pre- and postprandial changes in orexigenic and
TRANSCRIPT

General and Comparative Endocrinology 176 (2012) 231–239
Contents lists available at SciVerse ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
Pre- and postprandial changes in orexigenic and anorexigenic factorsin channel catfish (Ictalurus punctatus)
Brian C. Peterson a,⇑, Geoffrey C. Waldbieser a, Larry G. Riley Jr. b, Kelli R. Upton b, Yasuhiro Kobayashi c,Brian C. Small d
a USDA–ARS Catfish Genetics Research Unit, Thad Cochran National Warmwater Aquaculture Center, Stoneville, MS 38776, USAb Department of Biology, California State University at Fresno, Fresno, CA 93740, USAc Department of Biological Sciences, Fort Hays State University, Hays, KS 67601, USAd Fisheries and Illinois Aquaculture Center Department of Animal Science, Food and Nutrition, Southern Illinois University, Carbondale, IL 62901, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 18 November 2011Revised 25 January 2012Accepted 28 January 2012Available online 16 February 2012
Keywords:AppetiteGRLNCARTNPYCCKCatfish
0016-6480/$ - see front matter Published by Elsevierdoi:10.1016/j.ygcen.2012.01.022
⇑ Corresponding author. Fax: +1 662 686 3567.E-mail address: [email protected] (B.C.
Ghrelin (GRLN), cocaine and amphetamine regulated transcript (CART), neuropeptide Y (NPY), and cho-lecystokinin (CCK) are neuropeptides involved in the regulation of appetite and feeding in vertebrates.We examined pre- and postprandial changes in the expression of plasma GHRL and mRNAs encodingGRLN, CART, NPY, and CCK in channel catfish. Fish were entrained to eat at 0900 h for 2 weeks. Fish werethen sampled at 0700, 0800, and 0900 h. Remaining fish were either offered feed at 0900 h (Fed) or fasted(Unfed). Fish sampling continued at 0.5, 1, 2, and 4 h post feeding. Feeding increased abundance of wholebrain CART mRNA out to 4 h with no effect observed in unfed fish. Whole brain NPY expression peaked at0.5 h in both treatments. NPY expression then declined in fed fish but remained elevated in unfed fish. Nodifferences in plasma or stomach GRLN expression were observed. Two separate cDNAs for CCK wereidentified. Brain CCKa and CCKb expression increased after feeding. These results suggest CART, NPY,and CCK play roles in the regulation of channel catfish feeding. Taken together, these results providenew insights into the neural and gastroenteric mechanisms regulating appetite in channel catfish.
Published by Elsevier Inc.
1. Introduction
In fish and other vertebrates, feeding and satiation are con-trolled through complex integrations of stimulatory (orexigenic)and inhibitory (anorexigenic) signals from the central nervous sys-tem and peripheral signals that provide information to the brainabout body energy reserves and meal size [6]. These elaborateinteractions have been studied in detail in mammals and yet themechanisms through which they regulate appetite are not clearlydefined. Investigation into the neural regulation of appetite in dif-ferent fish species has only recently begun. Understanding howfood intake is regulated in fish is important not only from a com-parative neuroendocrine perspective but may also have importantimplications on how we manage populations of fish.
Many neuropeptides homologous to mammalian appetite-regulating peptides have been isolated or their sequence deducedfrom cloned cDNA sequences in fish. Key regulators shown to con-trol appetite in fish include cocaine and amphetamine regulatedtranscript (CART), ghrelin (GRLN), cholecystokinin (CCK), neuro-peptide Y (NPY), and others [63]. A complete picture of how these
Inc.
Peterson).
signals regulate appetite before and after a meal is not clear formost species.
Cocaine and amphetamine regulated transcript is an anorexi-genic peptide that was first discovered in rats as a transcript thatincreased following administration of cocaine and amphetamine[8]. This transcript was later shown to have a role in the regulationof feeding as centrally injected CART inhibited food intake in rats[13]. To date, one or more genes or alternatively spliced CARTvariants have been identified in goldfish (Carassius auratus) [62],Atlantic cod (Gadus morhua) [24], catfish (Ictalurus punctatus)[25], winter skate (Raja ocellata) [8], winter flounder (Pseudopleuro-nectes americanus) [65], and medaka (Oryzias latipes) [33]. Many ofthese fish studies provide evidence that CART also plays a role inregulating food intake as has been reported for mammals.
In mammals, NPY functions as a powerful enhancer of appetitein the hypothalamus [59]. Neuropeptide Y has been isolated andcharacterized in a number of fish species such as goldfish (Carassiusauratus) [2], Chinese perch (Siniperca chuatsi) [28], rainbow trout(Oncorhynchus mykiss) [9], Atlantic cod (Gadus morhua) [24],Atlantic salmon (Salmo salar) [34], and channel catfish [29]. Fooddeprivation increases NPY mRNA in the hypothalamus of goldfish,coho salmon (Oncorhynchus kisutch), Chinook salmon (Oncorhyn-chus tshawytscha), and channel catfish [37,52–54] and this effect

232 B.C. Peterson et al. / General and Comparative Endocrinology 176 (2012) 231–239
can be reversed by refeeding [38]. Thus, the orexigenic function ofNPY seems to be conserved in fish.
Ghrelin is a peptide hormone that is synthesized in the stomach[51] and brain [4] that is involved in the control of food intake andenergy homeostasis in mammals [36,68]. Ghrelin has been identi-fied in several species of fish including goldfish [57], Japanese eel(Anguilla japonica) [18], rainbow trout, [19], Nile tilapia (Oreochr-omis niloticus) [41], Mozambique tilapia (Oreochromis mossambi-ques) [20], channel catfish [21], sea bass (Dicentrarchus labrax)[56], Atlantic salmon [34], and Arctic charr (Salvelinus alpines)[11]. Several of these studies suggest that GRLN is involved in stim-ulating appetite, energy homeostasis, feeding, and metabolism infish. These functions of GRLN have been reviewed by Kaiya andcoworkers [22].
