prophylactic fibrinolysis through selective dissolution of nascent clots by tpa-carrying...
TRANSCRIPT

A RT I C L E S
Because of their long circulation time, RBCs have been proposed foruse as carriers to improve the delivery and to prolong the life span andbioavailability of drugs in the bloodstream1–5. For example, intracellu-lar loading of drugs and antigens in RBCs enhances the delivery ofthese molecules to macrophages and antigen-presenting cells in ani-mals6,7. Furthermore, loading steroids into RBCs enhances their anti-inflammatory effects after reinjection into human patients8. However,unless the compounds diffuse through the RBC membrane, the encap-sulated therapeutic enzymes will have no effect. In theory, this limita-tion could be overcome by coupling the enzymes to the RBC surface.This study tests this approach in animal models and in the context ofprophylaxis of vascular thrombosis.
Thromboprophylaxis helps reduce morbidity and mortality inmany diseases, but it shows a limited benefit/risk ratio in stroke andpostsurgical settings where there is a high risk of both thrombosis andbleeding9–11. Mechanical filters trap large emboli but do not preventcerebral microembolism and arterial reocclusion, and postoperativebleeding limits the safety and utility of systemic anticoagulants12–14.Available plasminogen activators such as tPA are unsuitable for pro-phylaxis because they have a short life span, dissolve both preexistingand nascent clots, and cause side effects in tissues (e.g., in the brain)15.The ideal thromboprophylactic delivery system would precludeextravasation and spare preexisting hemostatic clots, while selectivelydissolving potentially lethal nascent clots.
In this study we tested the hypothesis that coupling tPA to carrierRBCs might fulfill the above requirements. Preexisting clots are lesspermeable than nascent ones, even to soluble plasminogen activa-tors16,17. In theory, reinjection of a stable complex of RBCs and tPA
(after surgical hemostasis is suitably advanced) will deny the enzymeaccess to the relatively impermeable preformed hemostatic clots andextravascular tissues, thereby reducing the risk of bleeding. The com-plex would circulate for prolonged periods and be incorporated insidenascent intravascular clots, initiating their lysis from within (the‘Trojan horse’ fibrinolysis of nascent clots by the complex, Fig. 1a). Theresults of the in vivo studies reported here show that RBC carriage oftPA alters the fibrinolytic profile of tPA as hypothesized, thus provid-ing a strategy for prophylactic fibrinolysis.
RESULTSDesign and in vitro characterization of the RBC-tPA complexWe coupled tPA to RBCs using streptavidin (SA)-biotin as a cross-linker, capitalizing on its effectiveness, modular plasticity, feasibilityand clinical safety3,18–20. We adapted this technique to couplebiotinylated proteins to biotinylated RBCs (b-RBCs) and showedthat b-RBC–SA–biotinylated protein complexes are stable, do notactivate complement or phagocytes and circulate for prolonged peri-ods in vivo4,21.
The amount of biotinylated tPA (b-tPA) that can be coupled to ratb-RBCs ranges from 105 molecules per RBC to fewer than 200 mole-cules of nonbiotinylated tPA (Fig. 1b). Similar results were obtainedwith mouse and human RBCs (data not shown). Massive biotinylationof RBCs could lead to inactivation of the complement-controllingproteins (decay accelerating factor, DAF, and CD59) expressed on theRBC surface, as a result of aggregation by polyvalent streptavidin-biotin interactions, leading to hemolysis of b-RBCs–SA in autologousserum22,23. However, RBCs biotinylated at biotin ester concentrations
1Institute for Environmental Medicine, University of Pennsylvania, 3620 Hamilton Walk, Philadelphia, Pennsylvania 19104, USA. 2Division of Pediatric Hematology,Children’s Hospital of Philadelphia, 1 Civic Center, Philadelphia, Pennsylvania 19104, USA. 3Department of Medicine, University of Pennsylvania, 421 Curie Blvd.,Philadelphia, Pennsylvania 19104, USA. 4Department of Pathology and Laboratory Medicine, University of Pennsylvania, 422 Curie Blvd., Philadelphia, PA 19104,USA. 5Department of Pharmacology, University of Pennsylvania, 3620 Hamilton Walk, Philadelphia, Pennsylvania 19104, USA. Correspondence should be addressedto V.R.M. ([email protected]).
Prophylactic fibrinolysis through selective dissolution ofnascent clots by tPA-carrying erythrocytesJuan-Carlos Murciano1, Sandra Medinilla1, Donald Eslin2, Elena Atochina3, Douglas B Cines4
& Vladimir R Muzykantov1,5
A fibrinolytic agent consisting of a tissue-type plasminogen activator (tPA) coupled to the surface of red blood cells (RBCs) can dissolve nascent clots from within the clot, in a Trojan horse–like strategy, while having minimal effects on preexistinghemostatic clots or extravascular tissue. After intravenous injection, the fibrinolytic activity of RBC-tPA persisted in thebloodstream at least tenfold longer than did that of free tPA. In a model of venous thrombosis induced by intravenously injectedfibrin microemboli aggregating in pulmonary vasculature, soluble tPA lysed pulmonary clots lodged before but not after tPAinjection, whereas the converse was true for RBC-tPA. Free tPA failed to lyse occlusive carotid thrombosis whether injectedbefore or after vascular trauma, whereas RBC-tPA circulating before, but not injected after, thrombus formation restored bloodflow. This RBC-based drug delivery strategy alters the fibrinolytic profile of tPA, permitting prophylactic fibrinolysis.
