requirement for a peptidoglycan recognition protein (pgrp ...pages.mtu.edu/~twerner/choe pgrp-lc...
TRANSCRIPT

/ www.sciencexpress.org / 28 February 2001 / Page 1/ 10.1126/science.1070216
Components of microbial cell walls are potent activatorsof innate immune responses in animals. For example, themammalian TLR4 signaling pathway is activated bybacterial LPS and is required for resistance to infectionby gram-negative bacteria. Other components ofmicrobial surfaces, such as peptidoglycan, are also potentactivators of innate immune responses, but less is knownabout how those components activate host defense. Herewe show that a peptidoglycan recognition protein, PGRP-LC, is absolutely required for the induction ofantibacterial peptide genes in response to infection inDrosophila and acts by controlling activation of the NF-κBfamily transcription factor Relish.
In response to infection, Drosophila activates transcription ofa battery of antimicrobial peptide genes in cells of the fatbody (the insect analogue of the liver). Two major branchesof this humoral response have been identified; as inmammals, these responses require NF-κB transcriptionfactors (1). One branch activates antifungal responses andrequires the receptor Toll and the NF-κB family transcriptionfactor Dif (2–4). The second branch, which is primarilyantibacterial, requires the NF-κB protein Relish, an IκBkinase, a caspase, a MAPKKK and the death-domain proteinImd (5–11).
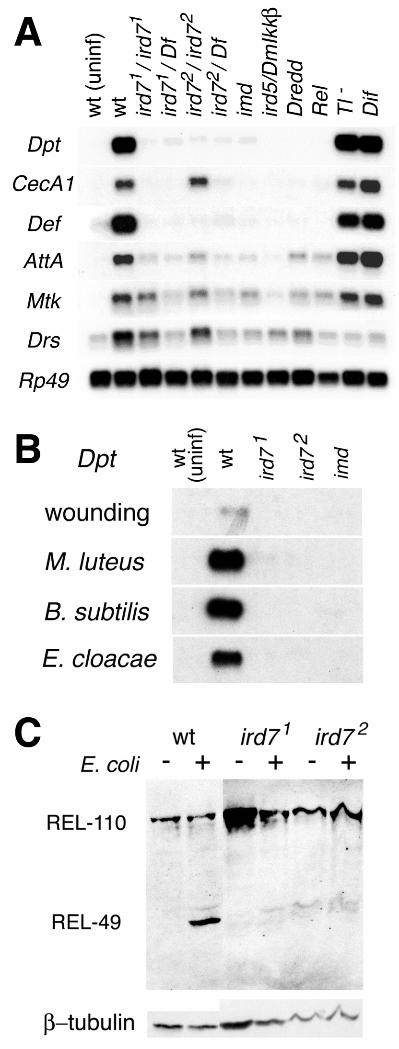
We have taken a genetic approach to identify genesrequired for the antibacterial response (12, 13). One gene thatis absolutely required for the induction of the antibacterialresponse is ird7 (immune response deficient 7). Twomutations in ird7, identified in an ethylmethane sulfonate(EMS) mutagenesis screen (12, 13), prevented induction ofthree antibacterial peptide genes, Diptericin, Cecropin andDefensin after infection by either gram-negative or gram-positive bacteria (Fig. 1, A and B). Three other antimicrobialpeptide genes, Attacin, Metchnikowin and Drosomycin, alsofailed to be induced to normal levels. The profile ofantimicrobial gene expression observed in the ird7 mutantswas similar to that observed in imd, DmIkkβ/ird5 and Relishafter bacterial infection, but distinct from that of Toll and Difmutants (Fig. 1A). This pattern suggests that ird7 is anessential component of the same signaling pathway thatrequires imd and Relish, but is not required for the Toll-Difpathway. Both ird7 mutants are homozygous viable andfertile, and blood cells from ird7 mutants can phagocytosebacteria (14), suggesting that ird7 is required specifically forthe humoral immune response.
