probiotic treatment reduces depressive-like behaviour in … · forced swim test (fst) is a...
TRANSCRIPT

Pi
ASa
b
c
d
e
a
ARRA
KDHPCHM
1
alatdMp
s
h0
Psychoneuroendocrinology 79 (2017) 40–48
Contents lists available at ScienceDirect
Psychoneuroendocrinology
jo ur nal ho me p ag e: www.elsev ier .com/ locate /psyneuen
robiotic treatment reduces depressive-like behaviour in ratsndependently of diet
nders Abildgaard a,b,∗, Betina Elfving a, Marianne Hokland c, Gregers Wegener a,d,ten Lund e
Translational Neuropsychiatry Unit, Aarhus University, Skovagervej 2, 8240 Risskov, DenmarkDepartment of Clinical Biochemistry, Aarhus University Hospital, Palle Juul-Jensens Boulevard 99, 8200 Aarhus N, DenmarkDepartment of Biomedicine, Aarhus University, Bartholins Allé 6, Build. 1242, 8000 Aarhuc C, DenmarkCenter of Excellence for Pharmaceutical Sciences, North-West University (Potchefstroom Campus), Hoffman Street, Potchefstroom, 2520, South AfricaDepartment of Medical Endocrinology (MEA), Aarhus University Hospital, Nørrebrogade 44, 8000 Aarhus C, Denmark
r t i c l e i n f o
rticle history:eceived 14 December 2016eceived in revised form 13 February 2017ccepted 13 February 2017
eywords:epressive-like behaviourigh-fat dietrobioticsytokinesPA axisetabolomics
a b s t r a c t
The gut microbiota has recently emerged as an important regulator of brain physiology and behaviourin animals, and ingestion of certain bacteria (probiotics) therefore appear to be a potential treatment formajor depressive disorder (MDD). However, some conceptual and mechanistical aspects need furtherelucidation.
We therefore aimed at investigating whether the habitual diet may interact with the effect of pro-biotics on depression-related behaviour and further examined some potentially involved mechanismsunderlying the microbe-mediated behavioural effects.
Forty male Sprague-Dawley rats were fed a control (CON) or high-fat diet (HFD) for ten weeks andtreated with either a multi-species probiotic formulation or vehicle for the last five weeks.
Independently of diet, probiotic treatment markedly reduced depressive-like behaviour in the forcedswim test by 34% (95% CI: 22–44%). Furthermore, probiotic treatment skewed the cytokine production bystimulated blood mononuclear cells towards IFN�, IL2 and IL4 at the expense of TNF� and IL6. In addition,probiotics lowered hippocampal transcript levels of factors involved in HPA axis regulation (Crh-r1, Crh-r2 and Mr), whereas HFD increased these levels. A non-targeted plasma metabolomics analysis revealed
that probiotics raised the level of indole-3-propionic acid, a potential neuroprotective agent.Our findings clearly support probiotics as a potential treatment strategy in MDD. Importantly, theefficacy was not attenuated by intake of a “Western pattern” diet associated with MDD. Mechanistically,the HPA axis, immune system and microbial tryptophan metabolism could be important in this context.Importantly, our study lend inspiration to clinical trials on probiotics in depressed patients.
© 2017 Elsevier Ltd. All rights reserved.
. Introduction
The emerging literature on the gut microbiota (GM) has outlined considerable crosstalk between the residing microbes and physio-ogical systems of the host, including brain and behaviour (Sampsonnd Mazmanian, 2015). This realization has led to research intohe possible importance of the GM in several psychiatric disor-
ers, including major depressive disorder (MDD) (Sampson andazmanian, 2015). Indeed, recent studies imply that the GM maylay a role in the aetiology of MDD. Firstly, studies have identified a
∗ Corresponding author at: Department of Clinical Biochemistry, Aarhus Univer-ity Hospital, Palle Juul-Jensens Boulevard 99, 8200 Aarhus N, Denmark.
E-mail address: [email protected] (A. Abildgaard).
ttp://dx.doi.org/10.1016/j.psyneuen.2017.02.014306-4530/© 2017 Elsevier Ltd. All rights reserved.
number of changes in GM composition in depressed patients (Jianget al., 2015; Naseribafrouei et al., 2014). Secondly, two intriguingstudies have shown that the depressive phenotype may be trans-ferred from depressed patients to rats and mice through the GM(Kelly et al., 2016; Zheng et al., 2016).
A number of pre-clinical studies have now evaluated whethermanipulations of the GM through ingestion of probiotics (definedas live microorganisms which when administered in adequateamounts confer a health benefit on the host (Hill et al., 2014)) canaffect behaviour in animals. Indeed, it appears that probiotics maybe used to modulate a wide range of behaviours related to psychi-
atric disorders, including depressive-like behaviour (Bravo et al.,2011; Desbonnet et al., 2010; Liang et al., 2015; Liu et al., 2016;Savignac et al., 2014). Consequently, the term “psychobiotics” was
uroen
cip
lbuai2scbaeldt(2coeefie
cotawhefiaio
erigatp
2
2
Dilaotc1c(cvw
A. Abildgaard et al. / Psychone
onceptualised by Dinan and colleagues and denotes a live organ-sm that produces a health benefit in patients suffering from asychiatric illness (Dinan et al., 2013).
However, several questions regarding the mechanisms under-ying the ability of certain bacteria to modulate depressive-likeehaviour remain unanswered. Firstly, several studies have eval-ated the activity of the hypothalamic-pituitary-adrenal (HPA)xis through plasma corticosterone measurements with conflict-ng results (Bravo et al., 2011; Desbonnet et al., 2008; Liang et al.,015; Liu et al., 2016; Savignac et al., 2014). Since the release oftress hormones occurs in rapid ultradian oscillations, follows aircadian rhythm and is easily affected by stress resulting from thelood sampling itself, crude plasma levels may be considered rel-tively insensitive in the evaluation of HPA axis regulation (Spigat al., 2014). So far, the effect of probiotics on central HPA axis regu-ation by hippocampus and hypothalamus has not been studied inetail. Secondly, several studies have evaluated immunomodula-ory properties of probiotics through changes in plasma cytokinesArseneault-Breard et al., 2012; Desbonnet et al., 2008; Liang et al.,015; Liu et al., 2016). However, the local and paracrine activity ofytokines holds a strong limitation to the functional interpretationf their plasma concentration. Thirdly, it has been shown that GMxerts a considerable influence on blood metabolite levels (Wikofft al., 2009). Unfortunately, no comprehensive metabolomic pro-ling has been carried out in relation to probiotic ingestion and itsffect on depressive-like behaviour.
Another issue regarding probiotic treatment may be the con-urrent diet. Importantly, MDD is associated with increased ratesf dysmetabolic conditions, including obesity and diabetes mellitusype II (Luppino et al., 2010; Rotella and Mannucci, 2013), and thisssociation may be partly caused by an unhealthy diet associatedith MDD (Jacka et al., 2011; Le Port et al., 2012). Interestingly, it
as been shown that diet has a major impact on the GM (Davidt al., 2014), and particularly, a “Western pattern” high-fat low-bre diet is associated with marked changes in GM compositionnd poor microbial diversity (Albenberg and Wu, 2014). However,t remains unknown whether a such diet interacts with the effectsf probiotics.
In the present study, we therefore aimed at evaluating theffect of probiotic treatment on depression-related behaviour inats on a control or high-fat diet (HFD). Furthermore, we exam-ned three mechanisms that may be involved in MDD and in theut microbiota-mediated effects on depression-related behaviours well; namely, hippocampal and hypothalamic HPA axis regula-ion, cytokine profile of stimulated immune cells and the completelasma metabolomic profile.
. Materials and methods
.1. Animals and diets
Forty male Sprague-Dawley (SD) rats (NTac:SD, SPF; Taconic,enmark) arrived at the age of four weeks and were pair-housed
n Eurostandard Type III H cages (37.5 × 21.5 × 18 cm) with raisedids, a shelter and nesting material. The environment was temper-ture (20 ◦C) and light-controlled (12/12 h light/dark cycle, lightsn at six AM). After arrival, the animals were randomly assignedo a control (CON) or high-fat diet (HFD) (CON cat no. E15000/HFDat no. E15741, Ssniff, Germany). The CON diet had a fat content of1 kJ% (soybean oil), a protein content of 23 kJ% and a carbohydrateontent of 66 kJ% (mainly corn starch). HFD had a fat content 60 kJ%
mainly beef tallow), a protein content of 20 kJ% and a carbohydrateontent of 20 kJ% (maltodextrin and sucrose). The diets were pro-ided ad libitum together with tap water. All rats and diet remnantsere weighed once a week. All procedures complied with the EUdocrinology 79 (2017) 40–48 41
Directive 2010/63/EU and with the Danish law regulating exper-iments on animals (permission ID 2012-15-2934-00254). Studydesign is depicted in Fig. 1.
