principles ofdesign offluid transport i, systems in zoology i · environment. the arebiteeture...
TRANSCRIPT

MICHAEL LABARBERA
I,
t
Why Must the Fluid Flow?
The m2-~[. basic transport pr~e~s J!1_.bio}QGY ts ~io1h.themovement of molecules from regions of high concentration .regions of low concentration resulting from thermally driven rotdom motion (7). On a cellular level, all transport processes ultimaIy involve diffUsion. On small spatial scales, diffusion is fa u,reliable, and cheap. If a cell or organism merely binds or chemi~ \'i
transforms a molecule available in its environment, more mole::will be delivered by random thcnnal morion. The relevant asdiffusion are given by Fi<;k'sfuuaw...;
dSld, = - D A dCldx
where dSldt is the rate of transport, A is the area through \V
diffusion occurs, dCldx is the concentration gradient, and D isdiffusion coefficient of the substance.
The direct dependence of diffusion rates on the area throwhich diffusion occurs and the inverse dependence of these ratdistance: imply that m:gani~J9.LP_~.9t9.!~is~)-!h!tdeon diffusion for exchange must be small or fiat (to minimizediffusion dlsrance) and 'shoulQ hive high surface-to-volume--- -"- -- .---
Ifluid plays In the biology of the organism or by the nature of tocirculation (that is, whether the fluid passes through the: system onlonce or is continually recirculated).
Transport Systems in BiologyOrganisms transport fluids for two analogous but distinct r~
sons. The most common form of fluid transport SYStCffi) anchange system) is found in the vast majority of mulricdluJar pl~and animals; this system sideste s the seve.re constraints that~
1fQ.P-!C dis~'!!1cCS place on the utility of diffusion. Such syst~
include c1assic~ circulatory systems, the uiiicrrrcffional and ~~convective systems that move fluids to the respiratory srruaure4cmetazoans, xylem dements in tr3cheophyres, and the body cavitfrof a variety of coclomate and pseudocoelomate animals. tj
The second functional class of fluid transport system, a tropHisystem, is restricted to suspension-feeding animals, organismsobtain nutrients by capruring organic particles that are in suspcnSt~
in a fluid. Trophic fluid transport systems are rare only in comp;t50n to the rear universality of exchange fluid transport systems.example, sponges, dams) brachiopods) a broad diversity of~
. pods, ascidians) many fishes, and baleen whales all transport 8we'through partS of their body, extracting suspended nutrients!.support their metabolism and growth.
Principles of Design of Fluid TransportSystems in Zoology
T HE DIVERSITY OF FORM:S IN THE BIOLOGICAL WORLD IS
dazzling, but there are limits to what m.~y be molded by theevolutionary process. Some of these limitations [intrinsic
constraints (1)] arc inhc:-cm [Q biological systems: the limits to thema..ximum forces animals can produce (arising from the maximumStress that an actin-myosin-bascd force-producing organ can generate) or developmencal consrraincs (2, J) which limit the di\'crsity offorms that evolution can produce from a given ta.'<on. Exrrinsicconstraints (1) originate from the physical and mathematical structure of the universe in which organisms evolve (4). EX3mpies range:from the dependence of the strength of materials on the characteristics of molecular bonds to the manifold effects a change in size canexert on the biology of an organism (5) some arising from strictlygeometrical considerations and others arising from changes in therelative magnirudes of physical forces such as surface tension,gravity, and viscosity (6).
To maintain their functional integrity) organisms must continually e.xchange materials with their environments and move materialswithin themselves. Such movement is almost imrariably mediated bythe convection of a fluid. In this article) I discuss the broadlimitations that physical laws place on the structure of the systemsl;." '~ch transport mese fluids within organisms; external flows are aseparate and much more complex topic. The advantage ofapproaching this subject from a physical perspective is that it allows one to
perceive and understand common elements (that is, "design principles") in the structure of organisms. The architecture of mcsesystems is not strongly influenced by the functional role mat the
The author is :IJ1 associate profc:ssor in the Dep:lrtlllcnt of Orgmismal Biology andAruromy at the Uniw;:-sil)' of Chieago, Chicago, IL 60637.
Fluid transport systems mediate the transfer of materialsboth within an organism and between an organism and itsenvironment. The arebiteeture offluid transport systemsis determined by the small distances over which transferprocesses are effective and by hydrodynamic and energeticconstraints. All fluid transport systems within organismsexhibit one of two geometries, a simple tube interruptedby a planar transfer region or a branched network ofvessels linking widely distributed transfer regions; each isdetermined by different morphogenetic processes. 1!LexploitiQ~the signal inIlc;r<;.IlL"-.t !9.<;al..~ear ~.l!~1..Q..n th~
vessel ~all~,-'!.'!LT\1als .h.ave. rel'.eatedly..£~oly"d.?_ ~~plex\fran~g ~erarchy of vessels approximating a gl()b_~
\ \ ",primal system that minimizes the costs of the constnl£don ~dmaintenanceof thdluid transport systc;tp.
SCIENCE, VOL.

,co maimi~ the ratio of the area available for diffusion to thedemand for materials). SmalJ organisms or partS of organisms arcdll:S well suitrd to use diffUsion as a transport mechanism. However,with an ir.:creasc in either organismal size or spatial scale.• thesimarion beeomes less favorable. Exchange transport systems haveevolv~ to circwnvent this problem.
The s,ame problem arises for suspension fceding animals. Theimperative for the maintenance of a fow of fluid past a suspensionfeeder's filtering structure is obvious: the vast majority of thebi9m;US suspended in the oceans lies in the lower portion of the sizes~etrum of particles (8), anc~ such particles have only limitedm6ti.lity. Many suspension feeders actively pump fluids past theirtil&ring structures; in most of these animals, the vulnerable filtersare hidden in thejr bodies, and fluids arc moved to and away from'the filters in trophic fluid transport systems, Particle capture insuspension feeders almOSt invariably involves direct contact betweenthe particle and an clement in the animal's filter; capmre distancesarc on the order of the particle radius, 10° to 102 IJ.m (9).
Despite the obvious differences in the central biological processes,the common constraint that links exchange and trophic fluidtransport systems is the small physical distance over which thesignificam biological phenomenon is effective.
Pn'lItiple 1: When a biologically relevant physical transfer processis limited to small spatial scales and local depletion of a necessaryresource is threatened, bulk flow of a fluid will be utilized for long-distance transport of that resource. ...
The Architecture of Fluid Transport Systems~What charaaeristics shouJd such fluid transport systems have?"First, to avoid locaJ exhaustion of the resource, the fluid muSt<;ircuJate (maintain dCldx or supply particles). The pattern ofc}Jculation is not tightly constrained; aJthough all recirculatingtran:iport systems are exchange systems and all trophic transportsyst~·are r,hrough·flow systems, the converses of these statementsarc not true and no other simple generalities hold.
second, distances must be minimized in the exchange sites (tokeep the concentration gradients high or to aJlow particle caprure).
TklbJe 1. Systemic circulation of a 13-kg dog. Radii and lengths arc average,values; the area given is the aggregate cross-sectional area of the vessels. Notethe increase in cross-sectional area and decrease in velocity in the rransferregion (the capillaries). The capillaries arc both small in radius and thin-walled; although the capillaries represent only about 0.07% of me length ofthe circuit. the blood spends about 4% of the circulation time there. ADumber of categories have been omined here from the original tabulation(90); therefore., circulation times do not a.dd to 100% (91).
Veloc- Tota.l
Vessel RadiusNumber Length Area iry . circu-
(on) (em) (em') (em latioo
j s-') time (%)
'Aom 0.5 1 40 0.8 50 2~ar- 0.15 40 20 3.0 13.4 4
tenesTerminal 2.5 x 10-3 1 X 106 0.1 19.6 2 0.1
arteriesAnerioles 1 x 10-3 4 X 107 0.2 125 0,32 2Capillaries 4 x 10-4 1.2 X 109 0.1 600 0.07 4Venules 1.5 X 10-3 8 X 107 0.2 570 0.07 8Terminal 6.5 x 10-3 1 X 106 0.1 132 0.3 1
veinsMain veins 0.12 600 10 27 1.48 19Large veins 0.3 40 20 11 3.6 15Vena cava 0.625 1 40 1.2 33.4 3
,}l AUGUST 1990
[bi.
Table 2. Circulatory s)'stem of Homo sapitrls (92). Numbers of \'essels werecalculated from average radii and aggregate areas ·given. Note the samefeatures as in Tabl: i.
