rajesh kumar n - shodhgangashodhganga.inflibnet.ac.in/bitstream/10603/6196/10/10_chapter 5.pdf ·...
TRANSCRIPT

Chapter V
Antiturnour and Anticarcinogenic activity of
Phyllanthus amarus and Mechanism of action

Antiturnour and anticarcinogenic activity of Phyllanthus amarus and i t . mechanism of action
1. Introduction
Hepatocellular carcinoma (HCC) is one of the most common and
devastating malignant tumours in some parts of the world, particularly in Far East and
Africa (Simonetti et al., 1991). HCC accounts for 90 % of all primary hepatic
malignancies (Okuda, 1992) and causes over 5 % of cancer deaths throughout the
world (WHO, 1997). The striking geographic differences in the incidence of HCC suggest
that environmental factors frequently contribute to its development. (Thung and Gerber,
1997). Its epidemologic frequency and pathogenesis are due to chronic viral infection with
hepatitis B and hepatitis C, consumption of aflatoxin contaminated foods, alcohol abuse
(Di Bisceghe et al., 1988), industrial chemicals, air and water pollutants (Peers and Linsell,
1973). Nitrosodiethylamine is a potent carcinogenic dialkyl nitrosamine present in tobacco
smoke and its carcinogenicity has already been demonsterated in several animal species.
Nitroso compounds has been associated with an increased risk of cancer development in
various organs (Pobel et al., 1995; Bartsch et a]., 1990). The presence of NDEA has been
detected in edible vegetable oils, in alcoholic drinks, steamed and fried fish and could be
formed endogenously in the stomach (Hedler et a]., 1979). The limited treatment options
and poor treatment success makes HCC a leading cause of death in developing countries.
Hence, a major effort should be directed towards its prevention.
Phy1lanrhu.s amarus (Plate 1 a) is widely used as an anti-viral agent (Thygarajan et
al., 1988; De et al., 1990; Jayram et al., 1997). P. amarus extract has been shown to
inhibit DNA polymerase of hepatitis B virus and related hepatitis viruses (Venkateswaran
et al., 1987; Blumberg et al., 1990) and down regulate hepatitis B virus mRNA
transcription and translation (Lee et al., 1996; Ott et al., 1997).The extract reversibly
inhibited cellular proliferation and suppressed HbsAg production in cultured hepatoma cell
line HepA2 (Yeh et al., 1993) and inhibited the release of HbsAg in Alexander cell line, a
human hepatocellular carcinoma derived cell line (Jayram and Thyagarrajan, 1996).
The present chapter deals with (a) the anti-tumour activity of P. amarus extract
against various transplantable tumours in mice. (b) the anti-carcinogenic potential of P.

amarus extract against chemical-induced liver tumours in rats and (c) the mechanism of
action of P. amarus extract
2. Materials and methods
2.1 Determination of the effect P. amarus on ascites tumours
Tumour cells (DLA I EAC) aspirated from the peritoneal cavity of mice were
washed with saline and 1 x 10 tumour cells were given intrapentoneally to four group of
animals (Male Swiss albino mice,lO-weeks-old, 20-25 g, 7 micelgroup). Animals in group
11-IV received P. amarus extract at a concentration of 60, 300, 1500 mgkg body weight
(p.o), respectively, 24 h after tumour inoculation and continued daily for 10 days. Animals
in the group I was kept as control mice which received the vehicle. Animals were observed
for the development of ascites tumour and deaths due to tumour burden were recorded. The
increase in life span (percent ILS) of treated group was calculated (Soudhamini and Kuttan,
1988).
2.2 Determination of the effect P. amarus on solid tumours
One million DLA 1 EAC cells were injected in to the right hind limb of male Swiss
albino mice (Group 1 - IV, 7 micelgroup). Animals in group 11-IV received P. amarus
extract at a concentration of 60, 300, 1500 mgtkg body weight (p.o), respectively, 24 h
after tumour inoculation and continued for 10 days. Animals in the group I was kept as
control which received only the vehicle. Diameter of the tumour was measured on every
fifth day using vernier calipers and volume was calculated (Kuttan et al., 1988).
2.3 Determination of the effect of P. amarus extract administration during induction
of hepatocarcinogenesis by NDEA
The study was performed on 8-10-week-old male Wistar rats weighing 120-140 g.
Rats were randomly divided into four groups (n=10 each). Animals in the group I were
kept as normal animals receiving vehicle alone. Animals in the group I1 to IV were
administered with 0.02 % NDEA, 2.5 d r a t , 5 days weekly for 20 weeks. Rats in the
group 111 to IV were administered with P. amarus extract 150 and 750 mg/ kg respectively
immediately after NDEA administration and continued for 20 weeks. After that animals
were kept without any drug treatment for one week and sacrificed by diethyl ether
anesthesia. Blood was drawn by cardiac puncture and serum was separated. Liver was
surgically excised, weighed and homogenate prepared was used for biochemical