Cholecystokinin is found in the brain and gastrointestinal tractand has multiple biologically active forms with CCK-8 being themost abundant form in the brain [32]. In fish and mammals, CCKhas many physiological roles but functions primarily as a satietysignal [5,31]. CCK-like immunoreactivity has also been shown inthe gut and nervous system of several fish species including Atlan-tic cod [17], goldfish [14], and halibut (Hippoglossus hippoglossus)[23]. CCK mRNA sequences have also been determined for a num-ber of fish species including goldfish [45], dogfish (Squalus acanth-ias) [16], pufferfish [41], Japanese flounder (Paralichthys olivaceus)[26], yellow tail (Seriola quinqueradiata) [35], winter skate [31],and red drum (Sciaenops ocellatus) [67]. Results of these studiessuggest that CCK plays a role in appetite regulation in teleost fish.
The functions of the above mentioned appetite stimulatory andinhibitory signals are slowly being defined in several species offish. In order to provide new information on the mechanisms thatregulate appetite in channel catfish, we assessed pre- and post-prandial changes in gene expression concentrations of GRLN, CART,NPY, and CCK. In addition, plasma levels of GRLN were measured.
2. Materials and methods
2.1. Research animals
Fish, the National Warmwater Aquaculture Center (NWAC) 103strain, used in this study were housed at the USDA–ARS CatfishGenetics Research Unit, Stoneville, MS aquaculture facility. Priorto randomization in tanks, approximately 25 fish from six differentspawns were placed into a 120-L holding tank for 1 day. One hun-dred and fifty NWAC103 catfish with a mean initial size of17.6 ± 0.1 g were randomly assigned to 6, 76-L tanks (25 fish/tank),and allowed to acclimate for 2 weeks under 14 L: 10 D h photope-riod at 27.1 �C flow-through well water. The acclimation period in-cluded feeding the fish to apparent satiation at 0900 h with acommercial feed (Delta Western, Indianola, MS). During experi-mentation, fish were reared in identical conditions as describedabove and fed once per day at 0900 h to apparent satiation. Appar-ent satiation was achieved by offering small quantities of feed tothe fish by hand until feeding activity stopped which was approx-imately 15 min. Water quality (pH �8.5 and dissolved oxygen lev-els > 5.0 mg/L) and flow rates were similar between tanks.
2.2. Tissue and plasma collection
Sample collection was conducted by using the protocol follow-ing the general guideline and procedure of the USDA InstitutionalAnimal Use and Care Committee (Approval # 64-F-006-0803).One group of three tanks of fish served as fed controls (fed) whilethe remaining three tanks of fish comprised a group that was fasted(unfed). For sampling, 0900 h was designated as the 0 h time point.
Prior to feeding, one fish from each tank was sampled at�2,�1, and0 h. The fed fish were fed at 0900 h while the unfed fish did not re-ceive food. Two fish from each tank were then sampled at 0.5, 1, 2,and 4 h post feeding. Fish from each group were euthanized with anoverdose of tricaine methanesulfonate (MS-222; Argent ChemicalLaboratories, Redmond, WA) and tissues were taken for analysis.Blood was collected from the caudal vasculature into syringescoated with heparin (20 I.U.). Plasma was isolated by centrifugationand stored at �20 �C until samples were analyzed for ghrelin.Whole brain (hypothalamus removed), hypothalamus, stomach,and the proximal small intestine (�100 mg) were taken for RNAextraction. Samples were immediately placed in 1 mL TRI-Reagent(Molecular Research Center, Inc., Cincinnati, OH), flash-frozen in li-quid nitrogen, and stored at �80 �C until RNA isolation.
2.3. RNA isolation, cloning, cDNA synthesis, and real time PCR
Total RNA was isolated using TRI-Reagent according to the man-ufacturer’s protocol and then quantified by measuring the absor-bance at 260 nm using a NanoDrop ND-1000 spectrophotometer(NanoDrop Technologies, Rockland, DE). The integrity of the RNAwas verified by visualization of the 18S and 28S ribosomal bandsstained with ethidium bromide after electrophoresis on 2.0% aga-rose gels.
Extracted RNA was treated with DNase I to remove co-extractedDNA using a TURBO DNA-free™ kit (Ambion, Austin, TX, USA). Analiquot of the extracted RNA (20 lL) was treated at 37 �C for30 min with 0.1 volume of buffer (1.5 lL) and 2 U (1 lL) of DNaseI. The enzyme was deactivated with 2 lL of inactivation reagent atroom temperature for 2 min. Samples were centrifuged at 10,000gfor 1 min and the supernatant (16 lL) was transferred to a cleanRNase-free microcentrifuge tube and then stored at �80 �C beforethe reverse transcriptase step.
Zebrafish CCK sequence was used to query catfish EST and full-length cDNA sequences (www.catfishgenome.org/cbarbel) usingBlastN and BlastX. The search revealed full-length coding sequencefor CCKa and partial coding sequence for CCKb. The catfish se-quences were used as queries to identify genomic sequence contigs(unpublished) for each gene and to determine exon–intron bound-aries and validate EST sequences. Amplification primers and probesequences (Table 1) were designed for quantitative real-time PCRbased on genomic contig sequence. Amplification products werecloned into the pCR�4-TOPO vector (Invitrogen, Carlsbad, CA)and the identity of the cloned inserts was verified by automatedSanger sequencing (ABI PRISM 3100 Genetic Analyzer; Applied Bio-systems, Foster City, CA). The DNA concentration of each recombi-nant plasmid was measured using the NanoDrop ND-1000spectrophotometer.
One microgram of total RNA from each tissue was reverse-tran-scribed using the iScript cDNA Synthesis Kit (BioRad, Hercules, CA)according to the protocol provided by the manufacturer. Quantita-tive real-time PCR was performed using the iCycler iQ real-timePCR detection system (BioRad) to quantify GHRL, NPY, CART, andalpha tubulin mRNA as previously described [21,25,43,55]. Primerand probe sequences for CCKa and CCKb are listed in Table 1. Eachamplification reaction mixture (12.5 lL) contained 400 ng ofcDNA; 1� iQ™ Supermix (Bio-Rad), 1.0 lM dual-labeled probe,and 10 lM (CCKa, CCKb) of each primer. The real-time PCR proto-col for CCKa and CCKb was 3 min at 95 �C; 45 cycles of 95 �C �15 s,60 �C �1 min. All amplifications were performed in triplicate. Thestandard curve showed a linear relationship between cycle thresh-old values and the logarithm of input gene copy number. All spe-cific quantities were normalized against the amount of alphatubulin amplified as this reference gene has been shown to be sta-ble in similar channel catfish studies [55].

