reef degradation and the loss of critical ecosystem goods and services provided by coral reef fishes
TRANSCRIPT

Reef degradation and the loss of critical ecosystem goods andservices provided by coral reef fishesMorgan S Pratchett1, Andrew S Hoey1 and Shaun K Wilson2,3
Available online at www.sciencedirect.com
ScienceDirect
Coral loss and declines in the physical structure of reef habitats
have marked effects on the abundance of coral reef fishes
which will be become even more pronounced as coral cover
continues to decline. Further, changes in the abundance and
composition of fish assemblages may have consequences for
ecosystem function and fisheries productivity. Even if fisheries
species are generally resilient to habitat degradation,
harvesting of larger fishes may threaten ecosystem functions
that are critical to coral reef resilience. Ultimately, managers will
need to explicitly address conflicts between increasing
demand for fish and the need to maintain ecosystem services.
Addresses1 ARC Centre of Excellence for Coral Reef Studies, James Cook
University, Townsville, QLD 4811, Australia2 Marine Science Program, Department of Parks and Wildlife,
Kensington, WA 6151, Australia3 Oceans Institute, University of Western Australia, Crawley, WA 6009,
Australia
Corresponding author: Pratchett, Morgan S
Current Opinion in Environmental Sustainability 2014, 7:37–43
This review comes from a themed issue on Environmental change
issues
Edited by Georgios Tsounis and Bernhard Riegl
For a complete overview see the Issue and the Editorial
Received 29 July 2013; Accepted 28 November 2013
Available online 21st December 2013
1877-3435/$ – see front matter, # Published by Elsevier B.V.
http://dx.doi.org/10.1016/j.cosust.2013.11.022
IntroductionHabitat degradation and habitat loss are the principal
causes of declining biodiversity [1], thereby jeopardizing
critical ecosystem goods and services. Habitat degra-
dation is largely manifest as loss of key habitat-forming
organisms (e.g. trees, kelp, and corals), which may be
replaced by smaller or less complex organisms, leading to
reduced habitat heterogeneity and structural (or topo-
graphic) complexity. Declines in habitat complexity
reduce the number and diversity of organisms that can
coexist, while fundamental shifts in habitat structure
often leads to localised extinctions of highly specialised
species. On coral reefs, scleractinian corals are major
contributors to both biological and physical habitat struc-
ture, and there is already evidence that sustained coral
loss in many geographical regions [2�,3] is leading to
www.sciencedirect.com
loss of biodiversity and erosion of important ecosystem
services [4,5�].
Coral reefs are among the most threatened ecosystems,
owing to a long-history of anthropogenic degradation and
exploitation [6], as well as disproportionate susceptibility
to emerging effects of global climate change [4,7]. Aside
from sustained coral loss, shifts in the biotic composition
of benthic habitats and declines in topographic complex-
ity are contributing to degradation of reef habitats
[2�,8�,9]. These changes are often accompanied by
declines in abundance of many reef-associated organisms,
especially coral reef fishes [10,11,12��] and non-coral
invertebrates [13].
The structural complexity of reef habitats is largely a
function of the abundance or cover of scleractinian corals
[14�]. However, coral cover and structural complexity may
vary independently when either the structural integrity of
erect coral skeletons is retained following extensive coral
mortality caused by bleaching or coral predators [10],
coral assemblages become dominated by growth forms
with low structural complexity [2�], or underlying com-
plexity is provided by geomorphic features, rather than
contemporary coral growth [15]. It generally appears that
loss of live coral independent of changes in habitat
structure have disproportionate effects on highly special-
ized fishes that live, feed, or settle on live corals [10,11],
whereas declines in topographic complexity may have
much broader impacts across a wide range of coral reef
fishes, including many of the larger reef fishes that sustain
tropical fisheries [16,50�].
Sustained and ongoing habitat degradation in coral reef
ecosystems will undoubtedly lead to the declines in
abundance of some reef fishes, but questions remain
about which species will be most affected and how the
structure and function of fish assemblages may change
with shifts in the biological and physical structure of
reef habitats. Of greatest importance is whether
ongoing coal loss and habitat degradation will lead to
the extinction of some species [17], undermine fisheries
productivity [5�], and/or erode key ecosystem functions
[18�,19�]. Using extensive data on the declines in
abundance of different coral reef fishes following acute
episodes of coral loss [12��], this study explores the
relative importance of first, body size, second, func-
tional roles of fishes, and third, ecological specialisation,
in explaining differential effects of reef degradation on
coral reef fishes.
