regional variation in mtdna of the lesser prairie-chicken · i the col/dar li2(1 l"29···j7...
TRANSCRIPT

I
The COl/dar li2(1 l"29···J7 lJ;) The Cooper OrIlidlO!C)gtcal SOCii:ly 201U
REGIONAL VARIATION IN MTDNA OF THE LESSER PRAIRIE-CHICKEN
CllRISTIAN r\. H';'GLN1.S, J';'!\'lIS C. PlT1vIAN I6 , BRETT K. SANDERCOCK 1, DON H, Wour 2, RUIJI'.RT.J. ROlm.. ,
ROGlR D. AI'PLEG!\TEl.7, /\I';D SAR/\ .J. OVLLR-McCANCE4
iDivisioll ofBiology, Kansas Stare University, ManllOtfan, KS 66506 1C;. M. StI[lO/1 Aviun Research Cemer, Untvel'siry oj'Oklahoma, Bartlesville, OK 74005
'Rcsewch and Survey O/fi(',~. Kansas /)epurlment of Wildlife lind Parks. Emparia, KS 6630/ 4Rocky MOllntuin Ceillerjlll' Conservation Genetics (Jild Systemmics. u.s. Geological Survcy,
Dl'par1rnen/ olBiological Sciences, U/liversit)' 0/ Deaver, Dem'er. CO 80208
Ab"rroer. Cumulative loss of halmat and long-term declJne in the populations of the Lesser Prairie-Ch lekcn l1.i;mpalil/(:hus pallidicinctu.l') ha\'e led kl concern" lbr lhe species' viability lhroughout il, range in the Sl'ul'bern nreat Plains. For more efficienl conservation pa~l and pre~elil distributIOns of gen~lic vanatlOn need to be unJer$load. We examined the distribution of mitochondrial DNA (mtDNA) variation in the Lesser Prairie-Chicken aeross Kansas, Colorado, Oklah()ma, and New Mexico. Throughout the range we found lillie genetic differentiation exc;cpt f,))' the populatIOn in Ncw lV!exlco, which was Significantly different from most other popnlanon~. We did, however, find signl1i.eant isolation by distance at the rangewide scale (r =0.(98). We fOllnd no relationship between haplotype phylogelly and geogrnphy, and our analyses provide evidence for a posr-glacial population expansion wirh in thc species that i, consistent with the idea that speciarion within Tvmpanuchus is recent. Conservalion actions lhat incrcase lhe likelihood of genetically viable populations in th~ fulure should be evalua(~d J~H
implemcntation.
Kev l1'ordr: DNA, geneilc diversity, Lesser Pmirie- Chicken, mitochondrial DNA, prairie groU51', TympiHlUchus palhdici.nct us.
Estructura Genctica Regional del ADNmt en Tvmpanuchus pallidicincrus
Resumen. La pcrdida de habitat y Ja disminuci()n poblacional de largo plazo sufricla por Tymponuehns p((llidicilll.'nls ha condueido 3 prellcupaciones en lorno a la viabilidad de la cspecie a !o largl) de su distribL1CI()]) geograflc,j t:n 1<1 parl"e ·,ur de las (jrand~s PlillllCle~. Para 4LI~ la comervaeion ,,:a ma~ dit:ienle, es necesano entcnc!<::r ia distribucion pasada y presente de la variaci6n gel1ctiea. Exalllinamos 13 distribucion de la variacion en el ADN mitocmlc1rial (ADNmt) en T pallidiell/ctus ,I traves de Kansas, CoJorado, Ok lahoma y Nuevo Mexico. Encontramos poea diferclleiacion genetiea a 10 largo de la distribucion, exeepto para la poblaei6n do Nuevo Mexico que fur sign;ficalivamcntc dlfcrcnte de la nllyoria de las Q[ras poblaeioncs Sin embargo, cncontramos un patron significatlvo de ill,lamicnh) POI' Jislancia a 1,1 eseala de tada la distnbuci6n (r =' 0.698). No ellcontrmno~ rclacion erltre la Ii logema de haplotipo~ y la gcografia. Ademas, nucstros anatiSls provccn eVldencia dc una expansion poblaeional posl-glacla! de la espccle, 1o que eoncuerda con la idea de que la especiaci6n en ?jompanuchl/s es rccicn1c. Es neccsario cvaluar 1a posible implementaeion de aeciOlle, de conservaeion que inerementcn Ja probabilldad de contar con poblaeiollcs gcnctic:.unente viables en d futuro.
reduced egg ferrility and productivity In al Ieasl two populaINTRODUCTION
tions (West.etneicr et 31. 1998. Stiver cl al. 2008). Throughout the Holarctie Region grouse have declined dra The Lesser Prairie-Chicken U,vmpallllchus pallldh 111(.matically in reccnt years as a result of habitat loss, degrada IUS) is a lek-mating grouse occupying the southern Great tion. and fragmentation resulting from human land lISCS (e.g., Plains (including parts of KanSilS, Colorado, Oklahoma. si Ivicu!turc. agriculture, recreation, urbanizarion, Storch Texas, alld New Mexico) vegetated primarily with sand sage:2007). Habilal modificalion ilnd loss have affeeled di,~per:,al brush (tllNmisiu jilifuliu) and sand shinnery oak (Querctts end gene flow ill several species of grouse (Bouzat el al. 1997, havu/'{Iii) lHagen and Giesen 2005). Although the rilnge of Segelbilchcr et <11. 2003, Johnson el al. 2003, 2004, Oyler the Lesser Prairie-Chicken before European seltlcmcll{ ,va.s McCancc er a1. 2005a), and population deellncs have led to relatively large and contiguous, rhe currcnt distribution oflhis
ManUSCript reccived 5 August 2009; received 7 October 2009.
'Current address: Oreg,,!! Dcpanment of Fish and Wildllte, 61~74 Panel! Rd .. Bel!rl, OR 97702. f.-mail: 6Currcllt address: Rescarch and Survey Office. Kansas Depanrnetl( of Wildlife and Parks. EmpOria. KS 66g01. "Current address: Tennessee WiJdlifi: Resource Agency, Ellington Agricullmal Center. P.O. Box 40747, Nashville. Tenne»ee 37204
Tff(' C·)f1tiOJ. Vol 112, Number I. [Ja~es 29·37. tSSN 0010-5422, etec1r.onlc ISSN 1938~S4:!.2 ,;) ~O1O by The (o0pe.1' OrJJl1hD\ogical S"'c;ely. All nght.;: resened Ple:~;.ie dirt:ct
:lil rcqe::.;~1~ for r-('nnl~s:()il 1'(' phD~(,,>l;i)PY or n:·i>mduc.'{.· article t:\)E!k.nL lhrough HH; U01VL:r.,ny nfCu)1 rorlH.:1 rn::-;~ -'i R19hL~ ami Permi ...slol1:-' wcb::ite .
