production of auxotrophic mutants of escherichiaaem.asm.org/content/9/5/419.full.pdf · production...
TRANSCRIPT

Production of L-Threonine by Auxotrophic Mutants ofEscherichia coli
H. T. HUANG
Biochemical Research Laboratories, Chas. Pfizer and Company, Inc., Groton, Connecticut
Received for publication January 6, 1961
ABSTRACT
HUANG, H. T. (Chas. Pfizer and Company, Inc.,Groton, Conn.). Production of L-threonine by auxo-trophic mutants of Escherichia coli. Appl. Microbiol.9:419-424. 1961.-Two auxotrophic mutants ofEscherichia coli have been shown to accumulate signifi-cant amounts of L-threonine in the culture medium.One mutant, 13071, is deficient in a,,E-diaminopimelicacid (DAP), and the other, 13070, is deficient in bothDAP and methionine.Accumulation of L-threonine by 13071 in a synthetic
medium with mannitol as the carbon source at 37 Cincubation temperature was greatly influenced byinitial DAP concentration. At an optimal level of DAPa yield of 1.5 g/liter L-threonine was obtained in 28 hr.L-Threonine accumulation by 13070 with sorbitol as thecarbon source at 28 C incubation temperature wasdependent upon the initial concentration of both DAPand methionine. At optimal levels of the two deficientamino acids, a yield of 2.0 g/liter L-threonine was ob-tained in 44 to 48 hr. By adding to the medium suit-able quantities of beet molasses or sucrose supplementedwith corn steep liquor the yield could be raised to al-most 4 g/liter.
L-Threonine of extremely high purity was isolatedfrom fermentation broth from both mutants. Theproduct was not contaminated with any detectableamounts of D-threonine or allothreonine.
The microbial production of amino acids has at-tracted considerable attention in the fermentationindustry in recent years (Kinoshita, 1959). The chiefadvantage inherent in a microbial process, in contrastto chemical syntheses, is the exclusive production of thenutritionally valuable L form of the amino acid.The most noteworthy successes scored in this field
have been achieved by the use of auxotrophic mutantsof bacteria. The processes investigated can be dividedinto two categories. In the first category, the productaccumulated is an intermediate which occurs justbefore the point of genetic block on the relevant bio-synthetic pathway. For example, a,,E-diaminopimelicacid (DAP) is accumulated by a lysineless Escherichiacoli mutant (Casida, 1956), and ornithine is accumu-
lated by a citrullineless Micrococcus (Kinoshita, Naka-yama, and Udaka, 1957). In the second category thecompound accumulated and the point of genetic blockoccur on separate biosynthetic pathways which are,nevertheless, interrelated in some subtle way. Thus, abiotin-deficient Micrococcus glutamicus accumulatesL-glutamic acid (Kinoshita, Udaka, and Shimono,1957), and a homoserine-deficient mutant of the aboveorganism is an excellent producer of lysine (Kinoshita,Nakayama, and Kitada, 1958).The present paper deals with the production of
L-threonine by DAP- and methionine-requiring mutantsof E. coli. It may provide yet another example in thesecond category of amino acid processes based on theuse of auxotrophic bacterial mutants. A preliminaryaccount of this work has appeared elsewhere (Huang,1960).
MATERIALS AND METHODS
DAP and L-lysine hydrochloride are products ofChas. Pfizer and Company, Inc. (Casida, 1956; Kitaand Huang, 1958). The particular lot of DAP used inthese experiments contains approximately 92 % of themeso and 8 % of the LL isomers by paper chromatog-raphy assay (Rhuland et al., 1955). L-Methionine is anoptically standardized sample from Schwartz Labora-tories, Inc. DL-Methionine and other materials are com-mercial products. "Casamino acids" is an acid hydroly-zate of casein made by Difco Laboratories.
Auxotrophic mutants of E. coli tested in the prelim-inary screening work are cultures which had previ-ously been isolated either by us or by others via ultra-violet (UV) ray treatment and penicillin selection(Davis, 1948, 1949; Lederberg and Zinder, 1948).Mutant D was obtained from Elizabeth Work. It wasoriginally derived from mutant 173-25 (Davis, 1952).
