protein - plant · pdf filewere used as test material. ... the index of activity adopted here...
TRANSCRIPT

ON THE MECHANISM OF ACTION OF GROWTH REGULATORS
ROBERT M. MUIR AND CORWIN HANSCHDEPARTMENT OF BOTANY, STATE UNIVERSITY OF IOWA, IOWA CITY, IOWA AND
DEPARTMENT OF CHEMISTRY, POMONA COLLEGE, CLAREMONT, CALIFORNIA
Received August 8, 1952
Introduction
Investigations of the effects of various analogues of indoleacetic acid,phenoxyacetic acid, phenylacetic acid and benzoic acid on the elongation ofsections of Avena coleoptiles (7, 8, 16, 17) have led to the conclusion thatplant growth-regulators react by a nucleophilic substitution at a position onthe ring ortho to the carboxyl group or the side chain carrying this groupand with the carboxyl group itself. The evidence at hand indicates tnat acysteinyl unit of a protein is the most likely substrate for the two-pointortho reaction as illustrated below with 2,4-dichlorophenoxyacetic acid.
0/CHi-C
NHO-CH2COOH 0 1 H
H-S-CH2-CH--CClCH2000H NHI \Protein /
C1 (z1There are nine important experimental requirements, in addition to the
requirement of an unsaturated ring and a carboxyl group first postulated byKOEPFLI et al. (11), which must be explained by any hypothesis which wouldcorrelate chemical structure of the growth regulators with physiologicalactivity. These are (a) the effect of optical isomerism of the side chain(18, 35, 36), (b) the effect of geometrical isomerism of the side chain (31,32, 33), (c) the ortho effect in benzoic acids (1, 16), (d) the ortho effect inphenoxyacetic acids (7, 13, 17), (e) the inhibiting effects of lactones (6, 32)and maleic hydrazide (4, 9, 14, 40), (f) the inhibiting effect of cysteine (2),(g) the inhibiting effects of reagents specific for sulfhydryl groups (28, 29),(h) the inactivity of phenoxyacetic acids without two open para positions(13), (i) the effect of electron-releasing and electron-attracting ring sub-stituents (16). It is the purpose of this paper to present data on the relativeactivity of some growth regulators and to consider recent proposals for thereaction mechanism of the growth regulators in relation to the requirementslisted above.
Methods
Seedlings of Avena plants grown in sand in darkness for 88 to 90 hoursunder conditions of constant temperature (24 ± 1° C) and humidity (85%)
218
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

MUIR AND HA.NSCH: ACTION OF GROWTH REGULATORS
were used as test material. The apical two millimeters of the coleoptilewere removed and after two hours a single section, 3 ± 0.02 mm. in length,was cut from the apex with the primary leaf included within the coleoptilesection. Fifteen such sections were floated on the surface of 25 ml. of a solu-tion of the chemical in distilled water in a Petri dish. The compounds weretested over a concentration range of 10-8 to 10-4 M, and in some cases thehigher concentrations included 5 x 10-4 M. When the concentration requiredto induce the growth response was great enough to give a pH of 5.4 or less,NaOH was added to raise the pH to 5.6 to 6.0. After 24 hours in the coveredPetri dish in the dark room, the sections were removed and their length wasmeasured with an ocular micrometer mounted in a binocular dissectingmicroscope at a magnification of 15 diameters.
Results and discussion
In formulating a measurement of the relative activity of chemical com-pounds in effecting the elongation of coleoptile sections several possibilitieshave been examined. The index of activity adopted here is the molar con-centration of the compound causing a growth response compared with themolar concentration of indoleacetic acid (IAA) inducing the same growthresponse. An average elongation of the sections which is greater by 0.15mm. (5%o of the original length) than the elongation occurring in distilledwater was made the standard of growth response. The concentration of IAAgiving this growth response in the tests reported here is 5 x 10-8 M.
The relative activities of 117 compounds are listed in tables I and II.The only compound which has been found to have a greater activity thanIAA is the 4-chloro-derivative, and the only compound with equal activityis a-naphthaleneacetonitrile. By the activity index used here a-naphtha-leneacetic acid and 2,3,5-triiodobenzoic acid have activities which are onehalf that of IAA, and 2,4-dichlorophenoxyacetic acid and 2,4,5-trichloro-phenoxyacetic acid have only one fourth the activity of IAA.
Following the report of JONES et al. (10) that indoleacetonitrile is activ-ein causing the elongation of Avena coleoptile sections a number of nitrileswere tested. As shown in table II a-naphthaleneacetonitrile is as activeas IAA and more active than naphthaleneacetic acid, and phenylacetonitrileis more active than phenylacetic acid. Benzonitrile is inactive as are a-naph-thonitrile and y-phenoxybutyronitrile. ZIMMERMAN and WILCOXON (39)found a-naphthaleneacetonitrile caused an epinastic response which wasslower than the response induced by other regulators and suggested thenitrile is hydrolyzed in the plant tissue to the acid. VELDSTRA (30) foundboth indoleacetonitrile and naphthaleneacetonitrile to be inactive in the peatest but active in root formation in cuttings and considered the latter ac-tivity to be due to hydrolysis. Since the only nitriles found to be active inthe present investigation are those which may be hydrolyzed to give an acidwhich is active, it might be supposed that hydrolysis does take place in thetissue of the coleoptile, yet the same results would be obtained if the nitrile
219
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
TABLE IRELATIVE ACTIVITY OF INDOLEACETIC ACID, PHENOXYACETIC ACID, ANDDERIVATIVES IN PROMOTING GROWTH (ELONGATION) OF PLANT CELLS.
