responses of high-elevation herbaceous plant assemblages to low glacial co2 concentrations revealed...
TRANSCRIPT

1 3
OecologiaDOI 10.1007/s00442-014-2982-y
PhysIOlOgIcal ecOlOgy - OrIgInal research
Responses of high‑elevation herbaceous plant assemblages to low glacial CO2 concentrations revealed by fossil marmot (Marmota) teeth
Bryan S. McLean · Joy K. Ward · Michael J. Polito · Steven D. Emslie
received: 18 December 2013 / accepted: 24 May 2014 © springer-Verlag Berlin heidelberg 2014
almost identical to that of nonherbaceous taxa from lower elevations. however, initial shifts in � aligned most closely with the onset of the late Pleistocene bipolar tempera-ture “seesaw” rather than cO2 increase, indicating unique limitations on glacial-age high-elevation plants may have existed due to both low temperatures and low cO2. Further development of system-specific faunal proxies can help to clarify this and other plant- and ecosystem-level responses to past environmental change.
Keywords atmospheric carbon dioxide · Plant carbon discrimination · Tooth enamel · Quaternary · stable isotopes
Introduction
Plant response to past concentrations of atmospheric cO2
Quaternary floral and faunal fossil material can offer valu-able perspectives on plant responses to changing concentra-tions of atmospheric cO2 ([cO2]). This is because [cO2] change during the Quaternary was substantial, ranging from ~170 to 280 parts per million volume between peak glacial and interglacial periods, respectively (Petit et al. 1999). shifts in [cO2] of this magnitude are expected to have important physiological consequences for c3 plants (gerhart and Ward 2010), which today account for roughly 95 % of earth’s plant species. For example, modern c3 taxa grown at glacial [cO2] levels show reductions in pho-tosynthetic rate and growth between 50 and 90 % relative to those grown under modern [cO2] (sage and coleman 2001; gerhart and Ward 2010). accordingly, effects of Quaternary-scale [cO2] fluctuations on c3 plants may have been manifested at broader scales as well, such as higher
Abstract atmospheric cO2 cycles of the Quaternary likely imposed major constraints on the physiology and growth of c3 plants worldwide. however, the measured record of this remains both geographically and taxonomi-cally sparse. We present the first reconstruction of physi-ological responses in a late Quaternary high-elevation herbaceous plant community from the southern rocky Mountains, Usa. We used a novel proxy—fossilized tooth enamel of yellow-bellied marmots (Marmota flaviventris)—which we developed using detailed isotopic analysis of modern individuals. calculated c isotopic dis-crimination (�) of alpine plants was nearly 2 ‰ lower prior to the last glacial Maximum than at present, a response
communicated by David r. Bowling.
Electronic supplementary material The online version of this article (doi:10.1007/s00442-014-2982-y) contains supplementary material, which is available to authorized users.
B. s. Mclean (*) Department of Biology and Museum of southwestern Biology, University of new Mexico, Msc03-2020, albuquerque, nM 87131-0001, Usae-mail: [email protected]
J. K. Ward Department of ecology and evolutionary Biology, University of Kansas, 1200 sunnyside avenue, lawrence, Ks 66045-7534, Usa
M. J. Polito Woods hole Oceanographic Institution, 266 Woods hole road, Ms #50, Woods hole, Ma 02543-1050, Usa
s. D. emslie Department of Biology and Marine Biology, University of north carolina Wilmington, 601 s. college road, Wilmington, nc 28403, Usa

Oecologia
1 3
trophic levels and across entire ecosystems (Prentice and harrison 2009). To understand the full history of each of these responses, analysis of actual fossil specimens that existed during past periods of low [cO2] is critical. This material has the potential to integrate signals of plant phys-iological adaptation to lower c availability and other per-turbations, long-term plant evolutionary change, and past ecosystem conditions.
Plant responses to Quaternary [cO2] fluctuations pre-viously have been documented via analysis of physically and chemically well-preserved plant macrofossil material. responses of c3 taxa in both north america and europe included various alterations in stomatal density (Beerling et al. 1993; Van de Water et al. 1994), shifts in c isotopic discrimination (Δ) that allow for inference of inter-cellular [cO2]/atmospheric [cO2] (ci/ca) (Beerling et al. 1993), and variation in levels of leaf ci (Van de Water et al. 1994; Ward et al. 2005). yet, to date, the evidence for these responses remains both geographically and taxonomically sparse, being available for only a few plant taxa from a very small num-ber of localities and ecosystems (Beerling et al. 1993; Van de Water et al. 1994; coltrain et al. 2004; Ward et al. 2005).
Vertebrate fossils, on the other hand, are prevalent in the Quaternary record and may help reveal the true diversity of ancient plant [cO2] responses across a broader represen-tation of ecosystems. This application remains relatively unexplored to date, despite increasing knowledge of natural variability in stable isotope ratios among plant photosyn-thetic pathways and across space and time, as well as the ways in which these are assimilated into vertebrate tissues. The development of system-specific vertebrate proxies that also account for the biology of those organisms (e.g., phe-nology, ontogeny, dietary breadth and flexibility) therefore has great—but mostly untapped—potential for document-ing plant responses to ancient environmental perturbations.
high-elevation studies are lacking for plant response to glacial [cO2]
One crucial gap in our knowledge of Quaternary plant physiology is that of high-elevation herbaceous plant taxa (annual and perennial forbs and grasses). Many of these species are adapted to unique climatic and environmen-tal envelopes that include cooler temperatures, shortened growing seasons, and reduced partial pressures of O2 and cO2, which in turn can affect stomatal conductance, Δ and ci/ca, as well as ci. some modern high-elevation taxa dis-play substantial local adaptation to these conditions such as varying Δ (Körner et al. 1988) and ci/ca (Marshall and Zhang 1994). Physiological adaptations to high elevation can be manifested in such evolutionarily relevant traits as increased reproductive performance under varying [cO2] (Ward and strain 1997).
reductions in cO2 partial pressure at high altitude dur-ing periods of already minimal atmospheric [cO2] (e.g., the last glacial period) should be particularly interesting to explore as this may represent the extremes of plant adapta-tion to low [cO2]. cO2 partial pressure is reduced at higher elevations as a function of reductions in total partial pres-sure, and even simultaneous reductions in O2 partial pres-sure and temperature (Terashima et al. 1995) that reduce photorespiration may not overcome the negative effects of limiting c substrate for photosynthesis. In modern stud-ies, reduced temperatures simulating those of the last gla-cial period did not compensate for severe negative effects of low glacial [cO2] on c3 plant physiology and growth, even with some reduction in rates of photorespiration/net photosynthesis (Ward et al. 2008). Owing to these fac-tors, responses of high-elevation taxa to Quaternary [cO2] fluctuations may have differed quantitatively from those at lower elevations, with correspondingly idiosyncratic effects on alpine ecosystems.
The paucity of information on high-elevation herba-ceous plant [cO2] response may be partly due to infrequent fossilization (particularly their lack of fossilizable woody structures) and the poorer (i.e., wetter) preservational con-ditions that typify many high-elevation regions. never-theless, expanding this record is critical, as responses of high-elevation plant taxa can be complex owing to spatial and temporal climatic and environmental variability (Pauli et al. 2012; Macias-Fauria and Johnson 2013). The avail-ability of high-resolution data from the Quaternary fos-sil record would enhance both understanding of past cli-matic and environmental effects on ancient high-elevation ecosystem function as well as the potential magnitude of plant responses to contemporary climate and [cO2] perturbations.
