resource utilization pattern of two species of tetranychid mites (acarina: tetranychidae)
TRANSCRIPT

Res. Popul. Ecol. (1985) 27, 145-157. �9 by the Society of Population Ecology
R E S O U R C E U T I L I Z A T I O N P A T T E R N O F T W O S P E C I E S O F
T E T R A N Y C H I D M I T E S ( A C A R I N A : T E T R A N Y C H I D A E )
Akira KONDO .1 and Akio TAKAFUJI *.2
* Laboratory of Applied Entomology, College of Agriculture, Okayama University, Okayama 700, Japan
** Entomological Laboratory, College of Agriculture, Kyoto University, Kyoto 606, Japan
INTRODUCTION
The two spotted spider mite, Tetranychus urticae K o ch and the kanzawa spider mite,
Tetranychus kanzawai KISHIDA are both important agricultural pests on a variety of crops
in Japan. Both of these closely related species are highly polyphagous, have many kinds
of host plants in c o m m o n (OSAKABE, 1967) and often occur in the same pear and peach
orchards (e.g. TAKAFUJI et al., 1982; UCHIDA, 1980). Most of the tetranychid mites of
the genus Tetranychus including these two species have very high capacity for population
increase and are typical colonizing species (e.g. TAKAFUJI and KAMIBA'ZASHI, 1984).
Although the two species have many similar ecological characteristics, observations
show that there is a marked difference between the patterns of injury caused by them on
their host plants. Injuring scars on leaves by T. urticae exhibit white-green spots and they
spread all over the leaves as the injury advances. With T. kanzawai, the injured leaves
often become brown and they tend to wither faster than those injured by T. urticae.
This difference seems apparent when we rear these mites on potted plants in the labora-
tory: it is much easier to maintain a population of T. urticae at high levels, than that of
T. kanzawai whose population often crashes because of severe deterioration of host plants.
This suggests that the pattern of resource utilization will differ considerably between the
two species.
There are a number of studies on the life history traits of T. urticae (e.g. LAING, 1969;
SAITO, 1979; HERBERT, 1981), whereas only a few are published on T. kanzawai (e.g.
OSAKABE, 1967). However, most of the data obtained are under different physical
conditions and on different host plants, so that comparison of their life history traits is
difficult.
The purpose of this study is to compare the resource utilization patterns of the two
species, based on their life history parameters and the interactions between the mites
and their host plant under the same experimental conditions.
1 Present address: Department of Pest Control, Okayama Prefectural Agricultural Experiment Station, Sanyo-cho, Okayama 709-08, Japan. Author to whom reprint request should be sent.

146
MATERIALS AND METHODS
The mites
The T. urticae population used in this study was collected from Lamium purpueum
growing in a pear orchard in the south of Okayama city in western Honshfl and the
T. kanzawai population from Clerodendron trichotomum around a pear orchard close to the
orchard where T. urticae was collected. The populations were maintained on the seedlings
of the mottled kidney bean, Phaseolus vulgaris L. under constant fluorescent illumination
at 25•176 70-80% R.H. The T. urticae population is a non-diapausing type, whereas
the population of T. kanzawai is a diapausing one (see TAKAFUJI and KAMIBAYASHI, 1984).
For ovipositing adult females from 4.75 to 5.25 days' postmaturation, the length and
width of idiosoma are both significantly longer for T. urticae than for T. kanzawai
(p<0.001).
Experiments were carried out under the same condition as where the populations
were maintained, except for the experiment in a glasshouse, and all replicates in each
experiment were run simultaneously. The host plant used in the experiments was also
the mottled kidney bean.
Life history parameters
One adult female of each species was introduced onto a 1.3-sq. cm leaflet placed on
water-soaked cotton in a plastic petri dish (9 cm in diameter and 2 cm in depth) and was
allowed to oviposit for 4 h. The eggs laid were removed leaving only one egg per leaflet.
The individual cultures were observed at intervals of 6 h until the first egg was laid by
the female of the next generation to determine the developmental times of each stage and
pre-oviposition period. Thereafter, observation was conducted at intervals of 24 h to
record ovisposition and mortality. The leaflets were replaced with new ones on the
10th day after the adult females were introduced and thereafter, every five days. One
adult male was introduced for mating onto each leaflet when females reached the adult
stage. The sex ratio and hatchability were determined from the eggs which were laid
during 24 h by 200 females of various ages.
Tolerance of starvation
This experiment was designed to determine the capacity of the mites to tolerate
deprivation of food and water and thereby mimic conditions during dispersal. Spider
mites which have low dispersal ability are thought to face severe mortality in the process
of their dispersal.
One adult female from 4.75 to 5.25 days' postmaturation was introduced into a
small plastic cage (1.1 cm in diameter and 0.5 cm in depth) covered with a cover glass.
Mortality was checked at intervals of 12 h.