Activity index =
Molar conc. of 3-indoleacetic acid inducing an elongation of 0.15 mm.
Molar conc. of growth regulator inducing an elongation of 0.15 mm.
Compound
3-Indoleacetic acid (5 x 10' M)4-Chloro-3-indoleacetic acid2-Methyl-3-indoleacetic acid4,7-Dichloro-2-methyl-3-indoleacetic acid5,7-Dichloro-2-methyl-3-indoleacetic acid3-Indolebutyric acid3-Indolepropionic acid
Phenoxyacetic acid2-Acetyl-5-methylphenoxyacetic acido-Bromophenoxyacetic acidm-Bromophenoxyacetic acidp-Bromophenoxyacetic acid2,4-Dibromophenoxyacetic acid2,6-Dibromophenoxyacetic acid2,4,6,-Tribromophenoxyacetic acido-Chlorophenoxyacetic acidm-Chlorophenoxyacetic acidp-Chlorophenoxyacetic acid2,4-Dichlorophenoxyacetic acid2,6-Dichlorophenoxyacetic acid3,5-Dichlorophenoxyacetic acid2,4,5-Trichlorophenoxyacetic acid2,4,6-Trichlorophenoxyacetic apid2,4-Dichloro-6-methylphenoxyacetic acid2,4-Dichloro-5-nitrophenoxyacetic acid2-Ethyl-4-chlorophenoxyacetic acido-Iodophenoxyacetic acidp-Iodophenoxyacetic acid2,4-Diiodophenoxyacetic acido-Isopropylphenoxyacetic acid2-Isopropyl-4-chloro-5-methylphenoxyacetic acido-Methoxyphenoxyacetic acidm-Methoxyphenoxyacetic acidp-Methoxyphenoxyacetic acido-Methylphenoxyacetic acidm-Methylphenoxyacetic acidp-Methylphenoxyacetic acid2,4-Dimethylphenoxyacetic acid2,5-Dimethylphenoxyacetic acid3,5-Dimethylphenoxyacetic acid2,4,6-Trimethylphenoxyacetic acido-Nitrophenoxyacetic acidm-Nitrophenoxyacetic acidp-Nitrophenoxyacetic acid2,4-Dinitrophenoxyacetic acid
Activity Index
100.0140.0
1.50.11.5
15.01.5
0.03Inactive
0.12.51.5
12.5InactiveInactive
0.062.05.0
25.0InactiveInactive25.0
InactiveInactive
0.2Inactive
0.1InactiveInactiveInactiveInactiveInactive
0.10.030.20.070.050.50.2
InactiveInactiveInactive
0.20.1
Inactive
220
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

MUIR AND HANSCH: ACTION OF GROWTH REGULATORS
TABLE IIRELATIVE ACTIVITY OF SOME R-EGULATORS IN PROMOTING GROWT7H
(ELONGATION) OF PLANT CELLS.Activity index =
Molar conc. of 3-indoleacetic acid inducing an elongation of 0.15 mm.x 100.
Molar conc. of growth regulator inducing an elongation of 0.15 mm.
Compound index Compound Activityindex-index
Benzoic acido-Aminobenzoic acidn-Aminobenzoic acid2-Amino-3,5-diiodobenzoic acido-Bromobenzoic acidm-Bromobenzoic acidp-Bromobenzoic -acido-Chlorobenzoic acidm-Chlorobenzoic acidp-Chlorobenzoic acid2-Chloro-5-nitrobenzoic acid2,4-Dichlorobenzoic acid2,5-Dichlorobenzoic acid,2,6-Dichlorobenzoic acidPentachlorobenzoic acid2,4,6-Triethylbenzoic acidm-Ethoxybenzoic acidp-Ethoxybenzoic acido-Hydroxybenzoic acidm-Hydroxybenzoic acidp-Iydroxybenzoic acid
Phenylacetic acida-Aminophenylacetic acidp-Aminophenylacetic acidm-Fluorophenylacetic acidp-Fluorophenylacetic acid2,5-Dihydroxyphenylacetic acidp-Iodophenylacetic acid2,5-Dimethoxyphenylacetic acid2,4-Dimethylphenylacetic acid3,5-Dimethylphenylacetic acid2,4,6-Trimethylphenylacetic
acidp-Nitrophenylacetic acid2,4-Dinitrophenylacetic acidp-Phenylphenylacetic acidDiphenylacetic acid
InactiveInactiveInactiveInactive
0.1InactiveInactive0.05
InactiveInactiveInactiveInact ive
1.00.1
InactiveInactiveInactiveInactiveInactiveInactiveInactive
1.0Inactive
0.051.51.50.02
InactiveInactive0.50.5
InactiveInactiveInactiveInactiveInactive
o-Fluorobenzoic acidm-Fluorobenzoic acidp-Fluorobenzoic acido-Iodobenzoic acidm-Iodobenzoic acidp-Iodobenzoic acid2,3,5-Triiodobenzoic acid3,4,5-'I'riiodobenzoic acid2,4,6-Triisopropylbenzoic acido-Methylbenzoic acidm-Methylbenzoic acidp-Methylbenzoic acid2,6-I)imethylbenzoic acid2,4,6-Trimethylbenzoic acid2,6-Dimethyl-4-methoxybenzoic2,6-Dimethoxy.benzoic acid2-Methoxy-3,5-dichloro-4,6,-
dimethylbenzoic acido-Nitrobenzoic acidm-Nitrobenzoic acidp-Nitrobenzoic acid3,5-Dinitrobenzoic acid
BenzonitrileBenzothiazyl-2-oxyacetic acida-Naphthaleneacetic acida-Naphthaleneacetonitrilea-Naphthonitrile,B-Naphthoxyacetic acid2-Phenanthreneacetic acid3-Phenanthreneacetic acidy-Phenoxybutyronitrilea-Phenoxypropionic acidPhenylacetonitr iley-Phenylbutyric acid2,4,6-Trimethylphenylbutyric
acidp-Chlorophenylglycinel3-Phenylpropionic acid
group reacts in similar fashion to the carboxyl group in the two-point reac-
tion. Further investigation of this problem is planned.One proposal concerning the chemical structure required to effect cell
elongation is that of WAIN (36) which emphasizes the presence of a hydro-gen atom in the alpha position of the aryloxyacetic acids and suggests thatthe growth regulator which effects cell elongation is adsorbed onto an active
InactiveInactiveInactiveInactiveInactiveInactive50.0
InactiveInactiveInactiveInactiveInactive
0.05InactiveInactiveInactive
Inactive0.1
InactiveInactiveInactive
Inactive0.5
50.0100.0
Inactive0.8
InactiveInactiveInactive
0.52.01.5
Inactive1.0
Inactive
221
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