The transcription factor Relish directly activatesantibacterial target genes in Drosophila. Relish is acompound protein similar to mammalian p100 and p105 (theprecursors of the p52 and p50 subunits of NF-κB), with an N-terminal Rel homology and a C-terminal ankyrin repeatdomain similar to IκB (15). In response to immune challenge,full-length Relish (REL-110) is endoproteolytically clipped togenerate the N-terminal REL-68 fragment, which translocatesinto the nucleus, and the C-terminal REL-49 ankyrin repeatfragment, which remains stable in the cytoplasm (16) (Fig.1C). In contrast to wild type animals, no processing of Relishwas detected in ird7 mutant larvae (Fig. 1C). The Rel domainof Relish failed to translocate to fat body nuclei in ird7mutants (17). These results indicate that ird7 is required forRelish processing and nuclear translocation.
Recombination and deficiency mapping localized ird7 to asmall interval on the third chromosome, 66F5-67A9 (Fig.2A). Based on the Drosophila genome sequence annotation,there are 12 genes in this region, including two genesencoding peptidoglycan recognition protein (PGRP) domains,PGRP-LA and PGRP-LC (18). Peptidoglycan is a strongactivator of innate immune responses in insects andmammals, and a PGRP was first identified in a silkmoth(Bombyx), based on its ability to bind peptidoglycan andactivate one aspect of the immune response, theprophenoloxidase cascade (19). Subsequent studies haveimplicated PGRPs in innate immune responses fromarthropods to mammals (20, 21).
We identified sequence changes that would disrupt thefunction of PGRP-LC in both ird7 alleles. The gene wasrepresented by several EST clones that encode a single spliceform, designated PGRP-LCa. In addition, sequencesencoding two additional exons encoding PGRP domains (“x”and “y”) were identified in an intron of PGRP-LC (18). Wescreened a larval-pupal cDNA library with the “x” and “y”exons and identified an alternatively spliced form of PGRP-LC that included the “x” exon; we call this isoform PGRP-LCx. Both PGRP-LC isoforms encoded type IItransmembrane proteins with common N-terminalcytoplasmic and transmembrane domains, but differentextracellular domains. The extracellular PGRP domains of thetwo isoforms were only 38% identical (55 of 145 residues).Northern hybridization with a common PGRP-LC exon proberevealed transcripts approximately 2.0kb in size in wild typelarvae, but no transcript of that size in ird71 mutant animals;instead a larger transcript of lower abundance was detected(Fig. 2B). Sequence analysis revealed an insertion of 858 bpof single copy sequence into exon 2, which is the first coding
Requirement for a Peptidoglycan Recognition Protein (PGRP) in Relish Activationand Antibacterial Immune Responses in DrosophilaKwang-Min Choe,1,2 Thomas Werner,3 Svenja Stöven,3 Dan Hultmark,3 Kathryn V. Anderson1,2*1Molecular Biology Program, Sloan-Kettering Institute, Memorial Sloan-Kettering Cancer Center, 1275 York Avenue, NewYork, NY 10021, USA. 2 Molecular and Cell Biology Program, Weill Graduate School of Medical Sciences, Cornell University,445 East 69th Street, New York, NY 10021, USA. 3Umeå Centre for Molecular Pathogenesis, Umeå University, SE-901 87Umeå, Sweden.
*To whom correspondence should be addressed. E-mail: [email protected]

/ www.sciencexpress.org / 28 February 2001 / Page 2/ 10.1126/science.1070216
exon in both isoforms, in the ird71 allele (Fig. 2C). Thisinsertion introduced a stop codon and would generate atruncated cytoplasmic protein. No sequence change in thePGRP-LCa isoform was identified in the ird72 allele.However, there was a G to A substitution in the “x” PGRPdomain in the PGRP-LCx isoform of ird72, which introduceda stop codon that makes a truncated protein lacking the last107 amino acids of this isoform (Fig. 2C). Because the ird72
allele alters only PGRP-LCx and has a profound effect onantimicrobial gene expression, this isoform must play acrucial role in vivo. The specific requirement for the PGRP-LCx isoform could be due its ability to specific ligands orbecause its expression is limited to specific cell types byregulated RNA splicing. Overexpression of either PGRP-LCcDNA rescued inducible expression of the Diptericin-lacZreporter gene in homozygous ird71 mutant animals (Fig. 3),confirming that the phenotype of ird7 mutants was the resultof the lack of PGRP-LC activity.
