rabies virus infection ofcultured rat sensory neuronsjvi.asm.org/content/61/9/2733.full.pdfrabies...
TRANSCRIPT

JOURNAL OF VIROLOGY, Sept. 1987, p. 2733-27410022-538X/87/092733-09$02.00/0Copyright © 1987, American Society for Microbiology
Rabies Virus Infection of Cultured Rat Sensory NeuronsERIK LYCKEt AND HENRI TSIANG*
Unite Rage, Institut Pasteur, Paris, France
Received 10 February 1987/Accepted 1 June 1987
The axonal transport of rabies virus (challenge virus strain of fixed virus) was studied in differentiated ratembryonic dorsal root ganglion cells. In addition, we observed the attachment of rabies virus to neuronalextensions and virus production by infected neurons. A compartmentalized cell culture system was used,allowing infection and manipulation of neuronal extensions without exposing the neural soma to the virus. Thecultures consisted of 60% large neuronal cells whose extensions exhibited neurofilament structures. Rabiesvirus demonstrated high binding affinity to unmyelinated neurites, as suggested by assays of virus adsorptionand immunofluorescence studies. The rate of axoplasmic transport of virus was 12 to 24 mm/day, including thetime required for internalization of the virus into neurites. The virus transport could be blocked bycytochalasin B, vinblastine, and colchicine, none of which negatively affected the production of virus in cellsonce the infection was established. It was concluded that, for the retrograde transfer of rabies virus by neuritesfrom the periphery to the neuronal soma, the integrity of tubulin- and actin-containing structures is essential.The rat sensory neurons were characterized as permissive, moderately susceptible, but low producers of rabiesvirus. These neurons were capable of harboring rabies virus for long periods of time and able to release virusinto the culture medium without showing any morphological alterations. The involvement of sensory neurons
in rabies virus pathogenesis, both in viral transport and as a site for persistent viral infection, is discussed.
The spread of rabies infection through the axoplasmic flowin peripheral nerves and neurons in the central nervous
system (CNS) represents an important and significant patho-genetic characteristic. Interruption of the peripheralaxoplasmic flow by ligature or sectioning (4, 15, 26) or bypharmacological impairment of the microtubulin functionprevents rabies virus transport from the peripheral virusinoculation site to the CNS (8, 43). Peripheral nerves of bothmotor and sensory neurons can be used for propagation ofrabies virus infection (10, 20), although sensory neurons are
most probably involved in the virus transfer from the site ofthe primary infection to the CNS (35). It is generally ob-served, by electron microscopy, that rabies virus buds frominfected neurons and subsequently enters into uninfectedcells (23, 24, 34, 39). However, the hypothesis of an axonaltransport of virus in the CNS has never been discarded (35).Virus inoculation into the eye has resulted in virus transportinto the CNS (31). Recently, experimental evidence has beenprovided for an axonal transport of rabies virus within theCNS. Stereotaxic inoculation of the virus into rat brainsgave patterns of virus spread which are consistent with thehypothesis of an axoplasmic flow-mediated transport (19).
Infection of cultured nerve cells (44, 47) suggested thatsusceptibility to rabies virus is likely to vary markedlydepending on the types of neuronal cells. In the presentstudy, we used rat dorsal root ganglia (DRG) neurons forevaluating axonal transport of rabies virus in vitro. Inaddition, we studied virus binding to neuritic extensions andits replication in sensory neurons. For this purpose, we tookadvantage of a compartmentalized cell culture system (11)which allowed us to infect and manipulate neuronal exten-sions without exposing the neuronal cell soma to the virus.
* Corresponding author.t On leave of absence from the Department of Virology, Institute
of Medical Microbiology, University of Goteborg, Goteborg, Swe-den.
MATERIALS AND METHODS
Virus. Challenge virus strain (CVS) fixed rabies virus wasobtained from supernatants of BHK-21 (baby hamster kid-ney)-infected cells. Cell monolayers were infected with a
multiplicity of infection of 5 PFU/ml and incubated for 3days at 37°C in Eagle minimal essential medium containing2% fetal calf serum (Flow Laboratories Ltd., Ayrshire,United Kingdom), 2 mM glutamine, penicillin (100 U/ml),and streptomycin (100 [Lg/ml). Infected supernatants werecentrifuged to remove cell debris, divided into aliquots, andstored at -70° until used.
Titration of infectious virus. The concentration of infec-tious virus was determined on cultures of chicken embryo-related cells as described by Smith et al. (38). Briefly, thesecells were grown in a medium containing 1.2 g of NaHCO3per liter and supplemented with 10% fetal calf serum, 2 mMglutamine, and antibiotics. After 6 days of incubation in a 5%C02, humidity-saturated incubator at 37°C, the cells werefixed with formaldehyde and stained with crystal violet.
Rabies virus antiserum. Anti-rabies virus antiserum was
prepared by immunizing rabbits with beta-propiolactone-inactivated, purified rabies virus emulsified in Freund adju-vant, as described earlier (3). Rabbits were given fourimmunizing injections at 1-week intervals. One week afterthe last injection, a blood sample was collected for titrationof the antibodies, using the plaque inhibition test. Theantibody titer was expressed as international units, using as
a reference an international rabies antibody standard. Thetiter of the serum used was 30 international units.Nerve cell cultures. DRG cells were obtained by dissecting
10 to 12 embryos of 15- to 17-day-pregnant Wistar rats.Ganglia were collected using forceps under a dissectionmicroscope, and cells were dissociated by treatment with0.25% trypsin in Ca2+- and Mg2+-free Hanks buffer at pH 7.2for 30 min at 37°C. The cells were then separated fromtrypsin by low-speed centrifugation and suspended in culture
2733
Vol. 61, No. 9
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2734 LYCKE AND TSIANG
FIG. 1. The two-chamber cell culture system, a collagen-coatedpetri dish with scratches made in the coat to direct neuritesextending from differentiating neurons. In the center of the dish, a
cloning cylinder is attached to its surface by silicon high-vacuumgrease and methylcellulose. Neuronal cells are seeded in the cloningcylinder (inner culture compartment); neuritic extensions will growacross the grease-cellulose seal and invade the outside of thecylinder (outer culture compartment).
