potential role of tissue transglutaminase in glaucoma...
TRANSCRIPT

Potential Role of Tissue Transglutaminase in GlaucomaFiltering Surgery
Siegfried G. Priglinger,1,2 Claudia S. Alge,1,2 Daniel Kook,1 Martin Thiel,1
Ricarda Schumann,1 Kirsten Eibl,1 Alice Yu,1 Aljoscha S. Neubauer,1 Anselm Kampik,1
and Ulrich Welge-Lussen1
PURPOSE. Scarring of the filtering bleb site is the main cause offailure in glaucoma filtration surgery. In the present study, therole of tissue transglutaminase (tTgase) in the accumulation ofextracellular matrix (ECM) proteins in these scars was investi-gated. Transglutaminases are enzymes capable of cross-linkingECM proteins to proteolysis-resistant complexes.
METHODS. Expression of tTgase, its reaction product �-(�-glu-tamyl)-lysine, and fibronectin and their colocalization wereinvestigated immunohistochemically in failed blebs and in anin vitro trabeculectomy model. Failed blebs were analyzed byRT-PCR for the presence of tTgase mRNA. Human Tenon fibro-blasts (HTFs) were treated with transforming growth factor-�2(TGF-�2). The effect was studied with immunohistochemistry,Northern blot analysis, and Western blot analysis. tTgase activ-ity was assayed by incorporation of biotinylated cadaverineinto fibronectin.
RESULTS. Expression of tTgase and �-(�-glutamyl)-lysine waspresent in all failed blebs. Staining was most prominent at therim of the Tenon cyst. In the in vitro trabeculectomy model,tTgase and �-(�-glutamyl)-lysine were barely present at theincision side of the flap but were perspicuously increased byTGF-�2 treatment. Enzyme and its reaction product were co-localized with fibronectin. Cultured HTFs contained a basallevel of tTgase mRNA. After treatment with TGF-�2, expressionand activity of tTgase significantly increased.
CONCLUSIONS. The findings demonstrated that tTgase is presentand functionally active in failed blebs. Expression and activityof tTgase appeared to be stimulated by TGF-�2, a growth factorknown to be increased in primary open angle glaucoma. Inter-vention at this pathway might open a new approach to preventscarring after glaucoma filtration surgery. (Invest OphthalmolVis Sci. 2006;47:3835–3845) DOI:10.1167/iovs.05-0960
The major determinant of the long-term outcome of glau-coma surgery is the wound-healing response. Excessive
postoperative scarring at the level of the conjunctiva and scler-ostomy sites is associated with poor postoperative pressure
control.1–4 The precise pathogenic mechanism involved inscar tissue formation after glaucoma surgery still is not com-pletely understood. However, human Tenon fibroblasts (HTFs)from the subconjunctival space are known to be the centralplayer in the wound repair and scarring processes after filtrat-ing glaucoma surgery.5 By proliferation, migration, production,and subsequent contraction of extracellular matrix (ECM) com-ponents, HTFs form the scar tissue responsible for filteringfailure of the bleb.5,6
Previous studies regarding scar formation in skin and inproliferative vitreoretinopathy demonstrated transglutaminase-induced irreversible cross-links of ECM that cannot be digestedby any known enzyme.7–10 In keeping with this and the findingthat tissue transglutaminase (tTGase) is basally expressed in theconjunctiva and sclera,11,12 excessive accumulation of ECM inbleb scars could result from the formation of irreversible ECMcross-links in the fibrocellular tissue.