estimations. A small piece of the liver was fixed in 10 % formalin. The formalin fixed
specimens were embedded in paraffin and sectioned (3-5~m), sections kern each group
were stained with haematoxylin and eosin, and histological sections were evaluated by
light microscopy.
Protein in liver homogenate was assayed by the method of Lowry et a1 (1951).
Serum y-glutamyl transpeptidase activity was assayed by using glutamyl para-nitroanilide
as the substrate (Szaz, 1976). Liver y-glutamyl transpeptidase activity was assayed by the
method described by Tate and Meister (1974). Cytosolic glutathione S-transferase activity
was determined by the method described by Haibig et a1 (1974). Tissue glutathione was
determined by dithio-his 2-nitrobenzoicacid method described by Moron et al (1979).
Alkaline phosphatase (ALP) was assayed by the method described by King and Armstrong
(1980). Glutamate pyruvate transaminase (GPT) was assayed by the method of Bergmeyer
and Bernt (1980). Bilirubin level in the serum was assayed by the method described by
Jendrassik and Jrof (1938). Serum lipid peroxidation was assayed by the method described
by Ohkawa et al (1979). Detailed procedures are given chapter 11.
2.4 Determination of the effect of P. amarus extract administration after induction of
HCC by NDEA
In another set of experiment, male Wistar rats (100-125 g) were divided into five
groups (7 ratslgroup). HCC was induced in animals in group 11-V by oral administration of
NDEA, 0.02%. 2.5 mllrat, 5 days weekly for 20 weeks (Narurker and Narurker, 1989).
Animals in group I were kept as normal animals receiving vehicle (distilled water) alone
and were sacrificed 20 weeks after the onset of the experiment. After 20 weeks animals in
group 11 were sacrificed and changes in liver weight and biochemical parameters were
recorded. Rats in group 111 were kept without any further treatment, sacrificed at the end of
the 291h week and changes in liver weight and biochemical parameters were noted.
Animals in group IV were observed for their survival time. Animals in group V received P.
amarus extract 150 mg /kg body weight orally, 5 days weekly, continuously for 54 weeks
or till they died. Surviving animals in group V were sacrificed at the end of the 54" week
and changes in liver weight and biochemical parameters were recorded.
Formalin fixed liver from each group was embedded in paraffin and sectioned (5
pm). Sections were dewaxed in xylene, rehydrated through graded alcohol, and stained

