review of Literature 41
commercialization of next-generation sequencing technology (Mardis, 2008) have
yielded powerful new tools in terms of time savings, cost effectiveness, and data
production capability. Methods such as 16S rDNA gene clone libraries, Fluorescence
In Situ Hybridization (FISH) or Denaturing Gradient Gel Electrophoresis (DGGE)
are being used to explore the bacterial diversity in waters have been reported
judiciously (Dewettinck et al. 2001; Zwart et al. 2002; 2005; Hoefel et al. 2005; Loy
et al. 2005; Bottari et al,. 2006; Wu et al. 2006; de Figueiredo et al. 2007; Von
Mering et al. 2007; Revetta et al.2010; Gabriel, 2010). More recently, the potential
of the high-throughput 454 pyrosequencing to explore the environmental diversity
has been emphasized (Roh et al. 2010).
16S rDNA gene diversity specifies the idea of species richness (number of
16S rDNA gene fragments from a sample) and relative abundance (structure or
evenness), which are reflective of relative pressures that construct diversity within
biological communities (Manefield et al. 2005; Paul et al. 2006). 16S rDNA based
molecular identification aims at identification, by virtue of it’s universal distribution
among bacteria and the presence of species-specific variable regions. Figure 12
represents characterization of microbial diversity by 16S rDNA sequencing strategy.
Figure 12: Flowchart depicting characterization of microbial diversity by 16S
rDNA analysis (Hobel, 2004)

Review of Literature 42
The selective pressures that shape diversity within communities are reflected
by richness and evenness of bacterial communities. These parameters need to be
measured for assessment of different treatment effects (e.g., physical disturbances,
pollution, nutrient addition, predation, climate change, etc.) on community diversity.
This molecular tool has been extensively used assessing the bacterial phylogeny and
to assign the right taxonomic position of an environmental isolate (Yumoto et al.
2001) it’s applied aspect aimed at bacterial identification of unculturable
microorganism, unique or novel isolates and collections of phenotypically identified
isolates (Drancourt et al. 2000). The polyphasic approaches for microbial diversity
are represented in Figure 13.
Molecular methods have been used to analyze the microbial diversity of a
wide range of environments, which has generated many beneficial findings.
Examples include solid waste composters (Nakasaki et al. 2009), wastewater
treatment plants (Wagner et al. 2002) agricultural soils (Ranjard et al. 2000) and
natural rivers (Brummer et al. 2000). Furthermore, culture-independent approaches
have been used to identify many novel bacterial and archaeal lineages from different
environments (Oren, 2004).
Figure 13: Assessment of microbial diversity through culture dependent and
culture independent approach (Jain et al. 2005)

Review of Literature 43
As a consequence, molecular approaches have lead to the interpretation, that
microbial world that is genetically and functionally more complex and diverse than
previously predicted from culture-dependent approach. However, the main
bottleneck of culture-independent approach lies in the fact that the strains in a
particular sample are indistinguishable and the unique properties of a particular
strain cannot be identified which requires intensive culture dependent approach to
assess microbial activites (Rapp & Giovannoni, 2003). Therefore, it would be very
difficult to use these culture-independent approaches to conduct a detailed study of
an individual strain for the development of an applied technology. In other words,
despite the widespread use of culture-independent approaches, cultural isolation will
continue to be an important but necessary method to generate new technologies.
Figure 14 represent the multidisciplinary approach to identify microbial diversity.
Figure 14: Multidisciplinary approach to analyze and characterize the microbial diversity through culture independent approach. (Hugenholtz, 2002)

Review of Literature 44
This approach of 16 S rDNA sequencing which utilize the molecular biology
tools have resulted in the discovery of entirely new lineages, some of which are
major constituents of environmental communities that were not detected by
traditional conventional systems which have lead to emergence of metagenomics to
access the gene pool of vast diversity of unculturable bacteria (Sharma et al. 2005).
The molecular phylogenetic analysis of bacterial communities by culture-
independent studies has resulted in increase of identifiable bacterial divisions to 40,
13 of which are characterized by environmental DNA sequences from unculturable
bacteria and are henceforth known as candidate divisions (Hugenholtz et al. 1998).
The microbial communities in ETP exist in dynamic consortial forms, the
understanding of which could be made through the knowledge of different co-
existing microbial populations (Manefield et al. 2005) which is non uniform with
changing operational conditions of the reactor (Kapley et al. 2007). Their
contribution to overall degradation of pollutants is likely to provide un paralleled
control over the bioremediation of the effluents (Manefield et al. 2005; Paul et al.
2006).
In wastewater treatment, microbial molecular ecology techniques have been
applied mainly to the study of flocs (activated sludge) and biofilms that grow in
aerobic treatment systems like trickling filters (Sanz and Kochling, 2007). Prokaryotes
are among the most important contributors to the transformation of complex organic
compounds in WWTP. Forster et al. in 2003 has reported the importance of bacterial
assemblages to the proper functioning and maintenance of treatment plants.
Molecular approaches such as 16S rDNA clone libraries (Blackall et al.1998 ;
McGarvey et al. 2004; Otawa et al. 2006), Ribosomal Intergenic Spacer Analyses
(RISA) (Yu and Mohn, 2001; Baker et al. 2003), 16S-restriction fragment length
polymorphism (16S-RFLP) (Baker et al. 2003) Repetitive Extragenic Palindrome
PCR (REP-PCR) (Baker et al. 2003) and fluorescent in situ hybridization (FISH)
( Bjornsson et al. 2002) have already been applied to the study of wastewater-
associated microbial communities.

Review of Literature 45
PCR–DGGE has been successfully implemented in many fields of microbial
ecology to assess the diversity and to determine the community dynamics in response
to environmental variations. DGGE-based approach has been used to study bacterial
diversity in wastewaters using reactors systems (Liu et al. 2002; Casserly and
Erijman, 2003; Kaksonen et al. 2003; Rowan et al. 2003) and activated sludge (Boon
et al. 2002; Ibekwe et al. 2003; Gilbride et al. 2006) revealing the presence of highly
complex bacterial communities. Bacterial diversity of aerated lagoons from WWTP
so far have not been substantially assessed where the degradation of organic matter
takes place. Cloning had been employed to establish with precision the phylogenetic
position of filamentous bacteria in granular sludge (Sekiguchi et al. 2001) or to
determine the prevalent sulfate reducing bacteria in a biofilm (Ito et al. 2002). The
study was extended by Yamada et al. 2005 who found the most prevalent sulfate
reducer belonging to Chloroflexi subphylum. This prokaryote has been associated
with bulking of sludge in treatment plants. The microbial communities residing in
reactors for treating several types of industrial wastewater have also been determined
by means of 16S rDNA cloning and sequencing. Egli et al. 2004 examined the
microbial composition and structure of a rotating biological contactor biofilm for the
treatment of ammonium-contaminated wastewaters. The study revealed the
sequences of several previously undetected and uncommon microorganisms.
Phylogenetic analysis of the sequences obtained showed a narrow range of diversity,
with most of the screened microorganisms belonging to the Methanosarcina sp.
Studies conducted by Zhang et al. in 2005 elucidated the efficacy of cloning
approach in conjunction with in situ hybridization analysis in methanogenic reactor
adapted to phenol degradation.
This essentially requires designing of new specific primers and gene probes
for detection and/or quantification of microorganisms. On the similar lines, Crocetti
et al. 2000 extracted genomic microbial DNA from a sequencing batch reactor
cloned the bacterial 16S rDNA and identified Rhodocyclus sp. and Propionibacter
pelophilus as the microorganisms responsible for the polyphosphate accumulation
taking place in the reactor. Futhermore, the same research group, designed probes for
these species that could correlate phosphorous removal and the number of hybridized
cells in different sludges. Alpha-proteobacteria, whose role in anaerobic/aerobic-

Review of Literature 46
activated sludge phosphorous removal plants has been analysed were characterized
by new probes for in situ hybridization with information provided by 16S rDNA
gene library sequences and DGGE analysis (Beer et al. 2004). A similar approach
had been considered three years earlier by the same authors, but Kong et al. 2001
utilised the combination of cloning, DGGE, and FISH for identification of
predominant microorganisms in an anaerobic sequencing batch reactor (SBR)
without probe design and phosphorus removal. A comparative analysis of further
insights into molecular biology tools for detection of microbial diversity are
represented in Table 8.
Table 8: Comparative analysis of further insights into molecular biology tools
for detection of microbial diversity (Sanz and Kochling, 2007)
S.No Method Outline Advantages Disadvantages
1 tRFLP 16 S based, relies on differences in polymorphism
Relatively simple procedures
Heterogenous size of fragments makes
phylogenetic analysis less confident
2 RISA Phylogeny with intergenic spacer region between 23S and 16S
rDNA sequences
High sensitivity, down to sub-species level
Database for comparative analysis small
in comparison to 16S sequences
3 PCR with genes
Microorganisms containing enzymes
involved in the biodegradation process are detected
Direct detection of the presence of degradative microorganisms.
Subtyping on strain level is possible
Global profiling of microbial
community missing
4 PLFA Profiling of microorganisms by
characteristic fatty acid content
Molecular characterization of microorganisms not relying on genes.
Complementary information to 16S based assays
Not a good choice as a standard
alone method
5 DNA microarray
Multi sample hybridization method
High sample throughput Parallel analysis of
different parameters
Expensive equipment, difficult handling

Review of Literature 47
Studies on bacterial diversity of waste water systems have disseminated
knowledge with respect to potential of indigenous microbes in attenuation of
pollutants from contaminated sites. Studies based on assessing microbial
communities of textile effluents by 16 S rDNA sequencing have been reported. To
comprehend the biochemical attributes of autochthonous strains, it becomes
quintessential to identify them and place them in appropriate divisons.
Karthikeyan and Anbusaravanan in 2013 conducted a study pertaining to
characterization of bacteria isolated from textile effluents influxed with sewage
released into the river Amaravathy, Karur, Tamilnadu , identified bacteria by 16 S
rDNA sequencing as Bacillus cereus AK1968 and Pseudomonas sp. AKDYE14.
Rajeswari et al. in 2013 screened textile effluent adapted bacteria for
degradation of different reactive dyes, from CETP, Tirupur and various textile units
and identified the potential isolate through sequencing of 16S rDNA gene as
Stenotrophomomonas maltophila RSV-2.
Ankita and Saharan in 2013 isolated a potential azo dye degrading bacterium
from grit tank of an ETP of tannery industries at Jajmau, Kanpur, Uttar Pradesh,
India and identified the strain by 16 S r DNA sequencing as Pseudomonas otitidis.
Khadijah et al. 2009 conducted a broad spectrum study for screening reactive
azo dye decolourising bacterial strains from effluent from local textile mill, batik
making site, textile laboratory, lake, streams, market, and domestic wastewater of
Shah Alam city. Of a remarkable 1540 bacterial isolates obtained, two potential dye
decolourisers were molecularly characterized as Chryseobacterium sp. and
Flavobacterium sp.
The isolation of microorganisms from highly contaminated environments
(textile effluents) indicates the natural adaptation of microorganisms to survive in the
presence of toxic dyes. The difference in their rate of decolourisation may be due to
the loss of ecological interaction, which they might be sharing with each other under
natural conditions (Bhimani, 2011) and offers novel bacteria of unique functionality
and potential applications in different biotechnological processes. It also entails a