We used RNA interference (RNAi) to test the role ofPGRP-LC in the response to bacterial components. Treatmentof blood cells from the mbn-2 line with peptidoglycan, E. colior LPS led to a robust induction of the antibacterial peptidegenes. Introduction of double stranded RNA of PGRP-LC,but not PGRP-LA, effectively blocked induction ofDiptericin, CecropinA1 and AttacinA in response to all threestimuli (Fig. 4). Thus PGRP-LC is required for the responseto both peptidoglycan and LPS in these cells.
Because PGRP-LC is predicted to encode atransmembrane protein with an extracellular PGRP domain,PGRP-LC may act as a pattern recognition receptor that linksrecognition of microbial components with host immuneresponses (22). Because PGRP-LC is required for responsesto both peptidoglycan and LPS, the extracellular domain ofPGRP-LC may bind both peptidoglycan and LPS and bindingof either ligand may activate downstream signaling events.Alternatively, PGRP-LC may bind peptidoglycan, and notLPS, and act as an essential subunit of a larger complex thatincludes other pattern recognition receptors that bind LPS. Inmammals, signaling by Toll-like Receptor 2 (TLR2) isactivated by peptidoglycan (23). PGRP-LC might act in acomplex with another transmembrane protein similar toTLR2.
Twelve PGRP genes have been identified in theDrosophila genome (18). Another Drosophila gene, PGRP-SA, encodes a soluble peptidoglycan recognition protein thatis essential for activation of the Toll signaling pathway inresponse to infection by gram-positive bacteria (21). FourPGRP genes have been identified already in the humangenome (24). Given the evolutionary conservation of manyproteins required for innate immune responses, it will beimportant to evaluate whether PGRPs function as a family ofpattern recognition receptors in human innate immuneresponses.
References and Notes1. R. S. Khush, F. Leulier, B. Lemaitre, Trends Immunol. 22,
260 (2001).2. B. Lemaitre, E. Nicolas, L. Michaut, J.-M. Reichhart, J. A.
Hoffmann, Cell 86, 973 (1996).3. X. Meng, B. S. Khanuja, Y. T. Ip, Genes Dev. 13, 792
(1999).4. S. Rutschmann et al., Immunity 12, 569 (2000).5. M. Hedengren et al., Mol. Cell 4, 827 (1999).6. Y. Lu, L. P. Wu, K. V. Anderson, Genes Dev. 15, 104
(2001).
7. M. Elrod-Erickson, S. Mishra, D. Schneider, Curr. Biol.10, 781 (2000).
8. S. Rutschmann et al., Nature Immunol. 1, 342 (2000).9. F. Leulier, A. Rodriguez, R. S. Khush, J. M. Abrams, B.
Lemaitre, EMBO Rep. 1, 353 (2000).10. S. Vidal et al., Genes Dev. 15, 1900 (2001).11. P. Georgel et al., Dev. Cell 1, 503 (2001).12. L. P. Wu, K. V. Anderson, Nature 392, 93 (1998).13. L. P. Wu, K.-M. Choe, Y. Lu, K. V. Anderson, Genetics
159, 189 (2001).14. K.-M. Choe, N. Matova, K. V. Anderson, unpublished
data.15. M. S. Dushay, B. Åsling, D. Hultmark, Proc. Natl. Acad.
Sci. U.S.A. 93, 10343 (1996).16. S. Stöven, I. Ando, L. Kadalayil, Y. Engström, D.
Hultmark, EMBO Rep. 1, 347 (2000).17. See supplemental material at Science Online
(www.sciencemag.org/cgi/content/full/1070216/DC1).18. T. Werner et al., Proc. Natl. Acad. Sci. U.S.A. 97, 13772
(2000).19. H. Yoshida, K. Kinoshita, M. Ashida, J. Biol. Chem. 271,
13854 (1996).20. D. Kang, G. Liu, A. Lundström, E. Gelius, H. Steiner,
Proc. Natl. Acad. Sci. U.S.A. 95, 10078 (1998).21. T. Michel, J. M. Reichhart, J. A. Hoffmann, J. Royet.
Nature 414, 756 (2001).22. C. A. Janeway, Cold Spring Harbor Symp. Quant. Biol.
54, 1 (1989).23. O. Takeuchi et al., Immunity 11, 443 (1999).24. C. Liu, Z. Xu, D. Gupta, R. Dziarski, J. Biol. Chem. 276,
34686 (2001).25. L. S. Hatton, K. O’Hare, Elsevier Trends Journals
Technical Tips Online T01816, http://tto.trends.com(1999).