. DOl ,ff I52S,oond 1010 n90lS0
29

30 CHRTST1A"N A. HAGEN ET AL.
/
OkJahqma ,
"'I New Mexico '->
Colorado
,_.-_...-.....;-~ , Texas
R.....';":;",....,t, '
'i \
" ;J
~
FIGURE 1. Current rangc (dark gray) and Ilisronc rallg~ (jighl ~ra\) of rhe Le>s~r PraIne-Chicken (adapred trum Hagen and GIesen 2005). SamplIng locations (black dOl;) arc depicr~d wnhlJ\ c.OUl1ly bmindanes. Okiahoma and New Mexico 'liles arc from Van Den Bu;sch.; el a.l. (2003).
species has bc~CIl reduced tv <8%, of the hislonc distribution and is highly fragmented because of agriculture ilnd other human develvpmcllt (Fig. I, Hagcn and Giesen 20051. Much of the habitar loss has occurred in the center of the Lesser PralrieChicken's range, the Texas panhandle, with vast areas ofprainc no longer suitable for the species (Fig. I). The cumulative loss
ofhabitat, declining population trends (Table I), and imminent thrcats led to a recent increase in priority ranking of the 1995 listing as "warranted but precluded" under the Endangel cd Specics Act (USFWS 2(08).
The divergencc of 0'mpai1uchus spp. and p<)st-glaclal cx.panslOn of thcir distribution help explalll contcmpor~ry
pauerns of population genetic structltring (John,on 2008). These patterns, combined with current knowLedgc of habitat fragmentation and movement corridors, can elucidate rates of gene tlow ilnd popu lat ions at risk of reductions ill gcncli..:: diversity. The distribution of genetic variation and the structure ofeertaiu populiltions of the Lesscr Prairie-Chicken havc been described from Oklahoma and l"cw Mexicv by Van den Bussche et al. (2003) and Bouzat and lohman (2004). Bounl and Johnson (2004) investigated the Lesser Prairie-Chickcn's genetic structure ofat four leks in 1\ew MexLco and found no differentiation in llllDNA sequence. In a separatc study, Van den Bllssehe ct al. (2003) used nllDNA sequen..::e data to document gendie variation at 20 leks in Oklahoma and New Mexico and found reasonably high leveLs of genetic variation. Thc latter study found some regional structuring among leks within cach state and detecred a high level of differentliltion between popuiations from Oklahoma and New Mexico in both the mitochondrial and nuclear genomes, Van den Busschc et a1. (2003) suggested that habitat fragmentation and loss may have contributed to the genetic structuring observed, Both studle~ provided importilnt inf()rJnation ab<1ut local distrtbutions of genetic variation. A rangewide perspeet'l ve of genetle variatIOn, however, cum:ntly unkno\vn for rh..c Lcs~er Prairi..cChicken. is necessary for conservauon decislons at th;;: le\el of rhe specics. Rllngcwide gcnetie analyses have eOl1trihutcd to conservation and recovery strategies for related species such as the Greater (Cenfrocercus urophasianus) and Gunnison Cc. minimus) Sage-Grollse lOyler-\.feCance el al. 2005a, b).
Here, uSlllg mitochondrial D1\A (mtDNA) sequence data, wc greatly extend previous sl udies' range and density of sampiing ro address the distribution of genetic variation across most of the Lesser Prairie-Chicken's range. We bni!d on the initial work ofVil!) den Bussche el al. (2003) in Oklahoma and New Mexico to include samples from the remaining: portion
l~"BLE l. The Lcss~r Prairie-Chicken's population status (size and t.rend) in the five states lhat constit.uTe the species' current mnge. ----~----_._._._._-~_._--------~-----------_._------_. __._._-_.~------------
Breeding popnlalion SlaTIIS
Slate Trend
1980-2008 Rdcrcnee,
Colorado <1000 Declining Giesen (2000), Davis etal. (20081 Kansas IR 000-29 000 Stable1lnereasing Johllsgard (2002), Davis d al. (200R) New 1\1.:xil.:o 6000 D~'clJl1lng Johnsgard (2002), Davis el <II. (2008) Oklahoma <3000 [)e~hnillg .!<)hnsgard (2002), Dnvis et ill. (20()~)
Texas nOOO Deehning S:ivy el a!. (200~), Davis ct a1. (2008)

11
ofthc species' range in Kansas :lnd Colorado. The combined information will a Ilow for the dcvelopment ofa morc cohesive and erilclent conservation .<.t.raregy based on (he rang:ewide distribution ofgcnctic variation in the Lesser Prairie-Chicken.
METIIODS
STUDY ARF.'\S
Jn 2000 to 2002. we caplured Lesser Prairie-Ouckcns and collcctcd blood samples from thcm in Baca and Prowcrs counties in Colorado and six counties in Kansas, three north (Gove, Trego, and Ness) and three somh of the Arkansas River (Comanche. Finney, Kearny; Fig. I). Samples from Trego and Ness counties were combined beeausc of their geographic proximity and smali sample sizes from these arcas. We compared tbe birds' mtD1\A to thal of populations previously sampled by Van lkn Busschc ct al. 1200.1). We used mtDNA data from Van den Busst:he et a1. (200.1) but made comparisons among coumies insread of among kks and included ~Idrper County (pooling Harper [IZ '" 8], and Ellis [n:. 53] counties) and Beavcr County (n = 27), Oklahoma. and Rooscvelt County, ~ew
\Iex ieo (n =63; hereafcer ROl)sevelc). We lacked samples from populations in Texas but sampled adjat:ent popularions in New Mexico and Oklahoma (Fig. I).