Review of Literature 48
variety of genes responsible for microbial tolerance or defense against extreme
conditions or xenobiotics present in the media (Gomaa and Momtaz, 2007). Bhimani,
2011 carried out an elaborate study to assess the bacterial diversity of a CETP of
Jetpur, located in heart of Saurashtra, Gujarat which is largely infiltrated by textile
effluents. Of total 37 isolates screened, three exhibited tremendous potential to
degrade textile based azo dye and was characterized by 16 S rDNA sequencing and
identified them as Alcaligenes faecalis JTP-07; Pseudomonas aeruginosa JTP-37;
Lysinibacillus fusiformis JTP-23.
Halophilic bacteria have also shown to be a good source of dye degradation
(Kowsalya et al. 2013; Asad et al. 2014). Prabhakar in 2012 isolated a total of 84
bacterial strains from Kelambakkam Solar Salt Crystallizer ponds (or salterns) and
screened for their ability to produce extracellular tannase and laccase enzymes and
eventually to decolourize three widely used textile dyes- Reactive Blue 81, Reactive
Red 111 and Reactive Yellow 44. Of these 84 strains, 18 strains exhibited tannase
activity and 36 strains showed positive laccase enzyme activity. The 11 bacterial
strains that displayed both tannase and laccase enzyme activity were screened for
their ability to decolourize the three textile azo dyes. The best isolate AMETH72 was
sequenced for it’s 16S rDNA gene and was identified as Halomonas elongate.
Bacillus sp is supposedly unique with respect to it’s adaptability to dye based
environments; both soil contaminated with dyes and effluents at large (Devassy,
2010) and this genus is ubiquitous in nature with ability to degrade a wide range of
substrates. Plausibly, comprehensive reports pertaining to textile dye decolourizing
potential of this inherent species are widely available and are indicative of it’s role in
bioremediation of textile effluents. A study was conducted to isolate a potential
bacterial strain from textile wastewater generated by textile industries in Cairo,
Egypt and establish it’s tolerance to pH, low temperature and hydrogen peroxide.
The partial 16S rDNA gene amplification and sequencing revealed the strain to be a
gram positive rod and were identified as Bacillus macroccanus with (Gomaa and
Momtaz, 2007). In a similar study conducted on soil contaminated with dye and
untreated textile mill effluent, Bacillus sp identified through culture dependent
approach has found to possess versatility in terms of azo dye decolourisation

Review of Literature 49
(Dubey et al. 2010). In an another study, bacteria from soil contaminated with dye
was screened for it’s potential to degrade reactive azo dye, the 16s r DNA gene
sequencing revealed the isolated organism as Enterococcus faecalis strain YZ66
(Sahasrabudhe and Pathade, 2011). Aerobic strains of Streptococcus faecalis and
Bacillus subtilis had been isolated and identified by 16S r DNA sequencing from
textile effluent and had been explored for metabolizing the textile effluent. This
study is represents the remarkable feature of strains adaptability to textile effluents
(Sivaraj et al.2011). In an another study, Bacillus subtilis RA-29 isolated from
garden soil in the vicinity of an industrial town in Himachal Pradesh had been
phylogenetically characterized (Kumar and Sawhney , 2011).
Kowsalya et al. 2013 explored bacterial diversity of effluent samples
collected from textile dye units from various identified sites in Kanchipuram,
Tamilnadu to establish their bioremediative properties with respect to dye
degradation. The isolates were characterized as Halomonas KB1; Bacillus KB2;
Bacillus subtilis KB 3; Bacillus cereus KB4; Staphylococcus sp. A bacterium
isolated and identified as Bacillus cereus from tannery effluent collected at Central
Leather Research Institute (CLRI), Chennai was employed for decolourisation of an
azo dye (Kanagaraj et al. 2011).
Shah et al. 2013 isolated dye decolourizing bacterial isolate from textile dye
effluent from textile industries of Ankleshwar, Gujarat, India and identified it as
ETL-79(Bacillus sp.) through culture dependent approach. An earlier study
conducted by Shah et al. 2013 also revealed the presence of ETL-1 (Klebsiella
oxytoca) and ETL-2 (Bacillus subtillis) out of 84 strains isolated from Common
Effluent Treatment Plant (CETP) of Ankleshwar, Gujarat, India for dye degradation
studies. Pradhan et al. in 2012 conducted a study on waste water samples generated
from sugar industry to screen for dye degrading bacteria and the best dye degrader
was characterized at molecular level was Bacillus sp. SCWS5.
Soil and effluent samples obtained from CETP, Ankleshwar, Gujarat had
been the source of an environmental isolate Pseudomonas steutzeri-ETL-4 as
characterized by 16 S rDNA sequencing. This strain had been judiciously utilsed for

Review of Literature 50
degradation of CR which is a sulphonated diazo direct dye (Shah, 2014). In a similar
study carried out earlier, activated sludge obtained from CETP, Naroda, Gujarat had
been a source of an indigenous bacterial strain Bacillus boroniphilus as identified by
16 S rDNA sequencing to degrade Reactive Yellow 145 dye (Derle et al. 2012).
Subhatra et al. 2013 investigated the potential of bacteria (Bacillus sp.)
isolated from effluent produced by textile industries in Madurai , Tamil Nadu to
establish a synergistic effect on heavy metal tolerance and dye degrading potential.
Milikli and Rao, 2012 investigated soil samples from contaminated sites of
Mangalagiri textile industry, Guntur, Andhra Pradesh, India to screen out the most
potential Bromophenol blue dye degrading bacteria. Through selective screening the
most potential isolate out of the 3 isolates screened, was identified by 16 S rDNA
sequencing as Bacillus subtilis.
Manivannan et al. 2011 explored the dye house effluent collected from a dye
processing industry, Arulpuram, Tirupur region, Tamil Nadu to assess it’s bacterial
diversity to explore the biodegradability of Orange 3G dye. The efficient strains
identifies were Bacillus sp., Escherichia coli and Pseudomonas fluorescens
Balakrishnan and Sowparnika, 2013 studied bacterial diversity of fresh textile
effluent released by textile industries in Erode district, Tamil Nadu, India in terms of
dye degradation. The most promising bacterial isolates identified by biochemical
tests were found out to be Pseudomonas sp., Bacillus sp and Alcaligenes sp.
Balgurunathan and Sudha, 2013 isolated and identified a bacterial strain as
Bacillus licheniformis from effluent sample of Infra Tex textile industry, Perundurai
in Erode district Tamil Nadu, India for it’s use as excellent biomass for removal of
reactive dyes. A similar study was conducted to explore the bacterial diversity at the
same site; Perundurai in Erode district of Tamil Nadu, India in which soil
contaminated with dye was collected at effluent discharge site and the isolates were
screened for a repertoire of textile dyes and the most efficient dye degraders were
Pseudomonas aeruginosa, Alcaligenes faecalis, Proteus mirabilis, Serratia
marcescens and Bacillus licheniformis as identified by a series of biochemical tests
(Neelambari et al. 2013).

Review of Literature 51
A combined study was carried out in which both the textile effluent and
sludge samples from Amanishah nullah, Jaipur were assessed for bacterial diversity
aimed at exploring the dye decolourising potential of the adapted bacterial strains.
Most potential light red dye degrading bacterial strains identified through culture
dependent approach were Pseudomonas sp, Klebsiella sp and Proteus sp from the
effluent and Shigella sp, Morganella sp and Klebsiella sp from the sludge sample
(Sethi et al. 2012). A similar study was carried out to explore potential dye
decolourizing bacterial strains from the textile industry waste located in Erode and
Tripur districts, Tamil Nadu, India. There was 96 morphologically distinct bacterial
isolates were isolated from 12 different sludge, textile effluent and dye contaminated
soil samples. Generic composition of the 96 isolates comprised of Bacillus sp.,
Enterobacteriaceae, Pseudomonas sp., Micrococcus sp.(Sahasrabhude et al. 2014)
Alcaligenes sp., Aeromonas sp., Staphylococcus sp., and Lactobacillus sp (Palani
Velan et al. 2012).
Not only does textile effluents and soil contaminated with dye have been the
source of Bacillus sp., it’s ubiquitous nature is well established from a report which
is suggestive of it’s source being the waste water generated from a carpet industry in
Khairabad, Uttar Pradesh, India (Dubey et al. 2010).
Marine bacteria are vital in recycling nutrients and could be useful for
innovative applications, which are helpful to human beings (Barakat, 2012). A study
was carried out to isolate the degrading bacterial strains from mangrove associated
sediment collected from nearby Roche Park, coastal area of Tuticorin and further to
assess the efficiency of selected bacterial strain in degradation of dyes and identified
the strain as Pseudomonas sp (Sponza et al. 2003) This report is unique in respect
that marine microorganisms have developed unique metabolic and physiological
capabilities that not only ensure survival in extreme habitats but also offer the
potential for the production of metabolites, which would not be observed, from
terrestrial organisms (Raja et al. 2013). Durve et al. 2012 isolated two potential dye
degrading strains from effluent samples of textile processing units in Maharashtra,
India and biochemically identified them as Pseudomonas aeruginosa and
Brevibacillus choshinensis (Surwase et al. 2013).

Review of Literature 52
Kannan et al. 2013 screened the dye-contaminated soil samples collected
from Kallidaikurichi, of Tirunelveli district, Southern Tamil Nadu, which is well
known for handloom production with flourishing dyeing units; to isolate and identify
the bacterial isolates which belonged to genera Pseudomonas sp. (32.5 %) followed
by Bacillus sp. (27.5 %), Aeromonas sp. (15.0 %), Micrococcus sp. (12.5 %) and
Achromobacter sp (12.5 %).
Globally, studies on bacterial diversity for dye degradation studies are also
reproducible. Bacterial strains were isolated from activated sludge (Corona
Wastewater Treatment Plant, CA, USA), turf grass soil (Chino loamy sand, pH 7.4,
Chino Park, CA, USA) and from a natural asphalt soil mixture obtained from the
Rancho La Brea tar pit’s in Los Angeles, CA, USA and characterized the strains
based upon their 16S rDNA sequence similarities. From activated Sludge AS7
Bacillus cereus CCM 2010; AS77 Pseudomonas nitroreducens 0802; AS81
Aeromonas punctata MPT4; AS96 Shewanella putrefaciens LMG 2369; Soil S46
Bacillus thuringiensis Al Hakam; S81 Massilia timonae were isolated (Khalid et al.
2008). A bacterial strain, CK3, with remarkable ability to 45 decolourize the reactive
textile dye Reactive Red 180, was isolated from the activated sludge collected from a
textile mill. Phenotypic characterization and phylogenetic analysis of the 16S rDNA
sequence indicated that the bacterial strain belonged to the genus Citrobacter CK 3
(Wang et al. 2009). Pahlaviani et al. 2011 isolated native bacterial strains from
activated sludge, turf grass soil in Iran for examining their azo dye degrading
potential. The strains were identified by 16 S rDNA sequencing as Bacillus cereus
CCM 2010, Pseudomonas sp. B13T, Shewanella algae ATCC 51192, Shewanella
algae ATCC 51192.
Olukanni et al. 2006 investigated the potential of effluent adapted and non-
adapted bacteria isolated from textile industries wastewater and outlet in Nigeria,
Africa. The effluents from textile industries and drains served as a source of effluent
adapted bacteria and isolates from a municipal landfill were effluent non-adapted
bacteria. Effluent adapted strains of Acinetobacter, Bacillus and Legionella with
potentials for colour removal and COD removal activities were characterized.
Likewise, a similar study conducted by Ajao et al. 2013 was based on dye effluent

Review of Literature 53
from International Textile Industry (Nig) Ltd, Odogunyan Industrial Estate Ikorodu,
Lagos State, Nigeria to explore dye decolourising potential of adapted strains under
immobilized conditions on agar-agar in a bioreactor. The strains were characterized
as Pseudomonas aeruginosa and Bacillus subtilis.
Barakat, 2012 isolated a novel azo dye-degrading bacterium T312D9 strain
from Abou Quir Gulf, Amya industrial pumping station considering enriched
location contaminated with various dyes in Alexandria, Egypt. The identification of
the isolate by 16S rDNA gene sequencing revealed to be Lysobacter sp T312D9.
This marine ecofriendly isolate was exploited for it’s ability to degrade two synthetic
azo dyes (CR and methyl red) considered as detrimental pollutants from industrial
effluents.
Mahmood et al. 2012 screened wastewater (Textile effluents), sludge and
affected soil samples from Hudiara drain near Nishat Mills Limited from Ferozepur
Road Lahore, Pakistan to isolate and identify potential dye degrading bacterial
strains. The isolates were identified as Bacillus subtilus (Isolate 20), Bacillus cereus
(Isolate 3), Bacillus mycoides (Isolate 1), Bacillus sp. (Isolate 5), Pseudomonas sp.
(Isolate 9) and Micrococcus sp. (Isolate 7) by standard physiological, morphological
and biochemical tests. The isolation of good dye-decolourizing species requires
screening, and these isolated strains should have ability to degrade and detoxify
textile dyes (Silveira et al. 2009). Yang et al. 2011 screened heavily dye
contaminated soil and wastewater samples from Changzhou dye manufacturing
industries and Qingtan wastewater treatment factory in Changzhou, China for
isolation of a potential dye degrading bacterial isolate The isolate was identified
through culture independent approach by 16 S rDNA sequencing and designated as
Psedomonas putida WLY. Pseudomonas aeruginosa , an effleuent adapted microbe
isolated from textile waste water selected from a group of dye degrading bacteria
obtained, was chosen for achieving the best decolourisation and for it’s broad
spectrum azo dye biodegradability potential (Hafshejani et al. 2013). Pseudomonas
sp. is widely distributed in a variety of habitats and is able to metabolize organic
contaminants in the environment (Ashiuchi et al. 1999; Poornima et al. 2010).

