relationship between integrated and nonintegrated viral dna in
TRANSCRIPT

JOURNAL OF VIROLOGY, June 1980, p. 615-6260022-538X/80/06-0615/12$02.00/0
Vol. 34, No. 3
Relationship Between Integrated and Nonintegrated ViralDNA in Rat Cells Transformed by Polyoma VirusSEBASTIANO GATTONI,t VITTORIO COLANTUONI, AND CLAUDIO BASILICO*
Department of Pathology, New York University School ofMedicine, New York, New York 10016
Fischer rat fibroblasts transformed by polyoma virus contain, in addition toviral sequences integrated into the host genome, nonintegrated viral DNA mole-cules, whose presence is under the control of the viral A gene. To understand themechanism of production of the "free" viral DNA, we have characterized theDNA species produced by several rat lines transformed by wild-type virus or byts-a polyoma virus and compared them with the integrated viral sequences. Everycell line tested yielded a characteristic number of discrete species of viral DNA.The presence of defectives was a very common occurrence, and these moleculesgenerally carried deletions mapping in the viral "late" region. The production ofmultiple species of free viralDNA was not due to heterogeneity ofthe transformedrat cell population, and its pattern did not change upon fusion with permissivemouse cells. Analysis of the integrated viral DNA sequences in the same cell linesshowed, in most cases, a full head-to-tail tandem arrangement of normal-size anddefective molecules. The free DNA produced by these lines faithfully reflectedthe integrated species. This was true also in the case of a cell line which containeda viral insertion corresponding to -1.3 polyoma genomes, with each of therepeated portions of the viral DNA molecule carrying a different-size deletion.These results support the hypothesis that the free DNA derives from theintegrated form through a mechanism of homologous recombination leading toexcision and limited replication.
Infection of rat fibroblasts with polyoma virusand selection for growth in agar result in theisolation of transformants which contain a vari-able number of polyoma DNA equivalents inte-grated into the host cell genome and whichexpress polyoma T antigen(s). In addition tointegrated viral DNA, the transformed rat celllines contain unintegrated "free" viral DNApresent in the nuclei mainly as supercoiled mol-ecules (15). The average number of these freeviral DNA molecules ranges from 10 to about 60per cell, but only a minority of the cell popula-tion contains free viral DNA at any given time(19).Rat fibroblasts transformed by a ts-a polyoma
mutant (6) which codes for a temperature-sen-sitive large T antigen (10, 11, 17) produce freeviral DNA at the permissive temperature(330C), whereas they do not express it at thenonpermissive temperature (39.50C). However,even after months of propagation at a high tem-perature, the shifting of ts-a transformants to33°C results in the rapid reappearance of freeviral DNA molecules (19). This experimentshowed that free viral DNA production was
t Present address: Institute of Cancer Research, ColumbiaUniversity College of Physicians and Surgeons, New York,NY 10032.
under the control of the A gene function andthat the free molecules derived from the inte-grated ones (19). In addition, the demonstrationthat loss of integrated viral genomes occurredwith high frequency in the rat transformantsunder conditions permissive for free viral DNAproduction suggested that this process involvedexcision of integrated viral DNA molecules (1).To understand the mechanism of production
of nonintegrated viral DNA molecules, we in-vestigated the structural relationship betweenfree and integrated forms of polyoma DNA inthe rat transformants. The results presented inthis paper show that the free viral DNA pro-duced in these cells faithfully reflects the mainspecies of integrated viral DNA, which are gen-erally present in a head-to-tail tandem arrange-ment (1, 3). Although the free DNA is oftenheterogeneous, every cell line produces a char-acteristic number of discrete species of viralmolecules. The results obtained are in agree-ment with the hypothesis that the free DNAderives from the integrated form rather thanbeing maintained in a nonintegrated "plasmidic"state.
MATERLILS AND METHODS
Cels and viruses. Swiss mouse 3T3 ceUs and615
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.

616 GATTONI, COLANTUONI, AND BASILICO
F2408 polyoma-transformed rat cells were used. Thegeneral properties of these cell lines have been de-scribed previously (15, 19). Py 54, Py 53, and Py 5 arepolyoma wild-type (WT) transformants; ts-a 13, H3,H5, and H6A were transformed by the ts-a mutant ofpolyoma virus (6). Cells were grown in Dulbecco-mod-ified Eagle medium containing 10% calf serum. Ratcells transformed by the WT virus were propagated at37°C, and both free viral and chromosomal DNAswere extracted from cells grown at this temperature.ts-a transformants, instead, were generally propagatedat 39.5°C. For the preparation of free viral DNA,subconfluent cultures were shifted to 33°C and keptat the lower temperature for 3 to 4 days, before ex-traction of the low-molecular-weight DNA. Chromo-somal DNA from ts-a transformants was extractedfrom cells grown at the higher temperature (39.5°C),which is nonpermissive for the production of free viralDNA. Polyoma virus infectivity was titrated by plaqueassay on 3T3D monolayers at 37°C for the WT virusand at 33°C for the ts-a mutant.
Preparation of DNA. For the analysis of nonin-tegrated viral DNA, low-molecular-weight DNA wasextracted from the cells by the method of Hirt (9).The Hirt supernatant was extracted twice with phenoland then precipitated twice with ethanol at -20°C.The DNA was then dissolved in 10 mM Tris-hydro-chloride (pH 7.9)-10 mM NaCl-0.5 mM EDTA anddialyzed against the same buffer before restrictionendonuclease digestion. Chromosomal DNA was pre-pared as described by Gros-Bellard et al. (8) withslight modifications.DNA blotting. The following restriction enzymes
(New England Biolabs) were used: BgllI, EcoRI,BamI, HaeII, HindIII, HincII, HgaII, PstI, HhaI, andPvuII. Digestion of DNA samples was carried outunder assay conditions suggested by the manufacturer,at 37°C for 4 to 5 h. The reaction was stopped byadding 10 mM EDTA-0.1% sodium dodecyl sulfate-5% glycerol-0.01% bromophenol blue. The sampleswere loaded on a 1% agarose slab gel, 0.3 cm thick forfree viral DNA and 0.6 cm thick for chromosomalDNA. The gel (14 by 15 cm) was run for 18 to 20 h at15 to 20 V. After alkali denaturation, the DNA wastransferred from the gel to nitrocellulose filter paper(18) as described by Kettner and Kelly (13). After thetransfer, the nitrocellulose paper was rinsed in 6x SSC(lx SSC is 0.15 M NaCl plus 0.015 M sodium citrate)and dried under vacuum at 80°C for 8 h. Beforehybridization, the filter was pretreated for 6 h at 65°Cin 6x SSC containing 0.2% each of polyvinylpyrroli-done (PVP 360; Sigma Chemical Co.), Ficoll 400 (Phar-macia Fine Chemicals, Inc.), and bovine serum albu-min as described by Denhardt (5). The filter was thenhybridized in Denhardt solution containing denatured32P-labeled nick-translated polyoma DNA (10 to 20ng/ml) in sealed plastic bags for 20 h at 65°C in thepresence of 50 ILg of sonicated and denatured calfthymus DNA per ml. The 32P-labeled, nick-translated(12) polyoma DNA had a specific activity ranging from5 x 107 to 2 x 108 cpm/yg and was prepared asdescribed by Rigby et al. (16). After hybridization, thefilter was rinsed at the same temperature with threechanges of 25 mM phosphate buffer-1.5 mM NaPPi-0.05% sodium dodecyl sulfate in 2x, lx, and 0.5x SSC,respectively.
