protein kinaseasuppresses srebp-1c · pdf fileprotein kinaseasuppresses srebp-1c expression...
TRANSCRIPT

1
PROTEIN KINASEA SUPPRESSES SREBP-1CEXPRESSION VIA PHOSPHORYLATION OF LXR IN THE
LIVER.Takashi Yamamoto1, *Hitoshi Shimano1,2, Noriyuki Inoue1, Yoshimi Nakagawa1,2,Takashi Matsuzaka1,2, Akimitsu Takahashi1,2, Naoya Yahagi2, Hirohito Sone1,2,
Hiroaki Suzuki1, Hideo Toyoshima1, and Nobuhiro Yamada1
From 1Department of Internal Medicine (Metabolism and Endocrinology),
Graduate School of Comprehensive Human Sciences, 2Center for Tsukuba
Advanced Research Alliance,Running Title: PKA Inhibits LXR Signaling
Address correspondence to Hitoshi Shimano, Department of Internal Medicine (Metabolism andEndocrinology), Graduate School of Comprehensive Human Sciences, 2 Center for Tsukuba AdvancedResearch Alliance, University of Tsukuba, 1-1-1 Tennodai, Tsukuba, Ibaraki, 305-8575, Japan; Fax. +81-29-853-317 E-Mail: [email protected]
SREBP-1c is a transcription factor thatcontrols synthesis of fatty acids andtriglycerides in the liver, and is highlyregulated by nutrition and hormones. In thecurrent studies, we show that PKA, a mediatorof glucagon/cAMP, a fasting signaling,suppresses SREBP-1c by modulating theactivity of LXRa, a dominant activator ofSREBP-1c expression. Activation of PKArepressed LXR-induced SREBP-1c expressionboth in rat primary hepatocytes and mouselivers. Promoter analyses revealed that theLXRa-binding site in the SREBP-1c promoteris responsible for PKA inhibitory effect onSREBP-1c transcription. In vitro and in vivo,PKA directly phosphorylated LXRa and thetwo consensus PKA-target sites (195,196serines and 290,291 serines) in its ligand-binding/heterodimerization domain werecrucial for the inhibition of LXR signaling.PKA phosphorylation of LXRa causedimpaired DNA binding activity by preventingLXRa/RXR dimerization, and decreased itstranscription activity by inhibiting recruitmentof co-activator SCR-1, and enhancingrecruitment of co-repressor NcoR1. Theseresults indicate that LXRa is regulated notonly by oxysterol derivatives, but also by PKA-mediated phosphorylation, which suggests thatnutritional regulation of SREBP-1c andlipogenesis could be regulated at least partiallythrough modulation of LXR.
In mammals, carbohydrates are an essentialenergy resource. When consumed in excess,carbohydrates are converted to lipids by lipogenicenzymes in preparation for times of energydeficiency. These processes are known to beregulated at the transcription level and severaldifferent transcription factors are known tocontribute to this regulation. SREBP (SterolRegulatory Element Binding Protein), atranscription factor belonging to the basic helix-loop-helix-leucine zipper family, regulatestriglyceride synthesis and cholesterolmetabolism(1). Three different SREBP isoformshave been identified. SREBP-1c plays a roleprimarily in triglyceride synthesis(2), whileSREBP-2 regulates cholesterol metabolism(3).SREBP-1a, is expressed in growing cells andactivates an entire array of genes involved in bothtriglyceride and cholesterol synthesis(4). In liver,SREBP-1c controls transcription of genesinvolved in fatty acid and triglyceridesynthesis(2,5). SREBP-1c expression is highlyregulated in response to the nutritional condition.Expression of SREBP-1c is undetectable duringfasting, while its expression is strongly induced ina fed state, followed by similar adapted nutritionalchanges in lipogenic genes(6).To understand the molecular mechanism of
nutritional regulation of SREBP-1c expression,the SREBP-1c gene promoter has beenextensively studied. In the proximal region ofthe mouse SREBP-1c promoter, Sp1, NF-Y, USF,SREBP, and LXR binding sites have beenidentified(7-9). It has been observed thatinsulin(10) and glucose(11,12) are factors that
http://www.jbc.org/cgi/doi/10.1074/jbc.M611911200The latest version is at JBC Papers in Press. Published on February 12, 2007 as Manuscript M611911200
Copyright 2007 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
induce hepatic SREBP-1c in vivo and in vitro,although precise mechanisms are yet to beelucidated. Adenoviral overexpression in ratprimary hepatocytes suggested that the signaltransducer, akt, could be a potential regulator ofSREBP-1c transcription(13).Liver X receptors (LXR) belong to a nuclear
receptor superfamily. The LXR subfamilyconsists of two members, LXRa and LXRbwhich are activated by oxysterols(14). Theexpression pattern of LXRa is restricted mainlyto liver, adipocytes, small intestine, andmacrophages, while LXRb is expressedubiquitously. Although early reports revealedinvolvement of LXRs in cholesterolhomeostasis(15,16), recent studies suggest thatLXR negatively regulates gluconeogenesis(17)and inflammatory responses(18,19). Whileinvestigating the pharmacological effect of LXRin rodent models, it has been observed that LXRligands are protective against the development ofatherosclerotic lesions (20) and amelioratesconditions of high blood glucose and impairedglucose tolerance(21). LXRs directly bind cis-element on the SREBP-1c promoter asheterodimers with RXR leading to transcriptionalactivation(8). Several studies have establishedLXRs as dominant activators of SREBP-1cexpression. LXRa/b double knockout micerevealed dramatically reduced expression ofSREBP-1c(9). We have used an expressioncloning strategy to show that LXRs are theprimary activators of the SREBP-1c promoter(8).Polyunsaturated fatty acids (PUFA) are the onlyknown dietary lipid capable of negativelyregulating hepatic SREBP-1c expression andlipogenesis(22). A portion of these effects aremediated at the transcriptional level throughrepression of LXR activity(23).Considering that hepatic SREBP-1c
expression is dominated by LXRs and eliminatedby fasting, it is probable that there is a mechanismby which LXR mediates the repressed SREBP-1cexpression during fasting. Consistently, it hasbeen reported that glucagons and its signalmediator, cAMP suppresses SREBP-1c in ratprimary hepatocytes(24). Protein kinase A(PKA) is a cAMP dependent protein kinase whichconsists of both a catalytic and regulatory subunitand regulates numerous cellular functions ineukaryotic cells by phosphorylating targetproteins. In regard to energy metabolism, PKAis subordinated to glucagon and adrenalin andtherefore is classically recognized as a fastingsignal to activate gluconeogenesis and b-oxidation, and to oppose triglycerides synthesis
and glucose utilization. It is known that severalnuclear receptors (e.g. ER, RAR, PPAR HNF-4a )are phosphorylated by PKA leading tomodification of their trans-activities via diversemechanism(25-28). In the current study, weinvestigated effects of cAMP / PKA on SREBP-1cexpression and LXR signaling system.