Review of Literature 54
Sun-Young An et al. 2006 isolated and identified a bacterial strain as
Citrobacter sp. from textile waste water capable of degrading both azo and
triphenylmethane group of dyes. An azo-dye-reducing, endospore-forming bacterium
DSM 13822T isolated from textile industry wastewater has been taxonomically
studied and identified as Paenibacillus azoreducens (McMullan et al. 2001).
Recently, a study focused at isolation of bacteria from activated sludge of a
textile plant wastewater treatment facility was reported (Franciscon et al. 2012)
which was aimed at sequential decolourisation and detoxification of the azo dyes
Reactive Yellow 107 (RY107), Reactive Black 5 (RB5), Reactive Red 198 (RR198)
and Direct Blue 71 (DB71). 16 S r DNA sequence of the isolate VN-15 revealed it to
be Brevibacterium sp.
Activated sludge obtained from obtained from denitrifying reactor of
wastewater treatment plant (WWTP) in textile factory “Giorgetti Bulgaria” AD –
Elin Pelin, was used as a source of high performance bacterial decolourizer AZO29
to reduce Amaranth dye. The isolate was characterized as Pseudomonas sp (Vasileva
et al. 2009). Previous studies had focused on the isolation of bacterial strains from
activated sludge. In the early years, (Horan et al. 1988) had attempted to isolate both
floc-forming and filamentous bacteria from activated sludge flocs. Until recent years,
a few bacterial strains especially filamentous bacteria were cultivated by the method
of dilution plate (Ramothokang et al. 2003) or micromanipulation (Blackall et al.
1996). However, it should be noted that a majority of microorganisms present in any
environment have not been cultivated through general media (Ellis et al. 2003) thus,
obtaining more unexplored microbes is an important prerequisite for better
understanding what functions these strains have in this complex environment (Lu et
al.2006) which showed that β-Proteobacteria was the most dominant group in sludge
samples. In addition, at the genus level, strains of the genera Klebsiella,
Pseudomonas, Bacillus, Aeromonas, Flavobacterium and Acinetobacter (12.7%,
12.7%, 12.7%, 11.1%, 7.9% and 7.9%, respectively) were the most predominant
microorganisms in the collection. REP-PCR has attracted a lot of attention in recent
years, for it’s highly discriminatory, accurate, fast and low-cost advantages. To date,
it has been applied for analyses of biodiversity among bacterial strains in water

Review of Literature 55
(Mohapatra and Mazumder, 2008), soil (Rejili et al. 2009), plants (Sikora and
Redzepovic, 2003). However, there were no reports on the analysis of isolated
bacteria diversity in activated sludge using this method. Meanwhile, it should be
noted that different strains belonging to the same genus may play different roles in
activated sludge. For this reason, it is urgent to obtain more detailed evaluation on
biodiversity among the same genus; REP-PCR can be a suitable candidate reference
method for this purpose (Jin et al. 2011).
In the past years, many researchers focused on the enumerating the bacterial
community presents in activated sludge by using molecular approaches, including
DGGE and 16S rDNA gene clone library analysis (Blackall et al. 1998; Boon et al.
2002; Eschenhagen et al. 2003; Choi et al. 2007). These culture-independent
technologies can reveal more abundance of microbial diversity than culture-
dependent approaches in the terms of bacterial composition. The bacterial
community of a bulking sludge from a municipal WWTP with anoxic-anaerobic-oxic
process was investigated by combination of cultivation and 16S rDNA gene clone
library analysis for understanding the causes of bulking. A total of 28 species were
obtained from 63 isolates collected from six culture media. The most cultivable
species belonged to Proteobacteria including Klebsiella sp., Pseudomonas sp.,
Aeromonas sp. and Acinetobacter sp. At the level of class, β -Proteobacteria (43.6%)
was the dominant group in the bulking sludge (Zhuibai et al. 2011) which was in
accordance with previous studies (Rani et al. 2008). Both the marker and functional
genes had been used in identification (Suizhou et al. 2006) of a dye-decolourizing
bacterial strain DN322 from activated-sludge of a textile-printing WWTP,
Guangzhou, China and was identified as Aeromonas hydrophila based on sequence
analysis of 16S rDNA gene and gyrase beta subunit (gyr). The diversity of bacterial
groups of activated sludge samples that received wastewater from four different
types of industries (Flanders, Belgium) i.e., domestic wastewater, carbohydrate rich
wastewater, wastewater from paper and starch related industries, protein and fat rich
wastewater from food and meat related industries and wastewater from textile
industry was investigated by a nested PCR- DGGE (Denaturing Gradient Gel
Electrophoresis) approach. Specific 16S rDNA primers were chosen for large
bacterial groups (Bacteria and α-Proteobacteria in particular), which dominate

Review of Literature 56
activated sludge communities, as well as for actinomycetes, ammonium oxidisers
and methanotrophs (types I and II) In addition primers for the new Acidobacterium
kingdom were used to observe their community structure in activated sludge. After
this first PCR amplification, a second PCR with bacterial primers yielded 16S rDNA
gene fragments that were subsequently separated by DGGE, thus generating `group-
specific DGGE patterns (Boon et al. 2002).
2.5 Dye removal techniques
The addressal of environmental issues pertaining to presence of colour in
wastewater needs some stringent laws to be reinforced so as to develop treatment
technologies so that the effluents prior to their release may be treated properly and
threat to the environment may be minimized upto certain extent. The important
methods useful in management of environmental contaminants in the environment
include the following (Rajendran and Gunasekran, 2006) (Figure 15).
Figure 15: Methods aimed at management of environmental contaminants
(Rajendran and Gunasekran, 2006)
Management of environmental contaminants
Ongoing strategies
In process treatment
End-of-pipe treatment
long term strategies
Remediation of polluted sites
Modification of existing process
Introduction of new processes and
products

Review of Literature 57
Though many technologies have been available for clean up purpose, only a
few of them have been proved to be of routine application value. The choice of
technology is influenced by many factors. The available technologies can be
categorized as potential and time proven trusted technologies. The available
technologies are assessed based on certain criteria (Rajendran and Gunasekran,
2006). Figure 16 represents criteria for available technologies for “clean up”.
Figure 16: Criteria for available technologies for “clean up” (Rajendran and
Gunasekran, 2006).
Several physical, chemical and biological methods for the dyes removal from
the wastewater are available (Pagga and Laboureur, 1983; Cooper, 1993;
Vandevivere, et al. 1998; Hao, et al. 2000; Robinson et al. 2001; Blumel, et al. 2002;
Ali, 2010; Archana et al. 2012). The physicochemical treatment include membrane
filteration, coagulation/ flocculation, precipitation, flotation, adsorption, ion exchange,
ion pair extraction, ultrasonic mineralization, electrolysis, advanced oxidation
(Chlorination, bleaching, ozonization, fenton oxidation and photocatalytic oxidation)
and chemical reduction.
Temporary or permanent solution
More conversion or total conversion
Cost effectiveness
Time taken for treatment
Regulatory standards

Review of Literature 58
The textile organic dyes must be removed from industrial wastewaters by
effective and viable treatments at treatment plant or on site following two different
treatment concepts as: (1) separation of organic pollutants from water environment, or
(2) the partial or complete mineralization or decomposition of organic pollutants.
Separation processes are based on fluid mechanics (sedimentation, centrifugation,
filtration and flotation) or on synthetic membranes (micro- ultra- and nanofiltration,
reverse osmosis). Additionally, physico-chemical processes (i.e. adsorption,
chemical precipitation, coagulation-flocculation, and ionic exchange) can be used to
separate dissolved, emulsified and solid-separating compounds from water
environment (Robinson et al. 2001; Anjaneyulu et al. 2005; Zaharia, 2005; Babu et
al. 2007; Suteu et al. 2009a; Suteu et al. 2011a; Zaharia et al. 2009; Zaharia et al.
2011). The partial and complete mineralization or decomposition of pollutants can be
achieved by biological and chemical processes (biological processes in connection
with the activated sludge processes and membrane bioreactors, advanced oxidation
with O3, H2O2, UV (Dos Santos et al. 2004 ; Oztekin et al. 2010 ; Wiesmann et al.
2007 ; Zaharia et al. 2009). A strategic approach pertaining to different separation
processes (sedimentation, filtration, membrane separation), and some physico-
chemical treatment steps (i.e. adsorption; coagulation-flocculation with inorganic
coagulants and organic polymers; chemical oxidation; ozonation; electrochemical
process) are integrated into a specific order in the technological process of
wastewater treatment for decolourisation or large-scale colour and dye removal
processes of textile effluents. To introduce a logical order in the description of
treatment methods for textile dye and colour removal, the relationship between
pollutant and respective typical treatment technology is taken as a reference. The first
treatment step for textile wastewater is the separation of suspended solids and
immiscible liquids from the main textile effluents by gravity separation (e.g., grit
separation, sedimentation including coagulation/flocculation), filtration, membrane
filtration, air flotation, and/or other oil/water separation operations.
The following treatment steps are applied to soluble pollutants, when these
are transferred into solids (e.g., chemical precipitation, coagulation/flocculation, etc.)
or gaseous and soluble compounds with low or high dangerous/toxic effect (e.g.,

Review of Literature 59
chemical oxidation, ozonation, wet air oxidation, adsorption, ion exchange, stripping,
nanofiltration/reverse osmosis). Solid-free wastewater can either be segregated into a
biodegradable and a nonbiodegradable part, or the contaminants responsible for the
non-biodegradable wastewater part that can be decomposed based on physical and/or
chemical processes. After an adequate treatment, the treated wastewater can either be
discharged into a receiving water body. An outline of different treatment processes
for textile effluents are outlined in Table 9.
2.6 Biodegradation of dyes: Applied aspect of bacterial diversity
Traditional wastewater treatment technologies are markedly ineffective for
handling wastewater of synthetic textile dyes because of the chemical stability of the
pollutants (Forgacs et al. 2004). Additionally, the water recycling issue remains
unaddressed (Soares et al. 2004). Physico-chemical methods have long been used to
treat textile wastewater influxed with synthetic dyes. The major disadvantage of
physicochemical methods is primarily the high cost, low efficiency, limited
versatility, need for specialized equipment, interference by other wastewater
constituents, and the handling of the generated waste (Van der Zee and Villaverde
2005; Kaushik and Malik, 2009). Physical methods can effectively remove colour,
but the dye molecules are not degraded, becoming concentrated and requiring proper
disposal. With the chemical techniques, although the dyes are removed,
accumulation of concentrated sludge creates a menace. A possibility of secondary
pollution problem always arises because of the excessive amounts of chemicals
involved (Amoozegar, 2011). Recently, other emerging techniques like advanced
oxidation processes, which are based on the generation of very powerful oxidizing
agents such as hydroxyl radicals- have been applied with success in pollutant
degradation (Arslan and Balcioglu, 1999). Although these methods are efficient for
the treatment of waters contaminated with pollutants, they are very costly and
commercially unattractive compounds (Alexander, 1994; Bennet et al. 2002). These
compounds may be biodegradable, persistent or recalcitrant (Nikaido and Glazer,
2007) which may be cleaved into smaller compounds by viable microbes (Marinescu
et al. 2009).

