proteomic analysis of set-binding proteins
TRANSCRIPT

RESEARCH ARTICLE
Proteomic analysis of SET-binding proteins
Jorge Vera1*, Josep M. Estanyol2*, Nuria Canela2, Franc Llorens3, Neus Agell1,Emilio Itarte3, Oriol Bachs1 and Montserrat Jaumot1**
1 Departament de Biologia Cel?lular i Anatomia Patològica, Facultat de Medicina,Universitat de Barcelona, Barcelona, Spain
2 Unitat de Proteòmica, Serveis Científico-tècnics, Universitat de Barcelona, Spain3 Departament de Bioquímica i Biologia Molecular, Universitat Autònoma de Barcelona, Bellaterra, Spain
The protein SET is involved in essential cell processes such as chromatin remodeling, apoptosisand cell cycle progression. It also plays a critical role in cell transformation and tumorogenesis.With the aim to study new SET functions we have developed a system to identify SET-bindingproteins by combining affinity chromatography, MS, and functional studies. We prepared SETaffinity chromatography columns by coupling the protein to activated Sepharose 4B. The pro-teins from mouse liver lysates that bind to the SET affinity columns were resolved with 2-DE andidentified by MS using a MALDI-TOF. This experimental approach allowed the recognition of anumber of SET-binding proteins which have been classified in functional clusters. The identifi-cation of four of these proteins (CK2, eIF2a, glycogen phosphorylase (GP), and TCP1-b) wasconfirmed by Western blotting and their in vivo interactions with SET were demonstrated byimmunoprecipitation. Functional experiments revealed that SET is a substrate of CK2 in vitroand that SET interacts with the active form of GP but not with its inactive form. These data con-firm this proteomic approach as a useful tool for identifying new protein–protein interactions.
Received: June 22, 2006Revised: October 23, 2006
Accepted: November 11, 2006
Keywords:
2-DE / Affinity chromatography / MALDI-TOF MS / SET / SET-binding proteins
578 Proteomics 2007, 7, 578–587
1 Introduction
SET is a phosphoprotein of 37 kDa that was first identifiedas a CAN-fusion gene in an acute undifferentiated leukemia[1]. It was further described as a potent inhibitor of theprotein phosphatase PP2A [2], which is involved in severalcellular processes such as metabolism, proliferation, differ-entiation, and DNA transcription and replication [3]. The
fusion protein SET-CAN does not efficiently regulate PP2Aactivity and promotes an abnormal accumulation of pro-teins in the nucleus due to an aberrant localization of thehCRM1 export factor [4]. SET has also been found asso-ciated with PP2A and the HRX leukemic fusion proteinpresent in myeloid cell extracts [5]. It has also been reportedthat SET activates the 17, 20 lyase activity of cytochromeP450c17 by inhibiting PP2A activity [6]. Increased expres-sion of the SET protein has been observed during renaldevelopment, in cells of the Wilms’ tumor [7], hepatomas,and during liver regeneration [8].
There is a lot of evidence to relate the SET protein withthe regulation of cell cycle progression. It has been reportedthat SET directly binds to p21CIP1 and reverts the inhibitoryeffect of p21CIP1 on cyclin E-cdk2 kinase activity [9]. Moreover,SET also binds to cyclin B [10] and inhibits cyclin B-cdk1 ac-
Correspondence: Professor Oriol Bachs, Departament de Biolo-gia Cel? lular, Facultat de Medicina, Universitat de Barcelona,Casanova 143, 08036-Barcelona, SpainE-mail: [email protected]: 134-93-402-19-07
Abbreviations: GAPDH, glyceraldehyde 3-phosphate dehydro-genase; GP, glycogen phosphorylase; hnRNPA2/B1, hetero-geneous nuclear ribonucleoprotein A2/B1; INHAT, inhibitor ofhistone acetyltransferases; JDP1A, jumonji domain-containingprotein 1A
* Both these authors have contributed equally to this work.** Additional corresponding author: Dr. Montserrat Jaumot,
E-mail: [email protected]
DOI 10.1002/pmic.200600458
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Proteomics 2007, 7, 578–587 Cell Biology 579
tivity, thus, regulating G2/M transition [11]. SET is also ableto interact with the cdk5 regulatory subunit p35nck5a and as aconsequence of this interaction the activity of cdk5-p35nck5a isincreased. Cdk5 plays a critical role in neuronal differentia-tion, cytoskeleton dynamics, neuronal degeneration, and celldeath [12]. SET might also negatively modulate the MEK/ERK pathway [13]. In general, the levels of expression of SETremain constant through the cell cycle [14].
SET is mainly located in the cell nucleus but is also foundin the ER where it is a component of the SET complex. Thisprotein complex comprises the endonuclease Ape1, theDNA-binding protein HMG-2, the transcriptional regulator,and nucleoside diphosphate kinase NM23-H1 and the in-hibitor of PP2A and tumor suppressor protein pp32. Thiscomplex is involved in the process of apoptosis induced byGranzyme B [15–17].
SET is an important modulator of chromatin remodeling.It is a component of the INHATcomplex (inhibitor of histoneacetyltransferases) which binds to histones and blocks theiracetylation by preventing the access of the acetylases p300/CBP and PCAF [18]. INHAT is constituted by TAF-1a, SET/TAF-1b, and pp32 subunits. It has recently been describedthat INHAT also binds to histone deacetylase classes I and II[19], thus functioning as a transducer of a transcriptionalrepressor signal. Likewise, INHAT prevents transcription byblocking the DNA dimethylation [20]. Nevertheless, SETmight function as a transcriptional activator by altering his-tone–DNA interaction [21], by binding to acetyltransferases[22] and by acting as a histone chaperone [23].
It has also been reported that SET, pp32, TAF1-a, andAPRIL, interact with the protein HuR which stabilizes themRNA by directly binding the AU-rich elements of the30UTR zones [24].
In summary, SET is a protein involved in essential cel-lular processes and also plays a critical role in cell transfor-mation and tumorigenesis. A way to study the new functionsof a protein is to identify its interacting proteins. With thisaim we have developed a system to identify SET-bindingproteins combining affinity chromatography, MS and func-tional studies. In the present work we have identified 36 SET-binding proteins and we have characterized the functionalrelevance of some of these interactions.
2 Materials and methods
2.1 Sample preparation
A mouse liver (1–1.5 g) was homogenized in 5 mL of bufferA (50 mM HEPES-KOH, pH 7.6, 50 mM KCl, 1 mM EGTA,1 mM MgCl2, 0.1% Triton X-100) containing 0.5 mg/mLaprotinin, 10 mg/mL leupeptin, and 1 mM PMSF. Homo-genates were filtered through four sheets of cheesecloth andsedimented twice at 14 0006g for 15 min each. The proteincontent of the supernatant was measured by the method ofBradford.
2.2 Proteins and plasmids
Purified glycogen phosphorylases (GPs) a and b and b-caseinwere obtained from Sigma. The expression vector pGEX-KG-SET was generated as previously described [11]. Recombi-nant human CK2 a (monomer) and CK2 a/b tetramer wereprovided by Dr. Itarte (Barcelona).