Radius Am WallVessel
(em)Number
(em')thickness
(ern)
Aorta 1.25 I 4.5 0.2Arteries 0.2 159 20 0.1Arterioles 1.5 x 10- 3 5.7 X 107 400 2 X 10- 3
CapiUarics 3 x 10-4 1.6 X 1010 4500 1 x 1O-~
Venulcs 1 x 10-3 1.3 x 10' 4000 2 X 10-4
Veins 0.25 200 40 0.05Vena cava 1.5 1 18 0.15
This requirement implies that the ~c:~~ls car!y'ing tl}sfl!:1i~.?~~s!.Qe
s01all_(10), tlO\~~<;£"'J).Qiseuille's Jaw (6) implies that any sucharrang~.!TI~nt _~U!~_~_~eD:_I:!!g~..~OS!. For steady'flow in a-·crrcular·pipe
Q = 7rdPr'18Lfl.
where Q is the volumetric Aow rate, dPIL is the pressure drop perunit length of pipe, IJ. is the dynamic viscosity of the fluid, and r isthe pipe radius. This equation is strictly applicable only in fully~~~..l.aJ.I.)jJ1ar flo., [that is, Re},"olds nwnber (Re) (11)< =2000 and ten diameters downstream ofan entrance or branch],but it will serve as a useful guide. The implied restriction is severe.For constant Q, resistance to flow is proportional to r -4; for aconstant flow velocity, resistance is proportional ~o r - 2. In bothcases, cost is direct1}' proportional to the length of pipe.
Pritlciple 2: Energetic considerations dictate that fluid transportsystems utilize both large and small vessels; small vessels occur atexchange sites and arc short, whereas all long-distance transportoccurs in large vessels.
Although this might seem a straightforward criterion to imple·ment. difficulties arise. Recall that diffusion occurs by the randommotion of molecules; it takes a finite time to achieve equilibrium orany fraction thereof. To ma..xiQ1i~ th~ effi'3-CY_oi.~~ystem. the.fI_uid.!~~~da significant fraction of the cirC::~!~Ei9E.time at the~~M~Implementation of principle 2 in its simpiest fonn,narrowing a large vessel down to a short, small vessel at theexchange site, would produce exaccly the opposite result. Becausethe volume of fluid that enters the large vessel must also exit thesmall onc, the lower cross·seruonal area in the small vessel impliesthat the flui9_!!1ust incrc3§C irs speed. The fluid then spends the leastrime in the small vessel, because of both the increase in flow speedand the short length of the small vessel. Again, the solution isstraightforward.
Pritlciple 3: In fluid transport systems, the total cross·scaionaJ area_of the ~ma~ yesse!s g~~rly ex~eeds "~at of rJlt;.l~rg~ y~~~~'so-flo~velocities in the small vessels are lower than those in the large vessels.
·The exchange fluid rransiJort systems in both plants ";;d-;.u~meet these three critcria. All systems consist of at least tWO distinasize classes of vessels. Exchange occurs in small vessels, which arcboth short and numerous. The aggregate cross·secrional area of thesmall vessels is always much greater than that of the large vessels; as aconsequence, flow velocities 3re lowest in the smallest vessels.Representative data for mammals are given in Tables 1 and 2; seealso (12).
The same constraints apply to trophic fluid transport systems(Table 3), albeit for slightly differenr reasons. Of the sLx particleeaprure mechanisms used by suspension feeders (9), one (inertialimpaaion) shows an increasing efficiency with increasing fluidvelocity, but this mechanism has such a low efficiency for thc low·
ARTICLES 993
,1! 'II .i
I I
:11r .I J
iII
I,

*Data from (85) for an "average" animal 3.5 an long pumping 1 an3 S-I. The vdocityin the mandc cavity given in (85} is obviously in error; It assumes that the aoss·scaionalarea in the mandc cavity perpendicular to the Bow is equal to the area of the acnidia[sec (86)~ tData from (&7) for a 15-an-~ong.animaI. A v~lumetric ~'! rate of177.7 em s-I was calculated, based on regressIons m (88). The highest vcloabcs occurin the incurrcnt siphon, contrary to the pancm gencrallv seen in bivalves. *Patafrom (89) for:1 0.412-g animal (dry weight). The uansl'cr region is the mucus sheetoverlying the gill bars in the pharynx.
Incurrent
Excurrent
Transfer
region
Incurrent
Site
Species
MollusC4 UMytilus 0.8 4.25 0.16 1.25 0.236 6.25 a
edulis* ~)
Crassostrea 0.85 78.2 2.43 3.48 0.038 1.22 rgigast '1Brachiopoda ~Terebratalia 0.970 11.340 0.293 0.31 0.027 1.03
transversa 1Laqueus 0.545 5.075 0.225 0.13 0.0'14 0.32californianus
ChordataStyela 0.503 25.3 0.196 1.51 0.03 3.87
clava:t:
transfer regions (Fig. 2), the exchange sites are distributed in three- :1dimensional space, typically throughout the body of the organism. ,jThe most familiar examples are the circulatory systems ofanimals. Injterrestrial plants, the exchange sites are the spongy mesophyll cells 1distributed through the body ofthe leaves, themselves distribUb:d in.lthree-dimensional space around the plant. A distributed geometry is 'jrarer in trophic than in exchange fluid transport systems but does ..occur-the choanocyres, which act as both pump and filter in ,sponges, are found in choanocyre chambers scattered throughout ;
t
f':Table 3. Aggregate cross-sectional areas and water velocities in the trophic ;fluid transport system of a 100-f,L1 segment of the sponge Haliclona pennollis r[abridged from (93)]. The collar slits of the choanocytes (the site of particle :capture) are the transfer region; because: the cross-sectional area is extremelyhigh, the velocity is very low. The reduced cross-sectional area of the Iosculum accelerates the excurrent jet and helps reduce re6luation.---------------------- .J
Area Velocii(mm2) (fJJIl s-tr) :!
-r-;-:-:-~-I:-~-al-a-pertu--r-es-----l-i-i-:~------I~-i-!---., :.•,•.1
ChoanocytesCollar bases 1500 4.9 \C
Collar slits 2400 3.1.jCollar apertures 350 21 i~~~~canal apertures 18~.3 57~~ I :~Osculum 0.14 5.13 x 1<>4 "t
-------------- t~l1
..... ·HTable 4. Cross-sectional areas and velocities in the trophic fluid transpon; t1systems of a variety of suspension-feeding marine invertebrates. "Incurrc:nt"iand "c:xcurrent" are those regions ofthe fluid transport system upstream an~L ~~downstream of the transfer region, respectively. For the mollusks amt1~chordate, these regions are the incurrent and c:xcurrcnt siphons; for the \1..r.
brachiopods, they are the lateral incurrcnt and median excurrcntgapes. Note: Iithe increase in area and decrease in velocity in the transfer regions; in general, l!the fluid leaves the systems with a high velocity. For the mollusks, the area Hgiven for the transfer region is the total area ofostia in the acnidium. For L'te Hbracbiopods, velocities at the lophophore (the transfer region) were calcu1at- lied from (84) on the assumption that stagger of the lophoph9re filaments .yields an area open to the flow equivalent to the projcetc:d area of thelophophore.
density organic particles and low Re typical of particle capture insuspension feeders that it is of minimal importance. Four of theparticle capture mechanisms (13) become less efficient at highervelocities. All five of these capture mechanisms depend on adhesiveinteractions between the particle and filter; higher velocities increasethe probability of particle loss. The efficiency of the remainingmechanism, sieving, is not directly dependent on fluid velocity, butsieving is rarely used by animals (9). There is a mild premium onhigher velocities in the incurrent section of a trophic fluid transportsystem (before the filter) to prevent particle sedimentation on thewalls, and a strong premium on high velocities in the excurrentsection to ensure that the momentum of the water is high enoughwhen it leaves the system to minimize the probability of refiltration.The trophic fluid transport systems of active suspension feeders fitthe paradigm (Table 4); the total cross-sectional area of the filtergreatly exceeds the cross-sectional area of the inflow and outflowpipes.Jor ~jJl tLqp.hi.c_.~st_~_g,~ge~~c!!L~~£QrtsY~~~~tlQ~velocities ilL.th~.Jrnn~f~L{~giQ~__i.mplyJo~~r_£~ts in movinK the.fluid.
The Geometries of Transfer RegionsFurther analysis of the architecture of fluid transport systems
depends on distinguishing between the two fundamentally differenttransfer-region geometries that occur in these systems. Planartransfer regions (Fig. 1), where the exchange sites are restricted to asingle plane, occur in a wide variety of fluid transport systems. Forexample, particle capture in the trophic fluid transport systems ofbivalved mollusks, brachiopods, and cephalochordates occurs on thectenidia, lophophores, and gills, respectively; in all three, thetransfer region is topologically a flat sheet. Respiratory exchange inthe exchange transport systems of teleost fishes, cephalopod andgastropod mollusks, and brachyuran crabs occurs in the gills(termed ctenidia in the mollusks); in these cases also, the topology isthat of a flat sheet.
