retinoids reprogramme pre-bud mesenchyme to give …biology.hunter.cuny.edu/cellbio/feinstein cell...
TRANSCRIPT

Development 100, 723-733 (1987)Printed in Great Britain © The Company of Biologists Limited 1987
723
Retinoids reprogramme pre-bud mesenchyme to give changes
in limb pattern
SUSAN M. WILDE, S. E. WEDDEN* and C. TICKLE
Department of Anatomy & Biology as applied to Medicine, The Middlesex Hospital Medical School, Cleveland St, London Wl P 6DB, UK
* Present address: Department of Physiology and Biophysics, Harvard Medical School, 25 Shattuck St, Boston, USA
Summary
Retinoic acid was locally applied to presumptive limbregions of chick embryos to find out the earliest timeat which the limb pattern can be reprogrammed.When beads soaked in retinoic acid were placed in theappropriate positions in embryos at stage 10 or older,duplicated or reduced leg patterns resulted. To pinpoint the time at which the cells in the limb rudimentrespond to the retinoid, beads were removed atvarious times and the lengths of exposure required toreprogramme limb development found. The earlylimb rudiments require longer exposures to give
duplications than late rudiments. The effective treat-ment periods last at least until stage 17 when the limbbud and apical ectodermal ridge develop. In contrast,the length of exposure to reduce the limb is constantat early stages. Retinoids first start acting to produceduplicated structures between stages 10 and 13.Therefore, retinoids appear to begin to reprogrammethe cells as soon as they are determined to give rise toa limb.
Key words: limb development, retinoids, patternformation, chick embryo.
Introduction
Experimental manipulations of developing chicklimbs can bring about changes in the pattern ofcellular differentiation across the anteroposterior axis(the axis running across the limb, for example, in thehuman hand from thumb to little finger). When tissuefrom the posterior margin of a chick wing bud (thepolarizing region) is grafted to the anterior margin ofanother bud, duplicated patterns result (Saunders &Gasseling, 1968; Tickle, Summerbell & Wolpert,1975). For example, six digits may develop instead ofthree; an additional set of digits 4 3 2 develops inmirror-image symmetry with the normal set 2 3 4.Local application of all-fram-retinoic acid to theanterior margin of chick wing buds has been foundto mimic the signalling of polarizing region grafts(Tickle, Alberts, Wolpert & Lee, 1982; Summerbell,1983). With appropriate concentrations of the reti-noid, mirror-image symmetrical wing patterns de-velop (Tickle, Lee & Eichele, 1985).
The changes in the pattern of cellular differen-tiation just described result from manipulations whenthe bud is already formed. The early bud consists of a
population of apparently homogeneous mesenchymecells encased in ectoderm which has a thickened rim,the apical ectodermal ridge. However, the cells thatwill form the limb appear to be set aside much earlierin development. Tissues from the appropriate lo-cation along the body axis from chick embryos asearly as stage 9, when neither bud nor thickenedepithelium are present, will develop into leg struc-tures when grafted to the coelomic cavity (Rudnick,1945).
It is possible that the signals that order the patternof cellular differentiation across the anteroposterior(a-p) axis operate before the definitive bud develops.Cells with detectable polarizing activity have beenfound in regions that will give rise to legs in embryosas early as stage 10 (Hornbruch & Wolpert, 1986).Alternatively, signalling may be required throughoutlimb development and may be linked to the pattern-forming process along the proximodistal (p-d) axis(running from the shoulder to the tips of the digits).The laying down of pattern along this axis is co-ordinated with bud outgrowth and depends on theapical ectodermal ridge (Saunders, 1948; Summer-bell, 1974). To gain insight into this problem, we have

724 S. M. Wilde, S. E. Wedden and C. Tickle
investigated whether retinoic acid can affect thepattern of cellular differentiation when applied topresumptive limb tissue before buds have developed.
Retinoids were applied to leg rudiments becausewe feared that application to the wing region of earlyembryos, near the developing heart, might causeembryo death (Jelinek & Kistler, 1981). We havefound that a duplicated leg can result when embryosas early as stage 10 (10-somite stage) are treated withretinoic acid. The length of exposure to retinoic acidrequired to cause duplication in a limb skeleton issimilar at stage 14 (22-somite stage) to the exposurerequired at a bud stage (20) (Eichele, Tickle &Alberts, 1985). However, embryos treated at stages10 and 13 require longer exposures in order to induceduplicated structures in the leg. These results arediscussed in relation to the timing of limb rudimentdetermination and the mechanism by which retinoidschange limb pattern and its relation to normal polar-izing activity.
Methods
EmbryosFertilized hen eggs (Needle Farm, Herts) were incubated at38 °C. The eggs were windowed on days 2 and 3 and theembryos were staged according to Hamburger & Hamilton(1951).