Review of Literature 60
Table 9: Various current and emerging dye separation and elimination treatments applied for textile effluents with their principal advantages and limitations (adapted from Robinson et al. 2001; Anjaneyulu et al. 2005; Babu et al. 2007)
S.No Treatment methodology Treatment stage Advantages Limitations
PHYSICO-CHEMICAL METHODS
1 Precipitation,
Coagulationflocculation
Pre/main treatment Short detention time and low capital costs. Relatively good removal efficiencies.
Agglomerates separation and treatment. Selected operating condition.
2 Electrokinetic Coagulation Pre/main treatment Economically feasible. High sludge production.
3 Fenton process Pre/main treatment Effective for both soluble and insoluble coloured contaminants. No alternation in volume
Sludge generation; problem with sludge disposal. Prohibitively expensive.
4 Ozonation Main treatment Effective for azo dye removal. Applied in gaseous state: no alteration of volume.
Not suitable for dispersed dyes. Releases aromatic dyes. Short half-life of ozone (20 min).
5 Oxidation with
NaOCl
Post treatment Low temperature requirement. Initiates and accelerates azo bond cleavage.
Cost intensive process. Release of aromatic amines.
ADSORPTION WITH SOLID ADSORBENTS
6 Activated carbon
Economically attractive.
Pre/post
Treatment
Good removal efficiency of wide variety of dyes.
Very expensive; cost intensive regeneration
Process.
7 Peat Pre treatment Effective adsorbent due to cellular structure. No activation required.
Surface area is lower than activated carbon.
8 Coal ashes Pre treatment Economically attractive. Good removal efficiency. Larger contact times and huge quantities are required.
Specific surface area of adsorption is lower than activated carbon.

Review of Literature 61
S.No Treatment methodology Treatment stage Advantages Limitations
9 Wood chips/
Wood sawdust
Pre treatment Effective adsorbent due to cellular structure. Economically attractive. Good adsorption
capacity for acid dyes.
Long retention times and huge quantities are
required.
10 Silica gels Pre treatment Effective for basic dyes Side reactions prevent commercial application
11 Irradiation Post treatment Effective oxidation at lab scale Requires a lot of dissolved oxygen
12 Photochemical process Post treatment No sludge production Formation of byproducts
13 Electrochemical oxidation Pre treatment No additional chemicals required and the end products are non-dangerous/hazardous.
Cost intensive process; mainly high cost of
Electricity
14 Ion exchange Main treatment Regeneration with low loss of adsorbents Specific application; not effective for all dyes
BIOLOGICAL TREATMENT
15 Aerobic process Post treatment Partial or complete decolourisation for all classes of dyes
Expensive treatment
16 Anaerobic process Main treatment Resistant to wide variety of complex coloured compounds. Bio gas produced is used for stream generation.
Longer acclimatization Phase
17 Single cell (Fungal, Algal & Bacterial)
Post treatment Good removal efficiency for low volumes and concentrations. Very effective for specific colour removal.
Culture maintenance is cost intensive. Cannot cope up with large volumes of wastewater.

Review of Literature 62
S.No Treatment methodology Treatment stage Advantages Limitations
EMERGING TECHNOLOGIES
18 Other advanced
oxidation process
Main treatment Complete mineralization ensured. Growing number of commercial applications Effective pre-treatment methodology in integrated systems and enhances biodegradability.
Cost intensive process
19 Membrane
filtration
Main
treatment
Removes all dye types; recovery and reuse of chemicals and water.
High running cost. Concentrated sludge production. Dissolved solids are not separated in this process
20 Photocatalysis
Post treatment Process carried out at ambient conditions. Inputs are no toxic and inexpensive. Complete mineralization with shorter detention times. Effective for small amount of coloured compounds.
Expensive Process
21 Sonication
Pre treatment Simplicity in use. Very effective in integrated systems.
Relatively new method and awaiting full scale application.
22 Enzymatic
Treatment
Post treatment Effective for specifically selected compounds. Enzyme isolation and purification is tedious.
23 Redox mediators Pre/ supportive
Treatment
Easily available and enhances the process by increasing electron transfer efficiency
Concentration of redox mediator may give antagonistic effect. Also depends on biological activity of the system..
24 Engineered
wetland systems
Pre/post treatment Cost effective technology and can be operated with huge volumes of wastewater
High initial installation cost. Requires expertise and managing during monsoon becomes difficult

Review of Literature 63
The high electrical energy demand and the consumption of chemical reagents
are common problems. The development of efficient, economic and eco friendly
technologies to decrease dye content in wastewater to acceptable levels at affordable
cost is of utmost importance (Couto, 2009). Biological methods are generally
considered environmentally benign because they lead to complete mineralization of
organic pollutants at effectively low cost (Pandey et al. 2007). They also dissipate
BOD, COD and suspended solids. The main limitation relates, in some cases to the
toxicity of some dyes and/or their degradation products to the organisms used in the
process.
Natural attenuation or bioattenuation is the dilution of contaminants in the
environment through biological processes (aerobic and anaerobic biodegradation,
plant and animal uptake), physical processes (advection, dispersion, dilution,
diffusion, volatilization, sorption/desorption), and chemical responses (ion exchange,
complexation, abiotic transformation) (Joutey et al. 2013). Figure 17 represents natural
attenuation of pollutants.
The resultant product may not be essentially desirable as the simpler
compounds may be more toxic than the parent compound (Nikaido and Glazer,
2007). Biodegradation in current scenario involves aerobic microorganisms, which
utilize molecular oxygen as reducing equivalent acceptor during respiration
(Anjanyelu, 2005; Zaharia, 2012). Under anaerobic environment (anoxic and
hypoxic environments) biodegradation also occurs, and survival of microorganisms
(Singh, 2008) is attained by using sulphates, nitrates and carbon dioxide as electron
acceptors (Birch et al. 1989). Mineralization is complete biodegradation process
(Joutey et al. 2013) in which the end products are CO2, water and other inorganic
compounds (Nikaido and Glazer, 2007) and other simpler forms like cleaved ring
structures (Vallero, 2010). An involvement of enzyme in biodegradation describes
the situation as gratuitous biodegradation in which an enzyme transforms a
compound other than it’s natural substrate, provided the unnatural substrate is able to
occupy the enzyme’s active site and the enzyme may exhibit it’s catalytic activity
(Nikaido and Glazer, 2007).

Review of Literature 64
Figure 17: Natural attenuation for reduction of environmental contaminants
Natural attenuation
Physical
Advection
Dispersion
Dilution
Diffusion
volatalization
Sorption/ desorption
Chemical
Ion exchange
Complexation
Abiotic tranformation
Biological
Aerobic Anaerobic
Plant/ animal uptake

Review of Literature 65
Biodegradation is one of the energy dependent (Alves and Pereira, 2012)
natural attenuation processes that helps to eliminate xenobiotic compounds from the
environment by microbially mediated catalyzed reduction in complexity of chemical.
Extracting a microbe from the environment and exposing it to a target
contaminant under controlled conditions is a strategic approach to break down target
component into simpler non –toxic forms. This can be achieved by bioremediation
(Vallero, 2010). Bioremediation process can be divided into three phases or levels.
First, through natural attenuation, contaminants are reduced by indigenous
microorganisms without any human augmentation. Second, biostimulation is
application of nutrients and oxygen to the systems to enhance their effectiveness
and to accelerate biodegradation. Finally, during bioaugmentation, microorganisms
are added to the systems. These supplemental organisms should be more efficient
than native flora to degrade the target contaminant (Diez, 2010).
A feasible remedial technology requires microorganisms being capable of
quick adaptation and efficient uses of pollutants of interest in a reasonable period of
time (Seo et al. 2009). Many factors influence microorganisms to use pollutants as
substrates or co metabolize (Nikaido and Glazer, 2007; Vallero, 2010) them, like, the
genetic potential and certain environmental factors such as temperature, pH, and
available nitrogen and phosphorus. The rate and extent of degradation is dependent
upon environmental factors (Frit’sche and Hofrichter, 2008). Therefore, applications
of Genetically Engineered Microorganisms (GEM) with high degradative capability in
bioremediation have received a great deal of attention. However, ecological,
environmental concerns and regulatory constraints are major obstacles for testing
GEM in the field (Menn et al. 2008). Figure 18 represents bioremediation of polluants
utilizing biodegradation approach.
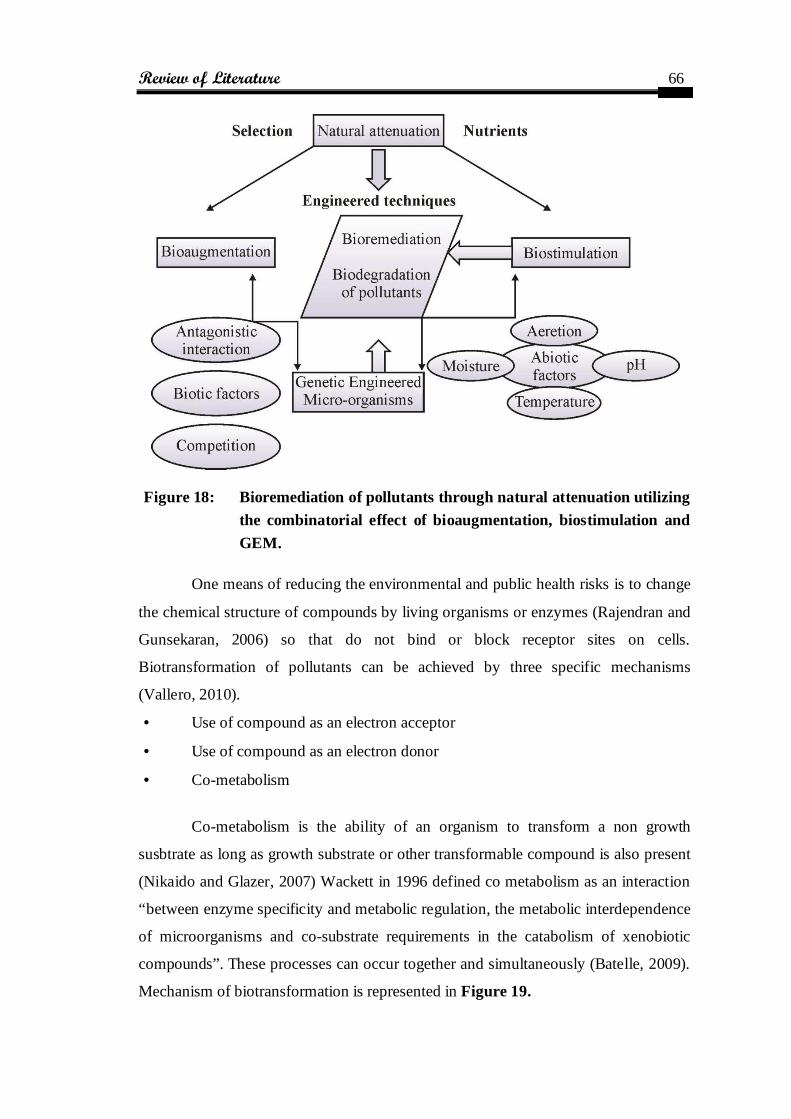
Review of Literature 66
Figure 18: Bioremediation of pollutants through natural attenuation utilizing the combinatorial effect of bioaugmentation, biostimulation and GEM.
One means of reducing the environmental and public health risks is to change
the chemical structure of compounds by living organisms or enzymes (Rajendran and
Gunsekaran, 2006) so that do not bind or block receptor sites on cells.
Biotransformation of pollutants can be achieved by three specific mechanisms
(Vallero, 2010).
• Use of compound as an electron acceptor
• Use of compound as an electron donor
• Co-metabolism
Co-metabolism is the ability of an organism to transform a non growth
susbtrate as long as growth substrate or other transformable compound is also present
(Nikaido and Glazer, 2007) Wackett in 1996 defined co metabolism as an interaction
“between enzyme specificity and metabolic regulation, the metabolic interdependence
of microorganisms and co-substrate requirements in the catabolism of xenobiotic
compounds”. These processes can occur together and simultaneously (Batelle, 2009).
Mechanism of biotransformation is represented in Figure 19.