medium. DRG cells were seeded into the inner compartmentof a two-chamber cell culture system originally developed byCampenot (11). The system, which was adapted for studieson nerve cell-virus interactions (32, 49), allows neuronalextensions which have outgrown in the outer culture cham-ber to be infected with the virus, without exposure of theneural cell soma in the inner compartment. I'he system (Fig.1) consisted of a 35-mm collagen-coated (Vitrogen 100,Collagen Corp., Palo Alto, Calif.) plastic plate (Falcon;Becton Dickinson, Grenoble, France), scratched across itscenter with stainless-steel needles placed in a brush 0.5 mmapart. An 8-mm-wide glass cylinder was put in the center ofthe plate and sealed with high-vacuum silicon grease. In thelower part of the glass cylinder, the trenches formed by thescratches in the collagen coat were sealed with 1%
methylcellulose (Methocel 400, Fluka AG, Buchs, Switzer-land) in Hanks buffer (pH 7.2). The glass cylinder serves asa tight diffusion barrier in the culture. After 1 day of culture,the cells were treated with 2.8 jig of cytosine arabinoside(Sigma Chemical Co., St. Louis, Mo.) per ml of medium for2 days to reduce the growth of cells exhibiting mitoticactivity. On day 4, the cultures were rinsed and freshmedium was added.We further modified the technique described above for the
present study. The position of the glass cloning cylinder wasidentified on the external side of the plate to facilitate themicroscope recognition of the areas observed, which are
either inside or outside the barrier even after removal of thecloning cylinder. We also used FIO cell culture medium(Boehringer, Mannheim, Federal Republic of Germany) sup-
plemented with 10%, fetal calf serum because this mediumappeared to restrict growth of contaminating fibroblasts. Tostimulate differentiation of neuronal cells and outgrowth ofneurites, UV light-sterilized GTlb ganglioside (the generous
gift of B. Hauttecoeur, Institut Pasteur) and nerve growthfactor (Collaborative Research Inc., Lexington, Mass.) wereadded as suggested by Hauw et al. (21) to final concentra-tions of 10 and 25 ng/ml, respectively.
It was repeatedly observed that the wall between the innerand outer culture compartments seemed a tight diffusionbarrier, as (i) the fluid level inside the glass cylinder re-mained at a relatively higher level than outside fluid; (ii)radiolabeled low-molecular-weight substances did not dif-fuse across the barrier; and (iii) the addition of concentratedH2SO4 outside the glass cylinder to destroy peripheralneurites did not result in any morphological cell changesinside the barrier.
Destruction of neuritic extensions. Neuritic extensions pre-sent in the outer culture chamber were destroyed by theaddition of 200 to 300 ,ul of concentrated H2S04. After a fewseconds of contact with the acid, the culture was rinsed withmedium until neutrality was restored.
Immunofluorescence. Rabies virus-infected cultures werestained with a rabbit anti-rabies virus nucleocapsid immuno-globulin G conjugated with fluorescein isothiocyanate(FITC; kindly prepared by P. Versmisse) as described earlier(46). Stained cultures were washed with phosphate-bufferedsaline, mounted in Elvanol, and examined with an invertedZeiss 1M32 microscope (Oberkochen, Federal Republic ofGermany).
Staining of neurites with monoclonal antibodies againstneurofilaments was performed with a mouse immunoglobu-lin Gl antibody (clone 2F11; Monosan, Uden, The Nether-lands).
RESULTS
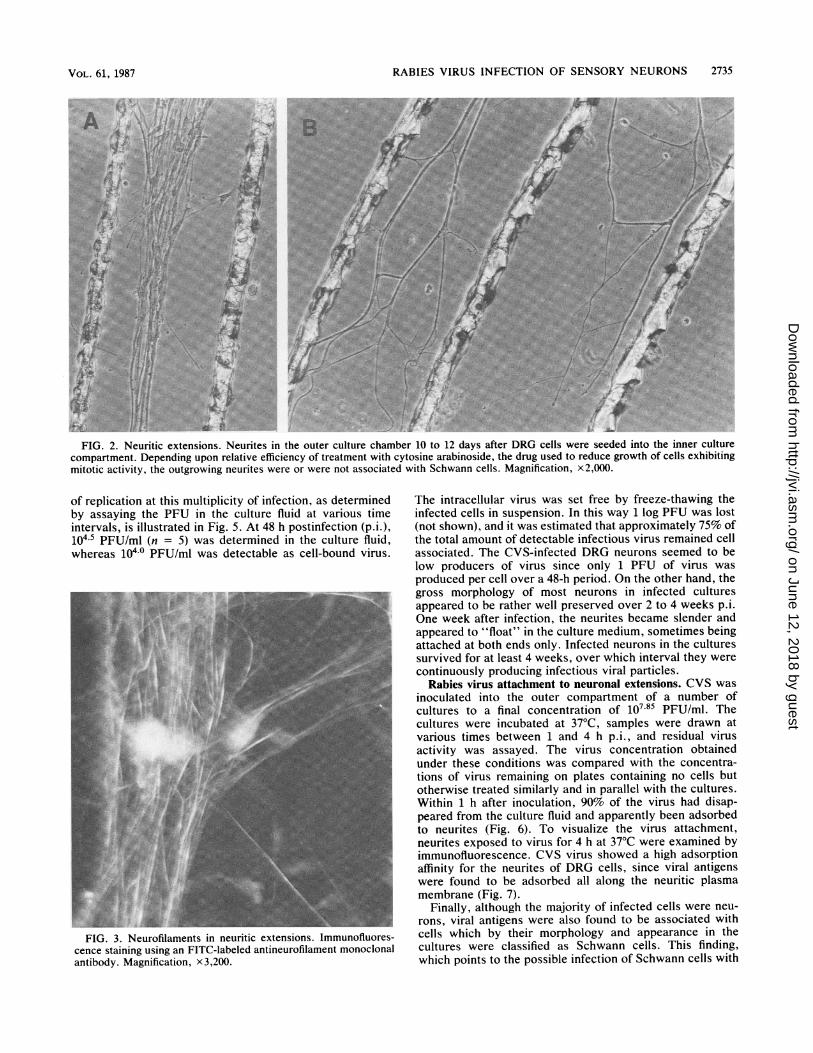
Uninfected DRG cell culture. The DRG cells seeded intothe inner culture compartment extended neurites whichwithin 8 to 12 days grew across the seal into the area outsidethe barrier. About 2 x 105 to 3 x 105 cells were present ineach cylinder as calculated from six cultures, of which 60%consisted of large neuronal cells. Neurons extendingneurites outside the barrier had no particular location in theculture; they were observed with the same frequency in thecenter of the culture as in the periphery. However, only aminor proportion of the neurons extended neurites capableof reaching the outer culture compartment. No more than236 neurites were counted in the outer compartment of aculture (the mean value was in the range of 68 to 96; n = 18).However, since the neuronal extensions seemed most oftento be able to penetrate the diffusion barrier only as fasciclesof extensions twisted together, the true number of neurites ina culture might be greater. Most frequently, the neuriticoutgrowth was restricted to the area between two trenches.However, some neurites were able to traverse the trenchesand form a neuritic network. Depending on the efficiency ofantimitotic treatment, neuritic extensions were found to beassociated or not with Schwann cells (Fig. 2). Staining theneurites with an FITC-labeled antineurofilament monoclonalantibody allowed us to demonstrate the presence of neuro-filaments in the extensions (Fig. 3). The neurites as well asthe neurons maintained their morphological and physiologi-cal characteristics during the 3 or 6 weeks of the culture.
