progressive infection in a subset of hiv-1–positive chimpanzees
TRANSCRIPT

1051
Progressive Infection in a Subset of HIV-1–Positive Chimpanzees
Shawn P. O’Neil,1,2,4 Francis J. Novembre,1,4
Anne Brodie Hill,2 Carolyn Suwyn,1,2 Clyde E. Hart,6
Tammy Evans-Strickfaden,6 Daniel C. Anderson,2
Juliette deRosayro,1 James G. Herndon,3
Michelle Saucier,1 and Harold M. McClure2,5
Divisions of 1Microbiology and Immunology, 2Research Resources,and 3Neuroscience, Yerkes Regional Primate Research Center,
Departments of 4Microbiology and Immunology and 5Pathologyand Laboratory Medicine, Emory University School of Medicine,
and 6Retroviral Diseases Branch, Division of AIDS, STD, and TBLaboratory Research, Centers for Disease Control and Prevention,
Atlanta, Georgia
Chimpanzees are susceptible to infection with human immunodeficiency virus (HIV)–1;however, infected animals usually maintain normal numbers of CD4+ T lymphocytes and donot develop immunodeficiency. We have examined 10 chronically infected HIV-1–positivechimpanzees for evidence of progressive infection. In addition to 1 animal that developedAIDS, 3 chimpanzees exhibit evidence of progressive HIV infection. All progressors have lowCD4+ T cell counts (!200 cells/mL), severe CD4:CD8 inversion, and marked reduction ininterleukin-2 receptor expression by CD4+ T cells. In comparison with HIV-positive nonpro-gressor chimpanzees, progressors have higher plasma and lymphoid virus loads, greater CD38expression in CD8+/HLA-DR+ T cells, and greater serum concentrations of soluble tumornecrosis factor type II receptors and b2-microglobulin, all markers of HIV progression inhumans. These observations show that progressive HIV-1 infection can occur in chimpanzeesand suggest that the pathogenesis of progressive infection in this species resembles that inhumans.
Recent studies show that the human immunodeficiency virus(HIV)–1 pandemic originated through cross-species transmis-sion of simian immunodeficiency virus (SIV)cpz from its naturalhost, the chimpanzee (Pan troglodytes), to humans in westequatorial Africa [1–3] . Like many other “unnatural” primatehosts, most HIV-positive humans experience progressive infec-tion, characterized by persistently elevated levels of virus inplasma and lymphoid tissues, chronic immune activation, re-lentless reductions in the number and function of CD41 T cells,and development of immunodeficiency [4–10]. A small per-centage of infected individuals, however, experience nonpro-gressive infections. These individuals, collectively referred to aslong-term nonprogressors (LTNPs), have substantially lowerlevels of virus in plasma, normal, stable numbers of CD41 T
Received 30 March 2000; revised 10 July 2000; electronically published8 September 2000.
Presented in part: 14th Annual Symposium on Nonhuman Primate Mod-els for AIDS, Portland, Oregon, 23–26 October 1996 (abstract 48).
All chimpanzees were housed at the Yerkes Regional Primate ResearchCenter, in accordance with the guidelines established by the Animal WelfareAct and the National Institutes of Health (NIH) guide for care and use oflaboratory animals. The Yerkes Center is fully accredited by the AmericanAssociation for the Accreditation of Laboratory Animal Care.
Financial support: NIH grants R01-AI40879, R01-AI38501, and RR-00165.
Reprints or correspondence: Dr. Shawn P. O’Neil, Yerkes Regional Pri-mate Research Center, 954 N. Gatewood Rd., Atlanta, GA 30329 ([email protected]).
The Journal of Infectious Diseases 2000;182:1051–62q 2000 by the Infectious Diseases Society of America. All rights reserved.0022-1899/2000/18204-0006$02.00
cells, and vigorous antiviral immune responses, and they donot develop AIDS [11–20] .
On the basis of a few documented cases of natural infection,SIVcpz appears to be nonpathogenic in its natural chimpanzeehost, recapitulating the relationship observed between otherSIV isolates and their respective natural, nonhuman primatehosts [2, 3, 21–23]. It is, therefore, not surprising that the vi-rologic and immunopathologic characteristics of progressiveHIV infection in humans are not typically observed in chim-panzees that have been experimentally infected with HIV-1 [24].Although chimpanzees are readily susceptible to infection[25–27], most HIV-1–positive chimpanzees mount vigorous an-tiviral humoral and cell-mediated immune responses, maintainnormal numbers of CD41 T lymphocytes, harbor low plasmavirus loads, and remain healthy, similar to human LTNPs[28–38]. With the exception of a single documented case of thedevelopment of AIDS [39], progressive HIV infection has notbeen described in chronically infected chimpanzees.
Several hypotheses have been advanced that attempt to ex-plain the apparent lack of disease progression in HIV-infectedchimpanzees [40–47]. Prevailing theories include the absence ofchronic immune activation, resistance of chimpanzee mono-cyte-macrophages to infection with primary HIV isolates, pres-ervation of CD41 T helper cell regenerative capacity, absenceof HIV-1–induced autoimmune phenomena, and absence ofcytotoxic CD81 T cell infiltration and degenerative changes inlymphoid follicles. It has been difficult to assess the relativesignificance of each of these pathogenic mechanisms to diseaseprogression, however, in the absence of a population of chim-panzees with pathogenic infections.
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021

1052 O’Neil et al. JID 2000;182 (October)
Table 1. Chronically infected human immunodeficiency virus (HIV)–positive chimpanzees at the Yerkes Regional PrimateResearch Center.
Animal Sex AgeaInoculation
date Inoculum Route Years HIV positiveb Current status
C463 M 5 8/2/84 Transfusion (whole blood) from C459 iv 14 ProgressorC499 M 3 4/8/85 HIV-1SF2, cell free iv 11 Progressorc
4 7/18/86 HIV-1LAV-1b, cell free iv5 1/20/87 HIV-1NDK, cell free iv
C550 F 6 3/4/85 HIV-1LAV-1b, cell free Intravaginal 14 Progressor7 4/8/86 LAV-2 (HIV-2EHO), cell free iv
C560 F 4 10/17/84 HIV-1LAV-1b, cell free iv 14 Progressor5 4/8/85 HIV-1SF2, cell free iv
C459 M 4 3/5/84 HIV-1LAV-1, cell associated iv 15 NonprogressorC487 M 3 7/30/84 HIV-1LAV-1b, cell associated iv 14 NonprogressorC497 M 5 1/20/87 HIV-1NDK, cell free iv 12 NonprogressorC542 F 8 9/8/86 HIV-1LAV-1b, cell free (following vaccination)d iv 12 NonprogressorC544 F 8 9/8/86 HIV-1LAV-1b, cell free iv 12 NonprogressorC552 F 7 9/8/86 HIV-1LAV-1b, cell free (following vaccination)d iv 12 Nonprogressor
NOTE. iv, intravenous; LAV, lymphadenopathy-associated virus.a Age in years at inoculation.b Years postinoculation, as of 1/1/99 (rounded off to nearest year).c Euthanatized 2/13/96 due to the development of AIDS.d Vaccinated with a recombinant vaccinia virus expressing the envelope glycoproteins of HIVLAV-1 prior to challenge with LAV-1b.
In this study, we examined 10 HIV-1–positive chimpanzeeswith chronic infections (seropositive for 110 years) for evidenceof progressive infection, applying virologic, immunologic, se-rologic, and histopathologic markers that have been used tocharacterize progressive infection in humans. Six of these an-imals exhibit evidence of stable, apathogenic HIV infection(HIV-positive nonprogressors [HIV/NPs]). However, in addi-tion to the animal that developed AIDS (C499), we have iden-tified 3 chronically infected chimpanzees that exhibit evidenceof progressive HIV infection (HIV-positive progressors [HIV/Ps]).
Materials and Methods
Lymphocyte subset analysis. Lymphocyte phenotype and ac-tivation state were determined in specimens of whole blood con-taining EDTA anticoagulant by 2- and 3-color flow cytometry,using fluorochrome-conjugated monoclonal antibodies that rec-ognize various lymphocyte surface antigens. Absolute numbers ofCD21/CD41 T lymphocytes were measured in all HIV-1–infectedchimpanzees from 1989 through 1990 and from 1993 through 1999,using the antibodies Leu-5b and Leu-3a (anti-CD2 and -CD4, re-spectively; Becton Dickinson/Pharmingen, San Diego). Three HIV-negative, age-matched chimpanzees were used as controls from1989 to 1996; however, 2 of these animals were subsequently in-oculated with HIV. Thus, blood specimens collected from 9 age-matched, HIV-negative chimpanzees in 1997 were used as controlsfor group comparisons from 1997 through 1999. CD31/CD41 Tcells were enumerated in blood collected from uninfected controlsin 1997 and from HIV-positive chimpanzees in 1998 and 1999, bygating on lymphocytes and using antibodies directed against CD3and CD4 (Becton Dickinson/Pharmingen). The activation state ofCD41 and CD81 T cells in blood specimens collected from HIV-positive chimpanzees in 1999 (i.e., excluding C459 and C499) was
compared with data collected in 1997 from 10 age-matched, HIV-negative controls. The antibodies used recognize CD8 (Leu-2a),major histocompatibility complex (MHC) class II (HLA-DR),CD25 (interleukin-2 receptor [IL-2R]), CD45RA, and CD38 (Bec-ton Dickinson) and CD29 (4B4; Coulter Corp., Miami).
Plasma virus load. The quantity of virion-associated RNA wasmeasured in plasma specimens by quantitative competitive poly-merase chain reaction, using a microplate-based detection system,as described elsewhere [48]. The lower limit of detection for thisassay is 800 RNA copies/mL plasma.
Assays for soluble immune activation markers. The concentra-tions of soluble tumor necrosis factor type II receptors (sTNF-RII)and b2-microglobulin (b2-M) were measured in serum specimensby commercial ELISA (Quantikine; R&D Systems, Minneapolis).