Screening experiments were carried out in 300-mlErlenmeyer flasks containing 25 ml medium. Afterinoculation from slants the flasks were incubated at 28or 37 C on a rotary shaker moving at 230 cycles/min.Single auxotrophs were tested in medium I (Table 1)supplemented by 20 mg/liter DAP and 0.5 g/literCasamino acids. For double auxotrophs, medium II,with 40 mg/liter DAP and 0.5 g/liter Casamino acids,was used. All media were sterilized at 120 C for 20 min.
419
on June 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

H. T. HUANG
A mutant was tested with at least two different carbonsources, glucose and sorbitol, at both incubating tem-peratures. Samples were taken at 20 and 44 hr of incu-bation.
Threonine was detected by paper chromatography.For rapid preliminary identification, 5-,ul aliquots ofbroths were applied on two sheets of Whatman no. 4paper, 13 by 20 cm. One sheet was developed by adescending front of methylethyl ketone, acetic acid,and water (90:25:30 by volume), the other by methyl-ethyl ketone, n-butanol, and 3 % aqueous NH40H(50:20:20 by volume). After 30 min, the sheets weredried in air and treated with ninhydrin in the usualmanner. In the acid system threonine (RF = 0.54)moves close to glutamic acid (0.51), glycine (0.47) andserine (0.46), but separates readily from valine (0.83).On the other hand, in the ammonia system threonine(RF = 0.83) is easily separated from glutamic acid(0.02), glycine (0.14), serine (0.26), although less easilyfrom valine (0.60). Thus, a comparison of the twochromatograms would immediately indicate whetherthreonine might be present in a sample. Confirmatoryevidence was then sought by more detailed paperchromatography.Inoculum for fermentation experiments was grown
in a Fernbach flask containing 1 liter of medium. For13071, the medium consisted of (per liter): minimalsalts (Davis and Mingioli, 1950); DAP, 20 mg; L-lysinehydrochloride, 20 mg; and glucose, 10 g. With 13070,the medium was (per liter): minimal salts; DAP, 50 mg,DL-methionine, 50 mg; and glycerol, 10 g. The flaskswere sterilized at 120 C for 20 min. After inoculation,they were incubated on the rotary shaker at 28 C for16 to 20 hr.
Fernentations were conducted with 2 liters ofmedium in a 4-liter glass fermentor (Shull and Kita,1955). Two milliliters of soybean oil were added to eachfermentor, which was then sterilized at 120 C for 30min. After inoculation with 2.5 to 5.0% inoculum byvolume, the fermentors were incubated in a water bathat the desired temperature, aerated at 1 volume of airper min, and stirred at 1,750 rev/min. The basal media,i.e., II for 13071 and III for 13070, are given in Table 1.
TABLE 1. Summary of media ingredients
MediumMaterial*
I II III
glliter glliter gllierK2HP04 7.0 7.0 5.0(NH4)2 HP04 3.0 13.0 10.0(NH4)2 S04 1.5 1.5MgS04 7H20 0.1 0.1 0.5Carbon source 10.0 20.0 20.0
* Supplements of amino acids are given in appropriate parts
Total threonine was determined by quantitativepaper chromatography, and L-threonine by micro-biological assay with Leuconostoc mesenteroides P60.