Review of Literature 67
Figure 19: Mechanisms underlying biotransformation of pollutants (Vallero,
2010)
Among the current pollution control technologies for textile effluents, an
advantage of biological treatment over certain physicochemical treatment methods is
biodegradation of synthetic dyes by different microbes. It is emerging as an effective
and promising approach (Charumathi and Das, 2010) as that > 70% of the organic
material measured by the COD test can be converted to biosolids (Forgacs et al.
2004; Anjaneyulu et al. 2005) and is found to be an economic, effective, biofriendly,
and environmentally benign process (Verma and Madamwar, 2003; Pearce et al.
2003; Chen et al. 2003; Jirasripongpun et al. 2007; Kalyani et al. 2008; Shedbalkar
et al. 2008; Ozdemir et al. 2008 ; Gopinath et al. 2009 ; Ramchandran et al. 2013)
amount of sludge produced is also very less (Carvalho et al. 2008; Saratale et al.
2009) as contrasted with physico- chemical methods (Zhang et al. 2004;
Amoozegar, 2011) which produce enormous amount of sludge and the emit toxic
substances (Johnson et al. 1978) efficiency for colour removal particularly for
sulphonated azo dyes is less (Banat et al. 1996). Therefore, bioremediation is a
publicly acceptable treatment technology (Hao et al. 2000; Supaka et al. 2004) which
may prove to be a green solution to the problem of environmental soil and water
pollution in future (Ali, 2010; Archana et al. 2012) and at the same time biological
treatment process for decolourisation of industrial effluents is ambiguous, different
and divergent (Anjaneyulu et al. 2005). Several excellent reviews have been
published on the biodegradation or bioremediation, both generally (Prescott et al.
2008; Chatterjee et al. 2008) or specifically, of xenobiotic compounds (Austin et al.

Review of Literature 68
1977; Chaudhry and Chapalamadugu, 1991; Chauhan et al. 2008; Chowdhury et al.
2008). Various microorganisms such as bacteria both gram positive and gram
negative (Sani and Banerjee, 1999; Kodam et al. 2005; Moosvi et al. 2005; Kalyani
et al. 2009; Wang et al. 2009) as well as fungi (Balan and Monteneiro, 2001; Verma
and Madamwar, 2005; Taskin and Erdal, 2010) , yeasts, algae have been reported to
remove dyes (Stolz et al. 2001; Saranraj et al.2013; Ramchandran et al. 2013).
Suitable strains of microorganisms must be adapted to textile effluents prior
decolourisation activity (Saratale et al. 2009; Phugare et al. 2011) further, identified
and characterized to optimize the best conditions for effective biological treatment of
azo dyes (Grossmann et al. 2014). Generally, removal of dyes by microorganisms
takes place by biosorption which is accumulation of chemicals and dyes by microbial
mass (Hu, 1992; Bras et al. 2001) primarily by cell membranes and/or cell walls
through physical adsorption, electrostatic interaction, ion exchange, chelation and
chemical precipitation and the structure remains intact (Ali, 2010) but differs by
biomass type (Barr and Aust, 1992; Ali et al. 2007) and dye biosorption or
biodegradation by microorganism is judged by the colour of cell mat (Chen et al.
2003). This option is viable in cases when conditions are not favourable for growth
and maintainance of microbial population (Miao. Y, 2010) Biosorption of dyes does
not eradicate the problem because the pollutant is not destroyed but instead
entrapped into the matrix of the adsorbent (the microbial biomass). The disposal of
the microbial biomass containing adsorbed dyes itself is a big hurdle in their
proposed role in biocleaning of coloured waters (Chander and Arora, 2007).
Biosorption by living fungi has found be an efficient mechanism of dye
removal (Fu and Viraraghavan, 2001) and the biosorption of dyes may be of interest
in biorecovery of these synthetic chemicals from spent dye baths.
The bioremediation process is a pollution control technology (Shah et al.
2013) mediated by bioengineering the capabilities of intrinsic microorganisms
(bacteria, fungi and yeasts) which completely decolourise and mineralize the
pollutants on site from the dye laden effluent (Nachiyar et al. 2012). The advantages
of such a decolourisation and degradation (of azo dyes) process arise from the fact
that it is an aerobic treatment system where toxic intermediates like aromatic amines

Review of Literature 69
produced in the effluent by abiotic and biotic means are completely mineralized by
the bacteria themselves (Barsing et al. 2011). The ubiquitous nature of bacteria
makes them invaluable tools in effluent biotreatment (Olukanni et al, 2006).
Microorganisms having high growth rate in polluted environment and minimal
nutritional requirements can be used to achieve good result in decolourisation as well
as dye degradation experiments (Chang et al. 2001). Bacterial strains selected by
adaptation or under selective pressure of environmental pollution (Chen et al. 2003;
Madamwar and Keharia, 2003) from textile effluents have been shown to
decolourize textile dyes (Saratale et al. 2009; Hemapriya et al. 2010; Phugare et al.
2011; Shah et al. 2013) under favourable conditions to optimize it’s degradation
potential (Novotny et al. 2004; Lucas et al. 2008).
Biodegradation of dyes is a biologically mediated energy-dependent process
which leads to breakdown the breakdown of dye into various byproducts through the
action of various enzymes (Kaushik and Malik 2009). Biodegradation of synthetic
dyes not only results in decolourisation of the dyes but also in fragmentation of the
dye molecules into smaller and simpler parts (breakdown products). Decolourisation
of the dye occurs when the chromophoric center of the dye is cleaved (Kaushik and
Malik, 2009). Bacterial decolourisation is normally faster (Kalyani et al. 2009). It is
well known that bacteria degrade azo dyes reductively under anaerobic conditions to
colourless aromatic amines can be toxic, mutagenic or carcinogenic (Isik and
Sponza, 2007). The carcinogenicity of an azo dye may be due to the dye it’self or
aryl amine derivatives produced during the reductive biotransformation of an azo
linkage (Dawkar et al. 2009). These colourless aromatic amines should be degraded
further because these may be toxic, mutagenic, and carcinogenic to humans and
animals (Chen, 2006). Aromatic amines formed during anaerobic cleavage of the azo
dyes could be further degraded in an aerobic treatment system (Kuhad et al. 2004;
Van der Zee and Villaverde, 2005).
According to the concept of combined anaerobic–aerobic treatment (Carvalho et
al. 2008; Lin et al. 2010) azo dyes should be removed from the water phase by
(anaerobic) reduction in which a reduction of the bond in the molecules takes place
by hydroxylation followed by (aerobic) oxidation of the dyes constituent aromatic

Review of Literature 70
amines and complete mineralization of the reactive dye molecule takes place
(Lavanya et al. 2014). This treatment holds promise as a method to completely
remove azo dyes from wastewater (Van der Zee and Villaverde, 2005). Conversely,
aerobic treatment followed by anaerobic treatment of biological declourisation has
been reported as a viable alternative (Ong et al. 2005) as certain dyes are susceptible
to anoxic/anaerobic decolourisation. Under aerobic conditions, most of the azo dye
metabolites are quickly degraded by oxidation of the substituents or of the side
branches (Zaharia et al. 2012). Anaerobic reduction of the azo bond by bacteria
seems to be better suited for the decolourisation of azo dyes in treatment systems
(Amoozegar et al. 2011).
The putative advantages of this method are:
(1) The depletion of oxygen is easily accomplished in static cultures, which
enables anaerobic, facultative anaerobic and aerobic bacteria to reduce the
azo dyes.
(2) The reactions take place at neutral pH values and are expected to be
extremely unspecific when low molecular redox mediators are involved.
(3) The reduction rates generally increase in the presence of other carbon
sources.
The reduction equivalents that are formed during anaerobic oxidation of these
carbon sources are used finally for the reduction of the azo bond. The main
microorganisms contributing to biodegradation of organic compounds are bacteria
(e.g. Bacillus subtilis, Aeromonas Hydrophilia, Bacillus cereus, Klebsiella
pneunomoniae, Acetobacter liquefaciens, Pseudomonas species, Pagmentiphaga kullae,
Sphingomonas), fungi (e.g., white-rot fungi: Phanerochaete chrysosporium,
Hirschioporus larincinus, Inonotus hispidus, Phlebia tremellosa, Coriolus versicolour),
algae (e.g. Chlorella and oscillotoria species). However, in the presence of specific
oxygen-catalysed enzymes called azo reductases, some aerobic bacteria are able to
reduce azo compounds and produce aromatic amines (Stolz, 2001). The factors
which govern the process of biodegradation are summarized in Table No 10.

Review of Literature 71
Table 10: Factors responsible for dye degradation by microbial process (Khan
et al. 2012)
Factors Description
pH The pH has a major effect on the effciency of dye decolourisation the optimal pH for colour removal in bacteria is often between 6.0 and 10.0. The tolerance to high pH is important in particular for industrial processes using reactive azo dyes, which are usually performed under alkaline conditions. The pH has a major effect on the effciency of dye decolourisation, the optimal pH for colour removal in bacteria is often between 6.0 and 10.0 (Chen et al. 2003; Guo et al. 2007; Kilic et al. 2007).
Temperature Temperature is also again a very important factor for all processes associated with microbial vitality, including the remediation of water and soil. It was also observed that the decolourisation rate of azo dyes increases uptothe optimal temperature, and afterwards there is a marginal reduction in the decolourisation activity.
Dye concentration
Earlier reports show that increasing the dye concentration gradually decreases the decolourisation rate, probably due to the toxic effect of dyes with regard to the individual bacteria and/or inadequate biomass concentration, as well as blockage of active sites of azo reductase by dye molecules with different structures.
Carbon and Nitrogen sources
Dyes are deficient in carbon and nitrogen sources, and the biodegradation of dyes without any supplement of these sources is very difficult. Microbial cultures generally require complex organic sources, such as yeast extract, peptone, or a combination of complex organic sources and carbohydrates for dye decolourisation and degradation.
Oxygen and agitation
Environmental conditions can affect the azo dyes degradation and decolourisation process directly, depending on the reductive or oxidative status of the environment, and indirectly, influencing then microbial metabolism. It is assumed that under anaerobic conditions reductive enzyme activities are higher; however a small amount of oxygen is also required for the oxidative enzymes which are involved in the degradation of azo dyes.
Electron donor Dyes with simpler structures and low molecular weights exhibit higher rates of colour removal, whereas the removal rate is lower in the case of dyes with substitution of electron withdrawing groups such as SO3H, SO2NH2 in the para position of phenyl ring, relative to the azo bond and high molecular weight dyes.
Redox mediator
Redox mediators (RM) can enhance many reductive processes under
anaerobic conditions, including azo dye reduction

Review of Literature 72
Utilization of microorganism consortia offers considerable advantages over
the use of pure cultures in the degradation of synthetic dyes (Banat et al. 1996;
Pearce et al. 2003; Alves and Pereira, 2012). The individual strains may attack the
dye molecule at different positions or may use decomposition products produced by
another strain for further decomposition (Forgacs et al. 2004). Furthermore, mixed
culture studies may be more comparable to practical situations. With the increasing
complexity of a xenobiotic, one cannot expect to find complete catabolic pathways in
a single organism; a higher degree of biodegradation and even mineralization n1ight
be accomplished when co-metabolic activities within a microbial community
complement one another (Nigam et al. 1996; Khadijah et al. 2009).
Using mixed cultures instead of monocultures, higher degrees of
biodegradation and mineralization can be achieved due to synergistic metabolic
activities of the microbial community (Ramalho et al. 2004; Kehehra et al. 2005; Ali,
2010). The individual strains can attack dye molecules at different positions, yielding
metabolic end products that may be toxic; these can be further metabolised as
nutrient sources to carbon dioxide, ammonia and water by another strain. Other
species present may not be involved in bioremediation at all, but can stabilise the
overall ecosystem (Kandelbauer and Gilibitz, 2005). This type of mineralization is
the safest way to assure that no potentially harmful and unrecognized intermediate
degradation products are released into the environment. Mixed consortia usually do
not require sterile conditions and have greater stability towards changes in the
prevailing conditions (pH, temperature and feed composition) compared with pure
cultures (Ramalho et al. 2004). Therefore, the use of mixed cultures is a good
strategy for bioreactors.
A synergistic action (enzymatic) of fungal-bacterial consortium leads to the
enhanced degradation and detoxification of azo dyes and, thus provides an alternate
way for efficient removal of contaminants (Khelifi et al.2009b; Su et al.2009; Qu et
al.2010). Moreover the high rates of dye decolourisation by fungal-bacterial
synergism suggest it to be an appropriate powerful tool for the efficient degradation
and detoxification of azo dyes as well as textile effluent (Khelifi et al. 2009; Su et al.
2009; Qu et al. 2010; Kadam et al. 2011). Eco-friendly, efficient and short

Review of Literature 73
degradation times are some of the highlights of fungal-bacterial synergism over
individual cultures. Such synergisms are more effective due to the concerted
metabolic activities, which might attack dye molecules at various positions or utilize
intermediate degradation metabolites for further mineralization into non-toxic forms
(Keck et al. 2002; Chen and Chang, 2007). It is known that, addition of intermediate
metabolites of dye decolourizing culture into another culture could enhance the azo
dye decolourisation rates (Chang et al. 2004). Different consortial approaches have
been studied due to their enhancing degradation abilities. Table 11 represents dye
decolourisation and dye degradation studies by adapted microbes under optimized
process parameters.
2.7 Enzymes involved in biodegradation
Microorganisms can decolourize the dyes with different enzyme systems.
Fungal enzymes are non-specific towards different structures of dyes and thus
oxidize a wide range of them (Aust, 1990). Fungi have been extensively studied to
degrade textile dyes due to their extracellular oxidoreductive, nonspecific and non
stereoselective enzyme system, including lignin peroxidase, laccase, manganese
peroxidase and tyrosinase (Stolz, 2001; Hofrichter, 2002; Kaushik and Malik, 2009).
The bacterial biodegradation is associated with it’s intracellular and extracellular
oxidoreductive enzyme system such as azo reductase, DCIP-reductase and laccase
(Chen et al. 2003; Kalyani et al. 2008; Telke et al. 2009a).
Biochemically, Azoreductases [NAD(P)H: 1-(4`-sulfophenylazo)-2-naphthol
oxidoreductase] are the non-specific cytoplasmic enzymes that catalyzes or
biotransform (Ramlan et al. 2011) the reductive cleavage of the azo bridge (-N=N-)
in azo dyes to produce colourless amine products (Stolz, 2001; Ramlan et al. 2011)
more toxic than the parent compound (Kumar et al. 2006; Stingley et al. 2010;
Mendes et al. 2011a ;Ramchandran et al. 2013; Kolekar et al. 2013). These enzymes
are found to be more efficient under anoxic and static conditions.