TISSUE COLLFCTTO"i AND Dr-: A EXTRACTlON
We sampl~d blood from l6l Lesser Prairie-Chickens (144 maies. ]7 females). captured during the spring <,nd fan of 2000 to 2002 H; funncllraps In Kansas and Coloradll (l-Iaukos ct al. 1990). Blol)d samples were obtained by ellppi ng a t,le
nail of each prairie-chicken and collccting tWO or three drops orblthld into a microfuge Wbe previollsly coated wiLh EDT.-"I (Brinkman). All blood samples werc stored frozen at -20"e. DN.-"I was extracted from blood by either a phenol- ch loroform method (Kahn ct al. 1999) or the Wizard Genomic DNA Purification System (Promega), according to the manullldurcr's instructions,
DI---A S[QUEI---Cr~G
Fol]o\,' ing QUill 11 (1992), we ampl died <l pOri ion anhe mlDNA eomral regi')1\ In 25-IlL rCadll)f[' vdh the primers L16755 (N-::dbal e\ al. 19(7) and OSU77J3 (Van den Bussche el al. 20(3). The procedure consisted of preheatiug ill 94 "C for 2 min followed by 35 cycles ofdenaturariun at 94"C !i.lr 40 $C'::,
annealing at 55 "C for 60 sec, and ex tension at 72 °C for 2 min. peR produt:ts were ckaned \\lith ~hrimp alkaline phosphatase and exonuclease 1 (OSB). Dye terminator cycle sequencing reactions were perfurmed with the Beckman CoultCT Quick Start Sequencing Kit according to the manufacturer's protocol. The products wc:rc preeipitatc:d an:ording to the manufaClura\ ~recification" resu,pcnde-d in30 ~LL offormumiJe, and run on ,I Beckman CUllltc:r CEQ 8000 XL Datn Analysis Sy~telll by method LFR-b. Sequences (466 base pairs) were
PRATRIE-CIllCKEN CiENETlCS
aligned with Sequcncher 4.2 (GeneCodes) and eompsrcd to tbe sequences previously defined by Van dL:ll Bussch~ cl al. (2003).
D/\T/\ ANALYSES
We calculated haplotype diversity (h) and nucleotlde dl I'ersn) (n) of mtDNA tor each population wlth the program ARLEQUIN (version 2.000: Sc:hncider c[ al. 2000). We eXlimined popul31i\)rl structure \.... ilh an analysis of molecular variance (AMOVA) by investigating bow much varia lion \vas explained by thc separation of populations into tbur geographic regions. sand sagebrush prairie (Finney, Kcarny, Prowers and Bat:a). mixed-grass prairie (Gove and Ncss), mixed sbrub (Comanche. Beaver, and Harper), and sand shinncry oak (Rooscvelt). We used a Kimura (1980) lwo-parameler nndeotide-subsLitulioll model for all calculations. Pairwise C!\. valucs, mca;;ure, of genetic differentiation between each group. were L·akulaled. and thcir sigl1lficanee wa~ detcrrnmed by random permuralIOns. Patrwise $ST values among 10 populatlom were Cllnsider~d to be slgllificam if their BontcrTOrll-corrected P-valuc was <0.00 I. We exami ned the relationship between gcographic and genetic distance with a Mantel (1967) test. Wc calculated probabililies for the Mantel test by using methods of Smouse eta I. (1986).
To investigate the relationship among haplotypL:s we gGnerated an unrooted haplotype network with the sratistical parsimony software TCS vcrsion 1.13 (Clement et al. 2000). We used the algorithm of'Templelon et al. (1992) ro consrruet rhe nct\-I'\lfk
We invesrigated the possibil ity of a posl-glacial 5pc,'le,·
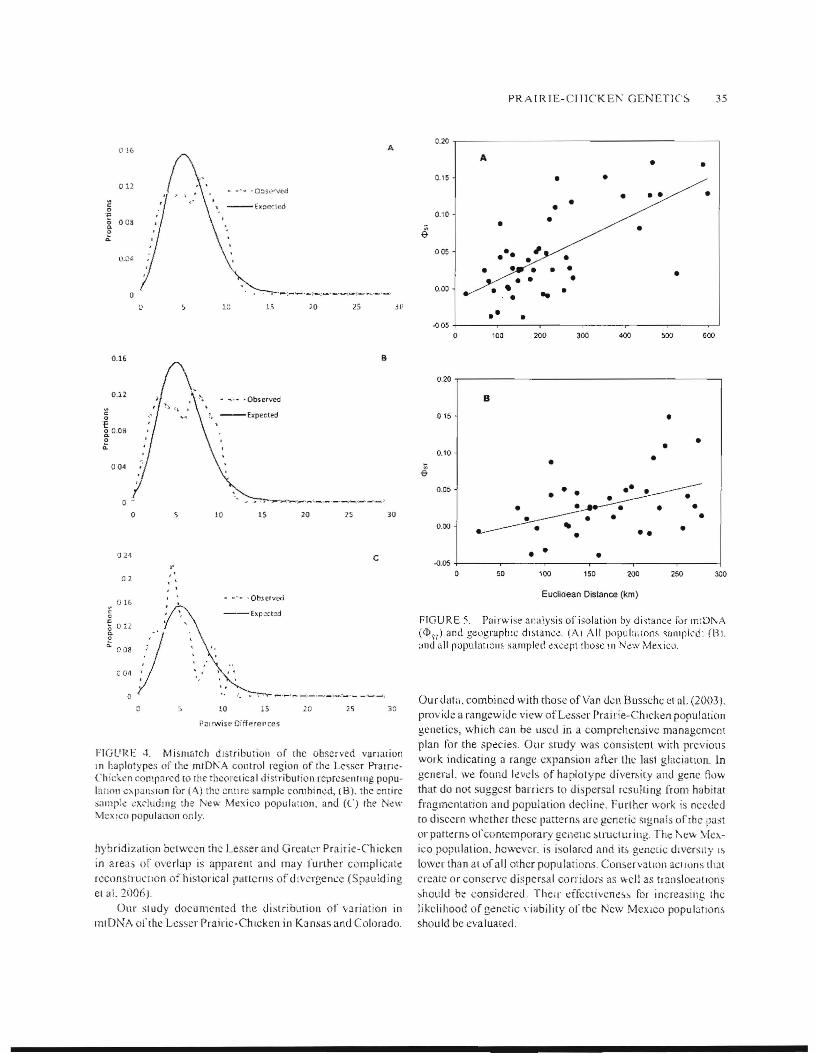
wide PllpUlal ion cxpansion by rneans of a rn ismateh distribullon ofpairwise genettc differencc~ in prognllm ARLFQUIN (Schneider ct al. 20(0) and DnaSP (version 3.4. Rozas and ROl.as 1999). DnaSP graphically compare~ the ()b~erYed and expected distriburions of populations at equilibriUln and cxpansions by using Rogers' (l995) method of moment,. We cxamined the mismatch (hstributiOIJ Ln three separate analy~es: the entire sample of birds. all samples but excluding )\e\\ il/rcx jeo, and New ~'1exico separately. 1t was important to scparale the New Mexico population because of its gClldic dlfferellces from Oklahoma populalions and its disjunct range (\',m den Bussehc Ct al 2003).