Transglutaminases are calcium-dependent enzymes that cata-lyze the posttranslational modification of proteins through an acyltransfer reaction between the �-carboxamide group of a peptide-bound glutaminyl residue and various amines. Covalent cross-linking using �-(�-glutamyl)-lysine bonds is stable and resistant toenzymatic, chemical, and mechanical disruption.13 Endopepti-dases capable of hydrolyzing the �-(�-glutamyl)-lysine cross-linksformed by transglutaminases have not been described in verte-brates, and even lysosomes do not contain enzymes capable ofsplitting the �-(�-glutamyl)-lysine bonds.14–16 Tissue transglutami-nase (tTgase, type 2) is the most widespread member of thisfamily and is present in many different cell types and tissues, withdiverse functions.17–19 The enzyme plays a role in programmedcell death16 and cell adhesion20 and in normal and abnormalwound healing.21 tTgase controls interaction between the celland its ECM through the cross-linking of proteins such as fibronec-tin,22 and collagen type 3.23 Fibronectin has been shown to beincreased in the ECM of scar tissue,24 and collagen type 3 iswidely recognized as the collagen in new scar tissue.25 Bothproteins play a major role in the healing process after glaucomafiltering surgery26,27 and are produced by HTFs in vitro.28
The present study was designed to investigate the possiblerole of irreversible �-(�-glutamyl)-lysine cross-link of ECM bytTgase in scar tissue of failed blebs. In addition to conductingimmunohistochemistry and molecular biology study on thescar tissue of failed blebs and on an in vitro trabeculectomymodel, we investigated the capacity of HTFs to synthesizetTgase. HTF cell culture is a well-accepted in vitro model forwound repair and scarring processes that occur in the anteriorsegment of the eye.5 In vitro HTFs have been shown to becapable of synthesizing ECM components28 and degrading en-zymes such as matrix metalloproteinases.29–31
Given that TGF-�2 is one of the most potent stimulators ofscarring in the eye and that it is involved in the pathogenesis ofconjunctival scarring and wound healing after filtration sur-gery,32–37 we investigated the influence of TGF-�2 on tTgasesynthesis of cultured HTFs. The activity of extracellular tTgasewas demonstrated by its ability to cross-link fibronectin, an
From the 1Department of Ophthalmology, Ludwig-Maximilians-University, Munich, Germany.
2These authors contributed equally to the work presented hereand should therefore be regarded as equivalent authors.
Supported by Grant DFG WE 2577/2–1.Submitted for publication July 23, 2005; revised January 23 and
February 20, 2006; accepted July 5, 2006.Disclosure: S.G. Priglinger, None; C.S. Alge, None; D. Kook,
None; M. Thiel, None; R. Schumann, None; K. Eibl, None; A. Yu,None; A.S. Neubauer, None; A. Kampik, None; U. Welge-Lussen,None
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be marked “advertise-ment” in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Corresponding author: Ulrich Welge-Lussen, Department of Oph-thalmology, Ludwig-Maximilians-University, Munich, Mathildenstrasse8, 80336 Munich, Germany; [email protected].
Investigative Ophthalmology & Visual Science, September 2006, Vol. 47, No. 9Copyright © Association for Research in Vision and Ophthalmology 3835
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018

ECM component that takes part in the initial healing processafter glaucoma filtering surgery.26,27
MATERIALS AND METHODS
Tissue Samples
Eight samples of scar tissue of failed blebs (Tenon cysts) were obtainedfrom patients undergoing trabeculectomy revision at the Departmentof Ophthalmology of the Ludwig-Maximilians-University Munich (Table1). Methods for securing human tissue were humane, included properconsent and approval, and complied with the Declaration of Helsinki.
Operations were performed by different surgeons using the sametechnique. Conjunctiva and Tenon cyst were carefully separated bysubconjunctival injection of phosphate-buffered saline (PBS; pH 7.4).After opening the Tenon cyst at its border area, the whole cyst over thetrabeculectomy flap and adjacent scar tissue were excised en bloc andput into PBS (pH 7.4) intraoperatively. Afterward Tenon cysts wereeither snap-frozen in liquid nitrogen for mRNA extraction or mountedin OCT mounting medium (Merck, Darmstadt, Germany) and thenstored in liquid nitrogen for cryostat sectioning.
In Vitro Trabeculectomy Model
Six eyes from donors between 35 and 65 years of age were obtainedfrom the Eye Bank of the Ludwig-Maximilians-University, Munich. Only
eyes from donors who gave specific consent for research purposes andwith contraindication for transplantation according to the guidelines ofthe German Federal Physicians’ Association were included.38 All eyeswere preserved in a moist chamber at 4°C within 45 hours after death.All experiments were performed in accordance with the Declaration ofHelsinki. Whole eyes were thoroughly cleansed in 0.9% NaCl solution,immersed in 5% polyvinyl pyrrolidone iodine, and rinsed again in thesodium-chloride solution.