Extensive folding of the transfer region often partially masks thefundamentally planar morphology. Folding increases the crosssectional area through which the flow passes and thus decreases theaverage velocity of the fluid at the exchange sites. For any given sizeof vessel in the transfer region, this velocity reduction implies alower cost in moving the fluid. This increase in cross-sectional areain the transfer region occurs both in systems with a low-velocityciliary pump (bivalves, brachiopods, ascidians, cephalochordates,and gastropods) and in systems with a high-velocity muscular pumpremote from the transfer region (fishes and cephalopods).
Typically, the planar transfer region partitions the fluid transportsystem into two sections, usually termed incurrent and excurrent,distinguished by whether the fluid has passed the exchange sites.The pipes in such a system rarely branch; the usual morphologyconsists ofa small number (typically one) of ineu:rrent openings anda small number (typically one or two) oflarge vessels leading to theexcurrent opening or openings. All fluid transport systems withplanar transfer regions have some feature that decreases the probability of recycling excurrent water [for example, down-currentrelease of the excurrent stream in fishes and some ascidians (14);constriction of the excurrent opening to accelerate the fluid inbivalves, brachiopods, and cephalopods]. Although all fluid transport systems with planar transfer regions are through-flow systems,not all through-flow systems have planar transfer regions [forexample, the fluid transport systems in sponges (trophic), bird lungs(exchange), and terrestrial plants (exchange) are all through-flowsystems with nonplanar transfer regions].
In the second major geometry of transfer regions, distributed
iff.\\O-,'$J
· .

where T is the radius of the vessel and k is a constant. Mayrovitz andRoy (39) measured flow ratcs and radii in 160 vessels in the ratarteriolar circulation; vcssel radii ranged from 3 to 54 J.1m. The
ARTICLES 995
Fig. 2. A fluid transportsystem with distribll[cdtransfer regiom (black),illustrating the branching hierarchy of the \'cs5<:ls. This highly schematiu:d drawing of thetropbic fluid transportsystem of a spongeshows the incurrem canals (dashed lines) andcxcurrcm canals (soHdlines) that carry watertoward and away from,respectively, the choanoeytc chambers. Flow directions are indicated by arrows.
8
Vessels from pathologic hearts yielded smaller values for the exponent, as did a heterogeneous grouping of other coronary arteries;none of these values wac significantly different from a value of 3(23). Zamir ef al. (24) compared the dimensions ofvcsscls at -500branch points in rat arterial systems to those predicted by Murray'slaw; agreement with theory appears to be good, but statisticalmeasures were not given.
In a variation on this approach, LaBarbera and Boyajian (25)measured vessels (diameters from 89 to 502 ~m) in the astrorhizalsystem of three species of Devonian stromatoporoids (an extinctgroup of sponges). The 3Strorhizac: arc usually interpreted as anetwork of e.xcurrenr canals (26). For each triplet of vessels at adichotomous branch point, an expected parem vessel diameter was
Iests of Murray's Law calculated from the daughter vessels' diameters, on the assumption",:. . that Murray's law was valid. Mean differences bctween observed and
There has been some controversy over the validity of Murray's law I calculated parem vessel diametcrs for 361 branch points in fiveU8, 20, 21), but it appears robust when appropriatc tests are distinct astrorhizal complcxes ranged from 0.6 to 5.8 J.1m (noneapplied. Hutchins et al. (22) measured vessel diamctcrs at 42 branch b. significantly diffcrcm from zero).points in normal human left main coronary arteries; the mcan Murray's law implies that, at any level in the hierarchy ofvcsscls,cxpOnent relating parent and daughter vessel radii was 3.2 (::to.2S). I the sum of the cubed radii should yield a constant value (16, 18).
The data presented here for m~3.lian cir~Jatory systems (Table
5)~ fluid tr~sr<:!!._srs!!=.!J1s.ofrO~T~p.E..cicsOf'S ng~. abiesA 6 through 8) utilize this relation. Except for the dioanoc:yte
~'.'... .i\<\',V,-';JEX'-:::, :)'.>:.. chambethrs an,d aPOthPYlcsc (discussf
e3'd bel01IW), the vessels in spodnges
WI': r; meet IS ru e WI 111 a lactQr 0 , exce em agreement conSI cnng d.-... that the values for vcssel diametcrs and numbers :Irc estimates from l~~
-.. IN .. ',:: «: small samples of tissue. The data for mammalian circulatory systems \~ \ '. ~,... :-:: ):<~ fit the model less well; arterial and venous vessels yield different~_",,_,Ex '" ~:: ... .. vaJues, whereas valucs for the arterioles and capillaries differ marked-
- =-. . . Iy from those of both arterial and venous vessels.This mode of analysis has a weakness, however. Any grouping of
vessels, be it anatomical (the human and dog data in Table 5),functional (the sponge data in Tables 6 through 8), or obtainedfollowing various vessel ranking methods (27-31) (the hamstermuscle in Table 5), necessarily groups vessels of different sizes intothe: same category. Calculation of a mean value for radius weights aUvessels equally, the sum of the number of vessels times the meanradius cubed will equal the sum of the individual cubed radii only ifthe number of vessels is inversely proportionaJ to the radius cubed.Precisely these arbitrary categoriz..1tions have been used in moststudies on the architecture of the bronchial and circulatory systems(32-38).
A more direct test utilizes a rclation implicit in Murray's law (18): IlQ = kr'
Ag. 1. Fluid rransportsystems with a planar geometry ofthe transfer re.gions. Dil'C'ction of flowis indicated by arrows.The. incurrent (IN) andexrurrc,nt (EX) partS ofme systems arc labeled;the ~fer regions arcindic:ued in solid black.(A) Schematic borizon-tal ~on through agenaalizcd rc:lc:ost fish,showing an exchangeBuia transport system.Flow is drivco by musroIar action of the buccalfloot and opercula; thet:nJ}Sfer region.:; arc megills. (8) Schematiccross section througb ageneralized eulameUibranch bivalvc mollusk.Although the rransfer regions (the ctenidia) areinvolved in respiration,their primary function isparticle Qprure:; bence,this is a trophic fluidtransport system. WaterBow in both the incurrcnt and excurrent regions is primarily perpendicuJar tothe plane of the drawing.
rht spOnge. Because the transfer regions in systems with this
giOmetry are widely dispcrsro, the fluid rransport sysrems them·selves must have a more q)mpkx geometry than in systems withplanar transfer regia.ns; in particular, they tend to have a hierarch)' ofsizes qfvcssc.ls, each of which is joined to the whole at branch pointsin qu::·systern. What rules govern the architecrurc ohhis branchinghicraicliy? Assuming that (i) it COSts energy to move the fluid; (ii)laminar, fully developed Poiseuille flow of a I:'lewtOnian fluid occursthroughout the system (Fig. 3A); (iii) there is some COst to buildingor,ry;aintaining the system (the vessels) and irs contents (the fluid);ana (iv) there is some function that maximizes rerurn on the systemfot the costs of construction, maintenance, and operation, Murray(15-17) derivcd the following relation:
Prilldplt. 4 (Murtay's Law): [n an optimally designed systeminvolving bulk laminar flow of a Ncwtonian fluid through pipes, atany branch point the tadius ofthe parent vessel (ro) cubed will equalthe swn of the cubes of the radii of the daughter vessels (r" r2,'3, ... '11):
Murray's original derivation minimized the costs of forcing thefluid (blood) through the pipes (blood vcssels) against the action ofthe fluid's viscosity and the COStS of building and maintaining thesystem (blood and blood vessels). However, the lattcr cost may bereplaced by" any cost associatcd with the volume of the system as a~~olc (18). ~is rdation holds only for laminar flow; for rurbulcml/Bow regimes, the exponent would be 2.33 rather than 3 (1&-20).
.31 AUGUST 1990
~~

exponent of r that they derived was 3.01 (95% confidence inter·val = 2.86 to 3.14). This result was robust to the genotype of therats and the vascular state; the mean exponent of six di.fferenttreaanents was 2.997 (±0.026). Kobari el al. (40), in a similar srudyof the flow in plial arteries (radius 10 to 100 ,.....m) in cats, obtainedan exponenr of 2.98 (41).