Table 1Nucleotide sequences of the PCR primers and probes used to assay gene expression by real-time quantitative PCR.
Gene Primer Sequence PCR productlength (base pair)
CCKa SenseAntisenseProbe1
TGT GCG CAG GCT GAA TGA ACGTC TCT GTC CTT TAT TCT GTG GACT CAT TTC CAG AAA AGG TTC ATACCG CAG
109
CCKb SenseAntisenseProbe1
TGC AGA TGG ACT CCA GCG TGCCT TGC TGT TCG CCA TCG AGCGA CGA ACA GAA CCT TTC CTG GAG ATA AGT C
126
1 The probes CCKa (GenBank Accession No. JQ028716) and CCKb (GenBank Accession No. JQ028715) were dual labeled with a reporter dye(FAM, 6-carboxyfluorescein) at the 50 end and a quencher dye (BHQ-1, Black Hole quencher-1) at the 30 end (Biosearch Technologies, Novato, CA).
414
453
54
726
252
197
105
755 255
3833+ CCK-B
CCK-A
Fig. 1. Structure of catfish CCKa and CCKb genes. Exons are denoted as boxes andintrons are denoted as lines. Exon sizes (basepairs) are denoted above line andintron sizes are denoted in italic below line.
B.C. Peterson et al. / General and Comparative Endocrinology 176 (2012) 231–239 233
2.4. Ghrelin RIA
Plasma GRLN levels were measured following the protocol de-scribed by Hosoda et al. [15] and validated for catfish following Ri-ley et al. [48] and Picha et al. [47]. Briefly, aliquots of plasma weretreated the following ways for validation: (1) 1/10 volume of 1 MHCl added to the plasma + purified by SepPak Light C18 cartridge(Waters, Milford, MA), (2) neat plasma + SepPak purification, (3)HCl added and no SepPak purification, (4) neat plasma (no HCland SepPak purification). SepPak cartridges were preconditionedwith 2 ml ethanol and 2 mL 0.1% aqueous trifluoroacetic acid(TFA) and eluted with 60% CH3CN/0.1% TFA. Samples were thendried and the pellet was re-hydrated in RIA buffer (50 mM Na2H-PO4; 25 mM EDTA–2Na; 80 mM NaCl; 0.05% NaN3; 0.5% Triton X-100; 0.5% BSA). Only the samples treated with HCl followed bySepPak purification exhibited a parallel slope to the standard curve(data not shown). Anti-rat GRLN [1–11] (a gift from Dr. HiroshiHosoda), at a final concentration of 1:750,000 was incubated withstandards (rat GRLN-C8) and plasma samples for 24 h at 4 �C. Theantibody detects both the C-8 and C-10 form of ghrelin, not theunacylated form. One hundred microliter of commercially avail-able 125I-human GRLN (Millipore, St. Charles, MO) was added andincubated for an additional 24 h at 4 �C. One hundred microliterof anti-rabbit IgG antibody (Sigma: 1:35 in RIA buffer) was addedand tubes incubated overnight at 4 �C. Free and bound tracers wereseparated by centrifugation at 3000g for 1 h at 4 �C. The superna-tant was aspirated and samples were counted using the Cobra IIgamma counter (Packard, Meriden, CT).
2.5. Statistical analysis
Data were analyzed by using the GLM procedures of StatisticalAnalysis System Version 9.1 software (SAS Institute, Inc., Cary,North Carolina, USA) for a repeated measures design. Tank servedas the experimental unit for each variable measured. Normalizedgene expression data passed Levene’s test for homogeneity of var-iance. The model included the main effect of time (�1, �2, 0, 0.5, 1,2, and 4 h). When the main effect was significant (P < 0.05), leastsquares means separation was accomplished by the PDIFF optionof SAS.
3. Results
Two CCK genes were identified in channel catfish with similarexonic structure. Both genes contained 3 exons based on compar-ison of the genomic and transcript sequences and the initiator co-don was found on exon 2 in both genes. However, the secondintron in CCKa was 255 bp compared with >3800 bp in CCKb(Fig. 1). Both genes encoded an identical CCK-8 peptide and dem-onstrated sequence similarity to other fish CCK coding sequences(Fig. 2).
Centrally, the abundance of CART mRNA expression in the brainincreased after feeding (0.5 h) and continued to be elevated 4 hpost feeding compared to unfed fish (P < 0.01) (Fig. 3). Concentra-tions of CART mRNA in the unfed fish did not change significantlyover the course of the study.
There were no differences in abundance of NPY mRNA expres-sion in the hypothalamus as a result of feeding (Fig. 4A). However,we observed a clear peri-prandial effect in the brain. NPY mRNAconcentrations within the brain were noticeably different over thetime course of the study (Fig. 4B) (P < 0.05). In the fed fish, abun-dance of NPY mRNA tended to peak at 0.5 h and then declined overthe next 4 h. In the unfed fish, NPY mRNA concentrations alsotended to peak at 0.5 h but continued to stay elevated at 1, 2 and 4 h.
Peripherally, there was no difference in abundance of stomachGRLN mRNA as a result of feeding and no peri-prandial changeswere observed (Fig. 5A). Similarly, plasma levels of GRLN did notchange over the time course of the study or with treatment(Fig. 5B).
Abundance of CCKa mRNA in the intestine increased (P < 0.001)in both the fed and unfed group of fish at 4 h compared to the othertime points (Fig. 6A). There was an increase (P < 0.05) in brain CCKamRNA in the fed group 1 h after feeding that continued out to 4 h(Fig. 6B). Abundance of CCKa in the hypothalamus was not differ-ent between the fed or unfed fish throughout the experiment(Fig. 6C).
Similar to CCKa, gene expression of CCKb increased (P < 0.001)in both the fed and unfed fish at 4 h compared to the other timepoints (Fig. 7A). Feeding increased (P < 0.05) abundance of brainCCKb mRNA at 4 h (Fig. 7B). Abundance of CCKb in the hypothala-mus was not different between the fed or unfed fish throughoutthe experiment (Fig. 7C).
4. Discussion
There is evidence that CART functions in the regulation offeeding in fish. Intracerebroventricular (ICV) administration ofhuman CART decreases food intake in goldfish [61] while fooddeprivation decreases expression of CART in a variety of speciesof fish [24,62–64] including catfish [25]. In addition, post-prandial

Fig. 2. Multiple alignments of cholecystokinin peptides in bony and cartilaginous fish and Xenopus.