Objectives of this study
We present a new record of plant response to changes in late Quaternary [cO2] from haystack cave, colorado, Usa. It is derived from stable c and O isotope signatures of tooth enamel from an herbivorous montane mammal, the yellow-bellied marmot (Marmota flaviventris), and repre-sents an archive of physiological response in the high-ele-vation herbaceous plant community at this site during the last glacial Maximum (lgM) and most of the last glacial-interglacial transition [~25,000–14,000 years before present (BP)]. We also present detailed isotopic analyses of mod-ern M. flaviventris tooth and bone that refine the utility of conspecific fossil specimens for paleoecological inference. Finally, we combine haystack cave data with results of previous work to yield a nearly continuous record of high-elevation herbaceous plant [cO2] response in the southern rocky Mountains spanning the past ~56,000 years BP.

Oecologia
1 3
Materials and methods
Marmota as a paleoecological proxy and model system
Our approach draws on an extensive knowledge of the biol-ogy and ecology of modern M. flaviventris. yellow-bellied marmots are large-bodied ground squirrels and quintessen-tial montane specialists distributed across the Intermoun-tain West (Frase and hoffmann 1980). The species was pre-sent in the vicinity of haystack cave throughout the late Pleistocene (emslie 1986; this study) and was at large in colorado’s Upper gunnison Basin (UgB) since at least the middle of the Wisconsin glacial period (emslie 2002). The biology of M. flaviventris has been studied intensively in the UgB for several decades [largely at rocky Mountain Biological laboratory; e.g., Ozgul et al. (2010)], providing a strong context for their use in high-elevation paleoeco-logical studies.
M. flaviventris is a generalist herbivore that consumes a variety of high-elevation forbs and grasses (armitage 1979; Frase and armitage 1989). as the c isotopic com-position of mammalian enamel reflects the stable c isotope ratio (δ13c) of bulk diet (ambrose and norr 1993), adult marmot tooth enamel should integrate δ13c of dietary her-baceous plant material occurring on nearby landscapes. Other life history characteristics of M. flaviventris, which include obligate hibernation (often for more than half of the year), result in a strict breeding phenology concentrated in the first several weeks of the growing season in colo-rado (Frase and hoffmann 1980). Breeding is followed by semi-independent foraging for the remainder of the grow-ing season to fuel growth and attain critical overwintering fat reserves (Frase and hoffmann 1980; Ozgul et al. 2010). Marmot body tissues formed during this interval (e.g., molariform teeth) should therefore offer an isotopic “snap-shot” of herbaceous plant δ13c constrained primarily to the growing season.
Furthermore, our use of M. flaviventris is justified by documentation of hibernation in fossil forms (rinaldi 1999) and previous analyses demonstrating dietary niche conservatism in this genus during the middle Pleisto-cene (Feranec et al. 2010), late Pleistocene and holocene (Mclean and emslie 2012). Perhaps the one remaining limitation for using Marmota as a paleoecological proxy is a poor understanding of how ontogenetic dietary changes across the growing season affect enamel isotopic signa-tures, which we addressed directly by isotopic analysis of modern M. flaviventris from our study area.
study site
haystack cave (elevation 2,450 m) is located in the UgB of central-southwestern colorado and has produced one
of the richest late Pleistocene vertebrate faunas known from the southern rocky Mountains (emslie 1986; nash 1987). The modern plant community surrounding hay-stack cave is characterized by an open sagebrush (Artem-esia) steppe/grassland. This community is widespread from low to mid-elevations in the interior UgB, fostered by the unique topographic and climatic setting found here. rela-tive to surrounding highlands, climates in the interior UgB are characterized by reduced annual precipitation (mean 28–42 inches less from 1981 to 2010) and higher maximum annual temperature (mean 8–15 °F warmer) (PrIsM cli-mate group 2013).
Paleoenvironmental constraint is critical for the use of mammalian fossils in reconstructing plant responses to [cO2], as environmental parameters (precipitation, nutrient availability) and plant phylogeny and ecophysiology (c3, c4, caM) influence δ13c of herbaceous taxa and, in turn, δ13cenamel. We were able to tightly control for these factors by analyzing M. flaviventris only from haystack cave and also by drawing from multiple lines of independent evi-dence that establish the dominance of steppe communities here since the late Pleistocene. Perhaps most importantly, the paleozoological record of haystack cave includes a range of mammalian and avian taxa with unambiguous grassland, steppe, and even tundra affinities that are docu-mented throughout cave strata (emslie 1986, 2002).
also, palynological analysis of haystack cave sedi-ments (scott 1981) indicated late Pleistocene plant assem-blages comprised high percentages of Artemisia, com-positae, cleomaceae, and chenopodiaceae/amaranthaceae. Pinus and other large conifers probably occurred only marginally, confined to isolated drainages or moist, shaded microhabitats as they are today. This scenario of late Pleis-tocene plant community stability is distinct from that at nearby higher elevation sites, where glacial lake sediment cores contain high percentages of arboreal pollen and clearly document dynamic late Pleistocene fluctuations of subalpine forests above 2,700 m (Briles et al. 2011).
last, botanical analysis suggests high levels of alpha-level endemism in the UgB, and that floral communities were well mixed here during the late Pleistocene (Barrell 1969). The available paleozoological, palynological and modern botanical records are thus in agreement that plant community structure and composition at haystack cave remained relatively stable on late Quaternary timescales, and that it is an ideal site for evaluating high-elevation plant physiological responses across late Quaternary [cO2] shifts.
stratigraphy and dating
Fossil M. flaviventris were collected from three different provenances in haystack cave. Five new radiocarbon (14c) dates were obtained from two provenances excavated by
Oecologia
1 3
nash (1987) and combined with a sixth date available from the third provenance (emslie 1986). Package clam version 2.1 (Blaauw 2010) in r (r Development core Team 2012) was used to calibrate 14c ages and construct an age-depth model for the combined 14c data set. all samples were com-bined due to limited age resolution within each provenance independently, and a linear deposition model was used as this best fit the 14c ages available from the stratigraphic col-umn with highest age resolution (n = 3 14c dates).
To permit comparisons between M. flaviventris from haystack cave and previous data from nearby cement creek cave (elevation 2,950 m), we constructed a calibrated age-depth model for the latter data set (n = 70 samples) using 16 previously reported 14c dates (Mclean and emslie 2012) and a smooth spline technique implemented in clam version 2.1 [(Blaauw 2010); smoothing level = 0.45]. The cement creek cave data set spans from >55,000 years BP to the present, but plant community change following the Pleistocene at this and nearby sites, as well as stratigraphic mixing, precludes detailed analyses of plant physiological parameters in this interval. samples from cave levels dating <30,000 years BP and without direct 14c dates were there-fore omitted from combined analysis, and subsequent com-bined analyses refer to the remaining subset with the com-plete haystack cave data set.