The mosreommon approach (20, 21,42,43) to resring the validieyof Murray's law (and alternative optimization models) utilizes thedependence of branching angle at a branch point on the relative sizeof the daughter vessels. A number of alternative fonnulations of therelation are possible; all predict that daughter vessels with smallerradii should make steeper angles to the parent vessel. Analysis ofbranching angles has. in general, shown poor agreement withtheoretical predictions. Zamir et at. (24) measured both diametersand angles of branches at 500 branch points in rat arterial systems;measured angles match theoretical predictions much less well thando diameter measurements from the same branch points. The anglesprediaed for different ratios of parent to daughter vessel cliaroetersby clifferent exponents arc. in general, not greatly different (43)jthus, this approach has not proved very useful. Zamir (44) contoured COSt functions for angles of branching, using minimumpower (minimum total drag) and combined minimwn power and
'j1
volume (Murray's law) models. Using data from a variety ofmammalian arterial systems, Zarnir found that., in all cases, Murray'~law fit the observed angles better than did any alternative fonnulations. More importantly, in virtually all cases the Q\·erwhe1m.ingmajority of the data points fell within the limits of a 5% COst abovethe oprimum (45). 1
Although the available data are not definitive, it would ap~that Murray's law is a useful approximation to the branchinghierarchy of fluid transport systems with distributed transfer rejgions. This agreement is not an accident of geometry. The uacheat;system in insects is superficially similar in architccrurc to the Builttransport systems discussed abovc. but bulk. flow of air is ci~absent or limited to the largest vessels; O2 is delivered to the tissuCJof the insect ~y diffusion. The vessels of ins:.ct ~eh~sysrcrns,follow the ~r- law expected for a diffusion-based sysrcm (46, 47):~~an the ~rJ of Murray's law. Murray's law better dcscritd
-- --. - - _.._--- ~~the fluid rransport system of sponges than the circulatory system 0 .
mammals. The bronchial system of mammals (33) matches less wethan systems that involve the movemcnt of liquids. probably bccaflow in the larger bronchi is rurbulenl (48) and bulk flow decrcases\in importance relative to diffusion in the smaller vessels. In thvessels. the assumptions lUlderlying Murray'S law arc violated.more general explanation for these cliscrepancics will be offerebelow.
·Oat3 from (92); numbers of ve.s.sds calculated from aggregate areas and radiigiven. tData from (90, 94); both attribute the basic data. to Mall (J2). *D.:I.t~from (J7). Vessel order was detennined wing Strahler ordermg; cemnfugal ordermgproduced no dear hierarchy of vessel sizes.
1
~,11
rlHow do biological systems of such wide phylogenetic distribu .
tion and differing functional roles all approach the architecture of ~system of global optimality? What cues are available to organisms to/usc in generating such systems? In a Murray's law system, the sheai{mess (7) on the walls of the vessels (49) is cnnsrant and equJLeverywhere throughour the sysrem (18). If the endothelial ~e~lining blood vessels had a mechanism for measuring local shear srrcs!!and comparing that value [0 some set point. growth or shrinkage ORvessels in which the shear stress departcd from the set point wouJiauromatically produce a sysrem that followed Murray's law (.5C0J!53)-local responses could produce a globally oprimal system. Ii
Evidence that endothelial cells in mammalian circulatory syst31do indced respond to local shear stress has accumulated in recenyears (54); effects include flow·induced membrane K+ currents a7 = 0.02 to 1.70 N m-' (55), protein secretion ar arrcrial sh Istresses (T = 1.5 to 2.5 N m-2) but not at venous shear str(7 = 0.4 N m-') (56), and membrane hyperpolarization that isfunction of local shear stress up to 12.0 N m-, (57). Long.rcm/.acconunodations to surgically altered flow dtrough the commoncarotid anery of dogs have been observed for both increased (up t~fourfold) and decreased (as much as 96%) flow rares (51); over 6 to.S months. changes in the vessels' diameters returned the wall sheJ.~stress to its original value (58). A similar regulation of wall sh~stress occurs in the iliac artery of Macaca fascicularis (59); 6 monarrer surgical intervention increased the blood flow by a faaor of 10the artery had doubled in cliamcter, returning wall shear stress to i :original value (-1.5 N m-'). Langille and O'Donnell (60) demon jstrated that wall shear stresses were regulated through diamerechanges in the common carotid aneries of rabbits. A 70% 8m:reduction yielded a diameter reduction of21% (raising shear sto 61% of their original value) in 2 weeks. Diameter change was nomerely due to wall thickening or chronic contraction of the smoomuscle of the vessel wall and was critically dependent on thpresence: of the endothelium; removing the endothelium abolishethe response (61). Interestingly, the response appeared to be local
The Developmental Mechanism forMurray's Law Systems
1.251.350.75
2.783.333.035.275.09
0.486Q.4000.7682.707.59
10.37
47614441122
18004 x 101
1.2 X 109
8 X 101
1800600
Callis pmitiaristI40600
0.50.150.05
1.82.854.27.613.65
0.30 40 10.800.625 I 1.53
Hmnstu cheek pOllch retractor mlude arurio/ar lIt'tlllorktVessel order
o1234
Vessel Avcrage Number 1,' 1.-' 1,'radius
Homo sapielU1>Aorta 1.25 1 1.56 1.95 2.44Aneries 0.2 159 6.36 1.27 0.25Anerioles 3 x 10-3 1.4 X 107 127.4 0.382 1.15 x 10-)Capillaries 6 x 10-4 3.9 X 109 1432 0.860 5.16 x 10- 04
Venules 2 x 10- 3 3.2 X 108 1273 2.55 5.09 x 10-3
Veins 0.25 200 12.9 3.18 0.80Vena cava 1.5 I 2.25 3.38 5.06
AomLarge aneriesMain arterial
branchesTenninal branches 0.03Anenoles 1 x 10-)Capillaries 4 x 10-'&Venules 1.5 x 10-3
Terminal veins 7.5 x 10-2
Main venous 0.12branches
Large veinsVena cava
Table 5. Exchange fluid transport systems in three mamma1s. For all threespecies, the 1,-3 (which should be constant by Murray's law) has beenca1culated. For Homo sapierls, values for I,.2 (proponiona1 to the area availablefor diffusion or inversc:ly proportional to flow velocity) and 1r4 (inverselyproportional to resistance) are given for comparison. In general, ~,J yic:ldsrelatively constant (but different) values for the artcrial and venous systems,but values depart widely from those of both the artcria1 and venous systcmsin the smaller vessels (arterioles and capillaries). See text for cxplanation. ForHomo and Callis, vessel radii ate in centimcters; the I~ values have beendivided by 0.1 em). For the hamster muscle, vessel radii arc in micrometers;I,J values have becn divided by 1000 IJ.m).
996 SCIENCE. VOL. 24

I'
\
, }
I J., I
'Y.',--:
--:
o
c
B
Fig. 3. Velocity profilesat branch points in vessels. In all cases, Re isassumed to be: weU bc::-low the critical value: forrurbulent flow (=2000);arrows indicate: the: dire:ction offlow. The: Rowin upstream vessels is as·swne:d to be fully devel·oped PoiscuiUe flow.The branching is assumedto be symmetrical; asymmetric:al branching willdiston the: patternsshown but will not affectthe qualitative: relations.Loca.I shear stTeS5 on thevc:ssc:1 wall will be: pro\Xlr·tional to the vc:locity grndiem; the closer the pointof maximum velocity liesto a wall. the higher thelocal shear stress. (A) Theidealized flow pattern im·plicit in the derivation ofMurray's law; in all ves·sds, the: vclocity profile isassumed to be fit1Iy dcvel·oped. The: maximum velocity is fOW'ld at the: center of the vc:sscls, and the:
-----+ velocity profiles arc par:J.ool:lSi the maximum ve:locity is twice: the ave:rage:velocity. (B) Velocityprofiles at branch pointsin large vcssds (100 < Re
< 15(0). Bc:causc the Row divider at the: branch JXlin[ intcrsects the region ofhighest velocity in the parrot vesscl, the vclocity profiles in the: daughter vesselsarc skewed roward the: median sides. (e) Velocity profiles at branch points insmall vc:ssds (Re: - 10- 1 to 10-2). The: influence of the branch divider is fc:ltupstreun so that the: \'docity profiles in the daughter vessels are skewed towardthe: lateral walls. (0) Velocity profiles at a confluence: of vc:sscls. The: profile ofthe: merged flows shows a double: peak with a minimum (but nonzero) velocityon the uis.
