rapid determination of quinolone resistance in acinetobacter spp
TRANSCRIPT

JOURNAL OF CLINICAL MICROBIOLOGY, May 2009, p. 1436–1442 Vol. 47, No. 50095-1137/09/$08.00�0 doi:10.1128/JCM.02380-08Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Rapid Determination of Quinolone Resistance in Acinetobacter spp.�
Kristine M. Hujer,1,2 Andrea M. Hujer,2 Andrea Endimiani,1,2 Jodi M. Thomson,3 Mark D. Adams,4Karrie Goglin,4 Philip N. Rather,5 Thuy-Trang D. Pennella,6 Christian Massire,6 Mark W. Eshoo,6
Rangarajan Sampath,6 Lawrence B. Blyn,6 David J. Ecker,6* and Robert A. Bonomo1,2,3,7*Department of Medicine, Case Western Reserve University School of Medicine, Cleveland, Ohio1; Research Service,
Louis Stokes Cleveland Department of Veterans Affairs Medical Center, Cleveland, Ohio2; Departments ofPharmacology3 and Genetics,4 Case Western Reserve University School of Medicine, Cleveland, Ohio;Departments of Microbiology and Immunology, Emory University School of Medicine, Atlanta, Georgia5;
Ibis Biosciences, Carlsbad, California6; and Molecular Biology and Microbiology,Case Western Reserve University School of Medicine, Cleveland, Ohio7
Received 11 December 2008/Returned for modification 3 February 2009/Accepted 11 March 2009
In the treatment of serious bacterial infections, the rapid institution of appropriate antimicrobial chemo-therapy may be lifesaving. Choosing the correct antibiotic or combination of antibiotics is becoming veryimportant, as multidrug resistance is found in many pathogens. Using a collection of 75 well-characterizedmultidrug-resistant (MDR) Acinetobacter sp. isolates, we show that PCR followed by electrospray ionizationmass spectrometry (PCR/ESI-MS) and base composition analysis of PCR amplification products can quicklyand accurately identify quinolone resistance mediated by mutations in the quinolone resistance-determiningregions of gyrA and parC, two essential housekeeping genes. Single point mutations detected by PCR/ESI-MSin parC (found in 55/75 of the isolates) and in gyrA (found in 66/75 of the isolates) correlated with susceptibilitytesting and sequencing. By targeting resistance determinants that are encoded by genes with highly conservedDNA sequences (e.g., gyrA and parC), we demonstrate that PCR/ESI-MS can provide critical information forresistance determinant identification and can inform therapeutic decision making in the treatment of Acin-etobacter sp. infections.
Acinetobacter baumannii is a gram-negative, aerobic, non-lactose-fermenting bacterium of increasing clinical importance(10, 14, 19, 21, 23). Since the 1980s, the number of reportsdescribing infections due to this organism has risen dramati-cally (5, 16, 20). Most A. baumannii infections occur withinintensive care units in patients with serious underlying disease(2, 19). Sites of infection include lung, bloodstream, urinarytract, and skin (20). In addition, A. baumannii has emerged asan important pathogen in injured United States military per-sonnel stationed in Afghanistan and Iraq as they return tomajor military medical treatment facilities (14, 24–26, 31).
An unwelcome feature of A. baumannii is the frequency inwhich isolates manifest resistance to the most commonly usedantibiotics (12, 14). When resistance to three or more classes ofantibiotics is demonstrated, A. baumannii is referred to asmultidrug resistant (MDR) (21). Among MDR A. baumanniiisolates, resistance to quinolones (e.g., ciprofloxacin and levo-floxacin), aminoglycosides (e.g., gentamicin and amikacin),ampicillin-sulbactam, extended-spectrum cephalosporins (e.g.,ceftazidime), and carbapenems (e.g., imipenem, meropenem,and doripenem) is escalating (21, 23). Concern exists that, ifthe incorrect antimicrobial agents are chosen to treat MDR A.baumannii infection, the outcome of patients may be poor (9,
22). Thus, a rapid assessment of antimicrobial susceptibilitycould have a significant impact on patient care.
In this report, we employ a rapid high-throughput method toidentify unique DNA changes associated with quinolone resis-tance in a collection of MDR A. baumannii isolates. Themethod was originally developed to identify and genotype var-ious species of bacteria from complex mixtures in respiratorysamples (6). This process uses electrospray ionization massspectrometry (ESI-MS) and base composition analysis of PCRamplification products derived from highly conserved genes(“housekeeping genes”). The primers that are designed forPCR/ESI-MS yield amplicons with unique mass signatures thatcan be measured by high-performance mass spectrometry andidentified by base composition analysis (7, 13).
The best-described mechanism of resistance to quinolonesin Acinetobacter spp. is mutations in the genes encoding DNAgyrase A (i.e., gyrA) and subunit A of topoisomerase IV (i.e.,parC). The most important mutations, resulting in changes atcodon 83 for gyrA and at codon 80 for parC, have been mappedto a unique location in each of these genes, the quinoloneresistance-determining region (QRDR) (27, 28). Additionalmutations in the QRDR of gyrA thought to enhance quinoloneresistance include mutations at codons for amino acids Gly81,Ala84, and Glu87 (1, 11, 17, 27, 28, 30). Another previouslydescribed mutation (at the Val101 codon) does not appear tohave an effect on the susceptibility profiles (1). Ancillarymutations in parC, at codons for amino acids Gly78 andGlu84, when combined with the Ser83 and Ser80 codonmutations, also contribute to high-level quinolone resistance(11, 27, 30).
Appreciating that mutations in the housekeeping genes gyrA
* Corresponding author. Mailing address for David J. Ecker: IbisBiosciences, 1891 Rutherford Road, Carlsbad, CA 92008. Phone: (760)603-2347. Fax: (760) 603-4653. E-mail: [email protected]. Mailingaddress for Robert A. Bonomo: Louis Stokes Cleveland Departmentof Veterans Affairs Medical Center, 10701 East Blvd., Cleveland, OH44106. Phone: (216) 791-3800, ext. 4399. Fax: (216) 229-8509. E-mail:[email protected].
� Published ahead of print on 18 March 2009.