Lymph nodes. Lymph node biopsy specimens were obtainedfrom all 10 HIV-infected chimpanzees and 6 HIV-negative, age-matched controls. Two separate specimens were obtained from eachHIV-positive animal except C459 (1 specimen). The duration oftime between biopsies was 2 years for all animals except C499, forwhich 1 lymph node biopsy specimen was available, as well aslymph nodes obtained 5 months later at necropsy. Specimens werefixed in 10% formalin and embedded in paraffin.
Tissue virus burden and pattern of virus distribution. HIV RNAwas localized in paraffin sections of lymph node tissue by in situhybridization (ISH), using methods described elsewhere for thelocalization of SIV RNA [49]. Chimpanzee lymph node sectionswere hybridized with a digoxigenin-labeled antisense riboprobecocktail that spans the entire length of the HIV genome (LofstrandLabs Ltd., Gaithersburg, MD). As positive controls, each assayincluded lymph node sections from C499, which had been incu-bated with HIV antisense riboprobe. Negative controls includedlymph node sections from C499 that had been incubated with HIVsense riboprobe, as well as lymph node sections from uninfectedchimpanzees that had been incubated with HIV antisense ribo-probe. The relative lymph node virus burden was determined by
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021
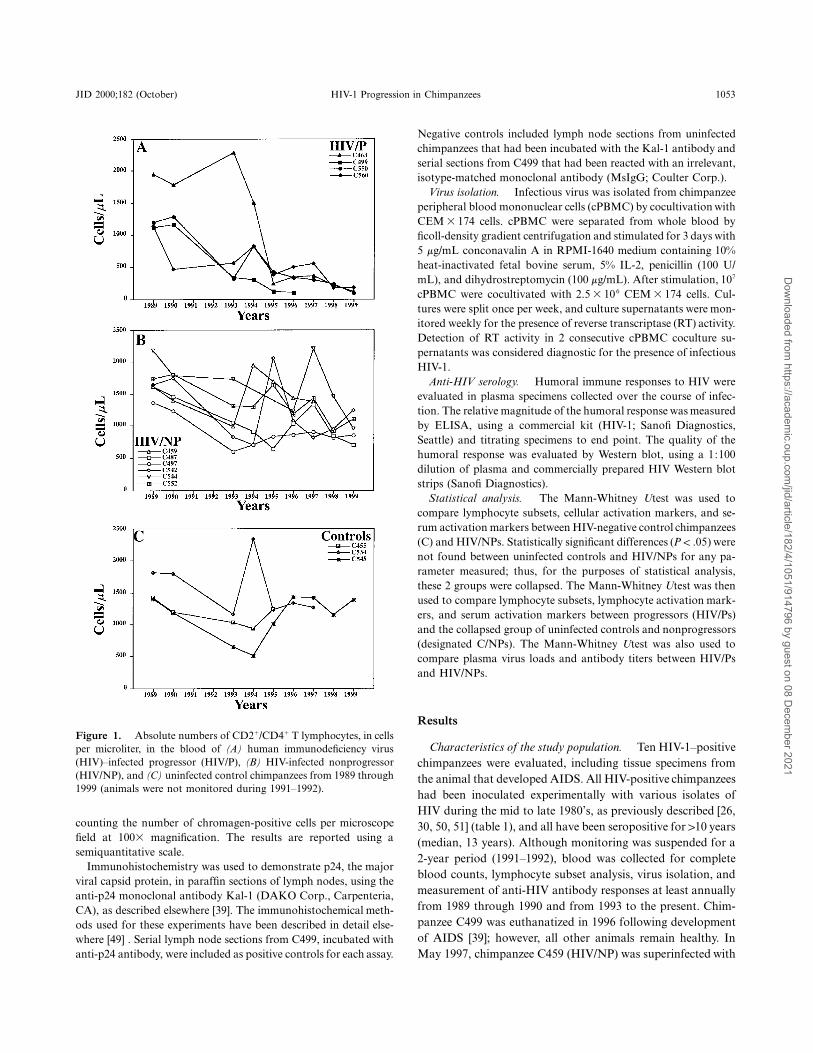
JID 2000;182 (October) HIV-1 Progression in Chimpanzees 1053
Figure 1. Absolute numbers of CD21/CD41 T lymphocytes, in cellsper microliter, in the blood of (A) human immunodeficiency virus(HIV)–infected progressor (HIV/P), (B) HIV-infected nonprogressor(HIV/NP), and (C) uninfected control chimpanzees from 1989 through1999 (animals were not monitored during 1991–1992).
counting the number of chromagen-positive cells per microscopefield at 1003 magnification. The results are reported using asemiquantitative scale.
Immunohistochemistry was used to demonstrate p24, the majorviral capsid protein, in paraffin sections of lymph nodes, using theanti-p24 monoclonal antibody Kal-1 (DAKO Corp., Carpenteria,CA), as described elsewhere [39]. The immunohistochemical meth-ods used for these experiments have been described in detail else-where [49] . Serial lymph node sections from C499, incubated withanti-p24 antibody, were included as positive controls for each assay.
Negative controls included lymph node sections from uninfectedchimpanzees that had been incubated with the Kal-1 antibody andserial sections from C499 that had been reacted with an irrelevant,isotype-matched monoclonal antibody (MsIgG; Coulter Corp.).
Virus isolation. Infectious virus was isolated from chimpanzeeperipheral blood mononuclear cells (cPBMC) by cocultivation with
cells. cPBMC were separated from whole blood byCEM 3 174ficoll-density gradient centrifugation and stimulated for 3 days with5 mg/mL conconavalin A in RPMI-1640 medium containing 10%heat-inactivated fetal bovine serum, 5% IL-2, penicillin (100 U/mL), and dihydrostreptomycin (100 mg/mL). After stimulation, 107
cPBMC were cocultivated with cells. Cul-62.5 3 10 CEM 3 174tures were split once per week, and culture supernatants were mon-itored weekly for the presence of reverse transcriptase (RT) activity.Detection of RT activity in 2 consecutive cPBMC coculture su-pernatants was considered diagnostic for the presence of infectiousHIV-1.
Anti-HIV serology. Humoral immune responses to HIV wereevaluated in plasma specimens collected over the course of infec-tion. The relative magnitude of the humoral response was measuredby ELISA, using a commercial kit (HIV-1; Sanofi Diagnostics,Seattle) and titrating specimens to end point. The quality of thehumoral response was evaluated by Western blot, using a 1:100dilution of plasma and commercially prepared HIV Western blotstrips (Sanofi Diagnostics).
Statistical analysis. The Mann-Whitney Utest was used tocompare lymphocyte subsets, cellular activation markers, and se-rum activation markers between HIV-negative control chimpanzees(C) and HIV/NPs. Statistically significant differences ( ) wereP ! .05not found between uninfected controls and HIV/NPs for any pa-rameter measured; thus, for the purposes of statistical analysis,these 2 groups were collapsed. The Mann-Whitney Utest was thenused to compare lymphocyte subsets, lymphocyte activation mark-ers, and serum activation markers between progressors (HIV/Ps)and the collapsed group of uninfected controls and nonprogressors(designated C/NPs). The Mann-Whitney Utest was also used tocompare plasma virus loads and antibody titers between HIV/Psand HIV/NPs.
Results
Characteristics of the study population. Ten HIV-1–positivechimpanzees were evaluated, including tissue specimens fromthe animal that developed AIDS. All HIV-positive chimpanzeeshad been inoculated experimentally with various isolates ofHIV during the mid to late 1980’s, as previously described [26,30, 50, 51] (table 1), and all have been seropositive for 110 years(median, 13 years). Although monitoring was suspended for a2-year period (1991–1992), blood was collected for completeblood counts, lymphocyte subset analysis, virus isolation, andmeasurement of anti-HIV antibody responses at least annuallyfrom 1989 through 1990 and from 1993 to the present. Chim-panzee C499 was euthanatized in 1996 following developmentof AIDS [39]; however, all other animals remain healthy. InMay 1997, chimpanzee C459 (HIV/NP) was superinfected with
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021
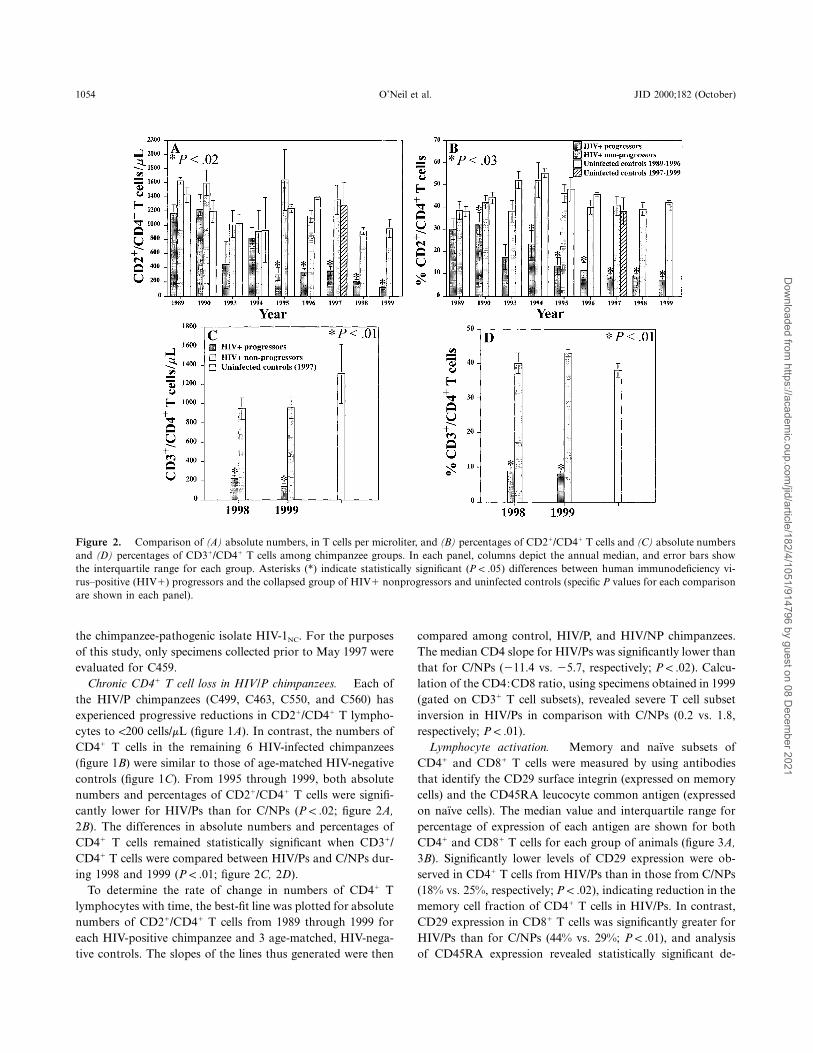
1054 O’Neil et al. JID 2000;182 (October)
Figure 2. Comparison of (A) absolute numbers, in T cells per microliter, and (B) percentages of CD21/CD41 T cells and (C) absolute numbersand (D) percentages of CD31/CD41 T cells among chimpanzee groups. In each panel, columns depict the annual median, and error bars showthe interquartile range for each group. Asterisks (*) indicate statistically significant ( ) differences between human immunodeficiency vi-P ! .05rus–positive (HIV1) progressors and the collapsed group of HIV1 nonprogressors and uninfected controls (specific P values for each comparisonare shown in each panel).
the chimpanzee-pathogenic isolate HIV-1NC. For the purposesof this study, only specimens collected prior to May 1997 wereevaluated for C459.
