psychoendocrinology and growth hormone: a review
TRANSCRIPT

Psychonewtoendocrinology, Vol. 3, pp. 131-153. 0306-4530/7810401--0131 $02,0010 O Pergamon Press Ltd. 1978. Printed in Great Britain
PSYCHOENDOCRINOLOGY AND GROWTH HORMONE" A REVIEW
GREGORY M. BROWN,* Jo A. SEGGIE,* JOHN W. CHAMBERS and PRAKASH G'. ETTIGI
Neuroendocrinology Research Section, Clarke Institute of Psychiatry, Toronto, Ontario, Canada M5T IR8
(Received 14 October 1977)
SUMMARY (1) Growth hormone is a pituitary hormone which is very largely dependent on central
nervous system regulation. Both inhibitory and excitatory hypothalamic hormones exist. Only the inhibitory hormone has been isolated and characterized. (2) In experimental animals, the regulation of growth hormone secretion has been shown to be dependent on afferent inputs to the hypothalamus from a variety of limbic and other brain areas including amygdala, septum, hippocampus and midbrain structures. (3) In man there is evidence for excitatory dopaminergic, noradrenergic and serotonergic neural mechanisms. (4) Growth hormone is primarily secreted in bursts of which there are approximately seven per day in the adult human male. Bursts can be either spontaneous or can be triggered by a variety of metabolic and stressful stimuli. Most of the effects of growth hormone appear to be mediated by another hormone, somatomedin, whose release is regulated by growth hormone. (5) Elevated levels of growth hormone are seen in a variety of conditions in which there axe no direct psycho- logical effects. Conversely, it is rather unlikely that there is a psychogenic origin for the hyperseeretion of growth hormone seen in aeromegaiy. In response to physical or psycho- logical stress, a rise in growth hormone is frequently seen which does not always correspond to adrenal activation (an accepted index of stress). GH can show a rise independent of cortisol and the reverse is also true. Conditions producing these divergent responses are only now beginning to be defined. (7) Elevation in growth hormone seen in anorexia nervosa is clearly related to nutritional status in that those patients with poor caloric intake have an elevated level which is reversed rapidly as the caloric intake increases. Reduced levels of growth hormone found in hypopituitarism or in single defects of pituitary function appear to produce no direct psychological effects. (8) In severe endogenomorphic depression, defective growth hormone responses to insulin hypoglycemia are seen in a high proportion of patients and frequently co-exist with the presence of growth hormone responses to TRH, a response which does not occur in normal subjects. These findings may be related to an underlying neurotransmitter abnormality. (9) In psychosociai deprivation, deficiency of growth hormone responses has also been reported which correlates with poor growth. It is important, however, to differenti- ate this condition from caloric deprivation in which there is inadequate food intake as many of such patients have elevated growth hormone resting levels.
Key Words--growth hormone; psychoneuroendocrinology; limbic system; mental illness.
INTRODUCTION
HUMAN growth ho rmone ( H G H ) is a prote in hormone consist ing of a l inear chain of 191 amino acids which is produced by the acidophil cells of the pi tui tary gland (Li & Bewley,
1976). I t is a un ique hormone and, contrary to earlier views, is clearly dist inguishable from
* Now at the Department of Neurosciences, McMaster University, 1200 Main St. W., Hamilton, Ontario, Canada L8S 4J9.
131

132 GREGORY M. BROWN et al.
prolactin. Growth hormone and prolactin share physiologic effects to a small extent which may represent some common chemical structure in these hormones. However, HGH differs not only in its structure but in the nature of its regulatory mechanisms and in its effects. Biologically active HGH fragments have been described (Li & Bewley, 1976; Lewis, Singh, Peterson & Vanderlaan, 1976; Kostyo, Bills, Reagan, Rudman & Wilhelmi, 1976) and it has been hypothesized that modification to a 2-chain structure during secretion may be essential for full biological activity (Lewis et al., 1976). There is evidence that there are two forms of HGH in plasma: one detectable by both radioimmuno- and bioassay and termed immunoreactive, the other biologically active only and thus termed bioactive (Ellis, Grindeland, Reilly & Yang, 1976). Because of the availability of sensitive radio- immunoassays for growth hormone, much is known of its physiologic and pharmacologic regulation and of the effects of various diseases on growth hormone release (Brown & Reichlin, 1972; Martin, 1973a; Reichlin, 1974; Brown & Kirpalani, 1975). It is now known that growth hormone is a very dynamic hormone whose levels change rapidly in response to a variety of factors.
HYPOTHALAMIC CONTROL The control of growth hormone is largely of hypothalamic origin. There are at least two
hypothalamic factors, one excitatory and the other inhibitory, which are involved in control of this hormone. What is the relative role of these two factors ? Information on this issue comes from studies of patients with section of the pituitary stalk (Glick, Roth, Yalow & Berson, 1965). In such individuals, blood levels of growth hormone remain very low indicating that in the human, GH control by the hypothalamus is primarily excitatory. Many and continued attempts have been made to isolate and purify the releasing factor(s). It was in the course of these studies that an inhibitory factor was isolated (Brazeau, Vale, Burgus, Ling, Butcher, Rivier & Guillemin, 1973). This growth hormone release inhibiting hormone, which has been named somatostatin has the structure
H-Ala-Cf s--Lys-~Asn-Phe-Phe ~ Trp- Ly-s-Thr-Phe--Thr-Ser-C~, s-OH. Using immunohistological techniques, neurons containing somatostatin have been
localized in the preoptic area and anterior periventricular hypothalamus between the anterior commissure and the optic chiasm and as far posteriorly as the anterior portion of the ventromedial nucleus (Alpert, Brawer, Patel & Reichlin, 1976). Localization of somato- statin in the anterior regions closely corresponds to one of the areas in which Krulich and coworkers originally localized growth hormone release inhibiting activity using a bioassay technique (Krulich, Illner, Fawcett, Quijada & McCann, 1972). The other area identified by Krulich and McCann as containing this activity was the median eminence itself. This site presumably represents nerve terminals of the above neurons. In addition to localization in the hypothalamus, numerous studies have shown that bioassayable and immunoassayable somatostatin is widely distributed in extrahypothalamic regions in brain and is also found outside of brain, presumably having a role separate from a role in pituitary regulation in these areas (Kato, Chihara, Ohgo & lmura, 1974).
Somatostatin is clearly a physiologic substance rather than one which has strictly pharma- cologic effects as injection of somatostatin antiserum and the subsequent neutralization of

PSYCHOENDOCRINOLOGY AND GROWTH HORMONE 1 33
200. C
1OO. C
75.( 50. C
25.c
~ lllO
~ 5.0
~ z.5
LO
0.5
dh..-----~Acid Saline 15 rni. I.~--.~mSomatostatin 30 pglkg
in 15 rnl,
I I I I I I I 0 30 60 90 120 150 180
TIME (min.)
Fie. 1. Growth hormone levels rise on cessation of somatostatin infusion in the rhesus monkey.
endogenous somatostatin leads to higher levels of growth hormone (Ferland, Labrie, Jobin, Arimura & Schally, 1976).
Under certain conditions, somatostatin can initiate release of GH (Fig. 1). During infusion of somatostatin, GH levels are low as expected while following the termination of infusion there is a rebound rise in GH (Chambers & Brown, 1976a). However, the rebound rise in GH produced by somatostatin withdrawal should not be interpreted as evidence that there is no GH releasing hormone. The rebound rise does not reach a sufficient height and does not remain sufficiently long to mimic the GH rise produced by a variety of provocative stimuli.
For example, in the rhesus monkey the infusion of 0.15 mg/kg of clonidine (Fig. 2) can produce a very large rise in GH (Chambers & Brown, 1976b). A response of this magnitude cannot be produced simply by cessation of somatostatin release. Thus, the existence of a GH releasing factor remains undisputed. The identity of this substance, however, is elusive, in part, because its activity in extracts is masked by somatostatin.
PHYSIOLOGIC REGULATION It is known that a number of metabolic stimuli trigger growth hormone secretion.
Hypoglycemia triggers growth hormone secretion and even the falling phase of elevated blood glucose following a glucose-rich meal induces growth hormone secretion (Glick et al., 1965). Several amino acids trigger growth hormone secretion (Koncz, Soeldner, Balodimos, Boden, Gleason & Younger, 1973) as does ingestion of protein-rich meals

134 GREGORY M. BROWN et al.
400
300
100
INFUSION
\ CLONIDINE HCi 0.15 mg/kg
_ _
0 30 60 90 120 150
TIME IN MINUTES
FIo. 2. In the rhesus monkey, infusion of ¢lonidine HC1 0.15 mg]kg produces a very large rise in GH.
(Burday, Fine & Schalch, 1968). Growth hormone is secreted during exercise (Schalch, 1967). There is also a secretory burst of growth hormone that occurs early after the onset of sleep, often in an association with the first non-REM phase of sleep (Finkelstein, Roff- warg, Boyar, Kream & Hellman, 1972). In addition to the above physiologic agents, growth hormone bursts occur in response to psychologic stimuli (Brown & Reichlin, 1972; Seggie & Brown, 1975) (see below).
Examination of growth hormone levels in blood samples taken through an indwelling cannula every 15 min for 24 hr shows that growth hormone is secreted episodically. In the adolescent, there are 6-7 secretory bursts of growth hormone throughout a 24-hr day while in older adults the number of episodes drops (Finkelstein et al., 1972). Some episodic bursts are spontaneous (Spitz, Gonen & Rabinowitz, 1972) while others are provoked by various agents. Why is growth hormone episodic? One simple reason is that episodic secretion seems to be characteristic of hormones that are under neural control. Growth hormone is tightly controlled by the hypothalamus. Growth hormone secretion occurs when hypothalamic tuberoinfundibular neurons turn on secretory processes either by phasic release of growth hormone releasing factor or by inhibition of somatostatin secretion. A second reason for this episodic release may be related to somatomedin. Somatomedin