1436
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 27
Dec
embe
r 20
21 b
y 18
8.21
4.24
3.74
.
and parC also define the quinolone resistance phenotype, wereasoned that PCR/ESI-MS and base composition analysis canbe applied to determine the sequence variability of theQRDRs of Acinetobacter spp. and rapidly predict a quinolone-resistant phenotype. Therefore, we chose to study 73 well-characterized clinical isolates of A. baumannii, one Acineto-bacter genome species strain 3 isolate, and one Acinetobacterjohnsonii isolate. This paper describes the novel application ofthis highly sensitive and specific method to the analysis ofresistance phenotypes in this clinically important pathogen.
MATERIALS AND METHODS
Bacterial isolates. The Acinetobacter sp. isolates were obtained from 75 civilianand military patients treated at the Walter Reed Army Medical Center in Wash-ington, DC. All isolates were collected from March 2003 through April 2005 andrepresent 16 unique Acinetobacter sp. clone types (14).
Determination of susceptibility to quinolones. MICs of ciprofloxacin and levo-floxacin were determined by the broth microdilution method using cation-ad-justed Mueller-Hinton medium according to the Clinical and Laboratory Stan-dards Institute (CLSI) standard criteria (3). Specific panels were “custom-made”by Trek Diagnostics (Cleveland, OH) to better measure the level of quinoloneresistance. Ciprofloxacin was tested in the range of 0.06 to 64 mg/liter, whereaslevofloxacin was tested in the range of 0.06 to 32 mg/liter. ATCC control strains,including A. baumannii isolates 9955 and 17961, Escherichia coli 25922, Pseudo-monas aeruginosa 27853, and Klebsiella pneumoniae 700603, were used. Suscep-tibility results were interpreted according to the guidelines recommended byCLSI (4). In this paper, A. baumannii isolates were defined as quinolone resistantwhen the ciprofloxacin or levofloxacin MIC or both MICs were in the nonsus-ceptible range (MIC � 1 mg/liter).
PCR amplification for RE digestion and DNA sequencing. All primers used forPCR amplification along with the product sizes are listed in Table 1. The re-striction endonuclease (RE) digestions with HinfI of the gyrA and parC QRDRshave been previously described (14).
Amplification of the gyrA and parC genes for direct sequencing was performed
using the forward and reverse primers listed in Table 1. A 1:10 dilution of anovernight lysogeny broth culture was boiled for 15 min. High-fidelity amplifica-tion using rTth polymerase (GeneAmp XL kit; Applied Biosystems, Foster City,CA) was performed with 5 �l of this dilution as the DNA template. PCRconditions included an initial denaturation at 95°C for 1 min followed by 30cycles of 95°C for 30 s and annealing for 45 s at 54°C for both gyrA and parC, withan extension at 72°C for 30 s. Cycling was followed by a final extension at 72°Cfor 5 min. The amplified products (25 �l) were purified with 5 U exonuclease Iand 1 U shrimp alkaline phosphatase (USB Corporation, Cleveland, OH) andwere incubated for 30 min at 37°C, 15 min at 45°C, and 80°C for 15 min todegrade the unincorporated primers and nucleotides. Sequencing reactions wereperformed using nested primers (Table 1) and employing Applied BiosystemsBigDye v1.1 sequencing kits. Sequence data were obtained on an Applied Bio-systems 3730xl DNA analyzer. Trace files were analyzed by Vector NTI (Invitro-gen, Carlsbad, CA), and the resulting consensus sequences were compared toGenBank (www.ncbi.nlm.nih.gov/GenBank) reference sequences X82165 forgyrA and X95819 for parC of A. baumannii.
DNA preparation and PCR amplification for PCR/ESI-MS. Template DNAwas prepared by making a 1:100 dilution of a lysogeny broth overnight culture inTE buffer (10 mM Tris, 1 mM EDTA [pH 8.0]) and boiling for 15 min. All PCRswere performed in 96-well plates. The PCR mixture consisted of 3 U of Ampli-Taq Gold polymerase (Applied Biosystems, Foster City, CA); 20 mM Tris (pH8.3); 75 mM KCl; 1.5 mM MgCl2; 0.4 M betaine; 800 �M equal mixture of dCTP,dTTP, dGTP, and dATP; and 250 nM of each primer. PCR cycling under thefollowing conditions was performed on an MJ Dyad 96-well thermocycler (Bio-Rad Inc., Hercules, CA): 95°C for 10 min, followed by 8 cycles of 95°C for 30 s,48°C for 30 s, and 72°C 30 s, with the 48°C annealing temperature increasing0.9°C each cycle. PCR was then continued for 37 additional cycles of 95°C for15 s, 56°C for 20 s, and 72°C for 20 s. The PCR cycle ended with a final extensionof 2 min at 72°C followed by a 4°C hold. Five microliters of the template DNAwas used in each PCR.