Experimental ProceduresMaterialsAnti-LXRa, anti-RXR and anti-SREBP-1antibodies were purchased from Santa CruzBiotechnology Inc. (Santa Cruz, CA). Anti-CREB and anti-phospho-CREB (Ser 133)antibodies were purchased from Cell SignalingTechnology Inc. (Beverly, MA). Anti-His 5antibody was purchased from Qiagen Inc.(Maryland, USA). Anti-HA and anti-mycantibodies were purchased from Roche. Anti-Flag antibody was purchased from Promega(Madison, WI). Dibutylyl cAMP was purchasedfrom Promega. T0901317 as synthetic LXRligand was purchased fromWAKO.Animals.Male mice (C57BL/6J) were obtained fromCLEA Japan (Tokyo, Japan). For fasting andrefeeding, mice were fasted for 24 h and then feda high sucrose /low fat diet for 12 h.Isolation and culture of hepatocytes.Primary hepatocytes were isolated form maleSprague-Dawley rats (200-300g) (Japan Clea,Tokyo, Japan). Cell were resupsended inDMEM containing penicillin and streptomycinsupplemented with 10 % fetal bovine serum(FBS) before being seeded on 100-mm collagen-coated dishes at 2x106 cells /dish. Afterincubation for 4 h to allow attachment, mediumwas replaced and experiments were performed.Northern blottingTotal RNA from mouse liver and rat primaryhepatocytes was isolated as describedpreviously(29). The cDNA probes for mouseSREBP-1, PEPCK, acidic ribosomalphosphoprotein PO (36B4), ABCA1 andLXRa were prepared as previouslydescribed(8,29,30).PlasmidsA series of mouse SREBP-1c promoter linked topGL2 basic were previously described(7).LXRE-enhancer construct linked to pGL2promoter vector was previously described(8).Human ABCA1 promoter (-919-+239) linked topGL3 basic were previously described(30).Expression vector for Gal4-DBD- LXRa LBDfusion protein and Gal4 RE Luc vector werepreviously described(8). Expression vectors for
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
mouse LXRa (wild type, 195 196 AA, 290 291AA) were constructed in pcDNA3.1(+).Expression vector for PKA c subunit wasconstructed in pFA. Expression vector for V5-tagged PKA regulatory subunit (dominantnegative form) was constructed in pcDNA3.1(-)according to previous report for mutated form ofthe regulatory type 1 subunit (RI) of PKA(31).Expression vector for HA-tagged human LXRawas constructed in pcDNA3. Expression vectorsfor Flag-tagged wild type human RXR and Flag-tagged mutant RXR in which 27-Serine residuewas substituted to alanine were constructed inpcDNA3.1(+). Expression vectors for Histidine-tagged recombinant mouse LXRa (Full length, 1-163 a.a., 162-326 a.a., 325-445 a.a.) wereconstructed in pET28a(+). Expression vector forGlutathione-S-transferase fusion protein wasconstructed in pGEX4T-2. For mammalian twohybrid system, mouse LXRa coding region wasligated into pACT. Mouse SRC-1 nuclearreceptor interaction domain (568-779 a.a.) andmouse NcoR1 nuclear receptor interaction domain(1944-2453 a.a.) were ligated into pM. For invitro translation, SRC-1 NID (576-779 a.a.) wasligated into pcDNA3.1(+). Expression vectorfor Flag-tagged CBP was previously described.Expression vector for HA-tagged mouse CREBwas constructed in pcDNA3.Transfections and Luciferase assay293 cells and 293T cells were grown in DMEMcontaining penicillin and streptomycinsupplemented with 10% FBS at 37 ºC in 24 wellplate overnight before transfection. Cells weretransfected with reporter vector, expression vectorusing Fugene 6. Total amounts of DNA wereadjusted to 0.5 mg /well using empty vector.After 24 h incubation, the amounts of fireflyluciferase activity in transfectants were measured.Firefly luciferase activity was normalized by theamounts of renilla luciferase activity expressedfrom CMV or SV40 promoter linked-renillaluciferase unless otherwise indicated.In vitro kinase assayHistidine-tagged recombinant mouseLXRa proteins were expressed in Escherichiacoli (BL21, DE3) and purified using standardtechniques and purification kit Mag Extractor(TOYOBO) according to the manufacture’sprotocol. Briefly, bacterial culture containingkanamycin were grown at 37 ºC. After inductionof recombinant proteins for 2-3 h, cells wereresuspended in lysis buffer (6 M guanidinehydrochloride, 5 M NaCl, pH 8.0) and lysed bysonication. The centrifuged supernatant wasmixed for 30 min at room temperature on a rotator
with magnetic Ni beads. Protein-absorbed beadswere washed with lysis buffer and recombinantLXRa proteins were eluted with elution buffer.After dialysis to exclude guanidine, theconcentration and size of histidine-tagged proteinswere estimated by SDS-PAGE followed byCoommasie Blue staining and immunoblottingusing a known quantity of molecular weightstandards. The purified proteins were stored at -80 ºC until experiments were performed.Bovine purified PKA catalytic subunit waspurchased from Promega. PKA c subunit,histidine-tagged LXRa and [g-32P]ATP weremixed in PKA buffer (50 mM Tris HCl (pH 7.5),10 mM NaCl, 10 mM MgCl2, 1 mM DTT) andincubated for 45 min at 30 ºC. Samples wereanalyzed by SDS-PAGE and phosphorylation wasvisualized by autoradiography. Quantity ofHistidine-tagged protein was confirmed byimmunoblotting using anti-His 5 antibody.Histone H1 protein (Calbiochem) was used aspositive control in kinase assays.In vivo kinase assayFor in vivo kinase assay, COS7 cells prepared
in 100-mm collagen-coated dishes at 1x106
cells/dish were transfected with control orexpression vector for HA-tagged human LXRavectors. After transfection, the cells werestarved in phosphate-deficient medium for 12 hand then incubated for 2 h in the same mediumcontaining 500 mCi /ml [32P]orthophosphate.Cells were treated with PKA activators (forskolin10 mM, dibutylyl cAMP 1 mM, IBMX 1 mM) for30 min before harvesting.At the end of the labeling period, the cells were
washed ice-cold PBS, harvested, lysed on ice.Protein samples were extracted in lysis buffermentioned elsewhere and centrifuged 15000 rpm4 C for 10 min. Supernatants were subjected toimmunoprecipitation assay used with anti HA-antibody (Roche) as previously described. Theimmunocomplex were eluted in Sample buffer,resolved by SDS-PAGE (8% gel), visualized withautoradipgraphy and immunoblotted.Electrophoretic mobility shift assayNuclear extracts were prepared from livers aspreviously described(32). RXR protein wasgenerated from expression vector using a coupledin vitro reticulocyte transcription/ translationsystem (Promega). Double-strandedoligonucleotides of LXR response element inSREBP-1c promoter using in EMSA wereprepared as previously described. In vitrosynthetic protein lysate or nuclear extracts(approximately 1 mg) were incubated. DNA-protein complex were resolved on a 4%
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