147
Oviposition pattern within a single leaf
In order to analyze the spatial distribution pattern of eggs oviposited by the mites,
one adult female from 4 to 5 days' postmaturation of each species was introduced onto
a 4-sq. cm leaf disc placed on water-soaked cotton in a plastic petri dish as described
above. A plastic cover with 2-sq. m m quadrats was placed on it so that when viewed
through the cover the leaf disc could be visualized as divided into quadrats. Eight holes
(about 8 m m in diameter) were made in the cover to allow for air circulation. The
number of eggs in each quadrat was recorded at intervals of 12 h for 84 h until the eggs
hatched.
The distribution pattern of the eggs on the leaf disc was analyzed by using the relative
index of aggregation ~/m (IwAo, 1968), where m is mean density and/~ is mean crowding.
In the present study, three spatial units were used; 2, 4 and 8-sq. mm. Although there
were eight replicates for each species, the number of eggs laid on leaf disc varied between
the two species. Therefore, in order to compare the values of ~n/m between the two
species, four replicates in which cumulative number of eggs laid for 84 h was nearly equal
were selected for the analysis.
The effect of leaf quality on dispersal
Three kinds of leaf discs (2.6-sq. cm in diameter) with different degrees of mite injury
were prepared for the experiment, i.e., one without any mite injury (G-leaf), one that
had been injured for 4 days by 10 adult females of each of T. urticae and T. kanzawai (Pl-leaf) and one that had been injured by the same number of each species but for 8 days
(P2-1eaf).
Each of the three leaf discs and one G-leaf were placed 1.5 cm apar t on water-soaked
cotton in a petri dish. Sixteen adult females from 4 to 5 days' postmaturation were
introduced onto the former leaf disc. Each pair of leaf discs was bridged with filter paper
(3 cm in length and 1.3 cm in width) which did not touch water in order to allow the
mites to disperse from the leaf disc to the G-leaf. Thereafter, the number of individuals
which dispersed to a G-leaf or drowned on the water-soaked cotton were counted at
intervals of 6 to 12 h for 120 h, to describe the dispersal tendency of the mites. The
individuals which dispersed to a G-leaf were removed at each census.
The patterns of population increase and dispersal on potted plants
This experiment was carried out from early March to mid-May, 1983 in a glasshouse
at the Faculty of Agriculture, Okayama University. The temperature in the glasshouse
was controlled between 15~ and 35~ by using an oil heater and a fan.
Mottled kidney plants cultivated individually in flower pots (24 cm in diameter and
19 cm in depth) were used for the experiment. A white foam plastic disc (34 cm in
diameter and 2 cm in thickness) with a small central hole to accomodate the plant stem
was placed on the surface of the flower pot. The gap between the stem and the hole

148
was filled with foam plastic and the leaves were prevented from touching the disc. The
flower pots were widely spaced in a large water-filled basin to prevent the mites f rom
migrat ing between the pots.
The bean plants used in the experiments were all about 15 cm tall with 11 leaves.
Sixteen adult females from 4 to 5 days ' pos tmatura t ion of each species were in t roduced
onto a single leaf of each plant. T h e y were confined on the leaf for 6 h to allow them
to settle by covering the stem with water-soaked cotton. To record the number of
dispersing adult females, a drawing paper (6 cm in width) coated with t ransparent adhe-
sive (Kinryfl| was fixed a round the white foam plastic disc and the paper was replaced
with new one at each census.
A populat ion census was made at intervals of 2 to 4 days, count ing the number o f
adult females on each leaf and the number of dispersing adult females which were t rapped
on the paper. T h e number of leaves, the number of defoliated leaves and the relative
injury level were also measured. The grade tbr visual assesment of foliage injury was as
follows: 0, no mite injury; 1, up to 25% surface area spotted; 2, 2 5 - 5 0 % ; 3, 5 0 - 7 5 % ;
4, 75 -100%; 5, dead or defoliated leaf. For each species, the experimet was replicated
five times.
RESULTS
Life history parameters
Table 1 shows the developmental times of the immature female stages of the two
species. For most of the stages, the developmental times were shorter for T. urticae than
for T. kanzawa i and the total developmental time of the former was slightly, but signifi-
cantly (p<0 .05) , shorter than that of the latter. Al though the or• period and
the total adult longevity were not significantly different between the two species (p > 0.05),
the pre-oviposition period was significantly shorter for T. urticae (p<0.001) than for T.
kanzawai (Table 2).
Table 1. Developmental times of the immature female stages of the two species of tetranychid mites in days.
Stage T. urticae a T. kanzawai b
Egg 4. 35• 03 4. 39• 02 Larva 0. 91 • 02 0. 94• 02 Protochrysalis 0. 83 • 0. 02 0. 82 • 0. 02 Protonymph 0. 65 • 02 0. 68 • 0. 02 Deutochrysalis 0. 85• 02 0. 84• 02 Deutonymph 0. 84+0. 02 0. 80• 02 Telochrysalis 1.11 • 02 1.15• 02
Total 9. 53• 02 9. 62• 03
a Average of 52 replicates~S.E. b Average of 37 replicates+S.E.