Isotopic analysis
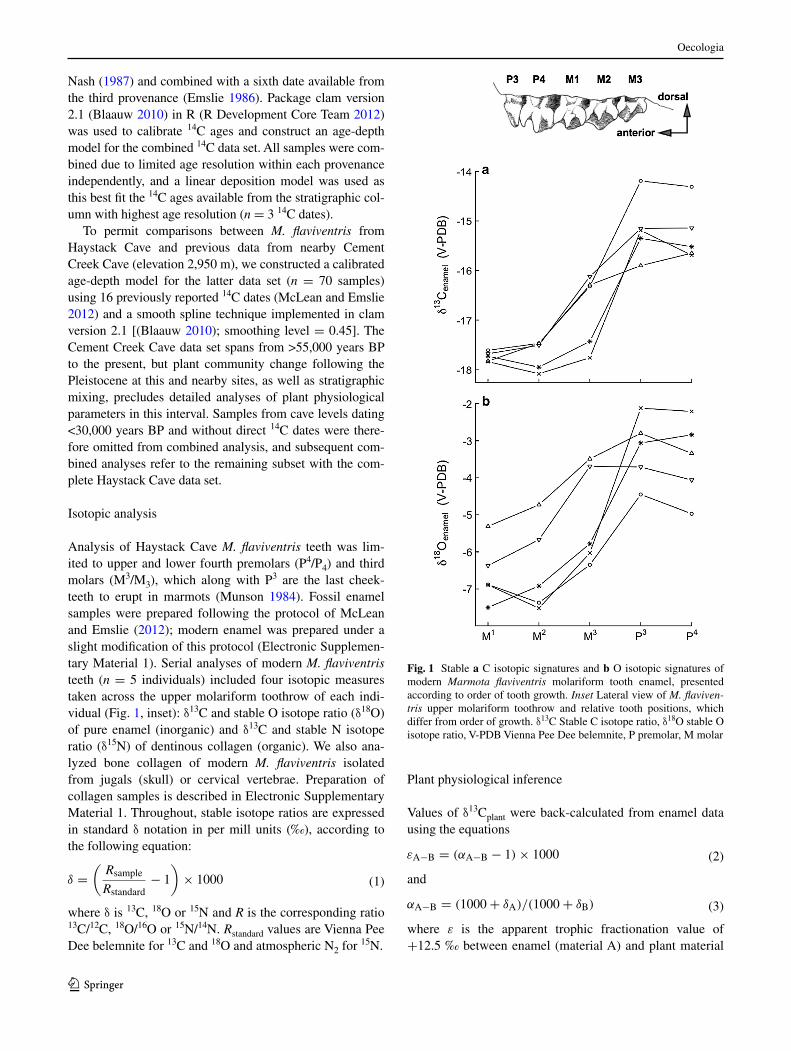
analysis of haystack cave M. flaviventris teeth was lim-ited to upper and lower fourth premolars (P4/P4) and third molars (M3/M3), which along with P3 are the last cheek-teeth to erupt in marmots (Munson 1984). Fossil enamel samples were prepared following the protocol of Mclean and emslie (2012); modern enamel was prepared under a slight modification of this protocol (electronic supplemen-tary Material 1). serial analyses of modern M. flaviventris teeth (n = 5 individuals) included four isotopic measures taken across the upper molariform toothrow of each indi-vidual (Fig. 1, inset): δ13c and stable O isotope ratio (δ18O) of pure enamel (inorganic) and δ13c and stable n isotope ratio (δ15n) of dentinous collagen (organic). We also ana-lyzed bone collagen of modern M. flaviventris isolated from jugals (skull) or cervical vertebrae. Preparation of collagen samples is described in electronic supplementary Material 1. Throughout, stable isotope ratios are expressed in standard δ notation in per mill units (‰), according to the following equation:
where δ is 13c, 18O or 15n and R is the corresponding ratio 13c/12c, 18O/16O or 15n/14n. Rstandard values are Vienna Pee Dee belemnite for 13c and 18O and atmospheric n2 for 15n.
(1)δ =
(
Rsample
Rstandard
− 1
)
× 1000
Plant physiological inference
Values of δ13cplant were back-calculated from enamel data using the equations
and
where ε is the apparent trophic fractionation value of +12.5 ‰ between enamel (material a) and plant material
(2)εA−B = (αA−B − 1) × 1000
(3)αA−B = (1000 + δA)/(1000 + δB)
Fig. 1 stable a c isotopic signatures and b O isotopic signatures of modern Marmota flaviventris molariform tooth enamel, presented according to order of tooth growth. Inset lateral view of M. flaviven-tris upper molariform toothrow and relative tooth positions, which differ from order of growth. δ13c stable c isotope ratio, δ18O stable O isotope ratio, V-PDB Vienna Pee Dee belemnite, P premolar, M molar

Oecologia
1 3
in the diet of marmots (material B), a figure based on con-trolled studies of rodent and lagomorph taxa (Passey et al. 2005; sare et al. 2005). Because no such studies exist for Marmota, and fractionation can vary significantly among taxa, this figure represents an approximation. It is, however, consistent with values used previously (Feranec et al. 2010; Mclean and emslie 2012).
c isotopic discrimination (Δ) of high-elevation herba-ceous plants was calculated as
where δ13cair represents δ13c of atmospheric cO2 and δ13cplant is that calculated above. Values for δ13cair were determined with yearly precision for both modern (Keeling et al. 2005) and fossil data to ~24,000 years BP (schmitt et al. 2012). For cement creek cave data older than this, a δ13cair value of −6.9 ‰ was assumed (leuenberger et al. 1992). equation 2 therefore accounts for both natural past changes in δ13cair and modern decreases due to anthropo-genic fossil fuel combustion (suess effect).
statistical analysis
The r package strucchange version 1.4-7 (Zeileis et al. 2002) was used to infer breakpoints in regressions of Δ on age. Bayesian information criterion (BIc) scores were used as the criteria for assessing different breakpoint scenarios (wherein number of breaks varied from 0 to 5). Package stats version 2.15.2 (r Development core Team 2012) was used for all further statistical analyses.
Results and discussion
serial sampling of modern Marmota teeth
serial isotopic analyses of sequentially accreted tissues (e.g., single teeth within toothrows or growth layers of sin-gle teeth) can reveal important and hard-to-observe features of mammalian ontogeny. however, previous results have been mixed (hobson and sease 1998; Jenkins et al. 2001; newsome et al. 2006). We employed serial analyses of modern M. flaviventris molariform toothrows (Fig. 1, inset) to understand effects of juvenile milk diets on adult enamel signatures and thus inferences of plant Δ. These analyses are novel as they include both inorganic and organic tooth fractions and are the most comprehensive from any mem-ber of the sciuridae, a diverse and near-globally distributed rodent family. The importance of this mammalian system has been demonstrated previously in paleoecological, soci-oecological and modern climate change research.
(4)� =δ13Cair − δ13Cplant
1 + δ13Cplant
We observed significant δ13cenamel variation across M. flaviventris tooth positions (anOVa, F = 28.97, P << 0.01; Fig. 1a), reflecting postnatal transition to a milk diet. low δ13cenamel values for M1 (the first permanent tooth to erupt) appear to reflect in utero diet and are followed by a mean increase of 2.5 ‰ (range 2.2–3.3 ‰ between M1 and P4). The magnitude of this “carnivore effect” agrees well with that observed in enamel of humans and marine mammals (hobson and sease 1998; Wright and schwarcz 1998, 1999), and thus Marmota toothrows may record the entire nursing (but not postweaning) phase. although inter-preted as primarily reflecting ontogenetic dietary change, we note that we presently are unable to assess additional effects of lipids in milk (which are 13c depleted) on mag-nitude of δ13cenamel shifts. however, the relatively low lipid content of milk from other hibernating ground squirrels (skibiel and hood 2013) suggests that these contributions could be small.
The δ13cenamel data (Fig. 1a) fall securely within the range expected given a pure c3 diet. This is consistent with known diets of M. flaviventris in the southern rock-ies and supports their utility as archives of high-elevation plant δ13c. Importantly, however, these data also demon-strate a delay between actual ontogenetic events and their isotopic record in enamel that was previously unrecog-nized. For example, P4 enamel [the last adult tooth to be accreted (Munson 1984)] retains a milk diet signal. This signal suggests further research on fractionation(s) during marmot nursing will be necessary to extract exact absolute dietary δ13c values from adult marmot δ13cenamel. however, we argue that this result does not affect our paleoecologi-cal inferences, as we are concerned with comparing rela-tive changes in plant Δ among time periods. By analyzing identical tooth positions from all time periods, these signals should be preserved unless developmental rates of M. fla-viventris have evolved on late Quaternary timescales.
We also observed significant variation in Marmota δ18Oenamel (anOVa, F = 10.59, P << 0.01; Fig. 1b) and a similar pattern of increase as δ13cenamel. We hypothesize this increase (mean 3.1 ‰) reflects the higher δ18O of milk relative to maternal body water (a form of trophic shift). similar differences in enamel signatures due to nursing/weaning have been demonstrated in ungulates and humans (Bryant et al. 1996; Wright and schwarcz 1998; Franz-Odendaal et al. 2003), but we present the first evidence that a δ18Oenamel increase occurs in Marmota and suggest the above effect is the most plausible explanation. yet we note that a multitude of extrinsic controls on meteoric water δ18O (from which mammalian body water is derived) also offer possible mechanisms for the change we observed.