RESULTSPreliminary Experiments
Screening experiments were carried out to study thepattern of amino acid accumulation in the culturemedium of a series of auxotrophic E. coli mutants,blocked at various points in the biosynthetic pathway oftryptophan, tyrosine, isoleucine, valine, threonine,methionine, lysine, DAP, histidine, and arginine. Mostof these accumulated glutamic acid, alanine, glycine,and one or two additional amino acids at a level of 50
to 100 mg/liter. Some of these accumulated largerquantities (i.e., 1 g/liter or more) of one particularamino acid; for example, a tyrosine-less mutant accu-
mulated phenylalanine and a threonineless mutantaccumulated valine. Only a DAP deficient mutant, D(Meadow, Hoare, and Work, 1957), produced a de-tectable spot which moved like threonine on paper
chromatograms.Mutant D was screened further in basal medium I
supplemented with DAP (20 mg/liter) and L-lysinehydrochloride (20 mg/liter), using a variety of carbonsources, viz: glucose, lactose, sucrose, maltose, glycerol,sorbitol, and mannitol. Presence of threonine at 100 to300 mg/liter was clearly indicated in glucose and manni-tol broths, which had been incubated at 37 C, or insorbitol or mannitol broths, incubated at 27 C. Glucoseat 27 C or sorbitol at 37 C produced practically no
threonine in the medium. The best broths were assayedmicrobiologically, and the results suggested that allthe threonine accumulated was the L isomer. Underthese conditions, glutamic acid, serine, glycine, alanine,and valine were also accumulated, although the sum ofthese amino acids did not exceed 200 to 300 mg/liter.Therefore, attempts were made to develop a more
desirable producer from mutant D by further mutationand selection.
Development of Mutants 13070 and 13071
Substrains of mutant D were obtained by treating a
young liquid culture with UV light, plating out sur-
vivors on a medium containing minimal salts, glucose,DAP, and L-lysine, and isolating colonies at random.They were screened in flasks as described earlier forL-threonine accumulation. One isolate, 13071, whichproduced less contaminating amino acids than theparent strain, was selected for further mutation, andfor fermentation studies.
In the mutation experiments, 13071 was exposed toUV light, and mutants with new nutritional deficienceswere selected via the penicillin technique (Davis, 1949)from a medium containing minimal salts, glycerol, DAP,and acid-hydrolyzed casein. A series of doubly auxo-
trophic mutants which required DAP and one otherof the text. Unless stated otherwise, the pH of the medium isadjusted to 7.8 to 8.0 with NH40H before sterilization.
420 [VOL. 9
on June 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
L-THREONINE PRODUCTION BY E. COLI
amino acid for growth were obtained. Special effortswere made, without success, to isolate a mutant inwhich the second nutritional deficiency was L-isoleucine.All the double auxotrophs were screened for L-threonineproduction. One mutant, 13070, was found to givehigher yields of L-threonine than 13071 when sorbitolwas the carbon source. It required, in addition to DAP,L-methionine for growth.'No growth was obtained whenL-methionine was replaced by either homocysteine or
cystathionine. The new genetic block must, therefore,occur between homocysteine and methionine.
L-Threonine Production by 13071
L-Threonine production by this mutant was investi-gated using basal medium II in 4-liter fermentors underthe conditions specified earlier. Of the combinations ofcarbon source and incubation temperature which hadbeen found in flask experiments to favor L-threonineaccumulation, mannitol at 37 C gave the best results.Next most effective was mannitol or sorbitol at 28 C,and lastly, glucose at 37 C.