NATURE BIOTECHNOLOGY VOLUME 21 NUMBER 8 AUGUST 2003 891
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy

A RT I C L E S
below 100 µM (bn-RBC, where n ≤ 100) do not activate complementand are stable in fresh homologous serum (b70-RBC-SA, Fig. 1b).
To test the activity of b10-RBC–SA–b-tPA complexes possessing6.5–7.5 × 104 tPA molecules per RBC (RBC-tPA), we used fibrin clotsformed by adding thrombin to 125I-labeled fibrinogen24. Clots thatformed in the presence of RBC-tPA were dissolved, releasing 125I intothe supernatant. RBC-tPA (but not control RBCs) dissolved clots pre-pared from pure fibrin more rapidly than those prepared fromplatelet-poor plasma or platelet-rich plasma (Fig. 1c), most likelybecause of the presence of plasminogen activator inhibitors in theplasma and because of mechanical retraction in the case of theplatelet-rich plasma. Similar results were seen with clots formed fromrat, mouse or human plasma using tPA coupled to homologous RBCs(data not shown). During a 24-h incubation in fresh serum at 37 °C,hemolysis of RBC-tPA by complement was comparable to that ofcontrol RBCs and only a minor fraction of tPA detached from theRBCs (data not shown).
Free tPA dissolved ∼ 50% of preexisting versus ∼ 90% of nascentclots (a 1.9-fold difference), whereas RBC-tPA dissolved fewerthan 5% of preexisting versus ∼ 80% of nascent clots (a 17-fold dif-ference) (Fig. 1d). Thus, RBC-tPA showed ten times greater selec-tivity towards nascent clots than free tPA, most likely as a result ofmore stringent limitations on RBC-tPA diffusion into the fibrinmeshwork.
Circulation of RBC-tPA in ratsWe injected dual-labeled 51Cr-RBC–125I-tPA complexes intravenouslyinto anesthetized rats. Coupling of tPA to RBCs marginally reducedthe biocompatibility of reinjected autologous RBCs: 80–85% of the
injected dose of control RBCs or RBC-tPAremained in the circulation 2 h after injection(Fig. 2a). Roughly 90% of 51Cr and 125I circu-lating in the blood was found in the cell pel-lets (data not shown), indicating that tPAremains associated with, and does not causehemolysis of, carrier RBCs in vivo.
The amount of RBC-bound tPA thatremained in the bloodstream after bolusinjection of RBC-tPA was markedly greaterthan that remaining after bolus injection offree tPA. A comparison of the respectiveareas under the curve (Fig. 2a) shows thatthe level of the circulating RBC-tPA was atleast an order of magnitude greater than thatof soluble tPA. The experiment was termi-nated at 2 h in compliance with the approvednonsurvival surgery protocol of the institu-tional animal care committee. However, thestable circulation of RBC-tPA throughoutthe duration of the study implies that RBCcarriage prolongs the life span of tPA wellbeyond this time frame.
We then tested the fibrinolytic activity ofcirculating RBC-tPA (Fig. 2b). Rats wereinjected with saline (control), 0.3 mg/kg freetPA or 0.06 mg/kg tPA in the form of RBC-tPA complexes. Blood was collected at varioustimes after injection, mixed with 125I-labeledfibrinogen, incubated for 20 min at 20 °C toform a clot and incubated further at 37 °C topermit ‘endogenous’ fibrinolysis. There was
only minimal enhancement of lysis when clots were formed fromblood obtained 10 min after injection of free tPA, and no enhancementwas observed in blood taken 1 h later. In sharp contrast, clots formedfrom blood obtained 10 min, 1 h or 3 h after injection of fivefold lesstPA in the form of RBC-tPA showed a higher degree of lysis.
Activity of RBC-tPA in a model of pulmonary embolismBased on the above results showing that RBC carriage markedly pro-longs the active life span of tPA, we tested the fibrinolytic efficacy ofRBC-tPA in a model of venous thrombosis in mice. We first confirmedthat RBC coupling prolongs the circulation of tPA in mice: free 125I-tPAdisappeared from the bloodstream within 20 min, whereas 50–60% ofinjected RBC–125I-tPA remained in the bloodstream 60 min after injec-tion (data not shown). We next confirmed in mice the observation firstmade in rats that the duration of tPA-mediated fibrinolytic activity inthe blood after bolus injection was substantially enhanced by RBC cou-pling (Fig. 3a). Fibrinolysis of clots formed from blood taken 2 minafter free tPA injection was facilitated, but this enhancement disap-peared within 20 min after injection. In contrast, clots formed fromblood taken 60 min after injection of RBC-tPA dissolved almost com-pletely. Therefore, as is the case in rats, active RBC-tPA, but not free tPA,circulates for a prolonged time after injection into mice.
