rapid recruitment of neutrophils containing prestored il ... · toward an invading micro-organism...
TRANSCRIPT

of November 24, 2018.This information is current as
InfectionMicrobialContaining Prestored IL-12 During
Rapid Recruitment of Neutrophils
Susan K. Bliss, Barbara A. Butcher and Eric Y. Denkers
http://www.jimmunol.org/content/165/8/4515doi: 10.4049/jimmunol.165.8.4515
2000; 165:4515-4521; ;J Immunol
Referenceshttp://www.jimmunol.org/content/165/8/4515.full#ref-list-1
, 21 of which you can access for free at: cites 33 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2000 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Novem
ber 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on N
ovember 24, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Rapid Recruitment of Neutrophils Containing Prestored IL-12During Microbial Infection 1
Susan K. Bliss, Barbara A. Butcher, and Eric Y. Denkers2
Neutrophils are well known to rapidly migrate to foci of infection, where they exert microbicidal functions. We sought to determinewhether neutrophils responding to in vivo infection with the protozoan pathogenToxoplasma gondiiwere capable of IL-12production as suggested by recent in vitro studies. Intraperitoneal infection induced a neutrophil influx by 4 h, accompanied byex vivo IL-12 p40 and p70 release. Approximately 85% of the neutrophils displayed intracellular stores of IL-12, as determinedby flow cytometry and confocal fluorescence microscopy. Neutrophils from IFN-g knockout mice also expressed IL-12, ruling outan IFN-g-priming requirement. Neither infected nor uninfected peritoneal macrophages displayed intracellular IL-12, but thesecells were strongly IL-101. Infection per se was unnecessary for IL-12 production because peritoneal and peripheral bloodneutrophils from uninfected animals contained IL-121 populations. Expression of the granulocyte maturation marker Gr-1(Ly-6G) was correlated with IL-12 production. Mice depleted of their granulocytes by mAb administration at the time of infectionhad decreased serum levels of IL-12 p40. These results suggest a model in which neutrophils with prestored IL-12 are rapidlymobilized to an infection site where they are triggered by the parasite to release cytokine. Our findings place neutrophils prom-inently in the cascade of early events leading to IL-12-dependent immunity toT. gondii. The Journal of Immunology,2000, 165:4515–4521.
N eutrophils are critical effector cells in the host’s responseto microbial invasion (1). They play a role in recogniz-ing and neutralizing bacteria, fungi, and parasites (2–6).
To accomplish this, neutrophils must possess a coordinated andspecialized set of functions that mediate detection and chemotaxistoward an invading micro-organism and effective killing mecha-nisms. Their ability to defend the body relies in part upon rapidmobilization of preformed compounds found in granules such asthe degradative enzymes cathepsin G, elastase, and lysozyme (7,8). Recent findings from several laboratories, including our own,have revealed that neutrophils also release several immunomodu-latory cytokines in response to in vitro microbial stimulation (3, 6,9, 10). Specifically, our work has focused on the role of neutrophilsin the early immune response to the microbial pathogen,Toxo-plasma gondii.
T. gondii is an obligate, intracellular protozoan parasite thatcauses morbidity and mortality in a broad range of host species.Indeed, a recent survey indicated thatT. gondiiis the leading causeof human death among food-borne diseases in the United States(11). As an opportunistic organism, its importance as a pathogenhas resurfaced with the AIDS epidemic, and it has been estimatedthat ;30% of AIDS patients suffer from reactivation of infection(12). Moreover, the parasite poses a serious threat to the unbornfetus when transmitted placentally during maternal infection (13).
Like many intracellular pathogens,T. gondii induces a strongprotective cell-mediated immune response that is driven by earlyIL-12 production (14, 15). The source of this cytokine duringT.gondii infection has been attributed to both macrophages and,more recently, dendritic cells (16, 17). Previously, we reported thathuman and murine neutrophils elaborate high levels of IL-12 uponin vitro stimulation with aT. gondiiAg extract (6, 18), results thatare in broad agreement with the findings of others investigatingCandida albicansinfection (19). Notably, experiments in cytokinegene and cytokine receptor knockout animals demonstrated thatparasite-triggered IL-12 release occurred independently ofendog-enous IFN-g and did not require signaling through the TNF p55receptor. We and others also found that C57BL/6 mice succumbacutely to infection when depleted of their granulocytes, con-firming the importance of these cells during in vivo infection (6,20, 21).
Given that neutrophils prestore and rapidly release microbicidalmolecules when appropriately stimulated and that these cells canbe triggered to release immunomodulatory cytokines during invitro stimulation, we sought to determine whether neutrophilsserve as an IL-12 source during in vivo infection. Furthermore, weexamined whether neutrophils prestore this cytokine in the absenceof an ongoing infection. Our results demonstrate that i.p. infectionwith tachyzoites induces a rapid influx of neutrophils into the peri-toneal cavity expressing the maturation marker Gr-1 (Ly-6G), andthese cells display high levels of intracellular IL-12 protein. Re-markably, we found that peripheral blood as well as peritonealneutrophils from noninfected mice contain a population of Gr-1-expressing neutrophils that also display high levels of intracellularIL-12. Mice depleted of their granulocytes at the time of infectionwith bradyzoite cysts have decreased levels of serum IL-12. Theseresults emphasize neutrophils as an important in vivo source ofIL-12 duringT. gondii infection. Moreover, the finding that intra-cellular IL-12 acquisition is associated with the normal neutrophilmaturation program places these cells prominently in the cascadeof early events that initiate cellular immunity to infection.
Department of Microbiology and Immunology, Cornell University College of Veter-inary Medicine, Ithaca, NY 14853
Received for publication April 18, 2000. Accepted for publication July 26, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grant AI40540.2 Address correspondence and reprint requests to Dr. Eric Denkers, Department ofMicrobiology and Immunology, Cornell University College of Veterinary Medicine,Ithaca, NY 14853-6401. E-mail address: [email protected]
Copyright © 2000 by The American Association of Immunologists 0022-1767/00/$02.00
by guest on Novem
ber 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Materials and MethodsMice
C57BL/6 female mice (6–12 wk of age) were obtained from The JacksonLaboratory (Bar Harbor, ME). IFN-g knockout female mice (GKO)3 on aC57BL/6 background (6–12 wk of age) were obtained through a NationalInstitute of Allergy and Infectious Diseases contract with Taconic Farms(Germantown, NY). The animals were housed under specific pathogen-freeconditions at the College of Veterinary Medicine animal facility at CornellUniversity; the college maintains an animal facility that is accredited by theAmerican Association for Accreditation of Laboratory Animal Care.