Application of retinoidsAG1-X2 beads (in formate form: Biorad) were used toapply all-fra/w-retinoic acid (Sigma batch No. 104F.0135) tothe leg buds or to regions of the flank where leg buds willdevelop. For stage-20 embryos, beads of 200,um diameterwere used; for earlier embryos the beads were 100fim indiameter. The beads were soaked in solutions of a\\-trans-retinoic acid dissolved in DMSO, rinsed in tissue culturemedium (see Tickle et al. 1985 for details of the procedure)and then implanted or positioned in the appropriate lo-cation in the embryo. In a short series of experiments,beads were soaked in solutions of a relatively stableanalogue, (E)-4-[2-(5,6,7,8-tetrahydro-5,5,8,8-tetramethyl-2-napthalenyl)-l-propenyl]-benzoicacid (TTNPB, Ro: 13-7410, a gift from Hoffman-LaRoche, Switzerland). Ascontrols, beads were taken through the same procedure butwere soaked in DMSO only.
Implantation of beads to leg buds of stage-20embryosThe extraembryonic membranes were removed and a slitwas made between the apical ectodermal ridge (AER) andthe anterior margin or the apex of the right leg bud. Thebead was placed between the free AER and the rest of thebud such that the bead was held in place. The procedure isdescribed in full by Tickle et al. (1985).
Implantation of beads to leg regions of stage-14 to-16 embryosThe vitelline membrane was removed and the bead wasplaced in a small hole made in the right flank using atungsten wire needle. Depending on the stage of theembryo, the hole was made in either somites 23-30 or in aposition in the unsegmented lateral plate where thesesomites will shortly form. The leg bud is first recognized as aswelling in the flank opposite somites 26-32 at stage 17. Itshould be noted that in the bead removal experimentsbelow, beads were placed on the vitelline membrane overstage-14 embryos.
Application of retinoids to stage 8-13 embryosThe bead was placed on the vitelline membrane over theright lateral plate in a position where the leg bud will laterform. The positioning of the bead was determined by aseries of preliminary experiments in which beads wereplaced in various positions along the lateral plate. Thepositions were recorded as the distance from the tail incamera-lucida drawings. In a few experiments, the beadwas placed in the region where the wing will develop.
Effective length of exposure to retinoidsTo find the length of exposure to retinoic acid required toaffect the development of the leg pattern, beads wereremoved at various intervals after being placed on thevitelline membrane over the presumptive leg regions instage-10 to -14 embryos. In control embryos, the beadswere not removed.
In all experiments, following application of the retinoid,the eggs were resealed and incubated until the embryoswere 10 days old.
Observation of the embryos following beadimplantationFollowing positioning of the bead in early embryos, acamera-lucida drawing was made before the egg wasresealed and returned to the incubator. The next day, theposition of the bead in relation to the early limb bud wasnoted.
Staining the embryos to show the pattern of cartilagedifferentiationOn the 10th day of incubation, the embryos were fixed in5% TCA, stained in 0-1% Alcian green in 70% acidalcohol, dehydrated in ethanol and then cleared in methylsalicylate.
Scanning electron microscopyIn a set of experiments to investigate the effect of retinoicacid on bud formation, 100^m beads soaked in 5mgml~'retinoic acid were placed on the vitelline membrane overthe leg region of embryos staged 10-13. The embryos werefixed 24-48 h later in half-strength Karnovsky fixative(Karnovsky, 1965) for 24 h at 4°C. They were washed in0-1 M-cacodylate buffer, placed in osmium tetroxide diluted1:1 with 0-2 M-cacodylate buffer for lh, rinsed in 0 1 M -cacodylate buffer and then dehydrated in ethanol. Theethanol was replaced with amyl acetate and the embryos

Retinoids reprogramme pre-bud limb mesenchyme 725
Table 1. The skeletal patterns that develop following implantation of beads soaked in all-trans-retinoic acidbeneath the apical ridge of leg buds of stage-20 embryos
Concentration
in which beadswere soaked(mgml"1)
Totalnumber
Bead implanted
% duplicateddigit
pattern
anteriorly
% reduceddigit
pattern
Meannumber of
digitsTotal
number
Bead implanted
% duplicated %digit
pattern
at apex
•> reduceddigit
pattern
Meannumber of
digits
0 11
10
679
1008667
01433
5-33-93-4
28
—
1000
—
0100—
20
—
were critical-point dried. The early leg buds were studiedusing a Jeol JSM-35 scanning electron microscope.
Results
Local application of retinoic acid to leg budsBeads soaked in retinoic acid were implanted to theleg buds of stage-20 embryos. These experimentswere done to provide data with which to compare theeffects of retinoids applied before the leg bud hasdeveloped. Both the dose of retinoid and the positionof application were varied. The results are summar-ized in Table 1.
When 200^m beads soaked in 0-1 mgml"1 retinoicacid were implanted at the anterior margin of the legbud, leg patterns were obtained with additional digitsin mirror-image symmetry to the normal set. Thepattern of digits in the majority of cases was III III IIIII IV or III HI IIII HI IV (the normal pattern is IIIHI IV, anterior to posterior, see Fig. 2A). Whenbeads soaked in higher concentrations were im-planted anteriorly, the number of digits progressivelydecreased. This was due to the loss of anterior digitseither in duplicated patterns (IV III HI IV, III III IIIIV and IV HI IV) or in reductions (II III IV and IIIIV). With the highest concentration of retinoic acidused, a duplicated fibula was often found in theforeleg and the tibia was missing. Local application ofretinoic acid to the apex of the leg bud led toduplicated patterns at low doses (e.g. IVIV and IIII)and complete truncations of the leg at higher doses.