Precipitation δ18O can increase seasonally (i.e., from winter to summer) by as much as 27 ‰ in the southern rocky Mountains (alstad et al. 1999; Welker 2000), a shift
Oecologia
1 3
more than sufficient to account for our toothrow δ18Oenamel increases. a sourcing shift from snowmelt (generally 18O depleted) to spring precipitation would produce analogous, although probably less prominent, seasonal δ18O increases in meteoric waters (alstad et al. 1999). Finally, δ18O of plant water is expected to increase throughout the growing season due to preferential transpiration of h2
16O relative to h2
18O from photosynthetic tissues (helliker and ehleringer 2000). however, because δ18Oenamel increases have been demonstrated in other mammalian taxa where development of young exceeds a single season, we maintain that a nurs-ing effect is the likely driver of patterns here. nevertheless, we cannot rule out contributions of the above extrinsic fac-tors. Their quantification will require seasonal sampling of meteoric waters and plant forage at our study area as well as marmot tissues with more rapid (annual or subannual) turnover.
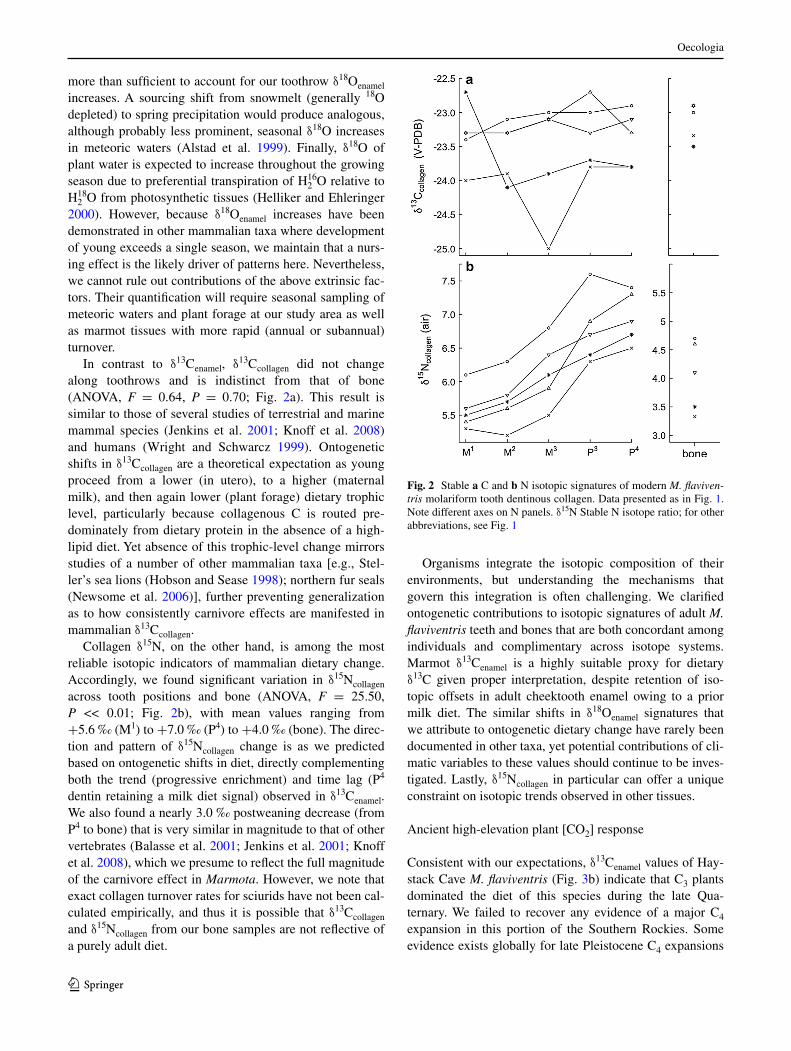
In contrast to δ13cenamel, δ13ccollagen did not change along toothrows and is indistinct from that of bone (anOVa, F = 0.64, P = 0.70; Fig. 2a). This result is similar to those of several studies of terrestrial and marine mammal species (Jenkins et al. 2001; Knoff et al. 2008) and humans (Wright and schwarcz 1999). Ontogenetic shifts in δ13ccollagen are a theoretical expectation as young proceed from a lower (in utero), to a higher (maternal milk), and then again lower (plant forage) dietary trophic level, particularly because collagenous c is routed pre-dominately from dietary protein in the absence of a high-lipid diet. yet absence of this trophic-level change mirrors studies of a number of other mammalian taxa [e.g., stel-ler’s sea lions (hobson and sease 1998); northern fur seals (newsome et al. 2006)], further preventing generalization as to how consistently carnivore effects are manifested in mammalian δ13ccollagen.
collagen δ15n, on the other hand, is among the most reliable isotopic indicators of mammalian dietary change. accordingly, we found significant variation in δ15ncollagen across tooth positions and bone (anOVa, F = 25.50, P << 0.01; Fig. 2b), with mean values ranging from +5.6 ‰ (M1) to +7.0 ‰ (P4) to +4.0 ‰ (bone). The direc-tion and pattern of δ15ncollagen change is as we predicted based on ontogenetic shifts in diet, directly complementing both the trend (progressive enrichment) and time lag (P4 dentin retaining a milk diet signal) observed in δ13cenamel. We also found a nearly 3.0 ‰ postweaning decrease (from P4 to bone) that is very similar in magnitude to that of other vertebrates (Balasse et al. 2001; Jenkins et al. 2001; Knoff et al. 2008), which we presume to reflect the full magnitude of the carnivore effect in Marmota. however, we note that exact collagen turnover rates for sciurids have not been cal-culated empirically, and thus it is possible that δ13ccollagen and δ15ncollagen from our bone samples are not reflective of a purely adult diet.
Organisms integrate the isotopic composition of their environments, but understanding the mechanisms that govern this integration is often challenging. We clarified ontogenetic contributions to isotopic signatures of adult M. flaviventris teeth and bones that are both concordant among individuals and complimentary across isotope systems. Marmot δ13cenamel is a highly suitable proxy for dietary δ13c given proper interpretation, despite retention of iso-topic offsets in adult cheektooth enamel owing to a prior milk diet. The similar shifts in δ18Oenamel signatures that we attribute to ontogenetic dietary change have rarely been documented in other taxa, yet potential contributions of cli-matic variables to these values should continue to be inves-tigated. lastly, δ15ncollagen in particular can offer a unique constraint on isotopic trends observed in other tissues.