I~YCHOF~DOCRINOLOGY AND GROWTH HORMONE 135
Somatostatin
/ HYPOTHALAMUS
I 1 Growth Hormone _ ~ + Releasing Hormone
PITUITARY
" / Growth Hormone $
LIVER
Somatomedin
FIG. 3. Growth hormone release from the pituitary somatotroph is controlled by two hypothalamic regulat- ing hormones, somatostatin and growth hormone releasing hormone. In response to growth hormone elevation, somatomedin is released into the circulation and mediat~ many of the effects attributed to
growth hormone.
[formerly called sulfation factor or thymidine incorporation factor, (Daughaday, Phillips & Herington, 1976)] is the protein factor which mediates most or all of the effects of growth hormone.
Somatomedin (Fig. 3) is released from liver in response to increased blood levels of growth hormone and it, in turn, mediates most or all of the effects attributed to growth hormone (Daughaday et al., 1976). It is conceivable that somatomedin is the hormone which is regulated rather than growth hormone. In that case, the enhanced secretion of growth hormone would be in response to a demand for an increase in somatomedin. Somatomedin in blood persists considerably longer than growth hormone so that periodic bursts of growth hormone release may be sufficient to maintain relatively constant blood levels of somatomedin.
PSYCHOLOGIC AND NEURAL CONTROL
What is known about the brain areas that regulate growth hormone release ? It has been established in numerous studies that the median eminence is essential for growth hormone control, and that diseases affecting this region will interfere with growth hormone regulation resulting in low GH levels with absent GH responses to provocative stimuli (Abrams, Parker, Blanco, Reichlin & Daughaday, 1966; Brown, Schalch & Reichlin, 1971b; Brown & Reichlin, 1972). The median eminence is the final common pathway by which hypo- thalamic systems regulate GH. In addition to the median eminence, a number of areas in the brain have been shown to participate in growth hormone regulation. Inhibition of growth hormone secretion appears to be related to a number of areas including the ventro- medial nucleus of the hypothalamus, the preoptic area and the cortico-medial amygdala. In contrast, release of growth hormone appears to involve the hippocampus, the base- lateral amygdala, the interpeduncular nucleus and the locus coeruleus (Martin, 1972;

136 GR~OORY M. BROWN et al.
Martin, 1973b, c; Martin, Kontor & Mead, 1973). Most of the evidence for these excitatory and inhibitory areas is from work by Martin and by Frohman (Frohman, Bernardis, Burek, Maran & Dhariwal, 1971) in the rat; however, some primate studies are also available which in general confirm work done in the rat. For instance, Smith & Root (1971) reported increases in plasma growth hormone following posterior hippocampal stimulation in the monkey while Ehle and coworkers found an increase in plasma growth hormone following amygdalar stimulation in the rhesus (Ehle, Mason & Pennington, 1977). Lesions of the midline optic chiasm in the monkey produce hyperresponsiveness to ether stress without altering basal GH (Brown et aL, 1971b).
It is worth noting that different neural areas are involved in control of plasma GH at rest as compared to conditions of stress. For instance, destruction of the septal nuclei in the rat has no effect on resting growth hormone levels nor on the 24 hr variation in these levels. However, this same lesion results in a potentiation of the growth hormone response to stressful stimulation (Seggie & Brown, 1976). This effect of septal lesions on growth hormone levels, namely, no effect on resting levels but potentiation of stress responses, is the same as the effect of septal lesions on corticosterone levels (Seggie, Uhlir & Brown, 1974a; Seggie, Shaw, Uhlir & Brown, 1974b; Seggie & Brown, 1976) suggesting that the mechanism of regulation of growth hormone release may be similar to that of other stress hormones under certain conditions of stimulation.
There is other data, however, to suggest that the mechanism of regulation of GH and corticosterone under other conditions differs. For instance, there are numerous instances in which the adrenal and growth hormone responses dissociate (see section on stress). Additionally, the previous experience of an animal can differentially affect the resting levels of these two hormones. Gentling and housing in groups are known to produce alterations in resting adrenal function in the rat (Ader, 1968). Since GH is known to be highly responsive to the same types of stimuli that produce adrenal activation, a study was done to determine whether housing in groups and gentling produce effects on resting plasma GH which are similar to those on corticosterone (Brown, Valadka & Seggie, 1975). Rats were assigned to 8 groups designed to elucidate the effects of three variables (a) housing individually vs housing in groups of 4; (b) gentled by handling and stroking for 10 min a day vs non-gentled and (c) effect of time of day: samples were collected at 2 times of day comparable to the crest and trough of the adrenal corticosteroid rhythm. At the 2 sampling times, the expected corticosterone differences were found. GH also showed crest vs trough differences (F = 7.4, d.f. = 1.56, p<0.01) in a reciprocal relationship to the corticosterone levels as has been reported in detail (Seggie & Brown, 1976). Housing in groups resulted in a lowering of growth hormone levels compared to individual housing (F = 6.5, d.f. ----- 1.56, p < 0.05). A similar effect was observed on corticosterone in agreement with previous observations. Gentling produced an elevation in resting corticosterone levels but had no significant effect on resting GH levels (F -- 3.1, d.f. --- 1.56, p > 0.05). Since in the rat the normal growth hormone response to stress is a drop in levels while the corti- costerone response is an elevation, the present data indicate that the effect of group housing on the resting level of these two hormones is not exerted via the mechanism involved in the stress response.
More recent studies in this lab have focused on the role of the amygdala in growth

PSYCHOENDOCRINOLOGY and GaOWTH HORMONE 137
E
c
10
0,-- ~ . ~0 Normal
I - , . . . . . . • Sham
• _- Amygdala i Lesion
_ T ~
2 4 6 8 10 12 14 16 18 20 22 24
Time of Day
F[O. 4. Mean growth hormone levels in groups of rats (n = 6-10) at various times of day. Each point represents the mean ~ standard error. Amygdala lesion refers to electrolytic destruction of the whole amygdala area including medial basal and cortical sections. Sham refers to being subj~ted to all the surgical
techniques up to but not including destruction of any brain tissue.
hormone regulation. Groups of rats were sacrificed under conditions of rest every 4 hr in the 24-hr day under conditions of 12 hr light, 12 hr dark. Experimental methodology was the same as described in Seggie et al. (1974b) with a lesion of the amygdala replacing a septal lesion as the experimental variable. Bilateral destruction of the whole amygdala complex including both medial and cortical nuclei had no effect on resting growth hormone levels as shown in Fig. 4 (F = 1.39, d.f. = 2 and 124, p>0.05). All groups demonstrated a significant (F = 3.05, d.f. ---- 5, 124, p < 0.01) 24 hr variation in growth hormone similar to that previously reported in rats under the same housing conditions (Simon & George, 1975; Scggie & Brown, 1976).
It is perhaps surprising that destruction of so large an area of the limbic system had no effect on resting growth hormone levels. However, the finding that stimulation of the corticomedial amygdala decreased growth hormone levels whereas stimulation of the basolateral amygdala caused release of growth hormone (Martin et al., 1973) suggests that because lesions in the present study destroyed both these areas there may have been no net effect on resting growth hormone levels. This suggestion does not hold up to experi- mental testing, however, as shown in the very recent data depicted in Fig. 5. These data were obtained in a study identical to the previous one except that destruction of the amyg- dala was confined to the basolateral area. Statistical analysis of these data has not yet been done; however, inspection of the standard errors suggest there will likely be no significant effect of the basolateral amygdala lesion on resting growth hormone levels although the expected diurnal variation is observed in all three groups.
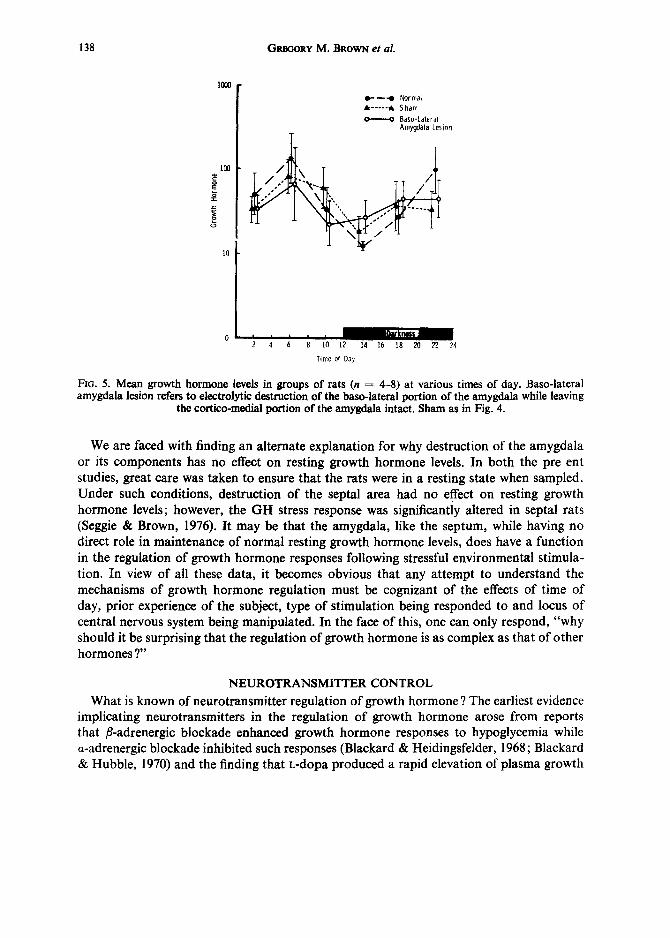
138 GREGORY M. BROWN et al.
1000
100
¢l*- m - i i Normal A. . . . . . ,L Sham
¢ 0 Baso-Lateral A mygdala Lesion
,,,'" "'x,. / t / / ,, /
2 4 6 8 10 12 14 16 18 20 ~ 24
Time of D~,
Fie. 5. Mean growth hormone levels in groups of rats (n = 4-8) at various times of day. Base-lateral amygdala lesion refers to electrolytic destruction of the base-lateral portion of the amygdala while leaving
the cortico-medial portion of the amygdala intact. Sham as in Fig. 4.
We are faced with finding an alternate explanation for why destruction of the amygdala or its components has no effect on resting growth hormone levels. In both the pre ent studies, great care was taken to ensure that the rats were in a resting state when sampled. Under such conditions, destruction of the septal area had no effect on resting growth hormone levels; however, the GH stress response was significantly altered in septal rats (Seggie & Brown, 1976). It may be that the amygdala, like the septum, while having no direct role in maintenance of normal resting growth hormone levels, does have a function in the regulation of growth hormone responses following stressful environmental stimula- tion. In view of all these data, it becomes obvious that any attempt to understand the mechanisms of growth hormone regulation must be cognizant of the effects of time of day, prior experience of the subject, type of stimulation being responded to and locus of central nervous system being manipulated. In the face of this, one can only respond, "why should it be surprising that the regulation of growth hormone is as complex as that of other hormones ?"
NEUROTRANSMITTER CONTROL
What is known of neurotransmitter regulation of growth hormone ? The earliest evidence implicating neurotransmitters in the regulation of growth hormone arose from reports that fl-adrenergic blockade enhanced growth hormone responses to hypoglycemia while a-adrenergic blockade inhibited such responses (Blackard & Heidingsfelder, 1968; Blackard & Hubble, 1970) and the finding that L-dopa produced a rapid elevation of plasma growth