with hematoxylin-eosin (H and E) for routine histopathology. Argyrophilic Nuclear
Organizer Region (AgNOR) staining was carried out by the method of Murray et al(1989)
with modifications as described by Lahshmi et al(1993
The values were expressed as means r standard deviations. The results were
analyzed statistically by use of Student's t-test. Values less than 5 % @ <0.05) were
considered to be indicative of statistical significance.
2.5 Aniline hydroxylase inhibitory assay
Aniline hydroxylase assay was performed by the method described by Maze1
(1971). The enzyme was induced in rats by the oral administration of phenobarbital (80
mgkg) for 5 days. A 10 % liver hornogenate prepared in 10 mM ice cold tris-HC1 buffer
(pH 7.4) was used for the assay. P-aminophenol formed during the enzyme action reacts
with phenol in alkaline medium to form a blue coloured product, which was measured at
630 nm. The percentage inhibition of aniline hydroxylase was calculated by comparing the
absorbency of control and that of drug treated samples.
2.6 Topoisomerase I and I1 inhibitory assays
Saccharomyces cervisiae mutant cell cultures JN 394, JN 394 and JN 394 1.2.5
used for topoisomerase assays. The cells from a fully grown plate of each organisms were
suspended in saline solution (10 rnl) and then diluted to obtain 5 X 10 6 ~ ~ ~ / m ~ . 50 pl of
this suspension was then used to inoculate petridishes containing YPDA media. The
aqueous extract of P. amarus was dissolved in DMSO and added to the inoculated plates
(20 pl) to give a final concentration of 250 pgl ml. These plates were inoculated at 27OC
for 72-96 h. At the end of incubation period the zones of inhibition were recorded for each
test organism. Controls were prepared by adding DMSO (20 pl) to inoculated plate (Chang
eta]., 1995; Roth et a]., 1998)
2.7 Determination of cdc25 tyrosine phosphatase and cdc2 kinase inhibitory assays
20 p1 of GST-cdc25A protein was mixed with 20 p1 100 mM dithiothreitol in Tris
buffer. Different concentrations of P. amarus extract were dissolved in Tris buffer (140
pl), in 96-we11 microtitration plates. The assay were initiated by addition of 20 pl of 500
mM p-nitrophenylphosphate phosphatase @-NPP). After 60 ;min incubation at 37' C
absorbance at 405 nm was measured in a BioRad microplate reader (Baratte et al., 1992).

Cdc2 kinase inhibitory assay was conducted by immunoblotting technique using anti-
antibodies (Rialt and Meijer, 1991).
3. Results 3.1 Effect of P. amarus on ascites tumours
P. amarus extract administration increased the life span of ascites tumour harboring
mice. All the untreated mice in the DLA tumour group died of tumour burden 21.00 + 1.91
days after tumour inoculation. Oral administration of P. amarus (60, 300 and 1500 mglkg)
increased the survival of animals to 23.00 k 2.2, 25.33 5 3.19 and 29.33 + 4.20 days and
the percentage increase in survival was found to be 8.69, 20.61 and 39.66 %, respectively
(Table V.l). Survival of animals inoculated with EAC was also found to be increased by
16.70, 19.66 and 29.45 % at concentrations given above (Table V.l).
3.2 Effect P. amarus on solid turnours
Administration of P. amarus extract reduced the tumour volume of both DLA and
EAC cell lines in a dose dependent manner. The tumour volume of animals without P.
amarus treatment on 301h day after tumour (DLA) inoculation was found to be 4.4 cc. The
tumour volume was reduced to 4.0, 3.3, and 2.4 cc by P. amarus extract administration at
concentrations of 60, 300 and 1500 mglkg body weight, respectively (Fig V.l). Similarly,
the tumour volume ot EAC inoculated animals on 3oLh day after tumour inoculation was
found to be 6.4 cc which was reduced to 6.1, 4.7 and 3.6 cc and the percent reduction in
tumour volume was found to be 4.9, 26.7 and 42.8 % in the group of animals treated with
P. amarus (Fig V. 1 ).
3.4 Effect of simultaneous administration of P. amarus on NDEA induced
hepatocarcinogenesis
All animals in the carcinogen-administered group developed liver tumours (Plate 7
a) by the end of 201h week. P. amarus administration was found to inhibit the tumour
development in liver tnduced by NDEA. At 150 mgkg dose, 40 % of animals developed
tumours while at 750 mgikg only 10 % of rats developed visible liver tumours (Table V.2).
The size of the liver and number of tumour nodules were reduced by P. amarus treatment
(Plate 7 e and g) V. I ) . Liver weight of NDEA treated animals were raised as compared to
normal rats. P. urnurus administration significantly lowered the liver weight (Table V.2).
Increased y-glutamyl transpeptidase, a marker of hepatocellular carcinoma (Hanigan and

Table V.1. Effect of P. amarus extract administration on the survival of ascites tumour harboring mice inoculated with DLA/EAC cell lines
b = p < 0.05, c = p< 0.01, d = p < 0.02 as compared to group I. Values are mean * SD, n = 7.
Group
I
I1
ID
IV
Ehrlich tumour Animal status
Without drug
P. amarus 60 mgtkg
P.amurus300mg/kg
P. amarus 1500 mgkg
Average life span (days)
19.28 4 2.86
22.50 k 4.27
24.00 i 5.76
27.33 i 4.64
DLA tumour
% increase in life span
0
16.70
19.66
29.45
Average life span (days)
21.00* 1.91
23.00 i 2.27
25 .33&3.19~
29 33 * 4.26 b
% increase in Life span
0
8.69
20.61
39.66