2.2. Probiotics
After five weeks on the diets, the rats were randomised to vehi-cle (VEH) or probiotic (PRO) treatment. The probiotics consistedof eight bacterial strains (B. bifidum W23, B. lactis W52, L. aci-dophilus W37, L. brevis W63, L. casei W56, L. salivarius W24, Lc. LactisW19, Lc. Lactis W58; “Ecologic Barrier”, Winclove Probiotics BV,The Netherlands) in a carrier matrix of maize starch, maltodextrinsand vegetable protein. Each cage received a bottle containing 4.5 g(2.5 × 109 CFU/g) of freeze-dried powder dissolved in 30 mL of tapwater. VEH groups were given the carrier matrix only. The bottleswere administered daily between four and six pm and completelyemptied by the animals during the night. Bottles were chosen overoral gavage to minimise the amount of stress inflicted on the ani-mals. The experimenter was blinded to treatment groups.
2.3. Barnes maze
The barnes maze (BM) is a test of hippocampus-dependent spa-tial memory which is often impaired in patients suffering fromMDD (Campbell and Macqueen, 2004). The maze consisted of ablack, circular table (Ø 120 cm) with 18 perimetric holes, and oneof the holes was connected to a hidden escape box. Black curtainsequipped with spatial cues (simple geometric symbols) surroundedthe maze. Each rat was tested twice daily with a 3-h interval forfour consecutive days (eight trials in total). In each trial, the ratwas allowed to spend a maximum of four minutes to locate theescape hole, otherwise it was gently moved to it by the experi-menter. An aversive environment was established by bright light(800 Lux) and loud background noise (White Noise Generator Soft-ware; www.sobolsoft.com). Just as the rat entered the escape hole,the noise was turned off, and the rat was collected after a 30-s delay.To exclude any local cues, the maze was turned 120◦ daily, and theescape box was moved proportionally to maintain the same phys-ical location in the room. Seven days after the last trial, a recalltrial was performed. The maze was wiped with an ethanol solutionbetween each trial. The sessions were video recorded and analysedby EthoVision XT9 software (Noldus IT, The Netherlands). Time (s)and distance moved (cm) before locating the escape hole were cal-culated. In the first trial, velocity (cm/s) was calculated as a measureof locomotor activity.
2.4. Forced swim test
The forced swim test (FST) is a commonly used screening toolfor depressive-like behaviour in rodents. Immobility (depressive-like behaviour) is defined as the rat making no movements beyondthose needed to keep its head above the water. We utilised themodified version of the test (Detke et al., 1995). Briefly, each ratwas immersed in a water-filled cylinder (H: 54 cm; Ø: 24 cm; waterdepth: 40 cm; 24–25 ◦C) and allowed to swim for 15 and sevenmin,respectively, on two consecutive days. The water was changedbetween each trial. All swim sessions were video-recorded. Thesecond day test trial was scored afterwards by an experimenter
blinded to group assignment using the time-sampling technique,i.e. the most predominant behaviour during each 5-s time bin wasregistered. Time spent on active behaviours (struggling and swim-ming) as well as immobility was calculated.
42 A. Abildgaard et al. / Psychoneuroendocrinology 79 (2017) 40–48
FT fat dieo
2
oFsse
2
NwacgmaosG
2
tcai
2
(Uas1ctbtaCs7s
2
Im
ig. 1. Study design and outline.he animals were four weeks old at study initiation. CON: control diet; HFD: high-ral glucose tolerance test.
.5. Open field test
To assess locomotor activity, the rats were subjected to a 10-minpen field test (OFT) immediately before the second day test trial ofST. Each animal was placed in a black 100 × 100 cm video-recordedquare box. The videos were analysed by Noldus Ethovision XT9oftware (Noldus IT, The Netherlands), and the distance moved byach animal (cm) was retrieved.
.6. Oral glucose tolerance test
Following a 6-h fast, a blood sample was taken from a tail snip.ext, the rat was given a glucose solution (2.5 g glucose/kg bodyeight; 500 g/L) by oral gavage, and blood samples were drawn
gain 30, 60 and 120 min later. For each time point, blood glucoseoncentration was measured in duplicate by use of a OneTouch Vitalucose monitor (Lifescan, Cilag GmbH, Switzerland). For insulineasurements, an additional amount of blood was collected in hep-
rinized capillary tubes, and the plasma was immediately frozenn dry ice. Insulin concentrations were measured with an ultra-ensitive rat insulin ELISA kit (DRG Diagnostics GmbH, Marburg,ermany).
.7. Blood and tissue collection
Animals were given a lethal dose of pentobarbital intraperi-oneally. At the absence of all reflexes (after 3–5 min), blood wasollected by cardiac puncture. After decapitation, hypothalamusnd hippocampus were dissected by an experienced technician andmmediately frozen on powdered dry ice.
.8. Anti-CD3/28 stimulation of PBMC
Peripheral blood (EDTA-anticoagulated) mononuclear cellsPBMC) were isolated by Ficoll-Paque 1.084 (GE Healthcare, Illinois,SA) density gradient centrifugation, washed twice in sterile PBSnd finally resuspended in RPMI-1640 (R8758; Sigma-Aldrich, Mis-ouri, USA) supplemented with 10% FBS, 100 U/mL penicillin and0 �g/mL streptomycin (P4333; Sigma-Aldrich, Missouri, USA). Aell count was obtained from a haemocytometer, and the concen-ration was adjusted to 106 cells/mL. Cell viability was determinedy trypan blue exclusion. Two hundred thousand cells were addedo each well of a U-bottom microplate that was pre-coated withnti-rat CD3 (5 �g/mL in 100 �L PBS; clone G4.18; eBioscience,alifornia, USA). Finally, soluble anti-rat CD28 (clone JJ319; eBio-cience, California, USA) was added to the wells at 1.5 �g/mL. After2 h of stimulated culture, the supernatants were collected andtored at −80◦.
.9. Measurement of cytokines
Six cytokines were measured in the PBMC culture supernatants.nterleukin 2 (IL2), IL4, IL6 and interferon gamma (IFN�) were
easured with a magnetic-bead based multiplex kit (Bio-Rad,
t; VEH: vehicle; PRO: probiotics; FST: forced swim test; OF: open field test; OGTT:
California, USA) according to the manufacturer’s instructions andassayed on a Luminex Bio-Plex 200 system (Bio-Rad, California,USA). IL10 and tumor necrosis factor alpha (TNF�) were measuredwith Quantikine Elisa kits (R&D Systems, Minneapolis, USA). Abso-lute levels as well as relative cytokine composition were analysed.
2.10. Plasma endotoxin
Lipopolysaccharide (LPS) was measured in sterile plasma(lithium heparin-anticoagulated) with a limulus amoebocyte lysateassay (Lonza AG, Switzerland). Spiked samples were made to deter-mine the dilution needed in order to avoid any inhibitory matrixcomponents. Consequently, plasma samples were diluted 25 timesand heated at 70 ◦C for 15 min. The assay was then done accordingto manufacturer’s instruction.
2.11. RNA extraction & cDNA synthesis
PARIS kits (Thermo Fisher Sci., Massachusetts, USA) were usedto extract RNA from left hippocampus and hypothalamus accordingto the manufacturer’s instructions. RNA concentration and puritywere determined by a Nanodrop spectrophotometer (ThermoFisher Sci., Massachusetts, USA), and the RNA concentration ofeach sample was adjusted to the lowest one observed (31 ng/�L(hippocampus) and 64 ng/�L (hypothalamus)). Reverse transcrip-tion PCR was performed to generate cDNA using random primersand Superscript IV Reverse Transcriptase (Thermo Fisher Sci., Mas-sachusetts, USA) according to the manufacturerı́s instructions. Theresulting cDNA was stored undiluted at −80 ◦C until quantitativereal-time polymerase chain reaction (real-time qPCR) analysis.
