respiration and osmoregulatio in rainbow trout with...
TRANSCRIPT

J. Exp. BioL (i 970), 53, 48i-494 481With 9 text-figures
m'rinted m Great Britain
RESPIRATION AND OSMOREGULATION IN RAINBOWTROUT WITH GILLS DAMAGED BY ZINC SULPHATE
BY J. F. SKIDMORE*Department of Zoology, University of Bristol
{Received 18 November 1969)
INTRODUCTION
Damage to the gill epithelium occurs when hatched fish are killed within a fewhours by solutions of a variety of substances, including detergents (Schmid & Mann,1962), phenols (Christie & Battle, 1963) and salts of heavy metals (Schweiger, 1957;Haider, 1964). When fish are exposed to solutions of the above poisons until the gillopercula have ceased to move, the epithelium covering the secondary lamellae issloughed off from the underlying pillar-cell system and forms a detached but usuallycontinuous layer of tissue lying above and between the desquamated lamellae. Theblood spaces outlined by the pillar cell flanges may be contracted. Mucus is secretedand precipitated over the body surfaces generally, but is not seen to affect the gills.There is no haemorrhage from the gills. Gill changes before the cessation of oper-cular movements have not been studied.
It has not been shown that the destruction of the gill epithelium causes death throughthe breakdown of any vital gill function. Gas exchange, extra-renal excretion and ionicexchange could be obstructed or facilitated as a result of these changes, since thelifting of the epithelial tissue would generally increase the diffusion distance fromwater to blood, whereas any breaks in the epithelium would locally reduce the diffusiondistance. Death might result primarily from asphyxia, or from osmotic stress. Thepresent paper describes the rate of oxygen uptake and the blood osmotic concentrationof rainbow trout exposed to toxic concentrations of zinc sulphate, in an attempt toidentify the physiological cause of death.
METHODS
Rainbow trout (Salmo gairdneri Richardson) from the Surrey Trout Farm, Nails-worth, Wilts., were held a t i 5 ± i ° C . ina mixture of Bristol tap water (Table 1) and de-ionized water. The total hardness was adjusted by dilution to 50 ± 5 p.p.m. as CaCOs.The pH was 7-1 to 7-5. The dissolved oxygen tension exceeded 130 mm. Hg. (Oa > 8p.p.m.). These conditions were maintained in all experiments unless otherwise stated.Fish were held for at least 7 days and were fed with fresh liver twice weekly. Theywere starved for 3 days prior to use in experiments.
Eleven trout, weight 210 ± 60 g. (mean ± standard deviation), total length 29 ± 2 cm.,were anaesthetized singly in 100 p.p.m. tricaine methanesulphonate (MS 222) andplaced on a portable operating table (Smith & Bell, 1964), modified and built by G. F.Holeton. The gills were irrigated with water containing 70 p.p.m. MS 222. The
• Present address: Dept of Biological Sciences, University of Aston in Birmingham.

482 J. F. SKIDMORE
buccal cavity was cannulated through the snout, posterior to the buccal valve,Portex PP 160 polythene tubing, internal diameter 1*14 mm., with a heat-flared(Saunders, 1961). One opercular cavity was cannulated through the centre of theoperculum, at the junction of the opercular and preopercular bones, using PP 90tubing, i.d. o-86 mm., with a heat-flared end (Saunders, 1961). Pairs of electrodesmade from Johnson and Matthey diamel-coated stainless-steel wire, SWG 45, wereimplanted under the skin below and anterior to the heart in ten of the eleven fish. Thedorsal aorta of four of the fish was also cannulated using PP 60 tubing tipped with anumber 21, short-bevel hypodermic needle (Smith & Bell, 1964). Approximately 100international units of heparin in 0-5 ml. Cortland teleost salt solution (Wolf, 1963)were injected into the aorta, and the cannula was plugged. Each fish was then revivedin water free of anaesthetic.
Canhulas
Fig. 1. Flow diagram of water circulation used in respiration experiments.
While still partially sedated, the fish was transferred, head downstream, to aPerspex chamber, 33 cm. by 6 cm. by 7 cm. high (Fig. 1). Water enteredthe chamberthrough a nozzle which directed the current evenly in all directions. The two (or three)cannulas left the chamber through a small side tube sealed with a rubber cap. The endof the chamber was plugged with a rubber bung. The electrode wires left the chamberbetween the bung and its Perspex seating, ending in terminals made of flexible springsattached to the outside of the fish chamber. The water inside the chamber was earthedby a stainless-steel wire grid.