Current Opinion in Environmental Sustainability 2014, 7:37–43

38 Environmental change issues
Figure 1
0
20
40
60
80
100
0 20 40 60 80 100
Div
ersi
ty a
nd a
bund
ance
of
fishe
s
Coral cover or structural complexity
(a)
0
20
40
60
80
100
0 20 40 60 80 100
Div
ersi
ty a
nd a
bund
ance
of
fishe
s
Coral cover or structural complexity
(b)
Current Opinion in Environmental Sustainability
Alternative positive relationships between fish abundance and diversity
against coral cover and structural complexity. The widely assumed linear
relationship (a) suggests that habitat degradation will have similar effects
(in terms of proportional loss in abundance and diversity of fishes) no
matter what the absolute level of coral cover or complexity. The
threshold relationship (b) meanwhile would mean that the additional
effects of habitat degradation are most pronounced when coral cover
and complexity are already low.
Linear or thresholds effectsThe first step in establishing responses of fishes to coral
loss and habitat degradation is to establish the nature of
these relationships (Figure 1). The abundance of fishes
tends to increase linearly with increases in topographic
complexity measured, at least over the limited range of
values reported for fine-scale rugosity [14�]. It is also
assumed that abundance of fishes will increase in direct
proportion to live coral cover (Figure 1a). However, some
studies have failed to detect a significant relationship
between live coral cover and abundance of fishes, even
for species (e.g. coral-feeding butterflyfishes) with strong
reliance on corals [20]. These fishes may be insensitive to
changes in coral cover over the current range sampled, but
may decline in abundance following even more pro-
nounced coral loss.
Holbrook et al. [21] revealed non-linear relationships with
live coral cover for both abundance and diversity of reef
fishes (Figure 1b). These threshold responses suggest
that reef fishes will be generally resilient to changes in
live coral cover, so long as coral cover remains above 10–20%. Very large changes in coral cover (>20%) can,
however, have measurable effects on abundance of fishes,
and are particularly noticeable if coral loss is combined
with declines in topographic complexity [10,22]. More-
over, changes in total coral cover may belie changes in
coral composition, which can have equally important
effects for reef fishes that have strong reliance on particu-
lar types of corals [23].
The extent to which live coral cover regulates the
abundance of different fish will ultimately depend upon
the basis for their reliance on corals (e.g. food versus
settlement), their specificity in coral use, and the relative
importance of other regulatory processes (e.g. limited
larval supply). Reliance on live corals by coral reefs fishes
is often greatest at settlement, where up to 65% of
species settle amongst live coral [24]. This includes
some fishes that have limited dependence on corals as
adults [25,26] and some important fisheries target
species [27–29,30�]. Preferred corals for newly settled
fishes, as well as for many obligate coral-feeding and
coral-dwelling adult fishes, are erect branching species,
and predominantly Acropora [29,30�]. Further declines
in the abundance of corals (especially Acropora) will,
therefore, greatly constrain abundance of many different
reef fishes, especially if coral cover continues to decline
below 10–20% [3].
Selective effects of coral loss on reef fishesPrimary functional groups
Declines in the abundance of fishes that perform key
ecological functions (e.g. herbivores) are important in that
they may limit the capacity of reefs to recover following
coral loss [18�,31,32��]. If these fishes are negatively
affected by degradation of reef habitats, then this will
Current Opinion in Environmental Sustainability 2014, 7:37–43
initiate a positive feedback that drives sustained and
accelerating degradation of reef habitats [18�]. Mainten-
ance of key ecological functions are often assumed based
on the high diversity of fishes within most primary func-
tional groups [33]. However, ongoing research is continu-
ing to unveil subtle differences in the ecological functions
of species placed within broad functional groups [18�,31].
Moreover, functional redundancy and the resilience of
coral reef ecosystems are critically dependant upon
response diversity among species with similar ecological
functions. Response diversity is rarely tested, but likely to
be low given that ecological function is often predicated
on strong reliance on specific resources [11].
Responses of fishes to coral loss and habitat degradation
do vary with trophic roles or functional groups [12��]. The
median response for all primary functional groups is
negative, though herbivores, carnivores and planktivores
www.sciencedirect.com

Effects of reef degradation on coral reef fishes Pratchett, Hoey and Wilson 39
Figure 2
0
+ve
-ve
Corallivores Carnivores Herbivores Planktivores
Res
pons
e to
cor
al lo
ss
Current Opinion in Environmental Sustainability
Response diversity apparent within primary functional groups of fishes, based on inter-specific differences in the responses of fishes to acute
disturbances that caused >10% declines in the cover or abundance of scleractinian corals. Data were compiled from multiple sources, as described in
Pratchett et al. [12��].
have higher response diversity compared to corallivores
(Figure 2), and some species within each of these groups
appear to benefit from coral loss. Importantly, marked
increases in abundance of some herbivorous fishes,
particularly grazing species [34�,35�], have been recorded
following extensive coral loss at some locations and are
probably critical to the prevention of phase shifts towards
macroalgal dominance [36]. However, response diversity
is substantially reduced when considering highly resolved
functional groups [12��]. The continued loss of live coral
is clearly going to lead to dramatic declines in the abun-
dance and diversity of corallivorous fishes and the func-
tions they perform. It remains to be seen, however, if
increases in a limited suite of species can offset the
extensive losses in other functional groups, thereby main-
taining key ecological processes as well as sustaining reef
fisheries. What is clear is that ongoing degradation of reef
habitats will further reduce functional redundancy [37]
making these systems even more vulnerable to future
disturbances.