Chronic CD41 T cell loss in HIV/P chimpanzees. Each ofthe HIV/P chimpanzees (C499, C463, C550, and C560) hasexperienced progressive reductions in CD21/CD41 T lympho-cytes to !200 cells/mL (figure 1A). In contrast, the numbers ofCD41 T cells in the remaining 6 HIV-infected chimpanzees(figure 1B) were similar to those of age-matched HIV-negativecontrols (figure 1C). From 1995 through 1999, both absolutenumbers and percentages of CD21/CD41 T cells were signifi-cantly lower for HIV/Ps than for C/NPs ( ; figure 2A,P ! .022B). The differences in absolute numbers and percentages ofCD41 T cells remained statistically significant when CD31/CD41 T cells were compared between HIV/Ps and C/NPs dur-ing 1998 and 1999 ( ; figure 2C, 2D).P ! .01
To determine the rate of change in numbers of CD41 Tlymphocytes with time, the best-fit line was plotted for absolutenumbers of CD21/CD41 T cells from 1989 through 1999 foreach HIV-positive chimpanzee and 3 age-matched, HIV-nega-tive controls. The slopes of the lines thus generated were then
compared among control, HIV/P, and HIV/NP chimpanzees.The median CD4 slope for HIV/Ps was significantly lower thanthat for C/NPs (211.4 vs. 25.7, respectively; ). Calcu-P ! .02lation of the CD4:CD8 ratio, using specimens obtained in 1999(gated on CD31 T cell subsets), revealed severe T cell subsetinversion in HIV/Ps in comparison with C/NPs (0.2 vs. 1.8,respectively; ).P ! .01
Lymphocyte activation. Memory and naı̈ve subsets ofCD41 and CD81 T cells were measured by using antibodiesthat identify the CD29 surface integrin (expressed on memorycells) and the CD45RA leucocyte common antigen (expressedon naı̈ve cells). The median value and interquartile range forpercentage of expression of each antigen are shown for bothCD41 and CD81 T cells for each group of animals (figure 3A,3B). Significantly lower levels of CD29 expression were ob-served in CD41 T cells from HIV/Ps than in those from C/NPs(18% vs. 25%, respectively; ), indicating reduction in theP ! .02memory cell fraction of CD41 T cells in HIV/Ps. In contrast,CD29 expression in CD81 T cells was significantly greater forHIV/Ps than for C/NPs (44% vs. 29%; ), and analysisP ! .01of CD45RA expression revealed statistically significant de-
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021
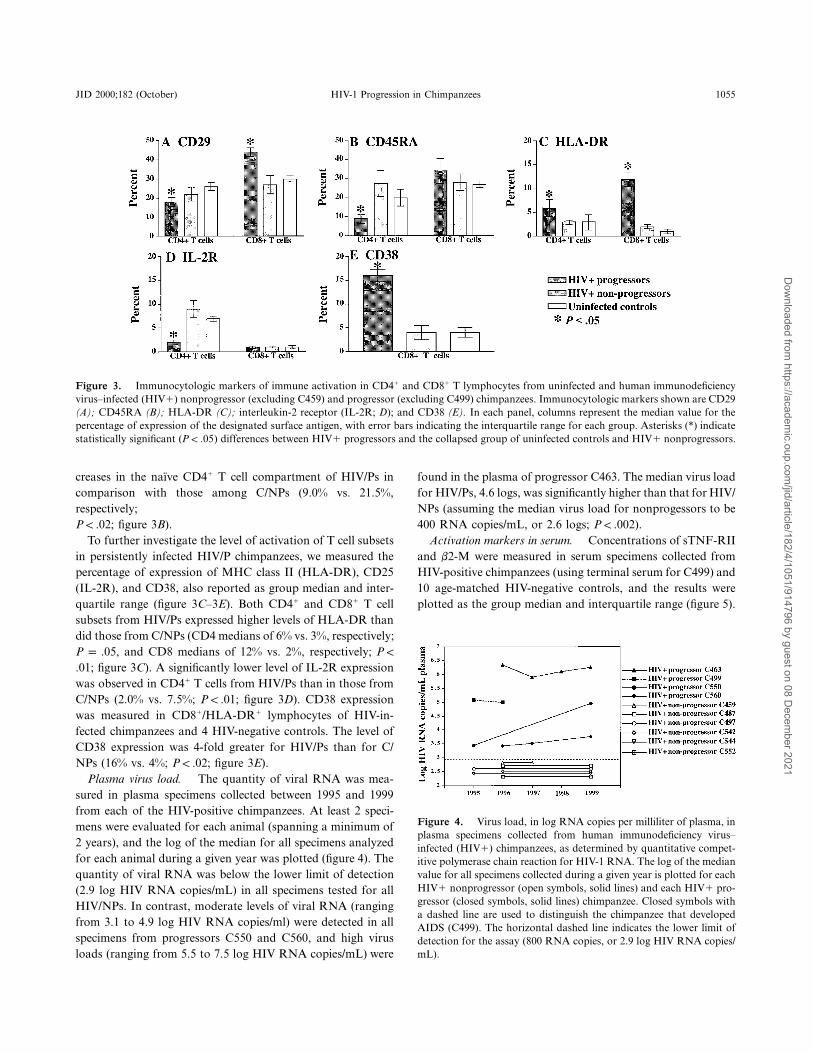
JID 2000;182 (October) HIV-1 Progression in Chimpanzees 1055
Figure 3. Immunocytologic markers of immune activation in CD41 and CD81 T lymphocytes from uninfected and human immunodeficiencyvirus–infected (HIV1) nonprogressor (excluding C459) and progressor (excluding C499) chimpanzees. Immunocytologic markers shown are CD29(A); CD45RA (B); HLA-DR (C); interleukin-2 receptor (IL-2R; D); and CD38 (E). In each panel, columns represent the median value for thepercentage of expression of the designated surface antigen, with error bars indicating the interquartile range for each group. Asterisks (*) indicatestatistically significant ( ) differences between HIV1 progressors and the collapsed group of uninfected controls and HIV1 nonprogressors.P ! .05
Figure 4. Virus load, in log RNA copies per milliliter of plasma, inplasma specimens collected from human immunodeficiency virus–infected (HIV1) chimpanzees, as determined by quantitative compet-itive polymerase chain reaction for HIV-1 RNA. The log of the medianvalue for all specimens collected during a given year is plotted for eachHIV1 nonprogressor (open symbols, solid lines) and each HIV1 pro-gressor (closed symbols, solid lines) chimpanzee. Closed symbols witha dashed line are used to distinguish the chimpanzee that developedAIDS (C499). The horizontal dashed line indicates the lower limit ofdetection for the assay (800 RNA copies, or 2.9 log HIV RNA copies/mL).
creases in the naı̈ve CD41 T cell compartment of HIV/Ps incomparison with those among C/NPs (9.0% vs. 21.5%,respectively;
; figure 3B).P ! .02To further investigate the level of activation of T cell subsets
in persistently infected HIV/P chimpanzees, we measured thepercentage of expression of MHC class II (HLA-DR), CD25(IL-2R), and CD38, also reported as group median and inter-quartile range (figure 3C–3E). Both CD41 and CD81 T cellsubsets from HIV/Ps expressed higher levels of HLA-DR thandid those from C/NPs (CD4 medians of 6% vs. 3%, respectively;
, and CD8 medians of 12% vs. 2%, respectively;P p .05 P !
; figure 3C). A significantly lower level of IL-2R expression.01was observed in CD41 T cells from HIV/Ps than in those fromC/NPs (2.0% vs. 7.5%; ; figure 3D). CD38 expressionP ! .01was measured in CD81/HLA-DR1 lymphocytes of HIV-in-fected chimpanzees and 4 HIV-negative controls. The level ofCD38 expression was 4-fold greater for HIV/Ps than for C/NPs (16% vs. 4%; ; figure 3E).P ! .02
Plasma virus load. The quantity of viral RNA was mea-sured in plasma specimens collected between 1995 and 1999from each of the HIV-positive chimpanzees. At least 2 speci-mens were evaluated for each animal (spanning a minimum of2 years), and the log of the median for all specimens analyzedfor each animal during a given year was plotted (figure 4). Thequantity of viral RNA was below the lower limit of detection(2.9 log HIV RNA copies/mL) in all specimens tested for allHIV/NPs. In contrast, moderate levels of viral RNA (rangingfrom 3.1 to 4.9 log HIV RNA copies/ml) were detected in allspecimens from progressors C550 and C560, and high virusloads (ranging from 5.5 to 7.5 log HIV RNA copies/mL) were
found in the plasma of progressor C463. The median virus loadfor HIV/Ps, 4.6 logs, was significantly higher than that for HIV/NPs (assuming the median virus load for nonprogessors to be400 RNA copies/mL, or 2.6 logs; ).P ! .002
Activation markers in serum. Concentrations of sTNF-RIIand b2-M were measured in serum specimens collected fromHIV-positive chimpanzees (using terminal serum for C499) and10 age-matched HIV-negative controls, and the results wereplotted as the group median and interquartile range (figure 5).