Primer design for PCR/ESI-MS and base composition analysis. The primerpairs used for mass spectrometry and base composition analysis of the QRDRsof gyrA and parC in Acinetobacter spp. are also listed in Table 1. To completelycover the QRDR, three sets of overlapping primers were constructed for bothtopoisomerase genes. Two sets of primers encompass the flanking regions as wellas the nucleotides in parC that encode amino acid Ser80 or those in gyrA that
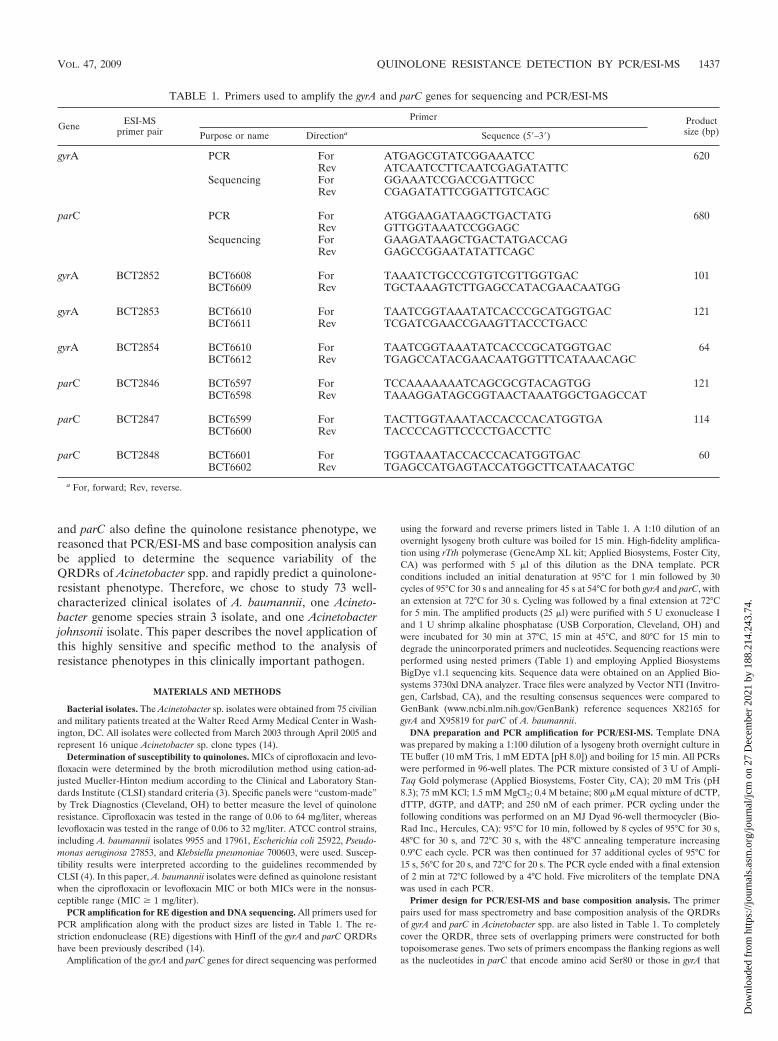
TABLE 1. Primers used to amplify the gyrA and parC genes for sequencing and PCR/ESI-MS
Gene ESI-MSprimer pair
Primer Productsize (bp)Purpose or name Directiona Sequence (5�–3�)
gyrA PCR For ATGAGCGTATCGGAAATCC 620Rev ATCAATCCTTCAATCGAGATATTC
Sequencing For GGAAATCCGACCGATTGCCRev CGAGATATTCGGATTGTCAGC
parC PCR For ATGGAAGATAAGCTGACTATG 680Rev GTTGGTAAATCCGGAGC
Sequencing For GAAGATAAGCTGACTATGACCAGRev GAGCCGGAATATATTCAGC
gyrA BCT2852 BCT6608 For TAAATCTGCCCGTGTCGTTGGTGAC 101BCT6609 Rev TGCTAAAGTCTTGAGCCATACGAACAATGG
gyrA BCT2853 BCT6610 For TAATCGGTAAATATCACCCGCATGGTGAC 121BCT6611 Rev TCGATCGAACCGAAGTTACCCTGACC
gyrA BCT2854 BCT6610 For TAATCGGTAAATATCACCCGCATGGTGAC 64BCT6612 Rev TGAGCCATACGAACAATGGTTTCATAAACAGC
parC BCT2846 BCT6597 For TCCAAAAAAATCAGCGCGTACAGTGG 121BCT6598 Rev TAAAGGATAGCGGTAACTAAATGGCTGAGCCAT
parC BCT2847 BCT6599 For TACTTGGTAAATACCACCCACATGGTGA 114BCT6600 Rev TACCCCAGTTCCCCTGACCTTC
parC BCT2848 BCT6601 For TGGTAAATACCACCCACATGGTGAC 60BCT6602 Rev TGAGCCATGAGTACCATGGCTTCATAACATGC
a For, forward; Rev, reverse.
VOL. 47, 2009 QUINOLONE RESISTANCE DETECTION BY PCR/ESI-MS 1437
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 27
Dec
embe
r 20
21 b
y 18
8.21
4.24
3.74
.

encode Ser83 (Fig. 1). The third set of primers specifically targeted only thenucleotides that encode the Ser83 or Ser80 amino acid. The length of eachamplified region is listed in Table 1.
ESI-MS. The study was performed using the T5000 Biosensor (Ibis Bio-sciences, Carlsbad, CA). After amplification, 15-�l aliquots of each PCR mixturewere desalted and purified using a weak-anion-exchange protocol as previouslydescribed (15). Accurate-mass (�1 ppm), high-resolution (M/�M � 100,000full-width half-maximal) mass spectra were acquired for each sample by usinghigh-throughput ESI-MS protocols described previously (13). For each sample,1.5 �l of analyte solution was consumed during the 74-s spectral acquisition. Rawmass spectra were converted to monoisotopic molecular masses. Unambiguousbase compositions were derived from the exact mass measurements of the com-plementary single-stranded oligonucleotides (18). Quantitative results were ob-tained by comparing the peak heights with an internal PCR calibration standardpresent in every PCR well at 100 molecules (13). The ESI-MS measurementsrequired approximately 45 s per PCR, so analysis of each Acinetobacter sp. isolaterequired less than 5 min of mass spectrometry time (338 min for 75 samples).
RESULTS
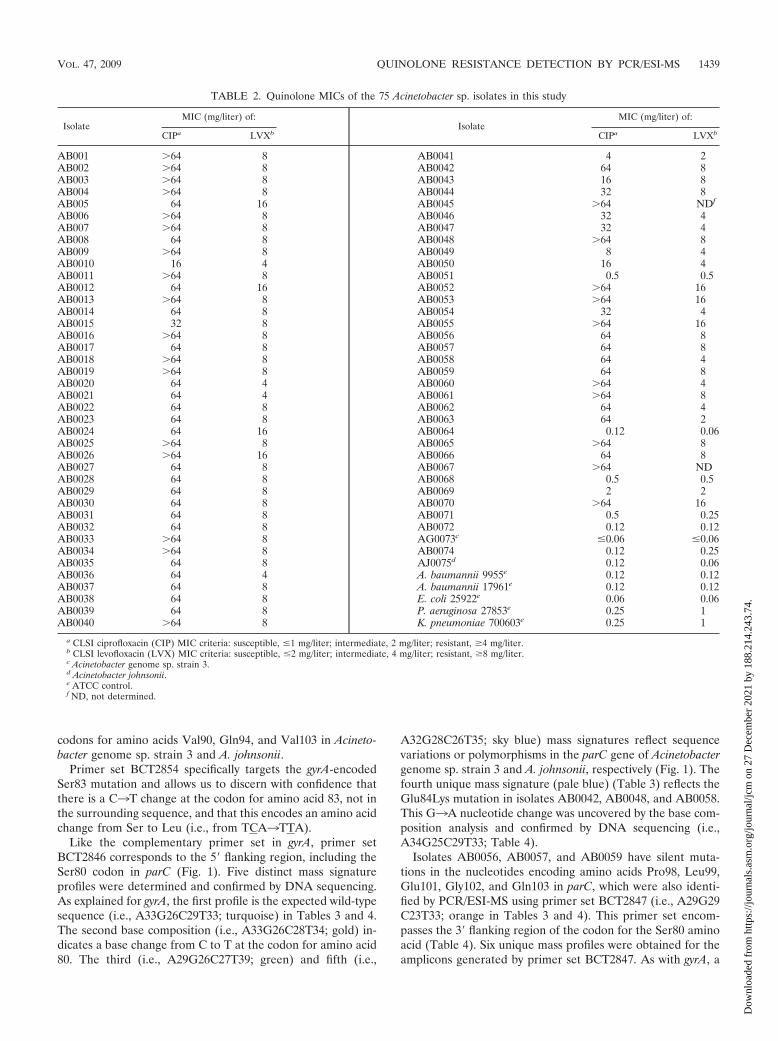
Susceptibility testing. The results of quinolone susceptibilitytesting for all 75 Acinetobacter sp. isolates are summarized inTable 2. Overall, 89.3% (67/75) of the isolates in this collectiondemonstrated MICs to quinolones in the resistance range (allof these isolates had ciprofloxacin MICs of �2 mg/liter). High-level ciprofloxacin resistance (i.e., �64 mg/liter) was seen in74.7% of the isolates. Similarly, 84.9% (62/73) of the isolateswere resistant (i.e., MIC � 4 mg/liter) to levofloxacin.