RESULTS
The mtDNA haplotypcs origiua1iy defined by Van den Bussche ct aJ. (2003) included a run of either seven or eight Ts aL tbe beginning of the sequence. Because of ambiguity of where to place the deletion, wc truncated the sequence by one' base pair. This deletion result.ed in our classifying a~ identical two hap]orypes (J and Z) originally defined as distinct by Van den Bussehe el al. (2003). Thus we refer to.T and Z a~ a single haplotype (1) (Table 2). From lndividuuls of the four st:ltcS eombi m:d (Ii ,~ 278). \.I·e ld<::nLJ fied 43 h<lplo(ypc~. 30 (,r LilL:tl\

32 CIlRISTfAN;\ fJAGEN ET AL
TABl.E 2. HJpktypes <llld dletr frequency [1\ JO populatwl1s from 4 regJOns throughoul thc range of the Lesser Prairie-Chicken Haplotypes J\ through EE previously dcscribcd by Van dCll l3use..:he Cl al. (2003)
Gove. Ne". fl11ney. Kearny, Pr()\\·ers, [3,lca, Comanche. II'lrper. Ruosevdr. KS KS KS KS CO CO KS OK NM Frcqueney
A J 3 1. 20 27 B 2 2 2 5 4 10 26 C 1 4 2 J 6 5 12 31 D 3 3 E 2 S 1 13 F 7 7 G 1. 2 5 II If I 5 J 8 [ 2 4 6 l3 J 6 7 1 2 2 2 22. K I 5 6 L 4 4 1\1 4 4 !\ 4 2 I:; o :; :; II p 3 3 Q 3 3 R 2 2 S 1 J T I :2 U V W X 2 6 y :\;\ I 8[3 5 2 2 " CC I DO I EE I FF I I GG 5 2 3 IJ fill I JJ KK :2 5 LL 1 1. 00 2 4 PP \ I S5 2 .) {,
UU 2 VV 4 4 XX 4 ZZ 1 I T,)fal 29 10 35 24 iO 14 61 27 63
._----------_._---------
had previously de;;cribed by Vau den Bussche et al. (2003) several ambiguous connections resolved by the frequency and (Tabk 2). Sequences of th~ 4J hap!otypc~ have been deposited topology criterIOn (Fig. 2). There was no relationshIp betw~cn
in Genbank (acccssiunllumbers GU2(i9l58-GU~69200).Hap haplotypes and geography, and oil haploL·ype relationships Imypcs ofmtDNA ","erc charaelCrizcd by 39 polymorphic sites were relatively shallow. Haplotypes C, A, B, and J were the con;;isting of 32 Lransitions, 3 lransversions. J polymorphic most common (found in 52% of individuals ;;equenc~d) WIth sites comaining both transitions emu inscrtionidektions, and I J bcing the most widcsprl'ad. Huplotypes E. 1, and GO were polymorphic site with hath a transtrion and a transversion. also relatively common (Fig. J). Haplotype (' was found in
According to statistlca! parsimony, the 95';/0 plausible set al1 populutions except Govc, Ness. anu Prowcrs. while hapof the neL,vork comprised many haploLypes anu comatned lolype B was found everywhere birds were sampled except

u
fl(;URE 2 Unrooted estimated Q5% p:mimony d,ldl)gr<1ll1 of 43 haplotype, detected in the Les~cr Prairie-Chickens. I'bpl()type~ are represented by kiter;. Lines represcnl single llIutllrrOIJS~ dots represent imCIlnedi<lIC haplot)'pe; not found i.n our salnple hut necessary t<J Iink the haplotypes ohs.::rved.
Gove. Prowers. and Baea (Fig. 3). Haplotype A was found in high freqllcney in Roo~evelt and was absent from N~ss, Prowcrs, Baca, and Comanche, while haplotypc.T was found everywhere except Ness and PIowers (Fig. 3).
Wirhin-roplJhninn haplmype diversiry (11). whIch represents the number and frequency of haplotypes, was on average 0.916 (Sf == 0.04), bUI Roosevelt, New Yfexicn. had the lowest
_c =A =a=J=E=1 c:::::J GQ r=::::J non
,. a:E rr."« G ~ :>u z '" co .<
'" ~ iL.. '" 0
0 " '" Vl
0 u '" FIGURE 3. Proportion or mdividuals in each population with common haplotypes: "llon" represents the number of uncommon hapilltypes. E.lt:h bar represenls the proponion oflJllljvictuals in each populmion with rhese commonllar1orypes. The pre\·nJenec ofhaplolyres IS rlinked fTOm g.reares't 10 leas! as C, A, B. J. L E. and (iG. the r~nk. ofdlc last three being equal.
'$ (II ,. >. ., Z.u ,~ i.:..' a:
z " '";;; ill "' '"
PRATRTE-CHtCKEN GENETTCS 3J
TABl.E 3. Summary statistics of mtDNA divcrSlty across rhe range of the Lesser Pr~liric-Chlcken.
Populanon 11" AI· h'(SEJ )('1 (SE)
Gove, KS 29 17 0.948 to On) 0.011 (ll.O061 Ness. KS 10 9 0.978 (0 05-1) O.tll:1 (O.lIUS I Flnney_ KS 35 14 0.919(0021) 0.011 (0 0071 Kearny, KS 24 II 0887 (0.045 J 0.012 (0007) PlOwers, CO 10 7 0.933(0.062) 0013 (0 0(7) Baea. CO 5 4 0.900 (0.J611 0.005 (0.004) CDmal1eh~, KS 14 9 0912 (0059) 0.010 (0.006) Harper. OK 61 22 0.944 (0012) 0.012 (0 DO"!) Beaver, OK 27 12 0.912 (0.028) (J.OII (OOOfi) Roo,;evelt, NM 63 9 0.828 (00271 0.008 (0004)
----------_._-----
•Sample size. "Number ofhapiOlypes C Haplorype diversity. d Nucleotide diversity.
diver<;it)' lO.828~ Table 3). Nueleotide diver,ity (11:) in New Mexico (0.008) and Buca (0.005) wa, lower than the average of all populations (i 0:= 0.0! 08, SE 0.004). Gellt.:IIC diver5lty7.;
attrjbutable to variation amnng regions. among popu lations within a region, and within a populations was partitioned as 6.52%, 0.43%. and 93.04%, respectively. Pairwise CV ST tests indic:ated low levels of population ditTerentiarion with sigl1lficant ditferellces between Roosevelt and all other popUlatIOns except Saca, Prowers, and Comanche Cfable 4).