Neuronal replication of CVS. Infection of DRG culturesdirectly in the cloning cylinder showed that the main targetcells of the CVS virus were the large sensory neurons (Fig.4). However, production of virus and its release into theculture fluid over a 24- to 30-h postinfection period requireda multiplicity of infection in the range of 20 PFU/ml. Kinetics
J. VIROL.
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
RABIES VIRUS INFECTION OF SENSORY NEURONS 2735
JI
FIG. 2. Neuritic extensions. Neurites in the outer culture chamber 10 to 12 days after DRG cells were seeded into the inner culturecompartment. Depending upon relative efficiency of treatment with cytosine arabinoside, the drug used to reduce growth of cells exhibitingmitotic activity, the outgrowing neurites were or were not associated with Schwann cells. Magnification, x2,000.
of replication at this multiplicity of infection, as determinedby assaying the PFU in the culture fluid at various timeintervals, is illustrated in Fig. 5. At 48 h postinfection (p.i.),104-5 PFU/ml (n = 5) was determined in the culture fluid,whereas 104-0 PFU/ml was detectable as cell-bound virus.
FIG. 3. Neurofilaments in neuritic extensions. Immunofluores-cence staining using an FITC-labeled antineurofilament monoclonalantibody. Magnification, x3,200.
The intracellular virus was set free by freeze-thawing theinfected cells in suspension. In this way 1 log PFU was lost(not shown), and it was estimated that approximately 75% ofthe total amount of detectable infectious virus remained cellassociated. The CVS-infected DRG neurons seemed to below producers of virus since only 1 PFU of virus wasproduced per cell over a 48-h period. On the other hand, thegross morphology of most neurons in infected culturesappeared to be rather well preserved over 2 to 4 weeks p.i.One week after infection, the neurites became slender andappeared to "float" in the culture medium, sometimes beingattached at both ends only. Infected neurons in the culturessurvived for at least 4 weeks, over which interval they werecontinuously producing infectious viral particles.
Rabies virus attachment to neuronal extensions. CVS wasinoculated into the outer compartment of a number ofcultures to a final concentration of 107.85 PFU/ml. Thecultures were incubated at 37°C, samples were drawn atvarious times between 1 and 4 h p.i., and residual virusactivity was assayed. The virus concentration obtainedunder these conditions was compared with the concentra-tions of virus remaining on plates containing no cells butotherwise treated similarly and in parallel with the cultures.Within 1 h after inoculation, 90% of the virus had disap-peared from the culture fluid and apparently been adsorbedto neurites (Fig. 6). To visualize the virus attachment,neurites exposed to virus for 4 h at 37°C were examined byimmunofluorescence. CVS virus showed a high adsorptionaffinity for the neurites of DRG cells, since viral antigenswere found to be adsorbed all along the neuritic plasmamembrane (Fig. 7).
Finally, although the majority of infected cells were neu-rons, viral antigens were also found to be associated withcells which by their morphology and appearance in thecultures were classified as Schwann cells. This finding,which points to the possible infection of Schwann cells with
VOL. 61, 1987
;NK'
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2736 LYCKE AND TSIANG
.I._1Es. n--<.:. .. i...:
I.~~~~~~~~~~WFI oio luoecn DR cll at1 72 hp withrabies viu (CS Imnfursecsting (A.a wth an FITlaele
anti-rabies virus nucleoprotein conjugate. (B) Phase-contrast microscope picture. Magnification, x3,200.
CVS virus, will not be discussed in the present paper but willbe dealt with elsewhere.
Transport of rabies virus in neuronal extensions. Neuriteswere infected by adding the virus (1066 PFU in a 0.5-mlvolume) to the outer compartment of DRG cell cultures. At1 h p.i., nonadsorbed virus was removed and the mediumwas replaced with fresh medium. The cloning cylinder wasfilled with medium containing 8 international units of anti-rabies virus serum per ml to exclude nonspecific extraneuralspread of virus. Incubation of the cultures at 37°C resulted inpropagation of the infection from the neurites to the cellsoma. The presence of neutralizing antibodies in the cellcultures did not significantly modify the infection; most ofthe neurons in the cylinder were found to be infected as theywere in untreated control cultures. At 24 to 30 h p.i., a few
5r
41E
U.IL
0)0-i
single neurons in the culture inside the diffusion barriershowed the presence of rabies antigen as revealed by immu-nofluorescence (Fig. 8). The infection spread from theseprimarily infected neurons to the neuritic network. Foci ofinfected nerve cells were observed which invaded the wholeculture (Fig. 9).The rate of uptake and transport of virus was subsequently
estimated from a series of experiments in which the timenecessary for establishing infection inside the cloning cylin-der was determined after the virus (107-85 PFU/ml) wasinoculated outside the diffusion barrier. The cultures wereincubated at 37°C for 1 h, and after various times rangingfrom 1 to 24 h, neurites outside the barrier were destroyed.