PSYCHOENIX~RINOLOOY and GROWTH Ho~or~ 139
hormone (Boyd, Lebovitz & Pfeiffer, 1970). Currently the best evidence indicates that there are excitatory inputs controlling growth hormone secretion from dopamine, noradrenergic and serotonin systems. Several studies have now shown a reliable growth hormone response to the dopamine receptor stimulant apomorphine in the human (Lal, de la Vega, Sourkes & Friesen, 1972; Brown, Krieger, Van Woert & Ambani, 1974; Lal, Guyda & Bikadoroff, 1977). Clonidine, a specific noradrenergic agonist, produces a rise in growth hormone in the human (Lal, Tolis, Martin, Brown & Guyda, 1975). These findings indicate that both dopamine and noradrenergie systems are involved in growth hormone secretion although dopaminergie control appears to predominate. Serotonin mechanisms are also involved in growth hormone regulation. 5-Hydroxytryptophan (the immediate precursor of serotonin) produces a prompt rise in plasma growth hormone in man (Takahashi, Kondo, Yoshimura & Ochi, 1974) while the serotonin antagonist, cyproheptadine, reduces GH responses to hypoglycemia (Bivens, Lebovitz & Feldman, 1973).
We will now focus on dopamine control of growth hormone. There are several pathways containing dopamine in brain (Ungerstedt, 1971). The majority of dopamine neurons originate in the midbrain and are distributed to different systems---the caudate nucleus receives the nigrostriatal component of the dopamine system and is apparently primarily related to movement. There is also the mesolimbic system with distribution to the nucleus accumbens and olfactory tubercle, parts of the limbic system. This system together with the mesocortical system may be related to emotional expression. There is a dopamine system located almost exclusively within the hypothalamus. These tuberoinfundibular fibres originate in and around the arcuate nucleus and terminate in the median eminence. The ineertohypothalamic fibres which originate in the midbrain terminate in the hypo- thalamus. There is also a dopamine system located within the retina. With respect to these different systems, the question may be posed, "which of them is involved in growth hormone regulation?" There is evidence that the nigrostriatal pathway is not primarily involved. Growth hormone responses to L-dopa occur promptly in severe Parkinsonian patients in whom a major depletion of striatal dopamine is found and there is no evidence that these responses differ to a major extent from control subjects. Such a response would be unlikely to occur if those nigrostriatal dopamine pathways which are known to be grossly deficient in Parkinsonian patients were primary in growth hormone regulation (Boyd et al., 1970). It is unlikely that the retinal system is directly involved in GH regulation. We are then left with the other systems--the mesolimbic system, the mesocortical system, the tuberoinfun- dibular system, or the incertohypothalamic system.
One peculiarity of the tuberoinfundibular system is that it lies partly outside the brain. The nerve terminals of these systems are in the median eminence, a structure in which the blood brain barrier is lacking. In order to assess the role of this system in regu- lation of pituitary, we have given a number of subjects carbidopa (Brown, G-arfinkel, Warsh & Stancer, 1976). Carbidopa is a peripheral decarboxylase inhibitor which blocks the enzymatic conversion of dopa to dopamine but which does not cross the blood-brain barrier. This agent, therefore, should block the production of dopamine within terminals in the median eminence without altering dopamine pro- duction either in the nigrostriatal dopamine systems or in the mesolimbic system. A dose of 300 mg of carbidopa was given daily for 10 days. The urinary output of tryptamine

140 GREGORY M. BROWN et al.
2:
Effect of Carbidopa (300 n9 daily) on Tryptamine and Prolactin
TRYPTAMINE PROLACTIN GROWTH HORMONE
2
Control Carbidoga Control Carbidoga Control Carbidopa
30
o-c.~
FIG. 6. Following 300 mg carbidopa daily to block production of dopamine, prolactin levels rise with no apparent change in growth hormone. Drop in urinary tryptamine demonstrates that decarboxylase blockade
is effective.
was used as an index of the effectiveness of carbidopa treatment. Tryptamine levels dropped to around 10~o of resting levels following carbidopa treatment indicating that there was effective decarboxylase blockade (Fig. 6). Prolactin levels showed a significant rise. This finding suggests that the tuberoinfundibular neurons are the brain dopamine neurons which participate in prolactin regulation. This finding has recently been independently confirmed by other workers using a different inhibitor (Pontiroli, Castegnaro, Vettaro, Viberti & Pozza, 1977). Growth hormone levels, however, showed no significant change, a finding which may or may not indicate that the tuberoinfundibular neurons have no effect on growth hormone. Since resting growth hormone levels were low and the expected response was a fall, it might be impossible to detect a fall under these conditions. A recent study by Burrow, May, Spaulding & Donabedian (1977),however, throws light on this issue. In this study, dopamine was infused intravenously in normal subjects. A significant drop in prolactin was found in normal men and women; simultaneously, there was a rise in growth hormone levels. Since dopamine infused peripherally can freely enter the median eminence and act at dopamine receptors in that area as well as acting at any potential dopamine receptors on the pituitary, these findings suggest that dopamine neurons terminating in the median eminence are involved in both prolactin and growth hormone regulation in the human. This being the case, it is possible to interpret findings of studies on dopamine regulation of growth hormone and prolactin as attributable to similar brain pathways. This is important for those who propose to use such studies to assess the function of these pathways. For instance, it has been proposed that prolactin regulation may be a good index for brain dopamine systems and that studies of this system may be a useful strategy in the investigation of schizophrenia (Meltzer, Sachar & Frantz, 1974: Sachar, 1975). Both GH and prolactin regulation involve tuberoinfundibular and/or incertohypothalamic

PSYCHOENDOCRINOLDGY AND ~'ROWTH HOI~MO~ 141
dopamine systems which are unlikely to be involved directly in processes involved in schizophrenia.
SCHIZOPHRENIA In schizophrenia, there is considerable evidence suggesting hyperactivity of a dopamine
system. All the neuroleptics which control psychotic symptoms produce blockade of dopamine receptors and those agents that are most potent in blocking dopamine are also most effective in controlling psychosis (Seeman, Lee, Chau-Wong & Wong, 1976). Several investigators have examined GH responses to apomorphine in schizophrenia. Ettigi, Nair, Lal, Cervantes & Guyda (1976), have examined GH responses to apomorphine in chronic schizophrenic patients withdrawn from neuroleptic therapy. Eight of seventeen schizo- phrenic patients showed inadequate GH response to apomorphine. The peak GH response was significantly less in patients withdrawn from chronic neuroleptic therapy compared to controls. Cervantes, Lal, Smith & Guyda (1977) showed no difference between GH response in 2 schizophrenic patients with catalepsy and healthy controls. In contrast, Pandey, Garver, Hengeveld, Ericksen, Gosenfeld & Davis (1977) have reported higher GH peaks in acute schizophrenic patients than in control subjects or in chronic schizophrenic patients. If confirmed, this report has major implications. It may provide the first clear evidence for the dopamine hypothesis of schizophrenia and supports the use of the GH response as a diagnostic tool.
To this point, we have briefly reviewed the regulatory mechanism controlling growth hormone secretion including the hypothalamic factors, physiologic regulators, neural pathways and neurotransmitter systems as well as studies in one patient population-- schizophrenics. We will now review some other situations in which growth hormone levels or responses are either enhanced or reduced.
STRESS
It is now banal to point out that growth hormone is a highly stress responsive hormone. What may not be so well realized, however, is that there are many situations in which growth hormone responses do not parallel the adrenal responses.
In a study done in Dr. Reichlin's laboratory, an instance of dissociation of growth hormone and adrenal stress responses was found (Brown, Schalch & Reichlin, 1971a). In this study, squirrel monkeys were taken from their cages and placed in restraining chairs. Blood samples obtained within 15 min of the transfer showed an elevation of both the adrenal steroids and growth hormone. However, following continued chair restraint there was a drop in growth hormone levels but a continued rise in cortisol, which was maintained for several hours indicating that the difference in responses was not just due to a lag in the cortisol response. Moreover, growth hormone release could be triggered at any point by appropriate stimuli indicating that secretion was not inhibited by the elevation in cortisol. More recently, Feldmann & Brown (1975, 1976) have shown dissociation of adrenal and growth hormone responses to chair adaptation in the rhesus monkey. Following chairing, there is a rise in the rhesus monkey in resting adrenal steroids for the first few days of chair adaptation; however, no GH elevation is seen. Mason, Mailer, Hartley, Mougey, Perlow & Jones (1976), have shown a similar finding with growth hormone levels in re-