2.3 Protein expression and purification
The Codon Plus® strain of Escherichia coli was transformedwith the vector pGEX-KG-SET. Expression of the recombi-nant GST-SETprotein was induced with 1 mM isopropyl-b-D-thiogalactopyranoside for 16 h at 377C. Cell pellets werelysed in NETN buffer (20 mM Tris-HCl, pH 8.0, 100 mMNaCl, 1 mM EDTA, 0.5% IGPAL, 0.5 mg/mL aprotinin,10 mg/mL leupeptin, and 1 mM PMSF). After sonication, celllysates were centrifuged and a supernatant containing GST-SET was obtained. The supernatant was then incubated withglutathione-sepharose beads (Amersham Biosciences) for2 h at 47C, spinned and washed twice with NETN buffer andonce with PBS. Next, beads were resuspended in 1 mL ofPBS and a thrombin protease digestion was performed toseparate GST from SET according to the manufacturer’sinstructions (Sigma).
2.4 SET-Sepharose 4B affinity chromatography
To prepare the SET-Sepharose 4B columns, 3–5 mg of puri-fied SET protein was coupled to 1 mL of CNBr-activatedSepharose 4B (Amersham Biosciences), following the man-ufacturer’s instructions. Twenty milligrams of the mouseliver lysate was then loaded on the SET-Sepharose 4B orSepharose 4B control columns. After picking up the flowthrough fraction, the columns were washed with 200volumes of buffer A. Then the bound proteins were elutedwith 2 mL of the same buffer but containing 1 M KCl insteadof 50 mM KCl; or with consecutive elutions of 2 mL buffer Acontaining increasing KCl concentrations (150, 300, 500, or1000 mM). Each elution was separated from the next one by5 mL wash of the same buffer.
2.5 Gel electrophoresis and Western blotting
The protein samples were analyzed by SDS-PAGE. The gelswere stained with silver nitrate [25] or transferred ontoImmobilon-P membranes (Millipore). The sheets were thenincubated with TBST (20 mM Tris-HCl, pH 7.5, 150 mMNaCl, 0.05% Tween-20) containing 5% defatted milk powderfor 1 h at room temperature. Western blots were probed for1 h at room temperature with the following antibodies: anti-SET (I2a-PP2A, AbCam ab1183, 1:100 dilution), anti-eukaryotic initiation factor 2a (eIF2a (C-20), Santa Cruz sc-7629, 1:200 dilution), anti-TCP-1b (Stressgen CTA-200,1:1000 dilution), anti-GP muscle form (kindly provided byDr. Guinovart, Barcelona, dilution 1:5000), anti-CK2
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

580 J. Vera et al. Proteomics 2007, 7, 578–587
(Upstate 06-873, 2 mg/mL), anti-CDK4 (H-303, Santa Cruz sc-749, 1:200 dilution), anti-CDK2 (UBI 06-505, 2 mg/mL), anti-cyclin D1 (72-13G, Santa Cruz sc-450, 1:200 dilution), anti-cyclin D2 (C-17, Santa Cruz sc-181, 1:200 dilution), anti-cyclin D3 (C-16, Santa Cruz sc-182, 1:200 dilution), anti-I1PP2A (PHAPI/PP32) (Stratagene B13006, 1:5000 dilu-tion), or anti-heterogeneous nuclear ribonucleoprotein A2/B1 (hnRNPA2/B1) [26] antibodies. After washing twice withTBST, sheets were incubated with the corresponding HRP-coupled secondary antibody (1:4000 dilution, BioRad) for45 min at room temperature. They were washed twice withTBST, once with TBS (20 mM Tris-HCl, pH 7.5, 150 mMNaCl) and visualized by enhanced chemiluminiscence(Amersham).
2.6 2-DE
Eluates obtained from the SET-Sepharose 4B columns wereprecipitated with 10% TCA and resuspended in 2-DE lysisbuffer (9.8 M urea, 2% NP-40, 1% ampholytes pH 3–10,100 mM DTT). Samples were then analyzed by 2-DE. Briefly,a prerunning without sample was performed at 200 V during10 min, then 15 min at 300 V, and finally 20 min at 400 V.The samples were subsequently run 10 min at 500 V and 3 hat 750 V, by using the mini-2D system from BioRad tube gelswith a pH range of 3–10 of Byolites (BioRad). The IEF gelswere equilibrated 30 min at 47C in 1 mL of equilibration so-lution (65 mM Tris-HCl, pH 6.8, 2.3% SDS, 10% glycerol,100 mM DTT, and 0.002% bromophenol blue). IEF gels werestored at 2807C until they were used for the second-dimen-sional run. SDS-PAGE was used as second dimension. Gelsobtained were further stained with silver nitrate [25].
2.7 Identification of proteins by MALDI-TOF MS
The spots from 2-DE were manually sliced from the gel anddigested with 100–150 ng of trypsin (Promega) at 377C over-night, using a Montage In-Gel DigestZP (Millipore) with thestandard manufacturer protocol. The resulting peptides weredried and resuspended in 0.1% TFA. About 0.5 mL of themwere mixed with the same volume of CHCA and were ana-lyzed by a MALDI-TOF Voyager DE Pro mass spectrometer(Applied Biosystems), operated in delayed extraction reflec-tor mode at 20 kV as accelerating voltage, 90 ns of pulse delaytime, 75% of a grid voltage, and a guide wire voltage of0.005%. Spectra were accumulated for 100 laser shots andwere visualized and analyzed using Data Explorer softwareversion 4.2 (Applied Biosystems). Monoisotopic peaks wereused for PMF, using the Protein Prospector MS-Fit softwareversion 3.2 [27] and the main databases that describe themouse proteome: Swiss-Prot and the National Center forBiotechnology Information (NCBI NIH, Bethesda, MD,USA).
2.8 Immunoprecipitation
Samples were lysed in buffer B (50 mM Tris-HCl, pH 7.4,250 mM NaCl, 5 mM EDTA, 0.1% Triton X-100) contain-ing 50 mM NaF, 1 mM PMSF, 10 mg/mL leupeptin,0.5 mg/mL aprotinin, and 0.1 mM Na3VO4 for 30 min onice. Lysates were clarified by centrifugation at 10 0006gfor 10 min at 47C. The supernatants (500 mg) were incu-bated with 3 mg of anti-SET overnight at 47C, followed byincubation with protein A beads for 1 h at 47C. Afterwashing in buffer B, the immunocomplexes were sub-jected to immunoblotting with the corresponding anti-bodies.
2.9 CK2 kinase assay
To determine CK2 kinase activity, 2 pmol of recombinanthuman CK2a (monomer) or 2 pmol of CK2a/b (tetrameractivated) and SET (30 pmol) were incubated with 5 mCi ofg-(32P)-ATP (Amersham; 3.000 Ci/mmol) in 10 mL of bufferTEST (50 mM Tris-HCl, pH 7.4, 1.5 mM EGTA, 1.5 mMEDTA, 25 mM MgCl2, 25 mM b-glycerol phosphate, 1 mMDTT, and 1 mM ATP) for 30 min at 307C. Then, the reactionswere stopped by adding Laemmli sample buffer and thesamples were electrophoresed in 10% SDS-PAGE gels. Thegels were dried and the phosphorylated proteins were ana-lyzed on a PhosphoImager (Molecular Image System GS-525; BioRad). The positive control was performed usingb-casein as a substrate.