To examine fibrinolysis of intravascular clots in vivo, we used amodel of pulmonary embolism induced by injecting radiolabeled fib-rin microemboli (125I-ME) 3–5 µm in diameter. Shortly after intra-venous injection, 125I-ME form aggregates invested with blood cells,which lodge in the lungs of mice and rats24,25. Measuring radioactivityin the lungs 1 h after 125I-ME injection provides a quantitative meas-urement of intravascular fibrinolysis of pulmonary emboli24.
892 VOLUME 21 NUMBER 8 AUGUST 2003 NATURE BIOTECHNOLOGY
b
d
a
c
Figure 1 The concept, design and in vitro characterization of RBC-tPA complex. (a) Free plasminogenactivators (PA, denoted by scissors) undergo elimination minutes after injection; residual circulating PA diffuses into preexisting hemostatic clots and into the tissues, resulting in a high risk of bleedingand tissue damage. RBC-coupled PA circulates for a prolonged time, has little access to preexistingclots or extravascular tissues, and is incorporated into nascent clots causing dissolution from within(‘Trojan horse’). (b) Coupling of biotinylated tPA to biotinylated RBCs via streptavidin. Inset shows thatstreptavidin conjugation does not cause hemolysis of modestly biotinylated carrier RBC by autologousserum. (c) Dissolution of clots formed from pure fibrin (PFC), platelet-poor plasma (PPP) and platelet-rich plasma (PRP) in vitro. RBC-tPA, but not control RBCs (lysis by RBCs is shown by dashed line),dissolve radiolabeled clots at 37 °C. (d) Comparison of in vitro fibrinolysis of preexisting (open bars)versus nascent (closed bars) fibrin clots by free tPAs versus RBC-tPA. All data in these figures arepresented as mean ± s.e.m., n = 4 per point; *, P < 0.05 (Student’s t-test, pair-wise comparisonbetween PFC and PPP or PRP groups).
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy

A RT I C L E S
Injection of free tPA 10 min after the 125I-ME administration dis-solved the clots almost completely. In contrast, less than 20% fibrinol-ysis was seen when tPA was injected even 5 min before the emboli,consistent with its rapid clearance from the bloodstream (Fig. 3b). Incontrast, RBC-tPA injected after the 125I-ME produced a modest (lessthan 40%) fibrinolysis of pulmonary emboli, whereas RBC-tPAinjected 20 min before the 125I-ME caused 80% fibrinolysis. Therefore,the ratio of fibrinolysis of nascent versus preexisting clots was 0.2 forfree tPA and 2.2 for RBC-tPA.
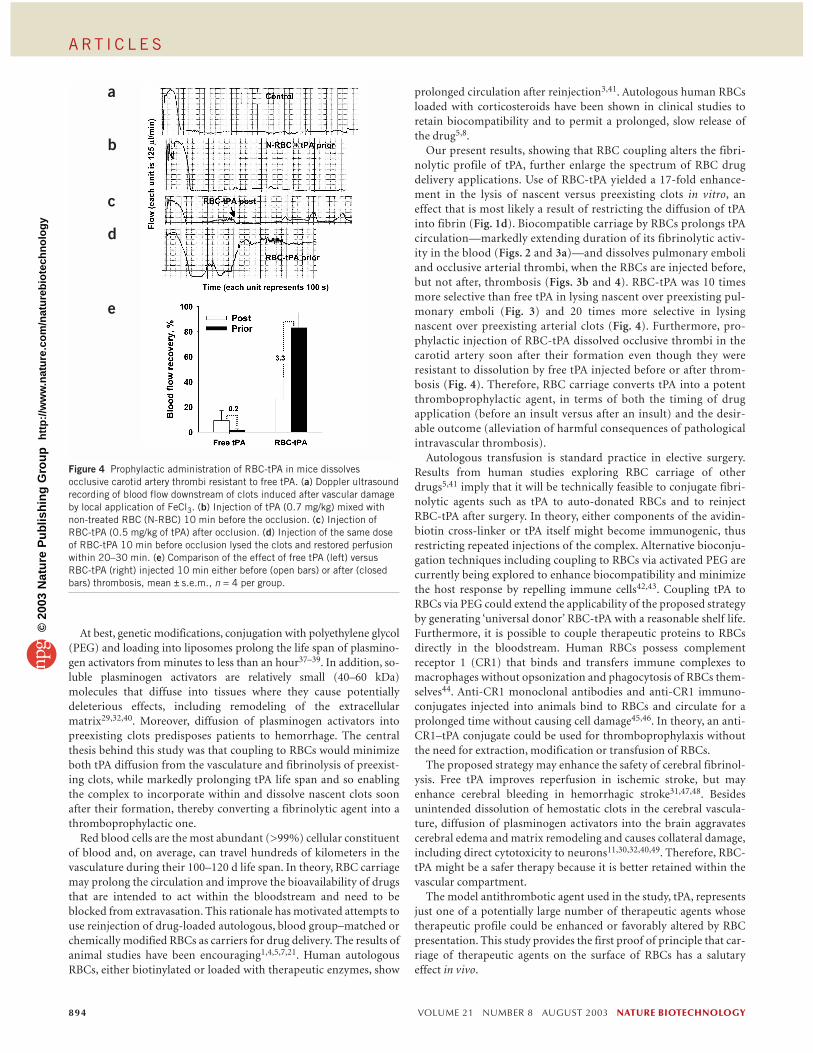
Activity of RBC-tPA in a model of arterial thrombosisLarge, life-threatening occlusive clots are less permeable and less sus-ceptible to fibrinolysis than microemboli because they have a lowersurface/mass ratio24. Therefore, we tested whether RBC-tPA thrombo-prophylaxis could effectively dissolve occlusive thrombi in a large ves-sel using the FeCl3 model of carotid arterial injury in mice.