The mismatch distributions calculated among hapiotypeifor the t'ntirc' sample, the New Mexico population alone. and the entire species excludIng New Mexi.;o 'Were all unimodal and consistent with post-glacial range expanston (Fig. 4/\-C). In all three analy,cs the goodness-ol:'Ai tesb with the model for expandt ng population growth showed no significant di ITerene6 (emire sample: stun of squared deviations = ().OO~. p::= 0.67; New .vfexieo ollly: sum of squared deviations = 0.017, P "'. 0.31; excluding Ne\\o Mexico: sum of squared deviations = 0.004. P "" 0.40). We tound a positivc condation betwcen genetic distance «1)s'() and geographic distance (I' ::= 0.692, P =.
0.004; Fig. 5A). Subsequently. we analyzed isolation by di~tan('e
to control for the extreme geographic ;Jlld genetic (hstanees to
the New MeXICO poptllatlon.., and found ~lgl1ificanr l"J!.ltlOn by
distance (r = 0.417. P = 0.017) in thIS subset ofdarn (llg. 5B).
DISCUSSION
In the late PleIstocene, praine grouse (Tvmpanuchus spp ) likely e>:pcriellced a rapid expansion and diverstlkallon (1.1ICchini et aL 2001, Dw\'elski 2003, Spaulding el aL 2006, .Tohnson 2008). Furthermore, on the basis ofmorphology, behavior, and geography, if has been shown that speciation within this genus is recent and Iineage sorting among. the three spee!cs j~ incomplete (Ellsworth et al. [994, Spau Idi ng el 31 2006. Johnson ~008). Our data arc consistent with previolls I1ndIng~, as w::: found relalively hIgh levels of gentilc di\crsily

34 CITRTSTTAN A. 1TAGE"K ET .\t..
TA BLE 4. Pftll'WIse <1.\'1 values tor mlDN ..'\ s~quencing data {roln JO populalions of the Lesser Pmi ric-Ch icken in Kansas, Colorarlo, Ok lahoma. <1))(1 New ,YlcA ieo: Values of cf>w in hold ar~ SignificanTly diffcrent (Bollfcrron i correction P S O.OOll in palf\vi,c compartsons ;lCroSS
lhe :ip~cies' range.
NelV Colomdo Oklal10ma vloieo
Gove N~ss Finney Kearney Comanche Prmvcr~ Baea Harper Beaver Rooscvd! ---------_._~_._------------------------------
Uu~e
Nc;;:s -U.03787 Finlley 0.04250 0.00203 Kearny 0.05067 0.D1057 -OJI0690 Comanche 0.09340 0.D2615 0.(m:32 0.04947 Prowers 0.02654 -0.DOi57 -003216 0.01012 002767 flaea 0.1481'1 0.11694 0.02444 0.00025 0.01449 0.08740 Harper O.()·1167 --.() OU<J66 001271 0.02514 --O.U0285 ---U.00234 0.02529 Benver (J 10979 0.04780 0.04547 0.02754 -() () 12(/l) O.OS395 --O.0391<'i 0.02483 Rno,c\'c!t (1.16793 G.12829 0.12683 G,1704G 002035 0.12572 0.15051 0.08142 0.12443_._._-_.__._._----_..._--_._--- ------------_.__._-----------
in mo,t population, and no relationship between haplotype phylogeny and geography (Fig. 2, Tahle 2). Also, 1001'43 hapl~itypes of'the Lesser Prairie-eh ieken exactly matched haplotypes ofrhc Gre3t~r Prairie-Chicken (T cupido) deposited in GenBank. while one haplmype matched a sequence identltled in both Ihe Sharp-tailed Grousc (T phasianel/us) and Greater Pramc-Chicken. One explanation for the lack of phylogeographic strnclun: is a populatioJl expansion following the Plei~rneel1e glaciatilw. Range expanSlOn after the Pleistocene glaciauon 11<1, been reported for several other dvian specles including tbe Greal Spollcd Woodpecker (DendT'ocvpo.l' l/I(1jor;
link et a!. 20(1), Marbled Murrckt (Brachvranzphus nlOrrrt()f(J
Ius; Congdon el al. 2000), King Eider lSomaleria ,\peullbilis;
Pearce ct al. 2004)_ and Mountain Plover (Charadrius /nonremus; Oyler-McCance et al. 200Se). Our re~ults art.: consistent with these other species, ilS our haplorY['Je phylogeny was shallow Md our mismatch di~tribution was unllnodill.
It has b"en argued that the Greflt Plains were sll~ieeted to massive ecologK;al pcrtLlrbation in the Pleistoccne, resulcing in many cxtlncilOns (Meng.~1 1970). A cornpanson of'species ('omp0sitions across North AJIl.:rica has suggcst.:d that the Great Plains' ilvifauna is 1m,\, in dl\·erslTY and relatively undit~
rcremialcd morphulogically. Tympanl/ellIIl may have divcrgcd around lhe Pleistocene Epoch and may have evolved rapidly into the three spccies rccognized today as glaciation constricted or shifred rhe grassland (Luechini et al. 200 I, Drovetski 2003, Spaltlding ct a1. 2006, Johnson 2(08). As glaciers receded, it appears lhal Le~~er Prairie-Chicken populalions expanded in eonecrl with the cxpilnding grasslands with the New Mexico ['Jo['Julation perhaps isolaled from the remalndc:r ofllle species.
Rangcwick mtD~A-scquellce dma suggest that gene flow 81ll(lrlg Lcssn Pr:llri~-Chicken popuiations persisls dcspit.:: popuilltion dcellT1es and habitat fragmcntlltion. Haplotype diversity in mo,,[of our populalions (h ;;. 0.8) 'A·as higher chan tn (;realer Praine-Chicken ['Jopulations ~h < 0.7) lhal have
experic:ne<>.d signdiellnt pO['Julalion horr!cne"ks (Bcl11l1ger el al. 2003, JohnsOll d al. 20()4). Our 3nalysl" rcvclIkd signilicant differences between New Mexico and most of the rest or the runge lind significant igolation by distance. As found in the Greater Prairie-Chicken by Johnson et al. (2003), however, this isolation by distance may bc reLnnanl of past sIgnals. Sim i/arly, given the fragmentation and isnlation of each popUlation we smnplcd (Fig. 1), the Illek of diHcrcntiatlOo among population~ we tou nd Inlly Ill)l retleet c()nt<~mp(lrary
gelle 1101,1,'. It is possible that lllnC 1'01' dnn to InJlllcnec ovol'all population mlDNA S1ru<;ture ha~ been In~\ll'nej,~nl, and ~Ignals ofp3M Isolalion by distance still persist. Additional analyses using nuclear markers more sensilive [0 reeen1 changes in population structure may be necessary for these competing hypothese~ co be addressed.