Fig. V.1. The effect of P, amarus extract administration on solid tumour volumes induced by DWEAC cell line in mice.
DLA tumour
Days post tumour inoculation
10 - EAC tumour 9 - 8 -
8 7 - - E 6 - 3 - 5 - L 4 - a
3 - z 2 -
1 - n,
Days post tumour inoculation
- Tumour cells alone; A - P. amarus 60 mglkg; - P. amarus 300rnglkg; 0 - P. amarus 1500 mglkg

Pitot, 1985). in serum as well as in liver was found to be 'effectively lowered by the
administration of P. amarus, indicating that P. amarus could reduce the proliferation of
tumour cells (Table V.2). Similarly, GSH and GST values which were increased after
NDEA treatment was found to be lowered by the administration of P. amarus (Table V.2
and V.3). ALP and GPT activity in the serum of NDEA administered group were raised as
compared to that of normal value. P. amarus administration significantly inhibited the rise
of ALP and GPT (Table V.3). Similarly, elevated levels of serum lipid peroxidation (LPO)
and bilirubin of NDEA treated group was also found to be lowered by the administration of
P. amarus (Table V . 3 ) .
Histopathological analysis of NDEA treated rat liver revealed that of well
differentiated hepatocellular carcinoma. The normal architecture of the liver was distorted
and the neoplastic cells often merge imperceptibly with liver cell plates. Nuclei were
enlarged and irregular in shape (Plate 7 b). Degenerating hepatocytes and necrotic areas
were detected in most of the areas. P. amarus treated rat liver (150 mgtkg) showed
neoplastic transformation but normal hepatocytes were detected in some foci (Plate 7 f).
Hepatocytes remained normal in the 750 mgtkg treated group (Plate 7 h) except for the
presence of a few alterations such as hepatocytomegalia and karyomegalia in certain areas.
3.5 The effect of P. amarus extract administration after induction of HCC by NDEA
Continued administration of P. amarus significantly increased the survival of
tumour bearing animals. All the untreated rats (Group IV) died of tumour burden at 33.7 +.
1.6 weeks, while four animals in the P. amarus administered group survived more than 54
weeks and average life span was 52.2 weeks (Table V.4). The percentage increase in
survival was found to be 54.78 % which is highly significant when compared to animals in
group 1V. Serum y-GT value of NDEA treated animals was found to be elevated to 152 ?
23 and 182 + 21 UII by 20 and 29 weeks, respectively as compared to the normal 1-GT
value 44.4 rt 5.8 Ulml. Further treatment with P. amarus extract effectively lowered the
elevated y-GT to 1 12 k 19 U/I, indicating that P. amarus could reduce the proliferation of
tumour cells (Table V.4). Similarly, GSH and GST values which were increased to 20.5 + 2.4 nmoVmg protein and 1534 -t 116 nmoVmin/mg protein respectively after NDEA
treatment were found to be lowered to 14.2 rt 2.2 nmoWmg protein and 1112 rt 89
nmol/min/mg protein respectively after continued administration of P. amarus (Table