PCR amplification, RE digestion, and DNA sequencing ofgyrA and parC. To determine if RE digestion patterns usingHinfI obtained in our laboratory correctly identified mutationsin the QRDR loci, we sequenced the QRDR and flankingregions of each topoisomerase gene. Analysis of the QRDRs ofgyrA and parC revealed mutations in this area previously de-scribed as resulting in ciprofloxacin resistance. In addition,DNA sequencing of the parC QRDR amplification productsshowed a Glu84Lys mutation in three isolates (i.e., AB0042,AB0048, and AB0058).
We further determined that isolates AB0056, AB0057, andAB0059 possess silent mutations in the codons of parC encod-ing Pro98, Leu99, Glu101, Gly102, and Gln103 (the impor-tance of these mutations is still unknown). Comparing ourresults with the reference sequence, we discovered that silentmutations were found in gyrA for isolate AG0073 (Acineto-bacter genome sp. strain 3) and AJ0075 (A. johnsonii) atcodons for amino acids Ala67, Val69, Pro79, Val90, Gln94, andVal103.
PCR/ESI-MS and base composition analysis. In Table 3, wepresent the target base composition analysis for all PCR/ESI-MS primer sets resulting from the generated monoiso-topic masses for each forward and reverse PCR ampliconstrand. Table 4 lists the ESI-MS data for both the gyrA andparC primer sets for each Acinetobacter sp. isolate analyzed.For ease of interpretation, the various base compositions de-tected by ESI-MS are color coded and correspond to substitu-tions from the wild-type sequence (Table 3). In every case,when base substitutions were detected using PCR/ESI-MS,standard DNA sequencing confirmed these findings (100%correlation). In addition, MIC determination identified 67 outof 75 (89.3%) strains as quinolone resistant. By PCR/ESI-MS88% of the isolates are predicted to be quinolone resistant, andoverall the accurate prediction of quinolone susceptibility orresistance was 98.7% (74/75 isolates).
By using three sets of overlapping primers, we were able todiscriminate the Ser83 and Ser80 point mutations as well asdetect mutations in the remaining nucleotides spanning theQRDRs of both gyrA and parC. The deduced overall net basechanges (addition or deletion of A, G, C, or T) from theexpected wild-type sequence are listed in Table 4. As statedabove, three overlapping primer sets were used to target theregions of interest in gyrA. Primer set BCT2852, which corre-sponds to the 5� flanking region through the Ser83 codonposition in gyrA (Fig. 1 and Table 4), yielded three distinctmass signature profiles. The wild-type sequence (i.e.,A25G23C22T31; turquoise) was found in strains AB0051,AB0064, AB0068, AB0069, AB0071, AB0072, and AB0074.The second base composition (i.e., A25G23C21T32; gold) in-dicates a base change from C to T. The third mass signature(i.e., A25G22C22T32; green) reflects sequence variations orpolymorphisms observed in the gyrA genes from different Acin-etobacter spp. (Acinetobacter genome sp. strain 3 and A. johnso-nii).
Primer set BCT2853 amplifies the nucleotide sequence ofthe gyrA QRDR that encodes amino acids 83 to 104. Thisprimer set also yielded three distinct mass signature profiles.As with primer set BCT2852, the wild-type signature (tur-quoise), the second distinct signature representing theSer83Leu mutation (gold), and the third signature (green)represent sequence variations that correspond to changes in
FIG. 1. Priming strategy used for PCR/ESI-MS of the QRDRs of gyrA and parC in Acinetobacter baumannii. Six primer pairs (BCT2852 toBCT2854 and BCT2846 to BCT2848) were developed to cover the entire length of the gyrA and parC QRDRs, respectively. The principalmutations captured by these primer pairs are indicated below the wild-type sequences. Colored boxes indicate the scope of these primer pairs, allof them covering the structurally homologous gyrA codon for Ser83 and parC codon for Ser80.
1438 HUJER ET AL. J. CLIN. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 27
Dec
embe
r 20
21 b
y 18
8.21
4.24
3.74
.
codons for amino acids Val90, Gln94, and Val103 in Acineto-bacter genome sp. strain 3 and A. johnsonii.
Primer set BCT2854 specifically targets the gyrA-encodedSer83 mutation and allows us to discern with confidence thatthere is a C3T change at the codon for amino acid 83, not inthe surrounding sequence, and that this encodes an amino acidchange from Ser to Leu (i.e., from TCA3TTA).