Finally, nitrocellulose paper was dried and exposedat -70°C against XR-2 X-ray film (Eastman KodakCo.) with intensifying screen (Lightning-plus; Du PontCo.).Fusion of transformed rat cells with permis-
sive cells. Transformed rat cells were mixed with 3T3mouse cells at a 1:1 ratio and plated in 100-mm petridishes at 2 x 106 cells per plate. After 1 day, the cellswere fused with polyethylene glycol 1000 (14). Cellswere incubated for variable times after fusion, andthen processed for virus or for DNA extraction asdescribed previously (1). Fusion was performed at33°C for cells transformed by the ts-a mutant (previ-ously grown at 39.5°C) or at 37°C for cells transformedby the WT virus.
RESULTSCharacterization of the free viral DNA
molecules. To gain information on the natureof the free viral DNA molecules, we screenedseveral lines of F2408 rat cells which had beentransformed by either theWT or the ts-a mutant(6) of polyoma virus. Although, in general, theresults obtained did not indicate any differencebetween WT- and ts-a-transformed cells withrespect to the type or number of species of freeviral molecules produced, our reasons for con-centrating on ts-a-transformed cells were thefollowing: (i) since ts-a transformants do notproduce free DNA at 39.5°C, but do so upon ashift to 330C (19), the production of free DNAin these cells could be induced rather synchro-nously by shift-down; (ii) WT-transformed cells,and ts-a-transformed cells at 33°C, upon pro-longed culturing, show a decreased productionof free viral DNA, a phenomenon also observedby Birg et al. (3). This decrease can be avoidedby maintaining ts-a transformants at 39.5°C.Under such conditions we observed that thesecell lines always produced constant amounts offree DNA upon shift-down.The DNA blot technique (18) was used to
analyze the free viral DNA. Low-molecular-weight DNA was extracted from the cells by themethod of Hirt (9), digested with restrictionenzymes, size fractionated by agarose gel elec-trophoresis, transferred to nitrocellulose paper,and hybridized with 32P-labeled polyoma DNA.Figure 1 shows a restriction map of polyomaDNA, indicating the coordinates of the cleavagesites of the restriction endonucleases that wehave mainly used (7). We will describe here indetail the results obtained with the most repre-sentative cell lines.
Figure 2 shows the comparison of the freemolecules derived from the ts-a H6A line (evenlanes) with a preparation of purified polyomavirus DNA used as a marker (odd lanes). Lanes1 and 2 show the pattern of untreated molecules,which migrated as expected from circular super-coiled form I (the fastest) or relaxed form II
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.

VIRAL DNA IN POLYOMA-TRANSFORMED RAT CELLS 617
1 2 3 4 5 6 7 8 9 10
3.4_ _- -- ::i.1.4_ ;i~
FIG. 1. Map ofpolyoma DNA (=3.4 x lt' daltons),showing the sites of cleavage of the restriction en-
zymes used in most of these experiments (from refer-ence 7).
molecules. When polyoma DNA was digestedwith EcoRI or BamI (lanes 3 and 5, respec-
tively), only one band corresponding to a fulllinear molecule (form Ill) of 3.4 x 106 in molec-ular weight was observed. HindIII (lane 9),which cuts polyoma DNA twice, gave two bands,corresponding to about 57 and 43% of the viralgenomes, respectively. The free DNA extractedfrom ts-a H6A gave the same pattern as themarker, and this could also be seen with otherenzymes, such as HindcI (lanes 7 and 8). Thiscell line, therefore, produces free moleculeswhich appear to have no gross alterations. Al-though the presence of some minor forms ofviral DNA was detectable in the blots, it is clearthat the predominant species of free viral DNAproduced by this line is a full-length polyomaDNA molecule.The free DNA of ts-a 13 (another cell line
transformed by the ts-a polyoma mutant) re-
vealed elements of further complexity (Fig. 3).In ts-a 13, there were at least two species offormI molecules (Fig. 3A, lane 2); the slower-migrat-ing one was indistinguishable from the marker(Fig. 3A, lane 1), whereas the second one mi-grated faster. By digestion with EcoRI, BamI,or HaeII (Fig. 3A, lanes 4, 6, and 8, respectively),we obtained two bands, one of which still mi-grated as did the marker, whereas the othermoved faster, indicating that the smaller mole-cule had a deletion. With HincII (Fig. 3A, lane10), only the A fragment originating from thefull-length molecule was visible and migrated as
did the marker (Fig. 3A, lane 9). The small
FIG. 2. Analysis of the free polyoma DNA ex-tracted from the ts-a H6A rat cell line. Low-molecu-lar-weight DNA was extracted from ts-a H6A cellsgrowing at 33°C, and the DNA preparation wasdigested with various restriction endonucleases asdescribed in the text. The digestion products werefractionated on 1% agarose gels run for 16 h at 15 V.At the end of the run, the DNA was denatured andtransferred to nitrocellulose filters, and specific pol-yoma DNA sequences were detected by hybridizationwith 32P-labeled polyoma DNA, followed by autora-diography. Odd lanes: purified polyoma DNAmarker. Even lanes: H6A low-molecular-weightDNA. (1 and 2) Undigested; (3 and 4) digested withEcoRI; (5 and 6) digested with BamI; (7 and 8)digested with HincII; (9 and 10) digested withHindIII. The arrows indicate the molecular weights(xl-6) ofsome ofthe linearDNA fragments contain-ingpolyoma DNA.
species remained uncleaved and was barely vis-ible as a supercoiled molecule migrating as didthe untreated ts-a 13 small molecule (Fig. 3A,lane 2). The HindIII digest (Fig. 3B, lane 4), inaddition to the two bands corresponding to themarker, showed a third band which appeared tohave exactly the same mobility as the faster oneobtained by EcoRI digestion (Fig. 3B, lane 3),suggesting that the small molecule had been cutby HindIII only once. Therefore, the smallerspecies of ts-a 13 DNA was missing both of theHincII sites and one of the HindIII sites.
In conclusion, ts-a 13 appears to produce two"families" of free DNA molecules, one of whichis indistinguishable from the polyoma marker,whereas the second one has a 35% deletion that,with the use of other restriction enzymes (datanot shown), has been mapped to extend fromapproximately map coordinates 12 to 47. Thisdeletion extends into the early DNA sequencecoding for the C-terminal of large T antigen.The free viral DNA described so far was pro-
duced from polyoma-transformed rat cell linesfrom which we have been able to rescue infec-tious polyoma virus by fusion with permissive3T3 mouse cells. However, this was not the casewith ts-a H3, a cell line transformed by ts-apolyoma. Figure 4 shows the restriction pattern
VOL. 34; 1980
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.