Water was pumped by an Eheim No. 383 pump through a flow meter to the fishchamber at a maximum rate of 800 rril./min. From the chamber it passed through athree-way tap to a fibre-glass filter. Water drained through the filter by gravity to a

Respiration and.osmoregulation in rainbow trout 483
fc6o 1. reservoir. The reservoir was connected to the pump by a second three-way tap. Asecond 60 1. reservoir containing zinc sulphate solution (Zn 40 p.p.m.) was also con-nected to the pump through the second tap. The circulation was completed by thefirst tap, which could direct water or zinc solution through a second filter to the secondreservoir. Both reservoirs were vigorously aerated (POl > 150 mm. Hg, O2 > 9/5p.p.m.). The water upstream and downstream of the fish chamber, and from the buccaland opercular cavities, could be sampled through cannulas a, b, c and d, respectively.All four cannulas were clamped to regulate water flow at 2 ml./min. The blood couldbe sampled through cannula e, being returned to the fish after test.
The cannulas were led in turn to a glass cuvette (dead space o-i ml.) surrounded bya water jacket maintained at the same temperature as the fish. The cuvette contained aBeckman oxygen macro-electrode connected to a Beckman physiological gas analyser,model 160. The oxygen tension was measured in the water entering and leaving thefish chamber, in the water entering the buccal cavity of the fish and leaving its opercularcavity and in the dorsal aortic blood. The readings were recorded on a Devices two-channel pen recorder after amplification by a Devices DC2C pre-amplifier.
The heart electrodes were connected to the second channel of the pen recorder afteramplification by a Devices AC 7 C pre-amplifier, to record heart rate. In six cases theopercular and coughing rates could also be distinguished on the same trace. In theother fish, opercular and coughing rates were timed direct, using a stop watch.
Water flow through the fish chamber was controlled to allow a reduction in POi of30 mm. Hg (2 p.p.m. O2). Thus the fish inspired water at POi 120 mm. Hg. Eachfish was maintained under these conditions overnight. The following day the waterflow was first reduced so that the inspired POi was 40 mm. Hg for 1 hi.-, then increasedto its former rate for 1 hr. Observations were made after 30 and 60 min. at each flowrate. Zinc_ sulphate solution was then circulated to the fish, the P o of the inspiredwater being 120 mm. Hg. Observations were continued at 15 to 30 min. intervalsuntil the gill opercula ceased moving and the heart stopped beating, survival timebeing defined as time to immobilization of the gill opercula. Because most fish struggledwhen the toxic solution was introduced, fish were exposed in later experiments towater containing 40 p.p.m. Zn plus 20 p.p.m. MS 222. This concentration of anaes-thetic in zinc-free water was sufficient to quieten the fish without depressing their rateof oxygen consumption, or affecting balance or changing their reaction to visualstimuli.
The following calculations were made on the observations with each fish, thesymbols being as far as possible those used in mammalian respiratory physiology.
(1)
VQ = Pan^nt)- F/Pa-E) W (2)U = iooPa.B)/Pi (3)
Terms: VOj, Routine* rate of oxygen uptake (mg./kg. fish per hr.); VG, ventilationvolume of water through gills (l./kg. fish per min.); U, utilization of oxygen, or dis-solved oxygen removed by fish (%); P, partial pressure of oxygen (mm. Hg); F, flowof water through fish chamber (ml./min.); W, wet weight of fish (g.).
• Defined as the rate of oxygen uptake of fish whose only activity is spontaneous (Fry, 1937, p. 46)..
484 J. F. SKDDMORE
Subscripts: I, Water inspired through buccal cavity; E, water expired through^opercular cavity; in, water entering fish chamber; out, water leaving fish chamber.
Eleven trout weighing 145116 g. were anaesthetized as before. Approximately075 ml. blood (15% of the estimated blood volume) was taken from the heart bycardiac puncture. The fish were returned to their holding tanks for 7 days to recu-perate. Seven of the fish were then transferred to 2001. of zinc sulphate solution (40p.p.m. Zn, POj > 130 mm. Hg) and four fish to a iool. tank of zinc-free water. Asecond blood sample was obtained from the unanaesthetized zinc-treated fish at gillopercula immobilization and from the control fish (under anaesthetic) after a similartime interval. The osmotic concentration of the blood was measured in a Mechano-Lab vapour-pressure osmometer.