A
(distance to reestablish a parabolic velocity profile) is a function ofRe, these effectS should be greater in larger vessels (which have both3. greater radius and Bow velocity). This is the pattc:rn seen in anglesof branching in the human pulmonary artery (radii from 0.84 em ro350 ~m); smaller parem vessels better fit Murray's law than largerparent vessels (21). In small arterioles (radius 10 to 35 ~m), theinverted pattern ofvclocicy profile distortion (70) (Fig. 3C) impliesthat the angle of branching should decrease. When Rows from twovessels merge (the venous system in mammals, the excurrcnr vesselsin sponges, exhalation in the lungs), higher velocities occur near the
ARTICLES 997
Vessel Radius Number ~"TIl'-(,-')
Affe:rent canals 40.6 8.1 x 104 5.43 123Cho:mocyte chambers 27.3 2.25 x 10' 46.5Efferent canals 123.7 5.2 x 103 9.84 68Arrium 2585 1 17.3 39Osculum 1405 1 2.77 242
Table 7. The trophic fluid ttaIlsport system of the sponge Uucalldra aspaa,after (95); vessel de:signations foUow the: original source:. Values for 1,J andT/~ (6-1) are relativc:ly constant; the: choanocyre: chamber value: is explained inthe: tot. Ave:rage: radius was calcu1ate:d from aggregate cross-sectional areasand cite:d numbers of e:ach element; the 1,3 values have: been divided by 10'~m3. Conventions are: as in Table: 6.
·',
regions 9f the arterial wall whcn~ the endothelium was incompletelyrtffiovcd showed local diameter reduction (60).
A mechanism in which local shear stress determines the diameterof the vessel rationalizes the data for human and dog circulatorysystc~ presented in Table 5. First, the shear rate set points for theanerial\;md venous endothelial edls apparently differ (56, 62); themechanism dcrcnnining this difference is llllknown. Second, bloodis a non-Newtonian fluid in which viscosity decreases when localsh1ear rates increase; the effect is most pronoWlccd in vessels withdilnfcters smaller than about 300 ~m (31). Those vessels in which};," departS most from the mean for the system, the arterioles andCapillaries, arc precisely those in which the effective viscosity of theblood should be most reduced. From the aorta through thecapillaries, the wall shear stresses in the human systemic circulationrange only from aOOm 1.04 to 2.6 N m-2 (52); values for the venouscirculation range from 0.14 to 0.63 N m-2 (63).
Approximare values for the shear rate (T/~) (64) in the vosscls oftwo species of sponges (Tables 6 and 7) all lie within a factor of 3for each sponge; here, tOO, vessel diameter may be detennin!=d bythe response ofthe cells lining the vessels to local shear stress. Such aresponse would also account for the extreme discrepancies in the IT)
values for the chemocyte chambers and apopylcs (excurrent openings of the choanocyte chambers) of the sponges in Tables 6 ro 8.Flow in sponges is driven by the choanocytcs' flagella; Bow in thechoanocyte chambers and apopyles is assuredly laminar(Re = 10-'), but it is also assuredly nor the parabolic vdocityprofile assumed in PoiseuiUe flow. In lieu of acruaJ measuremenrs ofthe vdocity profile or local shear Stress. these vessels cannot beincluded in the analysis. Do the cells lining the vessels in the fluid~port system of sponges respond to local shear stress? To date,only the endothelial cells in the vascular system of mammals havepeen investigated for sensitivity to shear stress./A mechanism dependent on local shear Stress clarifies some of the
conFWversies. At every branch point in a system of branching pipes,the ;vdocity. profile will be temporarily altered. Near a branch pointin large vessels. the medial sides of the daughter vessels willexperience higher velocities and the latcral sides lower velocities thanin fully developed flow (Fig. 3B) (65-68). Medial cells shouldrespond as if they were in a vessel tOO small (shear stresses too high)and lateral cells as if they were in a vessel too large (shear Stresses too.Jow); unless the response is averaged around the vesscI [see (69)].. the result would be to alter branching angles to higher values thanpredi~cd from Murray's law. Because entrance or inlet length
Ves,d Radius Number ~"TIl'-
(,-')
Ostia 10.3 3.93 x 10" 0.429 222Inhalant canal aperrures 97.5 1.44 x 102 1.34 70Prosodi 2.5 6.57 x 10' 1.03 92Prosopylcs 1.5 1.26 x lOT 0.425 222Otoanocyre ch:unbc:rs 15.5 1.20 x 106 44.7Apopylcs 7.0 1.17 x 106 4.01 24Exhalant canaJ apertUres 187.5 l1.n 0.776 122Os<:ulum 2100 0.Q1011 0.936 98
" 3I,AUGUST 1990O-
r:".,
Table 6. The troghic fluid tr.ulsport system of the sponge Haliclona prnnollis.Note that the Ir is constant with a factor of 3 for all vessel types exceptme choanocyte chambers and apopyles (cxcurrenr openings from the:choanocyte chambers}:'-Shear rate (,../~), which is proportional to shear srress
Ion the vessel walls, foUows a similar panem (64). The excursion of values inthe choanocyre chambers and apopyles is explained in the text. Data from
}
(9J) for a l00-.,u mature segment. Radii are: mean values (in micrometers);where: means were not given, the midpoint of the range was used. Numbersof elements were calculated from the mean radius and the: total cross·secrional areas wven; the: value: for the:o~~ is the proportional fractional
: pan of the whole:. Ir3 values have been diVided by 108 ~m3.
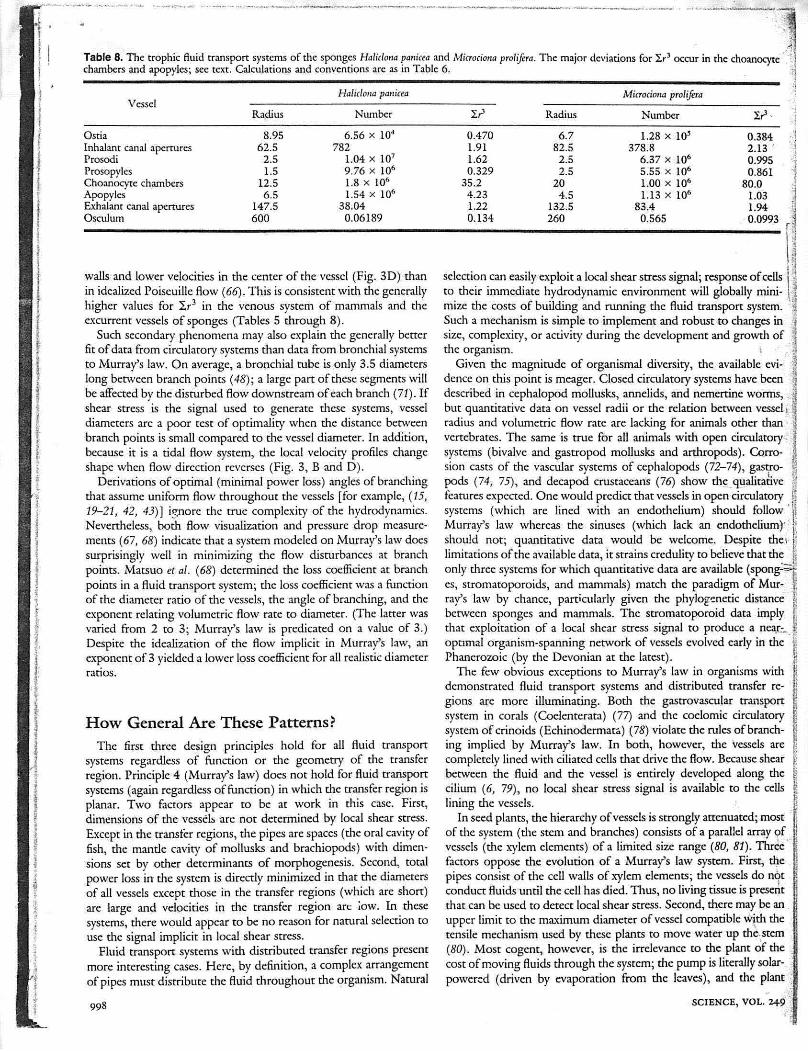
0.3842.13 '0.9950.861
80.01031.94
·00993
Number
1.28 x 10'378.8
6.37 x 106
5.55 X lOt>1.00 X 106
1.13 X 106
83.40.565
Mitrociotlrl proliJfTll
6.782.5
2.52.5
204.5
1325260
Radius
0.4701.911.620.329
35.24.231.220.134
.~;
SCIENCE, VOL. 2+9.~....;.