2.12. Real-time qPCR
The expression of seven genes related to structural plasticity(Bdnf, Creb1, Igf-1, VegfA, Traak, Trek2, TrkB) and six genes related toHPA axis regulation (Gr, Mr, 11ˇ-hsd1, Crh-bp, Crh-r1, Crh-r2) wasdetermined in hippocampus by use of real-time qPCR. In hypotha-lamus, only genes involved in HPA axis regulation were assessed(Gr, Mr, 11ˇ-hsd1, Crh-r1, Crh-r2, Pomc). In addition, eight refer-ence genes were included (ActB, Ywhaz, Hmbs, Hprt, Rpl13, CycA,18s rrna, Gapdh). Primers are listed in supplementary Table S1. Thereal-time qPCR reactions were carried out on a Mx3000P system(Stratagene, California, USA) as described previously (Elfving et al.,2008). Briefly, each well of a 96-well microplate was loaded withSYBR green master-mix (Bio-Rad, California, USA), 0.5 uM of primerpairs and 3 uL of diluted cDNA (hippocampus 1:11; hypothalamus1:20) (10 uL in total). A standard curve in duplicate was made oneach plate, and all samples were also run in duplicate. Individual
sample genes were normalised to the geometric mean of the twomost stable reference genes which were selected by Normfindersoftware (hippocampus CycA/Rpl13; hypothalamus Actb/18 s rRNA)(Bonefeld et al., 2008).
uroendocrinology 79 (2017) 40–48 43
2
pmagcus(tHiorno
2
s(uqlbTcdiBlmcidtedCerHoasU
3
3
(iwn
3
(3w
Fig. 2. Forced swim test.Mean ± SEM. Probiotics markedly lowered depressive-like behaviour (immobility)independently of diet. Panel A depicts minute-wise immobility during the 7-min
A. Abildgaard et al. / Psychone
.13. Metabolomics
A commercially available plasma metabolomic analysis waserformed by Metabolon Inc. (North Carolina, USA) to identifyetabolites associated with probiotic treatment. An untargeted
nalysis was chosen in order to take advantage of the hypothesis-enerating potential of the method. Ultrahigh performance liquidhromatography-tandem mass spectroscopy (UPLC–MS/MS) wassed. Briefly, proteins were precipitated with methanol, and eachample was analysed on four platforms: two separate reverse phaseRP)/UPLC–MS/MS with positive ion mode electrospray ioniza-ion (ESI), an RP/UPLC–MS/MS with negative ion mode ESI and aILIC/UPLC–MS/MS with negative ion mode ESI. Compounds were
dentified by comparison to library entries of purified standardsr recurrent unknown entities. Each compound was corrected inun-day blocks by registering the medians to equal one (1.00) andormalizing each data point proportionately. Metabolites that werenly detected in less than 75% of the animals were excluded.
.14. Statistical analyses
A significance level of � = 0.05 was used in all analyses. Groupizes were chosen on the basis of a power calculation for FST� = 0.2). Normality was assessed by QQ plots, and Bartlett’s test wassed to test for equal variances. Struggling time in FST and real-timePCR data were power-transformed, whereas LPS and cytokine
evels as well as data from BM and OGTT were log-transformedefore analysis. Trek2 was analysed using Wilcoxon rank-sum test.wo-way ANOVA was used in analysis of body weight, fasting glu-ose/insulin, FST, OF, LPS, BM recall trial and metabolomics withiet and probiotic treatment as independent factors. Significant
nteractions were followed up by use of pair-wise contrasts withonferroni-corrected p-values. Metabolomics data were also ana-
ysed by use of a principal component analysis (PCA) to detectajor changes in analyte composition. In analysis of BM, OGTT and
ytokines, a linear mixed model analysis was conducted with ratdentity as the random effect (intercept). Fixed effects includeday and session (BM), time (OGTT) or analyte (cytokines) andheir interactions with diet, treatment and diett × treatment. REMLstimation was used, and residuals were considered indepen-ent by trial (BM), time (glucose in OGTT) or analyte (cytokines).ontrasts of marginal linear predictions were used to test for differ-nces between groups after the models were built. False discoveryate (FDR) correction (Benjamini and Hochberg (Benjamini andochberg, 1995)) was applied to real-time qPCR data at a FDR levelf 0.10. For metabolomics, a less conservative FDR level of 0.20 waspplied because of the hypothesis-generating purpose of the analy-is. All analyses were performed with Stata 14 (StataCorp LP, Texas,SA).
. Results
.1. Body weight & caloric intake
Body weights at study initiation were similar between groupssupplementary Table S2). As expected, HFD led to a markedncrease in total caloric intake (F1,36 = 31.7; p < 0.001) and in body
eight at study end (F1,36 = 18.8; p < 0.001). Probiotic treatment didot affect these measures.
.2. Forced swim test
Probiotic treatment had a clear antidepressant-like effectFig. 2). Namely, the treatment reduced immobility time by4% (95% CI: 22–44%; F1,36 = 24.1; p < 0.001), and the effectas completely independent of diet (diet: F1,36 = 0.02; p = 0.9;
test trial, whereas panel B shows total immobility duration in the test.***) p < 0.001; (p) main effect of probiotics.CON: control diet; HFD: high-fat diet; VEH: vehicle; PRO: probiotics.
diet × treatment: F1,36 = 0.15; p = 0.7). An antidepressant-likeeffect under both dietary conditions was further confirmed byBonferroni-adjusted post-hoc comparisons (CON-VEH vs CON-PRO,p = 0.006; HFD-VEH vs HFD-PRO, p = 0.001). Correspondingly, pro-biotics increased the duration of swimming (F1,36 = 10.8; p = 0.002),whereas struggling remained largely unaffected (F1,36 = 2.64;p = 0.1).
3.3. Open field test
No differences between groups were observed in distance trav-elled during the 10-min open field session (diet: F1,36 = 0.81; p = 0.4;probiotics: F1,36 = 0.18; p = 0.7; diett × treatment: F1,36 = 0.15;p = 0.7).
3.4. Barnes maze
Data are presented in supplementary Fig. S3. Across the fourdays, a clear improvement was seen in distance travelled beforelocating the escape hole (day: �2(3) = 307; p < 0.001) and alsoin time spent to complete the trial (day: �2(3) = 557; p < 0.001).In addition, an advance was evident between the two sessionswithin the first and the second day in analysis of distance travelled(day × session: �2(3) = 9.38; p = 0.02) as well as in time to complete(day × session: �2(3) = 21.4; p < 0.001).
Interestingly, HFD was found to improve time to complete thetrials (diet × day: �2(3) = 8.2; p = 0.04). Specifically, HFD led to afaster completion on day 4 only (p = 0.02). Diet did, however, notaffect distance travelled (diet × day: �2(3) = 5.24; p = 0.2). No effectof probiotic treatment was observed (treatment × day: �2(3) = 0.84;
p = 0.8 (time)/�2(3) = 0.53; p = 0.9 (distance)).In the recall-session 7 days later, HFD lowered the distance trav-elled before locating the escape hole (F1,36 = 4.64; p = 0.04), whereasa significant diet × treatment interaction was seen for time to com-

44 A. Abildgaard et al. / Psychoneuroendocrinology 79 (2017) 40–48
Fig. 3. Metabolic measures.Mean ± SEM. HFD was associated with higher glucose (A) and insulin (B) levels dur-ing the oral glucose tolerance test (OGTT). In addition, HFD also led to higher plasmaendotoxin levels (C). Probiotic treatment did not affect any of these measures. Thir-t*C
pto
t
3
wsitO
Fig. 4. Cytokine production by PBMCs during 72 h of anti-CD3/28 stimulated cul-tures.A (absolute levels): Median and 25th/75th percentiles; whiskers represent totalrange. Probiotics increased the amount of IL2, IL4 and IFN� (p < 0.05), but no effectof diet was seen. B (relative distribution): Mean ± SEM. Probiotics increased thelevel of IL2, IL4 and IFN� in percent of the total cytokine amount, but lowered IL6and TNF�. Groups of similar treatment were merged to improve plainness. Absoluteand relative values for individual groups may be found in the supplementary TableS4. Data were missing from two rats because too few PBMCs were recovered (oneCON + VEH rat and one HFD + VEH rat). (p) main effect of probiotics; *) p < 0.05; **)p < 0.01. FDR correction applied.
een of the 160 insulin samples were missing due to haemolysis.) p < 0.05; **) p < 0.01; (d) main effect of diet.ON: control diet; HFD: high-fat diet; VEH: vehicle; PRO: probiotics.
lete (F1,36 = 0.20; p < 0.05). Here, post-hoc comparisons revealedhat HFD led to faster completion of the trial in VEH treated animalsnly (p = 0.03).