300
(J
* 200
I1
0
In water
>•--{---
In ZnSO* solution
i \ - • Struggling fish (x±t)
/ \ j Quiet ftth(x±t)
-f...j._.|.-i--f-;~i^j^-
i i
40 120 0Po, (mm. Hg)
50Survival time (%)
100
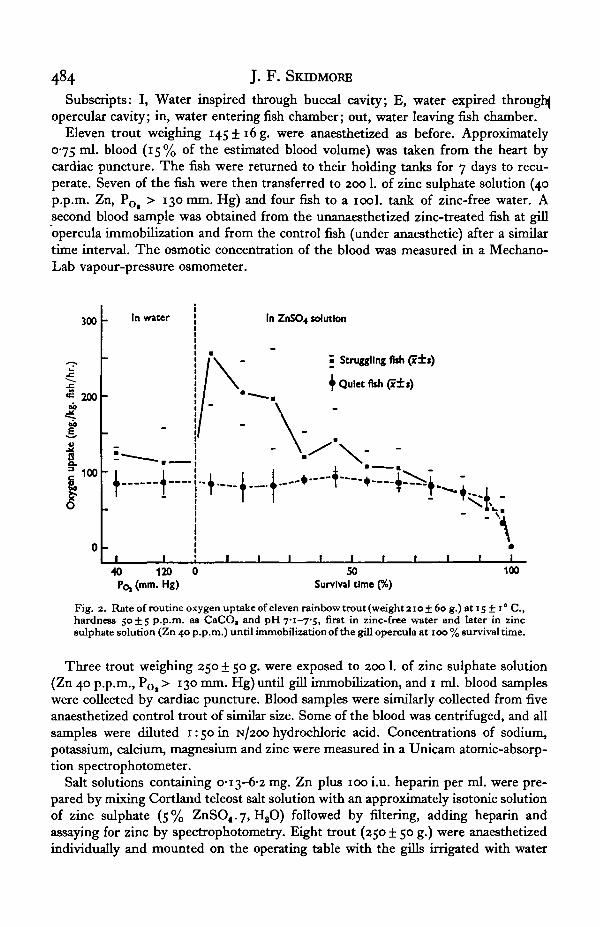
Fig. 2. Rate of routine oxygen uptake of eleven rainbow trout (weight aio± 6o g.) at 15 ± i° C ,hardness 50±5 p.p.m. as CaCO, and pH Ti-TS, first in zinc-free water and later in zincsulphate solution (Zn 40 p.p.m.) until immobilization of the gill opercula at 100 % survival time.
Three trout weighing 250 + 50 g. were exposed to 200 1. of zinc sulphate solution(Zn 40 p.p.m., POf > 130 mm. Hg) until gill immobilization, and 1 ml. blood sampleswere collected by cardiac puncture. Blood samples were similarly collected from fiveanaesthetized control trout of similar size. Some of the blood was centrifuged, and allsamples were diluted 1:50 in N/200 hydrochloric acid. Concentrations of sodium,potassium, calcium, magnesium and zinc were measured in a Unicam atomic-absorp-tion spectrophotometer.
Salt solutions containing O-I3-6-2 mg. Zn plus 100 i.u. heparin per ml. were pre-pared by mixing Cortland teleost salt solution with an approximately isotonic solutionof zinc sulphate (5 % ZnSO4.7, H2O) followed by filtering, adding heparin andassaying for zinc by spectrophotometry. Eight trout (250 ± 50 g.) were anaesthetizedindividually and mounted on the operating table with the gills irrigated with water

Respiration and osmoregulation in rainbow trout 485
containing 70 p.p.m. MS 222. The dorsal aorta was cannulated. One millilitre of bloodwas withdrawn and 1 ml. of solution injected. After initial dilution in the blood, thesetreatments were estimated to have produced concentrations of zinc ranging from 13 to620 p.p.m. Zn plus 10 i.u./ml. heparin. Approximately 0-75 ml. blood was collectedafter 3-60 min. Survivors were returned to their holding tanks. Blood collected beforetreatments was analysed by spectrophotometry for sodium, potassium, calcium,magnesium and zinc; blood collected after treatments was assayed for zinc only.
•
. m
il
:
ume
(vo
lIo
nitl
lat
>•
6
S
4
3
2
1
0
In water
o»1t
- \111
\ \
\ \
- \ l
\ \\ \
\ t
1 1
In ZnSO.4 solution _
/ \/ \
^y\ / \r—S \ / \I \ 1 \\ \ / / \ /
\ / ' v
/
• Struggling fUh
O Quiet fUh
l 1 1 1 1 1 1 1
\
\
\I\1 i
40 120 0Po, (mm. Hg)
SOSurvival time (%)
100
Fig. 3. Ventilation volume of five quiet fish and four struggling fish out of the eleven troutin Fig. 2, in zinc-free water and in zinc sulphate solution. Conditions as in Fig. 2.