Thrombus formation and fibrinolysis were monitored by Dopplerultrasound. Typical Doppler tracings from individual experimentsare shown in Figure 4a–d; data collected from ten experiments areshown in Figure 4e. Shortly after the application of the FeCl3 solutionto the tunica adventitia, blood flow in the carotid artery was practi-cally undetectable (Fig. 4a), and the vessel remained occluded for theentire experiment (60 min). Injection of free tPA before thrombusformation (Fig. 4b), or injection of free tPA (data not shown) or RBC-tPA 10 min after thrombus formation (Fig. 4c), did not cause clot lysisor facilitate reperfusion. However, RBC-tPA injected 10 min before
thrombosis, though not affecting the rate of clot formation, began todissolve the clots within 10–20 min and to restore the blood flow. Themean percentage of clot dissolution in all experiments was ∼ 80% ofbaseline by 20–30 min. Therefore, the ratio of fibrinolysis of nascentversus preexisting clots in this model was 0.2 for free tPA and 3.3 forRBC-tPA, respectively.
DISCUSSIONThis study shows the potential of RBC-coupled tPA for thrombopro-phylaxis in animal models of venous and arterial thrombosis.Plasminogen activators are used for therapeutic dissolution of preex-isting clots in settings where the danger from ischemic injury or thelonger-term sequelae from venous vascular damage or outflowobstruction are imminent15,26,27. Their application is restricted bytheir clearance within minutes28, side effects from extravascular activ-ities including tissue remodeling and neuronal toxicity29,30 and risk ofbleeding caused by dissolution of hemostatic clots31,32.
To overcome these limitations, diverse modifications of plasmino-gen activators have been proposed, including conjugating them toantibodies that recognize clot components and designing, through theuse of single-point mutations and deletion of nonprotease domains inplasminogen activators, genetic mutants with prolonged intravascularlife spans and enhanced resistance to plasma inhibitors11,26,33–36. Someof these modifications improve therapeutic fibrinolysis. However,available plasminogen activators lack the necessary properties for usein thromboprophylaxis.
NATURE BIOTECHNOLOGY VOLUME 21 NUMBER 8 AUGUST 2003 893
a
b
ID in
blo
od
, %
a
b
Figure 2 Prolonged circulation of active RBC-tPA complex in rats. (a) Tracing of radiolabeled control 51Cr-RBC, free 125I-tPA, and dual-labeled51Cr-RBC-125I-tPA complex after intravenous injection. (b) Fibrinolyticactivity recovered in blood samples obtained at the indicated times afterinjection of saline, 0.3 mg/kg of free tPA or 0.06 mg/kg of tPA coupled toRBCs (RBC-tPA). The data are shown as mean ± s.e.m., n = 4 per group; *, P < 0.05, **, P < 0.01 (a nonparametric Mann-Whitney assay betweensaline and drug-treated groups).
Figure 3 Active RBC-tPA circulates in mice for a prolonged time and dissolvespulmonary emboli injected after RBC-tPA. (a) Fibrinolytic activity recovered in blood samples obtained at the indicated times after injection of 0.5 mg/kg free tPA or tPA coupled to RBCs (RBC-tPA) into mice. (b) Dissolution of 125I-labeled fibrin microemboli lodged in the pulmonary vasculature in miceby bolus injection of 0.5 mg/kg of either free tPA or RBC-tPA. Fibrinolyticswere injected 20 min before (open bars) or 10 min after (closed bars) 125I-ME.The data are shown as mean ± s.e.m., n = 4–5 per group.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy
A RT I C L E S
At best, genetic modifications, conjugation with polyethylene glycol(PEG) and loading into liposomes prolong the life span of plasmino-gen activators from minutes to less than an hour37–39. In addition, so-luble plasminogen activators are relatively small (40–60 kDa)molecules that diffuse into tissues where they cause potentially deleterious effects, including remodeling of the extracellularmatrix29,32,40. Moreover, diffusion of plasminogen activators intopreexisting clots predisposes patients to hemorrhage. The centralthesis behind this study was that coupling to RBCs would minimizeboth tPA diffusion from the vasculature and fibrinolysis of preexist-ing clots, while markedly prolonging tPA life span and so enablingthe complex to incorporate within and dissolve nascent clots soonafter their formation, thereby converting a fibrinolytic agent into athromboprophylactic one.
Red blood cells are the most abundant (>99%) cellular constituentof blood and, on average, can travel hundreds of kilometers in thevasculature during their 100–120 d life span. In theory, RBC carriagemay prolong the circulation and improve the bioavailability of drugsthat are intended to act within the bloodstream and need to beblocked from extravasation. This rationale has motivated attempts touse reinjection of drug-loaded autologous, blood group–matched orchemically modified RBCs as carriers for drug delivery. The results ofanimal studies have been encouraging1,4,5,7,21. Human autologousRBCs, either biotinylated or loaded with therapeutic enzymes, show
prolonged circulation after reinjection3,41. Autologous human RBCsloaded with corticosteroids have been shown in clinical studies toretain biocompatibility and to permit a prolonged, slow release ofthe drug5,8.