Local application of retinoids to presumptive legregionsSmall 100 jiim beads soaked in retinoic acid wereimplanted to the somites or the unsegmented lateralplate, or were placed on the vitelline membrane, in,or anterior to, the presumptive leg regions of em-bryos ranging from stages 8-16. For the earlierstages, a series of pilot experiments showed that thebead must be placed on the membrane 1 mm from thetail to end up in the region of the leg bud. The aim ofthis series of experiments was to find the earliest time
at which beads soaked in the retinoid could lead tothe development of duplicated leg structures. Theresults for beads soaked in 5 mg ml"' retinoic acid areshown in Fig. 1.
When beads soaked in 5 mg ml"' retinoic acid wereplaced on stage-8/9 embryos, the embryos usuallydied. The legs of surviving embryos were normal.However, in one case (out of two) when beads soakedin the relatively stable retinoid TTNPB (0-005 mgmP1) were placed on stage-9 embryos, a leg with asingle additional digit (I III HI IV) resulted. Whenbeads soaked in 5mgml~' retinoic acid were placedin the appropriate position in embryos at stage 10,duplicated leg patterns developed (see Fig. 2). How-ever, many of the legs of embryos treated at this stagewere normal or had missing structures. Some had noleg at all. At stages 11-14, beads soaked in the sameconcentration of retinoid were more successful inproducing duplications and the percentage of normalor reduced/missing patterns fell (Fig. 1). Retinoidtreatment at stages 10-14 could also cause the loss ofthe tail and/or vertebrae. Apart from treatment at
100
•c 4 0
o
I 30
$ 20
10
Stage 8/9n = 5
10 11/12 13 14 15 16n = 42 n=17 n = 23 n = 2l n=16 n = 8
Fig. 1. A histogram showing the incidence of the classesof leg patterns resulting from experiments in which beadssoaked in 5mgml"1 all-www-retinoic acid were placed inpresumptive leg regions of embryos at a range ofdevelopmental stages 8-16. n = number of cases thatsurvived to 10 days. D, normal pattern; M, reduced ormissing; S, duplicated.

726 S. M. Wilde, S. E. Wedden and C. Tickle
2A
DFig. 2. Leg patterns resulting from placing beads soaked in 5 mgml ] all-/raw-retinoic acid on the vitelline membraneabove the leg rudiments of stage-10 embryos. (A) Normal pattern of digits: I I I III IV. Control bead soaked in DMSO.(B) Duplicated pattern of digits: IV III IV; tibia is absent. Bead at anterior of bud at stage 17. Note absence of tail andcaudal vertebrae. (C) Reduced pattern of digits: III IV; two fibulae in fore-leg. Bead anterior to bud at stage 17. Noteabsence of tail and caudal vertebrae. (D) Leg missing. Bead anterior to bud at stage 17. Note short tail.
stage 8/9, the survival rate of treated embryos wasmore than 80%.
Although the incidence of leg duplications gener-ally increased with stage at treatment, the quality ofthe duplications did not vary. Legs with an additionaldigit IV (the most posterior digit) could develop afterapplication of a bead soaked in 5 mg ml"1 at all stagesand the maximum number of digits usually attained
(6) was also the same irrespective of stage at treat-ment (see Fig. 4). Fig. 3 shows examples of dupli-cations following retinoid application to embryosbetween stages 13 and 20.
It should be noted that the greatest duplicatingeffect on the pattern was found in the digits. Theforeleg could be duplicated but no more than threeelements (fibula, tibia, fibula) (Fig. 3B) instead of the

Retinoids reprogramme pre-bud limb mesenchyme 727
normal two (tibia, fibula) were ever formed. Theforeleg elements could both be fibulae. However,frequently only one foreleg element, the fibula, waspresent (Fig. 2A). In no case was the femur dupli-cated even when the retinoid was applied at stage 10.
To determine whether the mixture of duplicationsand reduced leg patterns obtained at the earliest
Fig. 3. Duplicated leg patterns resulting from treatingstage-13 to -20 embryos. (A) Treated at stage 13; beadsoaked in 5mgml~' retinoic acid. Digit pattern: II III IIIIV. Bead at anterior of bud at stage 17. (B) Treated atstage 14; bead soaked in Smgrnl"' retinoic acid. Digitpattern: IV III IIII III IV; two fibulae and one tibia inforeleg. Bead at anterior of bud at stage 17. (C) Treatedat stage 20; bead soaked in 1 mgmP1 retinoic acidimplanted beneath the apical ridge at the anterior marginof leg bud. Digit pattern: IV HI III IV.
stages (10-13), was due to the position of the beadrelative to the limb field, the pattern of digits wascompared with the distance from the tail at which thebead was placed. This analysis revealed no consistenttrend. This was probably due to the variability ofbead displacement following positioning on themembrane over the leg region. However, when theposition of the bead in relation to the bud thatdeveloped 1-2 days later was compared with the digitpattern, a position-dependent effect was revealed(see Fig. 4A-C which were prepared from the data inFig. 1). When beads ended up at the anterior part ofthe early bud, duplicated leg patterns could develop,whereas when beads ended up in more posteriorpositions, usually the legs had structures missing.Tables 2A,B separate the data into experiments inwhich the bead ended up anteriorly or was foundposteriorly. The tables show that when beads soakedin Smgml"1 retinoic acid were found anteriorly, thepercentage of duplicated legs formed increased withstage at treatment to around 50 % at stages 13-16.