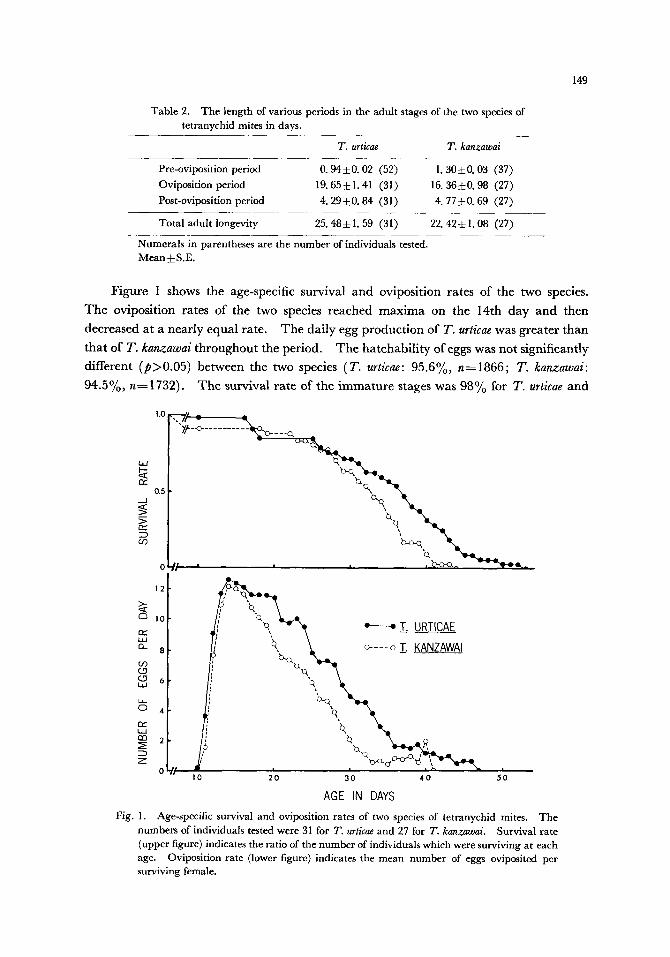
Table 2. The length of various periods in the adult stages of the two species of tetranychid mites in days.
149
T. urticae T. kanzawai
Pre-oviposition period 0. 94+0. 02 (52) 1.30• 03 (37) Oviposition period 19. 65 • 1.41 (31) 16. 36• 98 (27) Post-oviposition period 4. 29• 84 (31) 4. 77• 69 (27)
Total adult longevity 25. 48+ 1.59 (31) 22. 42• 1.08 (27)
Numerals in parentheses are the number of individuals tested. MeanzFS.E.
F igure t shows the age-specific survival and oviposi t ion rates of the two species.
T h e oviposi t ion rates o f the two species reached m a x i m a on the 14th d a y and then
decreased at a n e a r l y equa l rate. T h e da i ly egg p roduc t ion of T. urticae was grea te r t han
tha t of T. kanzawai t h roughou t the per iod. T h e ha t chab i l i t y of eggs was not s ignif icant ly
different ( p > 0 . 0 5 ) be tween the two species (T . urticae: 95.6%, n-=1866; T. kanzawai:
94 .5%, n-~1732). T h e survival ra te of the i m m a t u r e stages was 9 8 % for T. urticae a n d
1,0 , . , ~L -
O.S _J
lo * ; T. URTICAE W
8 ,
o r 10 2 0 30 ,40 5 0
AGE IN DAYS
Fig. 1. Age-specific survival and oviposition rates of two species of tetranychid mites. The numbers of individuals tested were 31 for T. urticae and 27 for T. kanzawai. Survival rate (upper figure) indicates the ratio of the number of individuals which were surviving at each age. Oviposition rate (lower figure) indicates the mean number of eggs oviposited per surviving female.
150
100 ...... .~_ . . . . .