Our present results, showing that RBC coupling alters the fibri-nolytic profile of tPA, further enlarge the spectrum of RBC drugdelivery applications. Use of RBC-tPA yielded a 17-fold enhance-ment in the lysis of nascent versus preexisting clots in vitro, aneffect that is most likely a result of restricting the diffusion of tPAinto fibrin (Fig. 1d). Biocompatible carriage by RBCs prolongs tPAcirculation—markedly extending duration of its fibrinolytic activ-ity in the blood (Figs. 2 and 3a)—and dissolves pulmonary emboliand occlusive arterial thrombi, when the RBCs are injected before,but not after, thrombosis (Figs. 3b and 4). RBC-tPA was 10 timesmore selective than free tPA in lysing nascent over preexisting pul-monary emboli (Fig. 3) and 20 times more selective in lysing nascent over preexisting arterial clots (Fig. 4). Furthermore, pro-phylactic injection of RBC-tPA dissolved occlusive thrombi in thecarotid artery soon after their formation even though they wereresistant to dissolution by free tPA injected before or after throm-bosis (Fig. 4). Therefore, RBC carriage converts tPA into a potentthromboprophylactic agent, in terms of both the timing of drugapplication (before an insult versus after an insult) and the desir-able outcome (alleviation of harmful consequences of pathologicalintravascular thrombosis).
Autologous transfusion is standard practice in elective surgery.Results from human studies exploring RBC carriage of otherdrugs5,41 imply that it will be technically feasible to conjugate fibri-nolytic agents such as tPA to auto-donated RBCs and to reinjectRBC-tPA after surgery. In theory, either components of the avidin-biotin cross-linker or tPA itself might become immunogenic, thusrestricting repeated injections of the complex. Alternative bioconju-gation techniques including coupling to RBCs via activated PEG arecurrently being explored to enhance biocompatibility and minimizethe host response by repelling immune cells42,43. Coupling tPA toRBCs via PEG could extend the applicability of the proposed strategyby generating ‘universal donor’ RBC-tPA with a reasonable shelf life.Furthermore, it is possible to couple therapeutic proteins to RBCsdirectly in the bloodstream. Human RBCs possess complementreceptor 1 (CR1) that binds and transfers immune complexes tomacrophages without opsonization and phagocytosis of RBCs them-selves44. Anti-CR1 monoclonal antibodies and anti-CR1 immuno-conjugates injected into animals bind to RBCs and circulate for aprolonged time without causing cell damage45,46. In theory, an anti-CR1–tPA conjugate could be used for thromboprophylaxis withoutthe need for extraction, modification or transfusion of RBCs.
The proposed strategy may enhance the safety of cerebral fibrinol-ysis. Free tPA improves reperfusion in ischemic stroke, but mayenhance cerebral bleeding in hemorrhagic stroke31,47,48. Besidesunintended dissolution of hemostatic clots in the cerebral vascula-ture, diffusion of plasminogen activators into the brain aggravatescerebral edema and matrix remodeling and causes collateral damage,including direct cytotoxicity to neurons11,30,32,40,49. Therefore, RBC-tPA might be a safer therapy because it is better retained within thevascular compartment.
The model antithrombotic agent used in the study, tPA, representsjust one of a potentially large number of therapeutic agents whosetherapeutic profile could be enhanced or favorably altered by RBCpresentation. This study provides the first proof of principle that car-riage of therapeutic agents on the surface of RBCs has a salutaryeffect in vivo.
894 VOLUME 21 NUMBER 8 AUGUST 2003 NATURE BIOTECHNOLOGY
a
b
c
d
e
Figure 4 Prophylactic administration of RBC-tPA in mice dissolvesocclusive carotid artery thrombi resistant to free tPA. (a) Doppler ultrasoundrecording of blood flow downstream of clots induced after vascular damageby local application of FeCl3. (b) Injection of tPA (0.7 mg/kg) mixed withnon-treated RBC (N-RBC) 10 min before the occlusion. (c) Injection ofRBC-tPA (0.5 mg/kg of tPA) after occlusion. (d) Injection of the same doseof RBC-tPA 10 min before occlusion lysed the clots and restored perfusionwithin 20–30 min. (e) Comparison of the effect of free tPA (left) versusRBC-tPA (right) injected 10 min either before (open bars) or after (closedbars) thrombosis, mean ± s.e.m., n = 4 per group.
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy

A RT I C L E S
METHODSMaterials, modification of proteins, animal protocols. RBCs and tPA werebiotinylated with Biotin-LC-NHS ester (Pierce) as described4,19. Proteinswere radiolabeled with 125I-Na (Perkin Elmer) using Iodogen (Pierce) andRBCs were radiolabeled with 51Cr (ref. 4). All animal experiments were per-formed according to protocols approved by the Institutional Animal Care andUse Committee (IACUC) of the University of Pennsylvania.