3 Results
3.1 Detection of SET-binding proteins from mouse
liver
To identify the SET-interacting proteins, we performed af-finity chromatography analysis by using SET-Sepharose 4Bcolumns and Sepharose 4B columns as a control. These col-umns were generated as described in Section 2. Then, 20 mgof mouse liver extract was loaded onto the columns and theproteins which were not retained were collected as the flow-through fraction. After washing, the proteins bound to thecolumns were eluted with a buffer containing high salt con-centration (1 M KCl). The proteins obtained in each step ofthe chromatography were then analyzed by SDS-PAGE andvisualized by CBB staining. As shown in Fig. 1A, more than20 proteins specifically interacted with the SET-Sepharose 4Bcolumn.
Next, with the aim of identifying these proteins by MS,we performed a more detailed analysis of 50 mg of the eluatesfrom the SET-Sepharose 4B column by 2-DE and silverstaining of the gels. A representative pattern, showing morethan 30 resolved spots, is shown in Fig. 1B.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
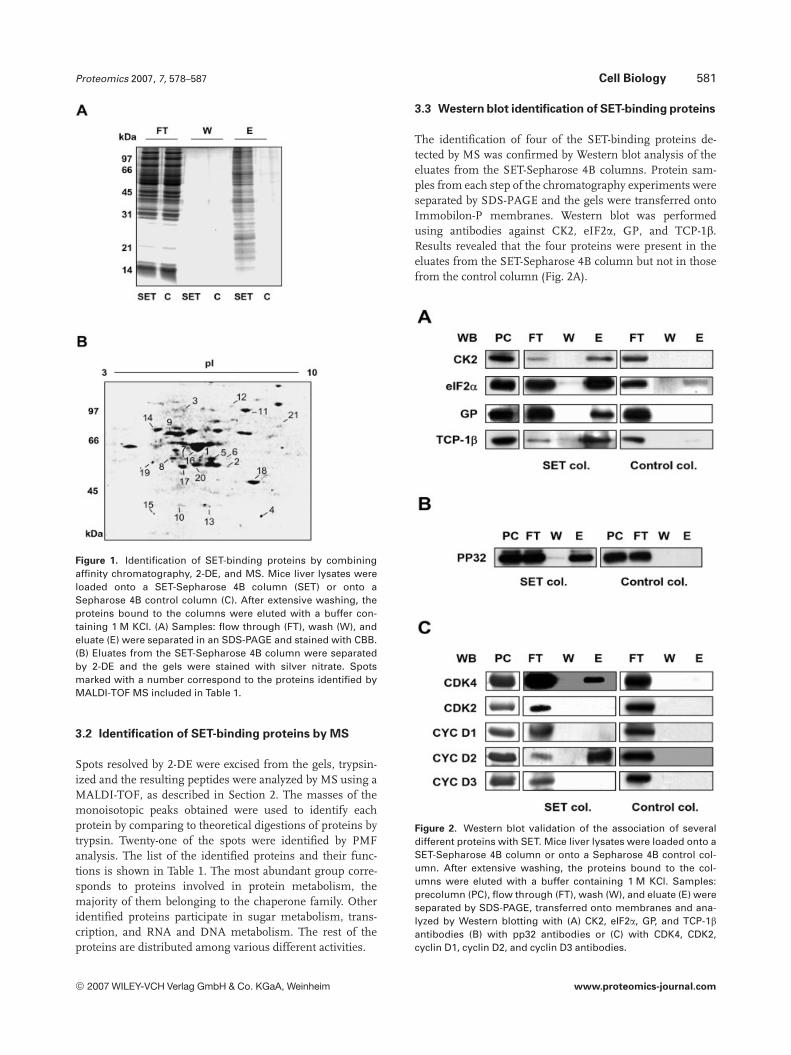
Proteomics 2007, 7, 578–587 Cell Biology 581
Figure 1. Identification of SET-binding proteins by combiningaffinity chromatography, 2-DE, and MS. Mice liver lysates wereloaded onto a SET-Sepharose 4B column (SET) or onto aSepharose 4B control column (C). After extensive washing, theproteins bound to the columns were eluted with a buffer con-taining 1 M KCl. (A) Samples: flow through (FT), wash (W), andeluate (E) were separated in an SDS-PAGE and stained with CBB.(B) Eluates from the SET-Sepharose 4B column were separatedby 2-DE and the gels were stained with silver nitrate. Spotsmarked with a number correspond to the proteins identified byMALDI-TOF MS included in Table 1.
3.2 Identification of SET-binding proteins by MS
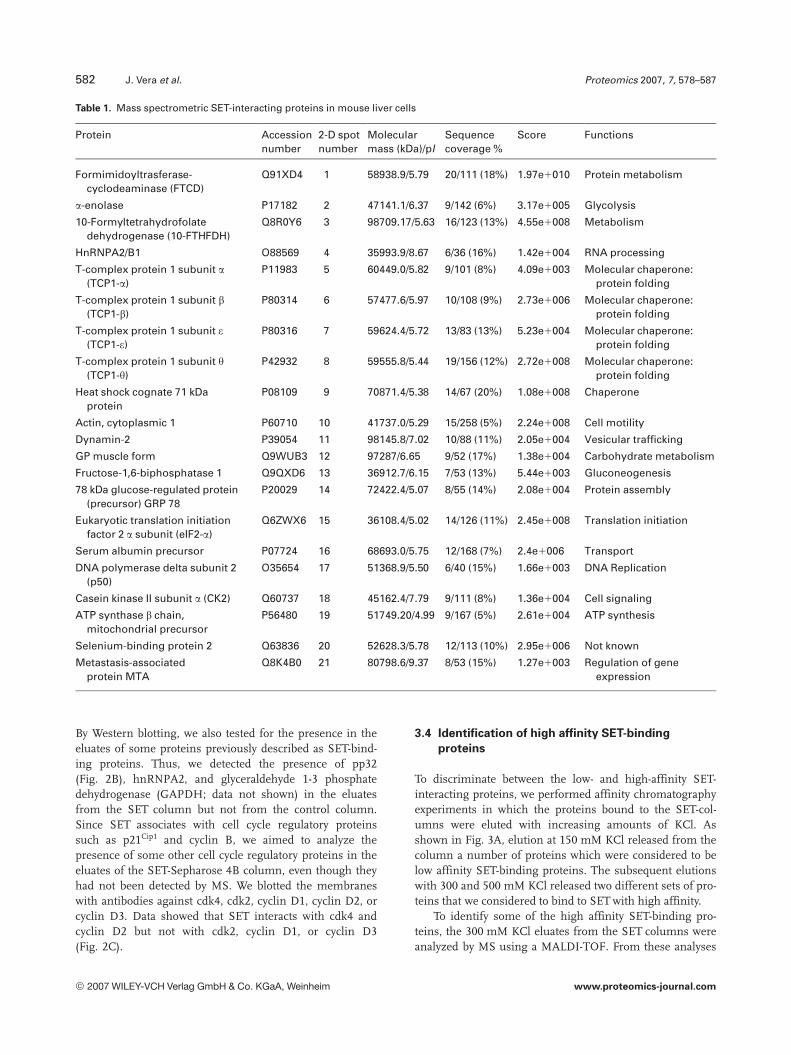
Spots resolved by 2-DE were excised from the gels, trypsin-ized and the resulting peptides were analyzed by MS using aMALDI-TOF, as described in Section 2. The masses of themonoisotopic peaks obtained were used to identify eachprotein by comparing to theoretical digestions of proteins bytrypsin. Twenty-one of the spots were identified by PMFanalysis. The list of the identified proteins and their func-tions is shown in Table 1. The most abundant group corre-sponds to proteins involved in protein metabolism, themajority of them belonging to the chaperone family. Otheridentified proteins participate in sugar metabolism, trans-cription, and RNA and DNA metabolism. The rest of theproteins are distributed among various different activities.