RESULTS
The mean routine rate of oxygen uptake (VOl) by eleven resting trout was aboutioo mg./kg. fish per hr. in zinc-free water at a POi of 120 mm. Hg and slightly moreat a POl of 40 mm. Hg (Fig. 2). At a POl of 120 mm. Hg in zinc sulphate solution(Zn 40 p.p.m.) the VOl followed one of two patterns, depending on the fishes' activity.In five unanaesthetized fish which struggled when first immersed in zinc, the VOi
trebled briefly before declining to the resting level after 40% of the survival timehad elapsed. Five lightly sedated fish and one unanaesthetized fish remained quietwhen zinc was introduced. Their oxygen consumption remained steady at the restinglevel until 80% of the survival time. Collapse of oxygen uptake by both groups wasthen rapid.
Ventilation volume (VG) was generally low in high P ^ and high in low POl in zinc-free water (Fig. 3). In five of the six quiet fish the VG increased steadily on exposure

486 J. F. SKIDMORE
to zinc. After 65-95 % °f the survival time it reached six times the rate at high POi inzinc-free water. In four out of five struggling fish the VQ fluctuated between four andten times the resting level in high POi throughout the first 95 % of the survival time.In the remaining two fish the opercular cannulas were blocked. In all eleven fish theventilation rate collapsed rapidly in the final 5 % of survival time., Oxygen utilization (U) was inversely correlated with VG, being high in high POl andlow in low POl, in zinc-free water (Fig. 4). In five quiet fish and four struggling fishit declined from 40 to 4% upon progressive exposure to zinc; but there was wideindividual variation largely depending on the fishes' activity.
•40
o
IS 20
10
• Struggling flih
O Quiet fish
In water
•40 120 0Po, (mm. Hg)
50Survival time (%)
100
Fig. 4. Oxygen utilization of the five quiet trout and four struggling trout in Fig. 3, in zinc-free water and in zinc sulphate solution. Conditions as in Fig. 2.
The mean opercular rate of the eleven fish in zinc-free water was about 70 beats/min. in high P0 | , and 100 beats/min. in low POf (Fig. 5). It rose steadily to 120 beats/min. after 65-85 % of the survival time in zinc, and declined rapidly after 95 % survivaltime. The graphs for individual fish were similar in form, and the large standarddeviations were due to consistent differences in the opercular rates of individual fish.
The mean coughing rate of the eleven fish was I-I coughs/100 opercular beats inhigh POi and almost nil in low POl (Fig. 6). It rose steadily on exposure of fish to zinc,reached a maximum of 20 coughs/100 beats after 85 % survival time, and then declinedrapidly.
The mean heart rate of ten of the eleven fish was 60 beats/min. at high POf and46 beats/min. at low POl (Fig. 7). The rate remained above 50 beats/min. on exposureof the fish to zinc until 50 % of the survival time had elapsed. It then declined to 29

Respiration and osmoregulation in rainbow trout 487
Table 1. Analysis 0/28 samples of Bristol tap water* collected in 1967-8
FactorTotal solids (p.p.m.)Total alkalinity (as pip.m. CaCO,)Total hardness (as p.p.m. CaCO,)Calcium (as p.p.m. CaCO,)Magnesium by subtraction (as p.p.m. CaCO,)Sodium (as p.p.m. Na)Potassium (as p.p.m. K)Iron (as p.p.m. Fe)Copper (as p.p.m. Cu)Zinc (as p.p.m. Zn)Chloride (as p.p.m. Cl)Sulphate (as p.p.m. SOJNitrate (as p.p.m. NO,)Silica (as p.p.m. SiO,)pHChemical oxygen demand (as p.p.m. O)Residual chlorine (as p.p.m. Cl.)
Mean value
246178203171
3*82-5
< o-iNilNil
172 2
1-72-4
7-7o-54O-I2
Range212-270145-205166-232168-174
——————
15-1920-24
1-3-2-0—
7-5-7-9——
• Data provided by Bristol Waterworks Company.
120
80
S3 40S.O
In voter
40 120 0
Po, (mm. Hg)
SOSurvival time (%)
100
Fig. 5. Opercular rate of the eleven trout in Fig. 2, in zinc-free water and in zinc sulphatesolution. Conditions as in Fig. 2.
beats/min. after 95% survival time, but accelerated to 38 beats/min. after the oper-cula became immobilized. Synchronization of opercular movements with heart beatoccurred in five fish out of six examined under low POi in zinc-free water and in fourfish out of six exposed for 65-85 % survival time in zinc solution.