5 0
<> -- URTICAE (3.11 �9 o.lo) 'b,,,, ~ _ C~
co 0 o---.o T. KANZAWAI~ (2.41 + 0.09)1 I " ' ' "O, . "O. n ~ ~
1 2 3 4 5 DAYS
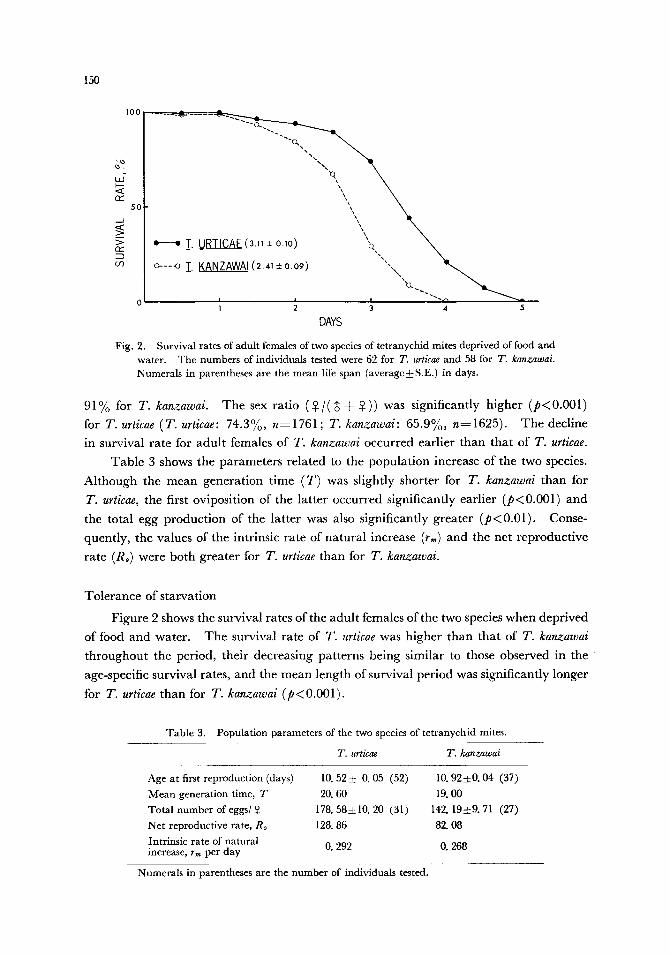
Fig. 2. Survival rates of adult females of two species of tetranychid mites deprived of food and water. The numbers of individuals tested were 62 for T. urticae and 58 for T. kanzawai. Numerals in parentheses are the mean life span (average--4-S.E.) in days.
91% for T. kanzawai. T h e sex ra t io ( ~ / ( ~ + ~ ) ) was s ignif icant ly h igher (p<0 .001)
for T. urticae (T. urticae: 74.3%, n = 1 7 6 1 ; T. kanzawai: 65.9%, n = 1 6 2 5 ) . T h e decl ine
in survival ra te for a d u l t females of T. kanzawai occur red ear l ier t han tha t of T. urticae.
T a b l e 3 shows the pa rame te r s re la ted to the popu la t ion increase of the two species.
A l though the mean genera t ion t ime (T) was sl ightly shor ter for T. kanzawai t han for
T. urticae, the first oviposi t ion of the la t te r occur red signif icant ly ear l ier (p<0 .001) and
the total egg p roduc t ion of the l a t t e r was also s ignif icant ly grea te r ( p<0 .01 ) . Conse-
quent ly , the values of the intr insic ra te of na tu ra l increase (r~,) and the net r eproduc t ive
ra te (Ro) were both g rea te r for T. urticae t han for T. kanzawai.
Tole rance of s ta rva t ion
F igure 2 shows the survival rates of the adu l t females of the two species when depr ived
of food and water . T h e survival ra te of T. urticae was h igher than tha t of T. kanzawai
t h roughou t the per iod, thei r decreas ing pa t te rns be ing s imi lar to those observed in the
age-specific survival rates, and the m e a n length of survival per iod was s ignif icant ly longer
for T. urticae than for T. kanzawai (p<0 .001) .
Table 3. Population parameters of the two species of tetranychid mites.
T. urticae 7". kanzawai
Age at first reproduction (days) Mean generation time, T Total number of eggs/ Net reproductive rate, Ro Intrinsic rate of natural increase, rm per day
10.52_+ 0.05 (52) 10.92:k0.04 (37) 20. 60 19. 00
178.58___10.20 (31) 142. 19:t:9.71 (27) 128. 86 82. 08
0. 292 0. 268
Numerals in parentheses are the number of individuals tested.
151
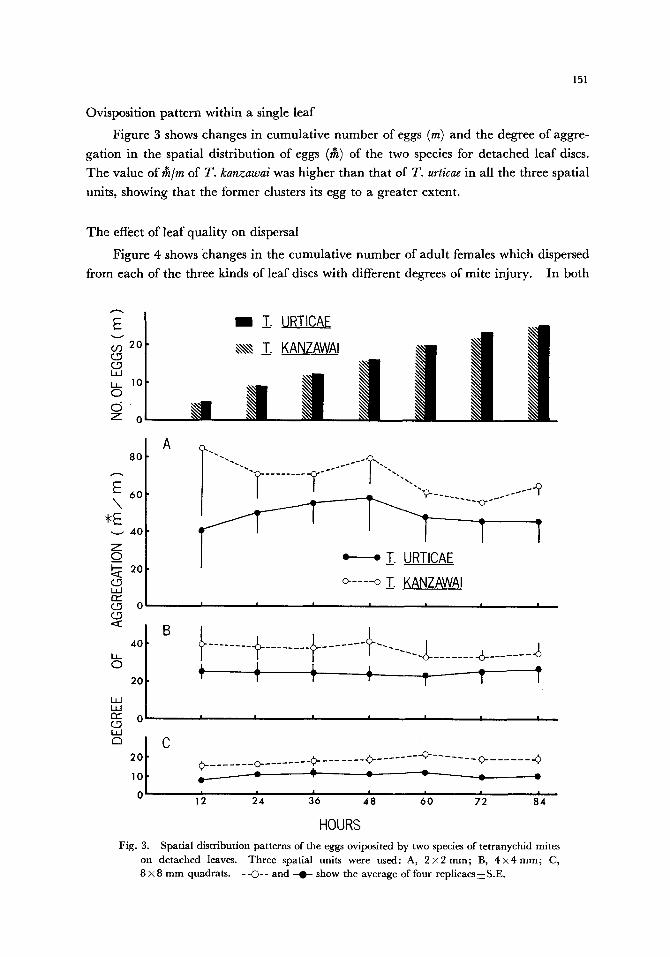
Ovisposition pattern within a single leaf
Figure 3 shows changes in cumulative number of eggs (m) and the degree of aggre-
gation in the spatial distribution of eggs (~) of the two species for detached leaf discs. The value of r~/m of T. kanzawai was higher than that of T. urticae in all the three spatial
units, showing that the former clusters its egg to a greater extent.