Like the complementary primer set in gyrA, primer setBCT2846 corresponds to the 5� flanking region, including theSer80 codon in parC (Fig. 1). Five distinct mass signatureprofiles were determined and confirmed by DNA sequencing.As explained for gyrA, the first profile is the expected wild-typesequence (i.e., A33G26C29T33; turquoise) in Tables 3 and 4.The second base composition (i.e., A33G26C28T34; gold) in-dicates a base change from C to T at the codon for amino acid80. The third (i.e., A29G26C27T39; green) and fifth (i.e.,
A32G28C26T35; sky blue) mass signatures reflect sequencevariations or polymorphisms in the parC gene of Acinetobactergenome sp. strain 3 and A. johnsonii, respectively (Fig. 1). Thefourth unique mass signature (pale blue) (Table 3) reflects theGlu84Lys mutation in isolates AB0042, AB0048, and AB0058.This G3A nucleotide change was uncovered by the base com-position analysis and confirmed by DNA sequencing (i.e.,A34G25C29T33; Table 4).
Isolates AB0056, AB0057, and AB0059 have silent muta-tions in the nucleotides encoding amino acids Pro98, Leu99,Glu101, Gly102, and Gln103 in parC, which were also identi-fied by PCR/ESI-MS using primer set BCT2847 (i.e., A29G29C23T33; orange in Tables 3 and 4). This primer set encom-passes the 3� flanking region of the codon for the Ser80 aminoacid (Table 4). Six unique mass profiles were obtained for theamplicons generated by primer set BCT2847. As with gyrA, a
TABLE 2. Quinolone MICs of the 75 Acinetobacter sp. isolates in this study
IsolateMIC (mg/liter) of:
IsolateMIC (mg/liter) of:
CIPa LVXb CIPa LVXb
AB001 �64 8 AB0041 4 2AB002 �64 8 AB0042 64 8AB003 �64 8 AB0043 16 8AB004 �64 8 AB0044 32 8AB005 64 16 AB0045 �64 NDf
AB006 �64 8 AB0046 32 4AB007 �64 8 AB0047 32 4AB008 64 8 AB0048 �64 8AB009 �64 8 AB0049 8 4AB0010 16 4 AB0050 16 4AB0011 �64 8 AB0051 0.5 0.5AB0012 64 16 AB0052 �64 16AB0013 �64 8 AB0053 �64 16AB0014 64 8 AB0054 32 4AB0015 32 8 AB0055 �64 16AB0016 �64 8 AB0056 64 8AB0017 64 8 AB0057 64 8AB0018 �64 8 AB0058 64 4AB0019 �64 8 AB0059 64 8AB0020 64 4 AB0060 �64 4AB0021 64 4 AB0061 �64 8AB0022 64 8 AB0062 64 4AB0023 64 8 AB0063 64 2AB0024 64 16 AB0064 0.12 0.06AB0025 �64 8 AB0065 �64 8AB0026 �64 16 AB0066 64 8AB0027 64 8 AB0067 �64 NDAB0028 64 8 AB0068 0.5 0.5AB0029 64 8 AB0069 2 2AB0030 64 8 AB0070 �64 16AB0031 64 8 AB0071 0.5 0.25AB0032 64 8 AB0072 0.12 0.12AB0033 �64 8 AG0073c �0.06 �0.06AB0034 �64 8 AB0074 0.12 0.25AB0035 64 8 AJ0075d 0.12 0.06AB0036 64 4 A. baumannii 9955e 0.12 0.12AB0037 64 8 A. baumannii 17961e 0.12 0.12AB0038 64 8 E. coli 25922e 0.06 0.06AB0039 64 8 P. aeruginosa 27853e 0.25 1AB0040 �64 8 K. pneumoniae 700603e 0.25 1
a CLSI ciprofloxacin (CIP) MIC criteria: susceptible, �1 mg/liter; intermediate, 2 mg/liter; resistant, �4 mg/liter.b CLSI levofloxacin (LVX) MIC criteria: susceptible, �2 mg/liter; intermediate, 4 mg/liter; resistant, �8 mg/liter.c Acinetobacter genome sp. strain 3.d Acinetobacter johnsonii.e ATCC control.f ND, not determined.
VOL. 47, 2009 QUINOLONE RESISTANCE DETECTION BY PCR/ESI-MS 1439
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 27
Dec
embe
r 20
21 b
y 18
8.21
4.24
3.74
.
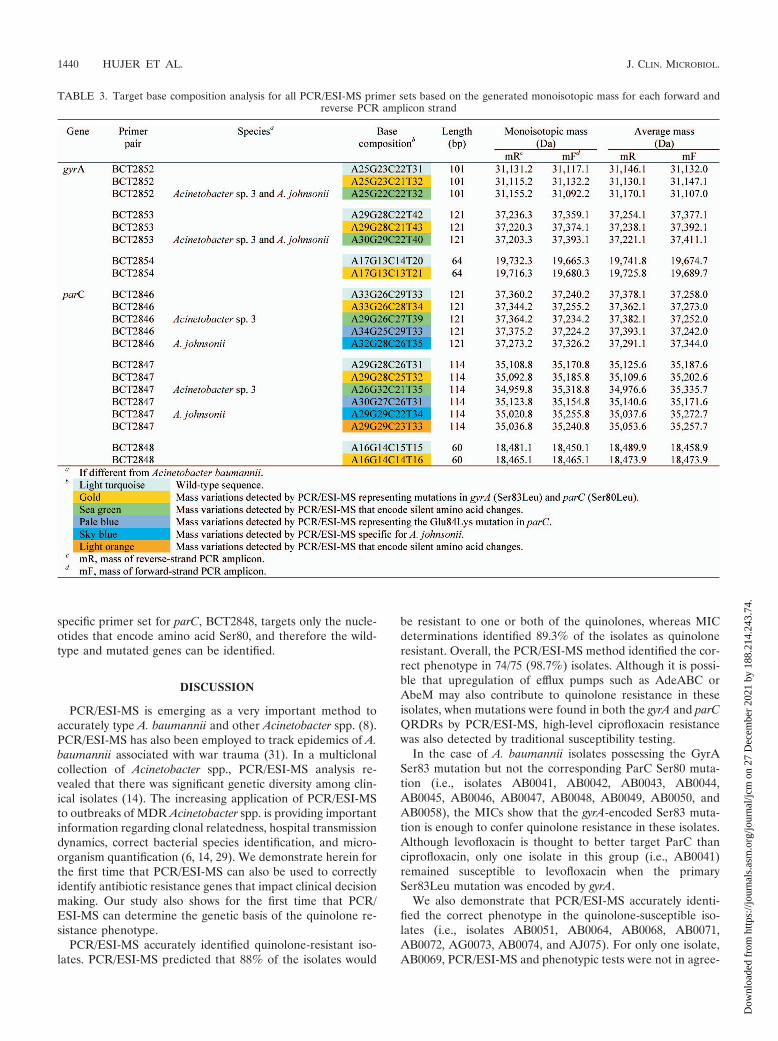
specific primer set for parC, BCT2848, targets only the nucle-otides that encode amino acid Ser80, and therefore the wild-type and mutated genes can be identified.