Coupling of tPA to carrier red blood cells. Biotinylated tPA was coupled to b-RBCs via streptavidin (RBC-tPA), as described4. A suspension of51Cr-RBC–125I-tPA in 3% (vol/vol) BSA in PBS was stable for at least 4 d at 4 °C. To test hemolysis by complement, control RBCs, b-RBCs –SA or b-RBC–SA–b-tPA were incubated in fresh serum for 1 h at 37 °C, cen-trifuged (5 min, 1,200g), and the amount of released hemoglobin was deter-mined spectrophotometrically at 405 nm.
In vitro clot lysis. Fibrin clots were formed by adding CaCl2 and thrombin(20 mM and 0.2 units/ml final concentrations, respectively) to fibrinogen (3 mg/ml) trace-labeled with 125I-fibrinogen. To simulate lysis of preexis-ting clots, clots were overlaid with each fibrinolytic agent or saline (control)for 20 min at 20 °C. To lyse nascent clots, fibrinolytics were added directlyto 125I-fibrinogen before adding CaCl2 and thrombin. To initiate fibrinoly-sis, clots were incubated at 37 °C and the radioactivity in the supernatantswas measured in a gamma counter (Perkin Elmer).
RBC, tPA and RBC-tPA circulation in vivo. 51Cr-RBC, 125I-tPA or 51Cr-RBC–125I-tPA were injected into the tail veins of anesthetized rats ormice. At selected times, 100–200 µl of blood was taken from animals, placedin heparin and centrifuged at 1200g.The radioactivity was measured inplasma supernatants and RBC pellets. The animals were killed 1–3 h afterinjection and radioactivity in the organs was analyzed in a gamma counter.
Dissolution of clots formed from blood obtained after injections of tPA orRBC-tPA. Control RBCs, free tPA, RBC-tPA or saline (control) were injectedintravenously into rats or mice. At the indicated times, 100- to 200-µl aliquotsof blood were drawn in the absence of anticoagulant, rapidly mixed with traceamounts of 125I-fibrinogen and allowed to clot in borosilicate tubes at 20 °C.After a 20-min maturation, clots were overlaid with saline and incubated at37 °C and the release of 125I was measured.
Dissolution of pulmonary emboli by tPA or RBC-tPA. Suspensions of 125I-fib-rin microemboli (125I-ME) were prepared as described24,25 and injected intoanesthetized mice. A 350-µl sample containing a 50% suspension of RBCs,RBCs mixed with saline or tPA (0.5 mg/kg) or RBC-tPA (carrying 0.5 mg/kg oftPA) was injected through the jugular vein either 20 min before or 10 minafter injection of 125I-ME. Animals were killed 1 h after injection of 125I-ME,the lungs were isolated and rinsed in saline, and the residual radioactivity wasmeasured in a gamma counter.
Dissolution of carotid arterial thrombi by tPA or RBC-tPA. We used a mousemodel of acute carotid embolization50. Occlusion of the vessel was deter-mined by Doppler ultrasound using a 0.5VB flow probe connected to arecording system (Transonic). Complete occlusion occurred within 8–10 minof the vessel injury by FeCl3, and in control animals vessels remainedoccluded until the experiment was ended at 60 min. Either before FeCl3 appli-cation or 10 min after complete occlusion, a 350 µl sample containing eitherPBS, tPA 0.7 mg/kg alone or mixed with 50% suspension of RBCs in PBS, tPA0.5 mg/kg as a complex with RBCs at 50% suspension (RBC-tPA) in PBS, or50% suspension of control RBCs in PBS, was injected through the oppositejugular vein.
ACKNOWLEDGMENTSThe authors thank D. Neschis and A. Joshi for valuable help in preliminary animal studies, R. Taylor and D. Sakharov for fruitful discussions and advice,S. Diamond for helpful discussions and reading of the manuscript and A. Fisherfor enthusiastic support at early stages of this research. This study was supported in part by a fellowship from the Spanish Ministry of Education and Science (J.C.M.),PENN Research Foundation, AHA Bugher-Stroke Award 0170035N, NationalInstitutes of Health grant nos. HL-66442 (V.R.M.) and HL-60169 (D.B.C.).
COMPETING INTERESTS STATEMENTThe authors declare that they have no competing financial interests.
Received 4 February; accepted 23 May 2003Published online 6 July 2003; doi:10.1038/nbt846
1. Ihler, G.M., Glew, R.H. & Schnure, F.W. Enzyme loading of erythrocytes. Proc. Natl.Acad. Sci. USA 70, 2663–2666 (1973).
2. Beutler, E., Dale, G.L., Guinto, D.E. & Kuhl, W. Enzyme replacement therapy inGaucher’s disease: preliminary clinical trial of a new enzyme preparation. Proc. Natl.Acad. Sci. USA 74, 4620–4623 (1977).
3. Cavill, I., Trevett, D., Fisher, J. & Hoy, T. The measurement of the total volume of redcells in man: a non-radioactive approach using biotin. Br. J. Haematol. 70, 491–493(1988).
4. Muzykantov, V.R., Murciano, J.C., Taylor, R.P., Atochina, E.N. & Herraez, A.Regulation of the complement-mediated elimination of red blood cells modified withbiotin and streptavidin. Anal. Biochem. 241, 109–119 (1996).