ancient high-elevation plant [cO2] response
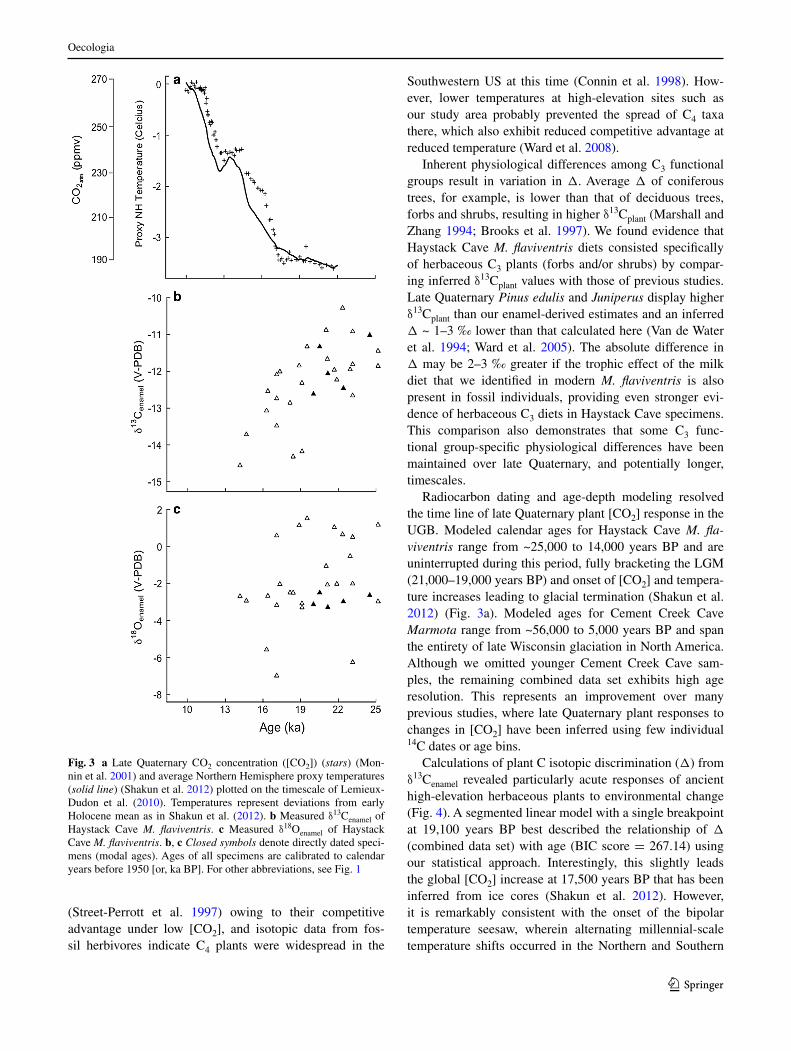
consistent with our expectations, δ13cenamel values of hay-stack cave M. flaviventris (Fig. 3b) indicate that c3 plants dominated the diet of this species during the late Qua-ternary. We failed to recover any evidence of a major c4 expansion in this portion of the southern rockies. some evidence exists globally for late Pleistocene c4 expansions
Fig. 2 stable a c and b n isotopic signatures of modern M. flaviven-tris molariform tooth dentinous collagen. Data presented as in Fig. 1. note different axes on n panels. δ15n stable n isotope ratio; for other abbreviations, see Fig. 1
Oecologia
1 3
(street-Perrott et al. 1997) owing to their competitive advantage under low [cO2], and isotopic data from fos-sil herbivores indicate c4 plants were widespread in the
southwestern Us at this time (connin et al. 1998). how-ever, lower temperatures at high-elevation sites such as our study area probably prevented the spread of c4 taxa there, which also exhibit reduced competitive advantage at reduced temperature (Ward et al. 2008).
Inherent physiological differences among c3 functional groups result in variation in Δ. average Δ of coniferous trees, for example, is lower than that of deciduous trees, forbs and shrubs, resulting in higher δ13cplant (Marshall and Zhang 1994; Brooks et al. 1997). We found evidence that haystack cave M. flaviventris diets consisted specifically of herbaceous c3 plants (forbs and/or shrubs) by compar-ing inferred δ13cplant values with those of previous studies. late Quaternary Pinus edulis and Juniperus display higher δ13cplant than our enamel-derived estimates and an inferred Δ ~ 1–3 ‰ lower than that calculated here (Van de Water et al. 1994; Ward et al. 2005). The absolute difference in Δ may be 2–3 ‰ greater if the trophic effect of the milk diet that we identified in modern M. flaviventris is also present in fossil individuals, providing even stronger evi-dence of herbaceous c3 diets in haystack cave specimens. This comparison also demonstrates that some c3 func-tional group-specific physiological differences have been maintained over late Quaternary, and potentially longer, timescales.
radiocarbon dating and age-depth modeling resolved the time line of late Quaternary plant [cO2] response in the UgB. Modeled calendar ages for haystack cave M. fla-viventris range from ~25,000 to 14,000 years BP and are uninterrupted during this period, fully bracketing the lgM (21,000–19,000 years BP) and onset of [cO2] and tempera-ture increases leading to glacial termination (shakun et al. 2012) (Fig. 3a). Modeled ages for cement creek cave Marmota range from ~56,000 to 5,000 years BP and span the entirety of late Wisconsin glaciation in north america. although we omitted younger cement creek cave sam-ples, the remaining combined data set exhibits high age resolution. This represents an improvement over many previous studies, where late Quaternary plant responses to changes in [cO2] have been inferred using few individual 14c dates or age bins.
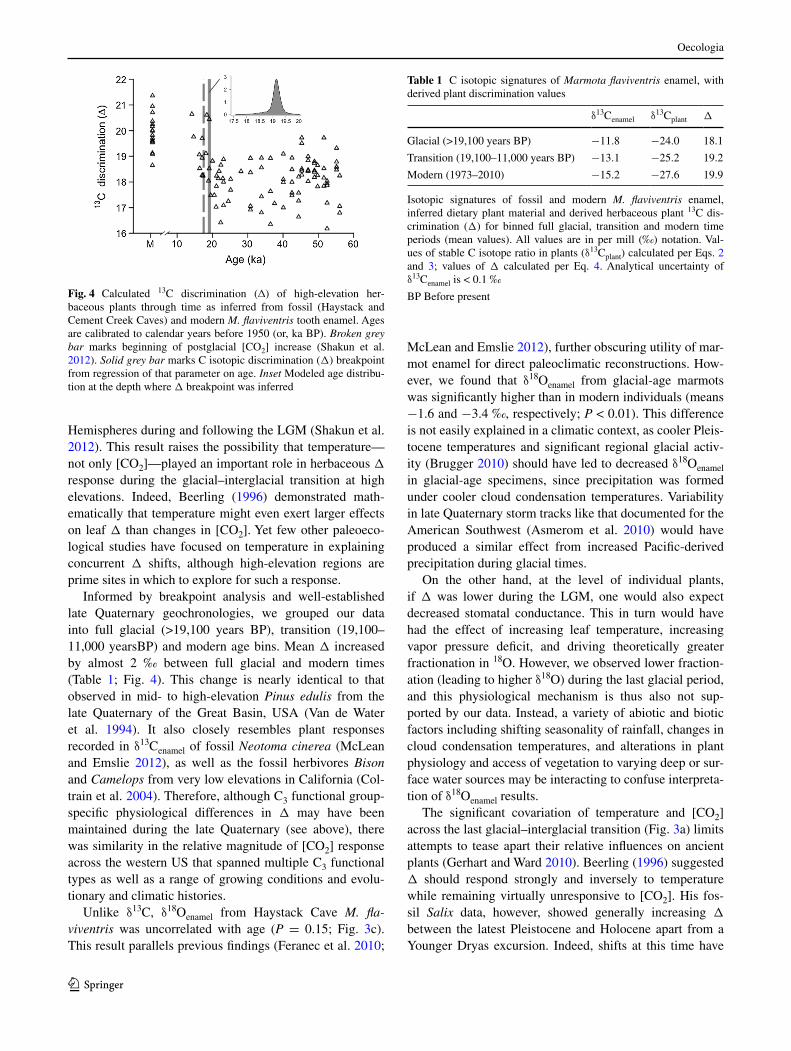
calculations of plant c isotopic discrimination (Δ) from δ13cenamel revealed particularly acute responses of ancient high-elevation herbaceous plants to environmental change (Fig. 4). a segmented linear model with a single breakpoint at 19,100 years BP best described the relationship of Δ (combined data set) with age (BIc score = 267.14) using our statistical approach. Interestingly, this slightly leads the global [cO2] increase at 17,500 years BP that has been inferred from ice cores (shakun et al. 2012). however, it is remarkably consistent with the onset of the bipolar temperature seesaw, wherein alternating millennial-scale temperature shifts occurred in the northern and southern
Fig. 3 a late Quaternary cO2 concentration ([cO2]) (stars) (Mon-nin et al. 2001) and average northern hemisphere proxy temperatures (solid line) (shakun et al. 2012) plotted on the timescale of lemieux-Dudon et al. (2010). Temperatures represent deviations from early holocene mean as in shakun et al. (2012). b Measured δ13cenamel of haystack cave M. flaviventris. c Measured δ18Oenamel of haystack cave M. flaviventris. b, c Closed symbols denote directly dated speci-mens (modal ages). ages of all specimens are calibrated to calendar years before 1950 [or, ka BP]. For other abbreviations, see Fig. 1
Oecologia
1 3
hemispheres during and following the lgM (shakun et al. 2012). This result raises the possibility that temperature—not only [cO2]—played an important role in herbaceous Δ response during the glacial–interglacial transition at high elevations. Indeed, Beerling (1996) demonstrated math-ematically that temperature might even exert larger effects on leaf Δ than changes in [cO2]. yet few other paleoeco-logical studies have focused on temperature in explaining concurrent Δ shifts, although high-elevation regions are prime sites in which to explore for such a response.