142 GREGORY M. BROWN et aL
strained monkeys returning to baseline within 2 hr of chair restraint while cortisol levels took 24 hr or more to return to baseline.
In the human, dissociation of growth hormone and adrenal responses is also seen. One situation in which growth hormone shows a reproducible rise, which may be independent of a cortisol rise, occurs following hypoglycemia (Luft, Cerasi, Madison, von Euler, Delia Casa & Roovete, 1966; Baylis, Greenwood, James, Jenkins, Landon, Marks & Samols, 1968; Yalow, Varsano-Aharm, Echemendia & Berson, 1969). In situations in which the stimulus is psychological, adrenal responses are usually found. Growth hormone responses may or may not accompany the adrenal responses. It is of considerable interest to observe and compare these situations.
TABLE I.
Cortisol alone Cortisol plus growth hormone
Cardiac catheterization ----engaged (Greene et al., 1970)
Anticipation of exhaustion (Mason, 1975)
Film-induced anxiety or sexual arousal (Brown & Heninger, 1975)
Cardiac catheterization --not engaged (Greene et aL, 1970)
Mirror drawing test in neurotics (Miyabo et al., 1976)
Catheterization in subjects with high "egotism" (Brown & Heninger, 1976)
Table I summarizes situations in which cortisol responses occur in the presence or absence of growth hormone responses. The first study to address this issue was by Greene, Conron, Schalch & Schreiner (1970). They showed that following cardiac catheterization for diagnosis of heart disease, the majority of patients showed a major cortisol elevation which was accompanied by growth hormone elevation in only certain subjects. The differ- ence between these two populations was related to the manner in which they coped with this stressful situation. Those patients without a growth hormone elevation talked to the operator and were obviosuly engaged with the environment while those who showed a growth hormone elevation did not engage with the environment. The degree of cortisol elevation did not differ between the two groups. This study points to coping style as the critical variable. Recently, Miyabo, Hisada, Asato, Mizushima & Ueno (1976) have examined the growth hormone and cortisol responses to a mirror drawing test in normal subjects and in neurotics. They found an elevation of growth hormone only in neurotic subjects with no consistent elevation in control subjects. Cortisol responses also tended to be greater in neurotics as a group but there was considerable overlap in individual responses. Miyabo e t al. (1976) concluded that (1) effective coping mechanisms exist in normal men which maintain hormonal responses at a minimum and (2) growth hormone responses were more adequate indicators of psychological stress responses in neurotic subjects. In another study, Rose & Hurst (1975) found that growth hormone responses to insertion of an intravenous catheter occurred in fewer subjects than cortisol responses. Brown & Heninger (1976) related the presence or absence of growth hormone responses following catheteriza- tion to the categories of field dependence or independence and to mood adjective check list scores. They found that growth hormone responses were more likely to occur in field inde- pendent subjects and that growth hormone responses correlated very strongly with higher

PSYCHOENDOCRINOLOOY AND GROWTH HORMO~m 143
"egotism" or "self-involvement" scores in the adjective check list. No examination of cortisol responses was made in this study. These studies further emphasize the importance of the characteristics of the population under study rather than the nature of the stimulus itself in determining growth hormone release. Mason, Maher, Hartley, Mougey, Perlow & Jones (1976) have also examined the response to venepuneture and compared growth hormone and cortisol responses. They found a close correlation between the two hormone responses; however, only a few subjects responded at all (3 of 8 subjects showed hormone response). All three responders had both growth hormone and cortisol elevation. It appears that in this situation, unlike cardiac catheterization or mirror drawing, no dissociation occurs. Mason (1975) has reported an additional study in which a striking elevation of cortisol was seen during an anticipatory period prior to the onset of a session in which subjects were to be exercised on a bicycle to the point of exhaustion. The majority of these subjects failed to show any growth hormone elevation despite a major rise in cortisol. Brown & Heninger (1975) have examined the response to film-induced anxiety and film- induced sexual arousal. Despite a consistent rise in cortisol in these situations, no consistent growth hormone elevation was seen. The demonstration of a cortisol response in the absence of a growth hormone response in this study suggests that the nature or intensity of the stimulus, although appropriate for producing cortisol release, was not appropriate for producing GH change. To summarize these findings, it would appear that in stress two factors may be important. The first factor is the nature and/or intensity of the stimulus to which the subject is exposed. Thus certain stimuli do not trigger growth hormone secretion, e.g. anticipation, film-induced anxiety, or film-induced sexual arousal while others are capable of triggering GH--such as venepuncture or cardiac catheterization. The second factor is the effectiveness of the coping mechanisms employed by the subject. These two factors combine in determining which subjects will have GH elevation in response to stress.
ANOREXIA NERVOSA Anorexia ncrvosa is another condition in which growth hormone elevation has been
reported. In anorexia nervosa, there is a constellation of pituitary findings, to such an extent that at one time it was confused with hypopituitarism (Sheehan & Summers, 1949). The firs t report of elevated growth hormone in anorexia nervosa was that of Landon, Greenwood, Stamp & Wynn, 1966 and subsequently growth hormone levels have either been reported to be normal or elevated by a number of different groups (Giordano, Marugio, Minute & Barreca, 1971 ; Lundberg, Walinder, Werner & Wide, 1972; Garfinkel, Brown, Stancer & Moldofsky, 1975; Frankel & Jenkins, 1975). We have examined growth hormone levels in 16 patients with anorexia ncrvosa and have found elevations in 8 of these 16 (Brown, Garfinkel, Jeuniewic, Moldofsky & Stancer, 1977).
The patient population that we studied included 15 females and one male. The degree of weight loss in the majority of subjects was greater than 259/o and nutritional status at the time of study as defined by caloric intake ranged from I000 to 4000 calorics per day and in the females, amenorrhea had lasted from 4 months to 70 months.
Eight of these sixteen patients showed growth hormone levels above 5 ng/ml. The growth hormone elevation did not correlate with the duration of amcnorrhea. It also did not correlate with degree of weight loss. However, a significant relationship was found with

144 GREGORY M. BROWN et al.
TABLE lI. OROWTH HORMONE LEVELS IN RELATION TO CALORIC INTAKE (ng/ml)
Poor Adequate (less than 1200 cal/day) (greater than 1500 cal/day)
59.0 2.6 13.5 0.4 6.0 1.0 3.4 0.6
19.3 1.7 16.7 2.3 4.0 4.3 9.2 5.6
Mean 16.4 2.3 S.E 6.4 0.6
t = 4.02 for iogGH.p < 0.001.
caloric intake (Table II). This relationship of elevated growth hormone to poor nutrition in patients with anorexia nervosa has been independently confirmed by Frankel & Jenkins (1975) and by Casper, Davis & Pandey (1977). Elevated levels of resting growth hormone have been shown in a variety of states in which there is decreased caloric intake including protein calorie malnutrition, kwashiorkor and marasmus (Pimstone, Becker & Hansen, 1973).
A second endocrine abnormality found in anorexia nervosa is immaturity of the circadian pattern of plasma luteinizing hormone (LH) (Boyar, Katz, Finkelstein, Kapen, Weiner, Weitzman & Hellman, 1974) so that the pattern is that of a prepubertal person. In addition, there are lowered daytime levels (Beumont, Carr & Gelder, 1973), reduced responses to clomiphene (Beumont et aL, 1973; Marshall & Fraser, 1971) and reduced responses to luteinizing hormone releasing hormone (LHRH) (Palmer, Crisp, Mackinnon, Franklin, Bonnar & Wheeler, 1975; Sherman, Halmi & Zamudio, 1975; Warren, Jewelewicz, Dyren- furth, Ans, Khalaf & Vande Wiele, 1975). We have examined resting levels of LH, responses to clomiphene and LHRH in comparison with GH levels in a population of patients with anorexia nervosa in order to determine whether abnormalities in growth hormone and in luteinizing hormone went together or whether they were related to different factors (Brown et al., 1977). We found in this study that there was a significant correlation between the percentage of loss in body weight from the original weight with resting plasma LH. In other words, the greater the degree of weight loss, the lower the resting plasma LH. In contrast, we found no relationship of resting plasma LH levels to caloric intake, to duration of amenorrhea or to the resting levels of any other of the hormones studied which included growth hormone, prolactin, TSH and cortisol as well as thyroxine. Low resting plasma LH is presumably correlated with an immaturity of the 24 hr secretory pattern in these patients as has been reported by Boyar et aL (1974). A similar relationship of plasma LH to reduced weight has been reported by several groups (Beumont et al., 1973; Beumont, George, Pimstone & Vinik, 1976; McArthur, O'Loughlin, Bectins, Johnson, Hourikan & Alonso, 1976).