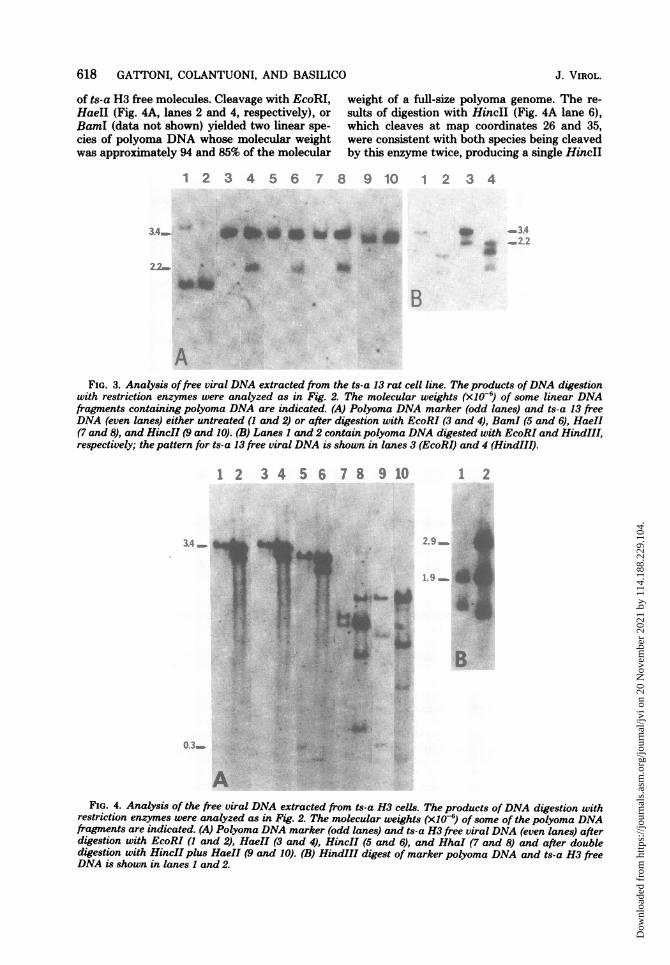
618 GATTONI, COLANTUONI, AND BASILICO
of ts-a H3 free molecules. Cleavage with EcoRI,HaeII (Fig. 4A, lanes 2 and 4, respectively), orBamI (data not shown) yielded two linear spe-cies of polyoma DNA whose molecular weightwas approximately 94 and 85% of the molecular
weight of a full-size polyoma genome. The re-sults of digestion with HincII (Fig. 4A lane 6),which cleaves at map coordinates 26 and 35,were consistent with both species being cleavedby this enzyme twice, producing a single HincII
1 2 3 4 5 6 7 8 9 10 1 2 3 4
**b. is-to| 4 j .. g _3411*_L.E~~0ahwdl
-' eb
B
AFIG. 3. Analysis of free viral DNA extracted from the ts-a 13 rat cell line. The products ofDNA digestion
with restriction enzymes were analyzed as in Fig. 2. The molecular weights (x10-6) of some linear DNAfragments containing polyoma DNA are indicated. (A) Polyoma DNA marker (odd lanes) and ts-a 13 freeDNA (even lanes) either untreated (1 and 2) or after digestion with EcoRI (3 and 4), BamI (5 and 6), HaeII(7 and 8), and HincII (9 and 10). (B) Lanes 1 and 2 contain polyoma DNA digested with EcoRI and HindIII,respectively; the pattern for ts-a 13 free viral DNA is shown in lanes 3 (EcoRI) and 4 (HindIII).
1 2 3 4 5 6 7 8 9 10 1 2
2.9.. . _
a
AFIG. 4. Analysis of the free viral DNA extracted from ts-a H3 cells. The products ofDNA digestion with
restriction enzymes were analyzed as in Fig. 2. The molecular weights (X10-) of some of the polyoma DNAfragments are indicated. (A) Polyoma DNA marker (odd lanes) and ts-a H3 free viral DNA (even lanes) afterdigestion with EcoRI (1 and 2), HaeII (3 and 4), HincII (5 and 6), and HhaI (7 and 8) and after doubledigestion with HincII plus HaeII (9 and 10). (B) HindIII digest of marker polyoma DNA and ts-a H3 freeDNA is shown in lanes 1 and 2.
.:V
."U
0.3
J. VIROL.
22. 0
Win0
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.
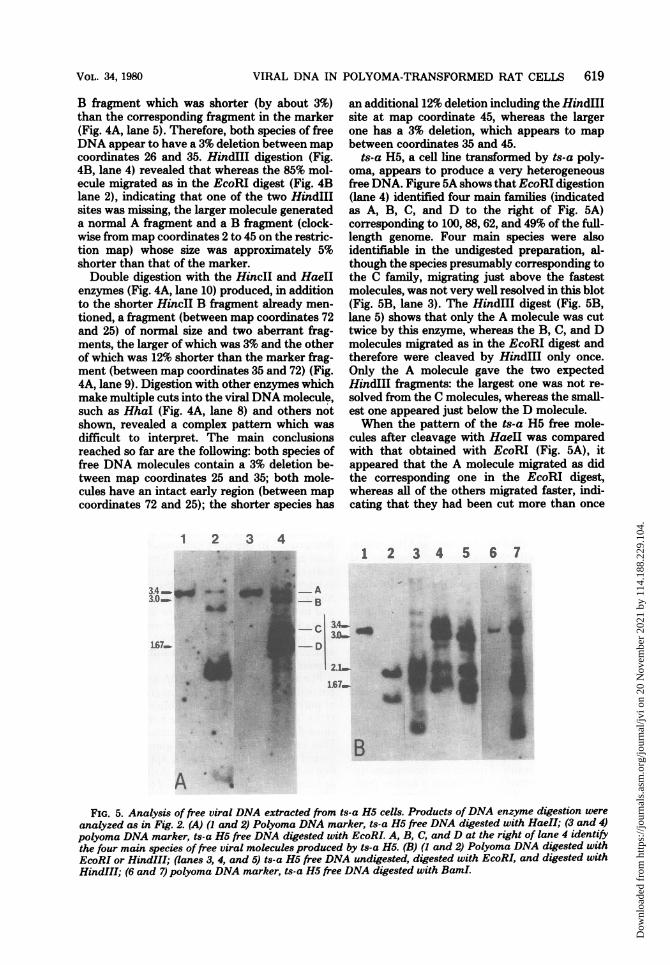
VIRAL DNA IN POLYOMA-TRANSFORMED RAT CELLS 619
B fragment which was shorter (by about 3%)than the corresponding fragment in the marker(Fig. 4A, lane 5). Therefore, both species of freeDNA appear to have a 3% deletion between mapcoordinates 26 and 35. HindIII digestion (Fig.4B, lane 4) revealed that whereas the 85% mol-ecule migrated as in the EcoRI digest (Fig. 4Blane 2), indicating that one of the two HindIIIsites was missing, the larger molecule generateda normal A fragment and a B fragment (clock-wise from map coordinates 2 to 45 on the restric-tion map) whose size was approximately 5%shorter than that of the marker.Double digestion with the HincII and HaeII
enzymes (Fig. 4A, lane 10) produced, in additionto the shorter HincII B fragment already men-tioned, a fragment (between map coordinates 72and 25) of normal size and two aberrant frag-ments, the larger of which was 3% and the otherof which was 12% shorter than the marker frag-ment (between map coordinates 35 and 72) (Fig.4A, lane 9). Digestion with other enzymes whichmake multiple cuts into the viral DNA molecule,such as HhaI (Fig. 4A, lane 8) and others notshown, revealed a complex pattem which wasdifficult to interpret. The main conclusionsreached so far are the following: both species offree DNA molecules contain a 3% deletion be-tween map coordinates 25 and 35; both mole-cules have an intact early region (between mapcoordinates 72 and 25); the shorter species has
1 2 3 4,I.:L '#
167.