Informed by breakpoint analysis and well-established late Quaternary geochronologies, we grouped our data into full glacial (>19,100 years BP), transition (19,100–11,000 yearsBP) and modern age bins. Mean Δ increased by almost 2 ‰ between full glacial and modern times (Table 1; Fig. 4). This change is nearly identical to that observed in mid- to high-elevation Pinus edulis from the late Quaternary of the great Basin, Usa (Van de Water et al. 1994). It also closely resembles plant responses recorded in δ13cenamel of fossil Neotoma cinerea (Mclean and emslie 2012), as well as the fossil herbivores Bison and Camelops from very low elevations in california (col-train et al. 2004). Therefore, although c3 functional group-specific physiological differences in Δ may have been maintained during the late Quaternary (see above), there was similarity in the relative magnitude of [cO2] response across the western Us that spanned multiple c3 functional types as well as a range of growing conditions and evolu-tionary and climatic histories.
Unlike δ13c, δ18Oenamel from haystack cave M. fla-viventris was uncorrelated with age (P = 0.15; Fig. 3c). This result parallels previous findings (Feranec et al. 2010;
Mclean and emslie 2012), further obscuring utility of mar-mot enamel for direct paleoclimatic reconstructions. how-ever, we found that δ18Oenamel from glacial-age marmots was significantly higher than in modern individuals (means −1.6 and −3.4 ‰, respectively; P < 0.01). This difference is not easily explained in a climatic context, as cooler Pleis-tocene temperatures and significant regional glacial activ-ity (Brugger 2010) should have led to decreased δ18Oenamel in glacial-age specimens, since precipitation was formed under cooler cloud condensation temperatures. Variability in late Quaternary storm tracks like that documented for the american southwest (asmerom et al. 2010) would have produced a similar effect from increased Pacific-derived precipitation during glacial times.
On the other hand, at the level of individual plants, if Δ was lower during the lgM, one would also expect decreased stomatal conductance. This in turn would have had the effect of increasing leaf temperature, increasing vapor pressure deficit, and driving theoretically greater fractionation in 18O. however, we observed lower fraction-ation (leading to higher δ18O) during the last glacial period, and this physiological mechanism is thus also not sup-ported by our data. Instead, a variety of abiotic and biotic factors including shifting seasonality of rainfall, changes in cloud condensation temperatures, and alterations in plant physiology and access of vegetation to varying deep or sur-face water sources may be interacting to confuse interpreta-tion of δ18Oenamel results.
The significant covariation of temperature and [cO2] across the last glacial–interglacial transition (Fig. 3a) limits attempts to tease apart their relative influences on ancient plants (gerhart and Ward 2010). Beerling (1996) suggested Δ should respond strongly and inversely to temperature while remaining virtually unresponsive to [cO2]. his fos-sil Salix data, however, showed generally increasing Δ between the latest Pleistocene and holocene apart from a younger Dryas excursion. Indeed, shifts at this time have
Fig. 4 calculated 13c discrimination (�) of high-elevation her-baceous plants through time as inferred from fossil (haystack and cement creek caves) and modern M. flaviventris tooth enamel. ages are calibrated to calendar years before 1950 (or, ka BP). Broken grey bar marks beginning of postglacial [cO2] increase (shakun et al. 2012). Solid grey bar marks c isotopic discrimination (Δ) breakpoint from regression of that parameter on age. Inset Modeled age distribu-tion at the depth where Δ breakpoint was inferred
Table 1 c isotopic signatures of Marmota flaviventris enamel, with derived plant discrimination values
Isotopic signatures of fossil and modern M. flaviventris enamel, inferred dietary plant material and derived herbaceous plant 13c dis-crimination (Δ) for binned full glacial, transition and modern time periods (mean values). all values are in per mill (‰) notation. Val-ues of stable c isotope ratio in plants (δ13cplant) calculated per eqs. 2 and 3; values of Δ calculated per eq. 4. analytical uncertainty of δ13cenamel is < 0.1 ‰
BP Before present
δ13cenamel δ13cplant Δ
glacial (>19,100 years BP) −11.8 −24.0 18.1
Transition (19,100–11,000 years BP) −13.1 −25.2 19.2
Modern (1973–2010) −15.2 −27.6 19.9

Oecologia
1 3
been attributed to changes in both temperature (Beerling et al. 1993) and [cO2] (Van de Water et al. 1994). Impor-tantly, recent empirical work demonstrates that physiologi-cal effects of temperature are subdued at glacial [cO2] lev-els in c3 plants (Terashima et al. 1995; Ward et al. 1999; sage 2002), implicating [cO2] as the major limiting physi-ological variable.
nevertheless, high-elevation taxa are capable of display-ing significant local physiological adaptation to low [cO2] (Körner et al. 1988; Marshall and Zhang 1994). conse-quently, they may have been more sensitive to postglacial temperature increases than were lower elevation taxa at low [cO2], even if these increases were transient (as in the bipolar seesaw). The magnitude of the post-lgM temper-ature increase was greatest at northern high latitudes and lower in temperate zones, but we note that few proxies exist from the interior of western north america. herbaceous plants in higher elevations of colorado may thus have been responding to a heretofore poorly resolved temperature increase within this vast and topographically complex area. such a response would have been further modified by sub-sequent increases in [cO2] beginning ca. 17,500 years BP (Fig. 4). Fully disentangling the nature and timing of plant physiological response at this critical juncture requires additional high-resolution climatic, floral and faunal prox-ies from both the southern rocky Mountains and other high-elevation locales.
Conclusion
We used stable isotopic analysis of fossil M. flaviven-tris, a well-studied, medium-sized montane herbivore, to assess physiological responses of high-elevation herba-ceous plants in colorado to environmental changes of the past ~56,000 years BP. We find that plant c isotopic dis-crimination (�), calculated from δ13cenamel of fossil yellow-bellied marmots, and representing an integrated measure across ancient herbaceous diets, was close to 2 ‰ lower prior to the lgM than in the present day. although likely driven by postglacial [cO2] increase, the initial shift in � inferred at 19,100 years BP aligns most closely with tem-perature fluctuations during the post-lgM bipolar ther-mal seesaw. We therefore hypothesize that physiological limitations on high-elevation herbaceous species may have been imposed not only by low [cO2], but also low late Pleistocene temperatures at high elevations. comparison with nonherbaceous macrofossil data reveals that func-tional group-specific differences in � were conserved on late Quaternary timescales, but that the relative magnitude of � shifts across the entire Pleistocene–holocene transi-tion were strikingly similar across herbaceous and nonher-baceous groups. Further development of additional faunal
proxies like that which we accomplish here should help clarify the generality of this observed [cO2] and tempera-ture response, permitting more complete reconstructions of plant- and ecosystem-level reactions to past environmental change.