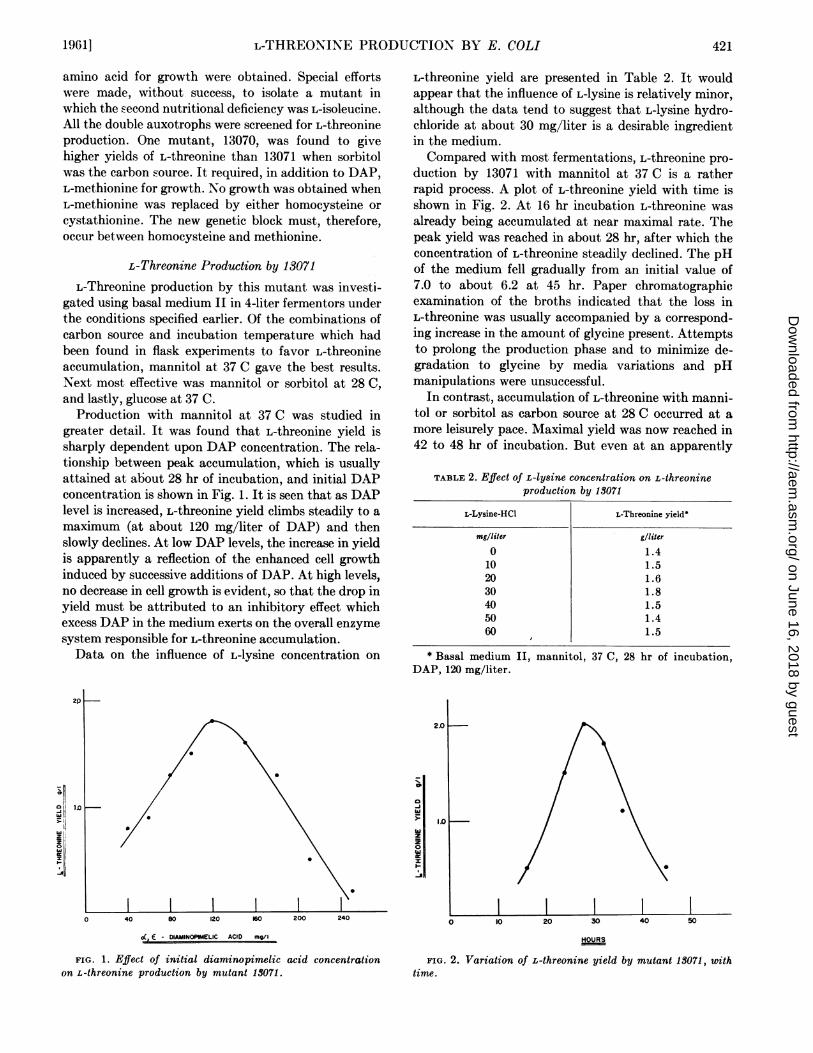
Production with mannitol at 37 C was studied ingreater detail. It was found that L-threonine yield issharply dependent upon DAP concentration. The rela-tionship between peak accumulation, which is usuallyattained at about 28 hr of incubation, and initial DAPconcentration is shown in Fig. 1. It is seen that as DAPlevel is increased, L-threonine yield climbs steadily to a
maximum (at about 120 mg/liter of DAP) and thenslowly declines. At low DAP levels, the increase in yieldis apparently a reflection of the enhanced cell growthinduced by successive additions of DAP. At high levels,no decrease in cell growth is evident, so that the drop inyield must be attributed to an inhibitory effect whichexcess DAP in the medium exerts on the overall enzymesystem responsible for L-threonine accumulation.Data on the influence of L-lysine concentration on
L-threonine yield are presented in Table 2. It wouldappear that the influence of L-lysine is relatively minor,although the data tend to suggest that L-lysine hydro-chloride at about 30 mg/liter is a desirable ingredientin the medium.Compared with most fermentations, L-threonine pro-
duction by 13071 with mannitol at 37 C is a ratherrapid process. A plot of L-threonine yield with time isshown in Fig. 2. At 16 hr incubation L-threonine was
already being accumulated at near maximal rate. Thepeak yield was reached in about 28 hr, after which theconcentration of L-threonine steadily declined. The pHof the medium fell gradually from an initial value of7.0 to about 6.2 at 45 hr. Paper chromatographicexamination of the broths indicated that the loss inL-threonine was usually accompanied by a correspond-ing increase in the amount of glycine present. Attemptsto prolong the production phase and to minimize de-gradation to glycine by media variations and pHmanipulations were unsuccessful.
In contrast, accumulation of L-threonine with manni-tol or sorbitol as carbon source at 28 C occurred at a
more leisurely pace. Maximal yield was now reached in42 to 48 hr of incubation. But even at an apparently
TABLE 2. Effect of L-lysine concentration on L-threonineproduction by 13071
L-Lysine-HCI L-Threonine yield*
mglliter g/liter
0 1.410 1.520 1.630 1.840 1.550 1.460 1.5
* Basal mediumDAP, 120 mg/liter.
II, mannitol, 37 C, 28 hr of incubation,
-J
CU
zz0
I.I-
-m
I I I
1D
I0 10 20 30 40 50
HOURS
FIG. 2. Variation of L-threonine yield by mutant 13071, withtime.