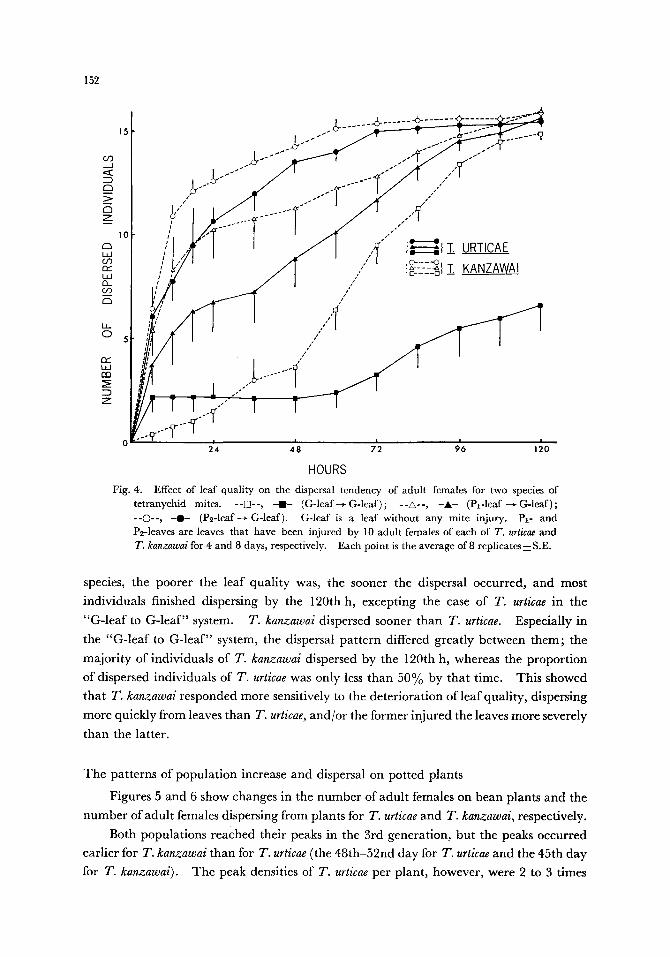
The effect of leaf quality on dispersal
Figure 4 shows Changes in the cumulative number of adult females which dispersed from each of the three kinds of leaf discs with different degrees of mite injury. In both
| BB T. URTICAE
~ [ ~ 20 ,x\\~, T. KANZAWA, ] ~ I
80
E 6O \ *E
v 40 Z o
20 (D laJ rr (D 0 (D
I_1_ 0
i , i LI_I
CD LI_I r'~
A
4ofB 20
0
2~ c 10
0 '
i .... i T. URTICAE
o- .... o T. KANZAWAI I I I I i I I
. . . . . . . . . + . . . . . . . . F . . . . . . . . . . . . . . . . . . . . . + . . . . . . . i I s i V T r T ~ ~ T
f I I I I I I
0 - . . . . . . . o - . . . . . . . . 0 - . . . . . . . O- . . . . . . . -o - . . . . . . . o - . . . . . . �9
12 24 ~6 4'8 gO ;2 84
HOURS Fig. 3. Spatial distribution patterns of the eggs oviposited by two species of tetranychid mites
on detached leaves. Three spatial units were used: A, 2 • B, 4 • C,
8 x 8 m m quadrats. - -O- - and - 0 - show the average of four replicaes ~z S.E.