Parasites and infections
Tachyzoites of the RH strain were maintained on human foreskin fibroblastmonolayers in DMEM (Life Technologies, Gaithersburg, MD), 1% FCS(HyClone, Logan, UT), and 100 U/ml penicillin and 0.1 mg/ml strepto-mycin (Life Technologies). Parasite cultures were free of contamination byMycoplasma spp.as determined by RT-PCR, ELISA (kits from Stratagene(La Jolla, CA) and Roche (Indianapolis, IN), respectively), microbiologicalassay, and fluorescent DNA staining (performed by the Mycoplasma Test-ing Laboratory, Coriell Institute for Medical Research, Camden, NJ). Be-fore infection, tachyzoites were washed in endotoxin-free PBS. Mice werei.p. infected with 23 106 tachyzoites or were injected with PBS.
ME49 bradyzoite cysts were maintained in Swiss-Webster mice as de-scribed previously (6). Mice were rendered neutropenic with an anti-Gr-1mAb (RB6C6.8C5 hybridoma originally provided by R. L. Coffman(DNAX Research Institute, Palo Alto, CA) and were infected with 100ME49 cysts i.p. as described previously (6).
Cell culture conditions
Peritoneal cells were obtained from mice 4 or 6 h postinfection (p.i.) bylavage with PBS. Cells were cultured in medium alone at 43 106/well for18 h at 37°C with 5% CO2. Medium consisted of DMEM with 10% FCS,1 mM sodium pyruvate (Life Technologies), 0.1 mM nonessential aminoacids (Life Technologies), 30 mM HEPES (Life Technologies), 100 U/mlpenicillin, 0.1 mg/ml streptomycin, and 53 1025 M 2-ME. Supernatantswere harvested and stored at220°C until assayed for IL-12 levels. Todeplete peritoneal cell populations of granulocytes, immunomagneticbeads (Dynal, Oslo, Norway) coupled to anti-Gr-1 mAb were used as de-scribed previously (6). To determine the composition of isolated cells,differential cell counts were performed on Diff-Quik (American ScientificProducts, McGraw Park, IL)-stained cytocentrifuge slides. A minimum of300 cells were counted per slide.
Cytokine measurement
IL-12 p40 was measured in cell-free supernatants or plasma by ELISA asdescribed in detail previously (6). To measure IL-12 p70, the protocol fordetermining p40 levels was followed, except that anti-IL-12 p70 (clone9A5, PharMingen, San Diego, CA) was used as the coating Ab at 5mg/ml.Detection sensitivities for IL-12 p40 were 10 and 50 pg/ml for IL-12 p70.
Confocal fluorescence microscopy
Cells (23 105/sample) were centrifuged onto 12-mm microscope cover-glasses (VWR Scientific, Rochester, NY) resting on glass microscopeslides (VWR Scientific) using cytofunnels (Shandon, Pittsburgh, PA) anda cytospin centrifuge (Shandon). Coverslips were then placed in wells of a24-well tissue culture plate (Corning Costar, Cambridge, MA) for subse-quent staining. Cells were fixed for 20 min at room temperature in PBScontaining 3% formaldehyde (Sigma, St. Louis, MO) with 0.1 mM CaCl2
(Sigma) and 0.1 mM MgCl2 (Sigma), then washed in permeabilizationbuffer (0.075% saponin dissolved in PBS). Blocking was subsequently per-formed (20 min at room temperature) using 5% normal mouse serum(NMS; Accurate, Westbury, NY) diluted in permeabilization buffer. Next,rat anti-mouse cytokine mAb were added (5mg/ml), and cells were incu-bated for 30 min at room temperature. Anti-IL-12 p40 (clone C17.8, pro-vided by G. Trinchieri, Wistar Institute, Philadelphia, PA) (22), anti-IL-10(clone JES5-16E3; PharMingen), and a control rat Ig (Accurate Chemical,Westbury, NY) were used to stain cells. After washing three times in per-meabilization buffer, FITC-conjugated mouse anti-ratk light chain (cloneOX-12; Serotec, Raleigh, NC) diluted 1/500 in permeabilization buffer wasadded, and cells were incubated for an additional 30 min at room temper-ature. In some experiments, an FITC-conjugated anti-Gr-1 mAb was em-
ployed (clone RB6C6.8C5; PharMingen). During the last 10 min of incu-bation, propidium iodide (Sigma) was added (15mg/ml finalconcentration). The cells were washed three times in permeabilizationbuffer, followed by four washes in PBS. Coverslips were mounted on glassmicroscope slides in ProLong Anti-Fade (Molecular Probes, Eugene, OR).The slides were examined with a Bio-Rad MRC600 confocal laser scan-ning microscope, and images were collected using Comos software (Bio-Rad, Hercules, CA).
Flow cytometric analysis
Peripheral blood was obtained by cardiac puncture, and erythrocytes werelysed by a method described previously (23). Briefly, 300ml of wholeblood was placed in 4 ml of lysis buffer (156 mM ammonium chloride, 20mM sodium bicarbonate, and 1 ml of 0.5 M EDTA at pH 8 in water;reagents from Sigma). The cells were incubated at room temperature for 5min and then spun at 1500 rpm for 5 min. The resulting leukocyte pelletwas washed once in wash buffer composed of 0.1% albumin (Sigma) and1 ml of 0.5 M EDTA at pH 8 in Ca12-, Mg12-, and phenol red-free HBSS(Life Technologies). Cells (13 106/sample) were fixed in 200ml of 3%formaldehyde with 0.1 mM CaCl2 and 0.1 mM MgCl2 at room temperaturefor 20 min. Cells were then washed twice in permeabilization buffer andblocked in 5% NMS diluted in permeabilization buffer for 15 min at roomtemperature. C17.8 mAb or control rat Ig were then added at 15mg/ml. AllAb were used at saturating concentrations. The cells were incubated atroom temperature for 20 min and then washed in permeabilization bufferthree times. Cells were resuspended in 5% NMS in permeabilization buffer,and 1mg/ml of PE-conjugated anti-ratk light chain (clone MRK-1, PharM-ingen) was added. The cells were incubated for 20 min at room tempera-ture, followed by two washes in permeabilization buffer. Cells were thenwashed once in PBS and resuspended in 5% NMS diluted in PBS witheither FITC-conjugated anti-Gr-1 or isotype control (clone A95-1; PharM-ingen) and incubated at room temperature for 20 min. Finally, cells werewashed three times in PBS and used for flow cytometric analysis. Cells(1 3 105/sample) were collected, and data were analyzed using CellQuestsoftware and a FACScalibur flow cytometer (Becton Dickinson Immuno-cytometry Systems, San Jose, CA). The lower than expected side scatter ofthe granulocyte population was a consequence of the fixation and perme-abilization protocol. It should be noted that no inhibitors of secretion, suchas monensin or brefeldin A, or cell activation factors, such as PMA orionomycin, were used in these studies.