Acknowledgments This research was funded by the national sci-ence Foundation (grant ear-0819678), rocky Mountain Biological laboratory (Dr lee r. g. snyder Fellowship), national speleologi-cal society (research grant), and University of new Mexico Biology Department (Joseph gaudin scholarship). carolyn landes (Bureau of land Management, anasazi heritage center) provided access to hay-stack cave samples. Personnel from Quest archaeological research Program (southern Methodist University) and the University of north carolina Wilmington assisted in excavation at cement creek cave. Valentina Ferretti illustrated the marmot maxilla shown in Fig. 1.
References
alstad KP, Welker JM, Williams s, Trlica MJ (1999) carbon and water relations of Salix monticola in response to winter brows-ing and changes in surface water hydrology: an isotopic study using δ13c and δ18O. Oecologia 120:375–385. doi:10.1007/s004420050870
ambrose s, norr l (1993) experimental evidence for the relation-ship of carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In: lambert J, grupe g (eds) Prehistoric human bone: archaeology at the molecular level. springer, Berlin, pp 1–37
armitage KB (1979) Food selectivity by yellow-bellied marmots. J Mammal 60:628–629. doi:10.2307/1380107
asmerom y, Polyak VJ, Burns sJ (2010) Variable winter moisture in the southwestern United states linked to rapid glacial climate shifts. nature 3:114–117. doi:10.1038/ngeO754
Balasse M, Bocherens h, Mariotti a, ambrose sh (2001) Detection of dietary changes by intra-tooth carbon and nitrogen isotopic analysis: an experimental study of dentine collagen of cattle (Bos taurus). J archaeol sci 28:235–245. doi:10.1006/jasc.1999.0535
Barrell J (1969) Flora of the gunnison Basin: gunnison, saguache and hinsdale counties, colorado; a study in the distribution of plants. national land Institute, rockford
Beerling DJ (1996) 13c discrimination by fossil leaves during the late-glacial climate oscillation 12–10 ka BP: measurements and physiological controls. Oecologia 108:29–37. doi:10.1007/BF00333211
Beerling DJ, Mattey DP, chaloner Wg (1993) shifts in the δ13c com-position of Salix herbacea l. leaves in response to spatial and temporal gradients of atmospheric cO2 concentration. Proc r soc B 253:53–60. doi:10.1098/rspb.1993.0081
Blaauw M (2010) Methods and code for “classical” age-model-ling of radiocarbon sequences. Quat geochronol 5:512–518. doi:10.1016/j.quageo.2010.01.002
Briles ce, Whitlock c, Meltzer DJ (2011) last glacial–interglacial environments in the southern rocky Mountains, Usa and impli-cations for younger Dryas-age human occupation. Quat res 77:96–103. doi:10.1016/j.yqres.2011.10.002
Brooks Jr, Flanagan lB, Buchmann n, ehleringer Jr (1997) carbon isotope composition of boreal plants: functional grouping of life forms. Oecologia 110:301–311. doi:10.1007/s004420050163
Brugger Ka (2010) climate in the southern sawatch range and elk Mountains, colorado, Usa., during the last glacial Maximum: inferences using a simple degree-day model. arct antarct alp res 42:164–178. doi:10.1657/1938-4246-42.2.164

Oecologia
1 3
Bryant JD, Froelich Pn, showers WJ, genna BJ (1996) a tale of two quarries: biologic and taphonomic signatures in the oxygen isotope composition of tooth enamel phosphate from modern and Miocene equids. Palaios 11:397–408. http://www.jstor.org/stable/3515249
coltrain JB, harris JM, cerling Te, ehleringer Jr, Dearing M, Ward JK, allen J (2004) rancho la Brea stable isotope biogeochemis-try and its implications for the palaeoecology of late Pleistocene, coastal southern california. Palaeogeogr Palaeoclimatol Palaeo-ecol 205:199–219. doi:10.1016/j.palaeo.2003.12.008
connin sl, Betancourt J, Quade J (1998) late Pleistocene c4 plant dominance and summer rainfall in the southwestern United states from isotopic study of herbivore teeth. Quat res 50:179–193. doi:10.1006/qres.1998.1986
emslie sD (1986) late Pleistocene vertebrates from gunnison county, colorado. J Paleontol 60:170–176. http://www.jstor.org/stable/1305104
emslie sD (2002) Fossil shrews (Insectivora: soricidae) from the late Pleistocene of colorado. southwest nat 47:62–69. http://www.jstor.org/stable/3672802
Feranec rs, hadly ea, Paytan a (2010) Isotopes reveal limited effects of middle Pleistocene climate change on the ecology of mid-sized mammals. Quat Int 217:43–52. doi:10.1016/j.quaint.2009.07.018
Franz-Odendaal Ta, lee-Thorp Ja, chinsamy a (2003) Insights from stable light isotopes on enamel defects and weaning in Pliocene herbivores. J Biosci 28:765–773. doi:10.1007/BF02708437
Frase Ba, armitage KB (1989) yellow-bellied marmots are generalist herbivores. ethol ecol evol 1:353–366. doi:10.1080/08927014.1989.9525505
Frase Ba, hoffmann rs (1980) Marmota flaviventris. Mamm species 135:1–8. http://www.science.smith.edu/msi/pdf/i0076-3519-135-01-0001.pdf
gerhart lM, Ward JK (2010) Plant responses to low [cO2] of the past. new Phytol 188:674–695. doi:10.1111/j.1469-8137.2010.03441.x
helliker Br, ehleringer Jr (2000) establishing a grassland signature in veins: 18O in the leaf water of c3 and c4 grasses. Proc natl acad sci Usa 97:7894–7898. doi:10.1073/pnas.97.14.7894
hobson K, sease J (1998) stable isotope analysis of tooth annuli reveal temporal dietary records: an example using stellar sea lions. Mar Mamm sci 14:116–129. doi:10.1111/j.1748-7692.1998.tb00694.x
Jenkins sg, Partridge sT, stephenson Tr, Farley sD, robbins cT (2001) nitrogen and carbon isotope fractionation between moth-ers, neonates, and nursing offspring. Oecologia 129:336–341. doi:10.1007/s004420100755
Keeling cD, Piper sc, Bacastow rB, Wahlen M, Whorf TP, heimann M, Meijer ha (2005) atmospheric cO2 and 13cO2 exchange with the terrestrial biosphere and oceans from 1978 to 2000: observations and carbon cycle implications. In: ehleringer Jr, cerling Te, Dearing M (eds) a history of atmospheric cO2 and its effects of plants, animals, and ecosystems. springer, new york, pp 83–113
Knoff a, hohn a, Macko s (2008) Ontogenetic diet changes in bottlenose dolphins (Tursiops truncatus) reflected through stable isotopes. Mar Mamm sci 24:128–137. doi:10.1111/j.1748-7692.2007.00174.x
Körner c, Farquhar gD, roksandic Z (1988) a global survey of car-bon isotope discrimination in plants from high altitude. Oecolo-gia 74:623–632. http://www.jstor.org/stable/4218519
lemieux-Dudon B, Blayo e, Petit J-r, Waelbroeck c, svensson a, ritz c, Barnola J-M, narcisi BM, Parrenin F (2010) consistent dating for antarctic and greenland ice cores. Quat sci rev 29:8–20. doi:10.1016/j.quascirev.2009.11.010
leuenberger M, siegenthaler U, langway cc (1992) carbon isotope composition of atmospheric cO2 during the last ice age from an antarctic ice core. nature 357:488–490. doi:10.1038/357488a0
Macias-Fauria M, Johnson ea (2013) Warming-induced upslope advance of subalpine forest is severely limited by geomorphic processes. Proc natl acad sci Usa 110:8117–8122. doi:10.1073/pnas.1221278110