It should be noted that when the bead releasingretinoid ended up well anterior to the leg bud, thedevelopment of the flank was normal. When beadssoaked in retinoic acid were placed on the membraneeven more anteriorly, over the presumptive wingregion, the pattern of the wing could be affected atleast as early as stage 10.
In addition to the position-dependent effects ofretinoid application, there was also a dose response(Table 2A,B). When beads soaked in lmgml"1
rather than 5 mg ml"1 retinoic acid were placed in thepresumptive leg region, the frequency and extent ofduplications were decreased. Reduced leg skeletonsrarely resulted with beads soaked in lmgmP 1 reti-noic acid. Control beads, soaked in DMSO alone, didnot affect the pattern of the legs.
The earliest time at which retinoids affect legdevelopmentThe data in Fig. 1 and Fig. 4A show that duplicatedlegs can develop when beads soaked in retinoic acidare placed on the vitelline membrane above thepresumptive leg regions in stage-10 embryos. How-ever, it is possible that the retinoid does not exert itseffect on leg development until some later stage. Toinvestigate this question, a series of experiments'wascarried out in which beads were placed on themembrane of stage-10 embryos and then removed atlater times. The length of exposure to retinoic acidrequired to bring about pattern changes after appli-cation at stage 10 was then compared with theexposure required at stages 13 and 14 (N.B. theresults of treatment of stage-14 embryos were verysimilar when beads were placed on the membrane orwere implanted to the embryos, these data are not

728 5. M. Wilde, S. E. Wedden and C. Tickle
44
434334
4334
43234
432234
43211234
3211234
a 211234
11234
Normal 1234
234
2 digits
I digit
0 digits
—D—
L.Position of bead Anterior Anterior Centre Posterior Posterior AnteriorAnterior Centre Posterior Anterior Anterior Centre Posterior
to bud i , i to bud to bud i , > to bud• 23-25 26/27 28/29 30/31leg bud leg bud
leg bud
Fig. 4. Beads soaked in 5 mgml ' retinoic acid were placed in presumptive leg regions of early embryos. Digit patternsin relation to the position of the bead at bud formation at stage 17. This figure was prepared from the data in Fig. 1.N.B. In this figure Arabic numerals rather than the usual Roman are used to denote the leg digits. (A) Stage-10embryos; (B) stage-13 embryos; (C) stage-14 embryos. • 10 cases, otherwise the side length of the square indicates thenumber of cases: the smallest square represents one case. * Somites.
Table 2. The effects of beads soaked in all-trans-retinoic acid applied to presumptive leg regions of early embryoson the skeletal pattern of the legs that developed
(A) Stage-10 to -13
Concentration
acid in which
embryos (beads placed on the membranes)
beads were soaked Stage(mgmr1)
01555
(B) Stage-14 to -16
Concentrationc\ T rp 11 n f\ i c\Jl 1 CL1IIUIL
acid in which
treated
10-1310-13
1011-12
13
Anterior to bud - centre
totalsurvivors
1124371419
%duplicated
021193653
embryos (beads placed in small holes in tissue)
beads were soaked Stage(mgml"1)
15
10
treated
14-1614-1614-16
Implantation to somites
totalsurvivors
18455
%duplicated
224960
:bud*
meannumberof digits
4-04-03-23-73-5
23-29
meannumberof digits
4-23-93-4
Centre
totalsurvivors
36
1658
bud - posterior
%reduced
017758075
to bud*
meannumberof digits
4-03-72-01-81-4
Implantation to somites 29-30
totalsurvivors
9195
%reduced
04280
meannumberof digits
4-03-11-4
Leg bud develops between somites 26-32.* Position of bead relative to leg bud at stage 17.