The pO['Julation in New Mexico was significantly different from all olhers.laeking of gene flow between Oklahoma, Kansas and Colorado (average ctJ ST "O.OXO ± O.OO?). rvloreo\,cr, in New Mexico haplotype dlversity was lower than In all other populations ,ampled, and Ihree ol'nine haplotY['Jes found there were unique, further supporting the idea that this population is Isoialed With the ['JOlential risk ofInbreeding (B0uzal and Johnson 20(4). Lesser Prall'ie-Chickens from <-iovc and t\css counties at the northern fringe 0f range bad the greatest haplOlype divcrsity (0.947 and 0.978, respeeri\-cly) and among the highest nucleotide diversity (0.011 and 0.Ot3, respecCively). HybridIzation between the Lesser and Greater Prairie-Chickens jn northtOrn Kansas has been documented by Bain and Farley (~002).
and we captured but did nor analyze llIo1cculariy five bm.b thal we considered 10 be hyhrids on the baSIS 01' morphology. AdditlOnillly, two hil['Jlotypes unique to the nonhern fnngc of'the range match Grealer PraIrie-Chicken hapIOlyp,;, in
GcnBank. [neom['Jlete lineage sorting ,n T\,nI!Janl.lchll.1 CGldd be due to recent range expansion ralher than {(l hybndll._ation (S['Jaulding ct 1I1. 2006, Johnson 200[;). However, current
35 PRATRTE-CJlTCKE1\" GENETlC~
A0,6
012 ~ ... '- . O:lsc'Ved
~ --fxoe<ted0
.~ 008 p.
e "
O.0~
10 IS 20 25 ';(1
0.16 B
0.11
c o t&. 0.08 o ~
004
>->....."'-,_._-•.-.,..-1.....- ..a a 10 15 10 lS 30
024 C
" a z
- .'. ·Ob,.,vod
--Exp~r.ted
J "
10 15 20 25 30
FIGl'RE 4. 1\1ismateh dl,;tribution of rhe observed vanation In haplotypes of the mlDI'A control region of the Le~scr Pralfl':Chickcn compared 10 rhe tllct)ret iea Idistribution rcprcsenllng popubnon C1o.pallSlon for (A) the ClltlfC sample eombinc,d, (B). the entire ;<llI1pi.: <:xduolng the N~w ,T\1~xico populaaon, and (C) rhe N.:w MexICO poplJlanoll only.
hybridizalion bctwc~n the Lesser and Grcater Prairie-Chicken in areas of overlap is apparent and !TIay funher complicat~
rccnflstniCflOIi of historical patterns of dtv.:;rgencc (Spaulding eUli. 2n06 J.
Our sl udy documcnted the distribution of variation in InIDN/\. oflhe Lesser Prairic-Chlcken in Kan~as and Colorado.
0.20
J
0.15
0.10
005
0.00
A
•
• • ••
••• ,
• •• • • • •
• • •• • •• ..
• •
•
• •
•
•
•
•
-005 •• •
0 100 200 300 400 500 600
0.20
ot5
B
•
J 0.10
0.05
0.00
-0.05
•
•
• •
• •
•
••
•• • •
••
•
••••
0 50 100 150 200 250
Euclidean Distance (km)
FIGU RE 5. PHirwist' analysis of i,olatioll by distancc lor mtD!'<A «p~7) and gCL'graphlc dlslance. (1\) All p\1pulalwn, 'iiJllIpkd: (HI. :md aTi popnlallom sampled cxcept. those 111 Ncw Mexico.
Our delta. combined with those of Van d,~n Busschc et al. (2003}, provide a rangewidc view ofLesser Prairie-Chicken population g.enetics, which can be uscd in a compn:hensivc management plan for the gpecies. Our study was consistent wirh prcvious work indicating a range expansion atkr the last glaciatlOll. In general. we found kvds of haplotype diverl>ily and gene fi,)\v that do not suggest barriers to dispersal resulting from habitat fragmenlarion and population declint'. Further work is nceded to discern whether thc~c patterns arc genetic SIgnals ofrhe past or patt~rns ofcomemporary genetic structuring. The ~ew ).t[e:\ico popnlalion, however. is isolared and it;, gencric diverSity I' lower than at uf all other populations. Consen'allol1 aClitlnS thal create or conscrve dispersal corridor; as \>,-cll as translocation, ,~hould be considered. Their em~cti\"en~ss for increa~i1ig (hc likclihood of genetic Yi8bility or tbe New MCXlCO popu latton5 should be evaluated.
300

]11 CTfRfSTL'\N A. flAGEN I:T A.l..
ACKNOWLEDGMENTS
;\"1. H. [laIn and T. Ficids provided valuable samples from Gove, '\ie~s. and Tre~o counlles \Ve thank J 0 Catlle Co .. ~unftowcr
Eleeinc Corp..-Bruokover C<lttl:: Co., P E. Beach, and R. Einsel for acccs, to !heH prnpeny C G. (;ri ffi n, (j. (' Sa Iter. R. Rodgas. T G. ShiUlC. C. SvvHnk. <Inti T. J. Whyte a~slSted wnh trapping of pram~
chicken; In Kama;,. \V. Bryant. K. M. Git'scn. and .I YOSl asslstcd with rhe collecrion l)f.,ampies 111 ColoraJo. R Van den Bnsschc pro· vl(kd dcfini!lon, of tnl D:"JA haplorype" enabling <.iJrecl comparison with our studies. hn;mcial and logistical SUPPL)r1 wa.< providcd hy the Kansa~ Department of\Vildlife and Parks (Federal A!d in WildHe restoration projcct W-53-R). Kansas Agncnltnral Experimenr Station (C'ontribu!lL)n No. 04-230-.1), Division of Biology at Kansas Siate University. and the Rocky !V!oulltaln Cenrer for (\'mervallon (Jcl1eties and Sy.,remaLics at the Unlversiry of Del1ver. Jeff John~on.