DISCUSSION
PCR/ESI-MS is emerging as a very important method toaccurately type A. baumannii and other Acinetobacter spp. (8).PCR/ESI-MS has also been employed to track epidemics of A.baumannii associated with war trauma (31). In a multiclonalcollection of Acinetobacter spp., PCR/ESI-MS analysis re-vealed that there was significant genetic diversity among clin-ical isolates (14). The increasing application of PCR/ESI-MSto outbreaks of MDR Acinetobacter spp. is providing importantinformation regarding clonal relatedness, hospital transmissiondynamics, correct bacterial species identification, and micro-organism quantification (6, 14, 29). We demonstrate herein forthe first time that PCR/ESI-MS can also be used to correctlyidentify antibiotic resistance genes that impact clinical decisionmaking. Our study also shows for the first time that PCR/ESI-MS can determine the genetic basis of the quinolone re-sistance phenotype.
PCR/ESI-MS accurately identified quinolone-resistant iso-lates. PCR/ESI-MS predicted that 88% of the isolates would
be resistant to one or both of the quinolones, whereas MICdeterminations identified 89.3% of the isolates as quinoloneresistant. Overall, the PCR/ESI-MS method identified the cor-rect phenotype in 74/75 (98.7%) isolates. Although it is possi-ble that upregulation of efflux pumps such as AdeABC orAbeM may also contribute to quinolone resistance in theseisolates, when mutations were found in both the gyrA and parCQRDRs by PCR/ESI-MS, high-level ciprofloxacin resistancewas also detected by traditional susceptibility testing.
In the case of A. baumannii isolates possessing the GyrASer83 mutation but not the corresponding ParC Ser80 muta-tion (i.e., isolates AB0041, AB0042, AB0043, AB0044,AB0045, AB0046, AB0047, AB0048, AB0049, AB0050, andAB0058), the MICs show that the gyrA-encoded Ser83 muta-tion is enough to confer quinolone resistance in these isolates.Although levofloxacin is thought to better target ParC thanciprofloxacin, only one isolate in this group (i.e., AB0041)remained susceptible to levofloxacin when the primarySer83Leu mutation was encoded by gyrA.
We also demonstrate that PCR/ESI-MS accurately identi-fied the correct phenotype in the quinolone-susceptible iso-lates (i.e., isolates AB0051, AB0064, AB0068, AB0071,AB0072, AG0073, AB0074, and AJ075). For only one isolate,AB0069, PCR/ESI-MS and phenotypic tests were not in agree-
TABLE 3. Target base composition analysis for all PCR/ESI-MS primer sets based on the generated monoisotopic mass for each forward andreverse PCR amplicon strand
1440 HUJER ET AL. J. CLIN. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 27
Dec
embe
r 20
21 b
y 18
8.21
4.24
3.74
.
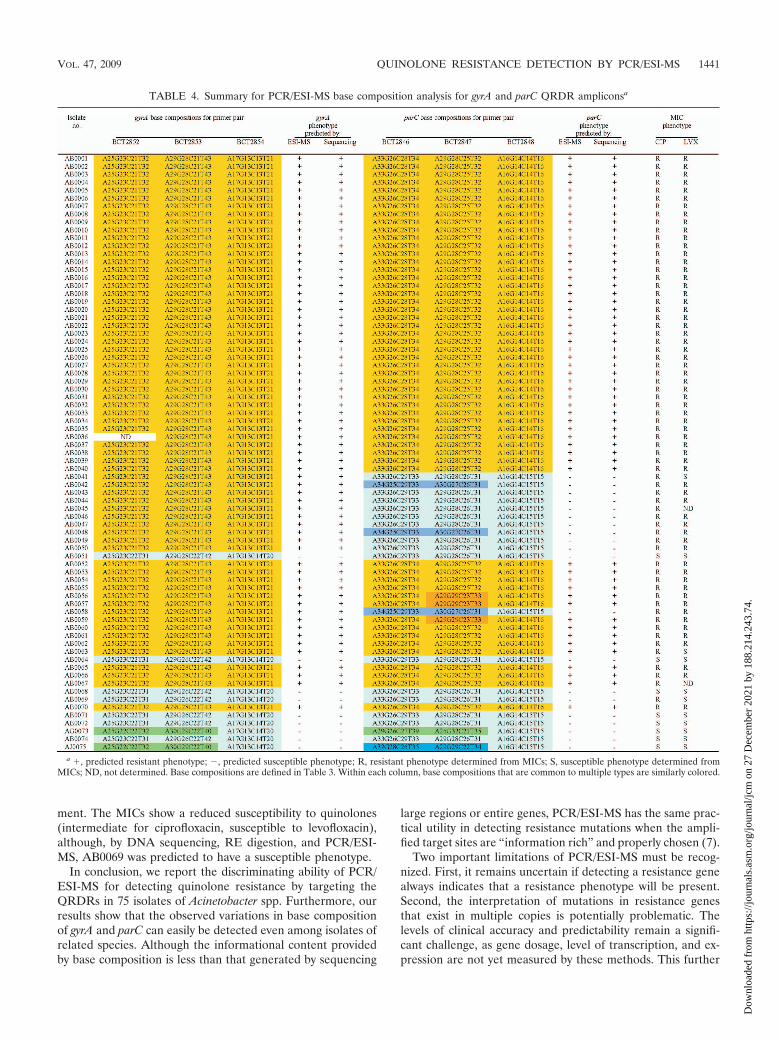
ment. The MICs show a reduced susceptibility to quinolones(intermediate for ciprofloxacin, susceptible to levofloxacin),although, by DNA sequencing, RE digestion, and PCR/ESI-MS, AB0069 was predicted to have a susceptible phenotype.