As regards locomotor activity, no differences in velocity or dis-ance travelled were observed in the first trial (data not presented).
.5. Oral glucose tolerance test
Baseline fasting levels of blood glucose and plasma insulinere not affected by diet or probiotic treatment, although HFD
trongly tended to increase insulin level (F3,33 = 3.9; p = 0.06). Dur-ng the OGTT, glucose levels rose to a higher level in HFD ratshan in CON rats (time × diet: �2(1) = 7.48; p = 0.006) (Fig. 3a).ver-all, HFD-fed rats also had higher insulin levels that CON
CON: control diet; HFD: high-fat diet; VEH: vehicle; PRO: probiotics.
rats (�2(1) = 5.83; p = 0.02), although diet did not interact withtime course (time × diet: �2(1) = 0.02; p = 0.9) (Fig. 3b). Probiotics-treated animals had lower glucose and insulin levels during the test,but the improvements were not statistically significant (glucose:treatment: �2(1) = 0.03; p = 0.9; time × treatment: �2(1) = 1.33;p = 0.2) (insulin: treatment: �2(1) = 0.93; p = 0.3; time × treatment:�2(1) = 0.14; p = 0.7).
3.6. PBMC cytokine production
The absolute cytokine levels in the supernatant of anti-CD3/28 stimulated PBMCs are shown in Fig. 4a. Over-all, probiotictreatment affected the cytokine levels (treatment × analyte:�2(5) = 25.38; p < 0.001), whereas diet had no such effect(diet × analyte: �2(5) = 6.67; p = 0.24). Specifically, probioticsincreased the level of IL2, IL4 and IFN; yet, the treatment didnot affect the total amount of cytokines in general (�2(1) = 2.07;p = 0.15). In analysis of each cytokine in percent of the totalamount produced, an overall effect of probiotics was observedagain (treatmentt × analyte: �2(5) = 25.68; p < 0.001). Namely, pro-biotics skewed the production towards IL2, IL4 and IFN� at the
expense of the highly pro-inflammatory cytokines IL6 and TNF�(Fig. 4b). Again, no effect of diet was observed (diett × analyte:�2(5) = 8.37; p = 0.14).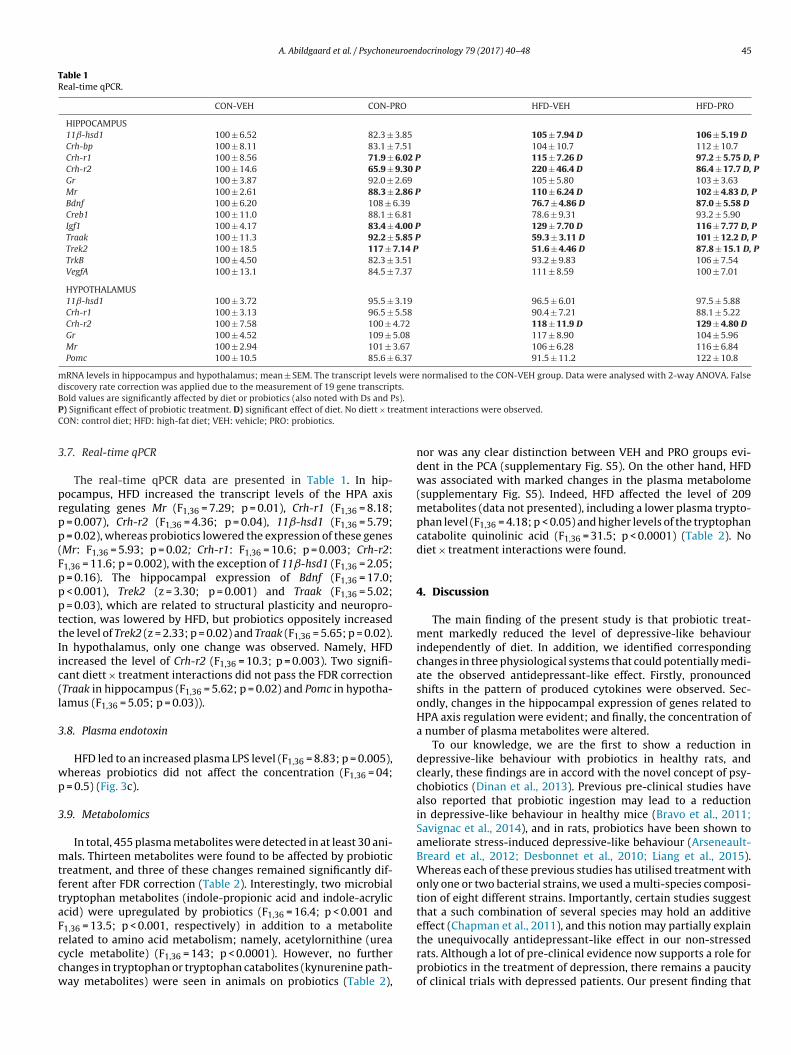
A. Abildgaard et al. / Psychoneuroendocrinology 79 (2017) 40–48 45
Table 1Real-time qPCR.
CON-VEH CON-PRO HFD-VEH HFD-PRO
HIPPOCAMPUS11ˇ-hsd1 100 ± 6.52 82.3 ± 3.85 105 ± 7.94 D 106 ± 5.19 DCrh-bp 100 ± 8.11 83.1 ± 7.51 104 ± 10.7 112 ± 10.7Crh-r1 100 ± 8.56 71.9 ± 6.02 P 115 ± 7.26 D 97.2 ± 5.75 D, PCrh-r2 100 ± 14.6 65.9 ± 9.30 P 220 ± 46.4 D 86.4 ± 17.7 D, PGr 100 ± 3.87 92.0 ± 2.69 105 ± 5.80 103 ± 3.63Mr 100 ± 2.61 88.3 ± 2.86 P 110 ± 6.24 D 102 ± 4.83 D, PBdnf 100 ± 6.20 108 ± 6.39 76.7 ± 4.86 D 87.0 ± 5.58 DCreb1 100 ± 11.0 88.1 ± 6.81 78.6 ± 9.31 93.2 ± 5.90Igf1 100 ± 4.17 83.4 ± 4.00 P 129 ± 7.70 D 116 ± 7.77 D, PTraak 100 ± 11.3 92.2 ± 5.85 P 59.3 ± 3.11 D 101 ± 12.2 D, PTrek2 100 ± 18.5 117 ± 7.14 P 51.6 ± 4.46 D 87.8 ± 15.1 D, PTrkB 100 ± 4.50 82.3 ± 3.51 93.2 ± 9.83 106 ± 7.54VegfA 100 ± 13.1 84.5 ± 7.37 111 ± 8.59 100 ± 7.01
HYPOTHALAMUS11ˇ-hsd1 100 ± 3.72 95.5 ± 3.19 96.5 ± 6.01 97.5 ± 5.88Crh-r1 100 ± 3.13 96.5 ± 5.58 90.4 ± 7.21 88.1 ± 5.22Crh-r2 100 ± 7.58 100 ± 4.72 118 ± 11.9 D 129 ± 4.80 DGr 100 ± 4.52 109 ± 5.08 117 ± 8.90 104 ± 5.96Mr 100 ± 2.94 101 ± 3.67 106 ± 6.28 116 ± 6.84Pomc 100 ± 10.5 85.6 ± 6.37 91.5 ± 11.2 122 ± 10.8
mRNA levels in hippocampus and hypothalamus; mean ± SEM. The transcript levels were normalised to the CON-VEH group. Data were analysed with 2-way ANOVA. Falsediscovery rate correction was applied due to the measurement of 19 gene transcripts.B s).P eatmeC
3
prpp(FpppttIic(l
3
wp
3
mtftaFrccw
old values are significantly affected by diet or probiotics (also noted with Ds and P) Significant effect of probiotic treatment. D) significant effect of diet. No diett × trON: control diet; HFD: high-fat diet; VEH: vehicle; PRO: probiotics.