polyacrylamide gel.ChIP assayThe chromatin immunoprecipitation (ChIP)assay was conducted with a chromatinimmunoprecipitation kit (Upstate Biotechnology,Lake Placid, NY). After H2.35 cells weretreated PKA activators (forskolin 10 mM,dibutylyl cAMP 100 mM) or vehicle underT0901317 (10 mM) for 1 h at 37 °C, H2.35 cellswere cross-linked for 10 min by addingformaldehyde directly to culture medium to afinal concentration of 1 %. Cross-linked cellswere washed ice-cold PBS containing proteaseinhibitors, scraped, pelleted, resuspended in 200ml SDS lysis buffer (1% SDS, 10 mM EDTA, 50mM Tris-HCl, pH 8.0) per 1x106 cells andincubated for 10 min on ice. The lysates werethen sonicated 30 s each by sonicator (sonifier250, BRANSON) with settings of duty cycle:20%, output power: 10%. The samples were onice for 1 min on ice between the cycles. Aftersonication, the samples were centrifuged andsupernatant were diluted 10-fold in ChIP dilutionbuffer containing protease inhibitors. 2 mLdilutants were precleared with 75 ml salmonsperm DNA /protein A spharose-50% slurry for 30min 4 °C and then incubated overnight with 3 mganti-RXR antibody or control IgG. Antibody-protein-DNA complexes wereimmunoprecipitated with 60 ml protein A.After intensive washing, pellets were eluted byfreshly prepared elution buffer (1% SDS, 0.1 MNaHCO3). Formaldehyde cross-linking wasreversed by overnight incubation at 65 °C afteradding 20 ml 5 M NaCl. Samples were purifiedwith phenol: chloroform: isoamyl alcohol andprecipitated with ethanol. DNA pellets weredissolved with TE buffer and used as templates inPCR. Each 8 ml of 20 ml DNA solution was usedfor PCR amplification (1st step is 95 C for 3 min(one cycle), 2nd step is 94 C for 1 min, 62.1 °C for30 s, 72 °C for 1 min (25 cycles)).ChIP primers: 5’-GAACCAGCGGTGGGAACACAGAGC-3’ and5’-GACGGCGGCAGCTCGGGTTTCTC.Immunoblotting and immunoprecipitationCOS7 cells were seeded on 100-mm dishes andtransfected with the expression vectors. Afterincubation for 48 h, dbcAMP (100 mM), forskolin(10 mM), and IBMX (100 mM) were added intomedium for 30 min and then cells were harvested.Protein lysate was extracted in lysis buffer (25mM HEPES (pH 7.9), 50 mM KCl, 6 % glycerol,5 mM MgCl2, 0.5 % TritonX 100, 1mM DTT, 50mM NaF, 40 mM b-glycerophosphate, 25 mMsodium pyrophosphate, protease inhibitor cocktail
(Roche)) on ice 20 min and centrifuge at 15000rpm at 4 ºC for 10 min. Supernatants weresubjected to immunoprecipitation assay aspreviously described(29).GST pulldown assayGST and GST-LXRa proteins were preparedusing standard techniques according to themanufacture’s protocol. Briefly, Escherichiacoli (BL21, DE3) transformed with the GST orGST-LXRa expression vector, were incubatedbefore induction at room temperature for 4 h with0.4 mM IPTG. The cells were harvested bycentrifugation and suspended in buffer A (25 mMHEPES (pH 7.9), 50 mM KCl, 6 % glycerol, 5mM EDTA, 5 mM MgCl2, 0.5 % TritonX 100,protease inhibitor cocktail, 1 mM DTT). Thecells were lysed by sonication and cellular debriswas removed by centrifugation at 15000 rpm at 4C for 10 min. Recombinant GST or GST-LXRaproteins were purified by glutathione sepharosebeads according to the manufacture’s protocol(Amersham Pharamcia Biotech). GST fusionproteins were eluted (50 mM Tris HCl (pH 8.0),50 mM NaCl, 10 mM Glutathione, proteaseinhibitor cocktail, 1 mM DTT). Amounts andsizes of GST or GST-LXRa proteins wereestimated by SDS-PAGE followed Commasieblue staining and immunoblotting using anti-LXRantibody. GST or GST-LXRa and PKA csubunit or equal amount BSA were mixed in PKAbuffer ( 50 mM Tris HCl (pH 7.5), 10 mM NaCl,10 mM MgCl2, 1 mM DTT, 50mM NaF, 20mMb-glycerophosphate) and incubated for 45 min at30 C. After adding Gluthatione beads and 35S-radio-labeled SRC-1 nuclear receptor interactiondomain (576-779 a.a.) synthesized in vitro tosamples and incubated for 2 hr at 4 ºC, beadswere washed three times with ice-cold Phosphatebuffed saline containing 0.1% TritonX 100.Proteins bound to beads were eluted with elutionbuffer (50 mM Tris-HCl (pH 8.0), 50 mMglutathione), resolved by SDS-PAGE andvisualized autoradiography.
Results
PKA represses expression of SREBP-1 in vivo.Expression of SREBP-1c in the liver is highlyregulated by nutrition; suppressed at fasting andinduced in a refed state as previously reported (6).One potential mechanism of this suppression atfasting could be mediated through cAMP/ ProteinKinase A (PKA) system that mediates theglucagon signal. C57BL/6J mice in a refed statewere administrated with dibutyl cAMP andtheophylline for activation of PKA to mimic
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
fasting signal. The SREBP-1c mRNA that washighly induced by refeeding was robustlyrepressed by cAMP/ theophylline administration(Figure 1A). Reciprocally, PEPCK, a gene thatcontrols gluconeogenesis and a well known targetof glucagons/cAMP/PKA signal, was suppressedby refeeding, but restored by these PKA activators.The reduction of SREBP-1 mRNA level by PKAactivation was associated with a robust decreasein active SREBP-1c protein in hepatic nuclearextracts (Figure 1B). SREBP-1c expression isdominantly controlled by LXR/RXR that bindsdirectly to the two LXREs in the SREBP-1cpromoter(8,9). As shown in Figure1B, theexpression of LXR/RXR was not affected by thePKA activation. Therefore, SREBP-1csuppression by PKA was not due to reduction inthe amounts of LXR/RXR.PKA activation represses LXR-mediatedSREBP-1 induction.The effect of PKA on LXR activation ofSREBP-1c was estimated. In rat primaryhepatocytes, SREBP-1c mRNA level wasessentially undetectable, but was highly inducedby T0901317, an LXR agonist. This inductionwas substantially suppressed by addition ofdibutyl cAMP while expression of LXR and RXRwas not increased at the mRNA level. (Figure 2).These data suggest that PKA repression ofSREBP-1c was mediated not through changes inLXR expression, but through modulating the LXRactivity. Meanwhile, LXR agonist-inducedexpression of ABCA1 (ATP binding cassettetransporter A1), an LXR target gene was onlyslightly affected by PKA activation. LXREresponse element is responsible for PKAinhibition of SREBP-1c promoter.To further investigate the PKA effect on SREBP-1 transcription, in vitro reporter assays forSREBP-1c promoter were performed.Luciferase reporters linked to different sizes ofSREBP-1c promoter region around the LXR-binding sites (LXREs) and expression vectors ofLXRa and PKA catalytic subunit for PKAactivation were transfected into HepG2 cells(Figure 3A). PKA expression did not affect thebasal level of SREPB-1c promoter activity inHepG2 cells. Meanwhile, PKA co-expressionstrongly suppressed LXRa-mediated activation ofSREBP-1c promoter. This was only observed inLXRE-containing promoters (-2.6 k and -550 bp).The shorter construct lacking LXREs (90bp) didnot show LXR activation or PKA inhibition.Thus, the inhibitory effect of PKA on SREBP-1cpromoter was conceivably mediated throughLXREs. This was supported by the LXREenhancer construct assays, demonstrating that
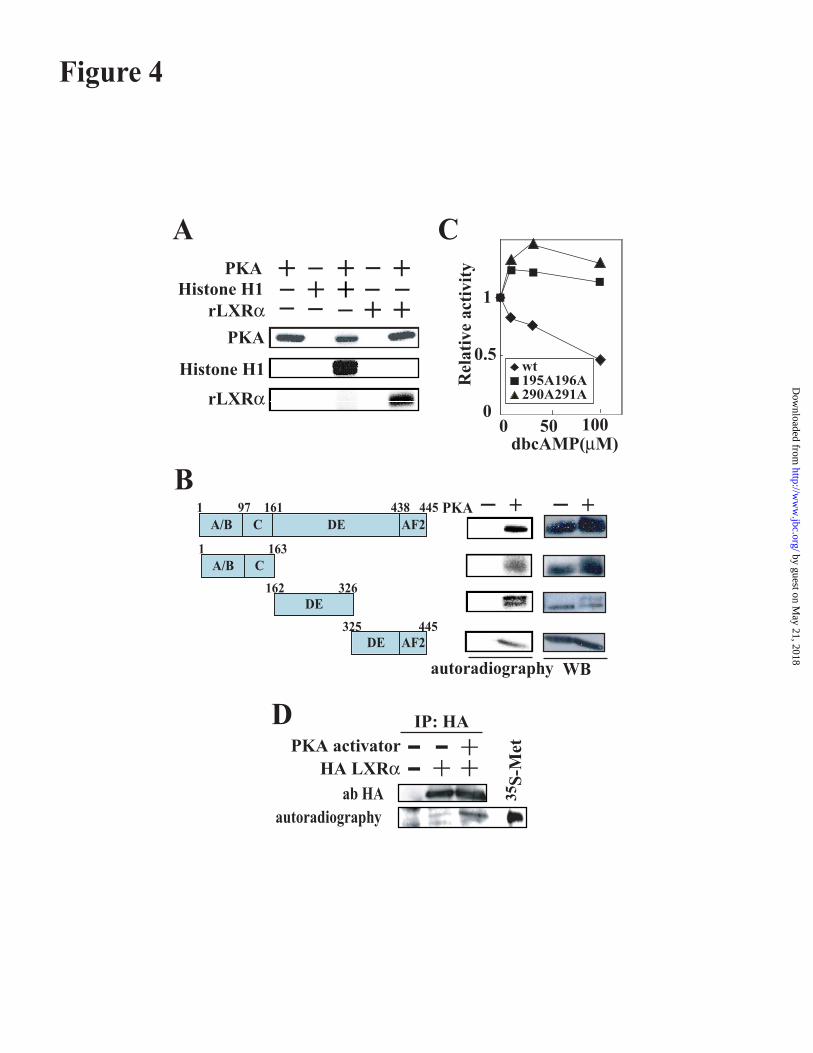