Body size
Variation in body size and corresponding changes in the
scale of home ranges or feeding territories are key factors
responsible for response diversity among species with
similar ecological functions [38]. For coral reef fishes,
body size varies greatly [39] and is inextricably linked
to patterns of habitat use and the scale at which the
structure is provided. Importantly, live corals provide
relatively fine-scale complexity on coral reefs, benefiting
mostly small bodied and/or juvenile fishes [30�]. How-
ever, larger fishes, which play a disproportionate role in
www.sciencedirect.com
key ecosystem processes [31,40] and contribute most to
fisheries catches, are expected to be less reliant on this
fine-scale complexity and relatively insensitive to coral
loss [15].
A prior meta-analysis that explored the extent to which
climate-induced coral bleaching compounds upon direct
threats to reef fishes from fisheries exploitation [41]
suggested that the large fishes targeted by reef fisheries
are generally immune from effects of coral loss and
habitat degradation. Alternative analyses suggest that
ongoing degradation of coral reefs and other coastal
habitats will certainly contribute to declines in pro-
ductivity (up to 20% by 2050) of tropical coastal fisheries
[5�,42], though effects of habitat degradation will be
compounded by several other more direct effects of
climate change on fishery species [43�]. Re-analysis of
the specific responses of 403 different coral reef species
[12��], comparing changes in abundance during acute
disturbances to species-specific records of maximum
length, revealed that mean responses were similar across
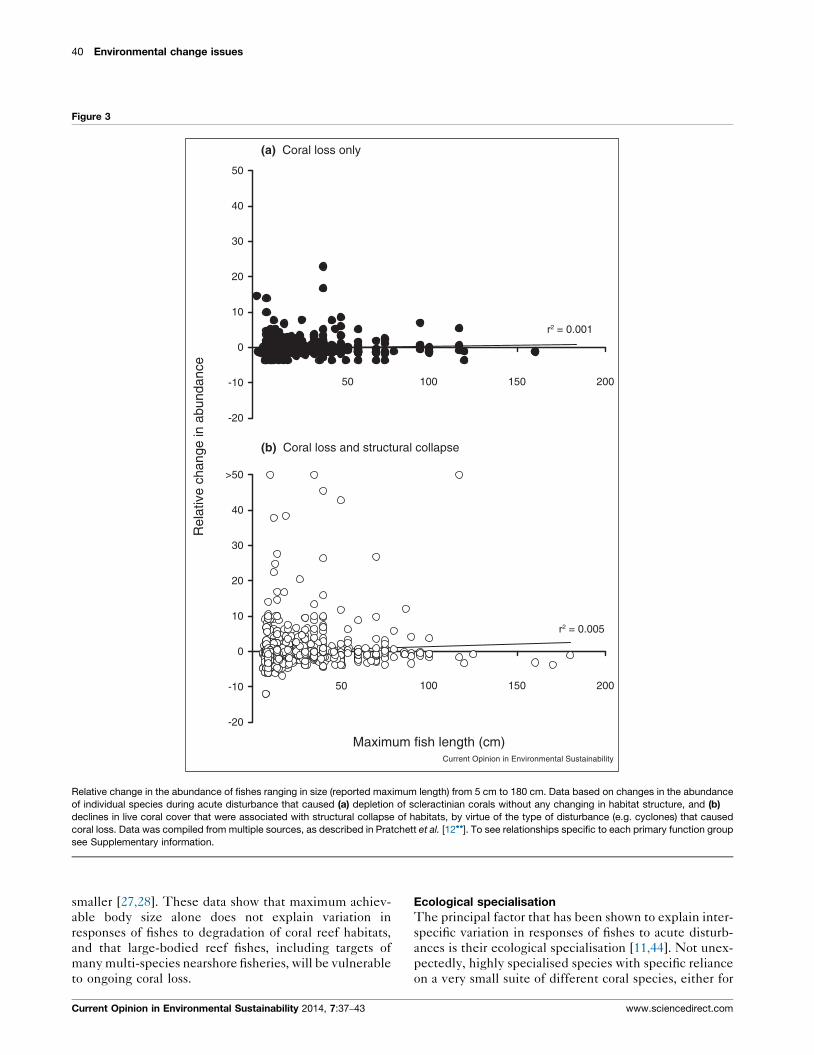
the full range of body sizes (Figure 3). Most of the fishes
that exhibited substantial declines in abundance were
small (20–50 cm in length). However, there were also
many small species that increased in abundance following
coral loss (with or without additional structural loss). For
larger fishes (>50 cm length), most species also declined
in abundance following coral loss, though some large
herbivorous and carnivorous species did exhibit increases
in abundance. The decline of larger bodied fish relates
partially to a higher dependence on live coral and associ-
ated structure during the early life history, when fish are
Current Opinion in Environmental Sustainability 2014, 7:37–43
40 Environmental change issues
Figure 3
100 150 200
-20
-10
0
10
20
30
40
50
0
100 150 200
-20
-10
0
10
20
30
40
>50
50
50
Rel
ativ
e ch
ange
in a
bund
ance
Maximum fish length (cm)
(a) Coral loss only
(b) Coral loss and structural collapse
r2 = 0.001
r2 = 0.005
Current Opinion in Environmental Sustainability
Relative change in the abundance of fishes ranging in size (reported maximum length) from 5 cm to 180 cm. Data based on changes in the abundance
of individual species during acute disturbance that caused (a) depletion of scleractinian corals without any changing in habitat structure, and (b)
declines in live coral cover that were associated with structural collapse of habitats, by virtue of the type of disturbance (e.g. cyclones) that caused
coral loss. Data was compiled from multiple sources, as described in Pratchett et al. [12��]. To see relationships specific to each primary function group
see Supplementary information.
smaller [27,28]. These data show that maximum achiev-
able body size alone does not explain variation in
responses of fishes to degradation of coral reef habitats,