Marshall JD, Zhang J (1994) carbon isotope discrimination and water-use efficiency in native plants of the north-central rock-ies. ecology 75:1887–1895. doi:10.2307/1941593
Mclean Bs, emslie sD (2012) stable isotopes reflect the eco-logical stability of two high-elevation mammals from the late Quaternary of colorado. Quat res 77:408–417. doi:10.1016/j.yqres.2012.02.001
Monnin e, Indermuhle a, Dallenbach a, Fluckiger J, stauffer B, stocker TF, raynaud D, Barnola J-M (2001) atmospheric cO2 concentrations over the last glacial termination. science 291:112–114. doi:10.1126/science.291.5501.112
Munson PJ (1984) Teeth of juvenile woodchucks as seasonal indi-cators on archaeological sites. J archaeol sci 11:395–403. doi:10.1016/0305-4403(84)90020-7
nash DT (1987) archaeological investigations at haystack cave, cen-tral colorado. curr res Pleistocene 4:114–116
newsome sD, Koch Pl, etnier Ma, aurioles-gamboa D (2006) Using carbon and nitrogen isotope values to investigate maternal strategies in northeast Pacific otariids. Mar Mamm sci 22:556–572. doi:10.1111/j.1748-7692.2006.00043.x
Ozgul a, childs DZ, Oli MK, armitage KB, Blumstein DT, Olson le, Tuljapurkar s, coulson T (2010) coupled dynamics of body mass and population growth in response to environmental change. nature 466:482–487. doi:10.1038/nature09210
Passey Bh, robinson TF, ayliffe lK, cerling Te, sponheimer M, Dearing MD, roeder Bl, ehleringer Jr (2005) carbon isotope fractionation between diet, breath cO2, and bioapa-tite in different mammals. J archaeol sci 32:1459–1470. doi:10.1016/j.jas.2005.03.015
Pauli h, gottfried M, Dullinger s, abdaladze O, akhalkatsi M, alonso JlB, coldea g, Dick J, erschbamer B, Fernandez calzado r, ghosn D, holten JI, Kanka r, Kazakis g, Kollar J, larsson P, Moiseev P, Moiseev D, Molau U, Mesa JM, nagy l, Pelino g, Puscas M, rossi g, stanisci a, syverhuset aO, Theuril-lat J-P, Tomaselli M, Unterluggauer P, Villar l, Vittoz P, grabherr g (2012) recent plant diversity changes on europe’s mountain summits. science 336:353–355. doi:10.1126/science.1219033
Petit Jr, Jouzel J, raynaud D, Barkov nI, Barnola J-M, Basile I, Bender M, chappellaz J, Davis M, Delaygue g, Delmotte M, Kotlyakov VM, legrand M, lipenkov Vy, lorius c, Pepin l, ritz c, saltzman e, stievenard M (1999) climate and atmos-pheric history of the past 420,000 years from the Vostok ice core, antarctica. nature 399:429–436. doi:10.1038/20859
Prentice Ic, harrison sP (2009) ecosystem effects of cO2 concen-tration: evidence from past climates. clim Past 5:297–307. doi:10.5194/cp-5-297-2009
PrIsM climate group, Oregon state University (2013) http://prism.nacse.org/normals/. accessed 10 October 2013
r Development core Team (2012) r: a language and environment for statistical computing. r Foundation for statistical computing, Vienna. http://www.r-project.org/
rinaldi c (1999) a record of hibernation in the incisor teeth of recent and fossil marmots (Marmota flaviventris). In: Mayhall J (ed) Proceedings of the 11th international symposium of dental mor-phology. Oulu University Press, Oulu, pp 112–119
sage rF (2002) Variation in the kcat of rubisco in c3 and c4 plants and some implications for photosynthetic performance at high and low temperature. J exp Biol 53:609–620. doi:10.1093/jexbot/53.369.609
sage rF, coleman Jr (2001) effects of low atmospheric cO2 on plants: more than a thing of the past. Trends Plant sci 6:18–24. doi:10.1016/s1360-1385(00)01813-6

Oecologia
1 3
sare DTJ, Millar Js, longstaffe FJ (2005) Tracing dietary protein in red-backed voles (clethrionomys gapperi) using stable isotopes of nitrogen and carbon. can J Zool 83:717–725. doi:10.1139/Z05-064
schmitt J, schneider r, elsig J, leuenberger D, lourantou a, chap-pellaz J, Kohler P, Joos F, stocker TF, leuenberger M, Fischer h (2012) carbon isotope constraints on the deglacial cO2 rise from ice cores. science 336:711–714. doi:10.1126/science.1217161
scott lJ (1981) Palynological investigations of three sites at cure-canti national recreation area, colorado. In: euler rT, stiger Ma (eds) 1978 test excavations at five archaeological sites in curecanti national recreation area, Intermountain colorado. Midwest archeological center, lincoln, pp 88–111
shakun JD, clark PU, he F, Marcott sa, Mix ac, liu Z, Otto-Bliesner B, schmittner a, Bard e (2012) global warming pre-ceded by increasing carbon dioxide concentrations during the last deglaciation. nature 484:49–54. doi:10.1038/nature10915
skibiel al, hood Wr (2013) Milk composition in a hibernating rodent, the columbian ground squirrel (Urocitellus columbi-anus). J Mammal 94:146–154. doi:10.1644/1-MaMM-a-078.1
street-Perrott Fa, huang y, Perrott ra, eglinton g, Barker P, Khelifa lB, harkness DD, Olago DO (1997) Impact of lower atmos-pheric carbon dioxide on tropical mountain ecosystems. science 278:1422–1426. doi:10.1126/science.278.5342.1422
Terashima I, Masuzawa T, Ohba h, yokoi y (1995) Is photosynthesis suppressed at higher elevations due to low cO2 pressure? ecol-ogy 76:2663–2668. http://www.jstor.org/stable/2265838
Van de Water PK, leavitt sW, Betancourt Jl (1994) Trends in stoma-tal density and 13c/12c ratios of Pinus flexilis needles during last glacial–interglacial cycle. science 264:239–243. doi:10.1126/science.264.5156.239
Ward JK, strain Br (1997) effects of low and elevated cO2 par-tial pressure on growth and reproduction of Arabidopsis
thaliana from different elevations. Plant cell environ 20:254–260. doi:10.1046/j.1365-3040.1997.d01-59.x
Ward JK, Tissue DT, Thomas rB, strain Br (1999) com-parative responses of model c3 and c4 plants to drought in low and elevated cO2. glob change Biol 5:857–867. doi:10.1046/j.1365-2486.1999.00270.x
Ward JK, harris JM, cerling Te, Wiedenhoeft a, lott MJ, Dearing M-D, coltrain JB, ehleringer Jr (2005) carbon starvation in glacial trees recovered from the la Brea tar pits, southern cali-fornia. Proc natl acad sci Usa 102:690–694. doi:10.1073/pnas.0408315102
Ward JK, Myers Da, Thomas rB (2008) Physiological and growth responses of c3 and c4 plants to reduced temperature when grown at low cO2 of the last ice age. J Integr Plant Biol 50:1388–1395. doi:10.1111/j.1744-7909.2008.00753.x
Welker JM (2000) Isotopic (δ18O) characteristics of weekly pre-cipitation collected across the Usa: an initial analysis with application to water source studies. hydrol Process 14:1449–1464. doi:10.1002/1099-1085(20000615)14:8<1449:aID-hyP993>3.0.cO;2-7
Wright le, schwarcz hP (1998) stable carbon and oxygen isotopes in human tooth enamel: identifying breastfeeding and wean-ing in prehistory. am J Phys anthropol 106:1–18. doi:10.1002/(sIcI)1096-8644(199807)106:3<411:aID-aJPa16>3.0.cO;2-3
Wright le, schwarcz hP (1999) correspondence between stable car-bon, oxygen and nitrogen isotopes in human tooth enamel and dentine: infant diets at Kaminaljuyu. J archaeol sci 26:1159–1170. doi:10.1006/jasc.1998.0351
Zeileis a, leisch F, hornik K, Kleiber c (2002) strucchange: an r package for testing for structural change in linear regression models. J stat softw 7:1–38. http://www.jstatsoft.org/v07/i02/paper