10 11/12 13 14
Retinoids reprogramme pre-bud limb mesenchyme 729
16 17 18 19 Stages
6h
29 h
bead notremoved
n = 44
13h
18h
24 h
bead notremoved
n = 5
n = 16
6h
18h
24 h
bead notremoved
n = 3
n = 2
n = 4
n = 11
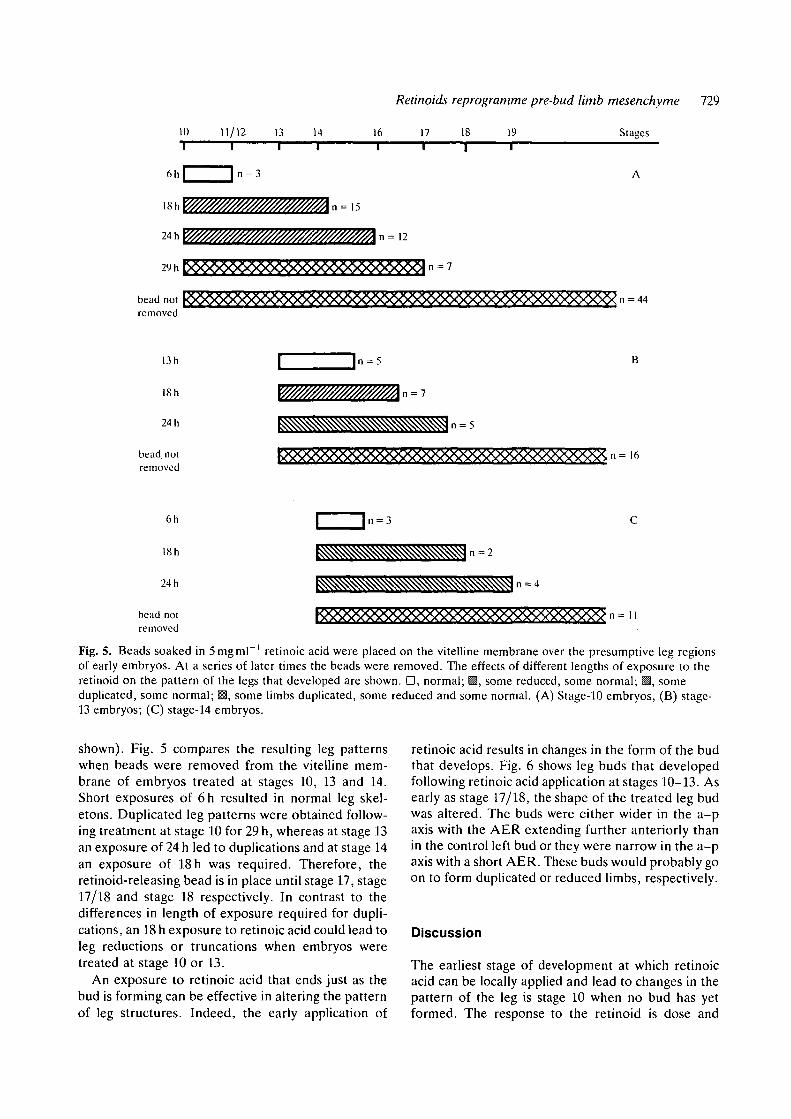
Fig. 5. Beads soaked in 5mgml ' retinoic acid were placed on the vitelline membrane over the presumptive leg regionsof early embryos. At a series of later times the beads were removed. The effects of different lengths of exposure to theretinoid on the pattern of the legs that developed are shown. D, normal; M, some reduced, some normal; ^, someduplicated, some normal; 13, some limbs duplicated, some reduced and some normal. (A) Stage-10 embryos, (B) stage-13 embryos; (C) stage-14 embryos.
shown). Fig. 5 compares the resulting leg patternswhen beads were removed from the vitelline mem-brane of embryos treated at stages 10, 13 and 14.Short exposures of 6h resulted in normal leg skel-etons. Duplicated leg patterns were obtained follow-ing treatment at stage 10 for 29 h, whereas at stage 13an exposure of 24 h led to duplications and at stage 14an exposure of 18h was required. Therefore, theretinoid-releasing bead is in place until stage 17, stage17/18 and stage 18 respectively. In contrast to thedifferences in length of exposure required for dupli-cations, an 18 h exposure to retinoic acid could lead toleg reductions or truncations when embryos weretreated at stage 10 or 13.
An exposure to retinoic acid that ends just as thebud is forming can be effective in altering the patternof leg structures. Indeed, the early application of
retinoic acid results in changes in the form of the budthat develops. Fig. 6 shows leg buds that developedfollowing retinoic acid application at stages 10-13. Asearly as stage 17/18, the shape of the treated leg budwas altered. The buds were either wider in the a-paxis with the AER extending further anteriorly thanin the control left bud or they were narrow in the a-paxis with a short AER. These buds would probably goon to form duplicated or reduced limbs, respectively.
Discussion
The earliest stage of development at which retinoicacid can be locally applied and lead to changes in thepattern of the leg is stage 10 when no bud has yetformed. The response to the retinoid is dose and

730 5. M. Wilde, S. E. Wedden and C. Tickle
Fig. 6. Leg buds that developed following treatment ofthe limb rudiment (on the right side of the embryo) atearlier stages with retinoic acid. (A) Treated stage 10.Drawing of tail end of embryo (photograph of specimenon right). Right bud at stage 17/18 is broad across theanteroposterior axis. Outgrowth is extended anteriorly.Compare with left bud. The anteroposterior extent of theright and left buds and their apical ridges are arrowed.(B) Treated stage 12. SEM of tail end of embryo viewedfrom ventral surface. Right bud at stage 23 is narrowacross the anteroposterior axis and the apical ectodermalridge is short.
position dependent as it is for leg-bud stages. Pro-gressively shorter exposures to retinoic acid arerequired by stage-10, -13 and -14 embryos to dupli-cate leg pattern. They end at stage 17/18 when thebud and apical ridge develop.