endogenous PKA activation by forskolin, anadenylate cyclase activator, dose-dependentlyinhibited LXRE enhancer activity induced by theLXR ligand and this inhibitory effect wasabolished by co-expression of PKA regulatorysubunit dominant negative form (Figure. 3B).The mechanism against the PKA suppression ofLXR activity was further explored the using Gal4DBD protein fused to LXRa ligand bindingdomain (LBD) in HepG2 (Figure 3C). LXR-LBD was highly activated in this system by theLXR synthetic ligand as previously reported.This activation was completely abolished by co-transfection of PKA, suggesting that PKAactivation inhibits ligand-mediated activation ofLXR. PKA-mediated inhibition of LXRtranscriptional activity was supported byluciferase assays using the promoter of ABCA1,another LXR target gene. Mouse ABCA1promoter activity was also dose-dependentlyinhibited by PKA expression in both basal andLXR-induced conditions (Figure.3D).LXRa is phosphorylated by PKA in vitro.These data led us to speculate that PKAmodulates LXR activity, most likely throughdirect phosphorylation of LXRa. To test thishypothesis, in vitro kinase assays of LXRa wereperformed. Purified recombinant protein ofHistidine-tagged LXRa was incubated with thePKA catalytic subunit. LXRa protein wasphosphorylated by PKA as well as PKAautophosphorylation and Histone H1phosphorylation as positive controls of PKAtarget protein (Figure 4A). Functional domainsof the LXRa protein were also tested using this invitro kinase assay (Figure 4B). Every domain ofLXRa was significantly phosphorylated. In theamino acid sequence of PKA, there are twopotential PKA-target sites (195,196 serines and290,291 threonine,serine) in theLBD/heterodimerization domain. Introductionof mutation in which these serine residues werereplaced by alanine caused resistance to PKAinhibition of SREBP-1c promoter activation(Figure 4C). To assess whether LXRa isphosphorylated by PKA in vivo, COS7 cells weretransfected with expression vector for HA-taggedLXRa and subsequently labeled with[32P]orthophosphate. After treatment with PKAactivators, the phosphorylation state of LXRawas examined by autoradiography. As evidentfrom autoradiogaphy and immunoblots (Figure4D), PKA stimulation induced thephosphorylation state of LXRa. These resultsindicate that PKA directly phosphorylates LXRprotein, which is crucial for PKA inhibition of
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
LXR activity.DNA binding activity of LXR/RXRheterodimer was decreased by PKAphosphorylation of LXRa.The effect of LXRa phosphorylation by PKA onDNA binding was tested by EMSA assays (Fig. 5A). Recombinant LXRa protein requires RXRfor its binding to LXRE. The signal ofLXR/RXR bound to LXRE was partiallydecreased by PKA treatment of LXR. Thiseffect required the incubation of LXR with PKAat 30 °C, indicating the PKA inhibition wasmediated through its kinase activity. To confirmthis effect in vivo, nuclear extracts of livers fromdbcAMP/theophylline-administrated mice wereprepared and subjected to EMSA assay (Figure5B). PKA activation in these nuclear extractswas confirmed by phosphorylation of CREB, awell known PKA target in mouse livers, as shownby immunoblot analysis. EMSA of these nuclearextracts demonstrated that the signal of LXREbinding mainly from LXR/RXR as confirmed bysupershift with LXR and RXR antibodies wasdose-dependently diminished by PKAadministration. Next, to determine whetherPKA activation reduce DNA binding activity ofLXR/RXR heterodimer, ChIP assay analysis onextracts from H2.35 cells, a mouse hepatocyte-derived cell line was conducted. Anti-RXRantibody for immunoprecipitation of LXR/RXRheterodimer, and primers to detect LXRE onSREBP-1c promoter for PCR were used. Asshown in Figure 5C, the interaction of LXR/RXRheterodimer with LXRE was decreased uponPKA activation in vivo. These data suggest thatthe modification of LXR activitiy by PKA isattributed to reduction of DNA binding activity ofLXR/RXR heterodimer via PKA phosphorylationof LXRa.Coactivator or corepressor recruitments ofLXRa are reciprocally modified by LXRaphosphorylation by PKA.It was reported that in LXRa or b-mediatedtranscription of SREBP-1c, SRC-1 works as acoactivator and NcoR1 as a corepressor,respectively(33). To test the possibility ofinvolvement of these cofactors in PKA inhibitionof LXR activity, mammalian two hybrid assayswere preformed. The system detects the bindingof Gal4 AD-LXRa fusion protein and Gal4-SRC-1-NID (a functional region that interacts withnuclear receptors(34)) in 293 cells. Addition ofthe LXR ligand markedly activated the signal, asevidenced by recruitment of SRC-1 to LXR.PKA coexpression markedly and dose-dependently inhibited the recruitment of SRC-1
(Figure 6A). Inversely, PKA coexpressionincreased the recruitment of NcoR1-NID to AD-LXRa (Figure 6B). Inhibition of the bindingbetween LXRa and SRC-1 NID by PKA wasdirectly confirmed in pull-down assays. Thelabeled SRC-1-NID was pulled down by GST-LXRa and the signal was reduced by PKAphosphorylation (Figure 6C). These datasuggest that PKA activation modifies LXR andenhances dissociation of co-activator andrecruitment of co-repressor, leading to repressionof LXR transcriptional activity.LXRa /RXR interaction is impaired by PKA.Finally, the effect of PKA on heterodimerizationof LXR/RXR was estimated by co-immunoprecipitation in vivo. Myc-taggedLXRa and RXR were co-expressed in COS7 withor without PKA activation (Figure 7A). RXRwas co-immunoprecipitated with the taggedLXRa. This binding between LXRa and RXRwas nearly eliminated by PKA activation. Incontrast, PKA increased thecoimmunoprecipitation of Flag-tagged CBP andHA-tagged CREB, well characterized targets ofPKA (Figure 7B). These data demonstrated thatthe interaction of LXRa and RXR is impaired byPKA activation, explaining at least partly for PKAinhibition of LXR transactivity.Contribution of RXR to PKA inhibition ofLXR/RXR effect on LXRE in SREBP-1cpromoterIn a previous report, RXR is a substrate of PKA
in vitro and Serine 27 is essential for cAMP-mediated downregulation of RXR transcriptionalactivation in COS cells by assessed in luciferaseassay(35). Therefore, the effect of PKA on RXRactivity to regulate activity of SREBP-1cpromoter was tested. HepG2 cells weretransfected with wild type RXR or mutant RXR(Ser27Ala). As a result (Figure 8), both wildtype and mutant RXRs activate the LXRE-luciferase reporter in response to LG268, asynthetic RXR agonist, with only slight(approximately 25%) decrease in both basal andligand-stimulated response by the RXR mutation.PKA activation dose-dependently inhibited thisRXR-mediated LXRE reporter. The RXRmutation modestly prevented this negative effectof PKA on RXR function especially at low dosedbcAMP(10, 30 mM) but less at high dosedbcAMP(100 mM). These results indicate thatRXR phosphorylation is at least in part involvedin suppression of SREBP-1c promoter activity bycAMP/PKA signals.