Plate 7
Gross morphology and histogathology of rat livers
(1) I\jDEA admir~ist~red rat liver at 20 weeks shows proliferrtion of tumour
norides
(0) Histopathology of NDEA administered rat liver at 20 weeks showing
distorted arrarrgement of liver cells with anisonuciensis and prominent
nuclec~lus ( W & E 40 X).
( c ) YDEA ad;r:i:listi:reci rat i i ~ e r at 29 weeks showing tuinogr rlodulez and
blcc,.ti filied areas
jd! r-iisto~ogy of NDEA administered rats at 29 weeks showing
Iymphophagocytosis and hepatocytes with prominent chroma.tin dots (II & E
41) X).
(e) Liver of rat treated with P;zyllanthus amarus 150 r@kg shaws reduced
liver size
(f) Histology of rat liver treated with Phyllanthus amarzs i50 mgkg
showing minimal anisonucieosis ( H & E 40 X).
(g) Liver of rats treated with Phyllunthus umarus 750 m a g showing reduced
size.
(h) Liver section of ra: tredteil with P!zyllanthus ilmarus 750 mgkg showing
hepatocytes of normai mxpholcg): (H & 30 X).
(i) &NOR stained liver sections 01 NDEA admir~istered rat (29 weeks
showing increased number AgNO? dots (1;30 X).
6 ) AgNOR stained liver sections cf :lit; treatci; wit!: 7Aylla.~.hu,r marus 150
mgkg (at 54 v.-cks) stlowing d?crc.ised number uf AgNOR c;ots ( 1 0 X ) .
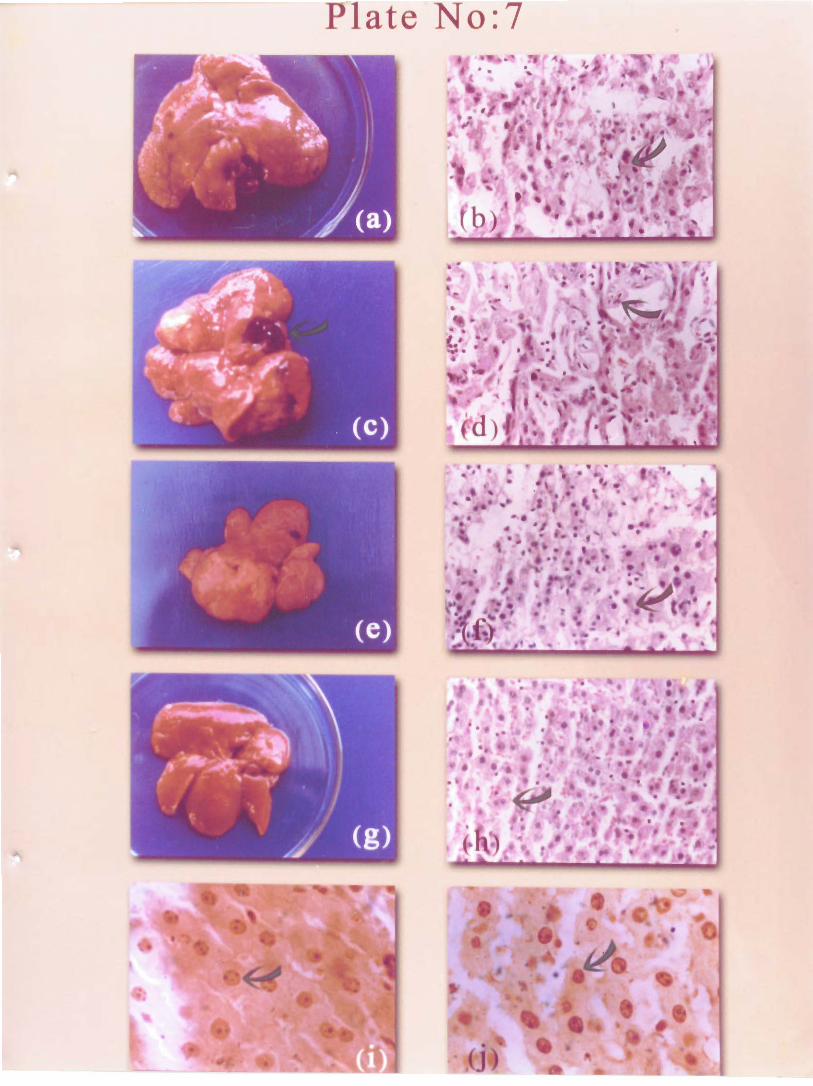