an additional 12% deletion including the HindIIIsite at map coordinate 45, whereas the largerone has a 3% deletion, which appears to mapbetween coordinates 35 and 45.
ts-a H5, a cell line transformed by ts-a poly-oma, appears to produce a very heterogeneousfree DNA. Figure 5A shows that EcoRI digestion(lane 4) identified four main families (indicatedas A, B, C, and D to the right of Fig. 5A)corresponding to 100, 88, 62, and 49% of the full-length genome. Four main species were alsoidentifiable in the undigested preparation, al-though the species presumably corresponding tothe C family, migrating just above the fastestmolecules, was not very well resolved in this blot(Fig. 5B, lane 3). The HindIII digest (Fig. 5B,lane 5) shows that only the A molecule was cuttwice by this enzyme, whereas the B, C, and Dmolecules migrated as in the EcoRI digest andtherefore were cleaved by HindIII only once.Only the A molecule gave the two expectedHindIII fragments: the largest one was not re-solved from the C molecules, whereas the small-est one appeared just below the D molecule.When the pattern of the ts-a H5 free mole-
cules after cleavage with HaeII was comparedwith that obtained with EcoRI (Fig. 5A), itappeared that the A molecule migrated as didthe corresponding one in the EcoRI digest,whereas all of the others migrated faster, indi-cating that they had been cut more than once
1 2 3 4 5 6 7
-AB
-D
2.1.
1.67.
B
FIG. 5. Analysis of free viral DNA extracted from ts-a H5 cells. Products ofDNA enzyme digestion wereanalyzed as in Fig. 2. (A) (I and 2) Polyoma DNA marker, ts-a H5 free DNA digested with HaeII; (3 and 4)polyoma DNA marker, ts-a H5 free DNA digested with EcoRI. A, B, C, and D at the right of lane 4 identifythe four main species of free viral molecules produced by ts-a H5. (B) (1 and 2) Polyoma DNA digested withEcoRI or HindIII; (lanes 3, 4, and 5) ts-a H5 free DNA undigested, digested with EcoRI, and digested withHindIII; (6 and 7) polyoma DNA marker, ts-a H5 free DNA digested with BamI.
VOL. 34, 1980
h.A
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.

620 GATTONI, COLANTUONI, AND BASILICO
by HaeII. In fact, at the bottom of the gel, aband of small molecular weight (about 4 x 105)appeared, suggesting that the defective mole-cules contain a reduplication of a small DNAsegment including the HaeII site. Since theHaeII site is very near the origin of replicationof polyoma DNA, reduplication of the origincould explain the unusual abundance of thesedefective molecules in the ts-a H5 cell line. InFig. 5B, the cleavage of ts-a H5 free DNA withBamI (lane 7) shows that only the A moleculeswere cleaved by this enzyme, as inferred fromtheir migration with respect to the polyomamarker, whereas the other molecules were prob-ably not cleaved at all and migrated as super-coiled form I DNA. In conclusion, it appearsthat ts-a H5 produces four main species of freeDNA, one of which is a full-size molecule,whereas the other three appear to contain areduplication of a 10% fragment containing theHaeII site and variable size deletions.We also analyzed the free viral DNA produced
by three cell lines transformed by WT polyomavirus. In all three cases, we found a patternsimilar to that described for ts-a 13, i.e., produc-tion of two main species of free DNA, consistingof full-size and defective molecules. The dele-tions in the defective molecules ranged from 18%for Py 5 to 50% for Py 53. As in the case of thets-a transformants, these deletions mappedmainly, but not exclusively, in the viral "late"region (Table 1). The general features of the freeDNAs obtained from both WT and ts-a trans-formants appear to be the same, and the resultsfrom all the cell lines examined are summarizedin Table 1.Constancy of the free DNA production.
We wanted to determine whether the multiplespecies of free DNA produced by some cell linescould have been the result of population heter-ogeneity, with each cell type producing one typeof molecule. Therefore, we isolated three sub-clones from ts-a H5, propagated them at 39.50C,and then shifted them to 330C. The species offree viral DNA produced was examined by blothybridization. All three clones produced multi-ple species of free viral DNA, whose pattern wasessentially identical to that shown for ts-a H5(Fig. 5) (data not shown). Therefore, the com-plexity of the free DNA produced by the ts-a H5line was not a result ofpopulation heterogeneity.We also wanted to determine whether poly-
ethylene glycol fusion (14) of transformed ratcells with permissive mouse cells (15) could havealtered the pattern offree viralDNA production.Under these conditions, the number of free mol-ecules increased up to 100-fold by 24 h afterfusion (data not shown). In all cases tested, the
TABLE 1. Species offree viral DNA produced bypolyoma-transformed rat cell lines
Mol wt (% of full-size poly-oma genome) of fre viral IntegatedCell line DNA species speciesI II III IV
ts-a H6A 100 Its-a 13 100 65a I, IIWT Py 54 100 60b I,1IWT Py 53 100 50 NDCWT Py 5 100 82d NDts-a H5 100 88e 62f 49f I, II, IVts-a H3 949 85h I-IIa Deletion from map coordinates 12 to 47.bDeletion from map coordinates 4 to 44.c ND, Not determined.d Deletion including the BamI site.'Deletion including the BamI site and the HindIII
site at map coordinate 45.f Deletion including the BamI site, the HindIII site
at map coordinate 45, and both HincII sites.g Two 3% deletions, one mapping between map co-
ordinates 35 and 45 and the other mapping betweenmap coordinates 25 and 35.
h A 3% deletion mapping between map coordinates25 and 35 and a 12% deletion including the HindIIIsite at map coordinate 45.