8.or
7.51
ILA U.67.00
6.5
3 F
1 24 48 72 96HOURS
FIG. 5. Rabies virus (CVS) replication in rat sensory neurons ofdorsal root ganglia. Amounts of virus recovered from culture liquidsat various times p.i.
---'A
II@I ***
. . . . .
2 4 6HOURS
FIG. 6. Attachment rate of CVS to to neurites of the outerculture chamber. Dotted line, Concentration of residual infectiousvirus, plotted against time. Solid line. Control displaying reductionof virus of a cell-fiee culture system.
J. VIROL.
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
RABIES VIRUS INFECTION OF SENSORY NEURONS 2737
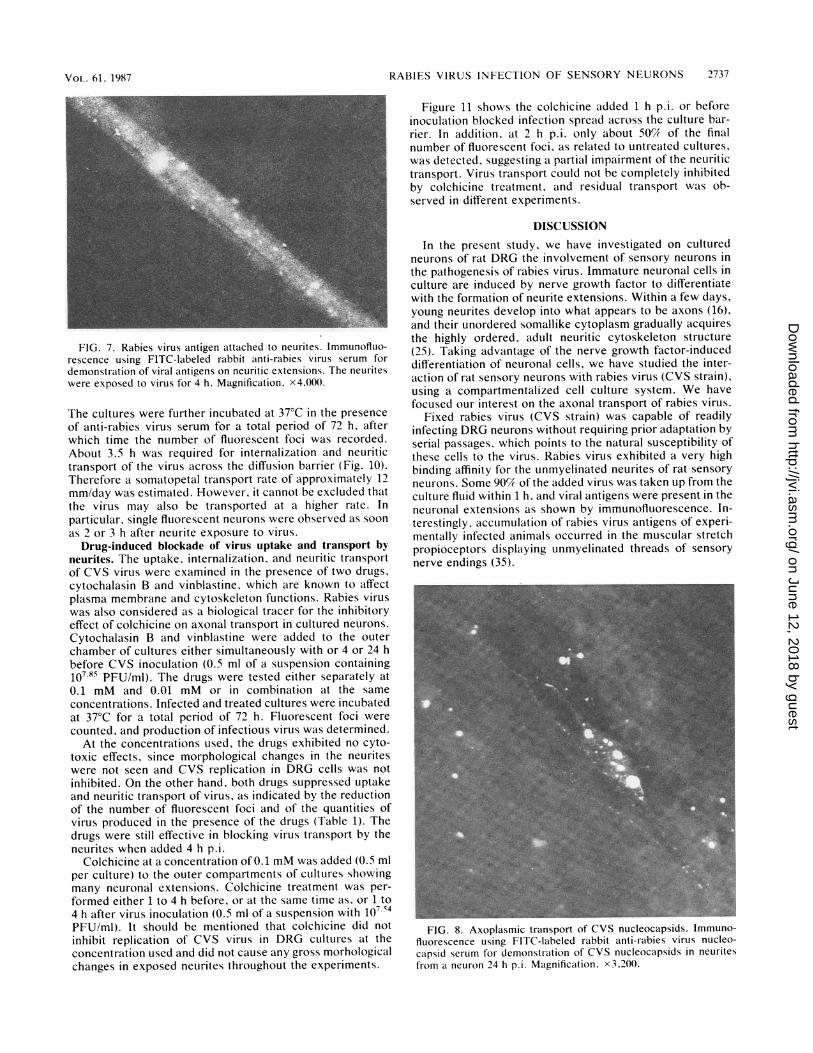
FIG. 7. Rabies virus antigen attached to neurites. Immunofluo-rescence using FITC-labeled rabbit anti-rabies virus serum fordemonstration of viral antigens on neuritic extensions. The neuriteswere exposed to virus for 4 h. Magnification. x4.O00.
The cultures were further incubated at 37°C in the presenceof anti-rabies virus serum for a total period of 72 h, afterwhich time the number of fluorescent foci was recorded.About 3.5 h was required for internalization and neuritictransport of the virus across the diffusion barrier (Fig. 10).Therefore a somatopetal transport rate of approximately 12mm/day was estimated. However, it cannot be excluded thatthe virus may also be transported at a higher rate. Inparticular, single fluorescent neurons were observed as soonas 2 or 3 h after neurite exposure to virus.
Drug-induced blockade of virus uptake and transport byneurites. The uptake, internalization, and neuritic transportof CVS virus were examined in the presence of two drugs,cytochalasin B and vinblastine, which are known to affectplasma membrane and cytoskeleton functions. Rabies viruswas also considered as a biological tracer for the inhibitoryeffect of colchicine on axonal transport in cultured neurons.Cytochalasin B and vinblastine were added to the outerchamber of cultures either simultaneously with or 4 or 24 hbefore CVS inoculation (0.5 ml of a suspension containing107.85 PFU/ml). The drugs were tested either separately at0.1 mM and 0.01 mM or in combination at the sameconcentrations. Infected and treated cultures were incubatedat 37°C for a total period of 72 h. Fluorescent foci werecounted, and production of infectious virus was determined.
At the concentrations used, the drugs exhibited no cyto-toxic effects, since morphological changes in the neuriteswere not seen and CVS replication in DRG cells was notinhibited. On the other hand, both drugs suppressed uptakeand neuritic transport of virus, as indicated by the reductionof the number of fluorescent foci and of the quantities ofvirus produced in the presence of the drugs (Table 1). Thedrugs were still effective in blocking virus transport by theneurites when added 4 h p.i.