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
Discussion
Our current studies demonstrate that PKA directlyphosphorylates LXRa protein and inhibits itssignaling, resulting in suppression of SREBP-1ctranscription, both in vitro and in vivo.Phosphorylation of LXRa, presumably throughits conformational change, causes twoconsequences: suppression of RXR dimerizationleading to decreased binding to LXRE, andsuppression of the ligand activation, followed bydecreased recruitment of co-activator SRC-1 andincreased recruitment of co-repressor NcoR1.Both events lead to repression of LXRtransactivation for SREBP-1c. PKA inhibitionwas more prominent for LBD activation of LXRthan for its binding to LXRE.
Post-translational modification oftranscription factors by phosphorylation providesa rapid cellular response to environmentalchanges. We now show that the ligand-inducedtransactivity of LXR can be regulated in vivo, exvivo, and in vitro by PKA-dependentphosphorylation. The catalytic subunit of PKAhas been identified in the nucleus andphosphorylates numerous transcription factorsmodulating their transcriptional activitiespositively such as HNF-4a and negatively such asL-PK(28). Now, LXR can be added to the list ofPKA-modulating factors. Although PKA-dependent phosphorylation sites (R-X(0-2)-S/T) liewithin all domains of LXR, the 195-196 serinesand 290-291 threonine and serine residues mightbe crucial for ligand-induced conformationalchange since these sites in humanLXRa completely match the most preferableconsensus sequence (R-R/K-X-S/T). It is alsopossible that phosphorylation states of thesecritical sites control other potentialphosphoryaltion sites. LXR/RXR alsotransactivate other genes such as the ABCA1(36)and the ABCG family(37,38). The LXRa-induced activity of the ABCA1 promotercontaining LXRE was also repressed by PKAsimilarly to the SREBP-1c promoter. However,the inhibitory effect of PKA on the ABCA1mRNA level was minimal in our sets ofexperiments in livers and hepatic cells.Conversely, expression of ABCA1 in murinemacrophage cell line RAW 264 cells was reportedto be up-regulated by dbcAMP(39,40), althoughits responsible regulatory motif in the ABCA1
promoter has not been identified. Thisundetermined cAMP-dependent activationmechanism might unmask LXR-mediatedsuppression of ABCA1 promoter by PKA in theliver.
It has been reported that theglucagon/cAMP signal suppresses the expressionof SREBP-1c in rat primary hepatocytes(24).Our current data clarify its molecular mechanism.The hypothesis that PKA suppresses SREBP-1cthrough LXR signal is supported by followingobservations. 1. LXR/RXR is a dominantactivator for SREBP-1c. 2. PKA inhibition ofSREBP-1c promoter activity was more prominentin its activation by the LXR agonist than in thebasal level. 3. Deletion studies with SREBP-1cpromoter luciferase constructs revealed thatLXRE is the responsible for PKA inhibition.Based upon our findings, glucagon/cAMP/PKAsignal could at least partially explain fastingsuppression of SREBP-1c through LXR/RXR andLXRE. Supportively, it was reported that effectof insulin on SREBP-1c expression could bemediated by LXR and LXRE(41). Furtherstudies are needed to fully determine the extent towhich glucagons/cAMP/PKA signal contributesto fasting regulation of hepatic metabolic genesincluding SREBP-1c.
In a recent report, it is observed that LXRaand b are phosphorylated in HEK293 cell byunkown kinase(s)(42). In their data, basal andligand-stimulated LXRa activity to induceABCA1 promoter were not altered by substitutionof phosphorylation redisue to alanine. However,the response of mutant LXRa to potential kinasesremains to be tested. Our study for the first timesuggests the possibility that LXR mediates anutritional signal via phosphorylation by PKA.Further study is needed to more precisely identifyphysiological function(s) in LXRphosphorylation.In summary, transcriptional activity of LXRa
on SREBP-1c promoter was decreased by PKA.Direct phosphorylation of LXRa by PKA resultedin decrease of DNA binding and coactivatorrecruitment of LXR. This first demonstration ofmodification of LXR activity by phosphorylationsuggest that reduction of mRNA level of SREBP-1c in fasting condition might be mechanisticallyat least in part through LXR phophorylation.
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
References
1. Brown, M. S., and Goldstein, J. L. (1997) Cell 89, 331-340
2. Shimano, H., Horton, J. D., Shimomura, I., Hammer, R. E., Brown, M. S., and Goldstein, J.
L. (1997) J Clin Invest 99, 846-854
3. Horton, J. D., Shimomura, I., Brown, M. S., Hammer, R. E., Goldstein, J. L., and Shimano,
H. (1998) J Clin Invest 101, 2331-2339
4. Shimano, H., Horton, J. D., Hammer, R. E., Shimomura, I., Brown, M. S., and Goldstein, J.
L. (1996) J Clin Invest 98, 1575-1584
5. Shimano, H., Yahagi, N., Amemiya-Kudo, M., Hasty, A. H., Osuga, J., Tamura, Y.,
Shionoiri, F., Iizuka, Y., Ohashi, K., Harada, K., Gotoda, T., Ishibashi, S., and Yamada, N.
(1999) J Biol Chem 274, 35832-35839
6. Horton, J. D., Bashmakov, Y., Shimomura, I., and Shimano, H. (1998) Proc Natl Acad Sci U
S A 95, 5987-5992
7. Amemiya-Kudo, M., Shimano, H., Yoshikawa, T., Yahagi, N., Hasty, A. H., Okazaki, H.,
Tamura, Y., Shionoiri, F., Iizuka, Y., Ohashi, K., Osuga, J., Harada, K., Gotoda, T., Sato, R.,
Kimura, S., Ishibashi, S., and Yamada, N. (2000) J Biol Chem 275, 31078-31085
8. Yoshikawa, T., Shimano, H., Amemiya-Kudo, M., Yahagi, N., Hasty, A. H., Matsuzaka, T.,
Okazaki, H., Tamura, Y., Iizuka, Y., Ohashi, K., Osuga, J., Harada, K., Gotoda, T., Kimura,
S., Ishibashi, S., and Yamada, N. (2001)Mol Cell Biol 21, 2991-3000
9. Repa, J. J., Liang, G., Ou, J., Bashmakov, Y., Lobaccaro, J. M., Shimomura, I., Shan, B.,
Brown, M. S., Goldstein, J. L., and Mangelsdorf, D. J. (2000) Genes Dev 14, 2819-2830