234 B.C. Peterson et al. / General and Comparative Endocrinology 176 (2012) 231–239
changes in CART mRNA expression have been observed in goldfish[62], Atlantic cod [24], and Atlantic salmon [60]. In the presentstudy, post-prandial changes in CART were also observed in channelcatfish. CART gene expression was increased post-prandially andcontinued to stay elevated for 4 h. In the unfed fish, levels of CARTmRNA were similar during the course of the study. In goldfish andAtlantic salmon, CART mRNA levels increased 2 and 1.5 h, respec-tively [62] while in Atlantic cod CART expression decreased 2 hpost-feeding and remained low after 7 days of withholding feed[6]. In another Atlantic salmon study, CART expression fell in fishfasted for 6 days [14]. In a previous catfish study, levels of CARTmRNA within the brain decreased after 30 days of fasting whilerefeeding for 15 days restored expression to a level similar to thefed control [25]. In a separate 7 week feeding study, CART mRNAlevels were lower in catfish that consumed more food and gainedmore weight [25]. Taken together, these results suggest that CARTis involved in the regulation of short and long term feeding, as wellas a central controller of body weight, similar to what has been re-ported in mammals [13].
It has been suggested that NPY is a key orexigenic peptide in theregulation of appetite in teleost fish [3,27,65]. In the present study,there were no significant differences in the levels of NPY mRNA inthe hypothalamus as a result of feeding. In contrast, NPY mRNAlevels within the whole brain were noticeably different over thetime course of the study. In fed fish, NPY mRNA levels peaked afterfeeding and then declined over the next 4 h. In the unfed fish, NPYmRNA levels also peaked at 0.5 h but continued to stay high overthe next 4 h. Previous studies with channel catfish showed thatinjection of porcine NPY increased food intake [19]. Similarly,injections of NPY also increased food intake in goldfish [37,7,30].In addition, NPY expression in the brain of goldfish increased 1–3 h ahead of feeding and fell 1–3 h after feeding [37].
Our results are similar to what has been observed for Atlanticcod and Atlantic salmon in which the expression of NPY increasesconcurrently with the onset of feeding [24,60]. NPY mRNA levelstended to decrease after a meal (2 h) in Atlantic cod [24] while lev-els were similar post feeding in Atlantic salmon [60]. In goldfish,NPY mRNA expression decreases in the telencephalon and in the

Fig. 2 (continued)
Fig. 3. Mean ± SE brain cocaine and amphetamine regulated transcript (CART)mRNA levels in Fed and Nonfed catfish (N = 6 per time point) sampled at �2, �1, 0,0.5, 1, 2 and 4 h. CART copy number was normalized as a ratio to the amount ofalpha tubulin. Significant differences (P < 0.05) are denoted by an asterisk.
B.C. Peterson et al. / General and Comparative Endocrinology 176 (2012) 231–239 235
hypothalamus 1 h after a meal but shows a small but significant in-crease in the optic tectum area [37]. Taken together, these resultssuggest an area specific regulation of NPY expression among tele-osts. Furthermore, our results suggest that NPY is involved in theregulation of food intake in channel catfish.
In a previous study we showed that there was no difference inabundance of NPY mRNA in the hypothalamus between fast andslow growing catfish [43]. The fast growing catfish consumed morethan 50% more food over the 7 week growth study and we hypoth-esized that higher levels of NPY mRNA would explain differences infood intake and thus growth. In another study we fed catfish once,twice, or three times a day for 6 weeks [44]. Fish fed three times aday consumed 53% more food and thus gained more weight thanfish fed only once a day. At the end of the study we took wholebrain tissue samples and measured mRNA levels of NPY. We didnot detect a difference in gene expression of whole brain NPY be-tween fish fed once a day compared to fish fed three times a day(Peterson et al., unpublished). In both studies, fish were sampled
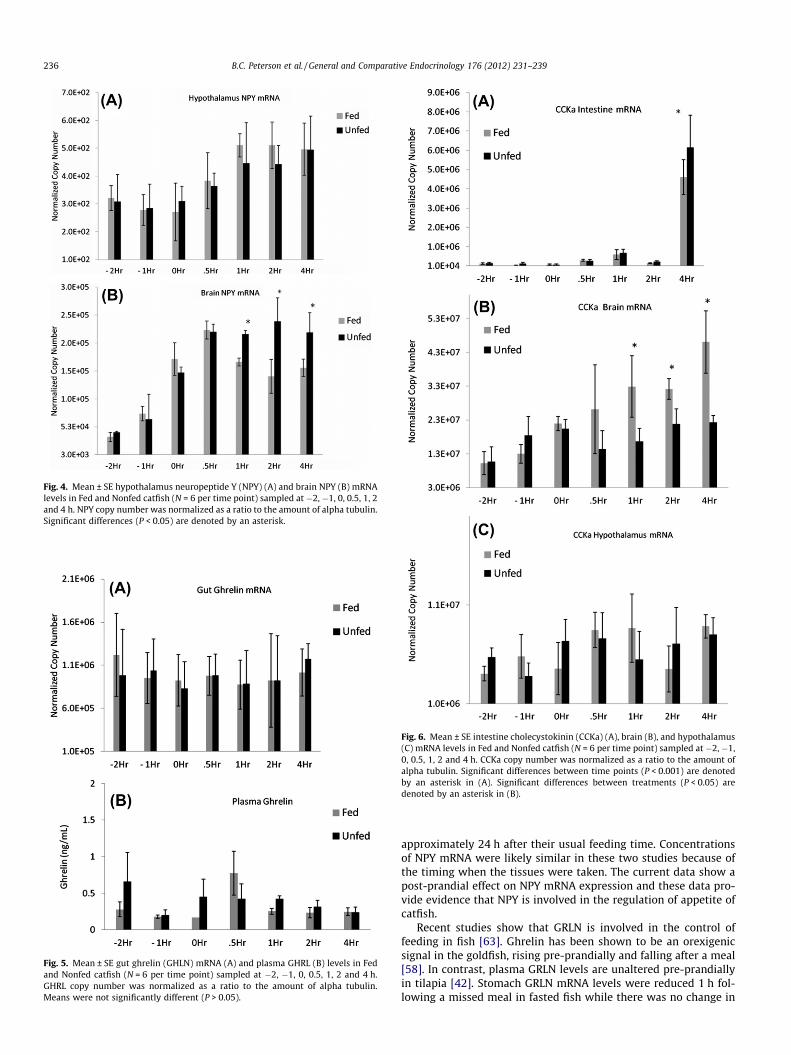
Fig. 4. Mean ± SE hypothalamus neuropeptide Y (NPY) (A) and brain NPY (B) mRNAlevels in Fed and Nonfed catfish (N = 6 per time point) sampled at �2, �1, 0, 0.5, 1, 2and 4 h. NPY copy number was normalized as a ratio to the amount of alpha tubulin.Significant differences (P < 0.05) are denoted by an asterisk.