site by three points: the carboxyl group, the unsaturated ring, and an alphahydrogen. Certain observations are not in accord with this hypothesis how-ever. Phenoxyacetic acid according to the hypothesis should be more activethan a-methylphenoxyacetic acid which is very active, yet as shown intable I phenoxyacetic acid has very low activity and many investigatorshave reported it to be completely inactive. The relative activities of sevenbenzoic acids which cause cell elongation although they do not have analpha hydrogen atom are given in table II. WAIN (36) believes the benzoicacids do not have the same mechanism of reaction as other growth regu-lators, but ZIMMERMAN and HITCHCOCK (38) have found that 2,3,6-trichloro-benzoic acid brings about the same responses of epinasty, proliferation oftissue, formation of adventitious roots, modification of leaves, and develop-ment of parthenocarpic fruit that are effected by other regulators. In thislaboratory 2,3,5-triiodobenzoic acid has been found to cause parthenocarpicfruit to develop when applied to ovaries of tomato in concentrations of 0.3and 0.03% in lanolin paste (22). The compound was also found to causegreater development of adventitious roots on cuttings of Euonymus at 10-6 Mand 10-7 M. SNYDER has reported similar effects of the compound on the root-ing of cuttings of Coleus when applied at a concentration of 1 p.p.m. (25).
In active compounds of the type of cis cinnamic acid, the alpha hydrogenatom, the benzene ring, and the carboxyl group do not have the same spatialrelation as in the saturated side chains of phenoxyacetic acids or phenyl-acetic acids and adsorption at precisely the same three points would there-fore not be possible. It also seems unlikely that the active compounds,y-phenylbutyric acid and 3-indolebutyric acid (table II), could be held bythe carboxyl group and an alpha hydrogen atom and still have the ringadsorbed at the same point which holds the ring of phenylacetic acid or phe-noxyacetic acid which have much shorter side chains.
The effect of alpha substitutions in the phenoxyacetic acids can beexplained by the steric hindrance of the carboxyl group by the alpha alkylgroups. It is a well known fact (5, 20) in organic chemistry that the intro-duction of substituents on the alpha carbon atom greatly slows down therate of reaction of the functional group, in this case the carboxyl group.This general problem has been discussed by WHELAND (37). Thus, if thecarboxyl group of a growth regulator reacts chemically then it is quitereasonable to expect that two groups on the alpha carbon atom would slowthis reaction to a point where the activity of the compound is negligible.This possibility is even more likely in the case of a biological reactionbrought about by large enzymatic catalysts which in themselves would re-quire considerable freedom of approach to the reaction site than it would bewith simple organic molecules.
Although K6GL and VERKAAIK (12) found that the D and L isomers ofa-methylindoleacetic acid are of equal activity, WAIN (36) and THIMANN(26) have reported that in a number of phenoxyacetic acids such as 2,4-dichloro-a-methylphenoxyacetic acid with one alkyl group on the alpha car-
222
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

MUIR AND HANSCH: ACTION OF GROWTH REGULATORS
bon, one optical isomer is active while the other is not. According to Wain'shypothesis one optical isomer would fit the three points of adsorption whileits mirror image would not and a compound with both hydrogens substitutedby alkyl groups would also not fit. However, the difference in activity ofoptical isomers resulting from an asymmetric carbon atom may also be ex-plained by differences in reaction rates resulting from interference in com-bining with another asymmetrical center. As an example of this interfer-ence, it has been shown (5) that the D form of mandelic acid reacts muchfaster than the L form With L-menthol in an esterification reaction. Thedemonstration by W\AIN and coworkers (24, 36) of the antagonism of theinactive di-alpha substituted phenoxyacetic acids toward the effect of suchregulators as 2,4-dichlorophenoxyacetic acid and the antagonism of the in-active isomer of 2,4,5-trichlorophenoxypropionic acid toward the effect ofthe acti-e isomer may be explained also on the basis of a two-poiint orthoreaction mechanism involving the carboxyl group. Despite the one or twoalpha substituents the carboxyl group would be able to form a salt linkagewith the basic group of the protein thus hindering the regulator from react-ing at this point. Although a salt linkage could be formed the subsequentamidization would be prevented by the methyl groups. Compounds withsteric hindrance of the carboxyl group but with open ortho positions couldreact at the ortho positions and prev-ent the regulator from completing thetwo-point attachment.