Table V.2. Effect of P. amarus extract treatment along with NDEA on tumour incidence, liver weight and y-glutamyl transpeptidase, hepatic GSH of rats.
a = p < 0.001, b = p < 0.005, c = p < 0.01 as compared to group 11. Values are mean* SD; n = 6 in group I - I1 and 10 in group 111 - IV
I
II
III
IV
Table V.3. Effect ofP. amarus extract treatment along with NDEA on hepatic, GST, serum ALP, GPT, LPO and bilirubin levels of rats.
a = p < 0.00 1, b = p < 0.005, d = p < 0.02 as compared to group 11. Values are mean i SD, n = 6 in group I - I1 and 10 in group 111 - IV.
GroupAnimaI status
Normal rats
NDEA alone
NDEA+P amarus
150 mgi kg NDEA+P
amarus 750 mg/ kg
Group
I
II
111
IV
GSH (nmoVmg proteim)
8.4 i 0.6
16.2 i 3.2
12.9 5 2.8
10.6 i 2.1
Tumour incidence
016
6/6
4/10
1'10
y -glutamyl transpeptidase activity
Animal status
Normal rats
NDEA alone
NDEA+P. amurus
150 mgi' kg NDEAiP,
arnaru.~ 750 mg: kg
Liver weight 100 g b.w
2.86 i 0.21
4.57 * 0.88
3.97 i 0.68
3.22 i 0.66
Serum Ufl at 30 " C)
35.3 i 4.8
151.5 i 21.4
98.8i16.1
68.8 i 14.2 a
Liver (nmoUminl mg protein)
0.09 i 0.01
1.50 * 0.41
*0,22 b
0.33 i 0 .26 a
GST (nmoVminImg
protein)
748 i 69
1738 i 222
1282i 171
1 0 2 0 i 1 0 2 a
GPT
(ulml)
225 i 17
874i 67
6 4 0 i 4 4 a
520 * 38 a
ALP (KAUIdI)
18.2 i 2.8
46.9 i 6.4
38.9i5 .9
2 8 8 t 4 . 4 a
LPO (nmoUml)
1.69 i 0.31
3.26 i 0.82
2.31 0.52 d
2.12 i 0.32
Total b i i b i n (mg/dl)
0.38 i0 .08
1.40 i 0.31
0.81 0.14b
0.66 i 0.22 a

V.5). Administration of P. amarus extract after induction of tumours was found ineffective
in controlling the rlse of liver weight. The liver weight of NDEA treated animals at the 2 0 ~
week was found to be 3.97 * 0.37 g1100 g b.w. The liver weight was found to be increased
to 5.57 0.99 g I100 g b. w. by the 29" week. P. amarus administration was also found to
be ineffective in controlling the increase in liver weight, tissue y-GT, serum ALP and
serum GPT (Table V.4 and V.5).
Grossly visible turnour nodules appeared in the liver by 20 weeks of NDEA
administration (Plate 7 a) and all lobes were perforated with nodules by 29 weeks (Plate 7
c). The shapes of the tumours were very variable. They were often multilobed and showed
irregular protuberances at their surfaces. H and E staining of NDEA administered rat liver
at 29 weeks showed well differentiated HCC (Plate 7 d). The section also revealed
coalescent nodular areas, which distort or replace the normal hepatic structure.
Proliferating bile ducts and hepatic cells compose almost the entire sections. The vessels
were enlarged and the~r walls became attenuated and distorted with indistinct outlines.
Degenerating hepatocytes with necrotic patches were detected in some areas. Sinusoides
were dilated in some portlons to form blood lakes. P. amarus treatment minimized the
liver damage and islands of normal liver cells were present in some portions. However,
inflammation and localized neutrophil infiltration were noticed in most of the areas.
AgNOR's dot and cluster of NDEA administered rats (Plate 7 i) were increased as
compared to normal rats. AgNOR dots and clusters of normal rats were found to be 1.68
and 0.52, which were ra~scd to 3.86 and 2.12, respectively by NDEA administration. P.
amarus treatment 150 rnglkg reduced the dot (Plate 7 j) value to 2.06 and cluster value to
1.04, respectively.
3.6 Effect of P. amarus on aniline hydroxylase
P. atnarus extract showed a dose dependent inhibition of aniline hydroxylase.
Concentration requlred fur 50 % inhibition of aniline hydroxylase was found to be 540
pglml (Fig V.2).
3.7 Effect of P. amarus on Topoisomerase I and I1
The aqueous extract of P. amarus demonstrated topoisomerase I1 inhibitory activity
when tested on Sarchromyc,~s cerevisiae mutant strains at 250 pglml. JN 394 is hyper
sensitive to topoisomerase I poisons, while JN 394.1 lack top 1 gene and therefore shows