3.3 Western blot identification of SET-binding proteins
The identification of four of the SET-binding proteins de-tected by MS was confirmed by Western blot analysis of theeluates from the SET-Sepharose 4B columns. Protein sam-ples from each step of the chromatography experiments wereseparated by SDS-PAGE and the gels were transferred ontoImmobilon-P membranes. Western blot was performedusing antibodies against CK2, eIF2a, GP, and TCP-1b.Results revealed that the four proteins were present in theeluates from the SET-Sepharose 4B column but not in thosefrom the control column (Fig. 2A).
Figure 2. Western blot validation of the association of severaldifferent proteins with SET. Mice liver lysates were loaded onto aSET-Sepharose 4B column or onto a Sepharose 4B control col-umn. After extensive washing, the proteins bound to the col-umns were eluted with a buffer containing 1 M KCl. Samples:precolumn (PC), flow through (FT), wash (W), and eluate (E) wereseparated by SDS-PAGE, transferred onto membranes and ana-lyzed by Western blotting with (A) CK2, eIF2a, GP, and TCP-1bantibodies (B) with pp32 antibodies or (C) with CDK4, CDK2,cyclin D1, cyclin D2, and cyclin D3 antibodies.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
582 J. Vera et al. Proteomics 2007, 7, 578–587
Table 1. Mass spectrometric SET-interacting proteins in mouse liver cells
Protein Accessionnumber
2-D spotnumber
Molecularmass (kDa)/pI
Sequencecoverage %
Score Functions
Formimidoyltrasferase-cyclodeaminase (FTCD)
Q91XD4 1 58938.9/5.79 20/111 (18%) 1.97e1010 Protein metabolism
a-enolase P17182 2 47141.1/6.37 9/142 (6%) 3.17e1005 Glycolysis
10-Formyltetrahydrofolatedehydrogenase (10-FTHFDH)
Q8R0Y6 3 98709.17/5.63 16/123 (13%) 4.55e1008 Metabolism
HnRNPA2/B1 O88569 4 35993.9/8.67 6/36 (16%) 1.42e1004 RNA processing
T-complex protein 1 subunit a(TCP1-a)
P11983 5 60449.0/5.82 9/101 (8%) 4.09e1003 Molecular chaperone:protein folding
T-complex protein 1 subunit b(TCP1-b)
P80314 6 57477.6/5.97 10/108 (9%) 2.73e1006 Molecular chaperone:protein folding
T-complex protein 1 subunit e(TCP1-e)
P80316 7 59624.4/5.72 13/83 (13%) 5.23e1004 Molecular chaperone:protein folding
T-complex protein 1 subunit y(TCP1-y)
P42932 8 59555.8/5.44 19/156 (12%) 2.72e1008 Molecular chaperone:protein folding
Heat shock cognate 71 kDaprotein
P08109 9 70871.4/5.38 14/67 (20%) 1.08e1008 Chaperone
Actin, cytoplasmic 1 P60710 10 41737.0/5.29 15/258 (5%) 2.24e1008 Cell motility
Dynamin-2 P39054 11 98145.8/7.02 10/88 (11%) 2.05e1004 Vesicular trafficking
GP muscle form Q9WUB3 12 97287/6.65 9/52 (17%) 1.38e1004 Carbohydrate metabolism
Fructose-1,6-biphosphatase 1 Q9QXD6 13 36912.7/6.15 7/53 (13%) 5.44e1003 Gluconeogenesis
78 kDa glucose-regulated protein(precursor) GRP 78
P20029 14 72422.4/5.07 8/55 (14%) 2.08e1004 Protein assembly
Eukaryotic translation initiationfactor 2 a subunit (eIF2-a)
Q6ZWX6 15 36108.4/5.02 14/126 (11%) 2.45e1008 Translation initiation
Serum albumin precursor P07724 16 68693.0/5.75 12/168 (7%) 2.4e1006 Transport
DNA polymerase delta subunit 2(p50)
O35654 17 51368.9/5.50 6/40 (15%) 1.66e1003 DNA Replication
Casein kinase II subunit a (CK2) Q60737 18 45162.4/7.79 9/111 (8%) 1.36e1004 Cell signaling
ATP synthase b chain,mitochondrial precursor
P56480 19 51749.20/4.99 9/167 (5%) 2.61e1004 ATP synthesis
Selenium-binding protein 2 Q63836 20 52628.3/5.78 12/113 (10%) 2.95e1006 Not known
Metastasis-associatedprotein MTA
Q8K4B0 21 80798.6/9.37 8/53 (15%) 1.27e1003 Regulation of geneexpression
By Western blotting, we also tested for the presence in theeluates of some proteins previously described as SET-bind-ing proteins. Thus, we detected the presence of pp32(Fig. 2B), hnRNPA2, and glyceraldehyde 1-3 phosphatedehydrogenase (GAPDH; data not shown) in the eluatesfrom the SET column but not from the control column.Since SET associates with cell cycle regulatory proteinssuch as p21Cip1 and cyclin B, we aimed to analyze thepresence of some other cell cycle regulatory proteins in theeluates of the SET-Sepharose 4B column, even though theyhad not been detected by MS. We blotted the membraneswith antibodies against cdk4, cdk2, cyclin D1, cyclin D2, orcyclin D3. Data showed that SET interacts with cdk4 andcyclin D2 but not with cdk2, cyclin D1, or cyclin D3(Fig. 2C).
3.4 Identification of high affinity SET-binding
proteins
To discriminate between the low- and high-affinity SET-interacting proteins, we performed affinity chromatographyexperiments in which the proteins bound to the SET-col-umns were eluted with increasing amounts of KCl. Asshown in Fig. 3A, elution at 150 mM KCl released from thecolumn a number of proteins which were considered to below affinity SET-binding proteins. The subsequent elutionswith 300 and 500 mM KCl released two different sets of pro-teins that we considered to bind to SET with high affinity.