Statistical analysis
Significant differences were determined using Student’st test in Figs. 1 and9. Values ofp # 0.05 were considered significant. All experiments wereperformed on a minimum of three independent occasions.
3 Abbreviations used in this paper: GKO, IFN-g2/2 mice; STAg, soluble tachyzoiteAg; p.i., postinfection; NMS, normal mouse serum.
FIGURE 1. Production of IL-12 byT. gondii-infected peritoneal cells.C57BL/6 mice were injected i.p. with 23 106 RH strain tachyzoites orPBS. After 4 h the peritoneal cells were obtained by lavage with PBS andincubated in medium alone at 43 106/well for 18 h. Cell-free supernatantswere collected for cytokine measurement by ELISA as described inMa-terials and Methods. The infected population was composed of 46% neu-trophils, 46.5% macrophages, 6.5% lymphocytes, and 1% eosinophils asdetermined by differential counts. The control population was composed of6% neutrophils, 83% macrophages, 10.5% lymphocytes, and 0.5% eosin-ophils. Results are expressed as the mean6 SD. ND, none detected. Dif-ferences between infected and uninfected cultures were statisticallysignificant.
4516 IL-12 EXPRESSION IN NEUTROPHILS
by guest on Novem
ber 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

ResultsIntraperitoneal tachyzoite infection induces rapid neutrophilinfiltration and release of IL-12 p40 and p70
We previously established that 18-h thioglycolate-elicited murineperitoneal neutrophils produce IL-12 p40 in response to in vitrostimulation with a soluble tachyzoite Ag (STAg) preparation (6).We also demonstrated that i.p. tachyzoite injection induced an in-flux of neutrophils that correlated with increased levels of IL-12p40 gene transcripts. However, these data did not indicate whetherIL-12 protein was released, nor did they directly demonstrate neu-trophil IL-12 production. To address these issues, mice were in-jected i.p. with 23 106 RH strain tachyzoites in PBS or PBSalone. After 4 h, peritoneal cells were obtained. Differential cellcounts revealed that infection induced a neutrophilic influx, suchthat 46% of the peritoneal cells were of this cell type (data notshown). In contrast, injection of PBS failed to elicit a similar in-flux. The unseparated cell populations were placed in culture andincubated for 18 h without further stimulation. Cells obtained frominfected animals released both IL-12 p40 and the bioactive p70form of the cytokine as determined by ELISA (Fig. 1). Approxi-mately one-third of the p40 contributed to the formation of p70. NoIL-12 was detected from uninfected cultures. Since this result rep-resents a mixed populational response, we next examined cytokineproduction at the single-cell level to definitively define the IL-12cell source during this model of early infection.
Mutually exclusive expression of IL-12 and IL-10 by neutrophilsand macrophages, respectively
Cells obtained 4 h after RH infection or PBS injection were im-mediately fixed and stained intracellularly for IL-12 or IL-10 byindirect immunofluorescence, and nuclei were counterstained withpropidium iodide. The resulting samples were examined by con-focal fluorescence microscopy. Fig. 2Ashows that the majority ofgranulocytes, here distinguished by their characteristic nuclearmorphology, possess IL-12. Although eosinophils display similarnuclear characteristics (24), Diff-Quik staining of parallel samplesrevealed that eosinophils only constituted 1% of the peritonealpopulation, ruling out the latter cells as the major IL-12 source.Notably, there is no evidence for expression of IL-12 in the re-mainder of the population, most of which is composed of macro-phages. Fig. 2Dshows an IL-121 cell within the population ofperitoneal cells from uninfected animals. Interestingly, while neu-trophils are much less common in this population, some stain forIL-12, suggesting that infection per se is not a prerequisite forneutrophil IL-12 expression.
Since neutrophils have been reported to be a source of IL-10 inC. albicansinfection (3, 19), we also examined RH-infected andPBS-injected populations for expression of this cytokine. Fig. 2Bdemonstrates that there is no evidence for expression of IL-10 inthe neutrophil population derived from infected animals. However,macrophages present in this population displayed strong stainingfor IL-10. FACS analysis indicated that IL-101 cells were alsoF4/801, confirming that these cells belong to the monocyte/mac-rophage lineage (data not shown). In no experiment did we findIL-12 expression by F4/801 cells. This is important because of arecent paper by Biermann et al. (24), who found that morpholog-ical distinction between a subset of granulocytes and monocytes/macrophages can be difficult due to similar ring-shaped nuclearmorphology. Of note, peritoneal macrophages from uninfectedmice appear to contain IL-10, although levels are clearly lowerthan those found in cells from infected mice (Fig. 2E, compare toFig. 2B). The isotype control mAb shown in Fig. 2,C and F,confirm the specificity of the cytokine mAb staining.
Fig. 3 demonstrates IL-12 staining in peritoneal cells 6 h afterinfection. Neutrophils remained strongly IL-121, while macro-phages continued to display little or no evidence for the presenceof intracellular IL-12 (Fig. 3A). This was true even for tachyzoite-infected macrophages, as shown in Fig. 3A. Additionally, macro-phages appear to be preferentially infected over neutrophils. Fig.
FIGURE 2. Reciprocal expression of IL-12 and IL-10 by peritonealneutrophils and macrophages, respectively. Cells from 4-h infected andPBS-injected mice were examined by confocal fluorescence microscopyafter staining for IL-12 (AandD), IL-10 (B andE), or isotype control (CandF). Nuclei were counterstained with propidium iodide.A–C, Infectedpopulations;D–F, control populations from uninfected animals.A, Thearrows point to IL-121 neutrophils.B, Numerous neutrophils (e.g., whitearrow) that do not display IL-10 in contrast to the surrounding macro-phages (e.g., green arrow).D, The white arrow indicates a neutrophil stain-ing for IL-12 in an uninfected population.E, The white arrow shows aneutrophil from an uninfected population that does not stain for IL-10. Thescale bar inA represents 20mm. The infected population was composed of36% neutrophils, 46% macrophages, 17% lymphocytes, and 1% eosino-phils as determined by differential counts. The control population was com-posed of 3% neutrophils, 76% macrophages, 19% lymphocytes, and 1%eosinophils.