Fig. 5. Mean ± SE gut ghrelin (GHLN) mRNA (A) and plasma GHRL (B) levels in Fedand Nonfed catfish (N = 6 per time point) sampled at �2, �1, 0, 0.5, 1, 2 and 4 h.GHRL copy number was normalized as a ratio to the amount of alpha tubulin.Means were not significantly different (P > 0.05).
Fig. 6. Mean ± SE intestine cholecystokinin (CCKa) (A), brain (B), and hypothalamus(C) mRNA levels in Fed and Nonfed catfish (N = 6 per time point) sampled at �2, �1,0, 0.5, 1, 2 and 4 h. CCKa copy number was normalized as a ratio to the amount ofalpha tubulin. Significant differences between time points (P < 0.001) are denotedby an asterisk in (A). Significant differences between treatments (P < 0.05) aredenoted by an asterisk in (B).
236 B.C. Peterson et al. / General and Comparative Endocrinology 176 (2012) 231–239
approximately 24 h after their usual feeding time. Concentrationsof NPY mRNA were likely similar in these two studies because ofthe timing when the tissues were taken. The current data show apost-prandial effect on NPY mRNA expression and these data pro-vide evidence that NPY is involved in the regulation of appetite ofcatfish.
Recent studies show that GRLN is involved in the control offeeding in fish [63]. Ghrelin has been shown to be an orexigenicsignal in the goldfish, rising pre-prandially and falling after a meal[58]. In contrast, plasma GRLN levels are unaltered pre-prandiallyin tilapia [42]. Stomach GRLN mRNA levels were reduced 1 h fol-lowing a missed meal in fasted fish while there was no change in

Fig. 7. Mean ± SE intestine cholecystokinin (CCKb) (A), brain (B), and hypothalamus(C) mRNA levels in Fed and Nonfed catfish (N = 6 per time point) sampled at �2, �1,0, 0.5, 1, 2 and 4 h. CCKb copy number was normalized as a ratio to the amount ofalpha tubulin. Significant differences between time points (P < 0.001) are denotedby an asterisk in (A). Significant differences between treatments (P < 0.05) aredenoted by an asterisk in (B).
B.C. Peterson et al. / General and Comparative Endocrinology 176 (2012) 231–239 237
stomach GRLN mRNA levels in fed tilapia (Oreochromis mossambi-cus) [42]. In burbot (Lota lota), fasting reduces plasma GRLN immu-noreactive peptide levels [39]. These studies suggest GRLNfunctions in the regulation of feed intake in these three speciesof fish. The role of GRLN in channel catfish is less clear. We foundthat gut mRNA levels did not change pre- or post-prandially ineither the fed or unfed catfish. Similarly, plasma levels of GRLNdid not change over the time course of the study.
In a previous catfish study, it was shown that gene expression ofGRLN was highest in the stomach followed by the pancreas andgall bladder [19]. These results suggest that the stomach (gut) isthe primary site and source of ghrelin being produced in catfish.We did not measure GRLN mRNA in any other tissue other thanthe gut as we hypothesized that GRLN mRNA in the gut and GRLNin the plasma would provide sufficient data to determine the roleof GRLN pre- and post-feeding. The role that GRLN plays in the reg-ulation of food intake in catfish is unclear at this time.
Studies in both mammals and teleosts have shown that CCKacts as a satiety signal in regulating food intake. The channel cat-fish, zebrafish, Japanese flounder, Atlantic salmon, rainbow trout,and Xenopus laevis genomes encode at least two CCK genes,although it is not known whether these arose due to localized orwhole genome duplication in a common ancestor. In contrast, onlyone CCK gene is annotated in the human and mouse genomes. Thehigh level of CCK-8 peptide sequence conservation within and be-tween fish species points to a conserved physiological role for CCKin these species. However, duplication of the CCK gene in fish couldpermit alternate transcriptional regulation that could alter tempo-ral or spatial patterns of CCK gene expression. In addition, two CCKgenes may function as back-up mechanisms to control satiety un-der different physiological situations. It is not clear if these genesevolved under separate natural selection pressures.
In teleosts, CCK participates in the regulation of digestive pro-cesses [1,10,40,49]. CCK also influences appetite regulation in fishas it has been demonstrated that both central and peripheral injec-tions of CCK cause a decrease in food intake in goldfish [17,66].Oral administration of CCK decreases food intake in sea bass [50],and oral administration of a CCK antagonist causes an increase infood consumption in trout and sea bass [12,50]. Furthermore,CCK mRNA levels increase after a meal in goldfish brain [46] andin the pyloric caeca of yellowtail (Seriola quinqueradiata) [35].
The present study is the first to examine feeding regulation oftwo CCK cDNAs in any species of fish. Both CCK cDNAs appearedto be regulated similarly in the three tissues we examined. Forexample, there was an increase in intestinal CCKa and CCKb mRNA4 h after feeding. In the brain, CCKa mRNA increased 1 h after feed-ing while CCKb mRNA increased 4 h after feeding. Expression ofboth genes appeared to be relatively stable in the hypothalamusin the fed and unfed groups of fish over the course of the study.It is interesting that intestinal CCKa and CCKb mRNA increased inboth the fed and unfed fish 4 h after the fish were either fed ornot fed. This is difficult to explain but it is possible that the avail-ability of feed at a regular time results in modifying the circadianpattern of hormone levels. It is also plausible that missing one mealis not enough to alter the entrained physiological response to beingfed at 0900 h, suggesting that CCK is not playing a role in reducingfood intake after a short-term fast. Further studies examiningabundance of CCKa and CCKb mRNA in the intestine under fedand fasted conditions over longer periods of time will aide in fur-ther understanding the roles of these genes in food regulation incatfish.