Review of Literature 74
Table 11: Dye decolourisation and dye degradation studies by adapted microbes under optimized process parameters
S.
No
Source Organism Dye used OPTIMIZATION PARAMETERS Mechanism % Colour
Removal
Reference
pH Temperature
(◦C)
Duration Carbon Nitrogen Dye concentration
(mg/l)
1 Dye contaminated sludge
Pseudomonas aeruginosa Remazol Orange 3R (RO3R)
7 30 15 minutes
Glucose and starch
Yeast extract
50 Enzymatic 98 Surwase et al.2013
2 Textile effluent Bacillus licheniformis Reactive Red 2
9 37 7 days Glucose NH4NO3 50 Enzymatic 80 Balagurunathan and Sudha et
al.2013
3 Environmental treatment Plant
Pseudomonas otitidis Reactive Blue
250
7 37 8 hr. Glucose Yeast extract
100 Enzymatic 96.98 Bhatt et al.2012
4 Textile effluent Bacillus boroniphilus Reactive Yellow
145
7 30 24 hr. Glucose Yeast Extract
50-150 Enzymatic 100 Derle et al.,2012
5 Dye contaminated site
Bacterial-fungal consortium(Pseudomonas sp .SUK 1 and Aspergillus
Ochraceus NCIM-1168)
Rubine GFL 8 37 30 hr. Lactose Yeast extract
100 Enzymatic 95 Govindwar et al. 2012
6 Effluent disposal sites
Pseudomonas aeruginosa Mordant Black 17
7 37 48 hr. Glucose Ammonium nitrate
100 Enzymatic 86 Karunya et al.2013
7 Waste water treatment plant
Bacterial consortium
SpNb 1
Reactive Red M8B
7.5 37 48 hr. Lactose Yeast Extract
300 Enzymatic 96.75
Bhatt et al. ,2012
8 Tannery effluent Shigella sp Acid Red 113 7 37 72 hr. Dextrose L-methionine
200 - 99 Sivranjani et al.2013

Review of Literature 75
S.
No
Source Organism Dye used OPTIMIZATION PARAMETERS Mechanism % Colour
Removal
Reference
pH Temperature
(◦C)
Duration Carbon Nitrogen Dye concentration
(mg/l)
9 Textile effluent Bacillus sp –ETL 79 Crystal Violet 8 35 24 hr. Dextrose Peptone 100 Enzymatic 95 Shah et al.2013
10 National chemical laboratory, Pune
Micrococcus glutamicus NCIM 2168
Reactive Red 195
5 37 18 hr. Sucrose Yeast extract
100 Enzymatic 94.25 Sahasrabudhe et al.,2014
11 Grit tank of Tannery effluent treatment plant
Pseudomonas otitidis SA 1 CR 7 37 24 hr. - - 100 Biosorption 94 Saharan et al.2013
12 Sugarcane waste water
Bacillus sp SCWS 5 CR
7 35 16 hr. Sucrose Beef extract 150 Enzymatic 100 Pradhan et al.2012
14 Soil contaminated by textile effluent
Staphylococcus hominis Acid orange 7 35 60 hr. Glucose Yeast extract
100 Enzymatic 90 Singh et al.2014
15 Tannery division, Central Leather Research Institute, Chennai
Bacillus subtilis Acid blue 113 7 37 50 hr. Starch peptone 200 Enzymatic 90 Gurulakshmi et al.2008

Review of Literature 76
On the contrary, a report suggests, azoreductases obtained from different
microorganisms are known to have broad specificities in their enzymatic reactions
(Nakanishi et al. 2001) and variations can exist among the same organism (Ghosh et
al. 1992). Identification, purification and characterization of azoreductase constitute
a straightforward approach for the development of azo dye biodegradation systems
(Syed et al. 2009; Milikli and Rao, 2012; Bhatt et al. 2013).
Classifiying an azoreductase based on it’s primary amino acid level is
difficult due to low homology. However, a classification scheme based on the
secondary and tertiary amino acid analysis has been developed (Abraham, 2007).
Based on function, another classification scheme is used in which azoreductases are
categorized as either flavin-dependant azoreductases (Nakanishi et al. 2001; Chen et
al. 2004; Chen et al. 2005) or flavin-independent azoreductases (Blumel and Stolz,
2003). The flavin-dependent azoreductases are further organized into three groups;
(1) NADH only (Nakanishi et al. 2001; Chen et al. 2004), (2) NAD(P)H only (Chen
et al. 2005) or (3) both (Ghosh et al. 1992) (Figure20).
Figure 20: Classification of azoreductases based upon function
The reaction catalyzed by azo reductases occurs only in presence of reducing
equivalents like FADH and NADH. Most of the azo dyes have sulphonate substituent
groups possessing high molecular weight which does not pass through cell
membranes. Therefore, the reducing activity of the dye does not depend on the
intracellular uptake of the dye (Robinson et al. 2001). Russ et al. 2000 suggested that

Review of Literature 77
bacterial membranes are almost impermeable to flavin containing cofactors and,
therefore, the transfer of reduction equivalents by flavins from the cytoplasm to the
sulphonated azo dyes is restricted. Thus, a mechanism other than reduction by
reduced flavins formed by cytoplasmic flavin-dependent azoreductases must be
responsible for sulphonated azo dye reduction in bacterial cells with intact cell
membranes. One such mechanism involves the electron transport-linked reduction of
azo dyes in the extra-cellular environment. To achieve this, the bacteria must
establish a link between their intracellular electron transport systems and the high
molecular weight, azo dye molecules.
For such a link to be established, the electron transport components must be
localized in the outer membrane of the bacterial cells (in the case of gram-negative
bacteria), where they can make direct contact with either the azo dye substrate or a
redox mediator at the cell surface (Myers and Myers, 1992). In addition, low
molecular weight redox mediator compounds can act as electron shuttles between the
azo dye and an NADH dependent azo reductase that is situated in the outer
membrane. These mediator compounds owe their origin to the metabolism of certain
substrates by the bacteria or they may be added externally from some other source
(Russ et al.2000). Many proteins and enzymes are not naturally present in an
aqueous either outside or inside of cells. In gram negative bacteria, periplasmic
proteins are present which are partially immobilized between outer and inner
membranes of the cell envelope (Scopes, 2004). Kudlich et al. 1997 support the
suggestion that the membrane-bound azo reductase activity, mediated by redox
compounds, is different from the soluble cytoplasmic azo reductase that is
responsible for the reduction of non-sulphonated dyes that permeate through the cell
membrane. Therefore, the membrane-bound and the cytoplasmic azo reductases are
two different enzyme systems (Kudlich et al. 1997). Although the final reduction of
the azo dyes in the cell supernatants is a dominantly chemical redox reaction, the
redox mediators depend on cytoplasmic reducing enzymes to supply electrons (Yoo
et al. 2001). It is also possible that this chemical redox reaction works in conjunction
with a direct enzymatic reaction involving an azo reductase, which may be a

Review of Literature 78
dehydrogenase enzyme that is synthesized throughout the cytoplasm and secreted
without accumulation inside the cell (Bragger et al. 1997).
Depending upon oxygen utilization, there are two broad classes of
azoreductases; - the true azoreductases produced under aerobic conditions and those
that are produced under anaerobic conditions. Aerobic azoreductases catalyze
reductive metabolism of azo dyes in the presence of molecular oxygen. Previous
studies conducted by Zimmermann and co-workers in 1982 and 1984 on
Pseudomonas strains K22 and KF46 revealed two azoreductases capable of
decolourizing Orange I and II respectively. The enzymes were purified, characterized
and compared. It was found that both azoreductases were monomeric flavin-free
enzymes that preferentially used NAD(P)H and to some extent NADH as co-factors.
They also demonstrated the ability to reductively cleave sulfonated substrates in
addition to the carboxylated growth substrates. Azoreductases transfer the reducing
equivalents originating from the oxidation of organic substrates to the azo dyes.
Studies with some aerobic and facultative aerobic bacteria demonstrated that azo
compounds can be utilized as the sole source of carbon, with azo bond cleavage
being facilitated by enzymes that are thought to be either intracellular or membrane
bound (Van der Zee et al. 2002). These specifically adapted strains synthesize true
azoreductases which reductively cleave the azo group in the presence of molecular
oxygen.
Azoreductase activity has been identified in several bacteria, such as
Xenophilus azovorarts KF46 (Blumel et al. 2001), Pseudomonas luteola (Hu, 1998),
Rhodococcus (Heiss et al. 1992), Shigella dysenteriae Type I (Ghosh et al. 1992),
Klebsiella pnumoniae RS-13 (Wong and Yuen, 1996) and Clostridium perfringens
(Rafii et al. 1997). Cloning of genes encoding azoreductase were also carried out
from various bacteria, such as Geobacillus stearothermophilus 0Y1-2 (Ohnishi et al.
2011), Xenophilus azovorans KF46F (Blumel et al. 2002) and Escherichia coli
(Nakanishi et al. 2001). Recently, such genes were found in Bacillus anthracis (Read
et al.2003) and Bacillus cereus ATCC 10987 (Rasko et al. 2004). Because of their
high biodegradation capacity, they are of considerable biotechnological interest and
their application in the decolourisation process of wastewaters has been extensively
investigated (Young and Yu, 1997).

Review of Literature 79
Enzymes have a number of features that make them more viable in relation to
conventional catalysts: they are biodegradable catalysts, allow the operation at low
and high substrate concentrations, allow the operation over a wide range of pH,
temperature and salinity, have no delays associated with the acclimatization of
biomass, have a reduced sludge formation, are simple and easy to control (Nicell et
al. 1993). Another advantage of using pure enzymes instead of the microorganism is
that the expression of enzymes involved in dye degradation is not constant with time,
but dependent on the growth phase of the organisms, possess greater specificity,
better standardization, easy handle and storage (Roges-Milgarejo et al. 2006).
Conversely, Roges-Milgarejo et al .in 2006 was of an opinion that the expression of
an azoreductase was not dependent on bacterial growth rates and is influenced by
inhibitors that may be present in the effluent. Synthetic or natural redox mediators
have, to be added many times to the enzymatic bath in order to achieve the total
capacity of the enzyme(s) or even to make their work possible (Alves and Pereira,
2012). In order to increase the potential use of enzymes in a wastewater
bioremediation process, their immobilisation is recommended for biochemical
stability and reuse, thereby reducing the cost (Duran and Esposito, 2000;
Kandelbauer et al. 2004). The major drawback of using enzyme preparations is that
once the enzymes become inactivated, it is of no use. Because enzymes can be
inactivated by the presence of the other chemicals, it is likely that enzymatic
treatment will be most effective in streams that have the highest concentrations of
target contaminants but the lowest concentration of other compounds that could
interfere with their action (Alves and Pereira, 2012). The recent biotechnological
advances have allowed the production of cheaper and more available enzymes
through improved purification and isolation processes. All these advantages add to
their high specificities and catalytic activities with the possibility of designing
enzymes with the exactly desired properties through genetic engineering and
computational design, suggest the potential application of this process in the
treatment of effluents (Call and Mucke, 1997).
2.8 Genes behind the Azoreductase
Azoreductase activity in azo dyes decolourisation has been extensively
examined to elucidate azo dye reduction mechanism (Chen et al. 2005, Deller et al.