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021
1056 O’Neil et al. JID 2000;182 (October)
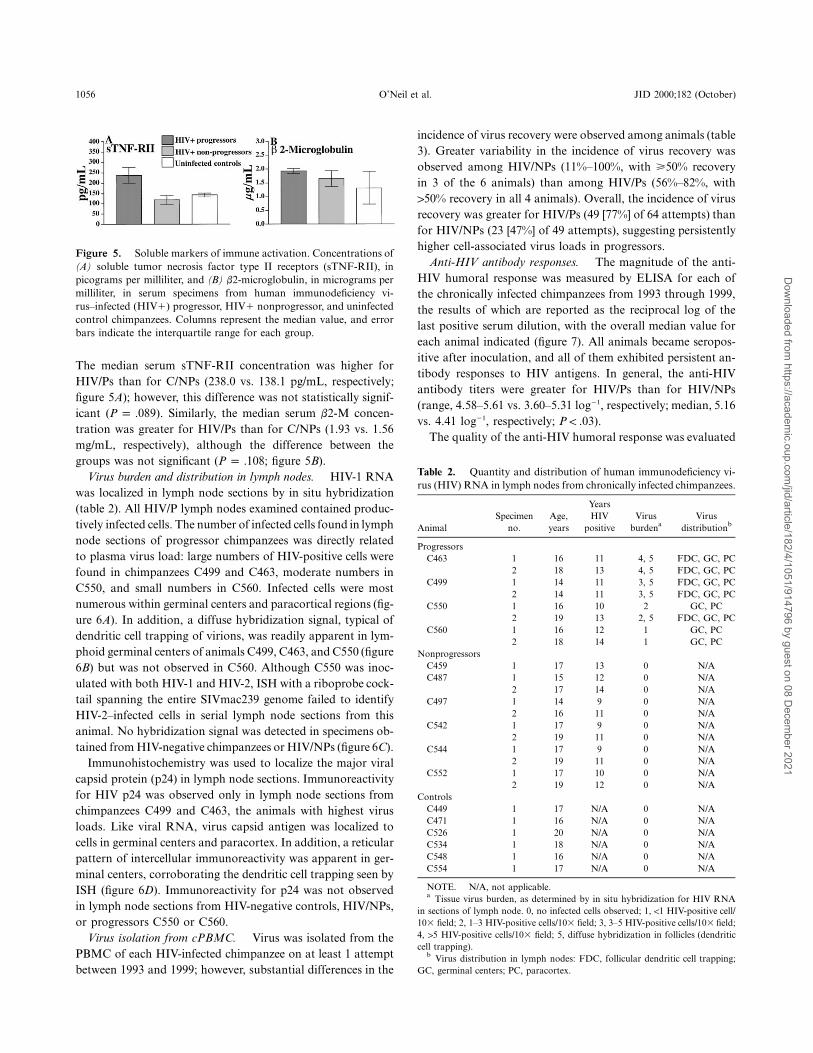
Figure 5. Soluble markers of immune activation. Concentrations of(A) soluble tumor necrosis factor type II receptors (sTNF-RII), inpicograms per milliliter, and (B) b2-microglobulin, in micrograms permilliliter, in serum specimens from human immunodeficiency vi-rus–infected (HIV1) progressor, HIV1 nonprogressor, and uninfectedcontrol chimpanzees. Columns represent the median value, and errorbars indicate the interquartile range for each group.
Table 2. Quantity and distribution of human immunodeficiency vi-rus (HIV) RNA in lymph nodes from chronically infected chimpanzees.
AnimalSpecimen
no.Age,years
YearsHIV
positiveVirus
burdenaVirus
distributionb
ProgressorsC463 1 16 11 4, 5 FDC, GC, PC
2 18 13 4, 5 FDC, GC, PCC499 1 14 11 3, 5 FDC, GC, PC
2 14 11 3, 5 FDC, GC, PCC550 1 16 10 2 GC, PC
2 19 13 2, 5 FDC, GC, PCC560 1 16 12 1 GC, PC
2 18 14 1 GC, PCNonprogressors
C459 1 17 13 0 N/AC487 1 15 12 0 N/A
2 17 14 0 N/AC497 1 14 9 0 N/A
2 16 11 0 N/AC542 1 17 9 0 N/A
2 19 11 0 N/AC544 1 17 9 0 N/A
2 19 11 0 N/AC552 1 17 10 0 N/A
2 19 12 0 N/AControls
C449 1 17 N/A 0 N/AC471 1 16 N/A 0 N/AC526 1 20 N/A 0 N/AC534 1 18 N/A 0 N/AC548 1 16 N/A 0 N/AC554 1 17 N/A 0 N/A
NOTE. N/A, not applicable.a Tissue virus burden, as determined by in situ hybridization for HIV RNA
in sections of lymph node. 0, no infected cells observed; 1, !1 HIV-positive cell/103 field; 2, 1–3 HIV-positive cells/103 field; 3, 3–5 HIV-positive cells/103 field;4, 15 HIV-positive cells/103 field; 5, diffuse hybridization in follicles (dendriticcell trapping).
b Virus distribution in lymph nodes: FDC, follicular dendritic cell trapping;GC, germinal centers; PC, paracortex.
The median serum sTNF-RII concentration was higher forHIV/Ps than for C/NPs (238.0 vs. 138.1 pg/mL, respectively;figure 5A); however, this difference was not statistically signif-icant ( ). Similarly, the median serum b2-M concen-P p .089tration was greater for HIV/Ps than for C/NPs (1.93 vs. 1.56mg/mL, respectively), although the difference between thegroups was not significant ( ; figure 5B).P p .108
Virus burden and distribution in lymph nodes. HIV-1 RNAwas localized in lymph node sections by in situ hybridization(table 2). All HIV/P lymph nodes examined contained produc-tively infected cells. The number of infected cells found in lymphnode sections of progressor chimpanzees was directly relatedto plasma virus load: large numbers of HIV-positive cells werefound in chimpanzees C499 and C463, moderate numbers inC550, and small numbers in C560. Infected cells were mostnumerous within germinal centers and paracortical regions (fig-ure 6A). In addition, a diffuse hybridization signal, typical ofdendritic cell trapping of virions, was readily apparent in lym-phoid germinal centers of animals C499, C463, and C550 (figure6B) but was not observed in C560. Although C550 was inoc-ulated with both HIV-1 and HIV-2, ISH with a riboprobe cock-tail spanning the entire SIVmac239 genome failed to identifyHIV-2–infected cells in serial lymph node sections from thisanimal. No hybridization signal was detected in specimens ob-tained from HIV-negative chimpanzees or HIV/NPs (figure 6C).
Immunohistochemistry was used to localize the major viralcapsid protein (p24) in lymph node sections. Immunoreactivityfor HIV p24 was observed only in lymph node sections fromchimpanzees C499 and C463, the animals with highest virusloads. Like viral RNA, virus capsid antigen was localized tocells in germinal centers and paracortex. In addition, a reticularpattern of intercellular immunoreactivity was apparent in ger-minal centers, corroborating the dendritic cell trapping seen byISH (figure 6D). Immunoreactivity for p24 was not observedin lymph node sections from HIV-negative controls, HIV/NPs,or progressors C550 or C560.
Virus isolation from cPBMC. Virus was isolated from thePBMC of each HIV-infected chimpanzee on at least 1 attemptbetween 1993 and 1999; however, substantial differences in the
incidence of virus recovery were observed among animals (table3). Greater variability in the incidence of virus recovery wasobserved among HIV/NPs (11%–100%, with >50% recoveryin 3 of the 6 animals) than among HIV/Ps (56%–82%, with150% recovery in all 4 animals). Overall, the incidence of virusrecovery was greater for HIV/Ps (49 [77%] of 64 attempts) thanfor HIV/NPs (23 [47%] of 49 attempts), suggesting persistentlyhigher cell-associated virus loads in progressors.
Anti-HIV antibody responses. The magnitude of the anti-HIV humoral response was measured by ELISA for each ofthe chronically infected chimpanzees from 1993 through 1999,the results of which are reported as the reciprocal log of thelast positive serum dilution, with the overall median value foreach animal indicated (figure 7). All animals became seropos-itive after inoculation, and all of them exhibited persistent an-tibody responses to HIV antigens. In general, the anti-HIVantibody titers were greater for HIV/Ps than for HIV/NPs(range, 4.58–5.61 vs. 3.60–5.31 log21, respectively; median, 5.16vs. 4.41 log21, respectively; ).P ! .03
The quality of the anti-HIV humoral response was evaluated
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021

JID 2000;182 (October) HIV-1 Progression in Chimpanzees 1057
Figure 6. Virus burden and distribution in lymph nodes from human immunodeficiency virus (HIV)–infected progressor and nonprogressorchimpanzees. In situ hybridization is shown for HIV-1 RNA in progressors C550 (A) and C463 (B) and nonprogressor C542 (C). A, Infectedcells (dark blue) are most numerous within lymph node germinal centers and surrounding paracortical zones of progressor chimpanzees. Nitrobluetetrazolium (NBT)/BCIP (5-bromo-4-chloro-3-indolyl-phosphate) chromagen with nuclear fast red counterstain (2003). B, HIV-1 RNA is alsolocalized to intercellular spaces of lymphoid germinal centers in progressor chimpanzees, creating the diffuse hybridization signal typical ofdendritic cell–trapped virions (apparent here as a pale blue blush overlying the germinal center). NBT/BCIP chromagen with nuclear fast redcounterstain (2003). C, HIV-1 RNA is not found within lymph nodes of nonprogressor chimpanzees. NBT/BCIP chromagen with nuclear fastred counterstain (2003). D, Immunohistochemistry for HIV-1 p24 in lymph node from progressor C499. Viral antigen (brown chromagen) islocalized in a reticular pattern over germinal centers, confirming the dendritic cell trapping observed by in situ hybridization. DAB (3, 3′-diaminobenzidine) chromagen with methyl green counterstain (4003).
by Western blot (data not shown). Serum from each of theHIV-infected chimpanzees recognized all major HIV antigens,including the precursor (gp160), surface (gp120), and trans-membrane (gp41) envelope glycoproteins; the precursor (p55),major capsid (p24), and matrix (p18) gag proteins; and the RT(p65) and endonuclease (p31) proteins. There was no discernibledifference in the quality of the anti-HIV humoral response gen-erated between HIV/Ps and HIV/NPs.