At first glance one might expect the di-ortho substituted benzoic acidsto contain a sterically hindered carboxyl group as in the di-alpha substi-tuted phenoxyacetic acids. Such benzoic acids are very slow to give manynormal reactions (37), but the mechanism postulated for 2,6-dichloro- and2,3,6-trichlorobenzoic acids (16) involves two reactions, one of which is anucleophilic substitution on the aromatic ring as follows:
NH+ H
{N+\ NProtein [ N-
112 Cl \C1 1 X \
Protein + I + Cl + H20A B C
To understand the steric significance of the intermediate, B, for the amidi-zation of A and C, one must consider the structure of B in detail. Theavailable chemical and theoretical evidence (3) shows tllat in nucleophilicsubstitution reactions an intermediate complex of type B is formed. MIanyof these complexes have considerable stability and some may even be isos.lated as such. When such a substitution occurs the usual aromatic reso-nance of the ring is momentarily destroyed and thus the carbon atoms 1 and6 may assume the tetrahedral structure. In the complex B above, carbon
223
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
atoms 1 and 6 may now possess the usual tetrahedral configuration with thecarboxyl group out of the plane of the ring. The first stage of the reactionis salt formation of the carboxyl group with an amine group, then nucle-ophilic displacement of the ortho chlorine atom. In the activated complexstate, the carboxyl group will be out of the plane of the ring and on the sameside of the ring from which the nucleophilic group, X, is attacking the chlo-rine atom. The important point to note is that in this position the carboxylgroup is attached to a ring carbon which in turn is attached to only twoother atoms, i.e., two ring carbon atoms, which means that the carboxylgroup will be relatively unhindered sterically for the amidization reactionwhich may now be promoted enzymatically. Thus, sterically, the structureof B would be analogous to a mono-alpha substituted phenoxyacetic acid,except that the latter would carry a hydrogen atom on the alpha carbonwhile the former would carry a momentarily unshared pair of electronswhich might be satisfied by a proton or an enzyme.
All of the benzoic acids listed in table II which cause cell elongation havea halogen atom or nitro group in the ortho position except 2,6-dimethyl-benzoic acid. The importance of the halogen atom at the ortho position indetermining the activity of the benzoic acids is demonstrated by the activityof 2,3,5-triiodobenzoic acid compared with the lack of activity of 2-amino-3,5-diiodobenzoic acid and 3,4,5-triiodobenzoic acid.
Although it was first shown by KOEPFLI et al. (11) that cis cinnamic acidcauses cell elongation and the trans form does not, the importance of thegeometrical isomerism of the side chain in determining the activity of allgrowth regulators with such forms has been emphasized by VELDSTRA andcoworkers (31, 32, 33, 34). The structure of the cis isomers is such that theside chain with the carboxyl group is held out of the plane of the ring be-cause of steric hindrance. Veldstra has proposed that the active compound,must have a non-polar part (ring system) with an acidic polar group (prefer-ably a carboxyl group) situated out of the plane of the ring as far as possi-ble or as frequently as possible.
The mechanism of action of growth regulators as interpreted by Veldstrais inadequate in several respects. In the first place, as THIMANN (26) haspointed out, if having the carboxyl group out of the plane of the ring is theprincipal requirement, then the most active compounds should be those suchas cis cinnamic acid in which this position is only one of many positionsattainable by free rotation, however such compounds are not the most active.Even more difficult to explain on the basis of Veldstra's hypothesis is theinactivity of certain benzoic acids. VELDSTRA (31) has shown spectrophoto-metrically that in 2,6-dichlorobenzoic acid and 2,3,6-trichlorobenzoic acidthe ortho substituents force the carboxyl group into a position at right anglesto the benzene ring and the activity of these compounds is predicted by hishypothesis. Benzoic acids with substituents in both ortho positions are notnecessarily active, however (16). Included in table II are several benzoicacids which have groups in both ortho positions and which are inactive.
224
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

MUIR AND HANSCH: ACTION OF GROWTH REGULATORS
These acids are 2,6-dimethoxybenzoic acid, 2,4,6-trimethylbenzoic acid,2,4,6-triethylbenzoic acid, 2,4,6-triisopropylbenzoic acid, 2,6-dimethyl-4-methoxybenzoic acid and 2-methoxy-3,5-dichloro-4,6-dimethylbenzoic acid.Certainly in these molecules there is no reason to doubt that the favoredposition of the carboxyl group is out of the plane of the ring and their lackof activity is not explained by Veldstra's hypothesis. It should be notedhere that 2,6-dimethylbenzoic acid is weakly active. The supposition thatthe hydrophilic/lipophilic character of the molecule effects its activity doesnot help in understanding the effect of ring substituents in benzoic acid.For example, in comparing the very active 2,3,6-trichlorobenzoic acid withthe completely inactive 2,4,6-trimethylbenzoic acid it seems highly unlikelythat these two molecules would differ greatly in their hydrophilic/lipophilicratio. Not only is the atomic volume of these groups the same (23), butthey are both also weakly polar, fat soluble groups. In both molecules thechlorine atoms and methyl groups in the ortho positions would hold thecarboxyl group in precisely the same position with respect to the ring.