PSYCHOENDOCRINOLOGY AND GROWTH HORMON~ 145
In seven of these patients, we also examined the LH response to LHRH. In these patients, we again found a correlation between per cent loss in body weight with the increase in plasma LH following LHRH, such that, those patients with the greatest loss in body weight were the ones with diminished LH responses to LHRH. These findings agree in general with previous reports (Palmer et al., 1975; Sherman et al., 1975; Warren et al., 1975; Beumont et al., 1976).
In the same patients, the increase in plasma LH was examined after administration of clomiphene daily for 5 days. Again, in agreement with the literature (Marshall et al., 1971 ; Beumont et al., 1973; Wakeling, Marshall, Beardwood, de Souza & Russell, 1976), there was a significant correlation between the increase in LH and the amount of weight loss so that those patients with the greatest loss in body weight had the smallest increase in LH. These findings then indicate that resting plasma LH, LH responses to LHRH and LH responses to clomiphene all show a significant correlation with body weight being less in those patients with the lowest body weight. None of these parameters show any relation- ship directly to caloric intake. In contrast, growth hormone resting levels showed clear cut increase in those patients with low food intake and showed no correlation with percentage of weight loss. We, therefore, conclude that the abnormality in growth hormone and the alterations in luteinizing hormone and LH reactivity are related to two separate factors.
DEPRESSION Depression is another disorder in which a variety of endocrine abnormalities have been
reported (Ettigi & Brown, 1977). Sachar's group as well as others have shown in several studies that the growth hormone
response to insulin-induced hypoglycemia is reduced in depressed patients (Sachar, Finkel- stein & Hellman, 1971; Gruen, Sachar, Altman & Sassin, 1975). Endo, Endo, Nishikubo, Yamaguchi & Hatotani (1974) have reported that the diminished growth hormone response to hypoglycemia reverts following recovery from depression indicating that this failure of growth hormone responsiveness is an integral part of the depressive illness. Depressed patients show an inadequate response to 5-hydroxytryptophan, a substance reported to cause a rise in plasma growth hormone levels in normal subjects (Takahashi et al., 1974). Thus, it appears that at least two growth hormone responses are decreased in depressed patients. In contrast to these responses, it appears that the growth hormone response to L-dopa is normal in depressed patients (Sachar, Altman, Gruen, Glassman, Halpern & Sassin, 1975). Sachar et al. (1975) showed that the responses to L-dopa were diminished in older men and women as compared to younger men and that the responses to L-dopa in depressed women in the age range of 48-70 did not differ from those in normal women of the same age group.
Recently, it has been reported that TRH which does not affect growth hormone levels in normal subjects causes an increase of growth hormone in depressed patients (Maeda, Kato, Ohgo, Chihara, Yoshimoto, Yamaguchi, Kuromaru & Imura, 1975).
In the study by Maeda et aL (1975), 8 of 13 depressed patients showed a significant increase in growth hormone levels following TRH. We also have some preliminary infor- mation on this response. A series of depressed patients were given a combined infusion of TRH and LHRH followed 2 hr later with a dose of insulin. Figure 7 shows findings in 4
~.Nza 312..-a

146 GREGORY M . BROWN e t al.
GH RESPONSE T0 TRH, LHRH, INSULIN-HYPOGLYCEMIA
40 ~ Contro l (5)
o - - - . o P r imary Depression (4l
3O
E
o =
10
. O - - ~ o ' ~ ~ o ~ ~ ~ , • ~ o ~ ~ ~ ~ .
0 15 30 45 60 90 120 15 30 45 60 ~ 120 Minutes
TRH ~ 0 pg I n s u l i n LHRH |(]0 pg 0.1 u/kg " p • 0.05
• " p • 0.025
FIG. 7. Patients with primary depression demonstrate a rise in GH following TRH that is not seen in normal subjects. In the same patients, reduced GH responses to hypoglycemia are seen.
unipolar depressed patients as compared to 5 normal subjects. None of the normal subjects showed an increase in growth hormone following combined TRH/LHRH while significant increases were seen in the 4 depressed patients. However, following insulin hypoglycemia, normal subjects all showed a rise in growth hormone while depressed patients showed a significantly smaller rise. From this preliminary evidence, it appears that the abnormal response to TRH occurs in the same subjects in which reduced responses to insulin are found suggesting that there may be a common mechanism underlying these abnormalities.
A number of other endocrine abnormalities have been repozted in depressed patients. A pervasive increase in plasma cortisol extending throughout the 24 hr daily cycle has been reported by a number of workers (Carroll, 1972a; 1972b; Sachar, Hellman, Roffwarg, Halpern, Fukushima & Gallagher, 1973). Depressed patients with elevated cortisol levels also failed to show normal suppression of dexamethasone (Carroll, 1972b; Stokes, 1972).
Yet another abnormality has been shown in depressed patients. Following the original reports that there was a brief antidepressant action of TRH, there was a flurry of studies in which TRH was infused into depressed patients (Kastin, Ehrensing, Schalch & Anderson, 1972; Prange, Wilson, Lara, Alltop & Breese, 1972). However, other workers have not been able to reproduce these results in a reliable way (Ehrensing, Kastin, Schalch, Friesen, Vargas & Schally, 1974; Hollister, Berger, Ogle, Arnold & Johnson, 1974; Furlong, Brown & Beeching, 1976). One finding, however, does stand out in many of these studies. The TSH response to TRH infusion is significantly lower in a number of depressed patients (Ehrensing et al., 1974; Brown et al., 1976) as compared with normal controls.
Maeda et aL (1975) also reported that in the depressed patients there was an exaggerated prolactin response following TRH as compared with normal subjects. Their study also shows abnormalities in growth hormone, prolactin and TSH in response to TRH in

PSYCHOENDOCRINOLOGY AND GROWTH HORMONE 147
depression. The findings with respect to the prolactin response are open to question, as in another study by Ehrensing et al. (1974), prolactin responses to TRH were found to be diminished in depressed patients. To summarize, it appears that in depression there is a constellation of pituitary findings with abnormalities in growth hormone, ACTH, TSH and perhaps in prolactin. At this time, it is not known which of these abnormalities co-exist and whether perhaps there are constellations of changes which occur in different subgroups of depressive patients. The altered endocrine profile seen in depressive disorder may well be secondary to a neurotransmitter disruption and may provide a key to understanding that abnormality.
PSYCHOSOCIAL DWARFISM We will now turn to another topic, that of psychosocial dwarfism. The entity itself is the
subject of dispute with some workers claiming that dwarfism is due to the effect of the environmental deprivation (PoweU, Brasel, Raiti & Blizzard, 1967; Brown, 1976) while Krieger (1973) has postulated that the growth failure is due to malnutrition on an organic basis. Endocrine findings of the syndrome were first described from Johns Hopkins (Powell et al., 1967).
In that study, a population of children was described who had a reduced height age for their chronological age. In searching for the cause of the height retardation, investigators examined thyroid, adrenal and growth hormone function. Thyroid function in general was normal. Adrenal function showed a reduced 17-hydroxysteroid response to metyrapone in the majority of the patients studied but the most striking finding of all was that growth hormone responses to hypoglycemia were low in 6 out of 8 patients. On the basis of these findings, the investigators postulated that due to the environmental deprivation there were abnormalities present both in adrenal regulation and in growth hormone regulation and that the reduction in growth was probably secondary to reduced secretion of growth hormone. The reduced growth hormone response to insulin-induced hypoglycemia was subsequently confirmed by many investigators (Powell et al., 1967) including two series of studies on quite large groups of patients (Kaplan, Abrams, Bell, C o n t e & Grumbach, 1968; Illig, 1972). In addition, the original Johns Hopkins series was expanded to a total of 22 patients from the original 13 (Brasel, 1973). In addition to abnormal responses to hypoglycemia, reduced responses to arginine have also been reported (IUig, 1972; Brasel, 1973; PoweU, Hopwood & Barratt, 1973). Studies on a single patient have shown that during the deprived phase there was no sleep-associated growth hormone rise despite a normal pattern of sleep (Powell et al., 1973).
What happens to these patients when they are admitted to hospital ? During periods of hospitalization, these patients exhibited catch-up growth (Powell et al., 1967). Following the period of catch-up growth, significant improvement was seen in the endocrine abnor- malities. The growth hormone response to insulin hypoglycemia returned to normal in 7 out of 7 patients studied in the original series (Powell et al., 1967) and responses to arginine as well as sleep-related changes are also reversed (Powell et al., 1973). It therefore appears that in psychosocial dwarfism there are reduced growth hormone responses to a variety of stimuli which are reversible upon removal from the deprived environment.
Krieger & Good (1970) and Krieger (1973) postulated that all the endocrine findings in

148 GREGORY M. BROWN et al.
patients with deprivation syndrome were related to malnutrition. Their study, however, differs from the previous studies. Patients were selected for study on the basis of poor environment together with retardation of language and of psychomotor development. They were not selected on the basis of growth retardation. Moreover, in reexamining their data, it is clear that the patients they studied fall into two groups. Infants under the age of 3 differed from those of age 3 and over. Children of age 3 or more showed defective growth hormone responses to insulin but had normal resting levels of growth hormone. In contrast, infants had elevated resting growth hormone but responses to hypoglycemia were not defective.
The GH profiles of infants under 3 were similar to patients with protein calorie mal- nutrition (either kwashiorkor or marasmus) (Pimstone, Becker & Hansen, 1972) or anorexia nervosa (Brown et al., 1977). The infants were largely dependent on their mothers for their feeding and they responded well to refeeding. Therefore, this group may have suffered from malnutrition rather than from psychosocial deprivation. In contrast, the older age group studied by Krieger appeared endocrinologically similar to children studied originally by Powell and coworkers and subsequently by others. These children may have been better able to counteract underfeeding by foraging for themselves so that, although their diet may have been unusual, they were more likely to maintain an adequate caloric intake. Moreover, the endocrine findings in the older groups were not those of malnutrition.
In summary, growth hormone is a pituitary hormone that is tightly controlled by the hypothalamus and importantly influenced by other brain areas. These brain areas include centers which are known to be important in the expression of emotions including areas such as the amygdala, the septal region, the hippocampus and the midbrain. Both excitatory and inhibitory hypothalamic regulatory factors are known to exist and at least 3 neuro- transmitters are involved in growth hormone regulation. In view of the foregoing, it is not surprising that growth hormone levels and responses are intimately tied in with psycho- logical functioning. Hypersecretion of growth hormone occurs during stress but only in certain types of stress. Elevated levels are also seen in states of poor nutrition. Unresponsive- ness of growth hormone is seen in some depressed patients and in the syndrome of psycho- social dwarfism. It is clear then that the examination of growth hormone resting levels and stress responses may serve as a useful index of central nervous system function. The use of growth hormone as such an index provides a tool which can be used to assess the integrity of the neural pathways involved in maintenance of the resting level or of the specific response which is examined. It can also provide information on the integrity of the neuro- transmitters in the specific neurons involved in those responses. As one example, the altered endocrine profile seen in depressive disorder is presumably secondary to a neuro- transmitter disruption and the endocrine alterations may provide a key to understanding that abnormality.
This work was supported by the Medical Research Council (M.R.C.) grant MT4749 and the Ontario Mental Health Foundation (O.M.H.F.) grants 729 and 484. G.M.B. is an O.M.H.F. Research Associate; J.A.S. is an O.M.H.F. Research Scholar; P.G.E. is an M.R.C. Fellow.