152
/ ....... T /"
/ / f ,,,,, KANZAWAI
/ /
/ / ' l I
i l! '
/
/
418 712 9/ I 0 24 6 120
HOURS
Fig. 4. Effect of leaf quality on the dispersal tendency of adult females for two species of tetranychid mites. - -if]--, - i - (G-leaf ~ G-leaf) ; --A- -, -A- (Pl-leaf ~ G-leaf) ; --�9 -0-- (P~-leaf--+ G-leaf). G-leaf is a leaf without any mite injury. P1- and P2-1eaves are leaves that have been injured by 10 adult females of each of T. urticae and 7'. kanzawai for 4 and 8 days, respectively. Each point is the average of 8 replicates•
,_.J
n
n z
10
i , I
I.x.I O_ 09
i_L. o 5
Cr t.O r'n
z
~�9 s / , ~ I~ s
/
/ / f .J
= = T. U R T I C A E 0 - - - - 0 i~___.~ T. I O-- - - -Q
species, the poorer the leaf quality was, the sooner the dispersal occurred, and most
individuals finished dispersing by the 120th h, excepting the case of T. urticae in the
"G-leaf to G-leaf" system. T. kanzawai dispersed sooner than T. urticae. Especially in
the "G-leaf to G-leaf" system, the dispersal pattern differed greatly between them; the
majority of individuals of T. kanzawai dispersed by the 120th h, whereas the proportion
of dispersed individuals of T. urticae was only less than 50% by that time. This showed
that T. kanzawai responded more sensitively to the deterioration of leaf quality, dispersing
more quickly from leaves than T. urticae, and/or the former injured the leaves more severely
than the latter.
The patterns of population increase and dispersal on potted plants
Figures 5 and 6 show changes in the number of adult females on bean plants and the
number of adult females dispersing from plants for T. urticae and T. kanzawai, respectively.
Both populations reached their peaks in the 3rd generation, but the peaks occurred
earlier for T. kanzawai than for T. urticae (the 48th-52nd day for T. urticae and the 45th day
for T. kanzawai). The peak densities of T. urticae per plant, however, were 2 to 3 times
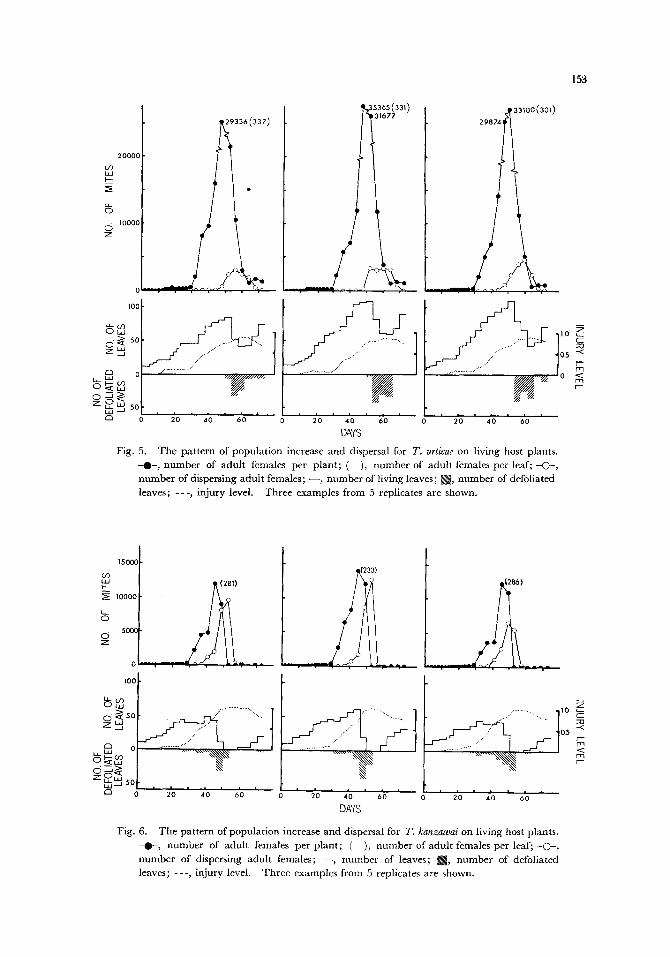
153
20000
Ld
C:; Ioooo Z
100
b~
t-', ~ L U 0
~C~ u J 5 0 LLI - J C l 0
6(33z/
. . . . . . . L J"
V / / ~ / / / H / / .
' 1 o " , ' o " go "
~L~s3~5(331) 31677
,, ..... ............. .Y
o ' 2'0 ' a'o ' 6'o '
DAYS
29874,! 33~~176176
1.0
Ct5
.................... o
20 40 60
Fig. 5. The pat tern of population increase and dispersal for T. urticae on living host plants. - 0 - , number of adult females per plant; ( ), number of adult females per leaf; -C~-, number of dispersing adult females; - - , number of living leaves; ~ , number of defoliated leaves; - - - , injury level. Three examples from 5 replicates are shown.
1 5 0 0 0
o ' )
:~ 10000
b 5 0 0 0
0 7
1 0 0
bm
_.J
123 0 i..u. IJ-I
I - - ( ~
t"-,
A~ I i ~
~233)
_/ . . . . . . I
.. ................... .......
i i 0 210 I 410 OlO I ' 2'o ' ,;o ' go ' o DAYS
Z . ............ 1.0 ~_
""' "" ' " " " t r-"
2'0 4'o 'go '
Fig. 6. The pattern of population increase and dispersal for 7". kanzawai on living host plants. -@-, number of adult females per plant; ( ), number of adult females per leaf; - O - , number of dispersing adult females; - - , number of leaves; [~, number of defoliated leaves; - - - , injury level. Three examples from 5 replicates are shown.