characteristic species produced by each cell linedid not change; thus, it appears that fusion in-duced the same type of free DNA which isproduced spontaneously with lower efficiency.Analysis of the polyoma DNA sequences
integrated in the host genome. From the celllines in which the free viral DNA molecules havebeen characterized, the chromosomal DNA wasextracted and analyzed, using the Southern (18)blotting technique, to determine the presence ofand to study the arrangement of integrated pol-yoma sequences. All ts-a-transformed cell lineswere grown at 39.5°C before DNA extraction toeliminate possible problems resulting from thepresence of free DNA.For some cell lines (ts-a H6A and ts-a 13, in
particular) an analysis of the integration patternof polyoma DNA has been published elsewhere(1, 2). Therefore, we summarize here the conclu-sions derived from those experiments. The ts-aH6A cell line, which contains about three inte-grated viral DNA equivalents, appears to haveone polyoma DNA insertion in the host genome,as indicated by the appearance of only one bandwhen the chromosomal DNA is digested withBglII or HpaI, neither of which cuts polyomaDNA. This insertion contains a tandem repeatof full-length polyoma molecules since EcoRI,HaeII, or BamI digestion reveals a band in theposition of the full linear molecule plus two
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.

VIRAL DNA IN POLYOMA-TRANSFORMED RAT CELLS 621
others presumably corresponding to "linker" se-quences. Digestion with a variety of enzymeswhich cut polyoma DNA more than once alwaysproduces normal viral fragments with two otherscorresponding to the linker sequences. Thus, ts-a H6A appears to have integrated full-lengthpolyoma molecules in a head-to-tail tandem ar-rangement.
ts-a 13 also probably contains one polyomainsertion with a tandem arrangement of severalfull-length genomes and defective ones. The de-fective molecule has a 35% deletion encompass-ing the two HincII sites and the HindIII site atmap coordinate 45, identical to the one found inthe shorter species of free DNA produced bythis cell line.
In an attempt to establish a structural corre-lation between integrated and free DNA, wefound a straightforward correspondence in bothts-a H6A and ts-a 13. The former had full-sizeintegrated genomes and produced full-size freemolecules; the latter produced two families offree viral molecules (one full size and the otherdefective), and we found the same molecularspecies integrated in the cell DNA in a tandemrepeat. From these data, it would seem obviousto infer that each free DNA species is a faithfulcopy of a corresponding integrated form, and theintegration pattern of Py 54 supports this con-clusion. Blotting analysis of chromosomal DNAof Py 54 (Fig. 6) showed that when it was di-gested with BglII (lane 2), two bands were ob-tained (both of which appeared to be more than107 in molecular weight), indicating an equiva-lent number of insertion sites. Upon digestionwith EcoRI, BamI, or HaeII (lanes 4, 6, and 8,respectively), two major bands of viral DNAwere resolved; one corresponded to a full-sizemolecule, whereas the other had a 40% deletionwhich extended approximately from map coor-dinates 4 to 44 on the physical map, as it lackedthe two HincII sites (data not shown) and twoHhaI sites, but not the HindIII sites at mapcoordinates 2 and 45. In fact, the HhaI digestionpattern (lane 10) showed that the full-lengthmolecule was cleaved in the same manner as themarker (lane 9), whereas the defective moleculeappeared to be cut only once and migratedequally to the EcoRI-, HaeII-, or BamI-digestedmolecules. In the HindIII digest (lane 12), it wasclear that, in addition to linker sequences, theonly fragments generated were identical to thoseof the marker. However, the A fragment was ofan abnormally high intensity, suggesting that itderived from the cleavage of both defective andfull-size genomes. Thus, this situation was simi-lar to that of ts-a 13, and, as in ts-a 13, thetandemly integrated molecules represented the
1 2 3 4 5 6 7 8 9 10 11 12
V.,,4
.,- I%w
1 .r"i
FIG. 6. Analysis ofpolyoma DNA sequences inte-grated in the DNA of the Py 54 cell line. Approxi-mately 10 pg of high-molecular-weight DNA per slotwas loaded on a 1% agarose slab gel after restrictionenzyme digestion. The gel was run overnight at 20 V;the DNA was denatured and transferred to a nitro-cellulose filter. Specificpolyoma DNA sequences weredetected by hybridization with denatured 32P-labelednick-translated polyoma DNA and visualized by au-toradiography. The positions and molecular weight(x10-6) of some linear DNA fragments containingpolyoma DNA are indicated. (1 and 2) Purified pol-yoma DNA marker and Py 54 DNA digested withBglII; (3 and 4) polyoma DNA marker, Py 54 DNAdigested with EcoRI; (5 and 6)polyomaDNA marker,Py 54 DNA digested with BamI; (7 and 8) polyomaDNA marker, Py 54 DNA digested with HaeII; (9and 10) polyoma DNA marker, Py 54 DNA digestedwith HhaI; (11 and 12) polyoma DNA marker, Py 54DNA digested with HindIII.
main species of free viral DNA.The integration pattern of ts-a H5 revealed
analogies with ts-a 13 and Py 54. Digestion withBglIl indicated that ts-a H5 had two and possi-bly three polyoma insertions (Fig. 7, lane 1)since, of the two bands, the slower-migrating onewas likely to have been a doublet. The EcoRIdigest (lane 3) showed several bands, two ofwhich appeared to hybridize more strongly. Thelarger one migrated as did the marker (lane 2);the smaller one had an apparent molecularweight of 3 x 106, identical to that of the Bmolecules in the free DNA (see above). TheHindIII digest (lane 5) gave the expected twofragments from the full monomer. (The faintnessof these two fragments is an artifact of thisparticular blot, since in other experiments thesebands always had the expected intensity.) How-ever, the defective molecule was apparently cutonly once by this enzyme (as was the case forthe free B molecules) and migrated as in the
VOL. 34, 1980
0 0 - 6I %
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.