PSYCHOENDOCRINOLOGY AND GROWTH HORMONE 149
REFERENCES ABRAMS, R. L., PARKER, M. L., BLANCO, S., RExcmaN, S. & DAUGI~d~AY, W. H. (1966) Hypothalamic
regulation of growth hormone secretion. Endocrinology 78, 605-613. AVER, R. (1968) Effects of early experiences on emotional and physiological reactivity in the rat. J. Comp.
Physiol. Psychol. 66, 264-268. ALPERT, L. C., BRAWER, J. R., PATEL, Y. C. & R~Icm.IS, S. (1976) Somatostatinergic neurons in anterior
hypothalamus: immunohistochemical localization. Endocrinology 98, 255-258. BAY~JS, E. M., GP~EmVOOD, F., JAMES, V., JENKINS, J., LANDON, J., MARKS, V. & SAMOLS, E. (1968) An
examination of the control mechanisms postulated to control growth hormone secretion in man. In Growth Hormone, A. Pecile and E. E. Miiller (Eds.), pp. 89-104. Excerpta Medica, Amsterdam.
BEUMONT, P. J. V., CARR, P. J. & GELDER, M. G. (1973) Plasma levels of luteinizing hormone and of immunoreactive oestrngens (oestradiol) in anorexia nervosa response to clomiphene citrate. Psychol. Med. 3, 495-501.
BEUMo~rr, P. J. V., GEORGE, G. C. W., ~MSTONE, B. L. & Vn~1~, A. I. (1976) Body weight and the pituitary response to hypothalamic releasing hormones in patients with anorexia nervosa. J. clin. Endocr. Metab. 43, 487-496.
BIvE~s, C. H., LEBOVITZ, H. E. & FELDMAN, J. M. (1973) Inhibition of hypoglycemia-induced growth hormone secretion by the serotonin antagonists cyproheptadine and methysergide. New Engl. Y. Med. 289, 236-239.
BLACKARD, W. G. & HEIDINGSFELDER, S. A. (1968) Adrenergie receptor control mechanism for growth hormone secretion. J. clin. Invest. 47, 1407-1414.
BLACKARD, W. G. & HUSH,L, G. J. (1970) Stimulatory effects of exogenous catecholamines on plasma HGH concentrations in presence of beta-adrenergic blockade. Metabolism 19, 547-554.
BOYAR, R. M., KATZ, J., FINKELSTEIN, J. W., KAPEN, S., WEINER, H., WEITZMAN, E. D. & HELLMAN, L. (1974) Anorexia nervosa: immaturity of the 24-hr luteinizing hormone secretory pattern. New Engl. J. Med. 291, 86!-865.
BOYD, A. E., LEBOVITZ, H. E, & PFEIFFER, B. (1970) Stimulation of human growth hormone secretion by L-dopa. New Engl. J. Med. 283, 1425-1429.
BRASEL, J. A. (1973) Review of findings in patients with emotional deprivation. In Endocrine Aspects of Malnutrition. Marasmus, Kwashiorkor and Psychosocial Deprivation, L. I. Gardner and P. Amacher (Eds.), pp. 115-127. Kroc Foundation, Santa Ynez, California.
BRAZEAU, P., VALE, W., BURGUS, R., LING, N., BUTCHER, M., RIVIER, J. & GUILLEMIN, R. (1973) Hypo- thalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science 179, 77-79.
BROWN, G. M. (1976) Endocrine aspects of psychosocial dwarfism. In Hormones, Behavior, and Psycho- pathology, E. J. Sachar (Ed.), pp. 253-261. Raven Press, New York.
BROWN, G. M. & I~tCrILIN, S. (1972) Psychologic and neural regulation of growth hormone secretion. Psychosom. Med. 34, 45-61.
BROWN, G. M. & KIRPALANI, S. (1975) A critical review of the clinical relevance of growth hormone and its measurement in the nuclear medicine laboratory. Semin. Nuclear Medicine 5, 273-285.
BROWN, W. A. & HENINGER, G. (1975) Cortisol, growth hormone, free fatty acids and experimentally evoked affective arousal. Am. J. Psychiat. 132, 1174-1176.
BROWN, W. A. & HENINGER, G. (1976) Stress-induced growth hormone release: psychologic and physio- logic correlates. Psychosom. Med. 38, 145-147.
BROWN, G. M., SC~tALCH, D. S. & REICHLIN, S. (1971a) Patterns of growth hormone and cortisol responses to psychological stress in the squirrel monkey. Endocrinology 88, 956-963.
BROWN, G. M., SCI-tALCH, D. S. & R~CHLIN, S. (1971b) Hypothalamie mediation of growth hormone and adrenal stress response in the squirrel monkey. Endocrinology 89, 694--703.
BROWN, G. M., VALADKA, M. & SEe, oIL, J. (1975) Effect of group housing and gentling on resting levels of prolactin, growth hormone and corticosterone. Psychosom. Med. 37, 83 (abstract).
BRowN, W. A., KRIEGER, D. T., VAN WOERT, M. H. & A~nA~, L. M. (1974) Dissociation of growth hormone and cortisol release following apomorphine. J. clin. Endocr. Metab. 38,1127-1130.
BROWN, G. M., GARVrNZEL, P. E., WARSH, J. J. & STANCER, H. C. (1976) Effect of carbidopa on prolactin, growth hormone and cortisol secretion in man. J. clin. Endocr. Metab. 43, 236-239.
BROWN, G. M., GASrn~r~L, P. E., JEUNIEWIC, N., MOLDOFSKY, H. & STANCER, H. C. (1977) Endocrine profiles in anorexia nervosa. In Anorexia Nervosa, R. A. Vigersky (Ed.), pp. 123-135. Raven Press, New York.

150 GREGORY M. BROWN et al.
BURDAY, S. Z., FINE, P. H. & SCHALCH, D. S. (1968) Growth hormone secretion in response to arginine infusion in normal and diabetic subjects: relationship to blood glucose levels. Jr. Lab. clin. Med. 71, 897-911.
BURROW, G. N., MAY, P. B., SPAULDING, S. W. & DONABEDIAN, R. K. (1977) TRH and dopamine inter- actions affecting pituitary hormone secretion. J. clin. Endocr. Metab. 45, 65-72.
CARROLL, B. J. (1972a) The hypothalamic-pituitary axis: functions, control mechanisms and method of study. In Depressive Illness: Some Research Studies, B. Davies, B. J. Carroll and R. M. Mowbray (Eds.), pp. 23-68. Charles C. Thomas, Springfield, Ill.
CARROLL, B. J. (1972b) Control of plasma cortisol level in depression: studies with the dexamethasone suppression test. In Depressive Illness: Some Research Studies, B. Davies, B. J. Carroll and R. M. Mow- bray (Eds.), pp. 87-148. Charles C. Thomas, Springfield, Ill.
CASPER, R. C., DAVIS, J. M. & PANDEY, G. N. (1977) The effect of nutritional status and weight changes on hypothalamic function tests in anorexia nervosa. In Anorexia Nervosa, R. A. Vigersky (Ed.), pp. 137-147. Raven Press, New York.
CERVArCrES, P., LAL, S., SMITH, F. & GUYDA, H. (1977) Dopaminergic function in two patients with catalepsy. Acta psychiat, neuroL scand. 55, 214-219.
CI-IAMaERS, J. W. & BROW'N, G. M. (1976a) Stimulation of rhesus monkey GH release: arginine vs plasma volume expansion. J. olin. endocr. Metab. 42, 169-172.
CHAMBERS, J. W. & BROWN, G. M. (1976b) Neurotransmitter regulation of growth hormone and ACTH in the rhesus monkey: effects of biogenic amines. Endocrinology 98, 42.0-428.
DAUGHADAY, W. H., PmLLIPS, L. S. & HERINOTON, A. C. (1976) Regulation of somatomedin generation. In Growth Hormone and Related Peptides, A. Pecile and E. E. Miiller (Eds.), pp. 169-177. Exeerpta Medica, Amsterdam.
EHLE, A. L., MASON, J. W. & PEm,UN~TON, L. L. (1977) Plasma growth hormone and cortisol changes following limbie stimulation in conscious monkeys. Neuroendocrinology 23, 52-60.
EHRENSING, R. H., KASTIN, A. J., SCI',ALCH, D. S., FRIESEN, H. G., VARGAS, J. R. & SCHALLY, A. V. (197 4) Affective state and thyrotropin and prolaetin responses after repeated injections of TRH in depressed patients. Am. d. Psychiat. 131,714-718.
ELLIS, S., GRINDELAND, R. E., REILLY, T. J. & YANG, S. H. (1976) Studies on the nature of plasma growth hormone. In Growth Hormone and RelatedPeptides, A. Pecile and E. E. Miiller (Eds.), pp. 75-83. Excerpta Medica, Amsterdam.
ENDO, M., ENDO, J., NISHIKUBO, M., YAMAGUCHI, T. & HATOTANI, N. (1974) Endocrine studies in depression. In Psychoneuroendocrinology, N. Hatotani (Ed.), pp. 22-31. S. Karger, Basel.
ETTIGI, P. G. & BROWN, G. M. (1977) Psychoneuroendocrinology of affective disorder: an overview. Am. J. Psychiat. 134, 493-501.
ETTIGI, P., NAIR, N. P. V., LAL, S., CERVANTES, P. & GUYDA, H. (1976) Effect of apomorphine on growth hormone and prolactin secretion in schizophrenic patients, with or without oral dyskinesia, withdrawn from chronic neuroleptic therapy, d. Neurol. Neurosurg. Psychiat. 39, 870-876.
FELOMANN, J. & BROWN, G. M. (1975) Cortisol and growth hormone responses to electric shock and avoidance conditioning in the rhesus monkey. First Pacific Congress o f Psychiatry, The Australian and New Zealand College o f Psychiatrists with the American Psychiatric Association, No. 118 (abstract).
FELDMANN, J. & BROWN', G. i . (1976) Endocrine responses to electric shock and avoidance conditioning in the rhesus monkey: cortisol and growth hormone. Psychoneuroendocrinology 1, 231-242.
FERLAND, L., LABRIE, F., JomN, M., ARIMURA, A. & SCHALLY, A. V. (1976) Physiological role of somato- statin in the control of growth hormone and thyrotropin secretion. Biochem. biophys. Res. Commun. 68, 149-156.
FINKELSTEIN, J. W., ROFFWARO, H. P., BOYAR, R. M., KREAM, J. & HELLMAN, L. (1972) Age-related change in the twenty-four-hour spontaneous secretion of growth hormone, d. clin. Endocr. Metab. 35, 665-670.
FRANKEL, R. J. • JENKINS, J. S. (1975) Hypothalamic pituitary function in anorexia nervosa. Acta Endocr. (Kbh) 78, 209-221.
FROHMAN, L. A., BERNARDIS, L. L., BUREK, L., MARAN, J. W. & DHARIWAL, A. P. S. (1971) Hypothalamic control of growth hormone secretion in the rat. The Second International Symposium on Growth Hormone, Milan, Italy, Abstract No. 21, p. 12.
FURLONG, F. W., BROWN, G. M. & BEECHING, M. F. (1976) Thyrotropin-releasing hormone: differential antidepressant and endocrinological effects. Am. d. Psychiat. 133, I 187-1190.
GARFINKEL, P. E., BROWN, G. M., STANCER, H. C. & MOLDOFSKY, H. (1975) Hypothalamic-pituitary function in anorexia nervosa. Archs gen. Psychiat. 32, 739-744.