Discussion
In this study, we provide conclusive proof of progressive in-fection in 3 surviving, chronically infected, HIV-1–positive
chimpanzees in the Yerkes cohort. As in humans, CD41 T cellloss in progressor chimpanzees was strongly correlated withplasma and lymphoid virus burden and with increases in cy-tologic and serologic markers of immune activation (summa-rized in table 4) [52–102]. We found the composition of T lym-phocyte subsets and the level of immune activation to beremarkably similar between HIV-positive nonprogressor andHIV-negative control chimpanzees. In contrast, studies of HIVdisease progression in humans have revealed distinct differencesbetween LTNPs and seronegative controls (table 4). This dis-crepancy could reflect fundamental differences in the mecha-nisms used to control HIV infection between an evolutionarily
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021

1058 O’Neil et al. JID 2000;182 (October)
Table 3. Virus isolation from chimpanzee peripheral blood mono-nuclear cells (cPBMC) of chimpanzees persistently infected with humanimmunodeficiency virus (HIV).
Animal
Virus isolation from cPBMC
Coculturesa Recovery, %
ProgressorsC463 23/29 79C499 7/9 78C550 5/9 56C560 14/17 82
Group total 49/64 77Nonprogressors
C459 4/6 67C487 2/9 22C497 1/9 11C542 8/8 100C544 4/9 44C552 4/8 50
Group total 23/49 47
a Ratio of positive cultures to total cultures attempted from 1993 to 1999.
Figure 7. Humoral immune responses to whole, disrupted humanimmunodeficiency virus type 1, as determined by ELISA. Scatter plotshows the range of responses for each individual, reported as the re-ciprocal log of the last positive serum dilution. Horizontal bars indicatethe median value for all measurements for each individual.
adapted host and an outlier population of a nonadapted host.Alternatively, this disparity may be the result of variation inthe parameters used by different investigators to define LTNPcohorts or may be due to heterogeneity in the LTNPpopulation.
Progressor chimpanzees had moderate (13 logs) to high (15logs) levels of virus in plasma, whereas viral RNA was belowthe lower limit of detection in plasma specimens from nonpro-gressor chimpanzees (table 4). These findings are in agreementwith several studies showing a direct association betweenplasma virus load and disease progression in HIV-infected peo-ple [13, 52–56, 58, 59]. Cell-associated virus loads were notmeasured in the present study; however, the frequency of virusrecovery from cPBMC was greater for HIV/Ps than for HIV/NPs. This suggests that progressor chimpanzees are more fre-quently viremic than nonprogressors, which has also been ob-served in studies of progressor versus LTNP humans [60, 62].Virus isolations were performed on bulk, rather than CD8-depleted, cPBMC, however, and one could alternatively inter-pret these results as indicative of higher levels of suppressivefactors in nonprogressor cPBMC cultures. In future studies, wehope to quantitate levels of b-chemokines in uninfected controland HIV-infected chimpanzees to help discriminate betweenthese 2 possibilities.
Lymphoid tissue virus burden in chimpanzees was directlycorrelated with plasma virus load. All lymph nodes examinedfrom progressor chimpanzees contained productively infectedcells within germinal centers and paracortex, similar to the dis-tribution of infected cells in HIV-infected humans with pro-gressive infection [63, 64, 66, 103–108]. Furthermore, the diffusehybridization signal characteristic of follicular dendritic cell(FDC)–trapped, immune-complexed virions [65, 66] was de-tected in lymphoid germinal centers of the 3 chimpanzees withthe highest virus loads. The persistent association of virionswith FDCs in lymphoid germinal centers is thought to be cru-cial to HIV immunopathogenesis, facilitating new infections of
activated CD41 T cells and precipitating dendritic cell loss andsubsequent follicular fragmentation and dissolution [4, 5, 7,109–111] . We found no evidence of HIV-1 RNA in lymphoidtissues from nonprogressor chimpanzees, in agreement with pre-vious studies investigating tissue virus burden during apatho-genic HIV-1 infection in this species [47].
Except for the animal that developed AIDS [39, 71], pro-gressive decreases in CD41 T lymphocytes have not been re-ported in chronically infected HIV-positive chimpanzees [31,35]. In this study we identified 3 additional HIV-1–positivechimpanzees in the Yerkes cohort that have experienced pro-found reductions in both absolute numbers and percentages ofCD41 T lymphocytes, similar to what has been described forhuman progressors [67–70, 72, 73, 112]. Furthermore, as withhuman progressors [67, 73, 76–78], progressor chimpanzeesshowed reductions in both memory (CD291) and naı̈ve(CD45RA1) subsets of CD41 T cells. In addition, the obser-vation of significant increases in memory (CD291), but notnaı̈ve (CD45RA1), subsets of CD81 T cells of progressor chim-panzees is in direct agreement with the CD81 T cell subsetchanges that have been observed to occur during progressiveHIV infection in humans [74, 83].
Persistent infection with HIV results in a chronic state ofimmune activation, which is reflected by increases in the ex-pression of activation markers on immune cells and increasedlevels of various soluble biomolecules in serum. We found in-creased levels of HLA-DR expression on CD41 and CD81 Tcells and increased CD38 expression on CD81 T cells of HIV/P as opposed to HIV/NP and HIV-negative chimpanzees. Assummarized in table 4, each of these changes has also beenassociated with HIV disease progression in humans [67, 81–83,85–87, 89–95]. HIV/Ps had significantly lower percentages ofCD251CD41 T lymphocytes than did HIV/NPs and uninfectedcontrols, an observation that has also been reported in HIV-infected people [87, 88]. Furthermore, HIV/Ps had greater se-
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021
JID 2000;182 (October) HIV-1 Progression in Chimpanzees 1059
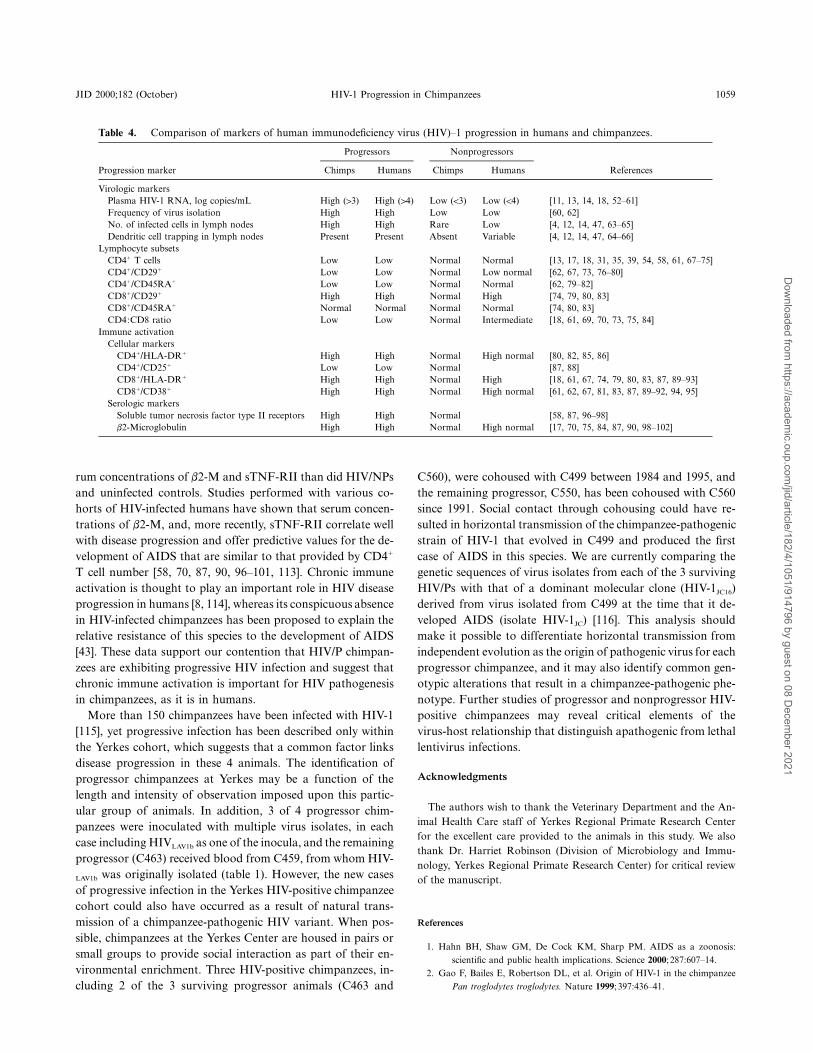
Table 4. Comparison of markers of human immunodeficiency virus (HIV)–1 progression in humans and chimpanzees.