622 GATTONI, COLANTUONI, AND BASILICO
1 2 3 4 5 6 7 8 9 10 digest identical to that shown by B and D freets-a H5 molecules upon digestion with the HaeIIenzyme (Fig. 5A, lane 2). We interpret this re-sult, as in the case of the free DNA produced bythis cell line, as indicating that these molecules
3.4_ _ are cleaved more than once by HaeII because3.0 ^* * 2.6 they contain a reduplication of the origin of
replication. The reduplicated DNA fragment is,1.67_. in fact, detectable as a faint band with an appar-
ent molecular weight of 4 x 105. This represents-_ _1. 25 the difference in molecular weight between the
HaeII-digested B and D molecules and the cor-responding EcoRI products. In conclusion, ts-aH5 contains two or three separate insertions oftandemly integrated viral DNA molecules, con-
}04 sisting of a full-length monomer and two distinctdefectives, both of which, however, have a re-duplication of an =11% fragment of the viralgenome containing the HaeII site. These three
FIG. 7. Analysis ofpolyoma DNA sequences inte- species correspond to the A, B, and D moleculesgrated in the DNA of the ts-a H5 cell line. The identified in the free DNA produced by this cellproducts of restriction enzyme digestion were ana- line but no species corresponding to the C freelyzed as in Fig. 6. (1) ts-a H5 DNA treated with BglII '(which does not cleave polyoma DNA); (2 and 3) molencea r to b integalto ed,ss n bypolyoma DNA marker, ts-a H5 DNA digested wi the absence of 2.1 x 106-dalton molecules n theEcoRI; (4 and 5) polyoma DNA marker, ts-a H5DNA EcoRI digest (Fig. 7, lane 3).digested with HindIII; (6 and 7) polyoma DNA The integration pattern of the ts-a H3 linemarker, H5 DNA digested with BamI; (8 and 9) was unusual in many respects, including the factpolyoma DNA marker, ts-a H5 DNA digested with that this was the only polyoma-transformed cellHaeII. The molecular weights (x10-6) of some of the line examined in which the viral DNA was notlinear DNA fragments containing polyoma DNA are integrated in full tandem. The BglII digest of ts-indicated by the arrows. Lane 10 contains HaeII- a H3 chromosomal DNA shows only one banddigested DNA extracted from a subline of ts-a H5which shows the 4 x 105-molecular-weight fragment (Fig. 8A, lane 4), idicating that only one poly-more clearly, because of what appears to be an am- oma insertion was present in the host genome.plification of sequences corresponding to the species With EcoRI and HaeII (Fig. 8A, lanes 2 and 4,with a molecular weight of 1.25 x 106 after HaeII respectively), no bands migrating at the positiondigestion (compare lanes 9 and 10). of a monomer were observed, but only two bands
of variable molecular weight were observed, pre-EcoRI digest. Another band with an apparent sumably the results of a single cleavage in themolecular weight of 1.6 x 106 (corresponding to viral DNA insertion, together with cleavage inthe molecular weight of the D molecules present the flanking host sequences.in the free DNA [Fig. 5]) showed the same Cleavage with HincII (Fig. 8A, lane 6), on thebehavior. The BamI digestion (lane 7) resolved, other hand, produced a band similar, but notof the three monomers, only the full-size one. identical, in size to the marker HincII A frag-Several bands of high-molecular-weight DNA ment (lane 5), suggesting that a continuousare also visible; some of them probably corre- stretch of viral DNA from map coordinates 35spond to the entire insertions of defective mol- to 26 is integrated in this cell line, but that thisecules, which, as expected from the characteri- fragment may contain a deletion. Double diges-zation of the ts-a H5 free DNA, appear to lack tion, with HincII plus HaeII (Fig. 8B), confirmedthe BamI site. BamI, therefore, behaves as a the existence of a 3% deletion between mapnoncutter for these molecules. The only band coordinates 35 and 72, and the HindIll digestion,with a molecular weight lower than 3.4 x 106 which generated a normal-size A fragment (mapappearing in the lane is likely to represent a coordinates 45 to 2; Fig. 8A, lane 12), allowed uslinker sequence of host and viral DNAs. to map this deletion more precisely betweenThe HaeII digest (lanes 9 and 10) showed map coordinates 35 and 45. In addition, Fig. 8C
three intense bands, only one of which migrated (lane 4) shows that the small HinclI fragmentas did the marker (lane 8). The other two bands was also present in the integrated viral DNA,exhibited an increased mobility with respect to but, as observed in the free DNA produced bythat shown by the other monomers in the EcoRI this cell line, was shortened by about 3% with
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.

VIRAL DNA IN POLYOMA-TRANSFORMED RAT CELLS 623
1 2 3 4 5 6 7 8 !
_~ _
A
1 2 1 2
d 1.83
.1.25
B
a
a
cFIG. 8. Analysis of polyc
grated into the DNA of tiproducts of restriction enz:
lecular-weight DNA werecposition and molecular weilinear polyoma DNA fragnand 2) Polyoma DNA marA
with EcoRI; (3 and 4) polycDNA digested with BglII;marker, ts-a H3 DNA digespolyoma DNA marker, ts-BamI; (9 and 10) polyoma ldigested with HhaI; (11marker, ts-a H3 DNA diges14) polyoma DNA marker, tHaeII. (B) (1 and 2) PolyoDNA after double digestioi(C) (1 and 2) Polyoma DAdigested with BamI; (3 andts-a H3DNA digested withDNA marker, ts-a H3 DNand 8) polyoma DNA ma
double digestion with Hinc.migrating bands in lanesof incomplete digestion.
respect to the marker (FThus, these data sugg
cell line contained an inquence corresponding t(genome (map coordinatbut containing a 3% deliordinates 26 and 35 an
910 11 12 13 14 between map coordinates 35 and 45, a situationsimilar to that found in the larger species of freeDNA produced by this cell line.The BamI digestion shown in Fig. 8A (lane 8)
V 34 produced a strong band, corresponding to -85%- 2.9 of the size of the viral genome, and a fainter
band of lower molecular weight. This could havebeen indicative of a situation like that observedfor the HaeII and EcoRI digestions (i.e., the
, 1.36 presence of only one site of cleavage for thisenzyme in the integrated viral DNA sequences),but further analysis with more sensitive probesand shorter gel runs (Fig. 8C, lane 2) revealedthe presence of a third band with a molecular
3 4 5 6 7 8 weight of -7 x 105. Since the largest viral DNAfragment produced by BamI was of the samemolecular weight (85% of the polyoma genome)as the shorter species of free DNA produced by
y ~ 231 the ts-a H3 cell line, it seemed likely that thew. , 0, ""integrated viral DNA sequences in this cell line
extended clockwise from map coordinates 35 to-0o78 35 (HincII sites) and continued to map coordi-
nate 58 (BamI), thus including two BamI sites* * O0.31 spanning a viral DNA sequence containing ap-* * proximately a 15% deletion. This was confirmed
by analysis with KpnI (Fig. 8C, lanes 5 and 6),which showed the presence of the 52% KpnI
oma DNA sequences inte- fragment (map coordinates 59 to 11), but insteadhe ts-a H3 cell line. The of the normal 48% fragment (map coordinates 11yme digestion of high-mo- to 59), the analysis showed a band approxi-znalyzed as in Fig. 6. The mately 34% of the size of the polyoma genome,ight (x10-6) of some of the as would be expected from the presence of therients are indicated. (A) (1 above-postulated deletion. Combined HincIIter, ts-a H3 DNA digested and BamI digestion (Fig. 8C) showed that the:ma DNA marker, ts-a H3 fragment between map coordinates 58 and 265 (5 and 6) polyoma DNA was normal, but that instead of a normal-size
a H3 DNA digested with (map coordinates 35 to 58) fragment, two bands)NA marker, ts-atH3DNA which were approximately 3 and 10% shorterand 12) polyoma DNA were detected.sted with HindIlI; (13 and These results and those obtained by digestinges-a H3DNA digested with ts-a H3 DNA with other enzymes shown in Fig.Fma DNA marker, ts-a H3 9 have confirmed this interpretation and allowedn with HincII and HaeII. us to construct a model of the arrangement ofTA marker, ts-a H3 DNA integrated viral DNA sequences in this cell line4) polyoma DNA marker, (Fig. 9). The joining points between viral andHincII; (5 and 6)polyoma host DNAs are on the left side approximately at
'rker gts-a H3 DNAP after map coordinate 30 and on the right side at mapland BamI. The slowest- coordinate 60. Thus, a limited repeat (&30%) of
5 through 8 are the result viral DNA sequences is present in the integratedsegment. The repeated portions of the integratedviral DNA (map coordinates 30 to 60) each
Pig. 8C, lane 3). contain a deletion of a different size, one of aboutrested that the ts-a H3 3% between map coordinates 35 and 45 and the[tegrated viral DNA se- other of 10 to 12% including the HindIII mapD at least one polyoma coordinate 45 site and the map coordinate 52es 26 to 26, or 35 to 35) SstI site (data not shown). The central part ofetion between map co- the viral DNA insertion is unique and containsId another 3% deletion an intact early region (map coordinates 70 to 25)
VOL. 34, 1980
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.