The variation in activity shown by the phenylacetic acid and phenoxy-acetic acid series is also not explained by Veldstra's hypothesis. In tablesI and II are found several derivatives of these acids in which both orthopositions are substituted, thus preferentially holding the side chain out ofthe plane of the ring, yet all of them are inactive. These compounds includethe 2,6-dichloro-, 2,6-dibromo-, 2,4,6-trichloro-, 2,4,6-tribromo-, 2,4,6-tri-methyl-, and the 2,4-dichloro-6-methyl- derivatives of phenoxyacetic acidand 2,4,6-trimethylphenylacetic acid. Also inactive is 2,4,6-trimethyl-y-phenylbutyric acid.
Any hypothesis attempting to correlate chemical structure and physio-logical activity must consider the important effect of di-ortho substitutionon the activity of benzoic acids, phenylacetic acids and phenoxyacetic acids.Slight differences in adsorption characteristics are not adequate to accountfor the effects on activity. The existence of the ortho effect is illustratedstrikingly by a comparison of the completely inactive 2,4,6-trichlorophe-noxyacetic acid with the active 2,4,5-trichlorophenoxyacetic acid (table I).Since these molecules are made up of exactly the same atoms their hydro-philic/lipophilic ratios would be the same, and the first compound which hasthe carboxyl group held out of the plane of the ring should, on the basis ofVeldstra's hypothesis, be more active, yet it is inactive.
VELDSTRA and BooiJ (33) attempted to obtain a correlation betweenactivity and the suppression of the polarigraphic oxygen maximum. Theyreasoned that the degree of adsorption of the organic compound to the mer-cury electrode (as measured by the suppression) would parallel its adsorp-tion to a protoplasmic membrane. This ingenious experiment seemed to bea way to measure the hydrophilic/lipophilic coefficient, and it appeared inthis first work that there was some correlation; however, a more extensiveinvestigation (19) using his polarigraphic technique shows no correlationbetween activity and adsorption on the mercury electrode. From this in-
225
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
vestigation and from a general consideration of the great difference in activ-ity of very similar compounds such as those mentioned above, it would seemthat knowledge of the hydrophilic/lipophilic ratio would be of little help inpredicting or explaining relative activity except in extreme cases.
Neither the hypothesis of Veldstra nor that of Wain takes sufficientaccount of the effects of the type or position of the groups attached to thebenzene ring. Neither view explains the effects of the compounds which areinhibitors of growth. The effects of lactones, cysteine, and reagents specificfor the sulfhydryl group have been discussed in an earlier paper (8). Itseems likely that the growth inhibitor, maleic hydrazide, reacts in the samefashion as the lactones by reaction with a sulfhydryl group as follows:
O 011 11C C
CH NH CH2 NHII I + H-S-R I ICH NH R-S-CH NH
C C11 11o 0
'here are many instances of reaction between sulfhydryl groups and acti-vated double bonds. MORGAN and FRIEDMANN (15) have shown that maleicacid will react with a variety of tlhiol compounds and that cysteine reactsin 93%o yield as follows:
0 011 11C C
CH OH CH2 OHII + H-S-CH2-CH-COOH - ICH OH I HOOC-CH-CH2-S-CH OH
c ~~~ ~~~~NH2I \C N CNH2
O 0
Undoubtedly a study of other stable organic compounds with double bondsactivated by groups such as amides, esters, ketones, and nitriles would leadto the discovery of many compounds quite toxic to plant cells.
Another recent study of the structure and activity of growth regulators isthat of LEAPER and BISHOP (13). They have prepared and tested all of thepossible mono-, di-, and trichlorophenoxyacetic acids. Their tests indicatedthat unless the compound contained an open ortho position and in additionan open position para to the ortho position, the molecule was inactive or ofvery low activity. Thus they found that 3,5-dichlorophenoxyacetic acidwas completely inactive. That this compound is inactive has been con-firmed by THIMANN (27) using the pea test, and in the investigations re-
226
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

MUIR AND HANSCH: ACTION OF GROWTH REGULATORS
ported here it was found to be inactive in the Avena section test. Leaperand Bishop have made the reasonable suggestion that the necessity for thetwo open para positions is explained by the oxidation of the growth regu-lator to quinoid compounds in plant cells as follows:
',.CH2COOHO-CH2COOH 0
C1 C1To test this hypothesis 2,5-dihydroxyphenylacetic acid was synthesized andtested for physiological activity. As shown in table II, this compound haslower activity than phenylacetic acid. Since the 2,5-dihydroxy- compoundshould readily form a quinone, the oxidation to quinoid compounds does notappear to be involved in the mechanism of action of phenylacetic acids.Additional evidence against the quinone hypothesis is found in the work ofTHIMANN (26) in which methylbenzoquinonethioacetic acid, trimethylbenzo-quinonethioacetic acid and 1,4-naphthoquinone-2-butyric acid were foundto be inactive. Another objection to the quinone hypothesis is raised bythe fact that 4-chloroindoleacetic acid is more active thain indoleacetic acidas shown in table I. Unless one postulates the displacement of the halogenin this compound, para quinone formation could result only through thedestruction of the five-membered ring with the quinone forming through the5 position and the point at which the nitrogen was attached. The quinonehypothesis fails to explain the activity of 2-methyl-5,7-dichloro-3-indole-acetic acid which has all positions blocked against quinone formation. Alsoa-naphthaleneacetic acid could not form a para quinone unless both ringsare involved. Thus at present it is unlikely that quinone formation explainsthe requirement of an open set of para positions in the phenoxyacetic acids.