.7. Real-time qPCR
The real-time qPCR data are presented in Table 1. In hip-ocampus, HFD increased the transcript levels of the HPA axisegulating genes Mr (F1,36 = 7.29; p = 0.01), Crh-r1 (F1,36 = 8.18;
= 0.007), Crh-r2 (F1,36 = 4.36; p = 0.04), 11ˇ-hsd1 (F1,36 = 5.79; = 0.02), whereas probiotics lowered the expression of these genesMr: F1,36 = 5.93; p = 0.02; Crh-r1: F1,36 = 10.6; p = 0.003; Crh-r2:1,36 = 11.6; p = 0.002), with the exception of 11ˇ-hsd1 (F1,36 = 2.05;
= 0.16). The hippocampal expression of Bdnf (F1,36 = 17.0; < 0.001), Trek2 (z = 3.30; p = 0.001) and Traak (F1,36 = 5.02; = 0.03), which are related to structural plasticity and neuropro-ection, was lowered by HFD, but probiotics oppositely increasedhe level of Trek2 (z = 2.33; p = 0.02) and Traak (F1,36 = 5.65; p = 0.02).n hypothalamus, only one change was observed. Namely, HFDncreased the level of Crh-r2 (F1,36 = 10.3; p = 0.003). Two signifi-ant diett × treatment interactions did not pass the FDR correctionTraak in hippocampus (F1,36 = 5.62; p = 0.02) and Pomc in hypotha-amus (F1,36 = 5.05; p = 0.03)).
.8. Plasma endotoxin
HFD led to an increased plasma LPS level (F1,36 = 8.83; p = 0.005),hereas probiotics did not affect the concentration (F1,36 = 04;
= 0.5) (Fig. 3c).
.9. Metabolomics
In total, 455 plasma metabolites were detected in at least 30 ani-als. Thirteen metabolites were found to be affected by probiotic
reatment, and three of these changes remained significantly dif-erent after FDR correction (Table 2). Interestingly, two microbialryptophan metabolites (indole-propionic acid and indole-acryliccid) were upregulated by probiotics (F1,36 = 16.4; p < 0.001 and1,36 = 13.5; p < 0.001, respectively) in addition to a metabolite
elated to amino acid metabolism; namely, acetylornithine (ureaycle metabolite) (F1,36 = 143; p < 0.0001). However, no furtherhanges in tryptophan or tryptophan catabolites (kynurenine path-ay metabolites) were seen in animals on probiotics (Table 2),nt interactions were observed.
nor was any clear distinction between VEH and PRO groups evi-dent in the PCA (supplementary Fig. S5). On the other hand, HFDwas associated with marked changes in the plasma metabolome(supplementary Fig. S5). Indeed, HFD affected the level of 209metabolites (data not presented), including a lower plasma trypto-phan level (F1,36 = 4.18; p < 0.05) and higher levels of the tryptophancatabolite quinolinic acid (F1,36 = 31.5; p < 0.0001) (Table 2). Nodiet × treatment interactions were found.
4. Discussion
The main finding of the present study is that probiotic treat-ment markedly reduced the level of depressive-like behaviourindependently of diet. In addition, we identified correspondingchanges in three physiological systems that could potentially medi-ate the observed antidepressant-like effect. Firstly, pronouncedshifts in the pattern of produced cytokines were observed. Sec-ondly, changes in the hippocampal expression of genes related toHPA axis regulation were evident; and finally, the concentration ofa number of plasma metabolites were altered.
To our knowledge, we are the first to show a reduction indepressive-like behaviour with probiotics in healthy rats, andclearly, these findings are in accord with the novel concept of psy-chobiotics (Dinan et al., 2013). Previous pre-clinical studies havealso reported that probiotic ingestion may lead to a reductionin depressive-like behaviour in healthy mice (Bravo et al., 2011;Savignac et al., 2014), and in rats, probiotics have been shown toameliorate stress-induced depressive-like behaviour (Arseneault-Breard et al., 2012; Desbonnet et al., 2010; Liang et al., 2015).Whereas each of these previous studies has utilised treatment withonly one or two bacterial strains, we used a multi-species composi-tion of eight different strains. Importantly, certain studies suggestthat a such combination of several species may hold an additiveeffect (Chapman et al., 2011), and this notion may partially explain
the unequivocally antidepressant-like effect in our non-stressedrats. Although a lot of pre-clinical evidence now supports a role forprobiotics in the treatment of depression, there remains a paucityof clinical trials with depressed patients. Our present finding that
46 A. Abildgaard et al. / Psychoneuroendocrinology 79 (2017) 40–48
Table 2Metabolomics.
Metabolite CON-VEH CON-PRO HFD-VEH HFD-PRO
Metabolites affected by probiotic treatmentN-delta-acetylornithine 0.69 ± 0.05 2.00 ± 0.22 P 0.62 ± 0.05 D 1.45 ± 0.10P, DIndolepropionate 0.85 ± 0.12 1.34 ± 0.22 P 0.67 ± 0.09 1.37 ± 0.14 PIndoleacrylate 0.92 ± 0.12 1.43 ± 0.24 P 0.86 ± 0.08 1.66 ± 0.14 PPhenyllactate (PLA) 0.80 ± 0.08 1.38 ± 0.19 0.87 ± 0.13 1.18 ± 0.12Caprate (10:0) 1.01 ± 0.05 1.16 ± 0.07 0.85 ± 0.03 1.11 ± 0.09N-methylproline 1.10 ± 0.11 1.31 ± 0.12 0.82 ± 0.05 D 1.14 ± 0.11 DTrigonelline (N′-methylnicotinate) 1.21 ± 0.20 2.33 ± 0.72 0.75 ± 0.09 D 1.00 ± 0.17 DPhenylacetate 0.81 ± 0.12 0.95 ± 0.09 0.96 ± 0.14 D 1.49 ± 0.21 D4-Ethylphenylsulfate 1.32 ± 0.17 0.96 ± 0.10 1.13 ± 0.13 0.85 ± 0.11N6-carboxymethyllysine 0.98 ± 0.08 1.32 ± 0.15 0.97 ± 0.09 1.14 ± 0.11Carnosine 1.54 ± 0.25 0.99 ± 0.15 1.32 ± 0.36 0.91 ± 0.14Dihomo-linolenate 1.35 ± 0.15 1.16 ± 0.15 1.01 ± 0.13 D 0.71 ± 0.07 DN-acetylproline 0.87 ± 0.09 1.51 ± 0.27 0.88 ± 0.09 1.06 ± 0.11
Tryptophan and tryptophan catabolites (kynurenine pathway)Tryptophan 1.05 ± 0.05 1.01 ± 0.04 0.95 ± 0.04 D 0.92 ± 0.04 DKynurenine 1.12 ± 0.09 1.08 ± 0.10 1.05 ± 0.09 0.92 ± 0.07N-acetylkynurenine 1.22 ± 0.13 1.13 ± 0.15 1.11 ± 0.14 0.88 ± 0.10Kynurenic acid 1.12 ± 0.09 1.00 ± 0.08 0.96 ± 0.09 0.99 ± 0.09Quinolinic acid 0.70 ± 0.15 0.66 ± 0.08 1.66 ± 0.16 D 1.34 ± 0.18 D
Bold values are significantly affected by diet or probiotics (also noted with Ds and Ps).Mean ± SEM (arbitrary units). In total, 455 plasma metabolites were identified in at least 30 animals. In the upper part of the table, the 13 metabolites affected by probiotict f the kd of proc
HoEv(
opIaotmiihcTpffiisdsrDmpadcatvl
e
reatment before FDR correction are presented. In the lower part, metabolites oiet × probiotics interaction was identified for any metabolite. P) Significant effect
ontrol diet; HFD: high-fat diet; VEH: vehicle; PRO: probiotics.
FD did not interact with the antidepressant-like effect of probi-tics only further expands the potential group of amenable patients.ncouragingly, a couple of trials on probiotic treatment in healthyolunteers show improvements on depression-related rating scalesMessaoudi et al., 2011; Steenbergen et al., 2015).