Progressors Nonprogressors
ReferencesProgression marker Chimps Humans Chimps Humans
Virologic markersPlasma HIV-1 RNA, log copies/mL High (13) High (14) Low (!3) Low (!4) [11, 13, 14, 18, 52–61]Frequency of virus isolation High High Low Low [60, 62]No. of infected cells in lymph nodes High High Rare Low [4, 12, 14, 47, 63–65]Dendritic cell trapping in lymph nodes Present Present Absent Variable [4, 12, 14, 47, 64–66]
Lymphocyte subsetsCD41 T cells Low Low Normal Normal [13, 17, 18, 31, 35, 39, 54, 58, 61, 67–75]CD41/CD291 Low Low Normal Low normal [62, 67, 73, 76–80]CD41/CD45RA1 Low Low Normal Normal [62, 79–82]CD81/CD291 High High Normal High [74, 79, 80, 83]CD81/CD45RA1 Normal Normal Normal Normal [74, 80, 83]CD4:CD8 ratio Low Low Normal Intermediate [18, 61, 69, 70, 73, 75, 84]
Immune activationCellular markers
CD41/HLA-DR1 High High Normal High normal [80, 82, 85, 86]CD41/CD251 Low Low Normal [87, 88]CD81/HLA-DR1 High High Normal High [18, 61, 67, 74, 79, 80, 83, 87, 89–93]CD81/CD381 High High Normal High normal [61, 62, 67, 81, 83, 87, 89–92, 94, 95]
Serologic markersSoluble tumor necrosis factor type II receptors High High Normal [58, 87, 96–98]b2-Microglobulin High High Normal High normal [17, 70, 75, 84, 87, 90, 98–102]
rum concentrations of b2-M and sTNF-RII than did HIV/NPsand uninfected controls. Studies performed with various co-horts of HIV-infected humans have shown that serum concen-trations of b2-M, and, more recently, sTNF-RII correlate wellwith disease progression and offer predictive values for the de-velopment of AIDS that are similar to that provided by CD41
T cell number [58, 70, 87, 90, 96–101, 113]. Chronic immuneactivation is thought to play an important role in HIV diseaseprogression in humans [8, 114], whereas its conspicuous absencein HIV-infected chimpanzees has been proposed to explain therelative resistance of this species to the development of AIDS[43]. These data support our contention that HIV/P chimpan-zees are exhibiting progressive HIV infection and suggest thatchronic immune activation is important for HIV pathogenesisin chimpanzees, as it is in humans.
More than 150 chimpanzees have been infected with HIV-1[115], yet progressive infection has been described only withinthe Yerkes cohort, which suggests that a common factor linksdisease progression in these 4 animals. The identification ofprogressor chimpanzees at Yerkes may be a function of thelength and intensity of observation imposed upon this partic-ular group of animals. In addition, 3 of 4 progressor chim-panzees were inoculated with multiple virus isolates, in eachcase including HIVLAV1b as one of the inocula, and the remainingprogressor (C463) received blood from C459, from whom HIV-
LAV1b was originally isolated (table 1). However, the new casesof progressive infection in the Yerkes HIV-positive chimpanzeecohort could also have occurred as a result of natural trans-mission of a chimpanzee-pathogenic HIV variant. When pos-sible, chimpanzees at the Yerkes Center are housed in pairs orsmall groups to provide social interaction as part of their en-vironmental enrichment. Three HIV-positive chimpanzees, in-cluding 2 of the 3 surviving progressor animals (C463 and
C560), were cohoused with C499 between 1984 and 1995, andthe remaining progressor, C550, has been cohoused with C560since 1991. Social contact through cohousing could have re-sulted in horizontal transmission of the chimpanzee-pathogenicstrain of HIV-1 that evolved in C499 and produced the firstcase of AIDS in this species. We are currently comparing thegenetic sequences of virus isolates from each of the 3 survivingHIV/Ps with that of a dominant molecular clone (HIV-1JC16)derived from virus isolated from C499 at the time that it de-veloped AIDS (isolate HIV-1JC) [116]. This analysis shouldmake it possible to differentiate horizontal transmission fromindependent evolution as the origin of pathogenic virus for eachprogressor chimpanzee, and it may also identify common gen-otypic alterations that result in a chimpanzee-pathogenic phe-notype. Further studies of progressor and nonprogressor HIV-positive chimpanzees may reveal critical elements of thevirus-host relationship that distinguish apathogenic from lethallentivirus infections.
Acknowledgments
The authors wish to thank the Veterinary Department and the An-imal Health Care staff of Yerkes Regional Primate Research Centerfor the excellent care provided to the animals in this study. We alsothank Dr. Harriet Robinson (Division of Microbiology and Immu-nology, Yerkes Regional Primate Research Center) for critical reviewof the manuscript.
References
1. Hahn BH, Shaw GM, De Cock KM, Sharp PM. AIDS as a zoonosis:scientific and public health implications. Science 2000;287:607–14.
2. Gao F, Bailes E, Robertson DL, et al. Origin of HIV-1 in the chimpanzeePan troglodytes troglodytes. Nature 1999;397:436–41.
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021

1060 O’Neil et al. JID 2000;182 (October)
3. Corbet S, Muller-Trutwin MC, Versmisse P, et al. env sequences of simianimmunodeficiency viruses from chimpanzees in Cameroon are stronglyrelated to those of human immunodeficiency virus group N from thesame geographic area. J Virol 2000;74:529–34.
4. Fauci AS. Multifactorial nature of human immunodeficiency virus disease:implications for therapy. Science 1993;262:1011–8.
5. Fauci AS. Host factors and the pathogenesis of HIV-induced disease. Nature1996;384:529–34.
6. Haynes BF, Pantaleo G, Fauci AS. Toward an understanding of the cor-relates of protective immunity to HIV infection. Science 1996;271:324–8.
7. Pantaleo G, Graziosi C, Fauci AS. The immunopathogenesis of humanimmunodeficiency virus infection. N Engl J Med 1993;328:327–35.
8. Pantaleo G, Fauci AS. New concepts in the immunopathogenesis of HIVinfection. Annu Rev Immunol 1995;13:487–512.
9. Perelson AS, Neumann AU, Markowitz M, Leonard JM, Ho DD. HIV-1dynamics in vivo: virion clearance rate, infected cell life-span, and viralgeneration time. Science 1996;271:1582–6.
10. Wei X, Ghosh SK, Taylor ME, et al. Viral dynamics in human immuno-deficiency virus type 1 infection. Nature 1995;373:117–22.
11. Cao Y, Qin L, Zhang L, Safrit J, Ho DD. Virologic and immunologiccharacterization of long-term survivors of human immunodeficiency vi-rus type 1 infection. N Engl J Med 1995;332:201–8.
12. Fauci AS, Pantaleo G, Stanley S, Weissman D. Immunopathogenic mech-anisms of HIV infection. Ann Intern Med 1996;124:654–63.
13. Hogervorst E, Jurriaans S, de Wolf F, et al. Predictors for non- and slowprogression in human immunodeficiency virus (HIV) type 1 infection:low viral RNA copy numbers in serum and maintenance of high HIV-1 p24–specific but not V3-specific antibody levels. J Infect Dis 1995;171:811–21.
14. Pantaleo G, Menzo S, Vaccarezza M, et al. Studies in subjects with long-term nonprogressive human immunodeficiency virus infection. N EnglJ Med 1995;332:209–16.
15. Pontesilli O, Klein MR, Kerkhof-Garde SR, et al. Kinetics of immunefunctions and virus replication during HIV-1 infection. Immunol Lett1997;57:125–30.
16. Rinaldo C, Huang XL, Fan ZF, et al. High levels of anti–human immu-
nodeficiency virus type 1 (HIV-1) memory cytotoxic T-lymphocyte ac-
tivity and low viral load are associated with lack of disease in HIV-
1–infected long-term nonprogressors. J Virol 1995;69:5838–42.
17. Sheppard HW, Lang W, Ascher MS, Vittinghoff E, Winkelstein W. The
characterization of non-progressors: long-term HIV-1 infection with sta-
ble CD41 T-cell levels. AIDS 1993;7:1159–66.
18. Greenough TC, Brettler DB, Kirchhoff F, et al. Long-term nonprogressive
infection with human immunodeficiency virus type 1 in a hemophilia
cohort. J Infect Dis 1999;180:1790–802.
19. Harrer T, Harrer E, Kalams SA, et al. Cytotoxic T lymphocytes in asymp-
tomatic long-term nonprogressing HIV-1 infection: breadth and speci-
ficity of the response and relation to in vivo viral quasispecies in a person
with prolonged infection and low viral load. J Immunol 1996;156:
2616–23.
20. Rosenberg ES, Billingsley JM, Caliendo AM, et al. Vigorous HIV-1–specific
CD41 T cell responses associated with control of viremia. Science
1997;278:1447–50.
21. Peeters M, Honore C, Huet T, et al. Isolation and partial characterization
of an HIV-related virus occurring naturally in chimpanzees in Gabon.
AIDS 1989;3:625–30.
22. Peeters M, Fransen K, Delaporte E, et al. Isolation and characterization
of a new chimpanzee lentivirus (simian immunodeficiency virus isolate
cpz-ant) from a wild-captured chimpanzee. AIDS 1992;6:447–51.
23. Kestens L, Vingerhoets J, Peeters M, et al. Phenotypic and functional pa-
rameters of cellular immunity in a chimpanzee with a naturally acquired
simian immunodeficiency virus infection. J Infect Dis 1995;172:957–63.
24. Vanden Haesevelde MM, Peeters M, Jannes G, et al. Sequence analysis of
a highly divergent HIV-1–related lentivirus isolated from a wild capturedchimpanzee. Virology 1996;221:346–50.
25. Alter HJ, Eichberg JW, Masur H, et al. Transmission of HTLV-III infectionfrom human plasma to chimpanzees: an animal model for AIDS. Science1984;226:549–52.
26. Francis DP, Feorino PM, Broderson JR, et al. Infection of chimpanzeeswith lymphadenopathy-associated virus. Lancet 1984;2:1276–7.
27. Gajdusek DC, Amyx HL, Gibbs CJ Jr, et al. Infection of chimpanzees byhuman T-lymphotropic retroviruses in brain and other tissues from AIDSpatients. Lancet 1985;1:55–6.
28. Balla-Jhagjhoorsingh SS, Koopman G, Mooij P, et al. Conserved CTLepitopes shared between HIV-infected human long-term survivors andchimpanzees. J Immunol 1999;162:2308–14.
29. Bogers WM, Koornstra WH, Dubbes RH, et al. Characteristics of primaryinfection of a European human immunodeficiency virus type 1 clade Bisolate in chimpanzees. J Gen Virol 1998;79:2895–903.