26. B. Chen, T. Chu, E. Harms, J. P. Gergen, S. Strickland,Genetics 149, 157 (1998).
27. A. H. Brand, N. Perrimon, Development 118, 401 (1993).28. A. C. Spradling, in Drosophila: A Practical Approach, D.
M. Roberts, Ed. (IRL, Oxford, 1986).29. D. A. Harrison, R. Binari, T. S. Nahreini, M. Gilman, N.
Perrimon, EMBO J. 14, 2857 (1995).30. S. M. Hammond, E. Bernstein, D. Beach, G. J. Hannon,
Nature 404, 293 (2000).31. K.-M. Choe, T. Werner, S. Stöven, D. Hultmark, K. V.
Anderson, data not shown.32. We thank R. Artero and P. Morcillo for technical advice,
B. Lemaitre for Drosophila and bacterial stocks, P. J.Lewis for bacterial stocks, D. Ferrandon, N. Perrimon andthe Drosophila Stock Center for Drosophila stocks, and T.Bestor for helpful comments on the manuscript. This workwas supported by grants from the NIH and the LitaAnnenberg Hazen Foundation to KVA and from the GöranGustafsson Foundation for Scientific Research, theSwedish Natural Science Research Council and theSwedish Medical Research Council to DH and theSwedish Natural Science Research Council to SS.
25 January 2002; accepted 19 February 2002Published online 28 February 2002;<zdoi;10.1126/science.1070216>Include this information when citing this paper.
Fig. 1. Phenotypes of ird7 mutants. (A) In ird7 mutants,Diptericin (Dpt), CecropinA1 (CecA1), Defensin (Def),AttacinA (AttA), Metchnikowin (Mtk), and Drosomycin (Drs)

/ www.sciencexpress.org / 28 February 2001 / Page 3/ 10.1126/science.1070216
transcription is not induced normally after E. coli infection, asassayed by Northern hybridization. ird71 is a very strong ornull allele, whereas ird72 behaves like a strong hypomorph.RNA was prepared from adult flies 6h after infection asdescribed (13). The loading control was Ribosomal protein49(Rp49). Similar results were obtained in larvae (31).Genotypes: wt: wild type (the parental P{w+ Dpt-lacZ} castock); Df: Df(3L)29A6; imd: imd1; ird5/DmIkkβ: ird51;Dredd: DreddD55; Rel: RelishE20; Tl-:Df(3R)Tl9QRX/Df(3R)roXB3; Dif: Dif1. For quantitation, see(17). (B) ird7 mutants fail to respond to both gram-negativeand gram-positive bacteria. Adult flies were pricked with asterile glass needle (wounding), or injected with Micrococcusluteus, Bacillus subtilis (gram-positive), or Enterobactercloacae (gram-negative), incubated for 6h, and total RNAswere prepared. Rp49 was the loading control (31). Theinduction of other antibacterial peptide genes by thesebacteria in ird7 and imd mutants was also very similar to thatshown in panel (A) (31). (C) Relish is not endoproteolyticallyprocessed after infection in ird7 mutants. Protein extractsfrom the wild-type parental stock (P[w+ Dpt-lacZ]ca), ird71
and ird72 were prepared from uninfected (-) or infected (+)wandering third instar larvae 30 minutes after E. coli injection(16). Protein from approximately 0.5 larva was loaded in eachlane. After blotting, Relish processing was detected with amonoclonal antibody that recognizes the C-terminal ankyrinrepeat domain of the protein. β-tubulin was the loadingcontrol.