The cytokine pattern was altered as a consequence of probi-tic treatment. Specifically, we found a distributional shift towardsroduction of the T lymphocyte-derived cytokines IL2, IL4 and
FN� and away from production of cytokines primarily associ-ted with macrophages, namely TNF� and IL6. Interestingly, thebserved immunomodulatory properties of probiotics appearedo be independent of the highly pro-inflammatory LPS. In several
eta-analyses, the levels of macrophage-derived plasma cytokines,ncluding TNF� and IL6, are consistently found to be modestlyncreased in depressed patients (Dowlati et al., 2010). On the otherand, a meta-analysis concluded that T cell activity is generallyompromised in MDD (Zorrilla et al., 2001). Importantly, certain
cell subsets have recently been found to support the structurallasticity of the CNS and to play a pivotal role in normal brain
unctioning of mice (Kipnis, 2016). Collectively, MDD may there-ore be associated with an overactivity of the innate arm of themmune system to the detriment of the adaptive arm. Interest-ngly, our present cytokine findings may reflect that probioticshifted the balance towards the adaptive arm. Furthermore, ourata are substantiated by the use of standardised ex vivo mea-ures, i.e. stimulated PBMC cultures. Although most similar studieseport crude plasma cytokine levels (Arseneault-Breard et al., 2012;esbonnet et al., 2008; Liang et al., 2015; Liu et al., 2016), sucheasurements remain challenging due to the low levels found in
eripheral blood; furthermore, plasma cytokines are prone to vari-tions caused by e.g. sample collection issues, diurnal rhythm andiet (Zhou et al., 2010). In stimulated cultures, the concentration ofytokines is higher than in plasma, and no environmental factorsffect the production. We therefore believe that our method easeshe measurement of cytokines and enhances their interpretativealue. Nevertheless, further studies are needed to evaluate a causal
ink between the cytokine pattern and the behavioural changes.Probiotic treatment dramatically altered the expression of sev-ral hippocampal genes related to HPA axis feedback. Clearly, our
ynurenine pathway are presented. Data were analysed with 2-way ANOVA. Nobiotics after FDR correction. D) Significant effect of diet after FDR correction. CON:
results are consistent with earlier studies that demonstrate animportant interplay between the gut microbiota and HPA axis(Desbonnet et al., 2008; Sudo et al., 2004). Interestingly, a studyin healthy human volunteers reported a reduction in urinary corti-sol after 30 days of probiotic treatment (Messaoudi et al., 2011).Our data therefore further justify the microbiota as a substan-tial regulator of the neuroendocrine stress axis at the level of theCNS. Momentously, it is well-established that a large proportion ofdepressed patients have a dysregulated HPA axis, and hippocampusis believed to play an important role in the negative feedback mech-anism (Pariante and Lightman, 2008). Noticeably, the hippocampaltranscript levels of Crh-r1 and Crh-r2 were found to be markedlylowered by probiotics in our study. Importantly, increased CRH-R1 signalling has been implicated in stress and depression (Daleet al., 2015), and receptor antagonists were shown to possessantidepressant-like effect in animals. Although clinical studies withsuch antagonists have been conflicting (Dale et al., 2015), thealtered expression may still be of pathophysiological importance.Nevertheless, our findings may serve to corroborate the observedantidepressant-like behavioural effect of the treatment.
HFD also changed the expression of several genes in hip-pocampus. Intriguingly, HFD and probiotics had oppositely directedeffects on all affected genes, and HFD thus increased the mRNAlevel of several factors involved in HPA axis regulation. Previousstudies have shown that chronic HFD consumption may lead toincreased basal and stress-induced plasma corticosterone levels(Abildgaard et al., 2014; Tannenbaum et al., 1997), and it is possi-ble that this observation also involves the gut microbiota. Namely,fatty acids generally possess antimicrobial effects (Kabara et al.,1972), and HFD is known to reduce gut microbiota diversity in mice(Hildebrandt et al., 2009). Similarly, a recent study that utilisedantibiotic treatment to induce gut microbial depletion found aconsequent increase in depressive-like behaviour associated withaltered hippocampal Gr and Crh-r1 transcript levels (Hoban et al.,2016). We also detected lower Bdnf, Traak and Trek2 mRNA levelsin animals on HFD. The latter two have been associated with pro-
tection against glutamatergic excitotoxicity (Lauritzen et al., 2000),a phenomenon that has been associated with depression. Althoughthese marked diet-induced changes in gene expressions were not
uroen
dso
bkrtttafuTtwnbaltt
iefic(tinipd
iepstphdeiirdrsec
rtaFncnhm
sai
A. Abildgaard et al. / Psychone
irectly mirrored in the level of depressive-like behaviour, it istill likely that animals on HFD are more susceptible to episodesf intercurrent stress.
The plasma metabolomic profile was only modestly affectedy probiotic treatment. Tryptophan and its metabolites, includingynurenine pathway metabolites, are believed to play an importantole in MDD, and two microbial tryptophan metabolites were foundo be upregulated by probiotics. One of these, IPA, has been showno possess neuroprotective properties and restrain CNS inflamma-ion (Hwang et al., 2009; Rothhammer et al., 2016). In addition,
recent study demonstrated that IPA improved intestinal barrierunction via the gut endothelial PXR receptor and, in turn, downreg-lated Toll-like receptor 4 (TLR4) signalling (Venkatesh et al., 2014).hus, IPA could potentially mediate some of the immunomodula-ory effects of the probiotics. Another important metabolite, 4-EPS,as lowered by probiotics in our study although the change did
ot pass FDR correction. Interestingly, 4-EPS was recently found toe responsible for some of the aberrant behaviours associated withn autism-like phenotype in mice, and the level of 4-EPS was simi-arly reduced by probiotic treatment (Hsiao et al., 2013). Our resultsherefore urge for intensified research into these metabolites andheir potential role in MDD.
HFD markedly affected the plasma metabolome, but nonteractions between diet and probiotic treatment were seen. Inter-stingly, HFD increased the level of plasma quinolinic acid, and thisnding is in line with a previous study in SD rats and is likelyaused by an increased need for NAD for fatty acid metabolismShibata and Murata, 1985). Interestingly, it has been hypothesisedhat microglia-derived quinolinic acid may hold a neurotoxic effectn MDD (Reus et al., 2015). However, since quinolinic acid doesot pass the blood-brain-barrier (Fukui et al., 1991), further stud-
es are needed to evaluate any cerebral implications of the alterederipheral tryptophan metabolism. Importantly, HFD did not affectepressive-like behaviour.
High-fat diet led to a clear metabolic stress as indicated byncreased body weight, caloric intake, glucose and insulin lev-ls. Although probiotics have been reported to improve metabolicarameters (Razmpoosh et al., 2016), these measures were notignificantly affected in the present study, and this may be dueo differences in probiotic species. Furthermore, no reductions inlasma LPS by probiotics were detected in our animals. Yet, LPSas been suggested to activate especially monocytes and induceepressive-like symptoms (Reichenberg et al., 2001). Unexpect-dly, we found that HFD improved memory in the BM, and thiss in obvious conflict with previous literature. The diet-inducedmprovement was, however, only seen on day 4 and in theecall trial. Since an aversive environment was utilised to expe-ite completion of the trials, an alternative explanation could beeduced fear extinction in rats on HFD. Interestingly, a recent studyuggested that increased CRH signalling may lead to impairedxtinction (Abiri et al., 2014), but more research is needed to con-lude on this topic.
A certain limitation of our study is related to the age of theats since only young adult rats were included. It is well-describedhat the gut microbiota changes with age (Claesson et al., 2011),nd the effect of probiotics may thus be different in older rats.urthermore, we examined the effect of probiotics in SD rats andot in a depression-related disease model. However, clinically effi-ient antidepressants traditionally exert an effect in the FST inon-diseased laboratory rats (Detke et al., 1995), and it may beypothesised that the effect size would be even larger in a diseaseodel owing to a reduced floor effect.
In conclusion, the multi-species probiotics used in the presenttudy clearly lowered depressive-like behaviour, and HFD did notffect the effectiveness of the treatment. The latter point is of greatmportance given the high metabolic comorbidity in depression,
docrinology 79 (2017) 40–48 47
and it further strengthens the generalisability of our findings. Onthe molecular level, probiotics were shown to interact with patho-physiological mechanisms believed to play an important role inMDD, including the immune system, hippocampal HPA axis regu-lation and microbial tryptophan metabolism. Our study thereforeconfirms probiotics as a promising candidate for the treatment ofdepression. Now we just need the clinical trials.
Financial disclosures
Gregers Wegener declares having received lecture fees from H.Lundbeck A/S, Servier SA, Astra Zeneca AB, Eli Lilly A/S, Sun PharmaPty Ltd, and Pfizer Inc. All other authors report no potential conflictsof interest.
Contributors
All authors contributed to formation of study concept/designand selection of methods. AA and BE contributed to acquisitionof data. AA performed data analysis and drafted the manuscript.All authors critically reviewed content and approved the finalmanuscript.