In conclusion, we report the discriminating ability of PCR/ESI-MS for detecting quinolone resistance by targeting theQRDRs in 75 isolates of Acinetobacter spp. Furthermore, ourresults show that the observed variations in base compositionof gyrA and parC can easily be detected even among isolates ofrelated species. Although the informational content providedby base composition is less than that generated by sequencing
large regions or entire genes, PCR/ESI-MS has the same prac-tical utility in detecting resistance mutations when the ampli-fied target sites are “information rich” and properly chosen (7).
Two important limitations of PCR/ESI-MS must be recog-nized. First, it remains uncertain if detecting a resistance genealways indicates that a resistance phenotype will be present.Second, the interpretation of mutations in resistance genesthat exist in multiple copies is potentially problematic. Thelevels of clinical accuracy and predictability remain a signifi-cant challenge, as gene dosage, level of transcription, and ex-pression are not yet measured by these methods. This further
TABLE 4. Summary for PCR/ESI-MS base composition analysis for gyrA and parC QRDR ampliconsa
a �, predicted resistant phenotype; �, predicted susceptible phenotype; R, resistant phenotype determined from MICs; S, susceptible phenotype determined fromMICs; ND, not determined. Base compositions are defined in Table 3. Within each column, base compositions that are common to multiple types are similarly colored.
VOL. 47, 2009 QUINOLONE RESISTANCE DETECTION BY PCR/ESI-MS 1441
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 27
Dec
embe
r 20
21 b
y 18
8.21
4.24
3.74
.

refinement will require other types of studies. Nevertheless,assessment of quinolone susceptibility by a rapid molecularmethod (in this case, less than 6 h) could have a significantimpact on clinical outcome by aiding in the proper selection ofantibiotics to treat infections.
This series of experiments served as a “proof of principle”for the application of PCR/ESI-MS to detect and evaluateresistance genes not only in A. baumannii but also in othergram-negative pathogens. Using this platform for the rapiddetermination of resistance phenotypes in hospital-associatedinfections, biodefense, and molecular epidemiology can have aprofound impact on clinical microbiology.
ACKNOWLEDGMENTS
The Veterans Affairs Merit Review Program and Geriatric ResearchEducation and Clinical Center VISN 10 supported R.A.B. The Na-tional Institutes of Health (RO1 AI072219) supported both R.A.B.and P.N.R. J.M.T. was supported in part by NIH grant T32 GM07250and the Case Medical Scientist Training Program.
REFERENCES
1. Adams-Haduch, J. M., D. L. Paterson, H. E. Sidjabat, A. W. Pasculle, B. A.Potoski, C. A. Muto, L. H. Harrison, and Y. Doi. 2008. Genetic basis ofmultidrug resistance in Acinetobacter baumannii clinical isolates at a tertiarymedical center in Pennsylvania. Antimicrob. Agents Chemother. 52:3837–3843.
2. Chastre, J. 2003. Infections due to Acinetobacter baumannii in the ICU.Semin. Respir. Crit. Care Med. 24:69–78.
3. CLSI. 2007. Methods for dilution antimicrobial susceptibility tests for bac-teria that grow aerobically. Approved standard M7-A7, 7th ed. CLSI,Wayne, Pa.
4. CLSI. 2007. Performance standards for antimicrobial testing; 17th informa-tional supplement. M100-S17. CLSI, Wayne, PA.
5. Dijkshoorn, L., A. Nemec, and H. Seifert. 2007. An increasing threat inhospitals: multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol.5:939–951.
6. Ecker, D. J., R. Sampath, L. B. Blyn, M. W. Eshoo, C. Ivy, J. A. Ecker, B.Libby, V. Samant, K. A. Sannes-Lowery, R. E. Melton, K. Russell, N. Freed,C. Barrozo, J. Wu, K. Rudnick, A. Desai, E. Moradi, D. J. Knize, D. W.Robbins, J. C. Hannis, P. M. Harrell, C. Massire, T. A. Hall, Y. Jiang, R.Ranken, J. J. Drader, N. White, J. A. McNeil, S. T. Crooke, and S. A.Hofstadler. 2005. Rapid identification and strain-typing of respiratory patho-gens for epidemic surveillance. Proc. Natl. Acad. Sci. USA 102:8012–8017.
7. Ecker, D. J., R. Sampath, C. Massire, L. B. Blyn, T. A. Hall, M. W. Eshoo,and S. A. Hofstadler. 2008. Ibis T5000: a universal biosensor approach formicrobiology. Nat. Rev. Microbiol. 6:553–558.
8. Ecker, J. A., C. Massire, T. A. Hall, R. Ranken, T. T. Pennella, C. AgasinoIvy, L. B. Blyn, S. A. Hofstadler, T. P. Endy, P. T. Scott, L. Lindler, T.Hamilton, C. Gaddy, K. Snow, M. Pe, J. Fishbain, D. Craft, G. Deye, S.Riddell, E. Milstrey, B. Petruccelli, S. Brisse, V. Harpin, A. Schink, D. J.Ecker, R. Sampath, and M. W. Eshoo. 2006. Identification of Acinetobacterspecies and genotyping of Acinetobacter baumannii by multilocus PCR andmass spectrometry. J. Clin. Microbiol. 44:2921–2932.
9. Falagas, M. E., S. K. Kasiakou, P. I. Rafailidis, G. Zouglakis, and P. Morfou.2006. Comparison of mortality of patients with Acinetobacter baumanniibacteraemia receiving appropriate and inappropriate empirical therapy. J.Antimicrob. Chemother. 57:1251–1254.
10. Gootz, T. D., and A. Marra. 2008. Acinetobacter baumannii: an emergingmultidrug-resistant threat. Expert Rev. Anti Infect. Ther. 6:309–325.
11. Hamouda, A., and S. G. Amyes. 2004. Novel gyrA and parC point mutationsin two strains of Acinetobacter baumannii resistant to ciprofloxacin. J. Anti-microb. Chemother. 54:695–696.
12. Higgins, P. G., H. Wisplinghoff, D. Stefanik, and H. Seifert. 2004. Selectionof topoisomerase mutations and overexpression of adeB mRNA transcriptsduring an outbreak of Acinetobacter baumannii. J. Antimicrob. Chemother.54:821–823.