5. Magnani, M. et al. Erythrocyte-mediated delivery of drugs, peptides and modifiedoligonucleotides. Gene Ther. 9, 749–751 (2002).
6. Magnani, M. et al. Targeting antiretroviral nucleoside analogues in phosphorylatedform to macrophages: in vitro and in vivo studies. Proc. Natl. Acad. Sci. USA 89,6477–6481 (1992).
7. Magnani, M. et al. Red blood cells as an antigen-delivery system. Biotechnol. Appl.Biochem. 16, 188–194 (1992).
8. Rossi, L. et al. Erythrocyte-mediated delivery of dexamethasone in patients withchronic obstructive pulmonary disease. Appl. Biochem. Biotechnol. 33, 85–89(2001).
9. Clagett, G.P., Anderson, F.A., Jr., Heit, J., Levine, M.N. & Wheeler, H.B. Prevention ofvenous thromboembolism. Chest 108, 312S–334S (1995).
10. Hirsh, J. & Hoak, J. Management of deep vein thrombosis and pulmonary embolism.A statement for healthcare professionals. Council on Thrombosis (in consultation withthe Council on Cardiovascular Radiology), American Heart Association. Circulation93, 2212–2245 (1996).
11. Gurwitz, J.H. et al. Risk for intracranial hemorrhage after tissue plasminogen activa-tor treatment for acute myocardial infarction. Participants in the National Registry ofMyocardial Infarction 2. Ann. Intern. Med. 129, 597–604 (1998).
12. Deklunder, G., Roussel, M., Lecroart, J.L., Prat, A. & Gautier, C. Microemboli in cere-bral circulation and alteration of cognitive abilities in patients with mechanical pros-thetic heart valves. Stroke 29, 1821–1826 (1998).
13. Brown, W.R., Moody, D.M., Challa, V.R., Stump, D.A. & Hammon, J.W. Longer dura-tion of cardiopulmonary bypass is associated with greater numbers of cerebralmicroemboli. Stroke 31, 707–713 (2000).
14. Imperiale, T.F. & Speroff, T. A meta-analysis of methods to prevent venous throm-boembolism following total hip replacement. JAMA 271, 1780–1785 (1994).
15. Holvoet, P. et al. Thrombolytic profiles of clot-targeted plasminogen activators.Parameters determining potency and initial and maximal rates. Circulation 87,1007–1016 (1993).
16. Anand, S. & Diamond, S.L. Computer simulation of systemic circulation and clot lysisdynamics during thrombolytic therapy that accounts for inner clot transport and reac-tion. Circulation 94, 763–774 (1996).
17. Sakharov, D.V. & Rijken, D.C. Superficial accumulation of plasminogen duringplasma clot lysis. Circulation 92, 1883–1890 (1995).
18. Kalofonos, H.P. et al. Imaging of tumor in patients with indium-111-labeled biotinand streptavidin-conjugated antibodies: preliminary communication. J. Nucl. Med.31, 1791–1796 (1990).
19. Muzykantov, V.R. et al. Targeting of antibody-conjugated plasminogen activators tothe pulmonary vasculature. J. Pharmacol. Exp. Ther. 279, 1026–1034 (1996).
20. Paganelli, G. et al. Two-step tumour targetting in ovarian cancer patients usingbiotinylated monoclonal antibodies and radioactive streptavidin. Eur. J. Nucl. Med.19, 322–329 (1992).
21. Smirnov, V.N. et al. Carrier-directed targeting of liposomes and erythrocytes todenuded areas of vessel wall. Proc. Natl. Acad. Sci. USA 83, 6603–6607 (1986).
22. Muzykantov, V.R., Smirnov, M.D. & Samokhin, G.P. Avidin attachment to biotinylatederythrocytes induces homologous lysis via the alternative pathway of complement.Blood 78, 2611–2618 (1991).
23. Zaltzman, A.B., Van den Berg, C.W., Muzykantov, V.R. & Morgan, B.P. Enhanced com-plement susceptibility of avidin-biotin-treated human erythrocytes is a consequenceof neutralization of the complement regulators CD59 and decay accelerating factor.Biochem. J. 307 (Pt 3), 651–656 (1995).
24. Murciano, J.C. et al. Platelets inhibit the lysis of pulmonary microemboli. Am. J.Physiol. Lung Cell. Mol. Physiol. 282, L529–L539 (2002).
25. Bdeir, K. et al. Urokinase mediates fibrinolysis in the pulmonary microvasculature.Blood 96, 1820–1826 (2000).
26. Collen, D. Fibrin-selective thrombolytic therapy for acute myocardial infarction.Circulation 93, 857–865 (1996).
27. Carmeliet, P. et al. Physiological consequences of loss of plasminogen activator genefunction in mice. Nature 368, 419–424 (1994).
28. Bu, G., Williams, S., Strickland, D.K. & Schwartz, A.L. Low density lipoprotein recep-tor-related protein/α2-macroglobulin receptor is an hepatic receptor for tissue-typeplasminogen activator. Proc. Natl. Acad. Sci. USA 89, 7427–7431 (1992).
29. Rogove, A.D., Siao, C., Keyt, B., Strickland, S. & Tsirka, S.E. Activation of microgliareveals a non-proteolytic cytokine function for tissue plasminogen activator in thecentral nervous system. J. Cell Sci. 112, 4007–4016 (1999).