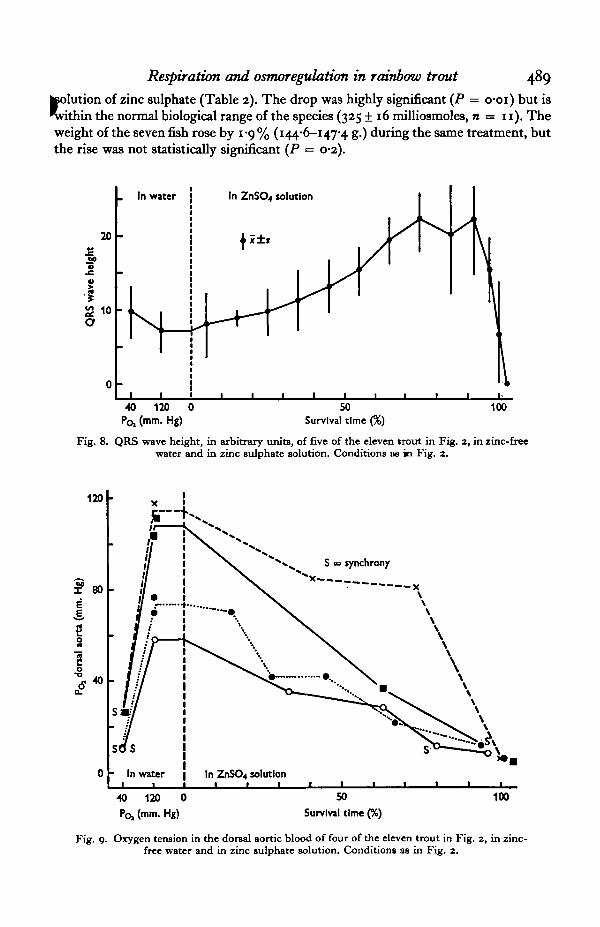
The height of the QRS wave from five of the eleven fish was low in high POj andslightly greater in low POf (Fig. 8). The QRS wave rose steadily on exposure of thefish to zinc, reaching a maximum of three-fold after 75-95 % survival time. Collapsewas then rapid.

The oxygen tension in the dorsal aortic blood (Fig. 9) was high in high water Po, and low in low Po,. ~ l o o d Po, declined steadily on exposure of fish to zinc sulphate solution.
The osmotic concentration of the b l d of seven fish (from a second batch) de- creased by 7'7% from 323 to 298 milliosmoles upon their immobilization in a toxic
Pig. 6. Coughing rate of the eleven trout in Fig. a, in zinc-free water and in zinc sulphate solution. Conditions aa in Fig. 2.
Fig. 7. Heart rate of ten of the eleven trout in Fig. z, in zinc-& water and in zinc sulphate solution Presence or absence of synchiony of cardiac and breathing cyclea indicated for aix of the ten fish. Conditions a s in Fig. a.
Respiration and osmoregulation in rainbow trout 489
keolution of zinc sulphate (Table 2). The drop was highly significant (P = o-oi) but isWithin the normal biological range of the species (325 ±16 milliosmoles, n = 11). Theweight of the seven fish rose by 1 -9 % (i44-6-i47'4 g.) during the same treatment, butthe rise was not statistically significant (P = 0-2).
20
In water | In ZnSO^ solution
102o
40 120 0Pft (mm. Hg)
50Survival time (%)
100
Fig. 8. QRS wave height, in arbitrary units, of five of the eleven trout in Fig. 2, in zinc-freewater and in zinc sulphate solution. Conditions ae in Fig. 2.
120
80
s1o
synchrony
- In water
-40 120 0
Po, (mm. Hg)
50
Survival time (%)
100
Fig. 9- Oxygen tension in the dorsal aortic blood of four of the eleven trout in Fig. 2, in zinc-free water and in zinc sulphate solution. Conditions as in Fig. 2.

490 J- F. SKIDMORE
The concentrations of sodium, potassium, calcium, magnesium and zinc in the|blood of three fish (from a third batch) immobilized in zinc sulphate solution weregenerally similar to concentrations of these ions in thirteen control fish (Table 3). Inparticular, there was no rise in the zinc concentration, and there was a fall in thesodium concentration, the latter being consistent with the observed decrease inosmotic concentration (Table 2).
The measured concentrations of zinc in the blood of four trout (from a fourthbatch) which survived injections of zinc sulphate solution were 27,35, 51 and 96 p.p.m.Zn. The concentrations in four trout which succumbed were 130, 140, 250 and 680p.p.m. Zn, compared with control values of 24115 p.p.m. Zn, ( n = 13).