2 1--
0 1D
z. Iz
w
0 40 so 120 IS0 200 240
oct E - DIAMINOPIMELIC ACID mg/I
FIG. 1. Effect of initial diaminopimelic acid concentrationon L-threonine production by mutant 13071.
1961] 421
on June 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
H. T. HUANG
optimal level of DAP (130 mg/liter), the highest yieldachieved was no more than 1 g/liter, and compares
rather unfavorably with that obtainable when doubleauxotroph 13070 was cultivated under similar condi-tions.
L-Threonine Production by 13070
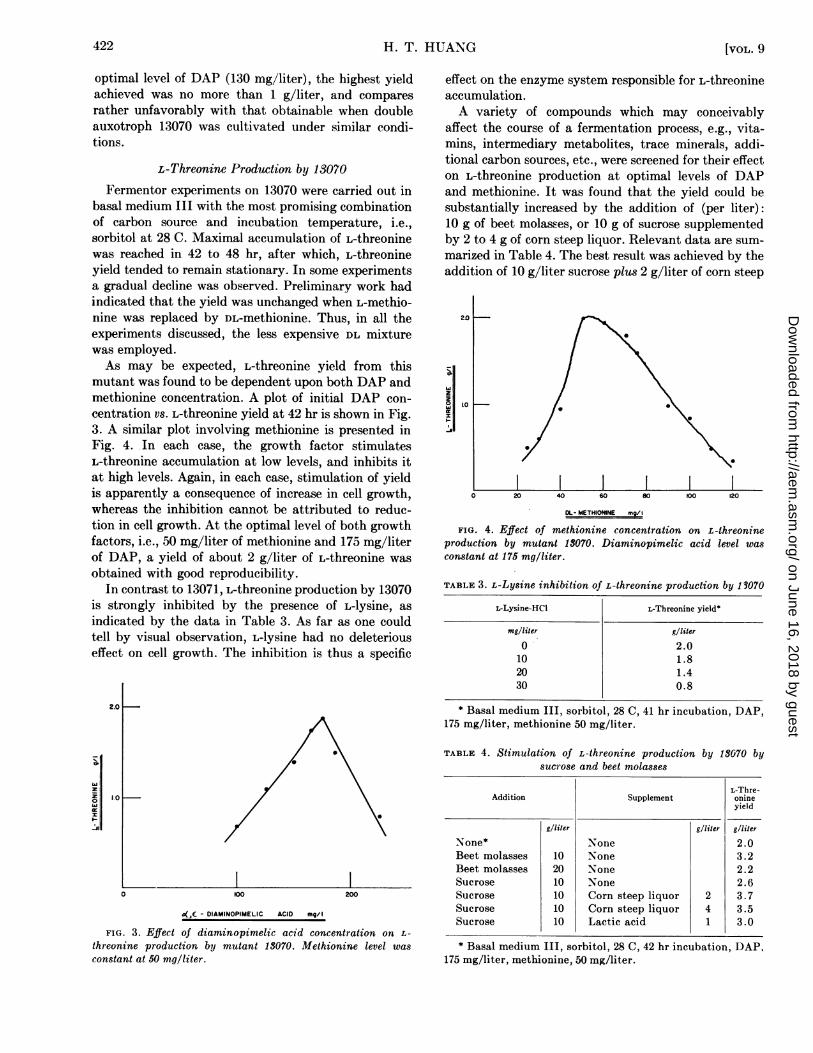
Fermentor experiments on 13070 were carried out inbasal medium III with the most promising combinationof carbon source and incubation temperature, i.e.,sorbitol at 28 C. Maximal accumulation of L-threoninewas reached in 42 to 48 hr, after which, L-threonineyield tended to remain stationary. In some experimentsa gradual decline was observed. Preliminary work hadindicated that the yield was unchanged when L-methio-nine was replaced by DL-methionine. Thus, in all theexperiments discussed, the less expensive DL mixturewas employed.As may be expected, L-threonine yield from this
mutant was found to be dependent upon both DAP andmethionine concentration. A plot of initial DAP con-
centration vs. L-threonine yield at 42 hr is shown in Fig.3. A similar plot involving methionine is presented inFig. 4. In each case, the growth factor stimulatesL-threonine accumulation at low levels, and inhibits itat high levels. Again, in each case, stimulation of yieldis apparently a consequence of increase in cell growth,whereas the inhibition cannot be attributed to reduc-tion in cell growth. At the optimal level of both growthfactors, i.e., 50 mg/liter of methionine and 175 mg/literof DAP, a yield of about 2 g/liter of L-threonine was
obtained with good reproducibility.In contrast to 13071, L-threonine production by 13070
is strongly inhibited by the presence of L-lysine, as
indicated by the data in Table 3. As far as one couldtell by visual observation, L-lysine had no deleteriouseffect on cell growth. The inhibition is thus a specific
20LzC,
IAJZ
z0Lhigrx1--
Li
1.0
0 200
o(,C - DIAMINOPIMELIC ACID mg/I
FIG. 3. Effect of diaminopimelic acid concentration on L-
threonine production by mutant 13070. Methionine level was