and that large-bodied reef fishes, including targets of
many multi-species nearshore fisheries, will be vulnerable
to ongoing coral loss.
Current Opinion in Environmental Sustainability 2014, 7:37–43
Ecological specialisation
The principal factor that has been shown to explain inter-
specific variation in responses of fishes to acute disturb-
ances is their ecological specialisation [11,44]. Not unex-
pectedly, highly specialised species with specific reliance
on a very small suite of different coral species, either for
www.sciencedirect.com

Effects of reef degradation on coral reef fishes Pratchett, Hoey and Wilson 41
food or habitat, experience greater declines in abundance
following extensive coral loss compared to generalist
counterparts that use a broader range of different corals
[44,45]. Severe or widespread depletion of specific coral
species may, therefore, lead to the extinction of highly
specialised reef fishes [17,45]. It is apparent, however,
that highly specialised species also have a number of
complementary traits that could offset their risk of extinc-
tion [17]. For example, highly specialised species often
dominate assemblages, reflective of the competitive
advantage conferred to specialists [46]. Even among
the most vulnerable fishes (e.g. Chaetodon trifascialisand other highly specialised coral feeding butterflyfishes)
there is some optimism for long-term persistence, given
high levels of gene flow apparent among widely dispersed
populations [17] such that it would require comprehen-
sive and simultaneous depletion of preferred prey (Acro-pora spp.) across virtually the entire Indo-Pacific to cause
global extinction of this species.
ConclusionsStrong associations between reef fishes and coral rich
habitats have been reported for many decades, though
the importance of these links has become particularly
apparent during recent and severe episodes of coral loss
[10,11,15,24]. Ongoing loss of live coral and associated
changes in the biological and physical structure of coral
reef habitats will unequivocally lead to declines in abun-
dance across a wide range of different coral reef fishes,
potentially undermining critical ecosystem processes,
and/or jeopardising sustainability of coral reef fisheries
[5�]. However, just as there are likely to be shifts in the
composition of coral assemblages, reflective of differen-
tial resilience among species [47�], there will be both
winners and losers among reef fishes [47�], leading to
changes in species composition. Highly specialised
species with a direct dependence on corals for food or
habitat are likely to be increasingly replaced with general-
ist species [48�]. The maintenance of ecosystem pro-
cesses, however, will depend on the functional
attributes/roles of the specific species that persist [49�].With the exception of corallivores, there appears to be
marked inter-specific variation in responses of species
within primary functional groups, providing some evi-
dence that high diversity and functional redundancy will
ensure continuity of ecosystem functions, at least during
moderate and short-lived declines of live coral cover. It
remains to be seen whether these increases in a limited
suite of species can offset reductions in the majority of
taxa in maintaining key ecological processes. Irrespective
of the compensatory potential of these species, reduced
diversity and hence functional redundancy within each
group will increase their vulnerability to future disturb-
ances and may lead to future ecological surprises.