FIGURE 3. Infected macrophages are apparent by 6 h p.i., but onlyneutrophils stain for IL-12. Six-hour infected peritoneal cells were obtainedby lavage and stained for IL-12 by indirect immunofluorescence as de-scribed in Fig. 2.A, Arrows indicate tachyzoites within macrophages.These cells display little or no evidence of IL-12 production, in contrast tothe surrounding neutrophils.B, Higher magnification of granulocytesshowing punctate cytosolic staining for IL-12. The bar is equivalent to 5mm. The population was composed of 26% neutrophils, 47% macrophages,27% lymphocytes, and 0% eosinophils.
4517The Journal of Immunology
by guest on Novem
ber 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

3B is a higher magnification image of IL-121 neutrophils. Thepunctate appearance of intracellular IL-12 suggests the possibilitythat the cytokine is located within cytoplasmic granules.
To better quantitate IL-12-producing cells, flow cytometric anal-ysis of peritoneal cells was performed. Infected and control peri-toneal cells were obtained 4 h p.i., fixed and permeabilized, andstained with anti-Gr-1 and anti-IL-12 mAb. Fig. 4,A andB, showthe forward and side scatter plots for control and infected popu-lations, respectively. The granulocyte populations, as determinedby Gr-1 back-gating, are demarcated. An influx of granulocyteswas apparent; 13% of the total population inA represented thegranulocyte population, which increased to 56% upon infection.Fig. 4,C andD, demonstrates IL-12 staining by these populations.In Fig. 4C the majority of cells were Gr-12 (80%). Of the Gr-11
subset, 75% of the cells stained for IL-12, while only 4.9% of theGr-12 cells possessed IL-12. Infected populations contained moreGr-11 cells, with 85% staining for IL-12 (Fig. 4D). To determinewhether injection of PBS alone provided sufficient stimulus to trig-ger the IL-12 expression shown in Fig. 4C, we also examinedperitoneal populations from noninjected animals. This cell popu-lation displayed virtually identical characteristics to those shownin Fig. 4C (data not shown).
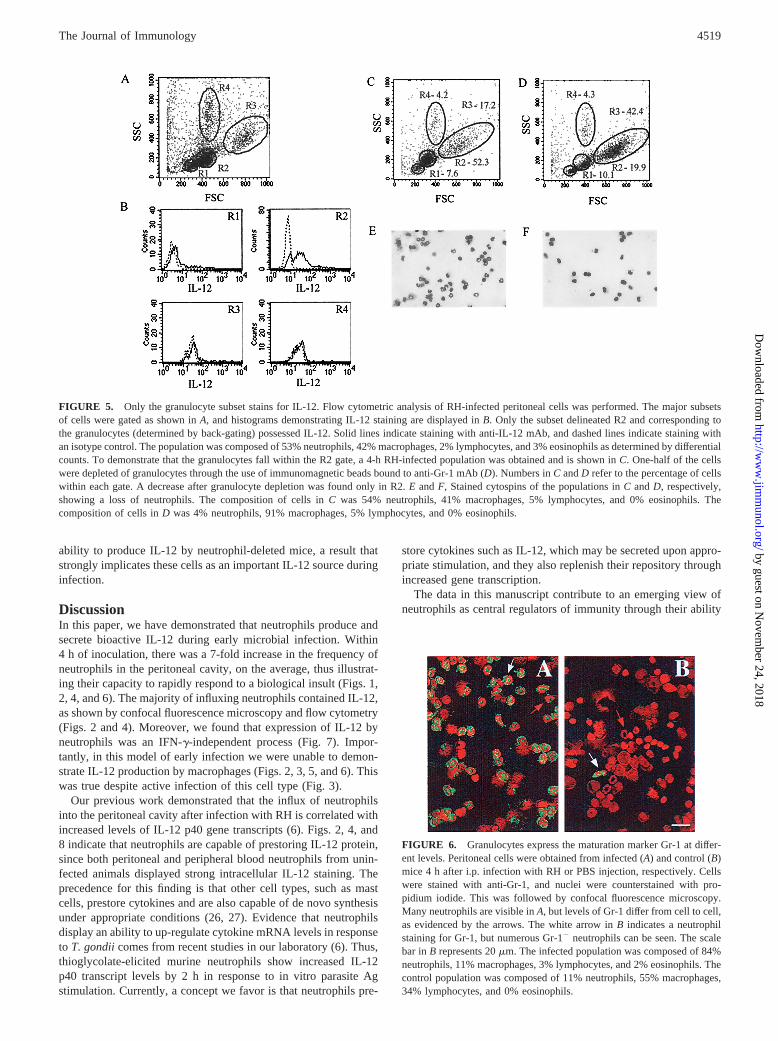
To confirm that only the granulocytes produced IL-12, all themajor subsets in an RH-infected population were gated, as dem-onstrated in Fig. 5B. Only the granulocyte population, labeled R2,stained for IL-12. To demonstrate that the granulocytes fell onlywithin the R2 gate, we obtained a 4-h-infected peritoneal cell pop-ulation (Fig. 5C) and depleted one-half of the population of gran-ulocytes with immunomagnetic beads coated with anti-Gr-1 mAb(Fig. 5D). Only cells contained within the R2 gate decreased inpercentage. Stained cytospin preparations of these populations,shown in Fig. 5,E andF, revealed a decrease in neutrophils (from54 to 4% after depletion). Based upon these data, we conclude that
RH infection induces rapid recruitment of neutrophils, the vastmajority of which contain intracellular IL-12 and express Gr-1.
Differential Gr-1 (Ly-6G) expression by granulocytes
Since gated granulocyte populations in Fig. 4 included Gr-12
cells, we examined Gr-1 expression in infected and control popu-lations (Fig. 6). Different levels of Gr-1 expression are evident inFig. 6A. The white arrow indicates a granulocyte that stainedstrongly for Gr-1, while the red arrow points to a cell displayingcomparatively lower levels. In Fig. 6B, the white arrow indicatesa strongly staining granulocyte within the uninfected population,but many Gr-12 granulocytes may be visualized in this field. Over-all, we found more Gr-12 granulocytes in control populations.