constant at 50 mg/liter.
effect on the enzyme system responsible for L-threonineaccumulation.A variety of compounds which may conceivably
affect the course of a fermentation process, e.g., vita-mins, intermediary metabolites, trace minerals, addi-tional carbon sources, etc., were screened for their effecton L-threonine production at optimal levels of DAPand methionine. It was found that the yield could besubstantially increased by the addition of (per liter):10 g of beet molasses, or 10 g of sucrose supplementedby 2 to 4 g of corn steep liquor. Relevant data are sum-
marized in Table 4. The best result was achieved by theaddition of 10 g/liter sucrose plus 2 g/liter of corn steep
2.0
a:
z.. LO .-
I20 40 60 80 100 120
DL- METHIONINE mg/
FIG. 4. Effect of methionine concentration on L-threonineproduction by mutant 13070. Diaminopimelic acid level wasconstant at 175 mg/liter.
TABLE 3. L-Lysine inhibition of L-threonine production by 1 3070
L-Lysine-HCI L-Threonine yield*
mg/liter gllier0 2.010 1.820 1.430 0.8
* Basal medium III, sorbitol, 28 C, 41 hr incubation, DAP,175 mg/liter, methionine 50 mg/liter.
TABLE 4. Stimulation of L-threonine production by 13070 bysucrose and beet molasses
L-Thre-Addition Supplement onine
yield
glliter glliter gl/tier
None* None 2.0Beet molasses 10 None 3.2Beet molasses 20 None 2.2Sucrose 10 None 2.6Sucrose 10 Corn steep liquor 2 3.7Sucrose 10 Corn steep liquor 4 3.5Sucrose 10 Lactic acid 1 3.0
* Basal medium III, sorbitol, 28 C, 42 hr incubation, DAP,175 mg/liter, methionine, 50 mg/liter.
422 [VOL. 9
on June 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

L-THREONINE PRODUCTION BY E. COLI4
liquor when a yield almost double that in the controlmedium was obtained. Practically no stimulation wasobserved when sucrose was replaced by glucose, glyc-erol, lactose, or sorbitol. The effect thus appears to bespecific for sucrose in combination with some factor (orfactors) present in both beet molasses and corn steepliquor.
In all these experiments, the pH of the medium fellvery gradually from an initial value of about 7.0 toabout 6.2 at 42 to 48 hr and about 6.0 at 60 hr. \Noimprovement in yield was observed when the mediumwas maintained at pH 7.0 during the incubation period.