Table 2. Mean osmotic concentration of blood and mean wet weights of seven rainbowtrout 1 week before treatment and at immobilization in zinc sulphate solution {Zn 40p.p.m.).
(Also mean osmotic concentration of the blood of four rainbow trout before and after a similar periodin tap water.)
Osmotic concentration of blood Wet weight
Before treatmentAfter treatmenttdfP
(milliosmoles)
Fish inwater
329339
0-4
3> 0-5
Fish inZnSO4 sol.
323298
3 76o-oi
of fish (g.)
Fish inZnSO4 sol.
1346147-4
1-46o-a
Table 3. Mean concentrations of sodium, potassium, calcium, magnesium and zinc in theserum, red blood cells and whole blood of three rainbow trout (250 ± 50 g.) immobilizedin zinc sulphate solution (Zn 40p.p.m.) and of 13 trout (250 ± 50^.) held in tap water.
(Sample sizes in parentheses.)
Concentration of metal (p.p.m.)
Fish immobilized inZnSO4 sol. Fish held in tap water
Serum RBC Blood Serum RBC Blood
Sodium 3100(2) 1300(2) 2300(2) 3200(2) 750(2) 2700(12)Potassium 400(2) 2700(2) 1500(3) 120(3) 2600(2) 1200(12)Calcium 320(2) 66(1) 91(3) '3° (3) 180(3) 140(13)Magnesium '30 (2) 60 (2) 44 (2) 29 (2) 60 (2) 57 (12)Zinc 19(1) 69(1) 22(3) 10(1) 50(1) 24(13)
DISCUSSION
Loss of osmotic control may.be ruled out as the cause of death in trout exposed to arapidly lethal solution of zinc sulphate because the changes in blood osmotic concen-tration, recorded centrally, were too slight to exceed the normal range. Similarly, thesmall changes recorded centrally in the blood concentrations of sodium, potassium,calcium, magnesium and zinc were unlikely to cause general physiological imbalance,

Respiration and osmoregulation in rainbow trout 491
blthough local changes in gill tissue concentrations may have produced structural gilldamage and loss of vital function.
When zinc sulphate solution was injected through the dorsal aorta, fish survived afour-fold increase in the normal zinc concentration in blood. Mortality in fish injectedwith higher concentrations of zinc was probably caused by mechanical obstructionof the blood vessels, since similar concentrations of zinc in vitro produced denseprecipitation when added to trout serum and to zinc-free Cortland teleost salt solution.
In contrast, drastic changes were noted in all observed respiratory parameterswhen trout were exposed to a rapidly lethal solution of zinc sulphate rich in oxygen.In quiet fish the routine rate of oxygen uptake remained unchanged until 80 % of thesurvival time had elapsed, and then declined rapidly. Ventilation volume, opercularrate, coughing rate and QRS wave height rose steadily to maximum values at 80-90%survival time. They then collapsed. Oxygen utilization, heart rate and the POi of dorsalaortic blood fell steadily throughout the period of exposure. Extreme bradycardiasometimes led to synchrony of the breathing and cardiac rhythms. In struggling fishsurvival time was shorter, oxygen uptake and ventilation volume were higher andoxygen utilization was lower than in quiet fish, but values of the other respiratoryparameters were similar in both groups.
Generally, fish exposed to zinc had to work increasingly hard to maintain an adequaterate of oxygen uptake. Failure to do so resulted fairly quickly in respiratory failure.Except for the large increases in coughing rate and in QRS wave height the observedchanges were similar to those observed when the same fish had been previouslyexposed to hypoxia. The similarities suggest that gas exchange became increasinglydifficult during exposure to zinc. The increased coughing rate during exposure to zinc(occupying up to 20% of the total opercular movements) probably reduced gillaeration even further since studies with ink showed that flow over the gills was usuallyreversed during coughs. The significance of the increase in QRS wave height is un-explained, though it suggests changes in the physiology of the heart.
Partial support for the view that gas exchange becomes difficult in zinc-poisonedfish is supplied by Jones (1938), who observed that the opercular rate of sticklebacksdoubled in toxic solutions of zinc sulphate and later demonstrated (1947) that an in-crease in opercular rate coincided with a decrease in rate of oxygen uptake in stickle-backs exposed to salts of lead, copper and mercury.
Although there is little literature on the physiological action of toxic substances onaquatic animals, there is more on the normal parameters measured in the presentstudy, under a variety of natural environmental conditions. These will now be dis-cussed in as far as they shed light on the problem of toxic action.