.,i1.~
'j]
![;
seleCtloncan easily exploit a local shear stress SIgnal;. response ofcells 11to their Immediate hydrodynamiC enVIronment Will globally rrurn- \~lffilZC the costs of building and running the fluid transport system. :~Such a mechanism is simple [0 implement and robust to changes in •size, complexity, or activity during the development and growth ofthe organism. I
Given the magnitude of organismal diversity, the available evi·dcnce on this point is meager. Closed circulatory systems have been ldescribed in cephalopod mollusks) annelids) and nemertine WOmlS) ~
but quantitative data on vessel radii or the relation between vessellIradius and volumetric flow rate arc lacking for animals other than 'vertebrates. 'TI1e same is true for all animals with open cirrulatory~:systems (bivalve and gastropod mollusks and arthropods). Corrosion casts of the vascular systems of cephalopods (72-74) gas~r<r ~
pods (74, 75), and decapod crustaceans (76) show th,_ q~alitative ~fcamres expected. One would predict that vessels in open circulatory ~~
systems (which arc lined with an endothelium) should follow' '11Murray's law whereas the sinuses (which lack an endothelium)": Is.ho,u!d. not; quanti~ative data. wou~d be we.leome. ~espite thel'Ilbmlt300ns of the aV31lable.data, It s~~s creduhty to ~heve that th~_~only three systems for whICh quantitative data arc avaIlable (spong-~l.
es, stromatoporoids, and mammals) match the paradigm of Mur·ray's law by chance, particularly given the phylog-:nctic distance jbetvleen sponges and mammals. The stromatOporoid data imply Iithat exploitation of a local shear stress signal to produce a neaJ':_ U.
opamal organism·spanning nctvlork of vessels evolved carly in the .hPhanerowic (by the Devonian at the latest). M
The few obvious exceptions to Murray's law in organisms with ndemonstr:ued fluid transport systems and distributed transfer re- !
gions arc more illuminating. Both the gastrovascular transportsystem in corals (Coelenterata) (77) and the coelomic cirrulatorysystem of crinoids (Echinodennata) (78) violate the rules of branching implied by Murray's law. In both, however, the vessels arecompletely lined with ciliated cells that drive the flow. Because shearbetween the fluid and the vessel is entirely developed along thecilium (6, 79), no local shear stress signal is available to the cellslining the vessels.
In seed plants, the hierarchy ofvessels is strongly attenuated; mostof the system (the stem and branches) consists of a parallel array C?f ,vessels (the xylem elements) of a limited size range (80, 81). Thr~e'
factOrs oppose the evolution of a Murray's law system. First, ~epipes consist of the cell walls of xylem elements; the vessels do nqtconduct fluids until the cell has died. Thus, no living tissue is prese~t
that can be used to detect local shear stress. Second, there may be an .upper limit to the maximum diameter of vessel compatible with thetensile mechanism used by these plants to move water up the.,stem(80). Most cogent) however, is the irrc:levance to the plant of thecost of moving fluids through the system; d,e pump is literally solarpower.:d (driven by evaporation from the I::aves), and the plant;
Haliclona pmricea
Number
6.56 X 104
7821.04 x 10'9.76 X 106
1.8 X 106
1.54 X 106
38.040.06189
Radius
8.9562.5
2.51.5
12.56.5
147.5600
Vessel
walls and lower velocities in the centcr of the vessel (Fig. 3D) thanin idealized Poiscuille flow (66). This is consistent with the generallyhigher values for I-rJ in the venous system of mammals and thec-xcurrent vessels of sponges (Tables 5 through 8).
Such secondary phenomena may also explain the generally betterfit ofdata from circulatory systems than data from bronchial systemsto Murray's law. On average, a bronchial rube is only 3.5 diameterslong between branch pointS (48); a large part of these segments willbe affected by the disru,bed flow downstream ofeach branch (71). Ifshear stress is the signal used to generate these systcms) vesseldiameters arc a poor test of optimality when the distance betweenbranch points is small compared to the vessel diameter. In addition,because it is a tidal flow system, the local velocity profiles changeshape when flow djrection reverses (Fig. 3, B and D).
Derivations of optimal (minimal power loss) angles of branchingthat assume uniform flow throughout the vessels [for cxanlple, (15,19-21, 42, 43)] ignore the true complexiry of the hydrodynamics.Nevertheless, both flow visualization and pressure drop measurements (67, 68) indicate that a system modeled on Murray's law docssurprisingly well in minimizing the flow dismrbances at branchpoints. Matsuo et al. (68) determined the loss coefficient at branchpoints in a fluid transport system; the loss coefficient was a functionof the diameter ratio of the vessels, the anglc of branching, and theexponent relating volumetric flow rate to diameter. (The latter wasvaried from 2 to 3; Murray's law is predicated on a vahie of 3.)Despite the idealization of the flow implicit in Murray's law, anexponent of 3 yielded a lower loss coefficient for all realistic diameterratios.
The first three design principles hold for all fluid transportsystems regardless of function or the geometry of the transferregion. Principle 4 (Murray's law) does not hold for fluid transportsystems (again regardless of function) in which the transfcr region isplanar. Two factors appear ro be at work in this case. First,dimensions of the vessels are not determined by local shear stress.Except in the transfer regions, the pipes are spaces (the oral cavity offish, the mantle cavity of mollusks and brachiopods) with dimensions set by other determinants of morphogenesis. Second, roralpower loss in the system is directly minimized in that the diametersof all vessels except those in the transfer regions (which arc short)arc large and velocitics in the transfer region art: iow. In thesesystems, there would appear to be no rcason for narural sclecrion touse the signal implicit in local shear stress.
Fluid transport systems with distributed transfer regions presentmore interesting cases. Here, by definition, a complex arrangementof pipes must distribute the fluid throughout the <?rganism. Narural
998
How General Are These Patterns?
OstiaInhalant canal aperturesProsodiProsopylcsChoanocytc chambersApopylc:sExhalant canal apcrturt::sOsculum
....~ _..•.--~.~~-.~-~.__.._---------_.._.~-_.__.._-----_.~_ .._--,---_._-- .
rable 8. The <wph.c flu.d "anspo" systems of the sponges H,/.c/,,,, p""'" and M""""" p"/Oft" The maiO' dcv>auons fo' l,' occu, in the choanocytlchambers and apopylcs; sec text. Calculations and conventions arc as In Table 6. -,1
1.,.,
iii
1<
j
t,
,t
j
11;
I

c:xFGs p" 'mcrabolic energy in driving the flow. Even if a localshe¥ stress signal were: available to the plant, there would be nosdc;crive pressure to exploit it (82)., The widespread presence of systems carrying fluids in laminar
,fio~ that meet the four principles given above is the result of.oonveyfent evolution. The first three rules arc dictated by the shortspatial ~tances over which the transfer processes arc effective, thehigh power requirement involved in forcing fluids through narrowspaces., and the necessity to limit flow vdocities past the transferregions. The solutions to these problems arc evolutionarily simpleanf! s'iraightforward. The problem of arranging a branching hierarch! ofpipes to deliver fluids to distributed rransfer regions appearsmuch more complex. Not only must the appropriate connections be:made (83), but the system must be tuned to a global optimum if theCOSts of building and running the system arc to be: reasonable. Thata simple signal. local shear stress, and local cellular responses to thatsignal can be used to construct a system that approximates minimalenergetic expenditure is fortuitous, given the complex hydrodynamics implicit in such systems, The pathways open to evolution arcindeed constrained, not because only one solution exists but becausea simple local signal that will gen~rally be available to tells in thevicinity of flowing fluids can be so easily utilized.
Comparative studies of fluid transport systems can be a powerfultool in analyzing how natural selection has c-xploitcd this opportunity. In the bronchial system of mammals, the confounding effects ofdiffusion in the lower pathways, turbulent flow in the lar.gcrbronchi, and the change in local velocity profiles during inhalationand exhalation should be "·parable. Srudy of the bronchial system ofsmaller mammals should eliminate the effectS of rurbulent flowbecause Re's are decreased. Quantitative data on the respiratory tree9( holothurians (Echinodermata) should allow isolation of ti,Carects of changing velocity profiles (resulting from tidal flow) fromthe effects of diffusion along the rubes. Whcther fluid transpertsystems simply use the signal of local shear stress or acruallyoptipUze the system to minimize COSts ofconstruction and operationcouJ,d also be determined by comparative srudics. Flow in the largearteries of mammals of greater body size than humans is certainlyturtfulenr; the exponent relating diameter to flow should be 2.33rather than 3 (19) if these systems are truly optimized by naruralselection.