Colchicine at a concentration of 0.1 mM was added (0.5 mlper culture) to the outer compartments of cultures showingmany neuronal extensions. Colchicine treatment was per-formed either 1 to 4 h before, or at the same time as. or 1 to4 h after virus inoculation (0.5 ml of a suspension with 107.54PFU/ml). It should be mentioned that colchicine did notinhibit replication of CVS virus in DRG cultures at theconcentration used and did not cause any gross morhologicalchanges in exposed neurites throughout the experiments.
Figure 11 shows the colchicine added 1 h p.i. or beforeinoculation blocked infection spread across the culture bar-rier. In addition. at 2 h p.i. only about 50% of the finalnumber of fluorescent foci, as related to untreated cultures,was detected, suggesting a partial impairment of the neuritictransport. Virus transport could not be completely inhibitedby colchicine treatment, and residual transport was ob-served in different experiments.
DISCUSSIONIn the present study, we have investigated on cultured
neurons of rat DRG the involvement of sensory neurons inthe pathogenesis of rabies virus. Immature neuronal cells inculture are induced by nerve growth factor to differentiatewith the formation of neurite extensions. Within a few days,young neurites develop into what appears to be axons (16),and their unordered somallike cytoplasm gradually acquiresthe highly ordered, adult neuritic cytoskeleton structure(25). Taking advantage of the nerve growth factor-induceddifferentiation of neuronal cells, we have studied the inter-action of rat sensory neurons with rabies virus (CVS strain),using a compartmentalized cell culture system. We havefocused our interest on the axonal transport of rabies virus.
Fixed rabies virus (CVS strain) was capable of readilyinfecting DRG neurons without requiring prior adaptation byserial passages, which points to the natural susceptibility ofthese cells to the virus. Rabies virus exhibited a very highbinding affinity for the unmyelinated neurites of rat sensoryneurons. Some 90% of the added virus was taken up from theculture fluid within 1 h, and viral antigens were present in theneuronal extensions as shown by immunofluorescence. In-terestingly, accumulation of rabies virus antigens of experi-mentally infected animals occurred in the muscular stretchpropioceptors displaying unmyelinated threads of sensorynerve endings (35).
FIG. 8. Axoplasmic transport of CVS nucleocapsids. Immuno-fluorescence using FITC-labeled rabbit anti-rabies virus nucleo-capsid serum for demonstr-ation of CVS nucleocapsids in neuritesfrom a neuron 24 h p.i. Magnification. x3,200.
VOL. 61. 1987
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2738 LYCKE AND TSIANG
FIG. 9. The neurons and neurites which constitute the neuronal network in the inner chamber of the cylinder were infected andsubsequently incubated (in the presence of anti-rabies virus antibodies). The cultures were stained with anti-rabies virus nucleoproteinfluorescent conjugate 72 h after virus inoculation. Magnification, x3,200.
At present it is difficult to evaluate to what extent thisbinding involves virus-specific receptors. The rabies virusreceptor has not been identified as yet. A role for glycolipidsin rabies virus binding (42), which would be mediated by
Q 100o A.,
CL4,,.
0
X~~~~~~:
A:
_
0 2 4 6 8 24
HOURSFIG. 10. Rate of axoplasmic transport of rabies virus in neurites.
Neurites in the outer culture compartment were exposed to CVS. Atvarious intervals ranging from 1 min to 24 h p.i., all neuriticextensions outside the diffusion barrier of the culture were elimi-nated by treating the outer culture compartment with H2S04. Thecultures were subsequently incubated at 37°C for a total period of 72h, after which time the number of fluorescent foci in the innerculture compartment was recorded. The results are the mean ofthree experiments. The number of fluorescent foci is expressed asthe percentage of the results obtained with an untreated but infectedcontrol.
their sialic acid residues (40), appears likely. Consequently,cellular gangliosides might be an integral part of the rabiesvirus receptor (41). It is worth mentioning that the nature ofgangliosides varies according to nerve cell types (2).
Although neurons were predominantly infected, infectionalso affected other cell types (23, 34, 39). In a previouspaper, we have reported that infection of dissociated cellsfrom the mouse DRG concerned 10% of nonneuronal cells,some of which were identified as Schwann cells (47). Itappears to us that rat Schwann cells can also be infected,although their relative susceptibility to rabies virus is dif-ficult to evaluate (unpublished data). The possible involve-ment of Schwann cells in rabies pathogenesis is still to beestablished.
It is well documented that the axonal transport machineryis also capable of directing movement of foreign organellesand particles such as injected synaptic vesicles and inertbeads with charged surfaces (1, 6, 37). The rate of axonaltransport of rabies virus in neuritic extensions was estimatedto be close to 12 mm/day, as 2 mm was the minimal distance
TABLE 1. Neuritic transfer of rabies virus after treatment ofneurites with cytochalasin B and vinblastine"
Treatment" Time p.i. No.sc f Log PFU/ml(h) fluorescent foci LgPUm
None 0 205 (100) 2.54Cyt 0 9 (4) 0Vin 0 12 (6) 0Cyt + Vin 4 22 (16) 0Cyt + Vin 24 190 (93) 2.17
" Rabies virus was inoculated into the outer culture compartment. Thepresence of viral antigens (fluorescent foci) or infectious virus in the innerculture compartment at 72 h p.i. was considered as an indication of neuritictransfer of virus across the barrier between the compartments.
' Final concentrations of cytochalasin B (Cyt) and vinblastine (Vin) were0.1 and 0.01 mM, respectively.