The routine rate of oxygen uptake in zinc-free water (Fig. 2) is similar to standard*and routine rates reported in several recent studies using salmonids (Table 4). Thissupports the view that spontaneous activity in the present study was low and that therate of oxygen uptake of quiet fish was not initially affected by exposure to zinc. Theoxygen uptake of struggling fish rose in zinc sulphate solution to three times theroutine rate, i.e. approximately to the activef rate, but the fish were unable to maintain. • Defined as the minimum rate of oxygen uptake of a figh consistent with its continued existence(Fry, 1957, p. 24).
t Defined as the maximum rate which will permit the highest continued level of activity (Fry, 1957,

492 J. F. SKEDMORE
this level of activity after 50% of their survival time had elapsed. Possibly theilinability to extract sufficient oxygen from the water was already the limiting factor.
Increases in ventilation volume in trout of up to thirteen times the minimum ratehave been measured by Holeton & Randall (1967) and up to five times by Stevens &Randall (1967). Present results gave a maximum ventilation volume of about eleventimes the minimum rate. This was associated with an increase of the opercular ratefrom 74 to 120 beats/min. and an increase in respiratory stroke-volume from 6-7 to48 ml. Hughes & Shelton (1962 ,Fig. 17) estimated that active fish spend such ahigh proportion of their total energy resources on respiration that the theoreticallimit of ventilation volume is about ten times that of the resting rate.
Table 4. Rates of oxygen uptake of salmonids at 150 C.
Species
Brook troutSockeye salmonRainbow troutBrook troutRainbow trout
Rainbow trout
Weight ofnsh(g.)
2 0 0
2 0 0
+oo± 100zoo26o±6o
aio±6o
Rate of oxygenuptake (mg./kg.
fish per hr.)
98, S»71, S
100, Rf107, S76, S
95, R
Reference
Beamish (1064), Table 1Brett (1965), Table 2Holeton & Randall (1967), Fig. 1Job (1955). Fig. 2CP. W. Webb, personal
communicationThis paper
• S = Standard rate of oxygen uptake; f R = routine rate.
Coughing has been studied by Schaumburg, Howard & Walden (1967) in salmonexposed to kraft pulp-mill effluent and DDT. They found a correlation between con-centration of pollutant and cough frequency. Their suggestion that the cough responsemay be related to the environmental POl at the gill surface was not confirmed in thepresent study, because the trout rarely coughed under hypoxia.
Bradycardia has been observed in diving mammals, birds, reptiles and in frogs(Andersen, 1966) as a response to low oxygen tensions in the blood. Holeton &Randall (1967) observed bradycardia in trout exposed to hypoxia, but G. F. Holeton(personal communication) did not observe it in trout exposed to carbon monoxide.The bradycardia in zinc-poisoned fish (Fig. 7) is believed to be the first report of thephenomenum in a fish uninfluenced by drugs and with a high concentration ofaccessible oxygen in the surrounding environment.
The main route of oxygen uptake in fish is believed to be from the water through thegills to the blood, passing through the epithelium of the secondary lamellae, the base-ment membrane and the flanges of the pillar cells (Hughes & Shelton, 1962). The lowoxygen tension observed in blood from the dorsal aorta of zinc-poisoned fish suggeststhat a block to this pathway occurs in the gills.
Histological examination of the gills of 50 g. trout, after exposure of the fish to zincsulphate solution (40 p.p.m. Zn), will be described in a later paper. In that studydamage to the gills was severe after at least 90 % of the survival time had elapsed, butonly slight after 75% of the survival time. This suggests that most gill damageoccurred between 75 % and 90 % of the survival time. Thus gill damage in small troutoccurred after a proportion of the survival time similar to that relating to respiratorychanges in large trout.

Respiration and osmoregulation in rainbow trout 493
k It seems probable that the most extreme changes in respiration physiology were theResult of gill damage modifying gas exchange and thereby imposing an internalhypoxia on the fish. However, less extreme respiratory changes probably precededstructural gill changes. A reduction in the rate of gas transfer through the gills couldresult principally from an increase in diffusion distance from water to blood, a decreasein effective respiratory area, a drop in permeability and changed flow patterns in waterand blood. None of these possibilities has been investigated, and this is considered tobe a suitable area for future work.
SUMMARY
1. Damage to the gill epithelium occurs when hatched fish are killed rapidly bysolutions of zinc sulphate.
2. The rate of routine oxygen uptake by lightly sedated, quiet, rainbow trout didnot alter on exposure to a rapidly toxic solution of zinc sulphate. However, oxygenutilization decreased seven-fold, gill ventilation volume increased six-fold, heart ratewas halved, coughing rate increased 18-fold and the POf of dorsal aortic blood declined.