THIMANN (26, 27) has advanced some suggestions for the correlation ofstructure and activity of plant growth-regulators. He accepts the view thatthe regulators react chemically with a substrate and states that the reactionoccurs at either the 2, 4, or 6 position on the ring of phenylacetic acids andphenoxyacetic acids with the nucleophilic influence divided between thesethree positions, with the result that no one of them is highly active. Thi-mann is led to the difficult position of this postulate by the observation thatphenoxyacetic acid is inactive and the introduction of one halogen atom inthe 2, 4, or 6 position increases the activity while the introduction of twohalogen atoms as in 2,4-dichlorophenoxyacetic acid increases activity to amaximum. Thus the occupation of one or more of the ortho or para posi-tions is believed to lead to intensification of the nucleophilic influence at theremaining positions. Not only does all theoretical and experimental workin organic chemistry point against such a hypothesis, but close examinationof the structure of active compounds offers little support for it.
227
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
THIMANN (27) has found by means of the pea test that 2,6-dichloro-phenoxyacetic acid is slightly active. A sample of this compound, kindlysupplied by Dr. Thimann, was found to be inactive in the Avena section test.in this investigation. LEAPER and BISHOP (13) also report the compoundto be completely inactive. At most, 2,6-dichlorophenoxyacetic acid must beregarded as of doubtful activity, although on the basis of Thimann's hy-pothesis one would expect it to be highly active. Thimann reports also that2,4,6-trichloro- and 2,4,6-tribromophenoxyacetic acids are slightly active,although all other investigators have found these compounds to be inactive.The suggestion that the activity found may be due to hydrolysis seemsunlikely for two reasons. First, it has been recognized for some time that an-OH group decreases rather than increases activity (27). Secondly, eventhough a chlorine atom were replaced by a less effective hydroxyl group, theposition would still be blocked for chemical reaction by the -OH group.
Thimann states that activation is by electron release to the 2, 4, and 6positions and thus alkyl groups and particularly methyl groups should beeffective in the activation of the phenoxyacetic acids and phenylacetic acids.Considerable evidence exists to show that such activation does not occur.The best procedure is to introduce the group in the acids without any othercomplicating groups attached to the ring. As is shown in table I, o-methyl-,p-methyl-, m-methyl-, 3,5-dimethyl-, 2,4,6-trimethyl-, o-ethyl-, and o-iso-propylphenoxyacetic acids are completely inactive or of low activity. Onthe basis of Thimann's hypothesis it is quite surprising that 2,4-dimethyl-phenoxyacetic acid has a very low activity since it is analogous to the veryactive 2,4-dichlorophenoxyacetic acid. The small activity of the p-methylcompound is also noteworthy. Just as interesting is the case of 2,4-di-methylphenylacetic acid which is shown in table II to be one half as activeas phenylacetic acid. Here the methyl groups actually lower the activity.These examples are compounds substituted with one type of group; withsubstitution of sufficiently strong electron-attracting groups of other typesthe effect could be reversed. So far the only groups which have been foundto confer activity individually on the phenoxyacetic acid ring are the halo-gens and the nitro group.
Thimann points out that resonance in the phenoxyacetic acids increasesthe electron density at the 2, 4, and 6 positions thus accounting for theactivity at these points. The increased electron density is supposed to indi-cate reactivity with a weakly charged positive substrate which does notseem unreasonable for the phenoxyacetic acid series. With the halogenatedbenzoic acids however- a weakly charged positive substrate would be in-capable of successful attack and substitution on the electron-deficient ring.Moreover, according to the hypothesis of Thimann, substitution of the ringof benzoic acid with electron-releasing groups such as -OH, -OR, and-NH2 should be more effective than the introduction of electronegativehalogen atoms, yet, as shown in table II, such substituted benzoic acids arecompletely inactive.
228
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.
MUIR AND HANSCH: ACTION OF GROWTH REGULATORS
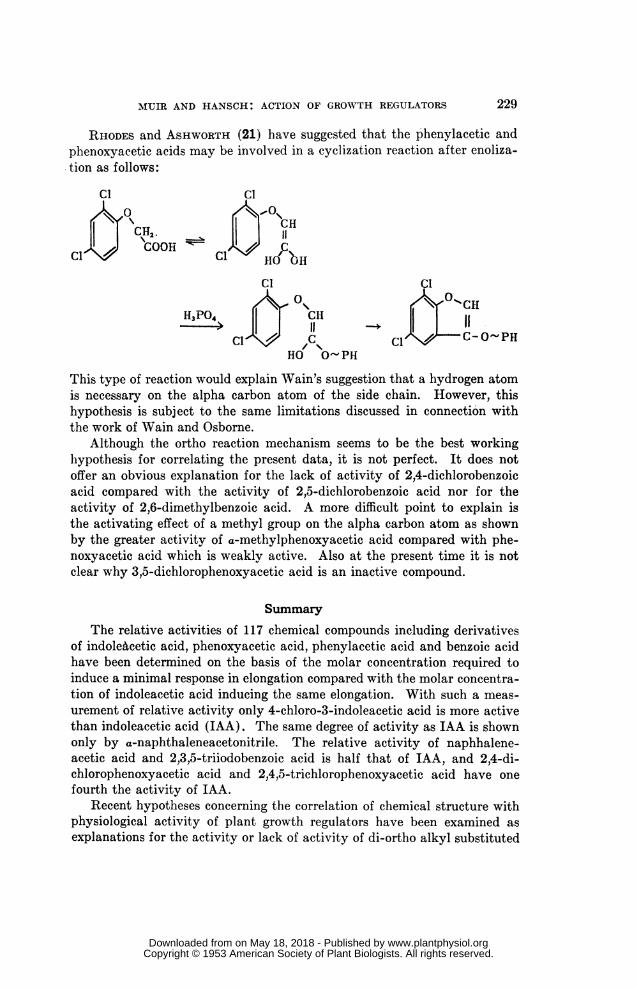
RHODES and ASHWORTH (21) have suggested that the phenylacetic andphenoxyacetic acids may be involved in a cyclization reaction after enoliza-tion as follows:
Cl Cl
CH
Cl\COOH CICF.