Recovery of L-ThreonineSamples of pure L-threonine have been isolated from
optimal broths of both mutants, i.e., 13071 with manni-tol at 37 C, and 13070 with sorbitol at 27 C. In eachcase the broth also contained glycine (<0.25 g/liter)and serine (< 0.1 g/liter), with minor quantities ofglutamic acid and valine. The procedure adopted wasas follows. The broth was adjusted to pH 2.0 withH2SO4 and filtered. The clear filtrate was passed througha column of IR 120 (H+), which absorbed all the aminoacids. After washing with water, the column waseluted with 1 N NH40H. Fractions were examined byrapid paper chromatography, and those containingamino acids were combined and evaporated in vacuoto a convenient volume. The neutral concentrate wasadjusted to pH 2.0 with H2SO4 and passed through acolumn of permutit Q (H+). The column was washedwith water and eluted with 0.5 N NH40H. Most of thethreonine came off between pH 4 and 9, together withlesser amounts of glycine. Subsequent fractions con-tained more glycine than threonine and were discarded.The threonine rich fractions were combined, concen-trated in vacuo, treated with Darco G 60, and filtered.Addition of methanol to the filtrate induced crystalliza-tion of threonine. One recrystallization of the productfrom aqueous methanol was sufficient to give an ana-lytically pure sample of L-threonine. The yield of purematerial from broth was about 50 %.A typical batch of recovered L-threonine was charac-
terized as follows: [a]25 = -29.3 (C, 2 in H20);
C4H9N03Found: C 40.1, H 7.7, N 11.8Calculated: C 40.3, H 7.6, N 11.8.
Extensive paper chromatographic analyses indicatethat it contained no allothreonine, and less than 0.1%of serine, glycine, and glutamic acid. In contrast, fourcommercial "analytical" grade samples of L-threoninewere found to contain from 1.5 to 7.2 % allothreonine.
DiscussioN-
The accumulation of threonine by a number of DAPrequiring mutants of E. coli, e.g., 173-25, the progenitor
of strain D, was first reported by Davis (1952). Ourwork thus provides unequivocal confirmation of Davis'soriginal observation.
In the mutants studied, the auxotrophic require-ments, i.e. DAP and methionine, and the productaccumulated, i.e. threonine, lie on separate biosyn-thetic pathways which are interrelated by having acommon origin. The interrelationship is shown sche-matically in the following diagram:
Aspartate --+ Compound X---j-+-- DAP --+ Lysine13070
l 13071Homoserine ---+ Threonine --+ Isoleucine
ICysteine --+ Cystathionine--+Homocysteine---J-1----+Methionine
13070
All three compounds are ultimately derived from as-partate, although threonine and methionine have amore immediate common precursor in homoserine.From our present knowledge of the biosynthetic inter-mediate between aspartate and homoserine (Black andWright, 1955) and of the mechanism of DAP biosyn-thesis from aspartate (Gilvarg, 1960; Rhuland, 1960)it is evident "compound X" is aspartic acid /3-semi-aldehyde.The positions at which genetic blocks occur are
indicated by vertical double lines. Consider first mutant13071. It is blocked just before DAP, although theactual point of block has not been determined. It mayreasonably be expected that one effect of the geneticblock would be a general piling up of the precedingintermediates, including compound X, on the DAP-lysine pathway. The excess amount of compound Xthus made available could overflow into the alternatepathway and result in an accumulation of threonine.Just why threonine, and not some other metabolite,say methionine, is actually accumulated evidentlydepends on factors of which we have, as yet, littleknowledge. The same mechanism apparently operatesin the reverse direction in the homoserine deficientMicrococcus (Kinoshita et al., 1958), which accumulateslysine. Similarly, in the case of double auxotroph 13070,the additional genetic block just before methionineshould lead to an increase in the availability of homo-serine. Thus, a marked improvement in threonine yieldwas achieved.
Since threonine is an obligatory precursor in E. coliof isoleucine (Umbarger, 1956) one would expect that ablock between these two amino acids could lead toaccumulation of threonine. So far no threonine accumu-lation was observed in single auxotrophs deficient inisoleucine. Our attempts to impose an isoleucine defi-ciency in 13071 were unsuccessful. However, this stillremains a desirable objective in further mutation work.The occurrence of threonine racemase activity in E.
coli has been reported (Amos, 1954). Particular care
1961] 423 on June 16, 2018 by guest
http://aem.asm
.org/D
ownloaded from

H. T. HUANG
was exercised in this work to detect any discrepanciesbetween microbiological and chromatography assayswhich might suggest the presence of D-threonine. Thedata indicated consistently that all the threonine inbroth or in the recovered product was of the L configura-tion.