10. Shimomura, I., Bashmakov, Y., Ikemoto, S., Horton, J. D., Brown, M. S., and Goldstein, J.
L. (1999) Proc Natl Acad Sci U S A 96, 13656-13661
11. Hasty, A. H., Shimano, H., Yahagi, N., Amemiya-Kudo, M., Perrey, S., Yoshikawa, T.,
Osuga, J., Okazaki, H., Tamura, Y., Iizuka, Y., Shionoiri, F., Ohashi, K., Harada, K.,
Gotoda, T., Nagai, R., Ishibashi, S., and Yamada, N. (2000) J Biol Chem 275, 31069-31077
12. Matsuzaka, T., Shimano, H., Yahagi, N., Amemiya-Kudo, M., Okazaki, H., Tamura, Y.,
Iizuka, Y., Ohashi, K., Tomita, S., Sekiya, M., Hasty, A., Nakagawa, Y., Sone, H., Toyoshima,
H., Ishibashi, S., Osuga, J., and Yamada, N. (2004) Diabetes 53, 560-569
13. Ono, H., Shimano, H., Katagiri, H., Yahagi, N., Sakoda, H., Onishi, Y., Anai, M., Ogihara,
T., Fujishiro, M., Viana, A. Y., Fukushima, Y., Abe, M., Shojima, N., Kikuchi, M., Yamada,
N., Oka, Y., and Asano, T. (2003) Diabetes 52, 2905-2913
14. Janowski, B. A., Grogan, M. J., Jones, S. A., Wisely, G. B., Kliewer, S. A., Corey, E. J., and
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
Mangelsdorf, D. J. (1999) Proc Natl Acad Sci U S A 96, 266-271
15. Peet, D. J., Turley, S. D., Ma, W., Janowski, B. A., Lobaccaro, J. M., Hammer, R. E., and
Mangelsdorf, D. J. (1998) Cell 93, 693-704
16. Alberti, S., Schuster, G., Parini, P., Feltkamp, D., Diczfalusy, U., Rudling, M., Angelin, B.,
Bjorkhem, I., Pettersson, S., and Gustafsson, J. A. (2001) J Clin Invest 107, 565-573
17. Cao, G., Liang, Y., Broderick, C. L., Oldham, B. A., Beyer, T. P., Schmidt, R. J., Zhang, Y.,
Stayrook, K. R., Suen, C., Otto, K. A., Miller, A. R., Dai, J., Foxworthy, P., Gao, H., Ryan, T.
P., Jiang, X. C., Burris, T. P., Eacho, P. I., and Etgen, G. J. (2003) J Biol Chem 278, 1131-
1136
18. Castrillo, A., Joseph, S. B., Vaidya, S. A., Haberland, M., Fogelman, A. M., Cheng, G., and
Tontonoz, P. (2003)Mol Cell 12, 805-816
19. Joseph, S. B., Castrillo, A., Laffitte, B. A., Mangelsdorf, D. J., and Tontonoz, P. (2003) Nat
Med 9, 213-219
20. Joseph, S. B., McKilligin, E., Pei, L., Watson, M. A., Collins, A. R., Laffitte, B. A., Chen, M.,
Noh, G., Goodman, J., Hagger, G. N., Tran, J., Tippin, T. K., Wang, X., Lusis, A. J., Hsueh,
W. A., Law, R. E., Collins, J. L., Willson, T. M., and Tontonoz, P. (2002) Proc Natl Acad Sci
U S A 99, 7604-7609
21. Laffitte, B. A., Chao, L. C., Li, J., Walczak, R., Hummasti, S., Joseph, S. B., Castrillo, A.,
Wilpitz, D. C., Mangelsdorf, D. J., Collins, J. L., Saez, E., and Tontonoz, P. (2003) Proc Natl
Acad Sci U S A 100, 5419-5424
22. Yahagi, N., Shimano, H., Hasty, A. H., Amemiya-Kudo, M., Okazaki, H., Tamura, Y.,
Iizuka, Y., Shionoiri, F., Ohashi, K., Osuga, J., Harada, K., Gotoda, T., Nagai, R., Ishibashi,
S., and Yamada, N. (1999) J Biol Chem 274, 35840-35844
23. Yoshikawa, T., Shimano, H., Yahagi, N., Ide, T., Amemiya-Kudo, M., Matsuzaka, T.,
Nakakuki, M., Tomita, S., Okazaki, H., Tamura, Y., Iizuka, Y., Ohashi, K., Takahashi, A.,
Sone, H., Osuga Ji, J., Gotoda, T., Ishibashi, S., and Yamada, N. (2002) J Biol Chem 277,
1705-1711
24. Foretz, M., Pacot, C., Dugail, I., Lemarchand, P., Guichard, C., Le Liepvre, X., Berthelier-
Lubrano, C., Spiegelman, B., Kim, J. B., Ferre, P., and Foufelle, F. (1999) Mol Cell Biol 19,
3760-3768
25. Chen, D., Pace, P. E., Coombes, R. C., and Ali, S. (1999)Mol Cell Biol 19, 1002-1015
26. Rochette-Egly, C., Oulad-Abdelghani, M., Staub, A., Pfister, V., Scheuer, I., Chambon, P.,
and Gaub, M. P. (1995)Mol Endocrinol 9, 860-871
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
27. Lazennec, G., Canaple, L., Saugy, D., and Wahli, W. (2000)Mol Endocrinol 14, 1962-1975
28. Viollet, B., Kahn, A., and Raymondjean, M. (1997)Mol Cell Biol 17, 4208-4219
29. Yamamoto, T., Shimano, H., Nakagawa, Y., Ide, T., Yahagi, N., Matsuzaka, T., Nakakuki,
M., Takahashi, A., Suzuki, H., Sone, H., Toyoshima, H., Sato, R., and Yamada, N. (2004) J
Biol Chem 279, 12027-12035
30. Sone, H., Shimano, H., Shu, M., Nakakuki, M., Takahashi, A., Sakai, M., Sakamoto, Y.,
Yokoo, T., Matsuzaka, K., Okazaki, H., Nakagawa, Y., Iida, K. T., Suzuki, H., Toyoshima,
H., Horiuchi, S., and Yamada, N. (2004) Biochem Biophys Res Commun 316, 790-794
31. Clegg, C. H., Correll, L. A., Cadd, G. G., and McKnight, G. S. (1987) J Biol Chem 262,
13111-13119
32. Ide, T., Shimano, H., Yahagi, N., Matsuzaka, T., Nakakuki, M., Yamamoto, T., Nakagawa,
Y., Takahashi, A., Suzuki, H., Sone, H., Toyoshima, H., Fukamizu, A., and Yamada, N.