The effects of coral loss and habitat degradation on the
overall productivity of reef-based fisheries are equivocal.
www.sciencedirect.com
It is clear that there will be shifts in abundance and
composition of fishes with changes in habitat structure,
but in some instances these novel assemblages may
actually have similar or even higher biomass (albeit lower
diversity) of fishes that are harvested [8�]. Many of the
predominant species targeted by contemporary reef fish-
eries will decline in abundance following extensive coral
depletion [5�], either due to their inherent reliance on
coral habitats [27,28] or on prey that predominate in coral-
rich habitats [42]. Declines in abundance and diversity of
fishes will also become even more pronounced following
extensive coral loss and major shifts in the structure of
reef habitats. However, corresponding increases in the
abundance of some fishes may offset these declines,
leading to changes in catch composition, but limited
net loss of productivity. Even for fisheries species that
are generally resilient to habitat degradation, ongoing
harvesting of fishes may jeopardise ecosystem functions
leading to feedbacks that will further accelerate reef
degradation [18�]. Ultimately, managers will need to
explicitly address conflicts between increasing demand
for fish and the need to maintain ecosystem services,
which will become increasingly difficult within degraded
systems.
Appendix A. Supplementary dataSupplementary material related to this article can be
found, in the online version, at http://dx.doi.org/10.1016/
j.cosust.2013.11.022.
References and recommended readingPapers of particular interest, published within the period of review,have been highlighted as:
� of special interest�� of outstanding interest
1. Hoekstra JM, Boucher TM, Ricketts TH, Roberts C: Confronting abiome crisis: global disparities of habitat loss and protection.Ecol Lett 2005, 8:23-29.
2.�
Alvarez-Filip L, Cote IM, Gill JA, Watkinson AR, Dulvy NK: Region-wide temporal and spatial variation in Caribbean reefarchitecture: is coral cover the whole story? Global ChangeBiol 2011, 17:2470-2477.
The latest in a series of papers recognizing that effects of ongoing coralloss are even more pronounced when combined with associated declinesin the structural (architectural) complexity of reef habitats. Interestingly,coral cover and complexity are not inextricably linked, due to differencesin the relative abundance of corals with varying contributions to habitatcomplexity.
3. De’ath G, Fabricius KE, Sweatman HPA, Puotinen M: The 27-yeardecline of coral cover on the Great Barrier Reef and its causes.Proc Natl Acad Sci USA 2012, 109:17995-17999.
4. Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS,Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ,Caldeira K, Knowlton N, Eakin CM, Inglesias-Prieto R, Muthiga N,Bradbury RH, Dub A, Hartziolos ME: Coral reefs under rapidclimate change and ocean acidification. Science 2007,318:1737-1742.
5.�
Bell JB, Ganachaud A, Gehrke PC, Griffiths SP, Hobday AJ,Hoegh-Guldberg O, Jonhson JE, Le Borgne R, Lehodey P,Lough JM, Matear RJ, Pickering TD, Pratchett MS, Sen Gupta A,Senina I, Waycott M: Tropical Pacific fisheries and aquaculturehave mixed responses to climate change. Nat Clim Change2013, 3:591-599.
Current Opinion in Environmental Sustainability 2014, 7:37–43

42 Environmental change issues
This is the most comprehensive, end-to-end, vulnerability assessmentundertaken to assess how fisheries productivity across the PacificIsland Countries and Territories (PICTs) will be effected by climatechange. Conclusions vary for different fisheries, but productivity ofcoastal (including reef based) fisheries is expected to decline due toongoing habitat degradation and emerging effects of increasing tem-perature and ocean acidification on both the fishes and habitats thatsupport them.
6. Jackson JBC, Kirby MX, Berger WH, Bjorndal KA, Botsford LW,Bourque BJ, Bradbury R, Cooke R, Erlandson J, Estes JA,Hughes TP, Kidwell S, Lange CB, Lenihan HS, Pandolfi JM,Peterson CH, Steneck RS, Tegner MJ, Warner R: Historicaloverfishing and the recent collapse of coastal ecosystems.Science 2001, 293:629-638.
7. Hughes TP, Baird AH, Bellwood DR, Card M, Connolly SR, Folke C,Grosberg R, Hoegh-Guldberg O, Jackson JBC, Kleypas J,Lough JM, Marshall PA, Nystrom M, Palumbi SR, Pandolfi JM,Rosen B, Roughgarden J: Climate change, human impacts, andthe resilience of coral reefs. Science 2007, 301:929-933.
8.�
Chong-Seng K, Mannering TD, Pratchett MS, Bellwood DR,Graham NAJ: The influence of coral reef benthic conditionon associated fish assemblages. PLoS ONE 2012, 7:e42167.