PSYCHOENDOCRINOLOGY AND GROWTH HORMONE 151
GXORDANO, G., MAgUOIO, M., ML~UTO, F. & BAmUSCA, T. (1971) Somatotropin increment in grave emacia- tion: preliminary results. Folia endocr. 24, 305-315.
GUCK, S. M., ROTH, J., YALOW, R. S. & BEaSON, S. A. (1965) The regulation of growth hormone secretion. Recent Prog. Horm. Res. 21, 241-283.
GREE~, W. A., CONgON, G., SCHALCH, D. S. & SCHgEL~,~R, B. F. (1970) Psychologic correlates of growth hormone and adrenal secretory responses of patients undergoing cardiac catheterization. Psychasom. Med. 32, 599-614.
GRUEN, P. H., SACHAR, E. J., ALTMAN, N. & SASSIN, J. (1975) Growth hormone responses to hypoglyce~nia in post-menopausal depressed women. Archs gen. Psychiat. 32, 31-33.
HOLLISTER, L. E., bEgGER, P., O~LE, F. L., ARNOLD, R. C. & JOHNSON, A. (!974) Prot/relin (TRH) in depression. Archs gen. Psychiat. 31, 468--470.
ILUO, R. (1972) Wachstumshormon-untersuchungen bei 225 Kindern mit Minderwuchs. Schweiz. reed. Wschr. 102, 760-765.
KAPLAN, S. L., AnR.~S, C. A. L., BELL, J. J., CONTE, F. A. & GRUIC,~CH, M. M. 0968) Growth and growth hormone. I. Changes in serum level of growth hormone following hypoglycemia in 134 children with growth retardation. Pediat. Res. 2, 43-63.
KASTIN, A. J., EH~NSlNO, K. H., SCHALCH, D. S. & ANVEgSON, M. S. 0972) Improvement in mental depression with decreased thyrotropin response after administration of thyrotropin-releasing hormone. Lancet 2, 740-742.
KATO, Y., Cm,ARA, K., OHGO, S. & IMURA, H. (1974) Effects of hypothalamic surgery and somatostatin on chlorpromazine-induced growth hormone release in rats. Endocrinology 95, 1608-1613.
KONCZ, L., SOELDNER, S., bALODL'~OS, M. C., BODEN, G., GLEASON, R. E. & YOtmGER, D. (1973) Human growth hormone secretion after double stimulation with arginine in normal and insulin-dependent diabetic women. Diabetes 22, 694-705.
KOSTYO, J. L., BXLLS, J. B., REAGAN, C. R., RUDMAN, D. & WILH~LMI, A. E. (1976) The nature of fragments of human growth hormone produced by plasmin digestion. In Growth Hormone and Related Peptides, A. Pecile and E. E. Mtiller (Eds.), pp. 33--40. Excerpta Mextica, Amsterdam.
KRIEGER, I. (1973) Endocrine and nutrition in psychosocial deprivation in the U.S.A.: comparison with growth failure due to malnutrition on an organic basis. In Endocrine Aspects of Malnutrition. Marasmus, Kwashiorkor and Psychosociai Deprivation, L. I. Gardner and P. Amacher (Eds.), pp. 129-162. Kroc Foundation, Santa Ynez, California.
KRXEGER, I. & GOOD, M. H. 0970) Adrenocortical and thyroid function in the deprivation syndrome: comparison with growth failure due to undernutrition, congenital heart disease, or prenatal influences. Am. J. Dis. Child. 120, 95-102.
K.gULICH, L., ILLNER, P., FAWCETT, C. P., QUUADA, M. & McC~u~t~, S. M. (1972) Dual hypothalamic regulation of growth hormone secretion. In Growth and Growth Hormone, A. Pecile and E.E. Miiller (Eds.), pp. 306-316. Excerpta Medica, Amsterdam.
LAL, S., GUYDA, H. & BIKADOgOFF, S. (1977) Effect of methysergid¢ and pimozide on apomorphine- induced growth hormone secretion in men. J. clin. Endocr. Metab. 44, 766-770.
LAL, S., DE LA VEGA s C. E., SOURK~, T. L. & FRIESEN, H. G. (1972) Effect of apomorphine on human growth hormone secretion. Lancet 2, 661.
LAL, S., TOLIS, G., MARTIN, J. B., bROWN, G. M. & GUYDA, H. (1975) EffeCt of clonidine on growth hormone, prolactin, luteinizing hormone, follicle-stimulating hormone and thyroid-stimulating hormone in the serum of normal men. J. clin. Endocr. Metab. 41, 827-832.
LANDON, J., GREENWOOD, F. C., STAMP, T. C. B. & WYNN, V. (1966) The plasma sugar, free fatty acid, cortisol, and growth hormone response to insulin, and the comparison of this procedure with other tests of pituitary and adrenal function. II. In patients with hypothalamio or pituitary dysfunction or anorexia nervosa. J. clin. Invest. 45, 437-449.
LEWIS, U. J., S1NGH, R. N. P., P~TERSON, S. M. & V~DERL~N, W. P. (1976) Human growth hormone: a family of proteins. In Growth Hormone and Related Peptides, A. Pecile and E. E. Mfiller (Eds.), pp. 64-74. Excerpta Medica, Amsterdam.
LI, C. H. & BEWLEY, T. A. (1976) Studies on plasmin-modified human growth hormone and its fragments. In Growth Hormone and RelatedPeptides, A. Pecile and E. E. Miiller (Eds.), pp. 14-32. Excerpta Medica, Amsterdam.
LUFT, R., CERASI, E., MADISON, L. L., VON EULER, V. S., DELLA CASA, L. & ROOVETE, A. (1966) Effect era small decrease in blood-glucose on plasma-growth-hormone and urinary excretion of catecholamines in man. Lancet 2, 254-256.