IFN-g is not required for IL-12 production by peritonealneutrophils
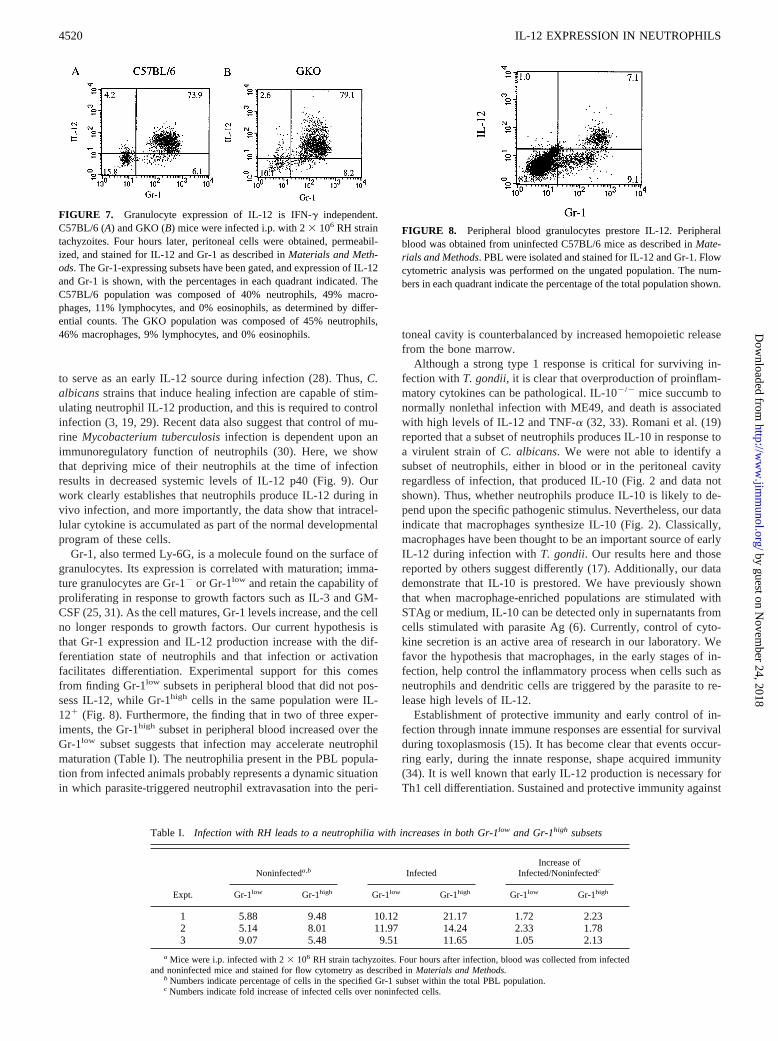
Our previous work has shown that thioglycolate-elicited neutro-phils derived from GKO mice secrete IL-12 p40 in response toSTAg at levels comparable to those in wild-type mice (6). Toconfirm that in vivo infected peritoneal neutrophils from GKOmice produce levels similar to those in wild-type mice, we ob-tained 4-h infected cells and performed a flow cytometric analysisfor IL-12 and Gr-1 expression. Granulocytes from C57BL/6 micewere gated, and staining for IL-12 and Gr-1 is shown in Fig. 7A.Once again, there were Gr-12 cells within this population thatstained poorly for IL-12. Ninety-two percent of the Gr-11 cellsstained for IL-12. Fig. 7Bshows similar results from GKO mice;91% of the Gr-11 cells stained for IL-12. This suggests that IFN-gis not required for IL-12 production by peritoneal neutrophils dur-ing in vivo infection.
A subset of Gr-11 PBL possess IL-12
Since our data indicate that a population of peritoneal neutrophilsfrom noninfected animals contains intracellular IL-12, we won-dered whether they synthesized and stored the cytokine before ex-travasation from the circulation into the peritoneal cavity. To an-swer this question, PBL from normal C57BL/6 mice weresubjected to intracellular cytokine staining and flow cytometricanalysis. As shown in Fig. 8, two populations of Gr-11 cells areapparent. Gr-1low cells, presumably the more immature granulo-cytes (25), did not stain for IL-12. However, the Gr-1high subsetclearly expressed a high level of intracellular IL-12, which indi-cates that Gr-1high granulocytes synthesize and prestore IL-12 aspart of their normal developmental program independently ofpathogenic stimulation. It is unlikely that the Gr-1low cells areeosinophils, as Diff-Quik-stained parallel samples revealed thatthis cell type represented only 5%.
When mice were infected with RH, and PBL were analyzed byflow cytometry 4 h after infection, there was leukocytosis withneutrophilia compared with uninfected mice (Table I and data notshown). Frequencies increased for both Gr-1low and Gr-1high sub-sets. In two of three experiments, the Gr-1high subset increasedmore dramatically than the Gr-1low subset.
Neutrophil-depleted mice have decreased levels of serum IL-12during the acute stage of disease
Our previous work and that of others established that neutrophilsare necessary to survive acute infection with the low virulencestrain ME49 (6, 20, 21). To ascertain whether their impact may beexplained at least in part by an immunoregulatory function, wemeasured serum IL-12 levels in neutrophil-depleted mice from thetime of infection until they became clinically ill, which occurred 8days p.i. Infected mice administered control rat Ig never appearedclinically ill. Fig. 9 demonstrates a systemic impairment in the
FIGURE 4. Flow cytometric analysis of IL-12 production by peritonealcells. C57BL/6 mice were injected with PBS or infected with 23 106 RHstrain tachyzoites. Four hours later peritoneal cells were obtained, perme-abilized, and stained for IL-12 and Gr-1 as described inMaterials andMethods.A and B, Forward and side scatter plots for uninfected and in-fected populations, respectively. The Gr-1-expressing subsets have beengated, and expression of IL-12 and Gr-1 are shown inC andD. The num-bers in each quadrant indicate the percentage of the total population shown.The infected population was composed of 56% neutrophils, 40% macro-phages, 2% lymphocytes, and 2% eosinophils, while the control populationwas composed of 13% neutrophils, 78% macrophages, 9% lymphocytes,and 0% eosinophils.
4518 IL-12 EXPRESSION IN NEUTROPHILS
by guest on Novem
ber 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
ability to produce IL-12 by neutrophil-deleted mice, a result thatstrongly implicates these cells as an important IL-12 source duringinfection.