J. VIROL.
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

RABIES VIRUS INFECTION OF SENSORY NEURONS 2739
FIG. 11.Neurites incolchicine ((after (-) inoinner cultur4cent foci derThe results;percentages
to be run fbarrier of th was the tis consider
simplex vii(11). The racetylcholimm/day indirection, caxonal traraxonal trarsince the tievents, i.e
port. In thetion by nefusion of ea rapid reais likely totransport.not yet krmicroscopeshortly aftenvelopedmay be meE. Lyke, a
pits wereinfection ocervical gainvolve co,some timerate of virn
Recentlyro-9-3-(2-hlintracellulavarious wa
axons were found to block the transport of the lectin wheatA germ agglutinin and of herpes simplex virus in neurites of
DRG cells (30). This effect was ascribed to the action of thedrugs on microtubular transport. Consistent with the prob-able importance of plasma membrane and cytoskeleton
A \ activities for neuritic uptake and transport of viruses, wefound that both cytochalasin B and vinblastine efficientlyblocked the neuritic transfer of rabies virus in the DRG cellcultures. Cytochalasin B inhibits microfilament formation by
v preventing actin from assembling and consequently blocksendocytosis (9, 33). Cytochalasin B could also block axonaltransport by abolishing microtubule formation (22). Micro-
A tubule inactivation resulting in transport dysfunction canA
also be caused by vinblastine, which is supposed to induceA microtubule aggregation (7). In addition, colchicine, which is
................................................. most commonly used for blocking axonal transport, exer-A cises a depolymerizing effect on microtubules. We verified
A that colchicine inhibited neuritic transfer of rabies virusl I when it was administered either before, or at the same time
-4 -2 0 +2 +4 as, or up to 1 h after virus inoculation. At 10 FM concentra-
HOURS tion. maximal inhibition of microtubule assembly inHOURS neuropils required 1 h (36). We conclude from these data thatEffect of colchicine on neuritic transport of rabies virus. both tubulin- and actin-containing structures appeared to bethe outer culture compartment were treated with involved in the neuronal transport of rabies virus. However,
D.1 mM) either before (+). or at the same time as (0), or the relative participation of each structure is not known atculation of CVS. The amount of virus transported to thee compartment was recorded as the number of fluores- present.nonstrable after a total incubation period of 72 h at 37C. The rat DRG sensory neuron can be described as beingare the mean of three experiments and are expressed as permissive but only moderately susceptible to rabies virus,of the results obtained with untreated control. which is suggested by the large multiplicity of infection
necessary for establishing a productive infection. This con-trasts with the high susceptibility usually found with primary
or transferring infection across the diffusion-tight cultures of neurons derived from the mouse and rat CNS (47;he compartmentalized culture system and 3.5 to 4 unpublished data). Furthermore, the rat sensory neuronime necessary for this transfer to occur. This rate appears capable of producing only relatively small amountsably less than that previously reported for herpes of virus. We estimated that there was about 1 PFU ofrus (50 mm/day), using the same culture system infectious virus produced per DRG neuron over a 48-hrapid phase of the axonal transport of molecular period, provided that no additional DRG cells in the cultureine esterase has been estimated to be close to 150 became productively infected. The observation of a rela-the retrograde and 410 mm/day in the anterograde tively low virus production by the neuronal cells correlatesand the corresponding slow phase of anterograde well with the scarcity of virus when infected neurons wereisport is estimated as 2.9 to 5.1 mm/day (13). The examined by electron microscopy (M. Derer, unpublishednsport rates are difficult to evaluate for viruses, data). Again, it is interesting to compare these results withme measured is usually the result of a sequence of the production of high yields of infectious virions by mouse., attachment, internalization, and axonal trans- and rat CNS neurons (10- to 100-fold) (47; unpublished data)e case of herpes simplex virus, whose internaliza- and to note the incapacity of superior cervical gangliaurites and nonneuronal cells seems to occur by neurons to replicate infectious rabies virions (45). Taken allnvelope and plasma membrane (18, 32), which is together. these results are in favor of the existence of variousction, the experimentally estimated transport rate levels in neurotropism. To what extent the low production ofreflect more accurately the actual rate of axonal virus is related to the stability of neurons in infected culturesThe neuritic uptake mechanism for rabies virus is and to the prolonged production of virus remains to beiown. However, from our preliminary electron specified. The finding that rabies virus-infected DRG neu-e observations, the virus found in the nerve cells rons can be maintained in culture for over 4 weeks demon-ter inoculation of the cultures appeared to be strates the capacity of this neuronal cell type to harbor theand internalized by coated pits, and thus uptake virus for long periods of time. Consequently, DRG neuronsdiated by an endocytosislike process (M. Derer, are good candidates as sites for the protracted harboring ofnd H. Tsiang, unpublished data). In fact, coated virus in naturally occurring cases of rabies exhibiting pro-observed in the early phases after rabies virus longed incubation times.of chicken embryo-related cells (40) and superior The observation that isolated neurons or isolated groupsLnglia neurons (45) If virus internalization were to of neurons can be infected through the neuritic networkated pits, virus uptake by neurites could require despite the presence of neutralizing antibodies deserves toand thus be responsible for the seemingly slow be discussed. This finding is to be related to the differential
is axonal transport. cell-to-cell virus transmission of pathogenic and apathogenic, three substances [taxol, nocodazole, and eryth- rabies virus strains (17). In our study, we observed theydroxynonyl)adenine] which are known to inhibit infection of neurons presumably connected to the neuritestr transport of endosomes and to interfere in extending in the external compartment, followed by thetys with the retrograde transfer of organelles in progressive infection of all the neurons in the inner compart-
1oo0
501F
U.)-
c0
n
00
U)
C.)0
0
Vol. 61, 1987
-
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2740 LYCKE AND TSIANG
ment. Since this infection occurred in the presence ofneutralizing antibody even when the neurons were notcontiguous, we postulate that viral spread occurs by neurite-to-cell contact. This clearly shows that once rabies virus ishidden in a neuronal circuitry, it escapes to the host defensesand can be transported from one neuron to the next. In thisrespect, the culture device described in this paper mimics a
brain model of rabies virus infection in which there is an
entry pathway and a neuronal network.The relatively low susceptibility of DRG neurons, coupled
with a low capacity for producing viral particles, can be an
advantage for this cell type, enabling it to sustain long-terminfection. DRG neurons have already been shown to repre-sent an excellent environment for persistent infection withother viruses, such as herpesvirus (29). It is tempting toassume that rabies virus preferentially infects central neu-
rons (47) and motor neurons (44), resulting in the productionof high yields of rabies virus, whereas DRG neurons shouldrather be considered as sites for virus entry in a persistentinfection cycle. We have already shown that myotubes,which allow an abortive rabies virus infection to develop,could be a site where rabies virus can persist extraneurally(44) and that the superior cervical ganglia neurons, which are
relatively resistant to rabies virus infection, could alsoharbor the virus for long periods of time without excreting it(45). Thus a variety of cell types which are naturally suscep-tible to rabies virus infection could harbor the virus andprotect it from the host's natural defenses until an eventtriggers viral replication.There is increasing interest in the investigation of virus
circulation in the nervous system. As for rabies virus,herpesvirus transport has been reported in the brain (5, 12,29) as well as in cultured neurons (32, 49). The involvementof the neural pathways in the transport of many other viruseshas been invoked, including the scrapie agent (28), poliovi-rus (14), reovirus (48), and Semliki forest virus (27). Thecompartmentalized cell culture system described here forrabies virus is a useful tool for the investigation ofneurotropic virus transport in cultured neurons.