152 GREGORY M. BROWN et al.
Lt~DB~R6, P. O., WALINDER, J., WERNP.R, I. & WIDE, L. (1972) Effects of thyrotrophin-releasing hormone on plasma levels of TSH, FSH, LH and OH in anorexia nervosa. Eur. J. din. Invest. 2, 150-153.
M~DA, K., KATO, Y., OHC, O, S., C ~ A , K., Yosm~lOTO, Y., YAMAOUCm, N., Ktmo~mu, S. & IitrRA, H. (1975) Growth hormone and prolactin release after injection of thyrotropin-releasing hormone in patients with depression. J. clin. Endocr. Metab. 40, 501-505.
MARSHALL, J. C. & F'Rm~S, T. R. (1971) Amenorrhoea in anorexia nervosa: assessment and treatment with clomiphene citrate. Br. reed. J. 4, 590-592.
MARTIN, J. B. (1972) Plasma growth hormone (GH) response to hypothalamic or extra-hypothalamic electrical stimulation. Endocrinology 91, 107-115.
MARTIN, J. B. (1973a) Neural regulation of growth hormone secretion, medical progress report. New Engl. J. Med. 288, 1384--1393.
MARTtN, J. B. (1973b) Functions of the hypothalamus and amygdala in regulation of growth hormone secretion. Trans. Am. neurol. Ass. 98, 229-232.
MARTrN, J. B. (1973c) The role of hypothalamic and extrahypothalamic structures in the control of growth hormone secretion. In Advances in Human Growth Hormone Research, S. Raiti (Ed.), pp. 223-255. National Institutes of Health, Bethesda.
MARTIN, J. B., KOm'OR, J. & MEAD, P. (1973) Plasma GH responses to hypothalamic, hippocampal and amygdaloid electrical stimulation: effects of variation in stimulus parameters and treatment with a- methyl-p-tyrosine (a-MT). Endocrinology 92, 1354-1361.
MASON, J. W. (1975) Emotion as reflected in patterns of endocrine integration. In Emotions--Their Para- meters and Measurement, L. Levi (Ed.), pp. 143-181. Raven Press, New York.
MAson, J. W., MA~ER, J. T., HARTLEY, L. H., MOUSEY, E. H., PERLOW, M. J. & JONES, L. G. (1976) Select- ivity of corticosteroid and catecholamine responses to various natural stimuli. In Psychopathology of Human Adaptation, G. Serban (Ed.), pp. 147-171. Plenum Publishing Corporation, New York.
McARTmm, J. W., O'LouG~IN, K. M., BECTINS, I. Z., JOHNSON, L., HOURIKAN, .I. & ALONSO, C. (1976) Endocrine studies during the refeeding of young women with nutritional amenorrhea and infertility. Mayo Clin. Prec. 5I, 607-616.
MELTZER, H. Y., SACHAR, E. J. & FRANTZ, A. G. (1974) Serum prolactin levels in unmedicated schizo- phrenic patients. Archsgen. Psychiat. 31,564-569.
MI",'ABO, S., HmADA, T., ASATO, T., MIzuSmMA, N. & UENO, K. (1976) Growth hormone and cortisol responses to psychological stress: comparison of normal and neurotic subjects. J. clin. Endocr. Metab. 42, 1158-1162.
PALMER, R. L., CRISP, A. H., MAC~I,a~ON, P. C. B., FRANKLIN, M., BONNAR, J. & WHEELER, M. (1975) Pituitary sensitivity to 50 ~g LH]FSH-RH in subjects with anorexia nervosa in acute and recovery stages. Br. med.J. 1,179-182.
PANDEY, G. N., GARVER, D. L., HENGEVELD, C., ERICKSEN, S., GOSENFELD, L. & DAVIS, J. M. (1977) Postsynaptic supersensitivity in schizophrenia. Am. J. Psychiat. 134, 518-522.
PIMSTONE, B. L., BECKER, D. J. & HANSEN, J. D. L. (1972) Human growth hormone in protein-calorie malnutrition. In Growth and Growth Hormone, A. Pecile and E. E. Mfiller (Eds.), pp. 389--401. Excerpta Medica, Amsterdam.
PIMSTONE, B. L., BECKER, D. J. & HANSEN, J. D. L. (1973) Human growth hormone and sulphation factor in protein-calorie malnutrition. In Endocrine Aspects of Malnutrition, Marasmus, Kwashiorkor and Psychosocial Dwarfism, L. I. Gardner and P. Amacher (Eds.), pp. 73-90. Kroc Foundation, Santa Ynez, California.
PONTIROLI, A. E., CASTEGNARO, E., VETTARO, M. e., VIBER'II, G. C. & POZZA, G. (1977) Stimulatory effect of the dopa-decarboxylase inhibitor Re 4-4602 on prolactin release; inhibition by L-dopa, metergoline, methysergide and 2-Br-a-Ergocryptine. Acta Endocr. 84, 36-44.
POWELL, G. E., HOPWOOD, N. J. & BARRATT, E. S. (1973) Growth hormone studies before and during catch-up growth in a child with emotional deprivation and short stature. J. clin. Endocr. Metab. 37, 674-679.
POWELL, G. E., BRASI/L, J. A., RAITI, S. & BLIZZARD, R. m. (1967) Emotional deprivation and growth retardation simulating idiopathic hypopituitarism. II. Endocrinologic evaluation of the syndrome. New Engl. d. Med. 2"/6, 1279-1283.
PRANGE, A. ft., WILSON, I. C., LARA, P. P., ALLTOP, L. B. & BREESE, G. R. (1972) Effect of TRH in depression. Lancet 2, 999-1002.
REICHIJN, S. (1974) Regulation of somatotrophic hormone secretion. In Handbook of Physiology, Section 7: Endocrinology Vol. IV The Pituitary Gland and Its Neuroendocrine Control, Part 2, R. O. Greep and E. B. Astwood (Eds.), pp. 405-447. American Physiological Society, Washington.

PSYCHOENDOCRINOI.DGY AND GROWTH HoP~4o~ 153
ROSE, R. M. & HURST, M. W. (1975) Plasma cortisol and growth hormone responses to intravenous catheterization. J. Human Stress 1, 22-36.
SACHAR, E. J. (1975) Neuroendocrine abnormalities in depressive illness. In Topics in Psychoendocrinology, E. J. Sachar (Ed.), pp. 135-156. Grune and Stratton, New York.
SACr~R, E. J., F r t~TEIN, J. & HELLMAr~, L. (1971) Growth hormone responses in depressive illness. Archs gen. Psychiat. 25, 263-269.
SACX~R, E. J., HELLMAN, L., ROrFWARG, H. P., HALPERN, F. S., FUKUSHIMA, D. K. & GALLAGHER, T. F. (1973) Disrupted 24-hr patterns of cortisol secretion in psychotic depression. Arehs gen. Psyehiat. 28, 19-24.
SACHAR, E. J., ALTMAN, N., GRUEN, P. H., GLASSMAN, A., HALPERN, F. S. & SASSIN, J. (1975) Human growth hormone response to levodopa. Relation to menopause, depression and plasma dopa concen- tration. Archs gen. Psyehiat. 32, 502-503.
ScI~a.CH, D. S. (1967) The influence of physical stress and exercise on growth hormone and insulin secretion in man. J. Lab. elin. Med. 69,256-269.
SeEMAN, P., LEE, T., CHAU-WONO, M. & WONG, K. (1976) Antipsychotic drug doses and neuroleptic/ dopamine receptors. Nature 261, 717-719.
SEGGIE, J. & BROWN, G. M. (1975) Stress response patterns of plasma corticosterone, prolactin and growth hormone. Can. J. Physiol. Pharmacol. 53, 629-637.
SEc~3m, J. & BROWN, G. M. (1976) Coping with stress: parallelism between the effects of septal lesions on growth hormone and corticosterone levels. Biol. Psychiat. 11, 583-597.
SEC~3m, J., UHLXR, I. & BROWN, G. M. (1974a) Adrenal stress responses following septal lesions in the rat. Neuroendocrinology 16, 225-236.
S~3XE, J., SHAw, B., UHLXR, I. & BROWN, G. M. (1974b) Baseline 24-hr plasma corticosterone rhythm in normal, sham-oporated and septally lesioned rats. Neuroendocrinology 15, 51-61.
SI~EHAN, H. L. & Stnv~RS, V. K. (1949) The syndrome of hypopituitarism. Q. Y. Med. 18, 319-378. StmRMAN, B. M., HAL~, K. A. & ZA~tJDIO, R. (1975) LH and FSH response to gonadotropin-releasing
hormone in anorexia nervosa: effect of nutritional rehabilitation. J. clin. Endocr. Metab. 41, 135-142. SIMON, M. L. & GEOROE, R. (1975) Diurnal variations in plasma corticosterone and growth hormone as
correlated with regional variations in norepinephrine, dopamine and serotonin content of rat brain. Neuroendocrinology 17, 125-138.
S~rti, G. P. & ROOT, A. W. (1971) Dissociation of changes in growth hormone and adrenocortical hormone levels during brain stimulation of monkeys. Neuroendocrinology 8, 235-244.
SPITZ, I., GO~mN, B. & RAmNOWITZ, D. (1972) Growth hormone release in man revisited: spontaneous vs stimulus-initiated tides. In Growth and Growth Hormone, A. Pecile and E. E. Mfiiler (Eds.), pp. 371-381. Excerpta Medica, Amsterdam.
STOKES, P. E. (1972) Studies on the control of adrenocortical function in depression. In Recent Advances in the Psychobiology of Depressive Illness, T. A. Williams, M. M. Katz and J. A. Shield (Eds.), pp. 199-220. U.S. DHEW Publication No. 70-9053.
TAKJa-L~m, S., KONDO, H., YOSm~nYRA, M. & Ocm, Y. (1974) Growth hormone responses to administration of L-5-hydroxytryptophan (L-5-HTP) in manic-depressive psychoses. In Psychoneuroendocrinology, N. Hatotani (Ed.), pp. 32-38. S. Karger, Basel.
UNOERSrEDT, U. (1971) Stereotaxic mapping of the monoamine pathways in the rat brain. Acta physiol. scand. Suppl. 367, 1-48.
WAKeUNG, A., MARSHALL, J. C., BEARDWOOD, C. J., DE SOUZA, V. F. A. & RUSSELL, G. F. M. (1976) The effects of elomiphene citrate on the hypothalamic-pituitary-gonadal axis in anorexia nervosa. PsychoL Med. 6, 371-380.
WARREN, M. P., JEWELEWXCZ, R., DYRENrURTH, I., ANS, R., KHALAF, S. & VANDE WmLE, R. L. (1975) The significance of weight loss in the evaluation of pituitary response to LH-RH in women with secondary amenorrhea. Y. clin. Endocr. Metab. 40, 601-611.
YALOW, R. S., VARSANo-Aru~M, N., ECH~r,rOL% E. & BERSON, S. A. (1969) HGH and ACTH secretory responses to stress. Horm. Metab. Res. 1, 3-8.