Sill1lilmha Vh~ely, and six anonynlOU~ re"lcwa, prOVided useful ('omm'::111& on earllCT dra fts ofth~ manuscripl.
LTTERATURE CITED
flAIN. MR., "NLJ (, IL I',\RLEY. 2002. Di&play of an apparent liybnJ prairie·chickens in 3 70ne of geographic overlap. COndl)r 104:683 687.
BELU\,Gl:R..\1. R., J. A. JOH"~ON, J. TOE:.PFER, AND P. DUNN. 2003. Ll)S, of gencrie variat ion in Greatcr Pralrie·Chickcn;; tollowing a population bottleneck In WisconSin, U.S.A. COllservaflon Biolo~v !7:717 724.
Ilf1i·i.~f, J. L, f-l. H CHfNG. 11. i\. LEWIN, J. D. Bn"w1'-, R. L. WESfI \1[1[:1<, AND K N. Pl\lGL. 1097. Genetic eval\laLion of a demographiC horrle[1eck In Greater I'ralfle-ChKken. ConscrvatIL)[l l3iology J2:336-843.
BOl;l·\[. J L., A\,D K. j{)I:N,O\,. 2004. Gcnetic Structure among c)o,c1y 3p'lCCd leks 111 a pcripheral population of Les,;er Pralnc· Chickcl1~. ML'leclitar Ecolugy 13:499··505
CU '11.N I. 1...1 D. PO,"D.", f,ND K. CRANI).\I I . 2000. TCS a eOrllputcr prO~fam 10 estimate gene genealoi!k". 1\·h)kcnlar E(;Olo:;]' 9:1657-·1660.
(;ON(;I)ON, B. C'., J. F. PIATT. 1<. M.\RTIN, :\'.r; V. L. Fnll·SFN. 2000. Mechanisms of pOpU!atIO(1 differenLiation in Marbled Murrelels: hislork~1 versus contcmp,wary proeess~;,. f:-,'...olution 54.974 \186.
DWI" D. M.• R. E. 1lORTON. E. A. OOELl, R. O. RODI,ERS, A'iD H. A.
Wili [LAII,. ~008. Lessc:r I'raine·Chieken conservatIOn initiative. Lcsscr Prairie Chicken Inlerstate Working Group. Colorado Oiv isiol1 of Wildlife. ForL Collin~, CO.
Dl\OV!·T<;Kl. S 2003. Plto-Plci&rocenc cltmatlc osc1I1atl,ms. hl)larctic bt(\geogr~phy and SpCCl<itTOn In an avIan ~ubfarni'y. J()urnal of IJlo;!L:ogrJphy 30. i l73.. 118 J.
b I s\\wnl. D L.. R L. []oNfvn" f. 1\. J. S\I V'Y, K D. RITTENH()I'SC. ~ND V,.!L S'·IITH. 1994. Mitochondnal-D!'<A and nude· ar-genc differenriation in Nonh '\mcncan prairie grouse. Auk 111:661 671.
GIl$I;N. K M. 2000. Popu[ati(lll statlis and management l'f Lesser I'rairie-Cbtckelllll Colorado. Praine NaturahsL 32:J37 J48.
['1 ... (;£;1', C A.. AND K. M. GIE<;EN (CJNt.lNEl 2005. Lesser PralrieChickcn (Tl'mpwlu,l1Us pallidi(.in,·!usj, no. 364. In A Poole (ED.]. The lmd_ of North America Cornell Lab ofOrnilhology, Ithaca. NY <. '" , ':>.
H·\IJKOS. D. ;\., L. (1.'1. S~lI1ll, .\ND G S. BRODA. 1990. Spnng trapplllg at' Les,er PrOlrie-Chlcken,; Journal ofField Ornithology 61 :20-·25.
J!)i1N~G."RD, P. A. 2002. Gras~land gro\i:;e llnd their con"enallon. Smlthsontal1ln:;\lllll;o:l Pr.;:s" \\'ashlngton. DC.
.lOiiNSO" ,I l,. 2f)O~. Rcc¢nr range expan;slon and chvcrgl2.n.;e alnong: t';onh ".merical) prairie grol1<;c. Journal o[ Hcredlty 99.] 65 173.
]OH"SON. J. t\ .. M. R. BCLLJNGf R, J. E. TOI·I"r::R. ,\ND P. O. DUN" 2004 Temporal chan ges in alido frequencies and low cftcctlve [l0pll!at 1011
size 111 (rreater Praine-Chickcm. Molecular Ecology 13:2617-26.10. JOIINSO'. J. A., J. E. Tnu'n,lt. ·,NO P. O. DUN'i. 2003. C'onlra>llllg
[latterns ofrllltochondnal and micrusald)n~ [l('pula:lon ..;tructure in fragmented populations ofGreatcr Prairie-Chickens. ;v10Ieeu· iarEeok1gy 12: 3335 3347
KAI!", N. W, C. E. BR.",U', J. R. Y<JLNI,. S. WOOD. D. R. :Vf.I·'A, '\:"0 T. \Ii QUIN"i 19\19. \-1olcenlar analysis ot g.enctle ."a[J:Hlor~
among large- <\I\d small-bodIed Sage Grousc llsing miiochondnal control·regiol1 seq\l~nce~. Auk J16:819-824.
KI~WR.\. M. 1980. t\ simple mcthod tor est IlllalJl1g evolutionary rare of base substirution through comparative studies of nucleotide st:qucnce~. Journal of!vlolecuJar Evolution 16:111 120.
LVX'HINI, \I.. J. HOGLU>.iO, S. KLAl,S . .1. SWE\iS(JN, ~ND E R,INDI. 2001. Historical biogeography and mirochondndl DNA phylog· eny ofgrousc and pLarJmgJIl. MO!eeuiar Phylogenetic, snd [v,)·
!lllion 20:149·· 162. f>v\.\, ,'U . N. I\167. The dcrc~tll)J) oflhsea,c c!u;rcring. and a genera!.
i7cd regression approach. Can~er Rescarch 27:209 220. MENGLL. R. M. )<)70. The Nllrtll Amt:ric3n central pla!n, as im I~O
lating agent in bird spcciatton. p. 230 .140.1/1 W. Dort allll.l K. Jone3 [EDS.]. Pleistocene and Recellt environments oflhe ccnlral Orear PJains. University Press of Kansas, Lawrencc. KS.