Stage, position and dose effectsLocal application of retinoic acid to embryos of awide range of developmental stages, including bothpre-bud and leg-bud stages leads to the developmentof duplicated leg structures. The incidence of dupli-cations obtained by treatment of pre-leg-bud mesen-chyme increases with developmental stage to around50 % for stage-13 to -16 embryos. The more effectivetreatment at these stages may reflect the reduction inthe length of exposure required to duplicate pattern.This increases the likelihood that the bead remains in
the proper position long enough for duplications toresult. Especially at the earliest stages, the bead maytranslocate on the membrane. 100 % duplications areformed when beads are implanted in contact with theAER at the anterior margin of stage-20 leg buds. Thissuggests that interaction of the AER and mesen-chyme may be required for changes in the pattern(see later).
The patterns of duplications appear to be indepen-dent of developmental stage. An additional digit IV isjust as likely to develop in a duplicated patternfollowing treatment at stage 10 as at stage 14.Therefore, the response to retinoids of the limbrudiment at different stages is equivalent.
The effect of retinoids is clearly position dependentwhen applied to limb buds. The duplications occurwhen the retinoid source is at the anterior of the bud.When the bead is at the apex or at the posterior of thebud, the limb can be reduced (Tickle et al. 1985; Lee& Tickle, 1985). The patterns of legs obtained follow-ing treatment at pre-bud stages similarly appear tocorrelate with the position of the bead relative to thebud that forms. Duplications mainly occur when thebead ends up anteriorly, whereas a mixture of dupli-cations and reductions occur when the bead ends upnear the centre of the bud. However, embryostreated at stages 10-13 are particularly susceptible tolimb reduction. This may relate to the small size ofthe limb rudiments at these early stages (Chaube,1959).
The extent of the changes in pattern depends onthe dose of retinoid applied at both pre-bud and leg-bud stages (for wing-bud stages, see also Tickle et al.1985). However, we cannot directly compare thedoses because the beads used for releasing the reti-noid into presumptive limb tissue are smaller thanthose used to treat the limb bud. In addition, thebeads are directly implanted into the leg tissue at budstages rather than being placed nearby or on themembranes.
Exposure time to retinoic acid required to registerpattern changes
When placed on stage-10 embryos, beads soaked inretinoic acid left in position for 29 h produce dupli-cated leg patterns; whereas a 24 h exposure is suf-ficient for stage-13 embryos and 18 h for stage-14embryos. The time for stage-14 embryos is the sameas that required to give full duplications in stage-20wing buds (Eichele et al. 1985). The longer exposureto retinoic acid that leads to duplications after treat-ment at stage 10 compared with stage 14 could suggestthat the limb rudiment may only respond to retinoicacid during the last 18 h of treatment. In this case, theeffective exposure would begin between 11-15 h afterstage 10 at stage 13(+). However, stage-13 embryos

Retinoids reprogramme pre-bud limb mesenchyme 731
require more than an 18 h exposure to duplicate thepattern. This argues against the cells first respondingat stage 13 and instead suggests a changing timerequirement to register pattern duplications duringearly development. The duplicating effect of TTNPBwhen applied at stage 9, rather earlier than retinoicacid, can be explained by the longer half life of thisretinoid in chick embryonic tissue (Eichele et al.1985).
In contrast to the changing exposures required forpattern duplications at stages 10-14, the length oftime required for limb reductions is 18 h for stages 10and 13. The different exposures required for thesetwo effects are difficult to reconcile with the assump-tion that the underlying mechanism of retinoid actionis the same irrespective of position: the differenteffects on limb pattern being due to interactions withthe signal from the native polarizing region (see Lee& Tickle, 1985).
Retinoid effects on bud outgrowthWhen a bead soaked in retinoic acid is positionedabove the leg region of stage-10 to -14 embryos, thebead must remain in place until at least stage 17 for aduplicated leg skeleton to result. Since the leg budfirst grows out from the flank at stage 17, the retinoidmay be required at this stage to affect the develop-ment of the apical ectodermal ridge. However, theamount of retinoid being released from the bead mustbe considerably reduced by this stage (Eichele et al.1984,1985). Even as the bud first grows out, the shapeof the bud and extent of the AER are alreadychanged compared with the contralateral controlbud. Changes in the extent of the apical ridge thatlead to the modification of bud shape can also bedetected as early as 24 h later when retinoic acid isapplied to limb buds (Lee & Tickle, 1985). Therefore,shape change occurs around 24h after treatment,independent of the stage at which retinoid is applied.The changes in shape may be closely linked to theregistering of the retinoid signal. This should becontrasted with the formation of additional digits thatcan occur at quite variable time intervals after reti-noid treatment.