624 GATTONI, COLANTUONI, AND BASILICO
L4550 60 70 lo 90 0 10 20 A 5
a
FIG. 9. Map of the viral DNA sequences integrated in the host genome in the ts-a H3 cell line. The mapwas constructed by analyzing the products of restriction enzyme digestion of high-molecular-weight DNAextracted from this line, as shown in Fig. 8. The restriction enzymes used and their sites of cleavage withinthe viral sequences are indicated. The heavy solid lines indicate cellular DNA sequences; the thin linerepresents integrated polyoma DNA. V indicates the regions of the polyoma map containing the junctionsbetween viral and host DNA. The map coordinate, indicated on the viral side of the junction, represents a
restriction enzyme site which is still present in the integrated viral sequence, whereas the map coordinate on
the host side of the junction represents the next restriction site ofpolyoma DNA which is not present in theintegrated sequences. A indicates a deletion. The thin lines (I and II) at the bottom indicate schematicallythe viralDNA sequences represented in the two major species offree DNA produced by this cell line. Symbols:0, BamI; 0, HaeII; A, HincII; A, HindIII; El KpnI; U, HhaI; O, PstI; *, PvuII.
and another 3% deletion between map coordi-nates 26 and 35.
Therefore, these data show that although a
single viral insertion can generate more than onespecies of free DNA, the characteristics of thefree DNA species are encoded in the integratedform, as the largest free DNA molecule corre-
sponds essentially to the unique integrated viralDNA sequences and the left side of the insertion,whereas the smaller species corresponds to a
sequence composed of the unique segment andthe right side of the insertion (Fig. 9).
DISCUSSIONThe results presented in this paper show that
polyoma-transformed rat cells, under conditionspermissive for large T antigen function, producediscrete species of free viral DNA. Each cell lineproduces a characteristic pattern of free viralmolecules, consisting of one or more (up to atleast four in the cell lines that we examined)major species. In the case of cell lines producingmultiple species, this is not due to heterogeneityin the cell population, although we cannot say
at this time whether each cell produces all pos-sible species of free DNA at the same time. Thefree molecules always reflect the molecular spe-
cies integrated in the same cell line, but in thecase of the ts-a H5 line, we observed thatwhereas three types of molecules were inte-grated and also expressed as free DNA, a fourthspecies was found only in the free DNA. Thesemolecules (designated as C species in the text)shared with the other defective species (B andD) a reduplication of an 11% segment ofthe viralDNA and contained the same-size deletion as
did the D molecules, but probably carried tworepeats of the above-mentioned reduplication(Fig. 5). Therefore, it is possible that they origi-nated from the D molecules, during either exci-sion or replication.
We have tested the effect of polyethyleneglycol-assisted fusion of the transformed rat cells
with permissive 3T3 mouse cells (14) on freeviral DNA production. In all cases tested, thepattern remained exactly the same, except forthe increased amounts of DNA produced. Thisresult could be due to the fact that fusion withpermissive cells either causes excision by a
mechanism identical to that operating at a lowlevel in the transformed rat cells or results onlyin the amplification of free viral DNA sponta-neously induced.As already reported, the most common feature
of polyoma integration in transformed rat cells
is the presence of a tandem, head-to-tail arrange-ment of full-length and defective viral DNAmolecules (1-3). In several cases, we have foundmore than one insertion. In the case of the ts-aH3 cell line, however, the polyoma sequenceswere integrated into the host DNA only as a
partial tandem. These integrated DNA se-
quences contained three deletions, located indifferent regions of the viral genome, that ac-
counted for the nonrescuability of the virus fromthis cell line after fusion with permissive mouse
cells. The location of these deletions, which werepresent in both the integrated and free viralDNA produced by this cell line, provided cluesas to the possible mechanism of production offree viral molecules from the integrated form.The two free viral DNA species had in commonthe 3% deletion mapping between map coordi-nates 26 and 35, whereas each of the others waspresent only in one of the two free viral DNAspecies. The arrangement of the integrated se-
quences was such that the shared deletion was
positioned centrally with respect to the others.Therefore, it appears that in ts-a H3 the largerfree DNA species is excised from the left side ofthe integrated sequences, and the shorter one isexcised from the right (Fig. 9). Ifwe assume that
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.

VIRAL DNA IN POLYOMA-TRANSFORMED RAT CELLS 625
the presence of homologous DNA sequences isnecessary for the excision event to occur, a par-tial tandem of integrated viral DNA would besufficient to allow homologous intramolecularrecombination and excision. When the peculiararrangement of integrated viral DNA sequencesin ts-a H3 is considered, the types of free viralmolecules produced would then depend onwhere the recombination event occurs. Theproperties of the ts-a H3 free DNA are consist-ent with this model.As stated before, the free DNA often contains
discrete species of deleted molecules. The pref-erential location of the deletions is in the lateregion of the polyoma genome, leaving un-touched both the origin of the replication andthe early region. In most cases this clearly de-rives from the integration of similarly defectivemolecular species. These defectives were notpresent in any substantial proportion in our viralpreparations (see the marker DNAs used in ourexperiments), and it is possible that they weregenerated during integration. The late localiza-tion of the deletions could be explained by thefact that most early deletions would probablynot confer a transformed phenotype to the cells.The only cell line which contained little morethan a single viral genome equivalent (H3 cellline) had a continuous stretch of integrated viralDNA spanning the origin of replication and theentire early region. In ts-a 13, the defective freemolecule had a 35% deletion which encompassedabout 15% of the early region, encoding the C-terminal part of the large T antigen. Accord-ingly, the 1OA-2 cell line, a partially cured re-vertant of ts-a 13 which has lost the insertion ofthe full monomers (1), is incapable of producingany free viral DNA and is T antigen negativewhen tested by immunofluorescence. It wouldappear that ts-a 13 is able to produce the defec-tive free molecules because a functional T anti-gen is produced by the inserted nondefectivemolecules.