Review of Literature 80
2006, Wang et al. 2007, Ryan et al. 2010a, Ryan et al. 2010b and Feng et al. 2012).
Only few reports have reported the regulation of azoreductase gene expression
(Töwe et al. 2007, Liu et al. 2009a; Ryan et al. 2010a). Ryan et al. 2010a reported an
increase in mRNA levels for azoreductase genes (ppazoR1, ppazoR2 and ppazoR3)
from Pseudomonas aeruginosa in the presence of azo dyes. A significant increase in
azoreductase mRNA levels including azoR1 and azoR2 have been observed in B.
subtilis in the presence of quinines (Töwe et al. 2007). It was reported that azoR1 and
azoR2 are negatively regulated by redox-sensing transcription factors YodB and
YkvE, respectively (Töwe et al. 2007; Leelakriangsak et al. 2008). Azoreductases
AzoR1 and AzoR2 not only have azoreductase activity but also have quinone
reductase activity that plays a role in bacterial protection thiol-specific stress
(Nishiya and Yamamoto, 2007; Töwe et al. 2007, Leelakriangsak et al. 2008,
Leelakriangsak and Borisut, 2012). More recently, evidence was presented that
azoreductase possess quinone reductase and nitroreductase activity (Rafii and
Cerniglia, 1993, Liu et al. 2008a, Liu et al. 2009a). The flavin-dependent
azoreductases AZR, AzoR from Rhodobacter sphaeroides and E. coli, respectively,
overexpressed in E. coli have quinone reductase activity by reducing quinone
compounds as substrate. Moreover, the quinone compounds were better substrates
for AzoR than the model azo dye substrate Methyl Red (Liu et al. 2009a).
Interestingly, the presence of quinone compound accelerated the azo dye
decolourisation of overexpressed azoreductase AZR (Liu et al. 2009b; Parshetti et al.
2010). A significant increase in the enzyme activities of azoreductase and NADH-
DCIP reductase over a period of methyl orange decolourisation by K. rosea MTCC
1532 was observed. A similar result of an increase in azoreductase and DCIP
reductase activity was also observed when Alishewamella sp. KMK6 exposed to dyes
(Kolekar et al. 2013). A putative azoreductase gene (so3585) of Shewanella
oneidensis was found to up-regulate in response to heavy metal (Murgerfeld et al.
2009). However, the results were suggestive reduction of an azo dye is not the
primary function of the SO3585 protein in vivo. Figure 21 represents a proposed
catalytic cycle for an azoreductase enzyme.
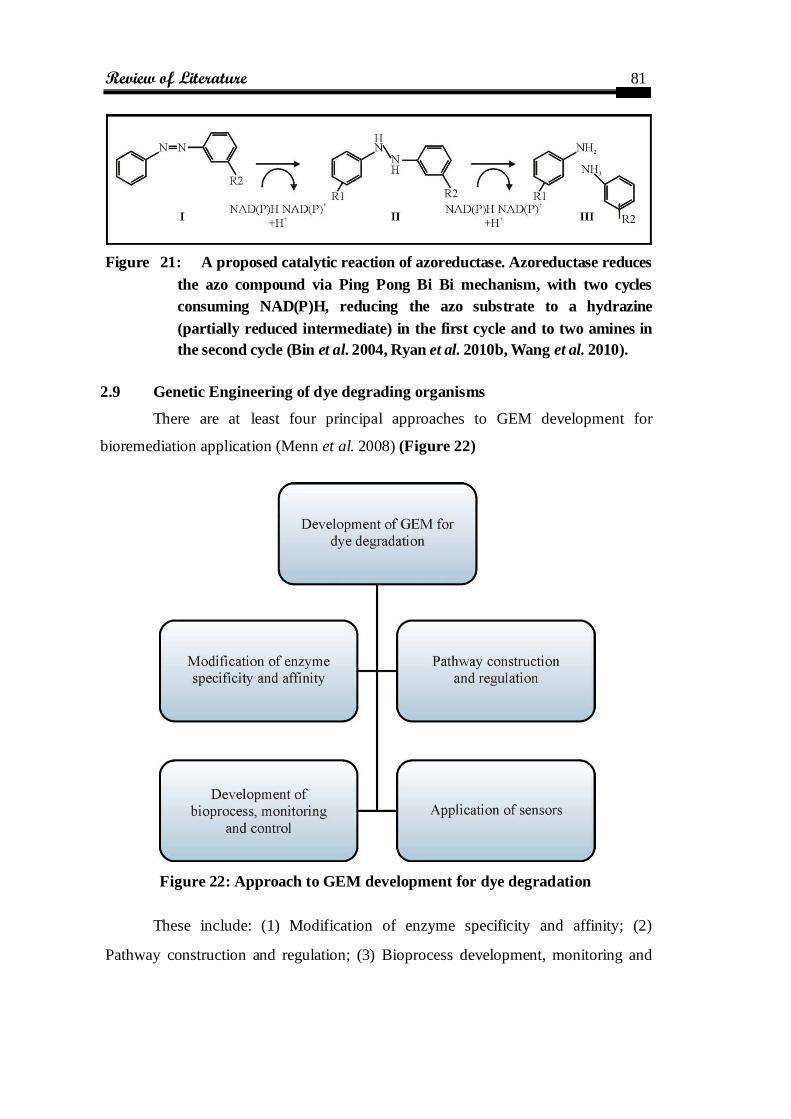
Review of Literature 81
Figure 21: A proposed catalytic reaction of azoreductase. Azoreductase reduces the azo compound via Ping Pong Bi Bi mechanism, with two cycles consuming NAD(P)H, reducing the azo substrate to a hydrazine (partially reduced intermediate) in the first cycle and to two amines in the second cycle (Bin et al. 2004, Ryan et al. 2010b, Wang et al. 2010).
2.9 Genetic Engineering of dye degrading organisms
There are at least four principal approaches to GEM development for
bioremediation application (Menn et al. 2008) (Figure 22)
Figure 22: Approach to GEM development for dye degradation
These include: (1) Modification of enzyme specificity and affinity; (2)
Pathway construction and regulation; (3) Bioprocess development, monitoring and

Review of Literature 82
control; (4) Bioaffinity, bioreporter sensor applications for chemical sensing, toxicity
reduction and end point analysis.
Identification, isolation, and transfer of genes encoding degradative enzymes
can greatly help in designing microbes with enhanced degradation capabilities
known as super degraders. Of the two approaches, acclimatization is natural, since
in this case the built-in genetic setup of the microorganism is not disturbed; only
some components are enabled. On the other hand, in genetic engineering, the natural
genetic set-up of a microorganism is changed by incorporating a new gene or genes.
By cloning and transferring genes encoding for dye degrading enzymes, organisms
could be designed that combine the abilities of mixed cultures within a single
species. Molecular cloning of the gene encoding azoreductase enzyme followed by
protein purification is likely to be crucial for further characterization and application
of this enzyme (Suzuki et al. 2001; Wang et al. 2007; Ryan et al. 2010a; Wang et al.
2010; Mendes et al. 2011a). A number of genes conferring the ability of dye
decolourising have been identified. Successful decolourisation of an azo dye using
Escherichia coli carrying the azoreductase gene from a wild-type Pseudomonas
luteola has been reported (Chang et al. 2000; Chang and Kuo, 2000). This approach
could become a useful alternative for shortening the extended time-periods otherwise
needed to adapt appropriate cultures and isolated strains, respectively. CotA-laccase,
a bacterial enzyme from Bacillus subtilis cloned and over-expressed in E. coli, has
proved to be a thermoactive and intrinsically thermostable enzyme with a high
capacity for the decolourisation of azo and anthraquinonic dyes (Pereira et al. 2009a,
b). The expression level of CotA-laccases in different E. coli host strains, growing
under different culture conditions, was compared and a high-throughput screenings
for the oxidation of dyes with high potential redox developed by (Brissos et al.
2009). Chen et al. 2010 described the cloning of azoreductase gene azoB from
Pigmentiphaga kullae K24. The recombinant azoreductase expressed in E. coli
exhibited optimal for activity of Orange I at pH 6.0 at temperatures between 37 and
45°C. Both NADH and NAD(P)H can be used as an electron donor but NAD(P)H is
preferred. The gene was characterized (Ooi et al. 2007). The recombinant
azoreductase expressed in E. coli exhibitted broad pH stability between 6 and 10

Review of Literature 83
with an optimal temperature of 60-80°C. AzrA effectively decolourized Methyl Red,
Orange I, Orange II and Red 88. No enzyme activity was detected for Orange G and
New Coccin. In addition, the enzyme activity of AzrA was oxygen insensitive and
required NADH as electron donor for dye reduction. Similar results have also been
described for azoreductase enzyme activity extracted from B. velezensis and P.
aeruginosa (Nachiyar and Rajakumar, 2005, Bafana et al. 2008). Furthermore, a
gene encoding NAD(P)H-flavin azoreductase (Azo1) from the skin bacterium
Staphylococcus aureus ATCC 25923 overexpressed in E. coli demonstrated that this
azoreductase is able to decolourize a wide range of structurally complex azo dyes
(Chen et al. 2005). The Azo1 cleaved the model azo dye Methyl Red and sulfonated
azo dyes Orange II, Amaranth and Ponceau. However, no enzyme activity was
observed when Orange G was used as substrate. Recently, the gene encoding an
FMN dependent NADH azoreductase AzrG from thermophilic Geobacillus
stearothermophilus was cloned and expressed in recombinant E. coli (Matsumoto et
al. 2010). The optimal temperature of AzrG was 85°C for Methyl Red degradation
and enzyme also showed a wide range of degrading activity towards several
tenacious azo dyes such as Acid Red 88 Orange I and CR.
Physiochemical properties, enzyme characterization and kinetic studies can
be investigated by obtaining purified azoreductase from whole cell extract from the
source organism or recombinant cell extract (Nachiyar and Rajakumar, 2005; Wang
et al. 2007; Gopinath et al. 2009; Punj and John, 2009; Mendes et al. 2011a;
Morrison et al. 2012). Purification from whole cell extracts from the source organism
employs classical purification procedures which require many steps such as
ammonium sulfate precipitation followed by ion exchange and affinity
chromatography methods (Maier et al. 2004; Nachiyar and Rajakumar, 2005; Punj
and John, 2009; Kolekar et al. 2013). However, in most cases recombinant DNA
techniques permit the construction of fusion proteins in which specific affinity tags
are added to the protein sequence of interest (Bin et al. 2004; Wang et al. 2007).
Therefore, the purification of the recombinant fusion proteins is simplified by
employing affinity chromatography methods. In addition, the expression and
purification of recombinant proteins facilitate the production and detailed