Cl Cl
H3P04 CHI
ci,¢ c ci C-O0PHHO o0 PH
This type of reaction would explain Wain's suggestion that a hydrogen atomis necessary on the alpha carbon atom of the side chain. However, thishypothesis is subj ect to the same limitations discussed in connection withthe work of Wain and Osborne.
Although the ortho reaction mechanism seems to be the best workinghypothesis for correlating the present data, it is not perfect. It does notoffer an obvious explanation for the lack of activity of 2,4-dichlorobenzoicacid compared with the activity of 2,5-dichlorobenzoic acid nor for theactivity of 2,6-dimethylbenzoic acid. A more difficult point to explain isthe activating effect of a methyl group on the alpha carbon atom as shownby the greater activity of a-methylphenoxyacetic acid compared with phe-noxyacetic acid which is weakly active. Also at the present time it is notclear why 3,5-dichlorophenoxyacetic acid is an inactive compound.
SummaryThe relative activities of 117 chemical compounds including derivatives
of indoleAcetic acid, phenoxyacetic acid, phenylacetic acid and benzoic acidhave been determined on the basis of the molar concentration required toinduce a minimal response in elongation compared with the molar concentra-tion of indoleacetic acid inducing the same elongation. With such a meas-urement of relative activity only 4-chloro-3-indoleacetic acid is more activethan indoleacetic acid (IAA). The same degree of activity as IAA is shownonly by a-naphthaleneacetonitrile. The relative activity of naphhalene-acetic acid and 2,3,5-triiodobenzoic acid is half that of IAA, and 2,4-di-chlorophenoxyacetic acid and 2,4,5-trichlorophenoxyacetic acid have onefourth the activity of IAA.
Recent hypotheses concerning the correlation of chemical structure withphysiological activity of plant growth regulators have been examined asexplanations for the activity or lack of activity of di-ortho alkyl substituted
229
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
analogues and the low activity or inactivity of mono-alkyl substituted ana-logues. It is concluded that the two-point ortho reaction mechanism affordsthe best working hypothesis although a few compounds at present do notseem to fit the hypothesis.
This investigation was made possible by National Science FoundationGrant no. 45. The authors acknowledge their gratitude to Professor K. V.Thimann of the Biological Laboratories, Harvard University, for the sampleof 2,6-dichlorophenoxyacetic acid; to Professor R. C. Fuson of the Depart-ment of Chemistry, University of Illinois, for the samples of 2,4,6-triethyl-benzoic acid, 2,6-dimethyl-4-methoxybenzoic acid, and 2-methoxy-3,5-di-chloro-4,6-dimethylbenzoic acid; to Professor R. L. Wain of Wye College,University of London, for the sample of a-phenoxypropionic acid; to Pro-fessor C. Vanderwerf of the Department of Chemistry, University of Kansas,for the samples of o-, m-, and p-fluorobenzoic acids; to Dr. J. Van Overbeekof the Agricultural Laboratory of Shell Oil Company for the sample ofpentachlorobenzoic acid; and to V. Prelog of the Federal Institute of Tech-nology, Zurich, for the sample of 3,4,5-triiodobenzoic acid.
LITERATURE CITED1. BENTLEY, J. A. Growth-regulating effect of certain organic compounds.
Nature 165: 449-450. 1950.2. BONNER, J. Limiting factors and growth inhibitors in the growth of the
Avena coleoptile. Amer. Jour. Bot. 36: 323-332. 1949.3. BUNNETT, J. F. and ZAHLER, R. E. Nucleophilic substitution reactions.
Chem. Rev. 49: 293-322. 1951.4. CURRIER, H. B. and CRAFTS, A. S. Maleic hydrazide a selective herbi-
cide. Science 111: 152-153. 1950.5. GILMAN, H. Organic Chemistry. Second Edition, Vol. 1, pp. 256-264.
John Wiley, New York. 1943.6. GOODWIN, R. H. and TAVES, C. The effect of coumarin derivatives on
the growth of the Avena coleoptile. Amer Jour. Bot. 37: 224-231.1950.
7. HANSCH, C. and MUIR, R. M. The ortho effect in plant growth-regu-lators. Plant Physiol. 25: 389-393. 1950.
8. HANSCH, C., MUIR, R. M., and METZENBERG, R. L. Further evidencefor a. chemical reaction between plant growth-regulators and aplant substrate. Plant Physiol. 26: 812-821. 1951.
9. HQFFMANN, 0. L. and SMITH, A. E. A new group of plant growth-regu-lators. Science 109: 588. 1949.
10. JONES, E. R. H., HENBEST, H. B., SMITH, G. F., and BENTLEY, JOYCE A.3-Indolylacetonitrile, a naturally-occurring plant growth hormone.Nature 169: 485-487. 1952.
11. KOEPFLI, J. B., THIMANN, K. V., and WENT, F. W. Phytohormones:Structure and physiological activity I. Jour. Biol. Chem. 122:763-780. 1938.
230
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

MUIR AND HANSCH: ACTION OF GROWTH REGULATORS
12. KOGL, F. and VERKAAIK, B. XXXVIII. Physiological activity of theoptical isomers of a- (B'-indolyl) propionic acid. Zeit. Physiol.Chem. 280: 167-176. 1944.