154
higher than those of T. kanzawai, and the peak densities per leaf were also higher for the
former, showing that the population size achieved on host plants was much greater for
T. urticae than for T. kanzawai. The peaks of the number of dispersing adults also
occurred earlier for T. kanzawai than for T. urticae (the 52nd-60th day for T. urticae and
the 49th-53rd day for T. kanzawai). This result corresponded well to the result from the
leaf disc experiment (Fig. 4) which showed that T. kanzawai dispersed sooner than T.
urticae.
Although the peak densities of T. urticae on plants were much higher than those of
T. kanzawai, conversely peaks in dispersing adult females were 2 to 4 times higher for
T. kanzawai than for T. urticae. In addition, the numbers of dispersing T. kanzawai
decreased abruptly after distinct peaks, whereas those of T. urticae populations showed
no distinct peaks but decreased gradually. This difference between species occurred
because most of the dispersing individuals of T. kanzawai dropped onto the white foam
plastic discs when heavily injured leaves defoliated, whereas some individuals of T. urticae
dispersed by spinning down from the tips of the leaves and drifting on air currents beyond
the edges of the discs where their dispersal was not recorded.
The growth of plants was much more heavily hindered by T. kanzawai than by
T. urticae. The maximum numbers of leaves on the plants with T. urticae were about
twice those with T. kanzawai. The numbers of leaves on the plants decreased with
increasing mite injury for each mite species. The leaves of the plants with T. kanzawai
were almost completely defoliated by about the 50th day, whereas those with T. urticae
began to decrease rapidly on about the 60th day but no complete defoliation occurred,
though most leaves deteriorated severely. The defoliation occurred more than 20 days
earlier on the plants with T. kanzawai than on those with T. urticae. Consequently, the
cumulative injury level of the plants was higher with T. kanzawai than with T. urticae
and its peaks occurred sooner with the former (around the 60th day for T. urticae and
around the 50th day for T. kanzawai). The leaves of the plants resurged after abrupt
decreases in mite density, but the new leaves were very small. During the course of this
experiment, there was no significant difference in mean leaf size between the plants with
the two species.
DISCUSSION
The feeding damage caused by the two closely related species were quite different
from each other. The growth of host plants was much more heavily hindered by T.
kanzawai than by T. urticae. It is known that cells punctured by T. urticae collapse and
then adjacent unpunctured cells show coagulated protoplasts and with further damage,
degenerative processes in the chloroplast structures (MoTHES et al., 1982). No such study
on the physiological mechanism in feeding damage by T. kanzawai has been reported.
However, it seems that there may be some qualitative differences between the feeding
mechanisms of the two species because the leaves injured by T. kanzawai become brown

155
and then wither, which seldom occurs in the leaves injured by T. urticae unless the injury
is extremely severe. This difference in feeding damage on host plants then resulted in
distinct differences between the patterns of population increase of the two. T. urticae
numbers increased to a maximum without severe defoliation, and then the mites dispersed.
This pattern in population increase for this species was quite different from that of T.
kanzawal which caused severe deterioration of host plants and dispersed before achieving
maximum population size. Thus, the peak density attained by T. urticae per plant was 2 to 3 times greater that of T. kanzawai and it is evident that the former utilizes a given
food resource much more efficiently, under the experimental conditions used in this study.
WANmUCm and SAITO (1983) studied the processes of population growth and resource
utilization pattern of two species of tetranychid mites, i.e., Olygonychus ununguis (JAcom)
and Panonychus citri (MeGREoOR) and showed that the boundary of the microhabitat
was a single leaf for O. ununguis but sapling or foliage for P. cirri. No such clear difference
is seen between the two species used in this study, except that T. kanzawai tends to cluster
its eggs more than T. urticae within a leaf. The difference in their resource utilization
pattern is therefore attributable to the difference in the interactive pattern with host
plants and the difference in their responses to crowding and/or the deterioration of host plants. The dispersal experiments on detached leaves with various degrees of injury
show that T. urticae is more tolerant than T. kanzawai of over-crowding and/or resource
deterioration. This is also supported by the fact that the former can survive longer when
deprived of food and water.
The intrinsic rate of natural increase, r,, is an index which shows the maximum rate
of potential increase for a species under given environmental conditions and is often used
as criteria when comparing the life history traits and habitat characteristics of species
(e.g. STEARNS, 1976; SOUTHWOOD, 1977). The value ofrm was higher for T. urticae than
for T. kanzawai but the difference was only slight. Nevertheless, the maximum popula-
tion size achieved on host plants was much greater in the former. This shows that for
comparing the capacity for population increase between species, it is essential to take
into consideration the interaction with their food resources.