Acknowledgements
This work was supported by donations from “SlagtermesterWörzners Mindelegat”, Vejle, Denmark; “Augustinus Fonden”,Copenhagen, Denmark; “Aase & Ejnar Danielsens Fond”, Lyngby,Denmark; “Brdr. Hartmanns Fond”, Copenhagen, Denmark; “Vil-helm Pedersen og Hustrus Mindelegat”, Slagelse, Denmark and “Th.Maigaards Eftf. Fru Lily Benthine Lunds Fond af 1.6.1978”, Copen-hagen, Denmark.
The probiotics used were kindly donated by Winclove ProbioticsBV, The Netherlands.
Kirsten M. Abildgaard is acknowledged for thorough proofread-ing.
Finally, we thank Birgitte H. Mumm and Per F. Mikkelsen forskilful technical assistance.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.psyneuen.2017.02.014.
References
Abildgaard, A., Lund, S., Hougaard, K.S., 2014. Chronic high-fat diet increases acuteneuroendocrine stress response independently of prenatal dexamethasonetreatment in male rats. Acta Neuropsychiatr. 26, 8–18.
Abiri, D., Douglas, C.E., Calakos, K.C., Barbayannis, G., Roberts, A., Bauer, E.P., 2014.Fear extinction learning can be impaired or enhanced by modulation of theCRF system in the basolateral nucleus of the amygdala. Behav. Brain Res. 271,234–239.
Albenberg, L.G., Wu, G.D., 2014. Diet and the intestinal microbiome: associations,functions, and implications for health and disease. Gastroenterology 146,1564–1572.
Arseneault-Breard, J., Rondeau, I., Gilbert, K., Girard, S.A., Tompkins, T.A., Godbout,R., Rousseau, G., 2012. Combination of Lactobacillus helveticus R0052 andBifidobacterium longum R0175 reduces post-myocardial infarction depressionsymptoms and restores intestinal permeability in a rat model. Br. J. Nutr. 107,1793–1799.
Benjamini, Y., Hochberg, Y., 1995. Controlling the false discovery rate – a practicaland powerful approach to multiple testing. J. R. Stat. Soc. B Met. 57, 289–300.
Bonefeld, B.E., Elfving, B., Wegener, G., 2008. Reference genes for normalization: a
study of rat brain tissue. Synapse 62, 302–309.Bravo, J.A., Forsythe, P., Chew, M.V., Escaravage, E., Savignac, H.M., Dinan, T.G.,Bienenstock, J., Cryan, J.F., 2011. Ingestion of Lactobacillus strain regulatesemotional behavior and central GABA receptor expression in a mouse via thevagus nerve. Proc. Natl. Acad. Sci. U. S. A. 108, 16050–16055.

4 uroen
C
C
C
D
D
D
D
D
D
D
E
F
H
H
H
H
H
J
J
K
K
K
L
L
541–547.
8 A. Abildgaard et al. / Psychone
ampbell, S., Macqueen, G., 2004. The role of the hippocampus in thepathophysiology of major depression. J. Psychiatry Neurosci. 29, 417–426.
hapman, C.M., Gibson, G.R., Rowland, I., 2011. Health benefits of probiotics: aremixtures more effective than single strains? Eur. J. Nutr. 50, 1–17.
laesson, M.J., Cusack, S., O’Sullivan, O., Greene-Diniz, R., de Weerd, H., Flannery, E.,Marchesi, J.R., Falush, D., Dinan, T., Fitzgerald, G., Stanton, C., van Sinderen, D.,O’Connor, M., Harnedy, N., O’Connor, K., Henry, C., O’Mahony, D., Fitzgerald,A.P., Shanahan, F., Twomey, C., Hill, C., Ross, R.P., O’Toole, P.W., 2011.Composition, variability, and temporal stability of the intestinal microbiota ofthe elderly. Proc. Natl. Acad. Sci. U. S. A. 108 (Suppl. 1), 4586–4591.
ale, E., Bang-Andersen, B., Sanchez, C., 2015. Emerging mechanisms andtreatments for depression beyond SSRIs and SNRIs. Biochem. Pharmacol. 95,81–97.
avid, L.A., Maurice, C.F., Carmody, R.N., Gootenberg, D.B., Button, J.E., Wolfe, B.E.,Ling, A.V., Devlin, A.S., Varma, Y., Fischbach, M.A., Biddinger, S.B., Dutton, R.J.,Turnbaugh, P.J., 2014. Diet rapidly and reproducibly alters the human gutmicrobiome. Nature 505, 559–563.
esbonnet, L., Garrett, L., Clarke, G., Bienenstock, J., Dinan, T.G., 2008. The probioticBifidobacteria infantis: an assessment of potential antidepressant properties inthe rat. J. Psychiatr. Res. 43, 164–174.
esbonnet, L., Garrett, L., Clarke, G., Kiely, B., Cryan, J.F., Dinan, T.G., 2010. Effects ofthe probiotic Bifidobacterium infantis in the maternal separation model ofdepression. Neuroscience 170, 1179–1188.
etke, M.J., Rickels, M., Lucki, I., 1995. Active behaviors in the rat forced swimmingtest differentially produced by serotonergic and noradrenergicantidepressants. Psychopharmacology (Berl.) 121, 66–72.
inan, T.G., Stanton, C., Cryan, J.F., 2013. Psychobiotics: a novel class ofpsychotropic. Biol. Psychiatry 74, 720–726.
owlati, Y., Herrmann, N., Swardfager, W., Liu, H., Sham, L., Reim, E.K., Lanctot, K.L.,2010. A meta-analysis of cytokines in major depression. Biol. Psychiatry 67,446–457.
lfving, B., Bonefeld, B.E., Rosenberg, R., Wegener, G., 2008. Differential expressionof synaptic vesicle proteins after repeated electroconvulsive seizures in ratfrontal cortex and hippocampus. Synapse 62, 662–670.
ukui, S., Schwarcz, R., Rapoport, S.I., Takada, Y., Smith, Q.R., 1991. Blood-brainbarrier transport of kynurenines: implications for brain synthesis andmetabolism. J. Neurochem. 56, 2007–2017.
ildebrandt, M.A., Hoffmann, C., Sherrill-Mix, S.A., Keilbaugh, S.A., Hamady, M.,Chen, Y.Y., Knight, R., Ahima, R.S., Bushman, F., Wu, G.D., 2009. High-fat dietdetermines the composition of the murine gut microbiome independently ofobesity. Gastroenterology 137, 1716–1724, e1711–1712.
ill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Morelli, L.,Canani, R.B., Flint, H.J., Salminen, S., Calder, P.C., Sanders, M.E., 2014. Expertconsensus document: the International Scientific Association for Probioticsand Prebiotics consensus statement on the scope and appropriate use of theterm probiotic. Nat. Rev. Gastroenterol. Hepatol. 11, 506–514.
oban, A.E., Moloney, R.D., Golubeva, A.V., McVey Neufeld, K.A., O’Sullivan, O.,Patterson, E., Stanton, C., Dinan, T.G., Clarke, G., Cryan, J.F., 2016. Behaviouraland neurochemical consequences of chronic gut microbiota depletion duringadulthood in the rat. Neuroscience 339, 463–477.
siao, E.Y., McBride, S.W., Hsien, S., Sharon, G., Hyde, E.R., McCue, T., Codelli, J.A.,Chow, J., Reisman, S.E., Petrosino, J.F., Patterson, P.H., Mazmanian, S.K., 2013.Microbiota modulate behavioral and physiological abnormalities associatedwith neurodevelopmental disorders. Cell 155, 1451–1463.
wang, I.K., Yoo, K.Y., Li, H., Park, O.K., Lee, C.H., Choi, J.H., Jeong, Y.G., Lee, Y.L., Kim,Y.M., Kwon, Y.G., Won, M.H., 2009. Indole-3-propionic acid attenuatesneuronal damage and oxidative stress in the ischemic hippocampus. J.Neurosci. Res. 87, 2126–2137.
acka, F.N., Kremer, P.J., Berk, M., de Silva-Sanigorski, A.M., Moodie, M., Leslie, E.R.,Pasco, J.A., Swinburn, B.A., 2011. A prospective study of diet quality and mentalhealth in adolescents. PLoS One 6, e24805.
iang, H., Ling, Z., Zhang, Y., Mao, H., Ma, Z., Yin, Y., Wang, W., Tang, W., Tan, Z., Shi,J., Li, L., Ruan, B., 2015. Altered fecal microbiota composition in patients withmajor depressive disorder. Brain Behav. Immun. 48, 186–194.