To identify some of the high affinity SET-binding pro-teins, the 300 mM KCl eluates from the SET columns wereanalyzed by MS using a MALDI-TOF. From these analyses
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
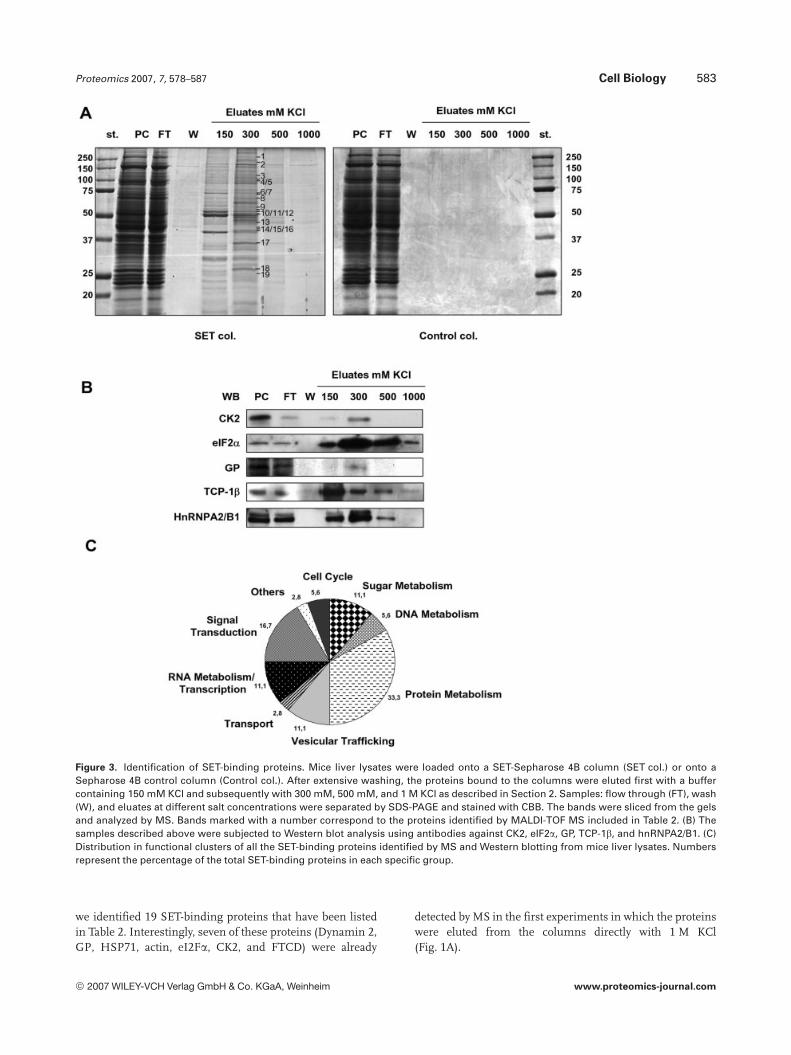
Proteomics 2007, 7, 578–587 Cell Biology 583
Figure 3. Identification of SET-binding proteins. Mice liver lysates were loaded onto a SET-Sepharose 4B column (SET col.) or onto aSepharose 4B control column (Control col.). After extensive washing, the proteins bound to the columns were eluted first with a buffercontaining 150 mM KCl and subsequently with 300 mM, 500 mM, and 1 M KCl as described in Section 2. Samples: flow through (FT), wash(W), and eluates at different salt concentrations were separated by SDS-PAGE and stained with CBB. The bands were sliced from the gelsand analyzed by MS. Bands marked with a number correspond to the proteins identified by MALDI-TOF MS included in Table 2. (B) Thesamples described above were subjected to Western blot analysis using antibodies against CK2, eIF2a, GP, TCP-1b, and hnRNPA2/B1. (C)Distribution in functional clusters of all the SET-binding proteins identified by MS and Western blotting from mice liver lysates. Numbersrepresent the percentage of the total SET-binding proteins in each specific group.
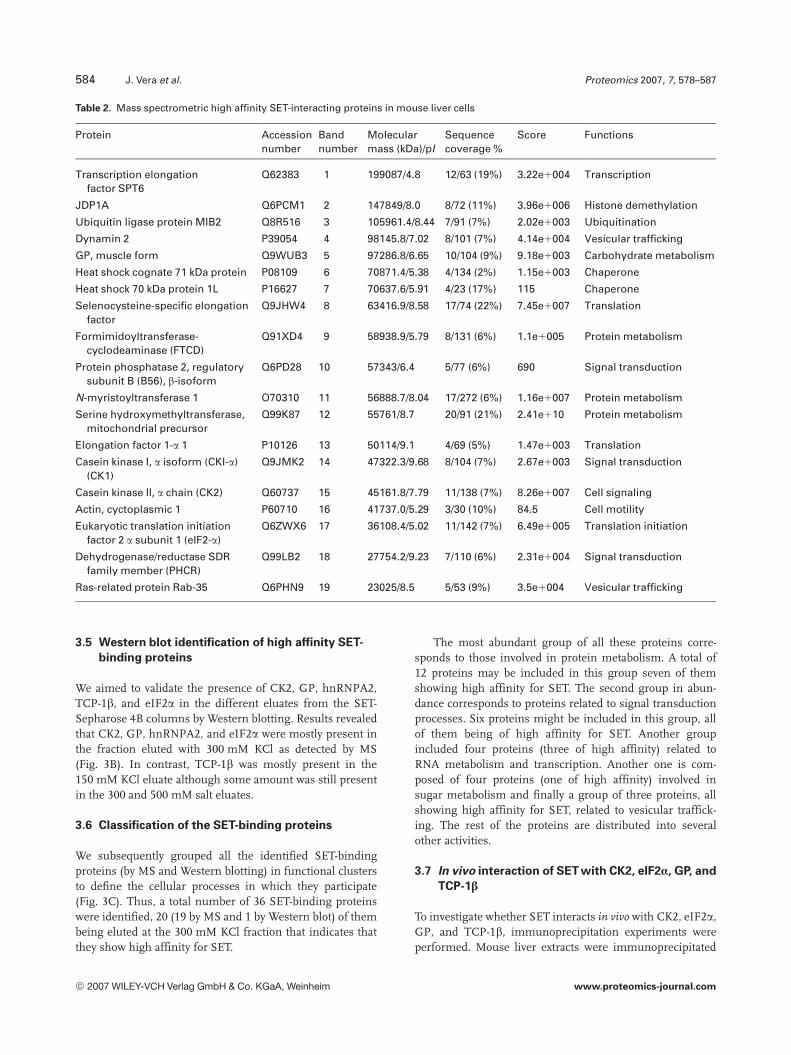
we identified 19 SET-binding proteins that have been listedin Table 2. Interestingly, seven of these proteins (Dynamin 2,GP, HSP71, actin, eI2Fa, CK2, and FTCD) were already
detected by MS in the first experiments in which the proteinswere eluted from the columns directly with 1 M KCl(Fig. 1A).