ACKNOWLEDGMENTS
We gratefully acknowledge the technical assistance of JanineCadinu and Simone Guillemer, and we thank Bernard Bizzini forreviewing the manuscript.The study was supported by grants from the Institut National de
La Santd et de la Recherche Medicale (CRE 846019) and from theSwedish Medical Research Council (4514).
LITERATURE CITED
1. Adams, R. J., and P. Bray. 1983. Rapid transport of foreignparticles microinjected into crab axon. Nature (London) 303:718-720.
2. Ando, S. 1983. Review. Gangliosides in the nervous system.Neurochem. Intern. 5:507-537.
3. Atanasiu, P., H. Tsiang, and A. Gamet. 1974. Nouveau vaccinantirabique humain de culture cellulaire primaire. Ann. Micro-biol. (Paris) 125B:419-432.
4. Baer, G. M. 1975. Pathogenesis to the central nervous system,p. 181-198. In G. M. Baer (ed.), The natural history of rabies.Academic Press, Inc., New York.
5. Bak, I. J., C. H. Markham, M. L. Cook, and J. Stevens. 1977.Intraaxonal transport of herpes simplex virus in the rat centralnervous system. Brain Res. 136:415-429.
6. Beckerle, M. C. 1984. Microinjected polystyrene beads exhibitsaltatory motion in tissue culture cells. J. Cell Biol. 98:2126-2132.
7. Bensch, K. G., and S. E. Malawista. 1969. Microtubular crystals
in mammalian cells. J. Cell Biol. 40:95-106.8. Bijlenga, G., and T. Heaney. 1978. Post-exposure local treat-
ment of mice infected with rabies with two axonal flow inhibi-tors, colchicine and vinblastine. J. Gen. Virol. 39:381-385.
9. Brown, S., and J. Spundich. 1981. Mechanism of action ofcytochalasin: evidence that it binds to actin filament ends. J.Cell Biol. 88:487-491.
10. Burrage, T. C., G. H. Tignor, and A. L. Smith. 1985. Rabiesvirus binding at neuromuscular junctions. Virus Res. 2:273-279.
11. Campenot, R. B. 1977. Local control of neurite development bynerve growth factor. Proc. Natl. Acad. Sci. USA 74:4516-4518.
12. Cook, M. L., and J. Stevens. 1973. Pathogenesis of herpeticneuritis and ganglionitis in mice: evidence for intraaxonal trans-port of infection. Infect. Immun. 7:272-288.
13. Couraud, J.-Y., and L. Giamberardino. 1982. Axonal transportof the molecular forms of AChE: its reversal at a nervetransection, p. 144-152. In D. G. Weiss (ed.), Axonal transport.Springer-Verlag, Berlin.
14. Dal Canto, M. C., R. L. Barbano, and B. Jubelt. 1986. Ultra-structural immunochemical localization of poliovirus duringvirulent infection of mice. J. Neuropathol. Exp. Neurol. 45:613-618.
15. Dean, D. J., W. M. Evans, and R. C. McClure. 1963. Pathogen-esis of rabies. Bull. WHO 29:803-811.
16. Dichter, M. A., A. S. Tischler, and L. A. Greene. 1977. Nervegrowth factor-induced change in electrical excitability and ace-tylcholine sensitivity of a rat pheochromocytoma cell line.Nature (London) 258:501-504.
17. Dietzschold, B., T. J. Wiktor, J. Q. Trojanowski, R. I. Macfar-Ian, W. H. Wunner, M. J. Torres-Anjel, and H, Koprowski.1985. Differences in cell-to-cell spread of pathogenic andapathogenic rabies virus in vivo and in vitro. J. Virol. 56:12-18.
18. Fuller, A. O., and P. G. Spear. 1985. Specificities of monoclonaland polyclonal antibodies that inhibit adsorption of herpessimplex virus to cells and lack of inhibition by potent neutral-izing antibodies. J. Virol. 55:475-482
19. Gillet, J. P., P. Derer, and H. Tsiang. 1986. Axonal transport ofrabies virus in the central nervous system of the rat. J. Neuro-pathol. Exp. Neurol. 45:619-634.
20. Harrisson, A. K., and F. A. Murphy. 1978. Lyssavirus infectionof muscle spindles and motor end-plates in striated muscles ofhamsters. Arch. Virol. 57:167-175.
21. Hauw, J. J., S. Fenelon, J. M. Boutry, Y. Nagai, and R.Escourolle. 1981. Effect of brain gangliosides on neurite growthin guinea pig spinal ganglia tissue cultures and on fibroblast cellcultures, p. 171-175. In M. M. Rapport and A. Gorio (ed.),Gangliosides in neurological and neuromuscular function, de-velopment and repair. Raven Press, New York.