NATURE BIOTECHNOLOGY VOLUME 21 NUMBER 8 AUGUST 2003 895
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy

A RT I C L E S
30. Tsirka, S.E., Gualandris, A., Amaral, D.G. & Strickland, S. Excitotoxin-induced neu-ronal degeneration and seizure are mediated by tissue plasminogen activator. Nature377, 340–344 (1995).
31. Figueroa, B.E., Keep, R.F., Betz, A.L. & Hoff, J.T. Plasminogen activators potentiatethrombin-induced brain injury. Stroke 29, 1202–1207; discussion 1208 (1998).
32. Gore, J.M. et al. Stroke after thrombolysis. Mortality and functional outcomes in theGUSTO-I trial. Global Use of Strategies to Open Occluded Coronary Arteries.Circulation 92, 2811–2818 (1995).
33. Runge, M.S. et al. Enhanced thrombolytic and antithrombotic potency of a fibrin-targeted plasminogen activator in baboons. Circulation 94, 1412–1422 (1996).
34. Krishnamurti, C., Keyt, B., Maglasang, P. & Alving, B.M. PAI-1-resistant t-PA: lowdoses prevent fibrin deposition in rabbits with increased PAI-1 activity. Blood 87,14–19 (1996).
35. Benedict, C.R. et al. New variant of human tissue plasminogen activator (TPA) withenhanced efficacy and lower incidence of bleeding compared with recombinanthuman TPA. Circulation 92, 3032-–3040 (1995).
36. Reed, G.L. et al. A catalytic switch and the conversion of streptokinase to a fibrin-targeted plasminogen activator. Proc. Natl. Acad. Sci. USA 96, 8879–8883(1999).
37. Abuchowski, A., van Es, T., Palczuk, N.C. & Davis, F.F. Alteration of immunologicalproperties of bovine serum albumin by covalent attachment of polyethylene glycol.J. Biol. Chem. 252, 3578–3581 (1977).
38. Langer, R. Drug delivery and targeting. Nature 392, 5–10 (1998).39. Heeremans, J.L. et al. Thrombolytic treatment with tissue-type plasminogen activa-
tor (t-PA) containing liposomes in rabbits: a comparison with free t-PA. Thromb.Haemost. 73, 488–494 (1995).
40. Chen, Z.L. & Strickland, S. Neuronal death in the hippocampus is promoted by plas-min-catalyzed degradation of laminin. Cell 91, 917–925 (1997).
41. Bax, B.E., Bain, M.D., Fairbanks, L.D., Webster, A.D. & Chalmers, R.A. In vitro and
in vivo studies with human carrier erythrocytes loaded with polyethylene glycol-con-jugated and native adenosine deaminase. Br. J. Haematol. 109, 549–554 (2000).
42. Blackall, D.P., Armstrong, J.K., Meiselman, H.J. & Fisher, T.C. Polyethylene glycol-coated red blood cells fail to bind glycophorin A-specific antibodies and are impervi-ous to invasion by the Plasmodium falciparum malaria parasite. Blood 97, 551–556(2001).
43. Scott, M.D., Murad, K.L., Koumpouras, F., Talbot, M. & Eaton, J.W. Chemical camou-flage of antigenic determinants: stealth erythrocytes. Proc. Natl. Acad. Sci. USA 94,7566–7571 (1997).
44. Cornacoff, J.B. et al. Primate erythrocyte-immune complex-clearing mechanism. J.Clin. Invest. 71, 236–247 (1983).
45. Taylor, R.P. et al. Use of heteropolymeric monoclonal antibodies to attach antigens tothe C3b receptor of human erythrocytes: a potential therapeutic treatment. Proc.Natl. Acad. Sci. USA 88, 3305–3309 (1991).
46. Muzykantov, V.R. & Taylor, R.P. Attachment of biotinylated antibody to red bloodcells: antigen-binding capacity of immunoerythrocytes and their susceptibility to lysisby complement. Anal. Biochem. 223, 142–148 (1994).
47. Kwiatkowski, T.G. et al. Effects of tissue plasminogen activator for acute ischemicstroke at one year. National Institute of Neurological Disorders and StrokeRecombinant Tissue Plasminogen Activator Stroke Study Group. N. Engl. J. Med.340, 1781–1787 (1999).
48. Nagai, N., De Mol, M., Lijnen, H.R., Carmeliet, P. & Collen, D. Role of plasminogensystem components in focal cerebral ischemic infarction: a gene targeting and genetransfer study in mice. Circulation 99, 2440–2444 (1999).
49. Wang, Y.F. et al. Tissue plasminogen activator (tPA) increases neuronal damage afterfocal cerebral ischemia in wild-type and tPA-deficient mice. Nat. Med. 4, 228–231(1998).
50. Farrehi, P.M., Ozaki, C.K., Carmeliet, P. & Fay, W.P. Regulation of arterial thromboly-sis by plasminogen activator inhibitor-1 in mice. Circulation 97, 1002–1008 (1998).
896 VOLUME 21 NUMBER 8 AUGUST 2003 NATURE BIOTECHNOLOGY
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reb
iote
chn
olo
gy