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
584 J. Vera et al. Proteomics 2007, 7, 578–587
Table 2. Mass spectrometric high affinity SET-interacting proteins in mouse liver cells
Protein Accessionnumber
Bandnumber
Molecularmass (kDa)/pI
Sequencecoverage %
Score Functions
Transcription elongationfactor SPT6
Q62383 1 199087/4.8 12/63 (19%) 3.22e1004 Transcription
JDP1A Q6PCM1 2 147849/8.0 8/72 (11%) 3.96e1006 Histone demethylation
Ubiquitin ligase protein MIB2 Q8R516 3 105961.4/8.44 7/91 (7%) 2.02e1003 Ubiquitination
Dynamin 2 P39054 4 98145.8/7.02 8/101 (7%) 4.14e1004 Vesicular trafficking
GP, muscle form Q9WUB3 5 97286.8/6.65 10/104 (9%) 9.18e1003 Carbohydrate metabolism
Heat shock cognate 71 kDa protein P08109 6 70871.4/5.38 4/134 (2%) 1.15e1003 Chaperone
Heat shock 70 kDa protein 1L P16627 7 70637.6/5.91 4/23 (17%) 115 Chaperone
Selenocysteine-specific elongationfactor
Q9JHW4 8 63416.9/8.58 17/74 (22%) 7.45e1007 Translation
Formimidoyltransferase-cyclodeaminase (FTCD)
Q91XD4 9 58938.9/5.79 8/131 (6%) 1.1e1005 Protein metabolism
Protein phosphatase 2, regulatorysubunit B (B56), b-isoform
Q6PD28 10 57343/6.4 5/77 (6%) 690 Signal transduction
N-myristoyltransferase 1 O70310 11 56888.7/8.04 17/272 (6%) 1.16e1007 Protein metabolism
Serine hydroxymethyltransferase,mitochondrial precursor
Q99K87 12 55761/8.7 20/91 (21%) 2.41e110 Protein metabolism
Elongation factor 1-a 1 P10126 13 50114/9.1 4/69 (5%) 1.47e1003 Translation
Casein kinase I, a isoform (CKI-a)(CK1)
Q9JMK2 14 47322.3/9.68 8/104 (7%) 2.67e1003 Signal transduction
Casein kinase II, a chain (CK2) Q60737 15 45161.8/7.79 11/138 (7%) 8.26e1007 Cell signaling
Actin, cyctoplasmic 1 P60710 16 41737.0/5.29 3/30 (10%) 84.5 Cell motility
Eukaryotic translation initiationfactor 2 a subunit 1 (eIF2-a)
Q6ZWX6 17 36108.4/5.02 11/142 (7%) 6.49e1005 Translation initiation
Dehydrogenase/reductase SDRfamily member (PHCR)
Q99LB2 18 27754.2/9.23 7/110 (6%) 2.31e1004 Signal transduction
Ras-related protein Rab-35 Q6PHN9 19 23025/8.5 5/53 (9%) 3.5e1004 Vesicular trafficking
3.5 Western blot identification of high affinity SET-
binding proteins
We aimed to validate the presence of CK2, GP, hnRNPA2,TCP-1b, and eIF2a in the different eluates from the SET-Sepharose 4B columns by Western blotting. Results revealedthat CK2, GP, hnRNPA2, and eIF2a were mostly present inthe fraction eluted with 300 mM KCl as detected by MS(Fig. 3B). In contrast, TCP-1b was mostly present in the150 mM KCl eluate although some amount was still presentin the 300 and 500 mM salt eluates.
3.6 Classification of the SET-binding proteins
We subsequently grouped all the identified SET-bindingproteins (by MS and Western blotting) in functional clustersto define the cellular processes in which they participate(Fig. 3C). Thus, a total number of 36 SET-binding proteinswere identified, 20 (19 by MS and 1 by Western blot) of thembeing eluted at the 300 mM KCl fraction that indicates thatthey show high affinity for SET.
The most abundant group of all these proteins corre-sponds to those involved in protein metabolism. A total of12 proteins may be included in this group seven of themshowing high affinity for SET. The second group in abun-dance corresponds to proteins related to signal transductionprocesses. Six proteins might be included in this group, allof them being of high affinity for SET. Another groupincluded four proteins (three of high affinity) related toRNA metabolism and transcription. Another one is com-posed of four proteins (one of high affinity) involved insugar metabolism and finally a group of three proteins, allshowing high affinity for SET, related to vesicular traffick-ing. The rest of the proteins are distributed into severalother activities.
3.7 In vivo interaction of SET with CK2, eIF2Æ, GP, and
TCP-1�
To investigate whether SET interacts in vivo with CK2, eIF2a,GP, and TCP-1b, immunoprecipitation experiments wereperformed. Mouse liver extracts were immunoprecipitated
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Proteomics 2007, 7, 578–587 Cell Biology 585
with anti-SET antibodies or with normal rabbit serum as acontrol. Then, the immunoprecipitates were immunoblottedwith antibodies against CK2, eIF2a, GP, or TCP-1b. Datarevealed that SET is able to interact in vivo with each one ofthe four studied proteins (Fig. 4).
Figure 4. In vivo interaction of SET with CK2, eIF2a, GP, and TCP-1b. Mice liver lysates were immunoprecipitated with SET anti-bodies or with normal rabbit serum (NRS) as a control. Thepresence of CK2, eIF2a, GP, and TCP-1b in the immunoprecipi-tates was investigated by Western blotting using CK2, eIF2a, GP,or TCP-1b antibodies, respectively. A sample of liver lysate wasadded as a control (L).
3.8 SET interacts with the active but not with the
inactive form of GP
GP is the key regulatory enzyme of glycogen metabolism,promoting the lysis of glycogen. We aimed to analyze herethe ability of SET to interact with both the active (phospho-rylated) and inactive (unphosphorylated) forms of this pro-tein. To this end, the purified phosphorylated active (GP-A)and the unphosphorylated inactive (GP-B) GP forms wereloaded separately onto SET-Sepharose 4B chromatographycolumns. After extensive washes, the protein bound to thecolumns was eluted with a buffer containing 1 M KCl. Thecollected fractions were electrophoresed and CBB stained. Asshown in Fig. 5A, only the active form of GP (GP-A) is able tointeract with SET.
3.9 SET is a CK2 substrate
CK2 is involved in the regulation of a great number of cel-lular processes including signal transduction, metabolism,proliferation, and differentiation. With the aim of studyingthe functional implication of CK2-SET interaction we per-formed in vitro phosphorylation assays in the presence ofSET using the monomeric (CK2a) or the tetrameric andmore active form of the enzyme (CK2ab). Thus, we deter-mined CK2 activity in the presence of SET. As a positivecontrol of the experiment the CK2 substrate b-casein wasused. As shown in Fig. 5B, we found that SET is a CK2 sub-strate.
Figure 5. SET binds to the active form of GP and is phospho-rylated by CK2. (A) 30 mg of purified active GP (GP-A) or inactive(GP-B) were loaded onto a SET-Sepharose 4B column (SET col.)or onto a Sepharose 4B column (Control col.). After extensivewashing, the proteins bound to the columns were eluted with abuffer containing 1 M KCl. Samples: precolumn (PC), flowthrough (FT), wash (W), and eluate (E) were electrophoresed andthe gels stained with CBB. (B) Phosphorylation of SET by CK2was performed by incubating 2 pmol of CK2a or 2 pmol of CK2a/band 30 pmol of SET in the presence of 5 mCi of g-(32P)-ATP asdescribed in Section 2. The samples were electrophoresed, thegels were dried and the phosphorylated proteins were analyzedon a PhosphoImager. The positive control was performed usingb-casein as a CK2 substrate.