22. Isenberg, G., P. Schubert, and G. Kreutzberg. 1982. Actin, aneuroplasmic T constituent requisite for axonal transport, p.314-321. In D. G. Weiss (ed.), Axonal transport. Springer-Verlag, Berlin.
23. Iwasaki, Y., and H. F. Clark. 1975. Cell to cell transmission ofvirus in the central nervous system. II. Experimental rabies inmouse. Lab. Invest. 33:391-399.
24. Iwasaki, Y., D. S. Liu, T., T. Yamamoto, and H. Konno. 1985.On the replication and spread of rabies virus in the humancentral nervous system. J. Neuropathol. Exp. Neurol. 44:185-195.
25. Jacobs, R. J., and J. K. Stevens. 1986. Changes in the organiza-tion of the neuritic cytoskeleton during nerve growth factor-activated differentiation of PC12 cells: a serial electron micro-scopic study of the development and control of neurite shape. J.Cell Biol. 103:895-906.
26. Johnson, R. T. 1965. Experimental rabies. Studies of vulnera-bility and pathogenesis using fluorescent antibody staining. J.Neuropathol. Exp. Neurol. 24:662-675.
27. Kaluza, G., G. Lell, M. Reinacher, L. Stitz, and W. R. Willems.1987. Neurogenic spread of Semliki forest virus in mice. Arch.Virol. 93:97-110.
28. Kimberlin, R. H., and C. A. Walker. 1986. Pathogenesis ofscrapie (strain 263K) in hamsters infected cerebrally, intraperi-toneally or intraocularly. J. Gen. Virol. 67:255-263.
J. VIROL.
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

RABIES VIRUS INFECTION OF SENSORY NEURONS 2741
29. Klein, R. J. 1982. The pathogenesis of acute, latent and recur-rent herpes simplex virus infection. Brief review. Arch. Virol.72:143-168.
30. Kristensson, K., E. Lycke, M. Ryotta, B. Svennerholm, and A.Vahlne. 1986. Neuritic transport of herpes simplex virus in ratsensory neurons in vitro. Effects of substances interacting withmicrotubular function and axonal flow (nocodazole, taxol anderythro-9-3 (2-hydoxynonyl) adenine). J. Gen. Virol. 67:2023-2028.
31. Kucera, P., M. Dolivo, P. Coulon, and A. Flamand. 1985.Pathway of the early progression of virulent and avirulent rabiesstrains from the eye to the brain. J. Virol. 55:158-162.
32. Lycke, E., K. Kristensson, B. Svennerholm, A. Vahlne, and R. J.Ziegler. 1984. Uptake and transport of herpes simplex virus inneurites of rat dorsal root ganglia cells in culture. J. Gen. Virol.65:55-64.
33. MacLean-Fletcher, S., and T. D. Pollard. 1980. Mechanisms ofaction of cytochalasin B on actin. Cell 20:329-341.
34. Matsumoto, S. 1975. Electron microscopy of central nervoussystem, p. 217-235. In G. M. Baer (ed.), The natural history ofrabies. Academic Press, Inc., New York.
35. Murphy, F. A. 1977. Rabies pathogenesis. Brief review. Arch.Virol. 54:279-297.
36. Reibman, J., K. A. Haines, A. M. Rich, P. Cristello, K. N.Giedd, and G. Weissmann. 1986. Colchicine inhibits ionophore-induced formation of leukotriene B4 by human neutrophils. Therole of microtubules. J. Immunol. 136:1027-1032.
37. Schroer, T. A., S. T. Brady, and R. B. Kelly. 1985. Fast axonaltransport of foreign synaptic vesicles in squid axoplasm. J. CellBiol. 101:568-572.
38. Smith, A. L., G. H. Tignor, K. Mifune, and T. Motohashi. 1977.Isolation and assay of rabies serogroup viruses in CER cells.Intervirology 8:92-99.
39. Sung, J. H., M. Hayano, A. R. Mastri, and T. Okagaki. 1976. Acase of human rabies and ultrastructure of the Negri body. J.Neuropathol. Exp. Neurol. 35:541-559.
40. Superti, F., M. Derer, and H. Tsiang. 1984. Mechanism of rabiesvirus entry into CER cells. J. Gen. Virol. 65:781-789.
41. Superti, F., B. Hauttecoeur, M. J. Morelec, P. Goldoni, B.Bizzini, and H. Tsiang. 1986. Involvement of gangliosides inrabies virus infection. J. Gen. Virol. 67:47-56.
42. Superti, F., L. Seganti, H. Tsiang, and N. Orsi. 1984. Role ofphospholipids in rhabdovirus attachment to CER cells. Arch.Virol. 81:377-382.
43. Tsiang, H. 1979. Evidence for an intraaxonal transport of fixedand street rabies virus. J. Neuropathol. Exp. Neurol. 38:286-296.
44. Tsiang, H., S. DeLaporte, D. J. Ambroise, M. Derer, and J.Koenig. 1986. Infection of cultured rat myotubes and neuronsfrom the spinal cord by rabies virus. J. Neuropathol. Exp.Neurol. 45:28-42.
45. Tsiang, H., M. Derer, and J. Taxi. 1983. An in vivo and in vitrostudy of rabies virus infection of the rat superior cervicalganglia. Arch. Virol. 76:231-243.
46. Tsiang, H., and J. C. Guillon. 1981. Presence of specific antigensin neuronal cells infected with fixed and street rabies virusstrains. Acta Neuropathol. 55:263-267.
47. Tsiang, H., A. Koulakoff, B. Bizzini, and Y. Berwald-Netter.1983. Neurotropism of rabies virus. An in vitro study. J. Neu-ropathol. Exp. Neurol. 42:439-452.
48. Tyler, K. L., D. A. McPhee, and B. N. Fields. 1987. Distinctpathways of viral spread in the host determined by reovirus S1gene segment. Science 233:770-774.
49. Ziegler, R. J., and R. E. Herman. 1980. Peripheral infection inculture of rat sensory neurons by herpes simplex virus. Infect.Immun. 28:620-623.
VOL. 61, 1987
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from