NWB:\L, M. A., R. L. ! IONfYCl;TT. S. G. EVAN';, R. M. WHITlN(; JR . .-\Ni)
D. R. DIl:.rz. 1997. NorLhern Bobwhite reslOckln~ ill easl Texas' ,1 generic assessment. Journai ofWtldlife Management: 61 :854 363.
OYLER·McC.~N(,E, S. .I.. S. E. TA., lOR, A"O 1'. W. Qut:'". :W05a. A multiloeus population genetic survey o[the Greakr Sagc-Gn'use aero" Their range. MoJecular Ecology j·1.1293 1310.
OYL[R-McC.... NC[, S. J., J. Sr. JO!lN. S. E. TAYLOR, ... NIJ T. VI... Qll~N.
~005b. PL'pulari')ll genetics ofGunl\ls()!1 Sagc-Grouse'lll1pllcmKms r,)r management. Journal of'Wildllfe t\·fal13gcment 69:630 -6.17.
OYLu<.-McC.'l.NCc. S. J, J. Sr. JOliN, F. L. K:-.oPI·. AND 1. W. Qu.'r-, 2005c. Popu)arion generic analysis of \10umnin Plovcr using mlloehondnal DNA sequence Jala. Condor iO"35J 362
i'Hl<Ct. J M., S L. T.·'i /lOi', B J. PII·R<',)"-. !\:1. R P,"I! "Sf N. K T SCRIBNeR, D L. OIO.SOl'<, AND A. MOSHI·.CH. 2004. Lack L)!' spa· ria] genetic slructllfc among lleslin~ and \Vlnlermg King Eiders. Condor 106:229 240,
QUINN. T. W. 1992. The g~netic legacy of Mother Go()se--phylog~.
netic patlcms of Lesser Snow (,oose Ch.,11 cum/ncens carlll,,:,crnsmarcrnililint:age" Molecular Ecology 1: IOS·-117.
ROGERS, .'\. J995. Genetic evidence for a Plel~IOCe!1e pOpUi,J110n explosion. Evolu11cn 49'008 615.
ROL.\S, J ..~ '<D R. ROZAS r999. Dn,ISP ver,lon J. an Illleg.rated riO' gram for molccular population gCl1etic~ and moJenJiar evolullon allal;·sis. Bioinfomatics IS: 174 175.
SCH"'[lD~R. S . D. R01:SSLl. AND L. Ex('O[ IIER. 2000. Arlequin \-er· sion 2.0: a ,,)ftware [or poplllarLOn genctlc' data analySIS. BlOm· erry LaboratOry, University ofGeneva, Geneva. Switzcr1<md.
S':GLLI3,\UH.R. G., J. H<xa.u"j), .~1'{) L STOR(:fl. 2003. From connectivity ro isolation: genetic consequencc., of population fragtn~II'
rallOn In Capercaillie <lcros~ Europe. Mole~ular Ecology 12: 1773 1780.
SIL\'>, N. 1., \oL J. PETERSON, "'ND R. R. LOPEZ. 200<1 Thcem:scofrhe decline orPinnared Grouse: thc Texas example. \),'t1dhfe Soeiery Bullctin32:IG 21.
SMOl.Sc, P. E., J. C. L01'<G. ANI) R. R. SOKAL. 1986. Mulrlpk regresSion and correia lion eXlensions of the Manrd leSI o[ matrix cor· respondence SystemarlC Z!l\)!ogy 35 627 632.
SPAlJLDING. A W.. K. E. MOCK. M. A. Sc HR.Ol:.l!lR.. ·\NI> K. I. w.',f(,. HEfT. 2006. Rccency, rangc cxpanSlOn, and uns,wtcd I!nc[;ge~'
IJ\lpllcatioH;; 1,11' interpreting nelltretl gel1etlc van,HHlIl III lhe:

;;, h~ rp-I<1 ilcd Grouse (Tynrp"nudlus phll.\I:aileUm). .\10 leeul ar Ecology 15:2317 2332.
Sllli;'R,.J. ft, A. D. AI'A. 1'. E Rl'.1JN0WN. ·\1';U R. M. GillSON. 2008. Polygyny anti female breeding failure reduce effecllve population size in the lekking Gunnison Sage-Gronse. Biological Con· servation 14l:472·-481.
STCjitCH, l. 20()7. Conservarion sraws of grouse worldwide: an update. Wddhl-" Biokgv 13 Supplemenl 1:5·-12.
S\IOrFoRD. D.L. 201l3. PAUP~. Phylogenetic analysis using parsiTllony ,'and other Inethods), Verslo1l4, Si nauer Associate;;, Sun· d.:rland, MA
Tnl!'! ETai'<, 1\. R., K ..>\. CR.\Nn·\u. ·INO C. F. SINe,. 1992.·'\ c1adisIi, "n.llv~I' "j phenotypic as,oeliHions With haplotypes interred from rcslr,CI ion cndonucica,e mapping and DNA scquence data. Ii I. CI,Hh'griHn C\tITJMilvn. GenetiCS 132 61'),,63J.
PRA m TE-CIlJeKE'\' GENETICS r
L'.S. FISI! .'INO WILDI..1I1 SU''';Cb. 2008. End.lnger~d and thr~a!cned
wildlife and plants; review of species that are calldrc!alC< Ih;l1 arc proposed f()r listing as endangered or lhn:31cned: the Le%er "rame·Chickcn. Federal Register 73: 75719 75180.
VAN DEN BUSSCHE. R. A.. S. R. ([OOFER, D. A. WIEDENF~.ll), D. H. \VOL i'E. .\1\l) S. K. SIII,RROIJ. 200l Genetic variatJOl1 witb1l1 and among fragmcmed populalioll5 for Lesscr Pralfle-ChiekellS (T}'mpanllchlls pallidicirletus) Moleeuh\r Ecology J2:675-6i1~.
WI,sn'.MI'IER. R. L, J. D. BIL~\\N, S. A S:\'IJ',ON, T. L. bKl:.R. R. W
JANSEN, J. W. \V.~lK, E L KERSHNE'l,. J 1... BOl!z·\T. ~"U K. N PIAGL 1<)98 Tracking the long-term declIne and recuverv 0l'3n
isolat"d popUlatIOn. Seic!1c·~ 282:1695 1698. liN .... R. \1., S. ROHWER, S. DROVLTSKI. R C BL:\CKWF.U.-RACC).
\l'il S. L. f:\HRLLi .. 2001. Holarctic phylogcography and ;,peclc, 1if)] its ofThrec-(ocd \Voodpeckers. Condor 104.167- 170