4 Discussion
A growing number of evidence indicate that the protein SETis a key regulator of several cellular processes including cellcycle control, chromatin remodeling, apoptosis, cell signal-ing, neuronal development, and RNA metabolism [2, 9, 11,13, 16, 18, 20, 24]. Moreover, it has also been reported thatSET is involved in tumorigenesis [1, 5, 7]. Thus, in order togain a better understanding of the molecular mechanismsinvolved in the cellular processes regulated by SET, we havedeveloped a proteomic approach to identify SET-bindingproteins by combining affinity chromatography and MStechniques which was complemented by Western blot anal-ysis.
Results presented here show the identification of 36 SET-binding proteins from mice liver extracts. From these, 20might be considered high affinity SET-binding protein be-cause they are eluted from the columns at 300 mM KCl, aconcentration higher than that present in the cells(150 mM). Three of these proteins, hnRNPA2/B1 and thephosphatases PP2A and pp32 have been previously identi-
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

586 J. Vera et al. Proteomics 2007, 7, 578–587
fied as SET-binding proteins [2, 28, 29]. Thus, the other 33identified proteins can be considered as new SET-interactingproteins. However, this does not mean that all of these pro-teins will directly interact with SET, but it is possible thatonly some of these proteins actually have the ability to di-rectly bind to SET, the others being indirectly associatedthrough protein complexes including SET and the directSET-interacting proteins.
The presence of five of the SET-binding proteins identi-fied by MS in the eluates from the SET-Sepharose 4B column(CK2, eIF2a, GP, hnRNPA2, and TCP-1b), was validated byWestern blotting using specific antibodies against these pro-teins. Moreover, the in vivo binding between four of theseproteins (CK2, eIF2a, GP, and TCP-1b) and SET was alsodemonstrated by immunoprecipitation experiments.
The classification of the SET-interacting proteins infunctional clusters revealed that most of the proteins belongto four major functional groups: (i) protein metabolism (12proteins); (ii) signal transduction (six proteins); (iii) RNAprocessing and transcription (four proteins), and (iv) sugarmetabolism (four proteins).
The first major group of SET-interacting proteins includesa number of chaperons Hsp70, HSP71, GRP78, and the sub-units a, b, e, and y of TCP-1 (T-complex) [30–32], a set of pro-teins involved in protein synthesis as eIF2a, eEF1A-1, andselenocystein-specific elongation factor [33–35] and two pro-teins involved in the synthesis or interconversion of aminoacids, the serine hydroxymethyl transferase (SHT), andFTCD. Binding to chaperons prevents the nascent polypep-tides from premature folding and incorrect interactions be-tween or within the amino acid chains. Once the protein isprepared for competent folding it does release from the chap-eron and in some cases it is processed by the cytosolic chap-eronin complex TCP-1 which completes the folding cycle. Theobservation that SET was bound to at least four of the TCP-1subunits and to other chaperones opens the possibility thatSET may be a chaperone substrate or alternatively that SETcould cooperate with chaperones in the folding process.
The initiation factor eIF2 mediates the binding of themethionil-tRNA to the ribosome during the initiation of thetranslation of the mRNAs [33–35]. Recently, eIF2 has alsobeen implicated in other cellular events as the response tostress [36]. The finding that SETnot only interacts with eIF2abut also with two elongation factors suggests a role of SETonthe regulation of protein synthesis. This putative role onprotein synthesis is also supported by the observation thatSET associates with SHT and FTCD enzyme participating inthe synthesis or interconversion of amino acids.
Six of the identified proteins, all of them showing highaffinity for SET, might be considered to participate in signaltransduction pathways. This group includes two proteinkinases, namely CK1 and CK2, the subunit B56 of the phos-phatase PP2A, the phosphatase pp32, a myristoyl transferaseand a dehydrogenase reductase. It merits the mention thatCK2 is a protein kinase that participates in many cellularprocesses such as cell division, transcription, RNA proces-
sing, and translation [37]. We observed that CK2 is able tophosphorylate SET in vitro a fact that is in agreement withthe observation that CK2 is also able to phosphorylate in vivoand in vitro the proteins NAP-1 and DEK [38, 39], bothstructurally and functionally similar to SET. Interestingly, ithas been reported that CK2 phosphorylates eIF2a a proteinthat we have also found in the eluates from the SET-Sephar-ose 4B columns (see above), suggesting that CK2 could beforming part of a protein complex that associates with theSET-columns. The interaction of SET with the phosphatasesPP2A and pp32 was already reported [2, 19].
We also report here that SET interacts with four proteinsinvolved in transcription or RNA processing (hnRNPA2,MTA1, SPT6, and Jumonji domain-containing protein 1A(JDP1A)).
The interaction between SET and hnRNPA2, a proteininvolved in RNA processing, confirms a previous report [29].The protein MTA1 displays histone deacetylase activity andbelongs to the nucleosome remodeling histone deacetylasecomplex (NuRD complex) involved in chromatin remodeling[40, 41]. The JDP1A protein is a histone dimethylase thatspecifically demethylates lys-9 of histone H3. As SET hasbeen reported to bind to histones and in that way blocking itsacetylation, the results reported here suggest that SET mightparticipate in chromatin remodeling by at least three differ-ent mechanisms: (i) by blocking histone acetylation; (ii) byassociating to MTA1 and as a consequence to the remodelingNuRD complex, and (iii) by regulating demethylation of his-tones through JDP1A. Interestingly, SET might also beinvolved in the regulation of transcription by its binding toSPT6 a protein that stimulates the transcriptional elongationby RNA polymerase II.
Three of the new SET-interacting proteins identified here(a-enolase, GP, and fructose 1,6-biphosphatase) participate inthe glycolytic pathway, strongly suggesting the involvement ofSET in the regulation of this metabolic pathway. These resultsare in agreement with the previous report showing thatGAPDH, another enzyme involved in the glycolytic pathwayis also a SET-binding protein and that it participates in cellcycle regulation [42]. Since alterations in the glucose metabo-lism are common in cancer cells, it could be possible that SETmight act as a connection between sugar metabolism and cellcycle through the interaction with GAPDH but also with GP,the enzyme which promotes the lysis of glycogen. It is knownthat protein kinase A phosphorylates phosphorylase kinasewhich in turn phosphorylates GP at ser-14 and as a con-sequence it is activated. In this work we have demonstratedthat SETdirectly interacts with the active and phosphorylatedform of GP but not with its unphosphorylated form, support-ing the hypothesis that SET could also be functionallyinvolved in glycogen metabolism.
Our results also revealed the interaction of SET withother proteins as belonging to the vesicular trafficking (actin,dynamin 2, MIB2, and Rab-35) suggesting its participationin these processes. We also found that SET interacts withDNA polymerase d and with 10-formyltetrahydrofolate
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Proteomics 2007, 7, 578–587 Cell Biology 587
dehydrogenase, enzymes involved in DNA synthesis andpurine synthesis, respectively, suggesting a direct role of SETin DNA replication.