REFERENCES AND NOTES
1. G. V. Lauder, Paltobiology 7, 430 (1981).2. P. Alberch and E. A. Gale, Evolution 39, 8 (1985).3. R..A. ~,DaI, Biol. 119,6 (1987).4. R.~M Aknnder, BioI.J. Linn, Soc, 26, 345 (1985),5. K. ·Schmidr-NicJscn. StllUng: IJl7ry 11 Animal Si.o:t So lmpDl'lIJnt? (Oxford Univ. Pt'l:U,
OOOnl, 1984).6. S, Vogd.Lij in Moving RuitlJ (Will.iud Grant, Bosron, 1981).7. H. C Berg, RJusdom IVIIlh in Biology (PrincClon Univ. Press, Princeton, 1'.1, 1983).8. J. Rodriguez and M... M. Mullin, LimnDl. Owmogr, 31,361 (1986).
,9. M. Wlub=, Am. Z«>I. 24,71 (198<). .ilO. Seconcb:ry considerations ouy reinforce this pattern. In animals thar ust cwa orI flagella [0 drive the Bow (for example, cilia on the respirarory ctenidia of
Igasaopods or me swpcnsion·fccding ctenidia of biv:l1ves), momennun transfer
. bctwec:n me cilia or flagella and the fluid is also a short·range phenomenon limiredI cssc:ntially [Q the length of the dri\'ing organeUe,).1. Reynolds nwnbcr is equal ro prUIp., where p is me density of the fluid, r is the
radius of the vc:ssc.l, U is the flow velocity, and "" is the dynamic \'iscosiry of thefluid. It is equivalenr to the ratio of the inertial [Q \'lscOUS forces in a flow siruuion(6).
U. M. La.B3Ibcn. and S. Vogel, Am. Sci. 70,54 (1982),13. The partide caprun: mechanisms of direct interception, gravitation3.1 deposition,
.. diffusive or motile·particle deposition, md~ md tr.lp c:apture all become lessefficient as IocaJ flow velocities inm:asc. See (9) for :a full descriprion anddiscussion.
H.' C. M. Young and L. F. Braithwaite, BioI. Bull. WoodJ Holt, Man, 159,428 (1980).15. CD. Murray, j. em. Physiof. 9, 835 (1926).16. -----. Proc. Na/I. AClId. Sci. U.S,A, 12,207 (1926).17. "Throughout the rest of rhis discussion I will use the phrase "Mumy's l:al¥" ;IS
. shorthand for an idC3Jizcd model that simultaneously minimizes the cmt of
'31 AUGUST 1990~.
~'"
pumping the fluid :md the tou.! volwne of the s)·stem.18. T. F. Sherman,}. Gm. Physiol. 78,431 (1981).19, H. B. M. Uylings, Bu/l, Ma/h. Bioi, 39, 509 (1977).20. A. Roy and M, J. Woldenberg, ibid. 44, 349 (1982).21. M. J. Woldenberg and K, Horsfield,]. 11rtor, Biel. 104,301 (1983).22. G. M. Hutchins, M. M. Miner, J, K. BoiOlon, Cin. Rts, 38. 572 (1976).23. R. A. Groat IFtd. Am. Sot, Exp. Biol. PrOf. 7,45 (1948) (:abstract)) n::poned an
exponent of 2.6.24. M. ZmUr, S. M. Wrigley, B. L. Langille,). Gm. Physiol. 81, 325 (1983).25. M. UBarber:1 and G, E, Bo)'ajian, in preparation.26. G. E. Boyajian and M:, La.Barber.l, uthlia 20, 223 (1987).27. Srnh.ler ordering preserves the hien.rchy of vcssc:1 sizes bener than centrifugal
ordering, but neither method is justifiable if the objective is to tcst some power lawrelation between levels in the branching hierarchy, whatever the preswnedexponent. Objca.ions to these vCSSoeI dassitication schemes have b«n rai~ onother grounds (28-J1).
28. M. Zamir and H. Chce, Blood Vtssds 24, 76 (1987).29. L. Koblingcr and W. Hofinann, Phys. Md. Bioi, 30, 541 (1985).30, ---. AMI, Rtl. 221, 533 (1988).31. R. Sblak, N. Ozhya, T C. Skalak, Annu, Rtv. Auiil Mtcn. 21, 167 (1989),32. J. r. Mall, Abh. Mlllh,-Phys. Koniglicb .$lIch. GtSJtI/. IV"wm. 14, 151 (1888).33, K. Horsfield and G. Cumming, Bull, Malh. Biophys. 29, 245 (1967).34. K. Hors.fidd and A. Thurlbcck, Bull. Mom. Bioi, 43, 681 (1981).35. E. T. Engelson, T C. Skalak, G. W, Schmid·Schonbein, A'1iaovllJc. Rts, 30, 29
(1985).36. E. T. Engelson, G. W. Schmid·Schonbein, B. W. Zweifach, ibid. 31,356 (1986),37. M, L. E1lswonh, A. Liu, B. Dawant, A, S, Popel, R, N. Pirunan, ibid, 34, 168
(1987).38. A. G. Hudetz It ai,,). Cmb. Blood Row I\-ltlab. 7, 342 (1987).39. H. N. Marrovi12. and J. Roy, Am.). Physio/. 245, HI031 (1983).40. M. Koban tt III" J. Ctrtb. Blood Row Mtfabol. 4, 110 (1984).oil. N, Suwa tl al. [Tohob j. £y;p. Mtd, 79, 168 (1963» rook a similar approach in
their analysis of the human circulatory system; their ~ues ror the exponent rangedfrom 2.51 to 2.87 (mean, -2.7). HowC\'er, they :acknowledge that their pl""'Ccdurefor producing vCSSoeI ClSts yielded differential dilation of vessels of different sizes; ap:>st hoc correction was attempted, but the data must be considen::d suspect.
42. M. Umir,}. Tnror. Bioi. 62, 227 (1976).43. M. J. Woldenberg, ibid. 122, 187 (1986),44, M. Umir, ibid. 120, 111 (1986).45, A 5% cost variation ro build and run the system represents :about :a lO% \·ari:ation
in vessel radius. In genenJ, the total power (osr is morc sensitive to decn:::ascs invessel rndius th:an to increases. See (19) and T. F. Shem13n tl III. [). Thtor. Bioi.136,245 (1989)]. .
46. V. B. Wigglesworth, Prot. R. SOL London Str. B 106, 229 (1930).47. T. Weis·Fogh, J. Exp. Bioi. 41, 207 (1964).48. T. J. Pedley, Amlll. Rtv. RuirJ Mtch. 9, 229 (1977).49. Shear stress is dr:ag :arising from the viscosiry of the Ruid. The shear stress in laminar
Row is given by T = ~rJU/dy, when:: dUldy is the velocity gr:1diem pcrpendicul:ar to
rhe wal1s. In Poiscuille flow in cylindric:u rubes, T:: 4~Qhfr).
50. S. Rodb:ard, Cardiology 60, 4 (1975).51. A. Kamiya and T. Togawa, Am.). Physiol. 239, HI4 (1980).52. A. K:amiy:a, R. Bukhari, T. Togawa, Bull, Malh. Bioi. 46. 127 (1984).53. A. Kamiya, J. Ando, M. Shibar:a, H. M;lSuda, Biorhtology 25, 271 (1988).54. P. F. Da...ies, Ntws PI'ysiol. Sci. 4, 22 (1989).55. S.·P. Olesen, D. E. Cl:aph:am, P, F, Davies, Nlltuft 331, 168 (1988).56, S. L. Diamond, S, G, Eskin, L. V. ~Idnrirc:. Stirn't 243, 1483 (1989),57, M. Nakache and H, E. Gaub, Proc Naif. Acad, Sci, USA. 85, 184:1 (1988),58. For increases in flow beyond 400%, regularion of wall shar streSS through
diameter change W;lS incomplete bur in the appropri:are direction. Initiation of thisn::guIatory response could be detected as early ;IS 1 week. after flow r.ltes werealtered.
59, C K. Zarins tt 01., J. VlIJ{. Surg. 5, 413 (1987),60. B. L. Langille and F. O'Donnell, Scimtt 231, 405 (1986).61. These long·tenn responses should not be confused with the acute response of
arteries and arterioles to changing flow effected by contr.llction of smooth musclecells (again apparently mcdi;lted b)' :a response of the endothelial cells to local shearstress); sec, for eumplc, P, M, Vanhoune, Hyptnnuion 13,658 (1989); J. A. VitaIt w.,). Am. Coil. Gmliol, 14, 1193 (1989).