Table V.5. Effect ofP. amarus extract administration on hepatic GSH, GST, serum ALP and GPT levels of HCC harboring rats.
Table V.4. Effect of P. amarus extract treatment on Liver weight, y-Glutamyl transpeptidase and survival of HCC harboring rats
Group
I
11
LU
N
V
a = p < 0.00 I , b p r 0.005 VS group 111. Values are mean i SD, n = 7 except group V (n = 4)
Group
I
II
IIl
N
V
a = p < 0.00 1 VS group I11 (survival compared to group IV). Values are mean i SD, n = 7 except group V (n - 4)
Animal status
Normal rats
NDLA alone (at 20"' week)
NDEA (at 29"" week)
NDE.4 (sunial) NDEA t
P ur11wu.s
(at 54Ih wcck)
Animal status
Nonnal rats
NDEA (at 20''' \vcek)
KD1:4 (at 29'" \\ eek)
NDEA (survival) KDl;.A +
I>. ~lrIlul~l(.\
Liver weight 100 g b.w
2.5 1 i 0.18
3.97 * 0.87
5.57 i 0.99
-
10.29 i 2.8
Survival in weeks
33.7 4 1.7
52.2 i 2.3 a
y-glutamyl transpeptidase activity
GSH (nmoV mg protein)
7.7 * 0.8
20.5 * 2 4
18.2 * 1.8
14.2 i- 2.2 b
Serum (Un at 30" C)
44.4 * 5.8
152 i 23
182 i 21
-
1 1 2 i 1 9 ~
Liver (nmoVmin/mgprotein)
0.09 rt 0.01
0.73k0.11
1.50 i 0.21
-
1.66 i 0.72
GST (nmoYmin/mgprotein)
733 i 49
1534h 116
1738 i 56
1 1 1 2 i 9 8 ~
ALP m U / d l )
1 7 . 4 i 1.8
54.9 i 2.7
46.9 i 6.1
64.8 i11.4
GPT (V/ml)
244 i 17
820 i 38
848 i 67
904 i 107
-

Fig V.2. Aniline hydroxylase inhibitory activity of P. amarus extract
0 0 100 200 300 400 500 600
Concentration of P. amams extract (vglrnl)

a lack of response to topoisomerase 1 poisons. JN 394 ,.2.5 carry top I1 gene, which is
resistant to topoisomerase I1 poisons but responds to topoisomerase I, poisons. The zones
of inhibition indicated that the extract inhibited topoisomerase I1 activity. However, the
concentration used in this study (250 pg/ml) was found to be ineffective in inhibiting
topoisomerase I activity.
3.7 The effect of P. amarus on cdc25 tyrosine phosphatase and cdc2 kinase
P. amarus extract was found to inhibit the activity of cdc25 tyrosine phosphatase.
Concentration required for 50 % inhibition of cdc25 tyrosine phosphatase activity was
found to be 25 pglml. P. amarus extract could inhibit the activity of cdc2 kinase only at
high concentration (IC so > 1000 pg/ ml).
4. Discussion
Primary liver cancer ranks fifth in frequency among all malignancies in the world
with an estimated number of 4,37000 new cases in 1990 (Bosch et al., 1999). The vast
majority of primary liver cancer is hepatocellular carcinoma. HCC is increasing throughout
the world, particularly in areas where hepatitis C virus infection is more common (Deuffic
et al., 1998). Over the past two decades, the incidence of hepatocellular carcinoma has
risen in the USA and younger Americans are developing the disease (Ei-Serag and Manson
1999). Hepatocellular carcinoma has become the most common malignancy associated
death in Asia (Hasegawa et al., 1999). As HCC typically develops in patients with
advanced chronic liver disease, radical surgery is often impractical (Okuda et al., 1987).
Inoperable HCC is an incurable malignancy with no accepted therapy (Stuart et al., 1999).
The prognosis of HCC is extremely poor, even with the remarkable progress in medical
science. The overall 5-yr survival rate worldwide is only 2 % because of the late diagnosis
(Kew., 1998). A complete cure is only possible with liver transplantation for an early HCC
without extrahepatic spread (Mazzaferro et al., 1996). This ideal therapy is rarely achieved
because of the very limited availability of donar liver. Local ablation therapy first with
ethanol (Livraghi et al., 1995), followed by microwave coagulation (Sao et al., 1996) and
radiofrequency ablation (Goldberg et al., 1998), prolongs S U N ~ V ~ somewhat, but
frequently recurrence occurs in the liver, the majority of the patients eventually succumb.
Liver-directed gene therapy is on the juvenile stage and the development is (Okuda., 2000;