abara, J.J., Swieczkowski, D.M., Conley, A.J., Truant, J.P., 1972. Fatty acids andderivatives as antimicrobial agents. Antimicrob. Agents Chemother. 2, 23–28.
elly, J.R., Borre, Y., O’Brien, C., Patterson, E., El Aidy, S., Deane, J., Kennedy, P.J.,Beers, S., Scott, K., Moloney, G., Hoban, A.E., Scott, L., Fitzgerald, P., Ross, P.,Stanton, C., Clarke, G., Cryan, J.F., Dinan, T.G., 2016. Transferring the blues:depression-associated gut microbiota induces neurobehavioural changes inthe rat. J. Psychiatr. Res. 82, 109–118.
ipnis, J., 2016. Multifaceted interactions between adaptive immunity and thecentral nervous system. Science 353, 766–771.
auritzen, I., Blondeau, N., Heurteaux, C., Widmann, C., Romey, G., Lazdunski, M.,2000. Polyunsaturated fatty acids are potent neuroprotectors. EMBO J. 19,
1784–1793.e Port, A., Gueguen, A., Kesse-Guyot, E., Melchior, M., Lemogne, C., Nabi, H.,Goldberg, M., Zins, M., Czernichow, S., 2012. Association between dietarypatterns and depressive symptoms over time: a 10-year follow-up study of theGAZEL cohort. PLoS One 7, e51593.
docrinology 79 (2017) 40–48
Liang, S., Wang, T., Hu, X., Luo, J., Li, W., Wu, X., Duan, Y., Jin, F., 2015.Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive,and biochemical aberrations caused by chronic restraint stress. Neuroscience310, 561–577.
Liu, Y.W., Liu, W.H., Wu, C.C., Juan, Y.C., Wu, Y.C., Tsai, H.P., Wang, S., Tsai, Y.C.,2016. Psychotropic effects of Lactobacillus plantarum PS128 in earlylife-stressed and naive adult mice. Brain Res. 1631, 1–12.
Luppino, F.S., de Wit, L.M., Bouvy, P.F., Stijnen, T., Cuijpers, P., Penninx, B.W.,Zitman, F.G., 2010. Overweight, obesity, and depression: a systematic reviewand meta-analysis of longitudinal studies. Arch. Gen. Psychiatry 67, 220–229.
Messaoudi, M., Lalonde, R., Violle, N., Javelot, H., Desor, D., Nejdi, A., Bisson, J.F.,Rougeot, C., Pichelin, M., Cazaubiel, M., Cazaubiel, J.M., 2011. Assessment ofpsychotropic-like properties of a probiotic formulation (Lactobacillushelveticus R0052 and Bifidobacterium longum R0175) in rats and humansubjects. Br. J. Nutr. 105, 755–764.
Naseribafrouei, A., Hestad, K., Avershina, E., Sekelja, M., Linlokken, A., Wilson, R.,Rudi, K., 2014. Correlation between the human fecal microbiota anddepression. Neurogastroenterol. Motil. 26, 1155–1162.
Pariante, C.M., Lightman, S.L., 2008. The HPA axis in major depression: classicaltheories and new developments. Trends Neurosci. 31, 464–468.
Razmpoosh, E., Javadi, M., Ejtahed, H.S., Mirmiran, P., 2016. Probiotics as beneficialagents in the management of diabetes mellitus: a systematic review. DiabetesMetab. Res. Rev. 32, 143–168.
Reichenberg, A., Yirmiya, R., Schuld, A., Kraus, T., Haack, M., Morag, A., Pollmacher,T., 2001. Cytokine-associated emotional and cognitive disturbances in humans.Arch. Gen. Psychiatry 58, 445–452.
Reus, G.Z., Jansen, K., Titus, S., Carvalho, A.F., Gabbay, V., Quevedo, J., 2015.Kynurenine pathway dysfunction in the pathophysiology and treatment ofdepression: evidences from animal and human studies. J. Psychiatr. Res. 68,316–328.
Rotella, F., Mannucci, E., 2013. Depression as a risk factor for diabetes: ameta-analysis of longitudinal studies. J. Clin. Psychiatry 74, 31–37.
Rothhammer, V., Mascanfroni, I.D., Bunse, L., Takenaka, M.C., Kenison, J.E., Mayo, L.,Chao, C.C., Patel, B., Yan, R., Blain, M., Alvarez, J.I., Kebir, H., Anandasabapathy,N., Izquierdo, G., Jung, S., Obholzer, N., Pochet, N., Clish, C.B., Prinz, M., Prat, A.,Antel, J., Quintana, F.J., 2016. Type I interferons and microbial metabolites oftryptophan modulate astrocyte activity and central nervous systeminflammation via the aryl hydrocarbon receptor. Nat. Med. 22, 586–597.
Sampson, T.R., Mazmanian, S.K., 2015. Control of brain development, function, andbehavior by the microbiome. Cell Host Microbe 17, 565–576.
Savignac, H.M., Kiely, B., Dinan, T.G., Cryan, J.F., 2014. Bifidobacteria exertstrain-specific effects on stress-related behavior and physiology in BALB/cmice. Neurogastroenterol. Motil. 26, 1615–1627.
Shibata, K., Murata, K., 1985. Comparison of the activity of the tryptophan-nadpathway between rats fed a fat-free diet and a fat diet. Agric. Biol. Chem. 49,2899–2904.
Spiga, F., Walker, J.J., Terry, J.R., Lightman, S.L., 2014. HPA axis-rhythms. Compr.Physiol. 4, 1273–1298.
Steenbergen, L., Sellaro, R., van Hemert, S., Bosch, J.A., Colzato, L.S., 2015. Arandomized controlled trial to test the effect of multispecies probiotics oncognitive reactivity to sad mood. Brain Behav. Immun. 48, 258–264.
Sudo, N., Chida, Y., Aiba, Y., Sonoda, J., Oyama, N., Yu, X.N., Kubo, C., Koga, Y., 2004.Postnatal microbial colonization programs the hypothalamic-pituitary-adrenalsystem for stress response in mice. J. Physiol. 558, 263–275.
Tannenbaum, B.M., Brindley, D.N., Tannenbaum, G.S., Dallman, M.F., McArthur,M.D., Meaney, M.J., 1997. High-fat feeding alters both basal and stress-inducedhypothalamic-pituitary-adrenal activity in the rat. Am. J. Physiol. 273,E1168–1177.
Venkatesh, M., Mukherjee, S., Wang, H., Li, H., Sun, K., Benechet, A.P., Qiu, Z., Maher,L., Redinbo, M.R., Phillips, R.S., Fleet, J.C., Kortagere, S., Mukherjee, P., Fasano,A., Le Ven, J., Nicholson, J.K., Dumas, M.E., Khanna, K.M., Mani, S., 2014.Symbiotic bacterial metabolites regulate gastrointestinal barrier function viathe xenobiotic sensor PXR and Toll-like receptor 4. Immunity 41, 296–310.
Wikoff, W.R., Anfora, A.T., Liu, J., Schultz, P.G., Lesley, S.A., Peters, E.C., Siuzdak, G.,2009. Metabolomics analysis reveals large effects of gut microflora onmammalian blood metabolites. Proc. Natl. Acad. Sci. U. S. A. 106, 3698–3703.
Zheng, P., Zeng, B., Zhou, C., Liu, M., Fang, Z., Xu, X., Zeng, L., Chen, J., Fan, S., Du, X.,Zhang, X., Yang, D., Yang, Y., Meng, H., Li, W., Melgiri, N.D., Licinio, J., Wei, H.,Xie, P., 2016. Gut microbiome remodeling induces depressive-like behaviorsthrough a pathway mediated by the host’s metabolism. Mol. Psychiatry 21,786–796.
Zhou, X., Fragala, M.S., McElhaney, J.E., Kuchel, G.A., 2010. Conceptual andmethodological issues relevant to cytokine and inflammatory markermeasurements in clinical research. Curr. Opin. Clin. Nutr. Metab. Care 13,
Zorrilla, E.P., Luborsky, L., McKay, J.R., Rosenthal, R., Houldin, A., Tax, A., McCorkle,R., Seligman, D.A., Schmidt, K., 2001. The relationship of depression andstressors to immunological assays: a meta-analytic review. Brain Behav.Immun. 15, 199–226.