The first study that unequivocally measures changes in the compositionand abundance of coral reef fishes along a gradient of reef degradation,from habitats with high coral cover and structural complexity to habitatsdominated by macroalgae and low structural complexity. While compo-sition and diversity (and therefore function redundancy) decline along thegradient, there were limited differences in the abundance of fishes withinkey functional groups.
9. McClanahan TR, Graham NAJ, Darling ES: Predicting the futureof coral reefs from the covarying sensitivities of fish and coralsto climate and fishing disturbances. Curr Opin EnvironSustainability 2014, 7:59-64.
10. Wilson SK, Graham NAJ, Pratchett MS, Jones GP, Polunin NVC:Multiple disturbances and the global degradation of coralreefs: are reef fishes at risk or resilient? Global Change Biol2006, 12:2220-2234.
11. Pratchett MS, Munday PL, Wilson SK, Graham NAJ, Cinner JE,Bellwood DR, Jones GP, Polunin NVC, McClanahan TR: Effects ofclimate-induced coral bleaching on coral-reef fishes:ecological and economic consequences. Oceanogr Mar BiolAnn Rev 2008, 46:251-296.
12.��
Pratchett MS, Hoey AS, Wilson SK, Messmer V, Graham NAJ:Changes in the biodiversity and functioning of reef fishassemblages following coral bleaching and coral loss.Diversity 2011, 3:424-452.
This study presents the first tests of response diversity among coralreef fishes, which is considered fundamental to the resilience of coralreef ecosystems. Response diversity was very low for corallivores,and moderate even among key functional groups of herbivores,omnivores and carnivores. It was concluded that high diversity willnot necessarily ensure the maintenance of key ecosystems functions,such that reefs with high diversity are no less vulnerable to majordisturbances.
13. Stella JS, Pratchett MS, Hutchings PA, Jones GP: Diversity,importance and vulnerability of coral-associatedinvertebrates. Oceanogr Mar Biol Ann Rev 2011, 49:43-104.
14.�
Graham NAJ, Nash KL: The importance of structural complexityin coral reef ecosystems. Coral Reefs 2013, 32:315-326.
This meta-analysis of quantitative assessments of structural complexityon coral reefs demonstrates that live coral is a major component of reefrugosity, although this relationship breaks down in the Caribbean wherebranching corals have been lost, emphasizing the importance of coralslike Acropora to reef structure. Analyses also found a stronger relation-ship between complexity and fish abundance in fished areas as there is ahigher occurrence of small bodied fish in fished areas due to removal ofpredators.
15. Graham NAJ, Wilson SK, Jennings S, Polunin NVC, Bijoux JP,Robinson J: Dynamic fragility of oceanic coral reefecosystems. Proc Nat Acad Sci USA 2006, 103:8425-8429.
16. Cinner JE, McClanahan TR, Graham NAJ, Pratchett MS,Wilson SK, Raina J: Gear-based fisheries management as apotential adaptive response to coral bleaching. J Appl Ecol2009, 46:724-732.
Current Opinion in Environmental Sustainability 2014, 7:37–43
17. Lawton RJ, Messmer V, Bay LK, Pratchett MS: High gene flowacross large geographic scales reduces extinction risk forhighly specialised coral feeding butterflyfishes. Mol Ecol 2011,17:3584-3598.
18.�
Hoey AS, Bellwood DR: Suppression of herbivory by macroalgaldensity: a critical feedback on coral reefs? Ecol Lett 2011,14:267-273.
While high densities of seaweed may be ‘bio-indicators’ of poor reefhealth, this study was the first to demonstrate that they are also potentialdrivers of positive feedback loops. Both grazing and browsing fishesavoided areas of high macroalgal biomass, a behaviour that may lead tothe growth and persistence of macroalgal stands on coral reefs, andincrease the stability of phase shifts to macroalgae.
19.�
Cheal AJ, Emslie M, MacNeil MA, Miller I, Sweatman HPA: Spatialvariation in the functional characteristics of herbivorous fishcommunities and the resilience of coral reefs. Ecol Appl 2013,23:174-188.
Examining the functional characteristics of herbivorous fish communitiesacross 92 reefs and 15 years on the Great Barrier Reef this study revealedthat many coastal reefs were most vulnerable to coral-macroalgal phaseshifts. The functionality of these reefs was compromised by lower abun-dance and diversity of fishes within each of four functional groups.
20. Lawton RJ, Pratchett MS: Influence of dietary specialisationand resource availability on geographical variation inabundance of butterflyfishes. Ecol Evol 2012, 2:1347-1361.
21. Holbrook SJ, Schmitt RJ, Brooks AJ: Resistance and resilienceof a coral reef fish community to changes in coral cover. MarEcol Prog Ser 2008, 371:263-271.