62. R. M. Zwolak, M. C Adams, A, W, Oowcs, J. VlIJt, Surg, 5, 126 (1987).63. Zarins tt al. (59) presented similar v.alues. H. H. Lipowsky and B. W. Zwcif.lch
[MimlV4Sr. RtJ. 7, 73 (1974» calculared ~ues for T in thc microvesscLs of catmc.scntary that varied almost 50·fold for vessels with diamcters from 7 ro 46 ~.However, their aru1ysis presumed that the viscosity or the blood was constant.
64. These shear r.ltes were calcul:a[ed on the usumption of Poiscuille flow in the\'csscLs. One cannot derive actual values for shar streSS without knowing the\i.Kosity of the seawater (dependent on both tcmpc:rarun: and saliniry). However,because se:awater is a Nc:wtoni:m fluid. she:ar sucss will be directly proportional to
shear rare.65. Secondary flows will also be established thn will further complicate the picture (JJ,
48,66), How streamlines for realistic models of arterial bnnch points arc prcsc:ntedin (67, 68) .
66. C. G. Caro, T. J. redley, R. C Schroter, W. A. Seed, Tht MuhlltliCJ ofthtCirtulaticm(Oxford Univ. Press, Oxford, 1978).
67. T. Karino and H. L. Goldsmith, Bierhtology 22, 87 (1985).68. T. Matsuo, R. Ok.eda, F. Higashino, ibid. 26, 799 (1989),69. P. F, Davies, S.·P. Olc:scn, D. E. Clapham, E. M. Morrc:l, F. J. Schoen, Hyprrttnsicm
11,563 (1988).70, M. Sam and N. Ohshima, Biorhtology 25, 279 (1988).
ARTICLES 999
7,
'.-

MICHAEL D. WARD* AND DANIEL A. BUTTRY
~iconceptual simplicity of these methods portend broad development 1
of commercial and research applications. In this article we discussthe fundamental properties, methodology, and examples of recent.applications that highlight the versatility of these mass-sensingpiezoelectric transducers.
PiezoelectricityIn 1880, ]acques and Pierre Curie discovered that a mechanical
stress applied to the surfaces of various crystals, includingquanz,rochelle salt (NaKC4H.,06· 4H20), and tourmaline, a1forded acorresponding electrical potential across the crystal whose m3gni- .tude was proportional to the applied stress (1). This behavior is:referred to as the piezoeleCtric effect, which is derived from the ~
SCIENCE, VOL. 149':
rtIIl~
H
. ~il$\ 1
for chemical agents. Advances in piezoe1ectric metbodolo~-llgy in the last decade now allow dynamic measurements of 1minute mass changes «10-9 grams per square centime- ~ter) at surfaces, thin films, and electrode interfaces in t'liqUid media as well. Mass measurements associated witha diverse collection of interfacial processes can he readilyperformed, including chemical and biological sensors,reactions catalyzed by enzymes immobilized on surfaces,electron transfer at and ion exchange in thin polymerfilms, and doping reactions of conducting polymers.
.jgill geometry may be the only option for these animals that increases gill surface ';1area over the phyUobrachiate condition and yet is still compatible with molting. On ;~
the other hand, here, too, no shear streSS signal is available; the epidc:rmal ccUs on Ithe gills arc covered by the cuticle. -j
83. The problem ofhow branching is controlled in these diverse sysn:ms has yet to be 'taddressed' l
84. M. laBarbera, J. Exp. Mar. BioI. &01. 55, 185 (1981). J85. C. B. Jorgensen tI al., Mar. Etol. Prog. Str. 34, 69 (1986). ·186. N. R. Silvester and M. A. Sleigh,]. Mar. Bioi. Assoc. U.K. 64,859 (1984).87. F. R. Bernard, Bioi. Bull. Woods Ho/~, Mass. 146, 1 (1974).88. P. R. WaIne, ). Mar. Bioi. Assoc. U.K. 52, 345 (1972).89. H. U. Riisgard. Mar. Etal. Prog. Str. 47, 129 (1988). .90. H. D. Green, in Mtdical Physics, O. Glasser, Ed. (Year Book, CJUcago, 1950), vol. ~
2, pp. 228-251.91. AS. Ibcra1I [Math. Biosa'. 1,375 (1967)J criticized the vessd dimensions prcscntal :
by Green (.90), in particular the valucs for the aorta and large arteries, and presenteda1ternativc values based on averages. As explained above, such av~gcs can hide the \pattern to be studicd. Some idea of the variation can be gathered from Iberall's6gure 2, where the diameters of the largest arterics vary by a faaor of 2.
92. W. F. Gan~n~, Rtvitw ofMtdiad Physiology (Lange Medical, Los Altos, CA, 1975). \ ..••93. H. M. RCtSWlg, J. Morphol. 145,493 (1975). :,94. A C. Bunon, Physiology and Biophysics of tht CiraJalion (Year Book, 0Uag0, :
1972). :95. G. P. Bidder, Q.J. Microsc. Sci. 67,293 (1923).96. I thank A. Bicwener, P. Davics, and S. Vogel for helpful comments on this
manuscript. This work was supported in part by NSF grant BSR 84-06731. .
In Situ Interfacial Mass Detection withPiezoelectric Transducers
-Present address: Department ofChcmical Engincering and Materials Science, Univer·sity of Minnesota, 421 Washington Avenuc, S.E., Minneapolis. MN 55455.
1000
M. D. Ward is in the Central Research & Development Department, E. I. duPont deNcmOlUS and Co., Inc., P. O. Box 80328, Wilmington, DE 19880-0328. D. A Buttryis in the Department of Chcmistry, Univccsity of Wyoming, Laramie, WY 820713838.
The converse piezoelectric effect, in which an electric fieldapplied across a piezoelectric material induces a stress inthat material, has spurred many recent developments inmass measurement teChniques. These methods commonlyrely on the changes in the vibrational resonant frequencyofpiezoelectric quartz oscillators that result from changesin .mass on the surface ofthe oscillator. The dependence offrequency on mass has been exploited extensively for massmeasurements in vacuum or gas phase, for example,thickness monitors for thin-film preparation and sensors
71. Vogel (6) recommended allowing a minimum of 1 pipe diameter downstream ofanentrance to a pipe to develop a recognizable approximation to a parabolic velocityprofilc; because the profilc in the entrance is not uniform. distanccs hcre would beeven greater. For full (99%) development of the profile, the entrance length isequal to 0.116 Re' r. J. K. B. Krijger t't al. [). Biomtch. 22, 1193 (1989)] fOWldthat vclocity profilcs did not return to a parabolic velocity distribution for 30 to 80pipe diameters downstream of a confluence of vcssels. .
72. L. S. Smith,). Morphol. 113,261 (1963).73. J. Browning, Zoomrorphology Btrl. 96, 243 (1980).74. G. B. Bourne, J. R. Redmond, D. D. Jorgensen, Physiol. Zool. 63, 140 (1990).75. J. Volaow, Zoomorphology B"'. 105,395 (1985).76. B. R. McMahon and L. E. Burnett, Physiol. Zool. 63, 35 (1990).77. E. H. Gladfelter, Bioi. Bull. Woods Holt. Mass. 165,619 (1983).78. J. C. Grimmer and N. D. Holland, Zoomorphology Btrl. 94,93 (1979).79. J. R. Blake, N. Liron, G. K. Aldis, ). Thtor. Bioi. 98, 127 (1982).80. W. F. Pickard, Prog. Biophys. Mol. Bioi. 37, 181 (1981).81. F. W. Ewers and J. B. Fisher, Am.). Bol. 76,645 (1989).82. Crustacea (Arthropoda) with dendrobrachiate or trichobrachiate gills [Po A.
Mclaughlin, in Tht Biology oftht Cnutacta, vol. 5, Inttntal Anatomy and PhysiologicalRtgulation, L. H. Mante~ Ed. (Academic Press, New York, 1983), pp. 1-52] havea distributed transfer region in which a Murray's law system has not evolved. Themost common gill type in the decapod crustaCcan5, the phyUobranchiate gill, showsa typical planar transfer geometry. The exchange surface of dcndrobrachiate andtrichobrachiate gills is clearly distributed in three·dimensional space, but there is noevidence of division of the channels between the filaments into some hierarchy ofbranched vessels. This example may be akin to straimng at gnats. Such a distributed
THE SIGNIFICANCE OF INTERFACIAL PROCESSES IN RE
search and commercial applications (such as sensors, electroplating, and c0FI'0sion) has stimulated the development of
methodologies that probe interfacial processes and chemistry atsurfaces and thin films. Advances in piezoelectric methods in the lastdecade now make possible in situ detennination of minute masschanges that occur at thin films and surfaces under a variety ofconditions, including liquid media. The low cost and procedural and
• •
iI
iII
j
'j .,I
1
J
III·t
t.1~