Ghosh et al., 2000). Because of these dismal experiences in treatment, more serious
thought should be given to the prevention of HCC.
The present study shows that P. amarus extract treatment could inhibit the
hepatocarcinogenesis induced by NDEA and could significantly increase the life span of
HCC harboring animals. There was also a decrease in serum y-glutamyl transpeptidase
activity indicating decreased proliferation of turnour cells. Tissue GST and tissue GSH also
showed a similar decrease. Increase in liver weight may be due to the oedema developed in
the liver, while the reason for unaltered ALP and GPT could be due to the irreversible
unrepairable damage to the liver induced by the carcinogen administration.
P. amarus extract possesses a broad spectrum of biological activities. Other than its
activity as an anti-viral agent, the extract and ingredients isolated from the extract have
been shown to inhibit the activity of HIV type 1 reverse transcriptase (Ogata et al., 1992).
P. amarus extract has been shown to reverse the chromosomal alterations induced by
genotoxic agents (Gowrishanker and Vivekanandan 1994). P. amarus extract has been
reported to posses antioxidant activities (Joy and Kuttan, 1995). A variety of hydrolysable
tannins purified from P. amarus were found to be potent inhibitors of wheat embryo Ca 2+
dependent protein kinase (CDPK), rat brain Ca 2+ protein kinase, phospholipid dependent
protein kinase C (PKC) and Ca 2+ calmodulin-dependent myosin light chain kinase (Polya
et al., 1995).
Deregulated cell proliferation is a key factor in carcinogenesis (Silvestrini, 1994).
Determination of cellular proliferation may thus be used to predict the proliferative
potential of tumour cells. Many proteins are known to be involved in the process of cell
proliferation and some of them have regulatory roles. Such proteins can be used as markers
of proliferative activity. Silver binding Argyrophilic Nucleolar Organizer Regions
(AgNOR) has been established as accurate estimates of tissue proliferation and therefore as
a valuable means of studying the biologic behaviour of tumour cells (Crocker et al., 1988).
AgNORs represent ribosomal DNA transcription activity and hence are an active reflection
of proliferative activity. especially the percentage of cells in the S phase (Eagan and
Croser, 1992; Mourad et al., 1992). AgNOR dots and clusters were reduced in the HCC
harboring rats treated with P. amarus extract. The extract was found to inhibit the activity
of topoisomerase I1 In Succhromyces cenmiacae mutant cell cultures. DNA topoisomerases

are enzymes that catalyze the interconversion of DNA topoisomers and play crucial role
during replication to relax the torsional strain generated when helicases unwind the duplex.
They perform similar function during transcription. Cancer cells have higher
topoisomerase I1 levels. Therefore, inhibition of this enzyme will lead to a decrease in cell
proliferation. The extract inhibited the activity of cdc25 tyrosine phosphatase, an important
enzyme involved in cell cycle regulation. This enzyme is a critical regulator of M phase
promoting factor (MPF), a universal intracellular factor, which removes inhibitory
phosphates in the catalytic domain of p34cdc2. Cdc25 tyrosine phosphatase thereby directly
controls entry of cells into M phase and is therefore a very suitable target to detect anti-
mitotic compounds. P. arnancs extract has been documented to scavenge oxygen free
radicals (Joy and Kuttan, 1995). Oxygen free radicals were implicated in the activation of
several carcinogens to their ultimate form as well as in tumour promotion (Kinsler and
Taffe, 1986). P. amarus extract was found to inhibit the P-450 enzyme aniline
hydroxylase, which is involved in the activation pathway of several carcinogens. The
inhibition of liver tumours may be due to the scavenging of noxious reactive oxygen
species and the inhibition carcinogen activation. The prolonged longevity observed in
ascites tumour harboring mice and HCC harboring rats may be due to the inhibition of,
DNA top I1 and cell cycle arrest of tumour cells thus leads to reduction in tumour cells and
their proliferative capacity.