ACKNOWLEDGMENTS
I thank Joan Griffin and Caroline Lasar for invalu-able assistance in the mutation and screening work,Peter Ashfield for conducting the fermentor experi-ments, Roberta Meyer for the paper chromatographydeterminations, and the Bioassay Section for micro-biological assays.
LITERATURE CITED
AMOS, H. 1954. A racemase for threonine in Escherichia coli.J. Am. Chem. Soc. 76:3858.
BLACK, S., AND N. G. WRIGHT. 1955. Homoserine dehydro-genase. J. Biol. Chem. 213:51-60.
CASIDA, L. E. 1956. Preparation of diaminopimelic acid andlysine. U. S. Patent 2,771,396.
DAVIS, B. D. 1948. Isolation of biochemically deficient mutantsof bacteria by penicillin. J. Am. Chem. Soc. 70:4267.
DAVIS, B. D. 1949. The isolation of biochemically deficientmutants of bacteria by means of penicillin. Proc. Natl.Acad. Sci. U. S. 35:1-10.
DAVIS, B. D. 1952. Biosynthetic interrelations of lysine,diaminopimelic acid and threonine in mutants of Escher-ichia coli. Nature 169:534-536.
DAVIS, B. D., AND E. S. MINGIOLI. 1950. Mutants of Escherichiacoli requiring methionine or vitamin B12. J. Bacteriol.60:17-28.
GILVARG, C. 1960. Biosynthesis of diaminopimelic acid. Federa-tion Proc. 19:948-952.
HUANG, H. T. 1960. Production of L-threonine. U. S. Patents2,937,121; 2,937,122.
KINOSHITA, S. 1959. The production of amino acids by fermen-tation processes. Advances in Appl. Microbiol. 1:201-214.
KINOSHITA, S., K. NAKAYAMA, AND S. UDAKA. 1957. The fer-mentive production of L-ornithine. J. Gen. Appl. Micro-biol. (Tokyo), 3:276-277.
KINOSHITA, S., S. UDAKA, AND M. SHIMONO. 1957. Studies onthe amino acid fermentation. I. Production of L-glutamicacid by various microorganisms. J. Gen. Appl. Microbiol.(Tokyo) 3:193-205.
KINOSHITA, S., K. NAKAYAMA, AND S. KITADA. 1958. L-Lysineproduction using microbial auxotroph. J. Gen. Appl.Microbiol. (Tokyo) 4:128-129.
KITA, D. A., AND H. T. HUANG. 1958. Fermentation processfor the production of L-lysine. U. S. Patent 2,841,532.
LEDERBERG, J., AND N. ZINDER. 1948. Concentration of bio-chemical mutants of bacteria with penicillin. J. Am.Chem. Soc. 70:4267-4268.
MEADOW, P., D. S. HOARE, AND E. WORK. 1957. Interrelation-ships between lysine and a, e-diaminopimelic acid andtheir derivatives and analogues in mutants of Escherichiacoli. Biochem. J. 66:270-282.
RHULAND, L. E. 1960. a,e-Diaminopimelic acid, its distribution,synthesis and metabolism. Nature 185:224-228.
RHULAND, L. E., E. WORK, R. F. DENMAN, AND D. S. HOARE.1955. The behavior of the isomers of a, E-diaminopimelicacid on paper chromatograms. J. Am. Chem. Soc. 77:4844-4846.
SHULL, G. M., AND D. A. KITA. 1955. Microbiological con-version of steroids. I. Introduction of the 11-,-hydroxylgroup into C21 steroids. J. Am. Chem. Soc. 77:763.
UMBARGER, H. E. 1956. L-Threonine, an obligatory precursor ofL-isoleucine in Escherichia coli. Federation Proc. 15:374.
[VOL. 9424
on June 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from