It has been suggested (3) that the free viralmolecules, in addition to being the result ofinduction, could also be maintained in a nonin-tegrated plasmid-like state, but the evidencethat we have collected tends to rule out ratherthan support this possibility. In this paper wehave shown that there are no free moleculescarrying any specific marker which is not alsopresent in the integrated viral sequences (whichwould constitute evidence for a plasmid-likestate).
In situ hybridization experiments performedin our system showed that the free DNA isinduced in a minority of the cell population(approximately 0.5%) rather than a few mole-cules being produced in every cell (19). It seems
possible that under these circumstances inducedcells might be killed (or cured of inducible viralDNA), and, in fact, under conditions permissivefor free DNA production, we have observed aslow decrease, in time, in the average number offree equivalents per cell as assessed throughreassociation kinetics experiments. This wouldindicate a slight selection against cells producingfree molecules.With regard to the mechanism leading to free
DNA production, Botchan et al. (4) have sug-gested a model in which activation of large Tantigen by permissive cell factors leads to therecognition of the origin of replication of theintegrated viral DNA, followed by repeatedrounds of in situ replication. In this polytenicstructure, viral DNA could be freed by peelingoff of the newly replicated DNA strands or byother mechanisms. However, we have presentedevidence that "curing" (i.e., loss of integratedviral DNA) occurs at a rather high rate in thepolyoma-transformed rat cells under conditionspermissive for free viral DNA production (1).This finding showed that curing involves exci-sion and loss of integrated molecules and thatsuch a process depends on the presence of anactive large T antigen (2) as in the case of theproduction of free viral DNA. In addition, thedata presented in this paper strongly suggestthat recombination of homologous sequencesplays an important role in the production of freeviral DNA molecules. It is thus possible that,after replication of integrated viral DNA, intra-molecular recombination of homologous viralsequences takes place, leading to circularizationand excision. On the other hand, recombinationof integrated homologous viral sequences couldtake place without prior in situ replication. Thisprocess could be facilitated by the presence ofan active polyoma large T antigen, if this viralprotein had recombination-promoting activity.Both of these models predict that, after excisionand replication, molecules which are faithfulcopies of the integrated ones would be produced,favored by the tandem arrangement of inte-grated viral DNA sequences.
ACKNOWLEDGMENTSWe thank Dimitris Zouzias for helpful discussions and
Nancy Porzio, Lisa Dailey, and Eva Deutsch for their expertassistance.
This investigation was supported by Public Health Servicegrants CA 11893 and CA 16239 forn the National CancerInstitute. V.C. is on leave from II Istituto Di Chimica Biolo-gica, II Facolta' Di Medicina, University of Naples, Naples,Italy.
LITERATURE CITED
1. Basilico, C., S. Gattoni, D. Zouzias, and G. DellaValle. 1979. Loss of integrated viral DNA sequences inpolyoma-transformed cells is associated with an activeviral A function. Cell 17:645-659.
VOL. 34, 1980
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.

626 GATTONI, COLANTUONI, AND BASILICO
2. Basilico, C., D. Zouzias, G. Delia Valle, S. Gattoni, V.Colantuoni, R. Fenton, and L. Dailey. 1980. Integra-tion and excision ofpolyoma virus genomes. Cold SpringHarbor Symp. Quant. Biol. 44:611-620.
3. Birg, F., R. Dulbecco, M. Fried, and R. Kamen. 1979.State and organization of polyoma virus DNA se-quences in transformed rat cell lines. J. Virol. 29:633-648.
4. Botchan, M., W. Topp, and J. Sambrook. 1979. Studieson SV40 excision from cellular chromosomes. ColdSpring Harbor Symp. Quant. Biol. 43:709-719.
5. Denhardt, D. T. 1966. A membrane filter technique forthe detection of complementary DNA. Biochem. Bio-phys. Res. Commun. 38:126-133.
6. Fried, M. 1965. Cell transforming ability of a temperaturesensitive mutant of polyoma virus. Proc. Natl. Acad.Sci. U.S.A. 53:486-491.
7. Fried, M., and B. E. Griffin. 1977. Organization of thegenomes of polyoma and SV40. Adv. Cancer Res. 24:67-113.
8. Gros-Bellard, M., D. Oudet, and P. Chambon. 1973.Isolation of high molecular weight DNA from mam-malian cells. Eur. J. Biochem. 36:32-38.
9. Hirt, B. 1967. Selective extraction of polyoma DNA frominfected mouse cell cultures. J. Mol. Biol. 26:365-369.
10. Hutchinson, M. A., T. Hunter, and W. Eckhardt. 1978.Characterization ofT antigens in polyoma-infected andtransformed cells. Cell 15:65-77.
11. Ito, Y., N. Spurr, and R. Dulbecco. 1977. Characteri-zation of polyoma virus T-antigen. Proc. Natl. Acad.Sci. U.S.A. 74:4666-4670.
12. Kelly, R. B., N. R. Cozzarelli, N. P. Deutschman, L. R.Lehman, and Z. Kornberg. 1970. Enzymatic synthesisof DNA. XXXII. Replication of duplex deoxyribonu-cleic acid by polymerase at a single strand break. J.Biol. Chem. 245:39-45.
13. Kettner, G., and T. J. Kelly. 1976. Integrated SV40sequences in transformed cell DNA. Analysis usingrestriction endonucleases. Proc. Natl. Acad. Sci. U.S.A.73:1102-1106.
14. Pontecorvo, G. 1975. Production of mammalian somaticcell hybrids by means of polyethylene glycol treatment.Somatic Cell Genet. 1:397-400.
15. Prasad, I., D. Zouzias, and C. Basilico. 1976. State ofthe viral DNA in rat cells transformed by polyomavirus. I. Virus rescue and the presence of nonintegratedviral DNA molecules. J. Virol. 18:436-444.
16. Rigby, D. W. J., M. Dieckmann, C. Rhodes, and P.Berg. 1977. Labelling deoxyribonucleic acid to highspecific activity in vitro by nick-translation with DNApolymerase I. J. Mol. Biol. 113:237-252.
17. Silver, J., B. Schaffhausen, and T. Benjamin. 1978.Tumor antigens induced by nontransforning mutantsof polyoma virus. Cell 15:485-496.
18. Southern, E. M. 1975. Detection of specific sequencesamong DNA fragments separated by gel electrophore-sis. J. Mol. Biol. 98:503-517.
19. Zouzias, D., L. Prasad, and C. Basilico. 1977. State ofthe viral DNA in rat cells transformed by polyomavirus. II. Identification of the cells containing noninte-grated viral DNA and the effect of viral mutations. J.Virol. 24:142-150.
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
4.18
8.22
9.10
4.