In summary, our results suggest that CART, NPY, and CCKa andCCKb are likely involved in the regulation of feeding in channel cat-fish. The function of GRLN during feeding is not known as bothplasma and mRNA concentrations were relatively unchanged dur-ing the peri-prandial experiment. Taken together, these resultsprovide new information on the mechanisms regulating appetitein channel catfish.
Acknowledgments
The authors wish to thank the assistance of the staff and tech-nicians of the USDA/ARS Catfish Genetics Research Unit. Mentionof trade name, proprietary product, or specific equipment does

238 B.C. Peterson et al. / General and Comparative Endocrinology 176 (2012) 231–239
not constitute a guarantee or warranty by the U.S. Department ofAgriculture and does not imply approval to the exclusion of otherproducts that may be suitable.
References
[1] G. Aldman, S. Holmgren, Intraduodenal fat and amino acids activategallbladder motility in the rainbow trout, Oncorhynchus mykiss, Gen. Comp.Endocrinol. 100 (1995) 27–32.
[2] A.G. Blomqvist, C. Soderberg, I. Lundell, R.J. Milner, D. Larhammar, Strongevolutionary conservation of neuropeptide Y: sequences of chicken, goldfish,and Torpedo marmorata DNA clones, Proc. Natl. Acad. Sci. USA 89 (1992) 2350–2354.
[3] J.M. Cerdá-Reverter, D. Larhammar, Neuropeptide Y family of peptides:structure, anatomical expression, function, and molecular evolution,Biochem. Cell Biol. 78 (2003) 371–392.
[4] M.A. Cowley, Hypothalamic melanocortin neurons integrate signals of energystate, Eur. J. Pharmacol. 480 (2003) 3–11.
[5] W.A. Cupples, Regulation of body weight, Am. J. Physiol. Regul. Integr. Comp.Physiol. 282 (2002) R1264–R1266.
[6] N. De Pedro, B.T. Björnsson, Regulation of food intake by neuropeptides andhormones, in: D. Houlihan, T. Boujard, M. Jobling (Eds.), Food Intake in Fish,Blackwell Science, Oxford UK, 2001, pp. 269–296.
[7] N. De Pedro, M.A. Lopez-Patino, A.I. Guijarro, M.L. Pinillos, M.J. Delgado, M.Alonso- Bedate, NPY receptors and opioidergic system are involved in NPY-induced feeding in goldfish, Peptides 21 (2000) 1495–1502.
[8] J. Douglass, A.A. McKinzie, P. Couceyro, PCR differential display identifies a ratbrain mRNA that is transcriptionally regulated by cocaine and amphetamine, J.Neurosci. 15 (1995) 2471–2481.
[9] C. Doyon, K.M. Gilmour, V.L. Trudeau, T.W. Moon, Corticotropin-releasing factorand neuropeptide Y mRNA levels are elevated in the preoptic area of sociallysubordinate rainbow trout, Gen. Comp. Endocrinol. 133 (2003) 260–271.
[10] L.G. Forgan, M.E. Forster, Effects of potential mediators of an intestinal brakemechanism on gut motility in Chinook salmon (Oncorhynchus tshawytscha),Comp. Biochem. Physiol. C 146 (2007) 343–347.
[11] E. Frøiland, K. Murashita, E.H. Jorgensen, T. Kurokawa, Leptin and ghrelin inanadromous Arctic charr: cloning and change in expressions during a seasonalfeeding cycle, Gen. Comp. Endocrinol. 165 (2010) 136–143.
[12] A. Gelineau, T. Boujard, Oral administration of cholecystokinin receptorantagonists increase feed intake in rainbow trout, J. Fish Biol. 58 (2001)716–724.
[13] M.H.A.G. Gorissen, G. Flik, M.O. Huising, Peptides and proteins regulating foodintake: a comparative view, Anim. Biol. 56 (2006) 447–473.
[14] B.A. Himick, R. Peter, CCK/gastrin-like immunoreactivity in brain and gut, andCCK suppression of feeding in goldfish, Am. J. Physiol. 36 (1994) R841–R851.
[15] H. Hosoda, M. Kojima, H. Matsuo, K. Kangawa, Ghrelin and des-acyl ghrelin:two major forms of rat ghrelin peptide in gastrointestinal tissue, Biochem.Biophys. Res. Commun. 279 (2000) 909–913.
[16] A.H. Johnsen, L. Jonson, I.J. Rourke, J.F. Rehfeld, Elasmobranchs expressseparate cholecystokinin and gastrin genes, Proc. Natl. Acad. Sci. USA 94(1997) 10221–10226.
[17] W. Jonsson, S. Holmgren, B. Holstein, Gastrin/CCK-like immunoreactivity inendocrine cells and nerves in the gastrointestinal tract of the cod Gadusmorhua and the effect of peptides of the gastrin/CCK family on codgastrointestinal smooth muscle, Gen. Comp. Endocrinol. 66 (1987) 190–202.
[18] H. Kaiya, M. Kojima, H. Hosoda, L.G. Riley, T. Hirano, E.G. Grau, K. Kangawa,Amidated fish ghrelin: purification, cDNA cloning in the Japanese eel and itsbiological activity, J. Endocrinol. 176 (2003) 415–423.
[19] H. Kaiya, M. Kojima, H. Hosoda, S. Moriyama, A. Takahashi, H. Kawauchi, K.Kangawa, Peptide purification, complementary deoxyribonucleic acid (DNA)and genomic DNA cloning, and functional characterization of ghrelin inrainbow trout, Endocrinology 144 (2003) 5215–5226.
[20] H. Kaiya, M. Kojima, H. Hosoda, L.G. Riley, T. Hirano, E.G. Grau, K. Kangawa,Identification of tilapia ghrelin and its effects on growth hormone andprolactin release in the tilapia, Oreochromis mossambicus, Comp. Biochem.Physiol. B 135 (2003) 421–429.
[21] H. Kaiya, B.C. Small, A.L. Bilodeau, B.S. Shepherd, M. Kojima, H. Hosoda, K.Kangawa, Purification, cDNA cloning, and characterization of ghrelin inchannel catfish, Ictalurus punctatus, Gen. Comp. Endocrinol. 143 (2005) 201–210.
[22] H. Kaiya, M. Miyazato, K. Kangawa, R.E. Peter, S. Unniappan, Ghrelin: Amultifunctional hormone in non-mammalian vertebrates, Comp. Biochem.Physiol. A 149 (2008) 109–128.