(2004) Nat Cell Biol 6, 351-357
33. Hu, X., Li, S., Wu, J., Xia, C., and Lala, D. S. (2003)Mol Endocrinol 17, 1019-1026
34. Glass, C. K., Rose, D. W., and Rosenfeld, M. G. (1997) Curr Opin Cell Biol 9, 222-232
35. Harish, S., Ashok, M. S., Khanam, T., and Rangarajan, P. N. (2000) Biochem Biophys Res
Commun 279, 853-857
36. Costet, P., Luo, Y., Wang, N., and Tall, A. R. (2000) J Biol Chem 275, 28240-28245
37. Kennedy, M. A., Venkateswaran, A., Tarr, P. T., Xenarios, I., Kudoh, J., Shimizu, N., and
Edwards, P. A. (2001) J Biol Chem 276, 39438-39447
38. Yu, L., Gupta, S., Xu, F., Liverman, A. D., Moschetta, A., Mangelsdorf, D. J., Repa, J. J.,
Hobbs, H. H., and Cohen, J. C. (2004) J Biol Chem
39. Abe-Dohmae, S., Suzuki, S., Wada, Y., Aburatani, H., Vance, D. E., and Yokoyama, S.
(2000) Biochemistry 39, 11092-11099
40. Takahashi, Y., Miyata, M., Zheng, P., Imazato, T., Horwitz, A., and Smith, J. D. (2000)
Biochim Biophys Acta 1492, 385-394
41. Chen, G., Liang, G., Ou, J., Goldstein, J. L., and Brown, M. S. (2004) Proc Natl Acad Sci U
S A 101, 11245-11250
42. Chen, M., Bradley, M. N., Beaven, S. W., and Tontonoz, P. (2006) FEBS Lett 580, 4835-4841
43. Wagner, B. L., Valledor, A. F., Shao, G., Daige, C. L., Bischoff, E. D., Petrowski, M., Jepsen,
K., Baek, S. H., Heyman, R. A., Rosenfeld, M. G., Schulman, I. G., and Glass, C. K. (2003)
Mol Cell Biol 23, 5780-5789
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
Acknowledgments
We are grateful to Alyssa H. Hasty for critical reading of this manuscript, Drs. Tomoaki Morioka and
Hidenori Koyama for a great help in generation of expression plasmid of mutated (dominant negative)
form of the regulatory type 1 subunit (RI) of cyclic AMP -dependent protein kinase. Correspondence
and requests for materials should be address to Hitoshi Shimano. This work was supported by grants-
in-aid from the Ministry of Science, Education, Culture, and Technology of Japan.
Figure Legends
Figure 1. Effects of PKA activation on expression of mouse hepatic SREBP-1.
C57BL6/J mice fasted for 24 hr (Fasted) followed by refeeding with a high sucrose high fat diet for 12 hr
(Refed) were intravenously treated with vehicle or a combination of PKA activators, dbcAMP (35mg/kg)
and theophylline (30mg/kg) (PKA). Two hr post- injection, mice were sacrificed and total liver RNA
and nuclear extracts prepared. Total RNA (10 mg) and nuclear extracts (40 mg protein) were subjected to
northern and immunoblotting, respectively.
Figure 2. Effects of PKA on mRNA level of SREBP-1 induced by the synthetic LXR ligand, T0901317.
Rat primary hepatocytes were incubated in medium containing vehicle or T0901317 (1 mM) in the
presence or absence of dbcAMP (30 mM) for 24 hr. Cells were harvested for extraction of total RNA,
followed by northern blotting as described in Materials and Methods Section.
Figure 3. Effects of PKA on SREBP-1c promoter activity induced by LXRa
A. HepG2 cells were transfected with SREBP-1c promoter Luc vector (300 ng/well), CMV renilla
luciferase vector as an internal reference (2.5 ng/well), expression vectors for LXRa (100 ng/well) and
PKA catalytic subunit (100 ng/well). After transfection, the cells were further incubated with the
medium containing vehicle or T0901317 (1 mM) for 20 hrs. B. HepG2 cells were transfected with
LXRE enhancer Luc vector (200 ng/well), CMV renilla luciferase vector (2.5 ng/well), and expression
vectors for LXRa (100 ng/well) and PKA regulatory subunit dominant negative form (DN PKA) (300
ng/well). After transfection, cells were incubated in medium containing vehicle or T0901317 (1 mM)
and forskolin (0, 0.1, 0.3, 1, 3 mM) for 20 hr. Assays were performed as described in the Materials and
Methods Section. C. HepG2 cells were transfected with expression vectors for Gal4-DBD-LXRa ligand
binding domain (150 ng/well), PKA Gal4 RE LUC vector (150 ng/well), PKA (3, 10, 30, 100, 200
ng/well) and CMV renilla luciferase vector (2.5 ng/well). After each transfection, cells were incubated
in medium containing vehicle or T0901317 (1 mM)) for 20 hr and were subjected luciferase and renilla
reporter assays as in described in the Materials and Methods Section. D. 293 cells were transfected with
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
hABCA1 promoter Luc vector (300 ng/well), expression vector for LXRa (100 ng/well), PKA (0, 3, 10,
30, 100 ng/well) and CMV renilla luciferase vector for internal reference (2.5 ng/well). After each
transfection, cells were incubated in medium containing vehicle or T0901317 (1 mM)) for 20 hr.
Figure 4. Direct phosphorylation of recombinant LXRa protein at the serine or threonine residues in the
ligand binding domain is crucial for inhibition of LXR signaling by dbcAMP.
A. In vitro kinase assay of full length LXRa. Histone H1 or recombinant histidine-tagged LXRa were
incubated with PKA and 32P-ATP and resolved by SDS-PAGE (8% gel). Phosphorylated protein was
visualized by autoradiography. Phosphorylation of Histone H1 and PKA itself was shown as positive
control of PKA target protein. B. In vitro kinase assay of several fragment of LXRa. 1-163 a.a., 162-
326 a.a., 325-445 a.a. corresponding amino acids sequences of LXRa protein, respectively were
incubated with PKA. Samples were resolved by SDS-PAGE (12 % gel) and electrophoretically
transferred to PVDF-membranes. Phosphorylated proteins were visualized by autoradiography. LXRa
proteins were estimated by immunoblotting with anti Histidine antibody. C. Substitution of serine or
threonine amino acid residues for alanine in LXRa abolished the PKA dependent repression of a
luciferase reporter drived by LXRE. Expression vectors for wild type LXRa (wt LXRa) or an
LXRa mutant with S195A and S196A (195A196A) or with T290 and S290A (290A 291A) (100 ng/well)
and CMV renilla luciferase vector (2.5 ng/well) were cotransfected with LXRE enhancer reporter plasmid
(100 ng/well) into 293 cells. After transfection, dbcAMP was added into the medium, followed by
incubation for 20 hr. Luciferase activity was measured. Relative activity compared to each control
was plotted. D. PKA activation increases phosphorylation of LXRa in vivo. COS7 cells transfected
with HA-tagged LXRa were metabolically labeled with [32P]orthophosphate and incubated for 30 min
with PKA activators (forskolin 10 mM, dibutylyl cAMP 1 mM, IBMX 1 mM) before harvesting. Protein
samples were immunprecipitated with anti-HA antibody. Labeled proteins resolved by SDS-PAGE were
autoradiographed and immunoblotted. 35S-Met is methionine labeled LXRa as used positive control.
Figure 5. Phosphorylation of LXRa by PKA decreases binding of LXRa/RXR to LXRE.
A. Decreased binding of LXRa/RXR to LXRE by PKA treatment with LXRa in electrophoretic
mobility shift assays. Increasing amounts of recombinant LXRa (left panel) were incubated with BSA
(used as nonspecific protein control) or PKA catalytic subunit and then incubated with radio-labeled
LXRE probe and in vitro synthesized RXR for EMSA as described in the Materials and Methods Section.
In right panel, recombinant LXRa were preincubated with BSA or PKA at 30 ºC or 0 ºC (on ice) (right
panel) and then incubated with radio-labeled LXRE probe and in vitro synthesized RXR for EMSA as
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
described in the Materials and Methods Section. B. In vivo PKA activation decreased the DNA binding
activity of LXR/RXR complex in EMSA (left panel). The various doses of PKA activators (vehicle,
dbcAMP 35 mg/kg + theophylline 30 mg/kg, dbcAMP 105 mg/kg + theophylline 30 mg/kg) were
administrated to refed C57BL6/J mice. Nuclear protein extracts prepared from livers were incubated
with the 32P-LXRE probe. The signal from LXR/RXR was confirmed by supershift assays after addition
of LXR and RXR antibodies to the incubations. Nuclear protein (30 mg/lane) were resolved by SDS-
PAGE (10 % gel) and subjected to immunoblotting with anti phospho-CREB, anti CREB and anti
LXRa antibodies (right panel). C. PKA activation decreases DNA binding of LXR/RXR complex to
LXRE on SREBP-1c promoter in vivo. H2.35 cells were treated with PKA activators (forskolin 10 mM,
dibutylyl cAMP 100 mM) or vehicle for 60 min. Since LXR antibody suitable for imuunoprecipitation is
not currently available(43), ChIP analysis was performed with anti-RXR antibody or control IgG.