The state of the limb cells when retinoids first begin toaffect the future limb patternOur data suggest that retinoids start to act on the limbrudiment between stages 10 and 13. This correlateswith the stage (11) at which Pinot (1970) found thatisolated prospective leg regions, plus adjacent axialmesenchyme, could first develop into leg structures.However, Rudnick (1945) found that leg structurescould develop from appropriate regions as early asstage 9. In this case, Hensen's node must be includedin the transplant. Since Hensen's node, at this stage,
has polarizing activity (Hornbruch & Wolpert, 1986),interactions may take place within the transplantedtissue that lead to the subsequent determination ofthe limb region. Retinoids appear to be able to startto reprogramme the pattern as soon as the mesen-chyme cells are determined. They appear unable toinduce the formation of a new limb field.
Mode of retinoid actionLeg duplications can arise without wounding thetissue. Treatment of stage-10 to -13 embryos involvedsimply placing the bead on the vitelline membraneover the presumptive limb area. Therefore, retinoidsdo not appear to act in concert with growth stimu-lated at cut surfaces as suggested by Scadding &Maden (1986).
From experiments on limb buds, it has been foundthat implantation of beads soaked in retinoids leadsto the establishment of a stable retinoid gradient(Tickle et al. 1985; Eichele et al. 1985). A gradeddistribution of retinoid is more effective in producingpattern changes than the same amount of retinoiddistributed evenly (Eichele et al. 1985). One possi-bility is that the gradient of retinoid may directlymodify pattern across the a-p axis of the limb. Asecond possibility is that only the high point of thegradient is effective and that this locally transformsadjacent mesenchyme into a new signalling region.
Morphogen gradients are believed to be involved inspecifying the anteroposterior axis in Drosophilaembryos. For example, a graded distribution of theproduct of a homeobox-containing gene has beenfound early in the development of the fertilizedDrosophila egg (Macdonald & Struhl, 1986). How-ever, there is the analogous problem about how suchgradients are read. In this example, the importantfactor in specifying pattern may not be the earlygraded distribution but instead the subsequent local-ization of the gene product at the posterior of theDrosophila embryo.
The results of the experiments here do not dis-tinguish between the two possible mechanisms forretinoid-induced changes in presumptive limb regionsthat lead to altered patterns in the legs. In the firstmechanism, the effects of retinoic acid would have tobring about stable changes in the limb rudiment suchthat extra structures are formed several days later. Inthe second mechanism, retinoids would transformpresumptive limb cells into a new polarizing regionwhich then signals along with the native polarizingregion.
When does the polarizing region signal in limbdevelopment?In amphibian embryos, grafts of regions of the flankposterior to the limb rudiment, when placed anterior

732 S. M. Wilde, S. E. Wedden and C. Tickle
to the rudiment can lead to the development ofduplicated limbs (Slack, 1976, 1977a,b). Hornbruch& Wolpert (1986) have found polarizing activity asearly as stage 10 in the leg region of chick embryos bygrafting tissue to wing buds. It is also well establishedthat posterior tissue from leg buds can cause dupli-cations when grafted to anterior positions in both legand wing buds (Summerbell & Tickle, 1977). There-fore, one interpretation is that the polarizing regionbegins signalling at the time that the limb rudiment isdetermined and continued signalling is requiredthroughout subsequent limb development. A secondpossibility is that the polarizing activity detected bygrafting experiments does not reflect signalling innormal limb development and the tissue only be-comes active when transplanted.
In bud stages, the requirement for the polarizingregion throughout limb development is suggested bythe defective wings that develop after removing aslice from the posterior of the bud (Hinchliffe &Gumpel-Pinot, 1981) and following insertion of animpermeable barrier between anterior and posteriorhalves of the bud (Summerbell, 1979). On the otherhand, removal of tissue from buds between stages 19and 24, normally used for polarizing region grafts,does not affect limb development (Fallon & Crosby,1975). Furthermore, only a short exposure to thesignal from the grafted polarizing region is sufficientto bring about pattern changes (Smith, 1979, 1980).
The time when signals operate is relevant to theanalysis of the mechanisms of signalling in normallimb development. For example, when are the genestranscribed that act in the pattern-forming processacross the anteroposterior axis of the limb? Theseexperiments with retinoids do not resolve this issue.However, retinoids can bring about pattern changeswith denned time/dose relationships. Therefore, theapplication of retinoids will allow the analysis ofevents when a signal that affects anteroposteriorpattern is registered.
This work is supported by the MRC. We thank A.Crawley for help with photography and Professor L.Wolpert and Dr G. Eichele for reading the manuscript.
References
CHAUBE, S. (1959). On axiation and symmetry intransplanted wing of the chick. J. exp. Zool. 140,29-77.
EICHELE, G., TICKLE, C. & ALBERTS, B. M. (1984).Microcontrolled release of biologically activecompounds in chick embryos: beads of 200 ^m diameterfor the local release of retinoids. Anal. Biochem. 142,542-555.
EICHELE, G., TICKLE, C. & ALBERTS, B. M. (1985).Studies on the mechanism of retinoid-induced pattern
duplications in the early chick limb bud: temporal andspatial aspects. J. Cell Biol. 101, 1913-1920.