[23] Y. Kamisaka, G.K. Totland, M. Tagawa, T. Kurokawa, T. Suzuki, M. Tanaka, I.Rønnestad, Ontogeny of cholecystokinin-immunoreactive cells in the digestivetract of Atlantic halibut, Hippoglossus hippoglossus, larvae, Gen. Comp.Endocrinol. 123 (2001) 31–37.
[24] A.S. Kehoe, H. Volkoff, Cloning and characterization of neuropeptide Y (NPY)and cocaine and amphetamine regulated transcript (CART) in Atlantic cod(Gadus morhua), Comp. Biochem. Phys. A 146 (2007) 451–461.
[25] Y. Kobayashi, B.C. Peterson, G.C. Waldbieser, Association of cocaine- andamphetamine-regulated transcript (CART) messenger RNA level, food intake,and growth in channel catfish, Comp. Biochem. Physiol. A 151 (2008) 219–225.
[26] T. Kurokawa, T. Suzuki, H. Hashimoto, Identification of gastrin and multiplecholecystokinin genes in teleost, Peptides 24 (2003) 227–235.
[27] D. Larhammar, Evolution of neuropeptides Y, peptide YY and pancreaticpolypeptide, Regul. Pept. 62 (1996) 1–11.
[28] X.F. Liang, G.Z. Li, W. Yao, L.W. Cheong, W.Q. Liao, Molecular characterizationof neuropeptide Y gene in Chinese perch, an acanthomorph fish, Comp.Biochem. Physiol. B 148 (2007) 55–64.
[29] J.B.K. Loenard, G.C. Waldbieser, J.T. Silverstein, Neuropeptide Y sequence andmessenger RNA distribution in channel catfish (Ictalurus punctatus), Mar.Biotechnol. 3 (2001) 111–118.
[30] M.A. Lopez-Patino, A.I. Guijarro, E. Isorna, M.J. Delgado, M. Alonso-Bedate, N.De Pedro, Neuropeptide Y has a stimulatory action on feeding behavior ingoldfish (Carassius auratus), Eur. J. Pharmacol. 377 (1999) 147–153.
[31] E. MacDonald, H. Volkoff, Neuropeptide Y (NPY), cocaine- and amphetamine-regulated transcript (CART) and cholecystokinin (CCK) in winter skate (Rajaocellata): cDNA cloning, tissue distribution and mRNA expression responses tofasting, Gen. Comp. Endocrinol. 161 (2009) 252–261.
[32] T.H. Moran, K.P. Kinzig, Gastrointestinal satiety signals II. Cholecystokinin, Am.J. Physiol. Gastrointest. Liver Physiol. 286 (2004) (2004) G183–G188.
[33] K. Murashita, T. Kurokawa, Multiple cocaine- and amphetamine-regulatedtranscript (CART) genes in medaka, Oryzias latipes: Cloning, tissue distributionand effect of starvation, Gen. Comp. Endocrinol. 170 (2011) 494–500.
[34] K. Murashita, T. Kurokawa, L.O.E. Ebbesson, S.O. Stefansson, I. Rønnestad,Characterization, tissue distribution, and regulation of agouti-related protein(AgRP), cocaine- and amphetamine-regulated transcript (CART) andneuropeptide Y (NPY) in Atlantic salmon (Salmo salar), Gen. Comp.Endocrinol. 162 (2009) 160–171.
[35] K. Murashita, H. Fukada, H. Hosokawa, T. Masumoto, Changes incholecystokinin and peptide Y gene expression with feeding in yellowtail(Seriola quinqueradiata): relation to pancreatic exocrine regulation, Comp.Biochem. Physiol. B 146 (2007) 318–325.
[36] M. Nakazato, N. Murakami, Y. Date, M. Kojima, H. Matsuo, K. Kangawa, S.Matsukura, A role for ghrelin in the central regulation of feeding, Nature 409(2001) 194–198.
[37] Y.K. Narnaware, P.P. Peyon, X. Lin, R.E. Peter, Regulation of food intake byneuropeptide Y in goldfish, Am. J. Physiol. Regul. Integr. Comp. Physiol. 279(2000) R1025–R1034.
[38] Y.K. Narnaware, R.E. Peter, Effects of food deprivation and refeeding onneuropeptide Y (NPY) mRNA levels in goldfish, Comp. Biochem. Physiol. B 129(2001) 633–637.
[39] P. Nieminen, A.-M. Mustonen, H. Hyvarinen, Fasting reduces plasma leptin andghrelin-immunoreactive peptide concentrations of the burbot (Lota lota) at2 �C but not at 10 �C, Zool. Sci. 20 (2003) 1109–1115.
[40] C. Olsson, G. Aldman, A. Larsson, S. Holmgren, Cholecystokinin affects gastricemptying and stomach motility in the rainbow trout Oncorhynchus mykiss, J.Exp. Biol. 202 (1999) 161–170.
[41] I.S. Parhar, H. Sato, Y. Sakuma, Ghrelin gene in cichlid fish is modulated by sexand development, Biochem. Biophys. Res. Commun. 305 (2003) 169–175.
[42] S.C. Peddu, J.P. Breves, H. Kaiya, E.G. Grau, L.G. Riley, Pre- and postprandialeffects on ghrelin signaling in the brain and on the GH/IGF-I axis in theMozambique tilapia (Oreochromis mossambicus), Gen. Comp. Endocrinol. 161(2009) 412–418.
[43] B.C. Peterson, B.C. Small, G.C. Waldbieser, B.G. Bosworth, Endocrine responsesin fast and slow growing families of channel catfish Ictalurus punctatus, NAJA70 (2008) 240–250.
[44] B.C. Peterson, B.C. Small, Effect of feeding frequency on feed consumption,growth, and feed efficiency in tank-reared Norris and NWAC103 channelcatfish (Ictalurus punctatus), JWAS 37 (2006) 490–495.
[45] P. Peyon, X.W. Lin, B.A. Himick, R.E. Peter, Molecular cloning and expression ofcDNA encoding brain preprocholecystokinin in goldfish, Peptides 19 (1998)199–210.
[46] P. Peyon, H. Saied, X.W. Lin, R.E. Peter, Postprandial, seasonal and sexualvariations in cholecystokinin gene expression in goldfish brain, Mol. Brain Res.74 (1999) 190–196.