Fig. 2. Molecular identification of the ird7 gene. (A) Geneticmapping of ird7. The ird7 mutation failed to complementDf(3L)29A6, but complemented Df(3L)Rdl-2 and Df(3L)AC1.Deficiency breakpoints were defined by single-embryo PCR(25). P-element induced male recombination mapping (26)placed ird7 locus between boule and EP(3)3043. Gray barsindicate the region that could include ird7. At all steps ofmapping, X-gal staining was used to monitor induction ofDpt-lacZ after E. coli infection. (B) Expression of PGRP-LCin wild type and ird7 mutants. 4 µg poly(A)+ RNAs preparedfrom wild-type (P[w+ Dpt-lacZ]ca) and ird7 adults wasloaded in each lane. Blots were hybridized with aradiolabeled probe from the second exon of PGRP-LC, whichis common to both splice variants. α-tubulin was the loadingcontrol. (C) Molecular lesions in PGRP-LC in ird7 mutants.The ird71 allele is associated with an insertion of 858 bp in acommon 5’ exon of PGRP-LC that introduces a stop codonand would generate a truncated cytoplasmic protein of 105amino acids. The ird72 is associated with a nonsense mutationin the “x” PGRP domain of the PGRP-LCx isoform, whichwould truncate this isoform. Shaded bars represent thetransmembrane domain. Speckled bars representpeptidoglycan recognition domains. To clone PGRP-LCx, alarval-pupal cDNA library (LP library from BerkeleyDrosophila Genome Project) was screened using a random-primed probe for putative exon “x” (18).
Fig. 3. Both PGRP-LCa and PGRP-LCx isoforms rescueinduction of Dpt-lacZ reporter gene in ird7 mutants. Fulllength PGRP-LCa and PGRP-LCx cDNAs were cloned intothe pUAST (w+) transformation vector (27) and introducedinto y w flies by P element-mediated transformation (28). Thesecond chromosome c564-GAL4 line, which is expressed inthe fat body and other tissues (29), was used to driveexpression of the UAS construct. Flies of indicated genotypeswere injected with E. coli (A to D), incubated for 6h, andassayed for β-galactosidase activity using X-gal. (A) c564-
GAL4/CyO; ird71 Dpt-lacZ/ ird71 Dpt-lacZ (no UAS-cDNA)animals did not express the reporter gene. (B) UAS-PGRP-LCx/CyO; ird71 Dpt-lacZ /ird71 Dpt-lacZ (no GAL4 driver)did not express the reporter gene. The same result wasobtained for UAS-PGRP-LCa/CyO; ird71 Dpt-lacZ /ird71
Dpt-lacZ animals. (C) c564-GAL4/UAS-PGRP-LCa; ird71
Dpt-lacZ/ird71 Dpt-lacZ expressed the reporter gene at highlevels after infection, as did c564-GAL4/UAS-PGRP-LCx;ird71 Dpt-lacZ/ird71 Dpt-lacZ animals (D). The GAL4-driventransgenes also showed a low level of constitutive expressionof Dpt-lacZ without E. coli injection (E and F). (E) c564-GAL4/UAS-PGRP-LCa; ird71 Dpt-lacZ/ ird71 Dpt-lacZ. (F)c564-GAL4/UAS-PGRP-LCx; ird71 Dpt-lacZ/ ird71 Dpt-lacZ.In four repetitions of this experiment, the level of X-galstaining in animals carrying both c564-GAL4 and the UAS-PGRP-LC transgene was greater in infected than inuninfected animals.
Fig. 4. Inactivation of PGRP-LC by transfection of doublestranded RNA (dsRNA) blocks induction of antibacterialgene expression in mbn-2 cells. Northern blot detection ofDiptericin, Cecropin A1 and Attacin A in mbn-2 cells aftertreatment with dsRNA from PGRP-LC, PGRP-LA or lacZand induction with the indicated elicitors. Ethidium bromidestaining of ribosomal RNA was used as a loading control.mbn-2 cells were plated at a density of 1 million cells/ml andtransfected one day later with 10 µg dsRNA (30). For PGRP-LA the dsRNA corresponded to 935 bp from exons 2-5; forPGRP-LC the dsRNA corresponded to 861 bp from thecommon exons 2 and 3. Three days after transfection, thecells were induced with insoluble peptidoglycan fromMicrococcus luteus for 6h, live Escherichia coli (O55:B5) for6h, LPS from Escherichia coli (O55:B5) for 2h, or sterileRinger (-) as control. The pellet of an E. coli overnightculture was resuspended 1:100 in sterile Ringer, and 15 µlwere used per induction. Peptidoglycan and LPS had a finalconcentration of 1 µg/ml. The cells were harvested after 2 or6 hours and total RNA was extracted. The loss of PGRP-LAand PGRP-LC mRNA due to RNA interference wasconfirmed by RT-PCR in a separate experiment. Drosomycinexpression is not inducible in this mbn-2 cell line, so theeffect of PGRP-LC RNAi on its expression could not beassessed in this experiment.