13. Hofstadler, S. A., R. Sampath, L. B. Blyn, M. W. Eshoo, T. A. Hall, Y. Jiang,J. J. Drader, J. C. Hannis, K. A. Sannes-Lowery, L. L. Cummins, B. Libby,D. J. Walcott, A. Schink, C. Massire, R. Ranken, J. Gutierrez, S. Manalili,C. Ivy, R. Melton, H. Levene, G. Barrett-Wilt, F. Li, V. Zapp, N. White, V.Samant, J. A. McNeil, D. Knize, D. Robbins, K. Rudnick, A. Desai, E.Moradi, and D. J. Ecker. 2005. TIGER: the universal biosensor. Int. J. MassSpectrom. 242:23–41.
14. Hujer, K. M., A. M. Hujer, E. A. Hulten, S. Bajaksouzian, J. M. Adams, C. J.Donskey, D. J. Ecker, C. Massire, M. W. Eshoo, R. Sampath, J. M. Thomson,P. N. Rather, D. W. Craft, J. T. Fishbain, A. J. Ewell, M. R. Jacobs, D. L.Paterson, and R. A. Bonomo. 2006. Analysis of antibiotic resistance genes inmultidrug-resistant Acinetobacter sp. isolates from military and civilian pa-tients treated at the Walter Reed Army Medical Center. Antimicrob. AgentsChemother. 50:4114–4123.
15. Jiang, Y., and S. A. Hofstadler. 2003. A highly efficient and automatedmethod of purifying and desalting PCR products for analysis by electrosprayionization mass spectrometry. Anal. Biochem. 316:50–57.
16. Joly-Guillou, M. L. 2005. Clinical impact and pathogenicity of Acinetobacter.Clin. Microbiol. Infect. 11:868–873.
17. Lee, J. K., Y. S. Lee, Y. K. Park, and B. S. Kim. 2005. Mutations in the gyrAand parC genes in ciprofloxacin-resistant clinical isolates of Acinetobacterbaumannii in Korea. Microbiol. Immunol. 49:647–653.
18. Muddiman, D. C., G. A. Anderson, S. A. Hofstadler, and R. D. Smith. 1997.Length and base composition of PCR-amplified nucleic acids using massmeasurements from electrospray ionization mass spectrometry. Anal. Chem.69:1543–1549.
19. Munoz-Price, L. S., and R. A. Weinstein. 2008. Acinetobacter infection.N. Engl. J. Med. 358:1271–1281.
20. Murray, C. K., and D. R. Hospenthal. 2005. Treatment of multidrug resistantAcinetobacter. Curr. Opin. Infect. Dis. 18:502–506.
21. Peleg, A. Y., H. Seifert, and D. L. Paterson. 2008. Acinetobacter baumannii:emergence of a successful pathogen. Clin. Microbiol. Rev. 21:538–582.
22. Peralta, G., M. B. Sanchez, J. C. Garrido, I. De Benito, M. E. Cano, L.Martinez-Martinez, and M. P. Roiz. 2007. Impact of antibiotic resistanceand of adequate empirical antibiotic treatment in the prognosis of patientswith Escherichia coli bacteraemia. J. Antimicrob. Chemother. 60:855–863.
23. Perez, F., A. M. Hujer, K. M. Hujer, B. K. Decker, P. N. Rather, and R. A.Bonomo. 2007. Global challenge of multidrug-resistant Acinetobacter bau-mannii. Antimicrob. Agents Chemother. 51:3471–3484.
24. Scott, P., G. Deye, A. Srinivasan, C. Murray, K. Moran, E. Hulten, J.Fishbain, D. Craft, S. Riddell, L. Lindler, J. Mancuso, E. Milstrey, C. T.Bautista, J. Patel, A. Ewell, T. Hamilton, C. Gaddy, M. Tenney, G. Chris-topher, K. Petersen, T. Endy, and B. Petruccelli. 2007. An outbreak ofmultidrug-resistant Acinetobacter baumannii-calcoaceticus complex infectionin the US military health care system associated with military operations inIraq. Clin. Infect. Dis. 44:1577–1584.
25. Sebeny, P. J., M. S. Riddle, and K. Petersen. 2008. Acinetobacter baumanniiskin and soft-tissue infection associated with war trauma. Clin. Infect. Dis.47:444–449.
26. Turton, J. F., M. E. Kaufmann, M. J. Gill, R. Pike, P. T. Scott, J. Fishbain,D. Craft, G. Deye, S. Riddell, L. E. Lindler, and T. L. Pitt. 2006. Comparisonof Acinetobacter baumannii isolates from the United Kingdom and theUnited States that were associated with repatriated casualties of the Iraqconflict. J. Clin. Microbiol. 44:2630–2634.
27. Vila, J., J. Ruiz, P. Goni, and T. Jimenez de Anta. 1997. Quinolone-resis-tance mutations in the topoisomerase IV parC gene of Acinetobacter bau-mannii. J. Antimicrob. Chemother. 39:757–762.
28. Vila, J., J. Ruiz, P. Goni, A. Marcos, and T. Jimenez de Anta. 1995. Mutationin the gyrA gene of quinolone-resistant clinical isolates of Acinetobacterbaumannii. Antimicrob. Agents Chemother. 39:1201–1203.
29. Whitman, T. J., S. S. Qasba, J. G. Timpone, B. S. Babel, M. R. Kasper, J. F.English, J. W. Sanders, K. M. Hujer, A. M. Hujer, A. Endimiani, M. W.Eshoo, and R. A. Bonomo. 2008. Occupational transmission of Acinetobacterbaumannii from a United States serviceman wounded in Iraq to a health careworker. Clin. Infect. Dis. 47:439–443.
30. Wisplinghoff, H., M. Decker, C. Haefs, O. Krut, G. Plum, and H. Seifert.2003. Mutations in gyrA and parC associated with resistance to fluoroquino-lones in epidemiologically defined clinical strains of Acinetobacter bauman-nii. J. Antimicrob. Chemother. 51:177–180.
31. Wortmann, G., A. Weintrob, M. Barber, P. Scott, S. T. Zoll, M. W. Eshoo, R.Sampath, D. J. Ecker, and C. Massire. 2008. Genotypic evolution of Acin-etobacter baumannii strains in an outbreak associated with war trauma.Infect. Control Hosp. Epidemiol. 29:553–555.
1442 HUJER ET AL. J. CLIN. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 27
Dec
embe
r 20
21 b
y 18
8.21
4.24
3.74
.