DiscussionIn this paper, we have demonstrated that neutrophils produce andsecrete bioactive IL-12 during early microbial infection. Within4 h of inoculation, there was a 7-fold increase in the frequency ofneutrophils in the peritoneal cavity, on the average, thus illustrat-ing their capacity to rapidly respond to a biological insult (Figs. 1,2, 4, and 6). The majority of influxing neutrophils contained IL-12,as shown by confocal fluorescence microscopy and flow cytometry(Figs. 2 and 4). Moreover, we found that expression of IL-12 byneutrophils was an IFN-g-independent process (Fig. 7). Impor-tantly, in this model of early infection we were unable to demon-strate IL-12 production by macrophages (Figs. 2, 3, 5, and 6). Thiswas true despite active infection of this cell type (Fig. 3).
Our previous work demonstrated that the influx of neutrophilsinto the peritoneal cavity after infection with RH is correlated withincreased levels of IL-12 p40 gene transcripts (6). Figs. 2, 4, and8 indicate that neutrophils are capable of prestoring IL-12 protein,since both peritoneal and peripheral blood neutrophils from unin-fected animals displayed strong intracellular IL-12 staining. Theprecedence for this finding is that other cell types, such as mastcells, prestore cytokines and are also capable of de novo synthesisunder appropriate conditions (26, 27). Evidence that neutrophilsdisplay an ability to up-regulate cytokine mRNA levels in responseto T. gondiicomes from recent studies in our laboratory (6). Thus,thioglycolate-elicited murine neutrophils show increased IL-12p40 transcript levels by 2 h in response to in vitro parasite Agstimulation. Currently, a concept we favor is that neutrophils pre-
store cytokines such as IL-12, which may be secreted upon appro-priate stimulation, and they also replenish their repository throughincreased gene transcription.
The data in this manuscript contribute to an emerging view ofneutrophils as central regulators of immunity through their ability
FIGURE 6. Granulocytes express the maturation marker Gr-1 at differ-ent levels. Peritoneal cells were obtained from infected (A) and control (B)mice 4 h after i.p. infection with RH or PBS injection, respectively. Cellswere stained with anti-Gr-1, and nuclei were counterstained with pro-pidium iodide. This was followed by confocal fluorescence microscopy.Many neutrophils are visible inA, but levels of Gr-1 differ from cell to cell,as evidenced by the arrows. The white arrow inB indicates a neutrophilstaining for Gr-1, but numerous Gr-12 neutrophils can be seen. The scalebar inB represents 20mm. The infected population was composed of 84%neutrophils, 11% macrophages, 3% lymphocytes, and 2% eosinophils. Thecontrol population was composed of 11% neutrophils, 55% macrophages,34% lymphocytes, and 0% eosinophils.
FIGURE 5. Only the granulocyte subset stains for IL-12. Flow cytometric analysis of RH-infected peritoneal cells was performed. The major subsetsof cells were gated as shown inA, and histograms demonstrating IL-12 staining are displayed inB. Only the subset delineated R2 and corresponding tothe granulocytes (determined by back-gating) possessed IL-12. Solid lines indicate staining with anti-IL-12 mAb, and dashed lines indicate staining withan isotype control. The population was composed of 53% neutrophils, 42% macrophages, 2% lymphocytes, and 3% eosinophils as determined by differentialcounts. To demonstrate that the granulocytes fall within the R2 gate, a 4-h RH-infected population was obtained and is shown inC. One-half of the cellswere depleted of granulocytes through the use of immunomagnetic beads bound to anti-Gr-1 mAb (D). Numbers inC andD refer to the percentage of cellswithin each gate. A decrease after granulocyte depletion was found only in R2.E andF, Stained cytospins of the populations inC andD, respectively,showing a loss of neutrophils. The composition of cells inC was 54% neutrophils, 41% macrophages, 5% lymphocytes, and 0% eosinophils. Thecomposition of cells inD was 4% neutrophils, 91% macrophages, 5% lymphocytes, and 0% eosinophils.
4519The Journal of Immunology
by guest on Novem
ber 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
to serve as an early IL-12 source during infection (28). Thus,C.albicansstrains that induce healing infection are capable of stim-ulating neutrophil IL-12 production, and this is required to controlinfection (3, 19, 29). Recent data also suggest that control of mu-rine Mycobacterium tuberculosisinfection is dependent upon animmunoregulatory function of neutrophils (30). Here, we showthat depriving mice of their neutrophils at the time of infectionresults in decreased systemic levels of IL-12 p40 (Fig. 9). Ourwork clearly establishes that neutrophils produce IL-12 during invivo infection, and more importantly, the data show that intracel-lular cytokine is accumulated as part of the normal developmentalprogram of these cells.
Gr-1, also termed Ly-6G, is a molecule found on the surface ofgranulocytes. Its expression is correlated with maturation; imma-ture granulocytes are Gr-12 or Gr-1low and retain the capability ofproliferating in response to growth factors such as IL-3 and GM-CSF (25, 31). As the cell matures, Gr-1 levels increase, and the cellno longer responds to growth factors. Our current hypothesis isthat Gr-1 expression and IL-12 production increase with the dif-ferentiation state of neutrophils and that infection or activationfacilitates differentiation. Experimental support for this comesfrom finding Gr-1low subsets in peripheral blood that did not pos-sess IL-12, while Gr-1high cells in the same population were IL-121 (Fig. 8). Furthermore, the finding that in two of three exper-iments, the Gr-1high subset in peripheral blood increased over theGr-1low subset suggests that infection may accelerate neutrophilmaturation (Table I). The neutrophilia present in the PBL popula-tion from infected animals probably represents a dynamic situationin which parasite-triggered neutrophil extravasation into the peri-
toneal cavity is counterbalanced by increased hemopoietic releasefrom the bone marrow.