3. Unsedated trout usually struggled on exposure to zinc. The survival time ofstruggling fish was reduced and oxygen uptake increased, but other physiologicalchanges were similar to those in quiet fish.
4. The respiratory changes in poisoned trout were generally similar to changesobserved earlier in the same fish under hypoxia.
5. The osmotic concentration and the concentrations of sodium, potassium, calcium,magnesium and zinc in blood were largely unaffected by immobilization in zincsulphate solution. Trout survived a four-fold increase in zinc concentration in theblood by injection.
6. The results suggest that epithelial damage decreased the permeability of thegills to oxygen, and did not increase their permeability to cations. Zinc was not a rapidinternal poison. Death was probably caused by tissue hypoxia, when maximum gillventilation was no longer sufficient to supply the oxygen needs of the fish.
I wish to thank Professor H. Heller for the use of the vapour-pressure osmometerand Professor G. M. Hughes for facilities and financial support through a grant fromthe Natural Environment Research Council.
REFERENCES
ANDERSEN, H. T. (1966). Physiological adaptations in diving vertebrates. Pkytiol. Rev. 46, 212-43.BEAMISH, F. W. H. (1964). Respiration of fishes with special emphasis on standard oxygen consumption.
Influence of weight and temperature on respiration of several species. Can. J. Zool. 43, 177-88.BRETT, J. F. (1065). The relation of size to rate of oxygen consumption and sustained swimming speed
of sockeye salmon (Oncorhynchus nerhd). J. Fish. Res. Bd Can. aa, 1491-1501.CHRISTIE, R. M. & BATTLE, HELEN, I. (1963). Histological effects of 3-trifluormethyl-4-nitrophenol
(TFM) on larval lamprey and trout. Can. J. Zool. 41, 51-61.FRY, F. E. J. (1957). The aquatic respiration of fish. In The Physiology of Fishes. Ed. Margaret E.
Brown. Vol. 1, pp. 1—63. New York: Academic Press.HAIDER, G. (1964). Zur Kenntnis von Schwermetallvergiftungen bei Fischen. I. Bleivergiftung bei
Regenbogenforellen (Salmo gatrdneri Rich.) und ihr Nachweis. Z. angeto. Zool. 51, 347-68.HOLBTON, G. F. & RANDALL, D. J. (1967). The effect of hypoxia upon the partial pressure of gases in
the blood and water afferent and efferent to the gills of rainbow trout. J. exp. Biol. 46, 317—27.

494 J- F- SKIDMORE
HUGHES, G. M. & SHELTON, G. (1962). Respiratory mechanisms and their nervous control in fishdAdvanc. comp. Physiol. Biochem. 1, 275-364. ™
JOB, S. V. (1955). The oxygen consumption of SalveUnus fontinalit. Univ. Toronto, Biol. Set. 61.Publication of Ontario Fisheries Research Laboratory. 73, 1-39.
JONES, J. R. E. (1938). The relative toxicity of salts of lead, zinc and copper to the stickleback (Gastero-steus aculeatus L.) and the effect of calcium on the toxicity of lead and zinc salts. J. exp. Biol. 15,304-407.
JONES, J. R. E. (1947). The oxygen consumption of Gasterotteus aculeatus L. in toxic solutions. J. exp.Biol. 33, 298-311.
SAUNDEKS, R. L. (1961). The irrigation of the gills in fishes. I. Studies of the mechanism of branchialirrigation. Can. J. Zool. 39, 637—53.
SCHAUMBURG, F. D., HOWARD, T. E. & WALDEN, C. C. (1967). A method to evaluate the effects ofwater pollutants on fish respiration. Water Res. 1, 731-7.
SCHMID, O. J. & MANN, H. (1962). Die Einwirkung von Dodecylbenzolsulfonat auf die Kiemen vonForellen. Arch. Fischerehoiss. 13, 41—51.
SCHWEIGER, GERTBAUD (1957). Die toxikologische Einwirkung von Schwermetallsalzen auf Fische undFi8chnahrtierc. Arch. Fuchereiwiss. 8, 54—78.
SMITH, L. S. & BELL, G. B. (1964). A technique for prolonged blood sampling in free swimming salmon.J. Fish. Res. Bd Can. ai , 711-17.
STEVENS, E. D. & RANDALL, D. J. (1967). Changes of gas concentrations in blood and water duringmoderate swimming activity in rainbow trout. J. exp. Biol. 46, 329—37.
WOLF, K. (1963). Physiological salines for freshwater teleosts. Prog. Fish. Cultur. 25, 135—40.