In the present study, resource utilization patterns of the two species were compared
on a single plant and on the host plant common to them. However, both species are
highly polyphagous and in the field they utilize various host plants as they are available
and suitable as food in different seasons (see TAKAFUJI and KAMIBAYASHI, 1984). There-
fore, in order to understand the significance of the difference in resource utilization
pattern between the two species, it is necessary to compare their population dynamics
in a habitat where the availability of various host plants fluctuates both temporally and spatially.
SUMMARY
Life history and resource utilization pattern were compared between two closely
related mite species of the genus Tetranychus.

156
1. Tetranychus urticae deve loped m o r e qu ick ly a n d h a d a h ighe r ovipos i t ion ra te t h a n
T. kanzawai. Co n s eq u en t l y , the p o t en t i a l for p o p u l a t i o n increase was g rea te r for T.
urticae t h a n for T. kanzawai b u t the di f ference was sl ight.
2. T . urticae was m o r e t o l e r an t o f a de t e r io r a t i ng food resource a n d / o r ove r -c rowding .
T. kanzawai r e s p o n d e d m o r e sensi t ively to food de t e r io r a t i on a n d dispersed m o r e quickly .
3. T h e r e was a cons ide rab le d i f ference b e t w e e n i n j u r y pa t t e rn s by the two species.
T. kanzawai d a m a g e d host p l an t s m o r e severely, caus ing ear l ie r a n d m o r e extens ive
defo l ia t ion of the p lants .
4. T h e m a x i m u m p o p u l a t i o n size ach ieved b y T. urticae was 2 to 3 t imes g rea te r t h a n
t h a t o f T. kanzawai o n po t t ed host p lants . T h i s showed tha t the fo rmer could ut i l ize
a food resource m u c h m o r e efficiently.
ACKNOWLEDGEMENTS: We wish to thank Dr. R. LAMB, of Agriculture Canada at Winnipeg, for reading
the manuscript and verbal correction. We are indebted to Prof. T. YOSHIDA of Okayama University,
Prof. E. KtlNO and Dr. H. TAKEDA of Kyoto University for valuable advice to this study. Lastly, we thank
Mrs. C. TAm-KONDO, and Mr. H. KAMEZAKI of Okayama University for their technical assistance in this
study.
REFERENCES
HERBERT, H.J . (1981) Biology, life tables, and innate capacity for increase of the twospotted spider mite,
Tetranychus urticae (Acarina: Tetranychidae). Can. Entomol. 113: 371-378.
IWAO, S. (1968) A new regression method for analyzing the aggregation pattern of animal populations.
Res. Popul. Ecol. 10: 1-20.
LAING, J. E. (1969) Life history and life table of Tetranychus urticae KocH. Acarologia 11: 32~,2.
MOTHES, U. and K. A. SEXTZ (1982) Fine structural alterations of bean plant leaves by feeding injury of
Tetranychus urticae Koch (Acari, Tetranychidae). Acarologia 23: 149-157.
OSAKABE, M. (1967) Biological studies on the tea red spider mite, Tetranychus kanzawai KXSmDA, in tea
plantation. Bul. Tea Res. Sta. Min. Agr. For. 4:35-156 (in Japanese with an English summary).
SAITO, Y. (1979) Comparative studies of life histories of three species of spider mites (Acarina:
Tetranychidae). Appl. Ent. Zool. 14: 83-94.
SOUTHWGOD, T. R. E. (1977) Habitat, the templet for ecological strategies? J. Anim. Ecol. 46: 337-365.
SamARNS, S. C. (1976) Life history tactics: A review of ideas. Quart. Rev. Biol. 51: 347.
TAKAFUJI, A., T. MAEDA, Y. EGASmRA, N. MORIMOTO and T. HENm (1982) Spider mite fauna in pear and
peach orchards in Okayama. Od~kon-Chagoku 24:9-12 (in Japanese).
TAKAFUJI, A. and M. KAMmAYASHI (1984) Life cycle of a non-diapausing population of the two-spotted
spider mite, Tetranvchus urticae KOCH in a pear orchard. Res. Popul. Ecol. 26: 113-123.
UCmDA, M. (1980) Distribution of three species mites on pear orchards in Tottori district (Acarina:
Tetranychidae). Od6kon-Chflgoku 22 : 50-53 (in Japanese).
WANmUCHI, K. and Y. SAITO (1983) The process of population increase and patterns of resource utilization
of two spider mites, Oligonychus ununguis (JAeom) and Panonychus citri (McGREGOR), under experimental
conditions (Acari: Tetranychidae). Res. Popul. Ecol. 25:116-129.

157
~ ~ ~ - . ~ ~ ~ ~
Tetranychus ~ OD ~U~ r 2~r (4- "~ ~ ~ ~'=- T. urticae ~ ,~ ~.~ ~ ".1 -~ ~7 ~ ~ ~ L= 7". kan zawai ) OD ~ ~
3. 2 ~ F ~ c ~ : ~ • b, ~ ",,+Y~,,9"=@1~5 ; ~ - - . ~ . , S ~ c ~ , ~ ,~