Proteins that have previously been described as SET-binding proteins as for instance p21Cip1 [9, 43] and GAPDH[42] or proteins that have been detected in the eluates fromthe SET-Sepharose 4B columns by Western blotting, as cyclinD2 and cdk4, have not been detected by MS. Thus, we con-clude that our experimental approach has permitted theidentification of a significant number of new SET-interactingproteins but still some of them escape from this approachpossibly due to the type of sample used for the experimentsor to the amount of these proteins present in these samples.Thus, in the near future, further investigations using differ-ent types of samples will probably increase the list of newSET-interacting proteins.
In summary, the combination of proteomic analysis andfunctional studies has been a successful tool to discover newcellular pathways in which the protein SET may be involved.
This work was supported by grants from the Spanish Minis-terio de Educación y Ciencia SAF 2002-00452, SAF 2003-08339, and GEN2003-20243-C08-01. Proteomic analysis wasperformed at the Proteomic Unit of the “Serveis Científico-tèc-nics” from the University of Barcelona.
5 References
[1] von Lindern, M., van Baal, S., Wiegant, J., Raap, A. et al, Mol.Cell Biol. 1992, 12, 3346–3355.
[2] Li, M., Makkinje, A., Damuni, Z., J. Biol. Chem. 1996, 271,11059–11062.
[3] Janssens, V., Goris, J., Van Hoof, C., Curr. Opin. Genet. Dev.2005, 15, 34–41.
[4] Saito, S., Miyaji-Yamaguchi, M., Nagata, K., Int. J. Cancer2004, 111, 501–507.
[5] Adler, H. T., Nallaseth, F. S., Walter, G., Tkachuk, D. C., J. Biol.Chem. 1997, 272, 28407–28414.
[6] Pandey, A. V., Mellon, S. H., Miller, W. L., J. Biol. Chem. 2003,278, 2837–2844.
[7] Carlson, S. G., Eng, E., Kim, E. G., Perlman, E. J. et al., J. Am.Soc. Nephrol. 1998, 9, 1873–1880.
[8] Fukukawa, C., Shima, H., Tanuma, N., Ogawa, K. et al., Can-cer Lett. 2000, 161, 89–95.
[9] Estanyol, J. M., Jaumot, M., Casanovas, O., Rodriguez-Vilarrupla, A. et al., J. Biol. Chem. 1999, 274, 33161–33165.
[10] Kellogg, D. R., Kikuchi, A., Fujii-Nakata, T., Turck, C. W. et al.,J. Cell. Biol. 1995, 130, 661–673.
[11] Canela, N., Rodriguez-Vilarrupla, A., Estanyol, J. M., Diaz, C.et al., J. Biol. Chem. 2003, 278, 1158–1164.
[12] Smith, D. S., Greer, P. L., Tsai, L. H., Cell Growth Differ. 2001,12, 277–283.
[13] Fukukawa, C., Tanuma, N., Okada, T., Kikuchi, K. et al., Can-cer Lett. 2005, 226, 155–160.
[14] Nagata, K., Saito, S., Okuwaki, M., Kawase, H. et al., Exp. CellRes. 1998, 240, 274–281.
[15] Martinvalet, D., Zhu, P., Lieberman, J., Immunity 2005, 22,355–370.
[16] Fan, Z., Beresford, P. J., Zhang, D., Lieberman, J., Mol. CellBiol. 2002, 22, 2810–2820.
[17] Fan, Z., Beresford, P. J., Oh, D. Y., Zhang, D. et al., Cell 2003,112, 659–672.
[18] Seo, S. b., McNamara, P., Heo, S., Turner, A. et al., Cell 2001,104, 119–130.
[19] Kutney, S. N., Hong, R., Macfarlan, T., Chakravarti, D., J. Biol.Chem. 2004, 279, 30850–30855.
[20] Cervoni, N., Detich, N., Seo, S. B., Chakravarti, D. et al., J.Biol. Chem. 2002, 277, 25026–25031.
[21] Okuwaki, M., Nagata, K., J. Biol. Chem. 1998, 273, 34511–34518.
[22] Shikama, N., Chan, H. M., Krstic-Demonacos, M., Smith, L. etal., Mol. Cell Biol. 2000, 20, 8933–8943.
[23] Gamble, M. J., Erdjument-Bromage, H., Tempst, P., Freed-man, L. P. et al., Mol. Cell Biol. 2005, 25, 797–807.
[24] Brennan, C. M., Gallouzi, I. E., Steitz, J. A., J. Cell Biol. 2000,151, 1–14.
[25] Blum, H., Beier, H., Gross, H., Electrophoresis 1987, 8, 93–99.
[26] Bosser, R., Faura, M., Serratosa, J., Renau-Piqueras, J. et al.,Mol. Cell Biol. 1995, 15, 661–670.
[27] Clauser, K. R., Baker, P., Burlingame, A. L., Anal. Chem. 1999,71, 2871–2882.
[28] Kutney, S. N., Hong, R., Macfarlan, T., Chakravarti, D., J. Biol.Chem. 2004, 279, 30850–30855.
[29] Vera, J., Jaumot, M., Estanyol, J. M., Brun, S. et al., Onco-gene 2006, 25, 260–270.
[30] Kiang, J. G., Tsokos, G. C., Pharmacol. Ther. 1998, 80, 183–201.
[31] Schroder, M., Kaufman, R. J., Annu. Rev. Biochem. 2005, 74,739–789.
[32] Spiess, C., Meyer, A. S., Reissmann, S., Frydman, J., TrendsCell Biol. 2004, 14, 598–604.
[33] Kimball, S. R., Int. J. Biochem. Cell Biol. 1999, 31, 25–29.
[34] Sonenberg, N., Dever, T. E., Curr. Opin. Struct. Biol. 2003, 13,56–63.
[35] Asano, K., Clayton, J., Shalev, A., Hinnebusch, A. G., GenesDev. 2000, 14, 2534–2546.
[36] Jiang, H. Y., Wek, R. C., Biochem. J. 2005, 385, 371–380.
[37] Meggio, F., Pinna, L. A., FASEB J. 2003, 17, 349–368.
[38] Li, M., Strand, D., Krehan, A., Pyerin, W. et al., J. Mol. Biol.1999, 293, 1067–1084.
[39] Kappes, F., Damoc, C., Knippers, R., Przybylski, M. et al., Mol.Cell Biol. 2004, 24, 6011–6020.
[40] Kumar, R., Wang, R. A., Bagheri-Yarmand, R., Semin. Oncol.2003, 30, 30–37.
[41] Zhang, Y., Ng, H. H., Erdjument-Bromage, H., Tempst, P. etal., Genes Dev. 1999, 13, 1924–1935.
[42] Carujo, S., Estanyol, J. M., Ejarque, A., Agell, N. et al.,Oncogene 2006, 25, 4033–4042.
[43] Carrascal, M., Carujo, S., Bachs, O., Abian, J., Proteomics2002, 2, 455–468.
© 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com