Although a strong type 1 response is critical for surviving in-fection withT. gondii, it is clear that overproduction of proinflam-matory cytokines can be pathological. IL-102/2 mice succumb tonormally nonlethal infection with ME49, and death is associatedwith high levels of IL-12 and TNF-a (32, 33). Romani et al. (19)reported that a subset of neutrophils produces IL-10 in response toa virulent strain ofC. albicans. We were not able to identify asubset of neutrophils, either in blood or in the peritoneal cavityregardless of infection, that produced IL-10 (Fig. 2 and data notshown). Thus, whether neutrophils produce IL-10 is likely to de-pend upon the specific pathogenic stimulus. Nevertheless, our dataindicate that macrophages synthesize IL-10 (Fig. 2). Classically,macrophages have been thought to be an important source of earlyIL-12 during infection withT. gondii. Our results here and thosereported by others suggest differently (17). Additionally, our datademonstrate that IL-10 is prestored. We have previously shownthat when macrophage-enriched populations are stimulated withSTAg or medium, IL-10 can be detected only in supernatants fromcells stimulated with parasite Ag (6). Currently, control of cyto-kine secretion is an active area of research in our laboratory. Wefavor the hypothesis that macrophages, in the early stages of in-fection, help control the inflammatory process when cells such asneutrophils and dendritic cells are triggered by the parasite to re-lease high levels of IL-12.
Establishment of protective immunity and early control of in-fection through innate immune responses are essential for survivalduring toxoplasmosis (15). It has become clear that events occur-ring early, during the innate response, shape acquired immunity(34). It is well known that early IL-12 production is necessary forTh1 cell differentiation. Sustained and protective immunity against
FIGURE 7. Granulocyte expression of IL-12 is IFN-g independent.C57BL/6 (A) and GKO (B) mice were infected i.p. with 23 106 RH straintachyzoites. Four hours later, peritoneal cells were obtained, permeabil-ized, and stained for IL-12 and Gr-1 as described inMaterials and Meth-ods. The Gr-1-expressing subsets have been gated, and expression of IL-12and Gr-1 is shown, with the percentages in each quadrant indicated. TheC57BL/6 population was composed of 40% neutrophils, 49% macro-phages, 11% lymphocytes, and 0% eosinophils, as determined by differ-ential counts. The GKO population was composed of 45% neutrophils,46% macrophages, 9% lymphocytes, and 0% eosinophils.
FIGURE 8. Peripheral blood granulocytes prestore IL-12. Peripheralblood was obtained from uninfected C57BL/6 mice as described inMate-rials and Methods. PBL were isolated and stained for IL-12 and Gr-1. Flowcytometric analysis was performed on the ungated population. The num-bers in each quadrant indicate the percentage of the total population shown.
Table I. Infection with RH leads to a neutrophilia with increases in both Gr-1low and Gr-1high subsets
Noninfecteda,b InfectedIncrease of
Infected/Noninfectedc
Expt. Gr-1low Gr-1high Gr-1low Gr-1high Gr-1low Gr-1high
1 5.88 9.48 10.12 21.17 1.72 2.232 5.14 8.01 11.97 14.24 2.33 1.783 9.07 5.48 9.51 11.65 1.05 2.13
a Mice were i.p. infected with 23 106 RH strain tachyzoites. Four hours after infection, blood was collected from infectedand noninfected mice and stained for flow cytometry as described inMaterials and Methods.
b Numbers indicate percentage of cells in the specified Gr-1 subset within the total PBL population.c Numbers indicate fold increase of infected cells over noninfected cells.
4520 IL-12 EXPRESSION IN NEUTROPHILS
by guest on Novem
ber 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

T. gondii requires a type 1 response with subsequent IFN-g pro-duction. Thus, it is important to understand how the immune re-sponse is initiated. Our hypothesis is that neutrophils play an im-munomodulatory role in infection, and furthermore, that they areinvolved in the earliest recognition of the parasite and generationof immunity. The capacity to prestore IL-12 may allow for rapidmobilization and secretion after stimulation. Also, the ability toincrease transcript levels suggests a sustained response to the par-asite. The mechanisms by which neutrophil-derived IL-12 is re-leased and its influence on T cell differentiation are currently beingexplored in our laboratory.
AcknowledgmentsWe are grateful to Drs. T. G. Clark and E. J. Pearce for critical review ofthe manuscript.
References1. Anderson, K. L., K. A. Smith, F. Pio, B. E. Torbett, and R. A. Maki. 1998.
Neutrophils deficient in PU.1 do not terminally differentiate or become function-ally competent.Blood 92:1576.
2. Rogers, H. W., and E. R. Unanue. 1993. Neutrophils are involved in acute, non-specific resistance toListeria monocytogenesin mice. Infect. Immun. 61:5090.
3. Romani, L., A. Mencacci, E. Cenci, G. Del Sero, F. Bistoni, and P. Puccetti.1997. An immunoregulatory role for neutrophils in CD41 T helper subset selec-tion in mice with candidiasis.J. Immunol. 158:2356.
4. Conlan, J. W., and R. J. North. 1994. Neutrophils are essential for early anti-Listeriadefence in the liver, but not in the spleen or peritoneal cavity, as revealedby a granulocyte-depleting monoclonal antibody.J. Exp. Med. 179:259.
5. Czuprynski, C. J., J. F. Brown, N. Maroushek, R. D. Wagner, and H. Steinberg.1994. Administration of anti-granulocyte mAb RB6-8C5 impairs the resistance ofmice toListeria monocytogenesinfection.J. Immunol. 152:1836.
6. Bliss, S. K., Y. Zhang, and E. Y. Denkers. 1999. Murine neutrophil stimulationby Toxoplasma gondiiantigen drives high level production of IFN-g-independentIL-12. J. Immunol. 163:2081.
7. Lehrer, R. I., and T. Ganz. 1990. Antimicrobial polypeptides of human neutro-phils. Blood 76:2169.
8. Tapper, H. 1996. The secretion of preformed granules by macrophages and neu-trophils.J. Leukocyte Biol. 59:613.
9. McDonald, P. P., S. Gasperini, F. Calzetti and M. A. Cassatella. 1998. Modula-tion by interferon-g of the production and gene expression of IL-1 receptor an-tagonist in human neutrophils.Cell. Immunol. 184:45.
10. Djeu, J. Y., D. Serbousek, and D. K. Blanchard. 1990. Release of tumor necrosisfactor by human polymorphonuclear leukocytes.Blood 76:1405.
11. Mead, P. S., L. Slutzker, V. Dietz, L. F. McCaig, J. S. Bresee, C. Shapiro,P. M. Griffen, and R. V. Tauxe. 1999. Food related illness and death in the UnitedStates.Emerg. Infect. Dis. 5:607.
12. Navia, B. A., C. K. Petito, J. W. M. Gold, E. S. Cho, B. D. Jordon, andJ. W. Price. 1986. Cerebral toxoplasmosis complicating the acquired immunedeficiency syndrome: clinical and neuropathological findings in 27 patients.Ann.Neurol. 19:224.