Review of Literature 84
characterization of virtually any protein. Native molecular weight of a protein can be
determined by native gel electrophoresis and/or size exclusion chromatography
(Moutaouakkil et al. 2003; Deller et al. 2006; Ooi et al. 2007; Wang et al. 2007).
The purification and characterization experiments of enzymes were conducted and
the results indicated that the enzyme activity differs in substrate specificity and
preferential coenzymes serving as electron donors. Concludingly, characterization of
recombinant azoreductases provide information for understanding these azoreductases
properties such as enzyme stability and activity, kinetic constants, cofactor
requirement, substrate profile, structure and mechanism (Wang et al. 2007, Ooi et al.
2009, Macwana et al. 2010, Ryan et al. 2010b, Mendes et al. 2011a). A broad range
of substrate specificity and thermostability are important factors in determining the
range of biologically degradable of azo dyes. Table 12 represents summarized
information of cloning and expression of recombinant azoreductase in Escherichia
coli.
Table 12: Cloning and overexpression of azoreductases (Leelakriangsak, 2013)
Organism Gene Expression vector
Molecular weight
Co-factor Reference
Pseudomonas putida MET94
ppAzoR pET-21a Homodimer 40 kDa
FMN, NAD(P)H
Mendes et al. 2011a
Pseudomonas aeruginosa
paazor1 paazor2 paazor3
pET-28b pET-28b pET-28b
Tetramer 110 kDa 23 kDa* 26 kDa*
FMN, NAD(P) H NADH NADH
Wang et al. 2007 Ryan et al. 2010b Ryan et al. 2010b
Pigmentiphaga kullae K24
azoB pET-11a Monomer 22 kDa
NAD(P)H Chen et al. 2010
Geobacillus stearothermophilus
azrG pET-3a Homodimer 23 kDa
FMN, NADH
Matsumoto et al. 2010
Bacillus sp. B29 azrA azrB azrC
pET-3a pET-3a pET-3a
Homodimer 48 kDa Homodimer 48 kDa Homodimer 48 kDa
FMN, NADH FMN, NADH FMN, NADH
Ooi et al. 2007 Ooi et al. 2009 Ooi et al. 2009
Bacillus subtilis yvaB, (azoR2)
p-Bluescript Homodimer 45 kDa
NADH Nishiya and Yamamoto, 2007

Review of Literature 85
Organism Gene Expression vector
Molecular weight
Co-factor Reference
Escherichia coli acpD (azo R)
pET-22b Homodimer 46 kDa
FMN, NADH
Nakanishi et al. 2001
Enterococcus faecium
acpD pET-15b 23 kDa* NAD(P)H Macwana et al. 2010
Enterococcus faecalis
azoA PET-11a Homodimer 43 kDa
FMN, NADH
Chen et al. 2004
Xenophilus azovorans KF46F
azoB pET-11a Monomer 30 kDa
NAD(P)H Blumel et al. 2002
Clostridium perfringens
azoC pET-15b Tetramer 90.4 kDa
FAD, NADH
Morrison et al. 2012
*Molecular weight determined by SDS-PAGE
2.10 Confirmatory studies on dye degradation
Dyes, as colourants, absorb in the visible region of the spectra and each one
has a maximal wavelength, depending on it’s visible colour therefore, the easiest way
to monitor dye degradation is by means of spectrophotometry, following the decrease
in it’s absorbance. By this technique, all the molecules present are quantified, and
intermediates and degradation products will contribute to the spectra absorbance.
Various basic and advanced instrumental techniques of chromatography such as Gas
Chromatography (GC), High Performance Liquid Chromatography (HPLC), Nuclear
Magnetic Resonance Spectroscopy (NMR), Mass Spectrometry (Ion-trap, MALDI)
and Capillary Electrophoresis (CE) are available to assist in the isolation and
characterization of the intermediates and products of dye degradation, thereby giving
new insight into the mechanism of biodegradation. Prior procedures of extraction of
the aqueous sample with an organic solvent or filtration are adopted when a
heterogeneous catalyst or solid reactant is employed, or when a pre-separation is
needed (Pereira and Alves, 2012).
Shah, 2014 conducted the study to investigate the decolourisation and
degradation of azo dyes using bacteria isolated from textile dye effluent. Three
different bacterial species were isolated and the isolates were identified as Bacillus
subtilis, Pseudomonas aeruginosa, and Psuedomonas putida. Decolourisation was
attained in the range from 65%-95% with 500 mg/l of dye amended with trace

Review of Literature 86
amounts of yeast extract, glucose and sucrose and then The degradation product after
decolourisation was examined by TLC and FTIR.
Sahasrabhude et al. 2014 studied the effect of Micrococcus glutamicus NCIM
2168 on degradation of Reactive red 195 and exhibited a tremendous decolourisation
potential of 94.25% in 18 hr. under static conditions. Decolourisation of dye was
attainable over a wide range of pH from 5-8 and in a narrow temperature from 37-
40°C. Effect of various carbon and nitrogen sources on decolourisation revealed
enhanced decolourisation in presence of sucrose followed by glucose, yeast extract
and peptone. Degradation of the dye was confirmed by UV-Vis spectroscopy, TLC,
HPLC and GC-MS.
Surwase et al. 2013 studied the biotransformation Remazol Orange 3R
(RO3R) by Pseudomonas aeruginosa strain BCH. 98 % decolourisation was attained
within 15 min. The RO3R was transformed to the N-(7 amino 8 hydroxynapthalen-
2yl) actamide (m/z, 198), Acetamide (m/z, 59) and Napthalen-1-ol (m/z, 144).
Pseudomonas sp. has been a focal point in many research investigations
pertaining to dye degradative studies. An effluent adapted Pseudomonas sp was
studied for decolourisation and degradation of azo dye and decolourisation attainable
was 98% at 50mg/L within five hr. in static anoxic condition. The optimum pH and
temperature for the decolourisation was 8.0 & 37°C respectively. The
biodegradation was monitored by FTIR analysis (Shah et al. 2013). Another study
was carried out to explore the decolourisation and degradation of Direct Blue 71 by
Pseudomonas aeruginosa. The bacterium was able to decolourize the dye medium to
70.43 % within 48 h under microaerophilic conditions. The degradation metabolites
formed were studied using UV–Vis techniques, HPLC, Fourier Transform Infra Red
(FTIR) spectroscopy and nuclear magnetic resonance spectroscopy analysis. Data
obtained provide evidence for the formation of aromatic amines and their subsequent
oxidative biodegradation (Hafshejani et al. 2013).
Bhatt and co-workers in 2012 screened bacteria for their dye decolourising
activity and the most promising bacterial isolate was used for further dye degradation
studies. The strain showed complete decolourisation of the selected dye (RB 250-100

Review of Literature 87
mg/L) within 8 hr in static condition. The optimum pH, temperature, inoculum size
and carbon and nitrogen sources for the decolourisation was studied at pH 7.0, 37°C,
glucose (0.2 %) and nitrogen (0.5 %) respectively. The biodegradation was
monitored by UV-Vis, HPTLC and FTIR analysis.
A bacterial isolate Bacillus licheniformis was able to degrade Reactive Red 2
dye, optimally at pH 9, temperature at 37°C, dye concentration of 50 mg/l at 20%
inoculum size. Glucose, NH4NO3 were found to be the best additional carbon and
nitrogen sources. The extracellular enzyme from Bacillus licheniformis was studied
for dye decolourisation potential. Biodegradation was confirmed by analyzing the
product using TLC an GC-MS analysis indicated the formation of 2, 4-dichloro-6-
[(1H-indazol-5-ylimino)-methyl]-phenol, benzene sulfonamide, 1H indole and urea
as final metabolites formed by Bacillus licheniformis (Sudha and Balgurunathan,
2013).
Derle et al.in 2012 isolated strain of Bacillus boroniphilus which showed
appreciable ability of decolourisation of Reactive Yellow 145 dyes and exhibited
maximum decolourisation 98% in static condition and supplemented with urea and
yeast extract. Further, biodegradation of azo dye was analyzed by TLC, UV-Vis
spectrophotometry and FTIR, results showed that -N=N- (azo bond) get converted
into –NH2 (amino group), which proves accomplishment of biodegradation of
Reactive Yellow 145.
The marine ecofriendly bacteria Lysobacter sp. T312D9 isolated from Abou
Quir Gulf, Alexandria, Egypt represent an inexpensive and promising marine
bacterium for removal of both methyl and CR (Barakat, 2012). The biodegradation
was analyzed by (GC-MS) analysis and FTIR. In another study, biodegradation of
CR by Fusarium sp TSF-01 was monitored by FTIR and generation of new peaks
other than the parent compound confirmed biodegradtion of the dye by fungus
(Shinde and Thorat, 2013).
GC-MS has been widely used to identify products of dyes degraded with
bacterial monoculture and consortia both in immobilized and non-immobilized forms
in textile effluents (Soundararajan et al. 2012).

Review of Literature 88
Not only does textile effluents or soil contaminated by dyes had proved to the
richest source dye degraders, tannery effluent does house some efficient dye
degrading bacteria belonging to Bacillus cereus, the decolourisation potential of
which has been well established for the Acid Black azo dye. The optimum activity of
Bacillus cereus was found at pH 7.3, temperature 37ºC, and duration of 4 days. The
decolourisation rate was found out to be 80% and 96% for the agitated and static
conditions. UV–Vis spectra analysis showed that the peaks which appeared in the
visible region of the treated dye disappeared, indicating complete decolourisation of
dye. The FTIR analysis for the Bacillus cereus treated sample showed the
transformation of azo linkage into N2 or NH3 or incorporated into complete biomass
(Kanagraj, 2011).
Bhimani, 2011 carried out a study on biodegradation of azo dyes (Reactive
Black 5 and Green B) by bacterial strains isolated from textile effluent obtained from
Jetpur, Gujarat. The biodegradation of azo dyes was preliminarily investigated by
TLC and further confirmed by (HPLC).
In another study aimed at monitoring biodegradtion of reactive dyes, bacteria
were screened for their biodegradative potential of commonly used textile dyes like
Reactive Lanasol Black B (RLB), Eriochrome Red B (RN) and 1, 2 metal complexes
I. Yellow (SGL) and an appreciable decolourisation ranging from 57%-100% was
attained in 15 hr. The biodegradation products of RLB formed during anaerobic and
sequential anaerobic/aerobic treatments were analyzed by HPLC. Peaks at different
retention times were observed in the anaerobic stage, and these peaks completely
disappeared at the end of anaerobic/aerobic incubation. This result clearly indicates
that the dye had been catabolized and utilized by isolates (Pahlaviani et al.2011).
Actinomycetes have also exhibited a tremendous potential to decolourise dye
stuffs. Streptomyces krainskii, SUK -5 was found to decolourize and degrade textile
dye Reactive blue–59.This azo dye was decolourized and degraded completely by
Streptomyces krainskii SUK–5 at 24 hour in shaking condition in the nutrient
medium at pH 8. Induction in the activity of Lignin Peroxidase (LiP) and NADH-
DCIP Reductase and MR reductase represents their role in degradation.

Review of Literature 89
2.11 Colour removal in Real Dye Waste Water (RDWW): An approach to
combat dye influxed wastewaters
RDWW not only comprises of dyes, but is also influxed with surfactants,
salts, chelators, precursors. Despite of immense decolourisation efficacy of microbial
strains, colour removal in RDWW remains a challenging task (Wesenberg et al.
2003). Bioremediation systems were commonly applied to treat industrial effluents
contaminated with different types of synthetic dyes by bacteria, fungi, yeast and
actinomycetes respectively by bioaccumulation (Ramchandran et al. 2013).
Bioaccumulation of chemicals/ pollutants process takes place by respiration or food
intake wherein the concentration of target pollutant is more in vivo than the external
environment (Mebrat, 2006). Fungal strains are preferred over bacterial counterparts
for remediating RDWW as they are capable of growing over a wide range of pH and
can resist dye toxicity even at high concentrations (Kaushik and Malik, 2009).
Treatment of RDWW collected from Tirupur and Erode districts of Tamil Nadu were
treated by dye degrading bacteria in monocultures and consortial forms (Mohan et al.
2013). A similar approach had been adopted to treat textile effluent collected from
CETP of Jetpur, Gujrat by Lysinibacillus fusiformis JTP-23 utilising a sequential static
and agitating incubation (Bhimani, 2011). Exiguobacterium sp. RD3 isolated from dye
contaminated site had be known to degrade textile effluent (Dhanve et al. 2014).
Exposing the target organism prior it’s degradation mechanism has been exemplified
has been a theme of research interest (Jalandoni –Buan et al. 2010). Likewise, a
similar approach to study biodegradation of RDWW, a comparison between effluent
adapted and effluent non-adapted strains has been established (Leena and Selva Raj,
2008).
Studies based on treating the effluents by industrial scale bioreactors have
also been stated in literature (Ali, 2010; Devassy, 2010). To monitor the exclusive
effect of fungal-bacterial consortium on biodegradation of RDWW, the approach
based at sterilizing the effluent. This strategy overcomes the inclusive effect of
residential microflora on biodegradation of real dye wastewater (Lade et al. 2012).