[47] M.E. Picha, C.N. Strom, L.G. Riley, A.A. Walker, E.T. Won, W.M. Johnstone, R.J.Borski, Plasma ghrelin and growth hormone regulation in response tometabolic state in hybrid striped bass: effects of feeding, ghrelin andinsulin-like growth factor-I on in vivo and in vitro GH secretion, Gen. Comp.Endocrinol. 161 (2009) 365–372.
[48] L.G. Riley, B.K. Fox, J.P. Breves, H. Kaiya, C.P. Dorough, T. Hirano, E.G. Grau,Absence of effects of short-term fasting on plasma ghrelin and brainexpression of ghrelin receptors in the tilapia, Oreochromis mossambicus, Zool.Sci. 25 (2008) 821–827.
[49] C.R. Rojas-Garcia, S. Morais, I. Rønnestad, Cholecystokinin (CCK) in Atlanticherring (Clupea harengus L.) – ontogeny and effects of feeding and diurnalrhythms, Comp. Biochem. Physiol. A (2011) 455–460.
[50] V.C. Rubio, F.J. Sanchez-Vazquez, J.A. Madrid, Role of cholecystokinin and itsantagonist proglumide on macronutrient selection in European sea bassDicentrarchus labrax, L, Physiol. Behav. 93 (2008) 862–869.
[51] I. Sakata, K. Nakamura, M. Yamazaki, M. Matsubara, Y. Hayashi, K. Kangawa, T.Sakai, Ghrelin-producing cells exist as two types of cells, closed- and opened-type cells, in the rat gastrointestinal tract, Peptides 23 (2002) 531–536.
[52] J.T. Silverstein, J. Breininger, D.G. Baskin, E.M. Plisetskaya, Neuropeptide Y-likegene expression in the salmon brain increases with fasting, Gen. Comp.Endocrinol. 110 (1998) 157–165.

B.C. Peterson et al. / General and Comparative Endocrinology 176 (2012) 231–239 239
[53] J.T. Silverstein, K. Shearer, W. Dickhoff, E.M. Plisetskaya, Regulation of nutrientintake and energy balance in salmon, Aquaculture 177 (1999) 161–169.
[54] J.T. Silverstein, E.M. Plisetskaya, The effects of NPY and insulin and food intakeregulation in fish, Am. Zool. 40 (2000) 296–308.
[55] B.C. Small, C.A. Murdock, A.L. Bilodeau-Bourgeois, B.C. Peterson, G.C.Waldbieser, Stability of reference genes for real-time PCR analyses inchannel catfish (Ictalurus punctatus) tissues under varying physiologicalconditions, Comp. Biochem. Physiol. B 151 (2008) 296–304.
[56] G. Terova, S. Rimoldi, G. Bernardini, R. Gornati, M. Saroglia, Sea bass ghrelin:molecular cloning and mRNA quantification during fasting and refeeding, Gen.Comp. Endocrinol. 155 (2) (2008) 341–351.
[57] S. Unniappan, X. Lin, L. Cervini, J. Rivier, H. Kaiya, K. Kangawa, R.E. Peter,Goldfish ghrelin: molecular characterization of the complementarydeoxyribonucleic acid, partial gene structure and evidence for itsstimulatory role in food intake, Endocrinology 143 (2002) 4143–4146.
[58] S. Unniappan, L.F. Canosa, R.E. Peter, Orexigenic actions of ghrelin in goldfish:feeding-induced changes in brain and gut mRNA expression and serum levels,and responses to central and peripheral injections, Neuroendocrinology 79(2004) 100–108.
[59] E. Valassi, M. Scacchi, F. Cavagnini, Neuroendocrine control of food intake,Nutr. Metab. Cardiovasc. Dis. 18 (2008) 158–168.
[60] R. Valen, A.-E.O. Jordal, K. Murashita, I. Rønnestad, Postprandial effects onappetite-related neuropeptide expression in the brain of Atlantic salmon,Salmo salar, Gen. Comp. Endocrinol. 171 (2011) 359–366.
[61] H. Volkoff, R.E. Peter, Effects of CART peptides on food consumption, feedingand associated behaviors in the goldfish, Carassius auratus: actions on
neuropeptide Y- and orexin A-induced feeding, Brain Res. 887 (2000) 125–133.
[62] H. Volkoff, R.E. Peter, Characterization of two forms of cocaine- andamphetamine-regulated transcript (CART) peptide precursors in goldfish:molecular cloning and distribution, modulation of expression by nutritionalstatus, and interactions with leptin, Endocrinology 142 (2001) 5076–5088.
[63] H. Volkoff, L.F. Canosa, S. Unniappan, J.M. Cerda-Reverter, N.J. Bernier, S.P.Kelly, R.E. Peter, Neuropeptides and the control of food intake in fish, Gen.Comp. Endocrinol. 142 (2005) 3–19.
[64] H. Volkoff, The role of neuropeptide Y, orexins, cocaine and amphetamine-related transcript, cholecystokinin, amylin and leptin in the regulation offeeding in fish, Comp. Biochem. Physiol. B 144 (2006) 325–331.
[65] H. Volkoff, M.Y. Xu, E. MacDonald, L. Hoskins, Aspects of the hormonalregulation of appetite in fish with emphasis on goldfish, Atlantic cod andwinter flounder: notes on actions and responses to nutritional, environmentaland reproductive changes, Comp. Biochem. Physiol. B 153 (2009) 8–12.
[66] H. Volkoff, A.J. Eykelbosh, R.E. Peter, Role of leptin in the control of feeding ofgoldfish Carassius auratus: interactions with cholecystokinin, neuropeptide Yand orexin A, and modulation by fasting, Brain Res. 972 (2003) 90–109.
[67] K. Webb, I.A. Khan, B.S. Nunez, I. Rønnestad, G.J. Holt, Cholecystokinin:molecular cloning and immunohistochemical localization in thegastrointestinal tract of larval red drum, Scianops ocellatus (L.), Gen. Comp.Endocrinol. 166 (2010) 152–159.
[68] A.M. Wren, C.J. Small, C.R. Abbott, W.S. Dhillo, L.J. Seal, M.A. Cohen, R.L.Batterham, S. Taheri, S.A. Stanley, M.A. Ghatei, S.R. Bloom, Ghrelin causeshyperphagia and obesity in rats, Diabetes 50 (2001) 2540–2547.