Primers specific to the LXRE-region of mouse SREBP-1c promoter were used for PCR analysis. As a
positive control (3% INPUT), the assay was performed with the same primers on the genomic DNA on
which the ChIP assays were performed.
Figure 6. PKAmodulates recruitment of coactivator and corepressor for LXRa.
A. Inhibition of recruitment of SRC-1 to LXRa by PKA in mammalian two hybrid system. 293 cells
were transfected with Gal4 RE Luc vector, CMV renilla luciferase vector (2.5 ng/well) and expression
vectors for VP-16-LXRa (1-445 a.a), Gal4-DBD-SRC-1: DNA binding domain (DBD) of Gal4 fused to
nuclear receptor interaction domain (NID) of SRC-1, and PKA. Four 4hrs after transfection, the cells
were further incubated with medium containing vehicle or T0901317 (1 mM) for 20 hrs. B. Activation
of recruitment of NcoR-1 to LXRa by PKA. 293 cells were transfected with Gal4 RE Luc vector, SV40
renilla luciferase vector (25 ng/well) and expression vectors for VP-16 LXRa (1-445 a.a), Gal4-DBD-
NcoR1, and PKA. 4 hrs after transfection, the cells were further incubated with medium containing
vehicle or T0901317 (1 mM) for 20 hrs. C. PKA treatment inhibited the interaction of LXRa with SRC-
1 in vitro. Recombinant GST-LXRa fusion protein was treated with PKA or BSA (used as nonspecific
control protein) at 30 ºC and then incubated with 35S-radio-labeled SRC-1-NID synthesized in vitro and
glutathione sepharose beads. Pull-downed proteins were resolved by SDS-PAGE (12 % gel) as
described in Materials and Methods.
Figure 7. PKA prevents interaction between LXRa and RXR in vivo.
A. PKA inhibition of co-immunoprecipitation of LXRa and RXR. B. PKA activation of co-
immunoprecipitation of CREB and CBP. Cos7 cells were transfected with expression vector for myc-
tagged LXRa and RXR (A), or for Flag-tagged CBP and HA-tagged CREB (B). After incubation for 48
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
hr, cells were treated with medium containing PKA activators (100 mM dbcAMP, 10 mM forskolin, 100
mM IBMX) for 30 min and harvested. Immunoprecipitation and immunoblotting with indicated
antibodies were performed as described in Materials and Methods. Proteins were extracted by lysis buffer
and resolved by SDS-PAGE (8 % gel for LXR and RXR, 4 % gel for CBP and 10 % gel for CREB).
There is different exposure time between Lysate and IP samples for detection of Flag-tagged CBP because
intensity of protein amount of Flag-tagged CBP in lysate was too strong.
Figure 8. PKAmodifies RXR activity induced by RXR ligand on LXRE in SREBP-1c promoter.
293T cells were transfected with LXRE enhancer Luc vector (300 ng /well), CMV renilla luciferase
vector (2.5 ng /well), and expression vector for wild type RXRa or mutant RXR (400 ng /well). After
transfection, cells were incubated for 20 h in medium containing vehicle or T0901317 (1 mM), LG268 (30
nM), dbcAMP (0, 3, 10, 30, 100 mM) and subjected to luciferase and renilla reporter assays.
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

SREBP-1LXRα
RXR
36B4
SREBP-1PEPCK
Fasted Refed PKAA
B
FastedRefedPKA
Figure 1
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

SREBP-1
LXRα
T0901317(1 μM)PKA activator
- +++--
PEPCK
36B4
RXR
Figure 2
ABCA1
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 3
0 5 10 15 20 25 30
LXRα+TPKA
LXRE LUC
LXRE LUC
LUC
SREBP-1c promoter LUC
-2.6Kbp
-550bp
-90bp
RLU
LXRα
DN PKA
+ + + + ++T(1 μM) + + + +
A
B
+ ++
+
0100200300400
500600700800900
Gal4-DBD
Gal4 RE LUC
LXRα-LBD
T
RL
U
T(1 μM)PKA - PKA
+-
- + + + + +
D
0
2
4
6
8
10
12
LXRαPKA(ng/well)
+ + + + +10 10030
T(1 μM) + + + ++++
010 100300
RL
U
+ + +++
+
+ + + +
+ + + +Forskolin(μM) 0.1 0.3 1 3 0.1 0.3 1 3
RL
U
C
+ ++
+
+ ++
+
0
2
4
6
8
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from
PKAHistone H1
rLXRα
+ ++
+++
+
PKA
Histone H1
rLXRα
A/B C DE AF2A/B C DE AF21 97 161 438 445
A/B C1631
DE162 326
DE AF2325 445
autoradiography WB
PKA + +
0
0.5
1
wt195A196A290A291A
50 1000dbcAMP(μM)
Relativeactivity
A
B
D
C
Figure 4
- -
----
-- --
HA LXRαPKA activator
+ +-- - +
35S-Met
ab HAautoradiography
IP: HA
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

rLXRαIVT RXR +
++ + + + + +
+PKAtreatment
rLXRαIVT RXR
preincubationtemperature
++ + + + ++ + + +
PKA + +
P-CREB
CREB
LXRα
ab LXRαab RXRPKA
activators
PKAactivators
Figure 5
A
B
C
---
- --- -
nosample + ++- + ++- + ++-
+ ++-
30 3000
lgG RXR INPUT
T0901317(10μM)PKA activators
+++
+++
+++
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

0
20
40
60
80
VP16-LXRα
RL
U
SRC-1-NID
T(1 μM)PKA - PKA-
- + ++- + + + +
+ +--
RL
U
T(1 μM)PKA - PKA
NcoR1-NIDGal4-DBD
LXRα VP16-AD
Gal4 RE LUC
T
+
0
2
4
6
8
10
+-
- + + +
A B
C
Figure 6
Gal4-DBD
LXRα VP16-AD
Gal4 RE LUC
T
35S SRC-1 NIDGST(μg)
GST-LXRα(μg)
po
siti
ve
con
t
2 4PKA - - + +
2 44
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

ab RXR
ab myc
Lysate IP (ab myc)
myc LXRRXR + ++ + +- +
+ ++ + +- +
PKA activators - - + + +--ab myc for IP + +--
WB
transfection
ab Flag
ab HA
Lysate IP (ab HA)
HA CREBFlag CBP + +-
+ +-PKA activators - - +
WB
transfection + +-+ +-
- - +
Figure 7
A
B
*
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

0
1
2
3
4
RLU
dbcAMP(μM)LG268(30nM)
- ---
----+ +- ++ + + ++ + + +
-3 10010 30 3 10010 30-
wild type RXRmutant RXR
+- ++ + +-- -+ - -- --+- ++ + +-- - +- -- --
T0901317(1μM) +- -- -- --- -- -- --
Figure 8
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Toyoshima and Nobuhiro YamadaMatsuzaka, Akimitsu Takahashi, Naoya Yahagi, Hirohito Sone, Hiroaki Suzuki, Hideo Takashi Yamamoto, Hitoshi Shimano, Noriyuki Inoue, Yoshimi Nakagawa, Takashi
liverProtein Kinase A suppresses SREBP-1c expression via phosphorylation of LXR in the
published online February 12, 2007J. Biol. Chem.
10.1074/jbc.M611911200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from