22. Wilson SK, Dolman AM, Cheal AJ, Emslie MJ, Pratchett MS,Sweatman HPA: Maintenance of fish diversity on disturbedcoral reefs. Coral Reefs 2009, 28:3-14.
23. Berumen ML, Pratchett MS: Recovery without resilience:persistent disturbance and long-term shifts in the structure offish and coral communities at Tiahura Reef, Moorea. CoralReefs 2006, 25:647-653.
24. Jones GP, McCormick MI, Srinivasan M, Eagle JV: Coral declinethreatens fish biodiversity in marine reserves. Proc Natl AcadSci USA 2004, 101:8251-8253.
25. Wilson SK, Depczynski M, Fisher R, Holmes TH, O’Leary RA,Tinkler P: Habitat associations of juvenile fish at Ningaloo Reef,Western Australia: the importance of coral and algae. PLoSONE 2010, 5:e15185.
26. Coker DJ, Graham NAJ, Pratchett MS: Interactive effects of livecoral and structural complexity on the recruitment of reeffishes. Coral Reefs 2012, 31:919-927.
27. Wen C, Pratchett MS, Almany G, Jones GP: Patterns ofrecruitment and microhabitat associations for three predatorycoral reef fishes on the southern Great Barrier Reef, Australia.Coral Reefs 2012, 32:389-398.
28. Nanami A, Sato T, Takebe T, Teruya K, Soyano K: Microhabitatassociation in white-streaked grouper Epinephelus ongus:importance of Acropora spp.. Mar Biol 2013, 160:1511-1517.
29. Cole AJ, Pratchett MS, Jones GP: Diversity and functionalimportance of coral-feeding fishes on tropical coral reefs. FishFish 2008, 9:286-307.
30.�
Coker DJ, Wilson SK, Pratchett MS: Importance of live coralhabitat for reef fishes. Rev Fish Biol Fish 2013 http://dx.doi.org/10.1007/s11160-013-9319-5.
This is the first comprehensive list of coral reef fishes that associate withlive coral, identifying 320 species from 39 families that utilise live coralhabitats, and may be vulnerable to extensive widespread coral loss.
31. Hoey AS, Bellwood DR: Limited functional redundancy in a highdiversity system: single species dominates key ecologicalprocess on coral reefs. Ecosystems 2009, 12:1316-1328.
32.��
Rasher DB, Hoey AS, Hay ME: Consumer diversity interactswith prey defenses to drive ecosystem function. Ecology 2013,94:1347-1358.
Investigating the susceptibility of various macroalgal types to herbivorousfish assemblages this study revealed a remarkable lack of overlap in themacroalgae that each herbivore consumed. This striking complementar-ity among herbivorous fishes was supported by strong negative relation-
www.sciencedirect.com

Effects of reef degradation on coral reef fishes Pratchett, Hoey and Wilson 43
ships between herbivore diversity and the abundance and diversity ofmacroalgae across the study reefs.
33. Folke C, Carpenter S, Walker B, Scheffer M, Elmqvist T,Gunderson L, Holling CS: Regime shifts, resilience, andbiodiversity in ecosystem management. Ann Rev Ecol Syst2004, 35:557-581.
34.�
Adam TC, Schmitt RJ, Holbrook SJ, Brooks AJ, Edmunds PJ,Carpenter RC, Bernardi G: Herbivory, connectivity, andecosystem resilience: response of a coral reef to a large-scaleperturbation. PLoS ONE 2011, 6:e23717.
This comprehensive and detailed study examined responses of herbivor-ous fishes (principally, Chlorurus sordidus and Scarus psittacus) to recentcoral loss in Moorea, French Polynesia. This study documents markedincreases in abundance of these herbivores in the aftermath of extensivecoral loss, which prevented the establishment of macroalgae. This studyreveals the critical importance of habitat structure in facilitating continuityof ecosystem function.
35.�
Gilmour JP, Smith LD, Heyward AJ, Baird AH, Pratchett MS:Recovery of an isolated coral reef system following severedisturbance. Science 2013, 340:69-71.
Isolated reefs are often assumed to be highly vulnerable to large dis-turbance events owing to strong reliance on local replenishment, but alsobenefit isolation from most chronic anthropogenic disturbances (e.g.fisheries exploitation). This study documents the resilience of a small,isolated, offshore reef, in the aftermath of significant climate-inducedbleaching. Settlement and survivorship of corals were facilitated by localabundance of herbivorous fishes throughout.
36. Hughes TP, Rodrigues MJ, Bellwood DR, Ceccerelli D, Hoegh-Guldberg O, McCook L, Moltchaniwskyj N, Pratchett MS,Steneck RS, Willis BL: Phase shifts, herbivory and theresilience of coral reefs to climate change. Curr Biol 2007,17:360-365.