13. LEAPER, J. M. F. and BISHOP J. R. Relation of halogen position tophysiological properties in the mono-, di- and trichlorophenoxy-acetic acids. Bot. Gaz. 112: 250-258. 1951.
14. LEOPOLD, A. C. and KLEIN, W. H. Maleic hydrazide as an antiauxinin plants. Science 114: 9-10. 1951.
15. MORGAN, E. J. and FRIEDMANN, E. Interaction of maleic acid withthiol compounds. Biochem. Jour. 32: 733-742. 1938.
16. MUIR, R. M. and HANSCH, C. The relationship of structure and plantgrowth activity of substituted benzoic and phenoxyacetic acids.Plant Physiol. 26: 369-374. 1951.
17. .vIUIR, R. M., HANSCH, C. H., and GALLUP, A. H. Growth regulationby organic compounds. Plant Physiol. 24: 359-366. 1949.
18. OSBORNE, D. J. and WAIN, R. L. Plant growth-regulating activity incertain aryloxyalkylcarboxylic acids. Science 114: 92-93. 1951.
19. PALEG, L. G. and MUIR, R. M. Surface activity as related to physio-logical activity of plant growth-regulators. Plant Physiol. 27: 285-292. 1952.
20. REMICK, A. E. Electronic Interpretations of Organic Chemistry. Sec-ond Edition, pp. 408-422. John Wiley, New York. 1949.
21. RHODES, A. and ASHWORTH, R. DE B. Mode of action of growth regu-lators in plants. N'ature 169: 76-77. 1952.
22. RUDMAN, SYLVIA V. A comparison of the effects upon plant growth of2,3,5-triiodobenzoic acid and indole-3-acetic acid. M.S. Thesis.State University of Iowa, Iowa City, Iowa. 1951.
23. SHRINER, R. L., ADAMS, R., and MARVEL, C. S. Stereoisomerism. In:Organic Chemistry, H. Gilman, Editor. First Edition, Vol. I, p.268. John Wiley, New York. 1938.
24. SMITH, M. S., WAIN, R. L., and WIGHTMAN, F. Antagonistic action of.certain stereoisomers on the plant growth-regulating activity oftheir enantiomorphs. Nature 169: 883-884. 1952.
25. SNYDER, W. E. Some responses of plants to 2,3,5-triiodobenzoic acid.Plant Physiol. 24: 195-206. 1949.
26. THIMANN, K. V. The synthetic auxins: Relation between structure andactivity. In: Plant Growth Substances, F. Skoog, Ed., pp. 21-36.University of Wisconsin Press, Madison. 1951.
27. THIMANN, K. V. The role of ortho-substitution in the synthetic auxins.Plant Physiol. 27: 392-404. 1952.
28. THIMANN, K. V. and BONNEIR, W. D. Experiments on the growth andinhibition of isolated plant parts. II. The action of several enzymeinhibitors on the growth of the Avena coleoptile and on Pisuminternodes. Amer. Jour. Bot. 36: 214-221. 1941.
29. THIMANN, K. V. and BONNER, W. D. Experiments on the growth andinhibition of isolated plant parts. I. The action of iodoacetate and
231
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
organic acids on the Avena coleoptile. Amer. Jour. Bot. 35: 271-281. 1948.
30. VELDSTRA, H. Researches on plant growth substances., IV. Relationbetween chemical structure and physiological structure. I. Enzy-mologia 11: 97-136. 1944.
31. VELDSTRA, H. On the growth-substance activity in the series of halo-genated benzoic acids and related compounds. Reported at theTwelfth International Congress of Pure and Applied Chemistry,New York. 1951.
32. VELDSTRA, H. Some chemical aspects of growth-substance action. Re-ported at the Seventh International Botanical Congress, Stockholm.1950.
33. VELDSTRA, H. and Booij, H. L. Researches on plant growth regulators.XVII. Structure and activity. On the mechanism of action. III.Biochem. Biophys. Acta. 3: 278-312. 1949.
34. VELDsTRA, H. and HAVINGA, E. On the physiological activity of unsatu-rated lactones. Enzymologia 11: 373-380. 1949.
35. WAIN, R. L. Plant growth regulating and systemic fungicidal activity:The aryloxyalkylcarboxylic acids. Jour. Science of Food and Agr.3: 101-106. 1951.
36. WAIN, R. L. Chemical structure in relation to fungicidal and plantgrowth-regulating activity of the aryloxy acids. ,Reported at theSeventh International Congress of Pure and Applied Chemistry,New York. 1951.
37. WHELAND, G. W. Advanced Organic Chemistry. Second Edition, pp.378-386. John Wiley, New York. 1949.
38. ZIMMERMAN, P. W. and HITCHCOCK, A. E. Growth-regulation effectsof chloro-substituted derivatives of benzoic acid. Contr. BoyceThompson Inst. 16: 209-213. 1951.
39. ZIMMERMAN, P. W. and WILCOXON, F. Several chemical growth sub-stances which cause initiation of roots and other responses inplants. Contr. Boyce Thompson Inst. 7: 209-229. 1935.
40. ZUKEL, J. W. Use of maleic hydrazide as a plant growth-inhibitor.Agr. Chem. 5: 35-36, 84-85. 1950.
232
www.plantphysiol.orgon May 18, 2018 - Published by Downloaded from Copyright © 1953 American Society of Plant Biologists. All rights reserved.