13. Remington, J. S., and G. Desmonts. 1990. Toxoplasmosis. InInfectious Diseasesof the Fetus and Newborn Infant.J. S. Remington and J. O. Klein, eds.W.B. Saunders, Philadelphia, p. 89.
14. Alexander, J., and C. A. Hunter. 1998. Immunoregulation during toxoplasmosis.Chem. Immunol. 70:81.
15. Denkers, E. Y., and R. T. Gazzinelli. 1998. Regulation and function of T cell-mediated immunity duringToxoplasma gondiiinfection. Clin. Microbiol. Rev.11:569.
16. Gazzinelli, R. T., S. Hieny, T. Wynn, S. Wolf, and A. Sher. 1993. IL-12 isrequired for the T-cell independent induction of IFN-g by an intracellular parasiteand induces resistance in T-cell-deficient hosts.Proc. Natl. Acad. Sci. USA 90:6115.
17. Reis e Sousa, C., S. Hieny, T. Scharton-Kersten, D. Jankovic, H. Charest,R. N. Germain, and A. Sher. 1997. In vivo microbial stimulation induces rapidCD40L-independent production of IL-12 by dendritic cells and their re-distribu-tion to T cell areas.J. Exp. Med. 186:1819.
18. Bliss, S. K., A. Marshall, Y. Zhang, and E. Denkers. 1999. Human polymorpho-nuclear leukocytes produce IL-12, TNF-a, and the chemokines macrophage in-flammatory protein-1a and -1b in response toToxoplasma gondiiantigens.J,Immunol. 162:7369.
19. Romani, L., A. Mencacci, E. Cenci, R. Spaccapelo, G. Del Sero, I. Nicoletti,G. Trinchieri, F. Bistoni, and P. Puccetti. 1997. Neutrophil production of IL-12and IL-10 in candidiasis and efficacy of IL-12 therapy in neutropenic mice.J. Im-munol. 158:5349.
20. Sayles, P. C., and L. J. Johnson. 1997. Exacerbation of toxoplasmosis in neutro-phil depleted mice.Nat. Immun. 15:249.
21. Scharton-Kersten, T., G. Yap, J. Magram, and A. Sher. 1997. Inducible nitricoxide is essential for host control of persistent but not acute infection with theintracellular pathogenToxoplasma gondii. J. Exp. Med. 185:1.
22. Wysocka, M., M. Kubin, L. Q. Vieira, L. Ozmen, G. Garotta, P. Scott, andG. Trinchieri. 1995. Interleukin-12 is required for interferon-g production andlethality in lipopolysaccharide-induced shock in mice.Eur. J. Immunol. 25:672.
23. Vowells, S. J., S. Sekhsaria, H. L. Malech, M. Shalit, and T. A. Fleisher. 1995.Flow cytometric analysis of the granulocyte respiratory burst: a comparison studyof fluorescent probes.J. Immunol. Methods 178:89.
24. Biermann, H., B. Pietz, R. Dreier, K. W. Schmid, C. Sorg, and C. Sunderkotter.1999. Murine leukocytes with ring-shaped nuclei include granulocytes, mono-cytes, and their precursors.J. Leukocyte Biol. 65:217.
25. Fleming, T. J., M. L. Fleming, and T. R. Malek. 1993. Selective expression ofLy-6G on myeloid lineage cells in mouse bone marrow.J. Immunol. 151:2399.
26. Arock, M., E. Ross, R. Lai-Kuen, G. Averlant, Z. Gao, and S. N. Abraham. 1998.Phagocytic and tumor necrosis factora response of human mast cells followingexposure to Gram-negative and Gram-positive bacteria.Infect. Immun. 66:6030.
27. Abraham, S. N., and M. Arock. 1998. Mast cells and basophils in innate immu-nity. Immunology 10:373.
28. Denkers, E. Y., and A. J. Marshall. 1998. Neutrophils as a source of immuno-regulatory cytokines during microbial infection.Immunologist 6:116.
29. Romani, L., F. Bistoni, and P. Puccetti. 1997. Initiation of T-helper cell immunityto Candida albicansby IL-12: the role of neutrophils.Chem. Immunol. 68:110.
30. Pedrosa, J., B. M. Saunders, R. Appelberg, I. M. Orme, M. T. Silva, andA. M. Cooper. 2000. Neutrophils play a protective nonphagocytic role in sys-temic Mycobacterium tuberculosisinfection in mice.Infect. Immun. 68:577.
31. Hestdal, K., F. W. Ruscetti, J. N. Ihle, S. E. W. Jacobsen, C. M. Dubois,W. C. Kopp, D. L. Longo, and J. R. Keller. 1991.Characterization and regulationof RB6-8C5 antigen expression on murine bone marrow cells.J. Immunol. 147:22.
32. Neyer, L. E., G. Grunig, M. Fort, J. S. Remington, D. Rennick, and C. A. Hunter.1997. Role of interleukin-10 in regulation of T-cell-dependent and T-cell-inde-pendent mechanisms of resistance toToxoplasma gondii. Infect. Immun. 65:1675.
33. Gazzinelli, R. T., M. Wysocka, S. Hieny, T. Scharton-Kersten, A. Cheever,R. Kuhn, W. Muller, G. Trinchieri, and A. Sher. 1996. In the absence of endog-enous IL-10, mice acutely infected withToxoplasma gondiisuccumb to a lethalimmune response dependent upon CD41 T cells and accompanied by overpro-duction of IL-12, IFN-g, and TNF-a. J. Immunol. 157:798.
34. Hoffman, J. A., F. C. Kafatos, C. A. Janeway, and R. A. Ezekowitz. 1999. Phy-logenetic perspectives in innate immunity.Science 284:1313.
FIGURE 9. Systemic levels of IL-12 p40 are decreased in neutrophil-depleted, infected mice. Mice (five per group) were administered 200mg ofanti-Gr-1 mAb (depleted, infected) or a control rat Ig (control, infected) i.p.on days22, 0,12, and14. Mice were infected with 100 ME49 cysts i.p.on day 0. Data from two uninfected mice were also included. Serum wasobtained every 2 days beginning on day 0, and levels of IL-12 p40 weredetermined by ELISA. Results are expressed as the mean6 SD. Differ-ences between control, infected, and depleted, infected mice were statisti-cally significant on days 4, 6, and 8.
4521The Journal of Immunology
by guest on Novem
ber 24, 2018http://w
ww
.jimm
unol.org/D
ownloaded from