30. Fultz PN, McClure HM, Swenson RB, et al. Persistent infection of chim-panzees with human T-lymphotropic virus type III/lymphadenopathy-associated virus: a potential model for acquired immunodeficiency syn-drome. J Virol 1986;58:116–24.
31. Fultz PN. Nonhuman primate models for AIDS. Clin Infect Dis 1993;17(Suppl 1):S230–5.
32. Fultz PN, Wei Q, Yue L. Rectal transmission of human immunodeficiencyvirus type 1 to chimpanzees. J Infect Dis 1999;179(Suppl 3):S418–21.
33. Goudsmit J, Smit L, Krone WJ, et al. IgG response to human immuno-deficiency virus in experimentally infected chimpanzees mimics the IgGresponse in humans. J Infect Dis 1987;155:327–31.
34. Heeney J, Jonker R, Koornstra W, et al. The resistance of HIV-infectedchimpanzees to progression to AIDS correlates with absence of HIV-related T-cell dysfunction. J Med Primatol 1993;22:194–200.
35. Johnson BK, Stone GA, Godec MS, et al. Long-term observations of humanimmunodeficiency virus–infected chimpanzees. AIDS Res Hum Retro-viruses 1993;9:375–8 (erratum: AIDS Res Hum Retroviruses 1993;9:579).
36. Santra S, Fultz PN, Letvin NL. Virus-specific cytotoxic T lymphocytes in
human immunodeficiency virus type 1–infected chimpanzees. J Virol
1999;73:7065–9.
37. Shibata R, Seimon C, Cho MW, et al. Resistance of previously infected
chimpanzees to successive challenges with a heterologous intraclade B
strain of human immunodeficiency virus type 1. J Virol 1996;70:4361–9.
38. Stephens DM, Eichberg JW, Haigwood NL, et al. Antibodies are produced
to the variable regions of the external envelope glycoprotein of human
immunodeficiency virus type 1 in chimpanzees infected with the virus
and baboons immunized with a candidate recombinant vaccine. J Gen
Virol 1992;73:1099–106.
39. Novembre FJ, Saucier M, Anderson DC, et al. Development of AIDS in
a chimpanzee infected with human immunodeficiency virus type 1. J
Virol 1997;71:4086–91.
40. Davis IC, Girard M, Fultz PN. Loss of CD41 T cells in human immu-
nodeficiency virus type 1–infected chimpanzees is associated with in-
creased lymphocyte apoptosis. J Virol 1998;72:4623–32.
41. Estaquier J, Idziorek T, de Bels F, et al. Programmed cell death and AIDS:
significance of T-cell apoptosis in pathogenic and nonpathogenic primate
lentiviral infections. Proc Natl Acad Sci U S A 1994;91:9431–5.
42. Gougeon ML, Garcia S, Heeney J, et al. Programmed cell death in AIDS-
related HIV and SIV infections. AIDS Res Hum Retroviruses 1993;9:
553–63.
43. Gougeon ML, Lecoeur H, Boudet F, et al. Lack of chronic immune acti-
vation in HIV-infected chimpanzees correlates with the resistance of T
cells to Fas/Apo-1 (CD95)–induced apoptosis and preservation of a T
helper 1 phenotype. J Immunol 1997;158:2964–76.
44. Schuitemaker H, Meyaard L, Kootstra NA, et al. Lack of T cell dysfunction
and programmed cell death in human immunodeficiency virus type
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021

JID 2000;182 (October) HIV-1 Progression in Chimpanzees 1061
1–infected chimpanzees correlates with absence of monocytotropic var-iants. J Infect Dis 1993;168:1140–7.
45. Heeney JL. AIDS: a disease of impaired Th-cell renewal? Immunol Today1995;16:515–20.
46. Zarling JM, Ledbetter JA, Sias J, et al. HIV-infected humans, but not chim-panzees, have circulating cytotoxic T lymphocytes that lyse uninfectedCD41 cells. J Immunol 1990;144:2992–8.
47. Koopman G, Haaksma AG, ten Velden J, Hack CE, Heeney JL. The relativeresistance of HIV type 1–infected chimpanzees to AIDS correlates withthe maintenance of follicular architecture and the absence of infiltrationby CD81 cytotoxic T lymphocytes. AIDS Res Hum Retroviruses1999;15:365–73.
48. Guenthner PC, Hart CE. Quantitative, competitive PCR assay for HIV-1using a microplate-based detection system. Biotechniques1998;24:810–6.
49. O’Neil SP, Mossman SP, Maul DH, Hoover EA. In vivo cell and tissuetropism of SIVsmmPBj14-bcl.3. AIDS Res Hum Retroviruses 1999;15:203–15.
50. Fultz PN, Srinivasan A, Greene CR, et al. Superinfection of a chimpanzeewith a second strain of human immunodeficiency virus. J Virol 1987;61:4026–9.
51. Hu SL, Fultz PN, McClure HM, et al. Effect of immunization with avaccinia–HIV env recombinant on HIV infection of chimpanzees. Nature1987;328:721–3.
52. Mellors JW, Kingsley LA, Rinaldo CR, Jr., et al. Quantitation of HIV-1RNA in plasma predicts outcome after seroconversion. Ann Intern Med1995;122:573–9.
53. Mellors JW, Rinaldo CR Jr, Gupta P, et al. Prognosis in HIV-1 infectionpredicted by the quantity of virus in plasma. Science 1996;272:1167–70(erratum: Science 1997;275:14).
54. de Wolf F, Spijkerman I, Schellekens PT, et al. AIDS prognosis based onHIV-1 RNA, CD41 T-cell count and function: markers with reciprocalpredictive value over time after seroconversion. AIDS 1997;11:1799–806.
55. Galetto-Lacour A, Yerly S, Perneger TV, et al. Prognostic value of viremiain patients with long-standing human immunodeficiency virus infection:Swiss HIV Cohort Study Group. J Infect Dis 1996;173:1388–93.
56. Mofenson LM, Korelitz J, Meyer WA III, et al. The relationship betweenserum human immunodeficiency virus type 1 (HIV-1) RNA level, CD4lymphocyte percent, and long-term mortality risk in HIV-1–infected chil-dren: National Institute of Child Health and Human Development In-travenous Immunoglobulin Clinical Trial Study Group. J Infect Dis1997;175:1029–38.
57. Spijkerman IJ, Prins M, Goudsmit J, et al. Early and late HIV-1 RNA leveland its association with other markers and disease progression in long-term AIDS-free homosexual men. AIDS 1997;11:1383–8.
58. Fahey JL, Taylor JM, Manna B, et al. Prognostic significance of plasmamarkers of immune activation, HIV viral load and CD4 T-cell mea-surements. AIDS 1998;12:1581–90.
59. Iuliano R, Forastieri G, Brizzi M, et al. Correlation between plasma HIV-1 RNA levels and the rate of immunologic decline. J Acquir ImmuneDefic Syndr Hum Retrovirol 1997;14:408–14.
60. Candotti D, Costagliola D, Joberty C, et al. Status of long-term asymp-tomatic HIV-1 infection correlates with viral load but not with virusreplication properties and cell tropism: French ALT Study Group. J MedVirol 1999;58:256–63.
61. Ferbas J, Kaplan AH, Hausner MA, et al. Virus burden in long-term sur-vivors of human immunodeficiency virus (HIV) infection is a determi-nant of anti-HIV CD81 lymphocyte activity. J Infect Dis 1995;172:329–39.
62. Clerici M, Balotta C, Meroni L, et al. Type 1 cytokine production and lowprevalence of viral isolation correlate with long-term nonprogression inHIV infection. AIDS Res Hum Retroviruses 1996;12:1053–61.
63. Pantaleo G, Graziosi C, Butini L, et al. Lymphoid organs function as majorreservoirs for human immunodeficiency virus. Proc Natl Acad Sci U SA 1991;88:9838–42.
64. Pantaleo G, Graziosi C, Demarest JF, et al. HIV infection is active andprogressive in lymphoid tissue during the clinically latent stage of disease.Nature 1993;362:355–8.
65. Haase AT, Henry K, Zupancic M, et al. Quantitative image analysis ofHIV-1 infection in lymphoid tissue. Science 1996;274:985–9.
66. Fox CH, Tenner-Racz K, Racz P, et al. Lymphoid germinal centers arereservoirs of human immunodeficiency virus type 1 RNA. J Infect Dis1991;164:1051–7 (erratum: J Infect Dis 1992;165:1161).
67. Giorgi JV, Detels R. T-cell subset alterations in HIV-infected homosexualmen: NIAID Multicenter AIDS cohort study. Clin Immunol Immuno-pathol 1989;52:10–8.
68. Polk BF, Fox R, Brookmeyer R, et al. Predictors of the acquired immu-nodeficiency syndrome developing in a cohort of seropositive homosex-ual men. N Engl J Med 1987;316:61–6.
69. Taylor JM, Fahey JL, Detels R, Giorgi JV. CD4 percentage, CD4 number,and CD4:CD8 ratio in HIV infection: which to choose and how to use.J Acquir Immune Defic Syndr 1989;2:114–24.
70. Fahey JL, Taylor JM, Detels R, et al. The prognostic value of cellular andserologic markers in infection with human immunodeficiency virus type1. N Engl J Med 1990;322:166–72.
71. Fultz PN, Siegel RL, Brodie A, et al. Prolonged CD41 lymphocytopeniaand thrombocytopenia in a chimpanzee persistently infected with humanimmunodeficiency virus type 1. J Infect Dis 1991;163:441–7.
72. Lang W, Perkins H, Anderson RE, et al. Patterns of T lymphocyte changeswith human immunodeficiency virus infection: from seroconversion tothe development of AIDS. J Acquir Immune Defic Syndr 1989;2:63–9.
73. Nicholson JK, Spira TJ, Aloisio CH, et al. Serial determinations of HIV-1titers in HIV-infected homosexual men: association of rising titers withCD4 T cell depletion and progression to AIDS. AIDS Res Hum Retro-viruses 1989;5:205–15.
74. Watret KC, Whitelaw JA, Froebel KS, Bird AG. Phenotypic characteri-zation of CD81 T cell populations in HIV disease and in anti-HIVimmunity. Clin Exp Immunol 1993;92:93–9.