FALLOW, J. F. & CROSBY, G. M. (1975). Normaldevelopment of the chick wing following removal ofthe polarizing zone. J. exp. Zool. 193, 449-455.
HAMBURGER, V. & HAMILTON, H. L. (1951). A series ofnormal stages in development of the chick embryo. J.Morph. 88, 49-92.
HINCHLIFFE, J. R. & GUMPEL-PINOT, M. (1981). Controlof maintenance and anteroposterior skeletaldifferentiation of the anterior mesenchyme of the chickwing bud by its posterior margin (the ZPA). J.Embryol. exp. Morph. 62, 63-82.
HORNBRUCH, A. & WOLPERT, L. (1986). Positionalsignalling by Hensen's node when grafted to the chicklimb bud. J. Embryol. exp. Morph. 94, 257-265.
JELINEK, R. & KISTLER, A. (1981). Effect of retinoic acidupon chick morphogenetic systems. I. Theembryotoxicity dose range. Teratology 23, 191-195.
KARNOVSKY, M. J. (1965). A formaldehydeglutaraldehyde fixative of high osmolarity for use inelectronmicroscopy. /. Cell Biol. 27, 137a (Abstr).
LEE, J. & TICKLE, C. (1985). Retinoic acid and patternformation in the developing chick wing: SEM andquantitative studies of early effects on the apicalectodermal ridge and bud outgrowth. J. Embryol. exp.Morph. 90, 139-169.
MACDONALD, P. M. & STRUHL, G. (1986). A moleculargradient in early Drosophila embryos and its role inspecifying the body pattern. Nature, Lond. 324,537-545.
PINOT, M. (1970). Le role du mesoderme somitique dansla morphogenese precoce des membres de l'embryonde Poulet. J. Embryol. exp. Morph. 23, 109-151.
RUDNICK, D. (1945). Limb-forming potencies of the chickblastoderm: including notes on associated trunkstructures. Trans. Conn. Acad. Arts Sci. 36, 353-377.
SAUNDERS, J. W. (1948). The proximo-distal sequence ofthe origin of the parts of the chick wing and the role ofthe ectoderm. J. exp. Zool. 108, 363-404.
SAUNDERS, J. W. & GASSELING, M. T. (1968).Ectodermal-mesenchymal interactions in the origin oflimb symmetry. In Epithelial-Mesenchymal Interactions(ed. R. Fleischmajer & R. E. Billingham), pp. 78-97.Baltimore: Williams & Wilkins.
SCADDING, S. R. & MADEN, M. (1986). Comparison ofthe effects of vitamin A on limb development andregeneration in Xenopus laevis tadpoles. J. Embryol.exp. Morph. 91, 35-53.
SLACK, J. M. W. (1976). Determination of polarity in theamphibian limb. Nature, Lond. 261, 44-46.
SLACK, J. M. W. (1977a). Determination ofanteroposterior polarity in the axolotl forelimb byinteraction between limb and flank rudiments. J.Embryol. exp. Morph. 39, 151-168.
SLACK, J. M. W. (1977ft). Control of anteroposteriorpattern in the axolotl forelimb by a smoothly gradedsignal. J. Embryol. exp. Morph. 39, 169-182.
SMITH, J. C. (1979). Evidence for a positional memory inthe development of the chick wing bud. /. Embryol.exp. Morph. 52, 105-113.

Retinoids reprogramme pre-bud limb mesenchyme 733
SMITH, J. C. (1980). The time required for positionalsignalling in the chick wing bud. J. Embryol. exp.Morph. 60, 321-328.
SUMMERBELL, D. (1974). A quantitative analysis of theeffect of excision of the AER from the chick limb-bud.J. Embryol. exp. Morph. 32, 651-660.
SUMMERBELL, D. (1979). The zone of polarizing activity:evidence for a role in normal chick limbmorphogenesis. J. Embryol. exp. Morph. 50, 217-233.
SUMMERBELL, D. (1983). The effect of local application ofretinoic acid to the anteriormargin of the developingchick limb. J. Embryol. exp. Morph. 78, 269-289.
SUMMERBELL, D. & TICKLE, C. (1977). Pattern formationalong the antero-posterior axis of the chick limb bud.In Vertebrate Limb and Somite Morphogenesis (ed. D.
A. Ede, J. R. Hinchliffe & M. Balls), pp. 41-53.Cambridge University Press.
TICKLE, C , SUMMERBELL, D. & WOLPERT, L. (1975).Positional signalling and specification of digits in chicklimb morphogenesis. Nature, Lond. 254, 199-202.
TICKLE, C , ALBERTS, B., WOLPERT, L. & LEE, J. (1982).Local application of retinoic acid to the limb budmimics the action of the polarizing region. Nature,Lond. 296, 564-566.
TICKLE, C , LEE, J. & EICHELE, G. (1985). A quantitativeanalysis of the effect of all-fra/w-retinoic acid on thepattern of chick wing development. Devi Biol. 109,82-95.
(Accepted 16 April 1987)