37. Hoey AS, Brandl SJ, Bellwood DR: Diet and cross-shelfdistribution of rabbitfishes (f. Siganidae) on the northern GreatBarrier Reef: implications for ecosystem function. Coral Reefs2013, 32:973-984.
38. Allen CR, Gunderson L, Johnson AR: The use of discontinuitiesand functional groups to assess relative resilience in complexsystems. Ecosystems 2005, 8:958-966.
39. Munday PL, Jones GP: The ecological implications of smallbody size among coral-reef fishes. Oceanogr Mar Biol Ann Rev1998, 36:373-411.
40. Bellwood DR, Hoey AS, Hughes TP: Human activity selectivelyimpacts the ecosystem roles of parrotfishes on coral reefs.Proc R Soc B 2012, 279:1621-1629.
41. Graham NAJ, Chabanet P, Evans RD, Jennings S, Letourneur T,MacNeil MA, McClanahan TR, Ohman MC, Polunin NVC,Wilson SK: Extinction vulnerability of coral reef fishes. Ecol Lett2011, 14:341-348.
42. Wilson SK, Fisher R, Pratchett MS, Graham NAJ, Dulvy NK,Turner RA, Cakacaka A, Polunin NVC: Habitat degradation andfishing effects on the size structure of coral reef fishcommunities. Ecol Appl 2010, 20:442-451.
www.sciencedirect.com
43.�
Cheung WWL, Watson R, Pauly D: Signature of ocean warmingin global fisheries catch. Nature 2013, 497:365-369.
Using information on the temperature preferences of fish, combined withtemporal changes in fisheries catch data from around the world, thisstudy showed that global climate change and increasing ocean tempera-ture are affecting the composition, if not overall productivity, of temperatefisheries.
44. Pratchett MS, Coker DJ, Jones GP, Munday PL: Specialisation inhabitat use by coral reef damselfishes and their susceptibilityto habitat loss. Ecol Evol 2012, 2:2168-2180.
45. Munday PL: Habitat loss, resource specialization, and extinctionon coral reefs. Global Change Biol 2004, 10:1642-1647.
46. Blowes S, Pratchett MS, Connolly SR: Heterospecificaggression, dominance and specialization in a guild ofcorallivorous reef fishes. Am Nat 2013, 128:2-13.
47.�
Hughes TP, Baird AH, Dinsdale EA, Moltschaniwskyj NA,Pratchett MS, Tanner JE, Willis BL: Assembly rules of reef coralsare flexible along a climatic gradient. Curr Biol 2012, 22:736-741.
This study explores spatial variation in the abundance of different coraltaxa along a latitudinal, and therefore, temperature gradient, to infer likelychanges in the composition of coral assemblages as a consequence ofongoing ocean warming. Given marked differences in the distributionof corals it is concluded that climate change will lead to reassortment ofcoral reef taxa rather than wholesale loss of reef ecosystems.
48.�
Bellwood DR, Baird AH, Depczynski M, Gonzalez-Cabello A,Hoey AS, Lefevre C, Tanner JK: Coral recovery may not heraldthe return of fishes on damaged coral reefs. Oecologia 2012,170:567-573.
The diversity of small bodied crypto-benthic coral reef fish does notchange after coral bleaching; however, the composition of the fishcommunity becomes characterized by habitat generalists and there isa decline in coral specialists. These changes to fish community persist,even after coral recovery, implying that rapid changes in fish communitiesdue to coral loss are preserved for up to 13 years, even when specieshave short life-spans.
49.�
Cheal AJ, MacNeil MA, Cripps E, Emslie MJ, Jonker M,Schaffelke B, Sweatman H: Coral–macroalgal phase shifts orreef resilience: links with diversity and functional roles ofherbivorous fishes on the Great Barrier Reef. Coral Reefs 2010,29:1005-1015.
Comparing the trajectories of reefs following extensive coral mortality,this study highlighted the importance of the composition of herbivorousfish assemblages, with a shift to macroalgal-dominance being associatedwith low herbivore diversity, and low abundances of browsing andcropping species.
50.�
Wilson SK, Babcock RC, Fisher R, Holmes TH, Moore JAY,Thomson DP: Relative and combined effects of habitat andfishing on reef fish communities across a limited fishinggradient at Ningaloo. Mar Environ Res 2012, 81:1-11.
One of the few studies that explicitly assesses the interaction betweenhabitat condition on coral reefs and the effects of fishing, demonstratingthat the positive correlation between abundance of fish and structuralcomplexity is stronger on fished reefs than those where fishing is pro-hibited and that structural complexity is a better predictor of fish diversitythan live coral cover.
Current Opinion in Environmental Sustainability 2014, 7:37–43