75. Buchbinder SP, Katz MH, Hessol NA, O’Malley PM, Holmberg SD. Long-term HIV-1 infection without immunologic progression. AIDS 1994;8:1123–8.
76. Gupta S. Subpopulations of CD41 (T41) cells in homosexual/bisexual menwith persistent generalized lymphadenopathy. Clin Exp Immunol1987;68:1–4.
77. Nicholson JK, Jones BM. Lymphocyte immunophenotyping at the Centersfor Disease Control: the program and special studies. Clin ImmunolImmunopathol 1989;52:61–7.
78. Vuillier F, Lapresle C, Dighiero G. Comparative analysis of CD4-4B4 andCD4-2H4 lymphocyte subpopulations in HIV negative homosexual, HIVseropositive and healthy subjects. Clin Exp Immunol 1988;71:8–12.
79. Brostrom C, Sonnerborg A, Lindback S, Gaines H. Low relative frequenciesof CD26(1) CD4(1) cells in long-term nonprogressing human immu-nodeficiency virus type 1–infected subjects. Clin Diagn Lab Immunol1998;5:662–6.
80. Ginaldi L, De Martinis M, D’Ostilio A, et al. Activated naive and memoryCD41 and CD81 subsets in different stages of HIV infection. Patho-biology 1997;65:91–9.
81. Giorgi JV, Hultin LE, McKeating JA, et al. Shorter survival in advancedhuman immunodeficiency virus type 1 infection is more closely associatedwith T lymphocyte activation than with plasma virus burden or viruschemokine coreceptor usage. J Infect Dis 1999;179:859–70.
82. Peakman M, Mahalingam M, Pozniak A, et al. Markers of immune cellactivation and disease progression: cell activation in HIV disease. AdvExp Med Biol 1995;374:17–26.
83. Prince HE, Jensen ER. Three-color cytofluorometric analysis of CD8 cellsubsets in HIV-1 infection. J Acquir Immune Defic Syndr 1991;4:1227–32.
84. Lifson AR, Buchbinder SP, Sheppard HW, et al. Long-term human im-munodeficiency virus infection in asymptomatic homosexual and bisex-
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021

1062 O’Neil et al. JID 2000;182 (October)
ual men with normal CD41 lymphocyte counts: immunologic and vi-rologic characteristics. J Infect Dis 1991;163:959–65.
85. Kestens L, Vanham G, Vereecken C, et al. Selective increase of activationantigens HLA-DR and CD38 on CD41 CD45RO1 T lymphocytesduring HIV-1 infection. Clin Exp Immunol 1994;95:436–41.
86. Mahalingam M, Peakman M, Davies ET, et al. T cell activation and diseaseseverity in HIV infection. Clin Exp Immunol 1993;93:337–43.
87. Plaeger S, Bass HZ, Nishanian P, et al. The prognostic significance in HIVinfection of immune activation represented by cell surface antigen andplasma activation marker changes. Clin Immunol 1999;90:238–46.
88. Hofmann B, Nishanian P, Fahey JL, et al. Serum increases and lymphoidcell surface losses of IL-2 receptor CD25 in HIV infection: distinctiveparameters of HIV-induced change. Clin Immunol Immunopathol1991;61:212–24.
89. Levacher M, Hulstaert F, Tallet S, et al. The significance of activationmarkers on CD8 lymphocytes in human immunodeficiency syndrome:staging and prognostic value. Clin Exp Immunol 1992;90:376–82.
90. Bass HZ, Nishanian P, Hardy WD, et al. Immune changes in HIV-1 infec-tion: significant correlations and differences in serum markers and lym-phoid phenotypic antigens. Clin Immunol Immunopathol 1992;64:63–70.
91. Giorgi JV, Liu Z, Hultin LE, et al. Elevated levels of CD381 CD81 T cellsin HIV infection add to the prognostic value of low CD41 T cell levels:results of 6 years of follow-up. The Los Angeles Center, MulticenterAIDS Cohort Study. J Acquir Immune Defic Syndr 1993;6:904–12.
92. Kestens L, Vanham G, Gigase P, et al. Expression of activation antigens,HLA-DR and CD38, on CD8 lymphocytes during HIV-1 infection.
AIDS 1992;6:793–7.
93. Stites DP, Moss AR, Bacchetti P, et al. Lymphocyte subset analysis to predict
progression to AIDS in a cohort of homosexual men in San Francisco.
Clin Immunol Immunopathol 1989;52:96–103.
94. Liu Z, Cumberland WG, Hultin LE, et al. Elevated CD38 antigen expression
on CD81 T cells is a stronger marker for the risk of chronic HIV disease
progression to AIDS and death in the Multicenter AIDS Cohort Study
than CD41 cell count, soluble immune activation markers, or combi-
nations of HLA-DR and CD38 expression. J Acquir Immune Defic
Syndr Hum Retrovirol 1997;16:83–92.
95. Yagi MJ, Chu FN, Jiang JD, et al. Increases in soluble CD8 antigen in
plasma, and CD81 and CD81CD381 cells in human immunodeficiency
virus type-1 infection. Clin Immunol Immunopathol 1992;63:126–34.
96. Stein DS, Lyles RH, Graham NM, et al. Predicting clinical progression or
death in subjects with early-stage human immunodeficiency virus (HIV)
infection: a comparative analysis of quantification of HIV RNA, soluble
tumor necrosis factor type II receptors, neopterin, and beta2-microg-
lobulin. Multicenter AIDS Cohort Study. J Infect Dis 1997;176:1161–7.
97. Godfried MH, van der Poll T, Jansen J, et al. Soluble receptors for tumour
necrosis factor: a putative marker of disease progression in HIV infection.
AIDS 1993;7:33–6.
98. Godfried MH, van der Poll T, Weverling GJ, et al. Soluble receptors for
tumor necrosis factor as predictors of progression to AIDS in asymp-
tomatic human immunodeficiency virus type 1 infection. J Infect Dis
1994;169:739–45.
99. Hofmann B, Wang YX, Cumberland WG, et al. Serum beta 2-microglobulin
level increases in HIV infection: relation to seroconversion, CD4 T-cellfall and prognosis. AIDS 1990;4:207–14.
100. Schwartlander B, Bek B, Skarabis H, et al. Improvement of the predictivevalue of CD41 lymphocyte count by beta 2- microglobulin, immuno-globulin A and erythrocyte sedimentation rate: The Multicentre CohortStudy Group. AIDS 1993;7:813–21.
101. Prince HE, Kleinman S, Czaplicki C, John J, Williams AE. Interrelation-ships between serologic markers of immune activation and T lymphocytesubsets in HIV infection. J Acquir Immune Defic Syndr 1990;3:525–30.
102. Ashton LJ, Carr A, Cunningham PH, et al. Predictors of progression inlong-term nonprogressors: Australian Long-Term Nonprogressor StudyGroup. AIDS Res Hum Retroviruses 1998;14:117–21.
103. Baroni CD, Pezzella F, Mirolo M, Ruco LP, Rossi GB. Immunohistochem-ical demonstration of p24 HTLV III major core protein in different celltypes within lymph nodes from patients with lymphadenopathy syn-drome (LAS). Histopathology 1986;10:5–13.
104. Biberfeld P, Chayt KJ, Marselle LM, et al. HTLV-III expression in infectedlymph nodes and relevance to pathogenesis of lymphadenopathy. Am JPathol 1986;125:436–42.
105. Embretson J, Zupancic M, Ribas JL, et al. Massive covert infection of helperT lymphocytes and macrophages by HIV during the incubation periodof AIDS. Nature 1993;362:359–62.
106. Schuurman HJ, Krone WJ, Broekhuizen R, Goudsmit J. Expression of RNAand antigens of human immunodeficiency virus type-1 (HIV-1) in lymphnodes from HIV-1 infected individuals. Am J Pathol 1988;133:516–24.
107. Tenner-Racz K, Racz P, Bofill M, et al. HTLV-III/LAV viral antigens inlymph nodes of homosexual men with persistent generalized lymphad-enopathy and AIDS. Am J Pathol 1986;123:9–15.
108. Ward JM, O’Leary TJ, Baskin GB, et al. Immunohistochemical localizationof human and simian immunodeficiency viral antigens in fixed tissuesections. Am J Pathol 1987;127:199–205.
109. Piris MA, Rivas C, Morente M, et al. Persistent and generalized lymphad-enopathy: a lesion of follicular dendritic cells? an immunohistologic andultrastructural study. Am J Clin Pathol 1987;87:716–24.
110. Schuurman HJ, Joling P, van Wichen DF, et al. Follicular dendritic cellsand infection by human immunodeficiency virus type 1: a crucial targetcell and virus reservoir. Curr Top Microbiol Immunol 1995;201:161–88.
111. Zhang ZQ, Schuler T, Cavert W, et al. Reversibility of the pathologicalchanges in the follicular dendritic cell network with treatment of HIV-1 infection. Proc Natl Acad Sci U S A 1999;96:5169–72.
112. Stein DS, Korvick JA, Vermund SH. CD41 lymphocyte cell enumerationfor prediction of clinical course of human immunodeficiency virus dis-ease: a review. J Infect Dis 1992;165:352–63.
113. Moss AR, Bacchetti P. Natural history of HIV infection. AIDS 1989;3:55–61 (erratum: AIDS 1989;3:following A100).
114. Copeland KF, Heeney JL. T helper cell activation and human retroviralpathogenesis. Microbiol Rev 1996;60:722–42.
115. Chimpanzees in research: strategies for their ethical care, management, anduse. National Research Council report. Washington, DC: NationalResearch Council, 1997 (www.nap.edu/readingroom/books/chimp/execsum.htm).
116. Mwaengo DM, Novembre FJ. Molecular cloning and characterization ofviruses isolated from chimpanzees with pathogenic human immunode-ficiency virus type 1 infections. J Virol 1998;72:8976–87.
Dow
nloaded from https://academ
ic.oup.com/jid/article/182/4/1051/914796 by guest on 08 D
ecember 2021