After intravitreal injection of 0.9% NaCl to normalize the intraocularpressure, estimated by palpation to be between 15 and 20 mm Hg,conventional trabeculectomy26,39 was performed in vitro (Fig. 1A).Afterward corneoscleral disks, including the trabeculectomy side,were excised and put into 50-mL tissue culture flasks (Primaria TissueCulture Flask, Falcon; Becton Dickinson Labware, Franklin Lakes, NJ)fixed on a cornea disk holder40 (Figs. 1B, 1C). Organ culture mediumconsisted of Eagle minimum essential medium (MEM) with Earle saltsand 25 mM HEPES buffer (Gibco Life Technologies, Paisley, Scotland,UK), containing 2% fetal calf serum (FCS; Biochrom, Berlin, Germany),2 mM glutamine (Gibco), 100 U/mL penicillin, 0.1 mg/mL streptomy-cin (Gibco), and 0.25 �g/mL amphotericin B (Gibco). After cornealtransplantation, the residual corneoscleral ring with the in vitro trab-eculectomies was kept in organ culture for another 2 months. Three ofthe six trabeculectomy samples now were also treated with 2 ng/mLTGF-�2 (R&D Systems, Wiesbaden, Germany), the maximal total con-centration previously found in the aqueous humor of patients with
TABLE 1. Clinically Relevant Data of Patients Who Underwent Trabeculectomy Revision with TenonCyst Excision
Patient Age, y Diagnosis Antimetabolites Time after TET Local Therapy, y
1 66 PEX Yes 2 y 42 68 POAG No 1 y 153 32 Congenital glaucoma Yes 6 mo 204 55 POAG No 3 mo 55 70 POAG Yes 1.5 y 206 60 POAG No 2 y 107 67 POAG No 2 mo 158 59 POAG No 7 wk 8
TET, trabeculectomy; PEX, pseudoexfoliation glaucoma; POAG, primary open-angle glaucoma.
FIGURE 1. In vitro trabeculectomy.(A) Conventional trabeculectomywas performed in vitro on a humandonor eye before preparation of thecorneoscleral disk. (B) Corneoscleraldisks, including the trabeculectomy,were excised and put into a cultureflask fixed on a cornea disk holder.40
Corneas were stored under sub-merged organ culture conditions in astandard incubator at 37°C and 5%CO2. (C) Magnification of the trab-eculectomy in organ culture.
3836 Priglinger et al. IOVS, September 2006, Vol. 47, No. 9
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018

different types of glaucoma.41 Corneas were stored under submergedorgan culture conditions in a standard incubator at 37°C and 5% CO2,and the medium was changed every 3 days as described pre-viously.42,43
Immunohistochemistry of Tissue Sections
Immunohistochemical double staining for extracellular tTgase andfibronectin and combined staining of �-(�-glutamyl)-lysine and fi-bronectin were performed in sections obtained from scarred blebs(Tenon cysts) and from the in vitro trabeculectomy model. Cryostatsectioning of unfixed scar tissue with a thickness of 8 �m was per-formed. After washing in Tris-buffered saline (TBS, pH 7.2–7.4) andpreincubation with TBS containing 3% bovine serum albumin (BSA;Roche, Mannheim, Germany) to minimize nonspecific staining, thesections were incubated for 2 hours at room temperature (RT) withmouse anti–tissue-transglutaminase (Cub7402; Quartett, Berlin, Ger-many), mouse anti–�-(�-glutamyl)-lysine (CovalAb Oullins, Cedex,France), and rabbit anti-fibronectin (Sigma, Deisenhofen, Germany). Allantibodies were diluted 1:100 (1 �g/100 �L) in TBS containing 3% BSA.After washing in TBS, the sections were incubated with Cy-2–conju-gated goat anti–mouse IgG (Dianova, Hamburg, Germany) and Cy-3–conjugated swine anti–rabbit IgG (Dianova) diluted 1:100 (1 �g/100�L) in blocking buffer for 2 hours at RT.
Control sections were incubated with TBS-BSA without primaryantibody or with a combination of 1:100 (1 �g/100 �L) diluted primaryantibody plus a fivefold (5 �g/100 �L) weight excess of guinea pigtTgase (Sigma). A fluorescence microscope (Leica, Bensheim, Ger-many) was used for visualization of the stained sections. For standardmorphology, either hematoxylin/eosin or phase-contrast microscopyimaging of the tissue sections was performed.
RNA Isolation and RT-Polymerase Chain Reactionof Scarred Blebs and Normal Conjunctiva
Total mRNA of five scarred blebs was extracted using a micro RNA kit(pepGOLD RNAPure; Peqlab, Erlangen, Germany). After confirming thestructural integrity of the total RNA samples of each failed bleb (Tenoncyst) by electrophoresis on 1% agarose gels and subsequent staining with0.5 �mol/mL ethidium bromide, RNA samples were treated with 3 U RQRNase-free DNAse (Promega, Madison, WI) for 35 minutes at 37°C toremove traces of contaminating genomic DNA. The content of RNA wasmeasured by photometric measurement, and the RNA concentration wasadjusted. Using Moloney murine leukemia virus (MMLV) reverse transcrip-tase and oligo(dT)-17 primer (Gibco), first-strand complementary DNA(cDNA) was prepared from total RNA. Quality of RNA and cDNA synthesiswas proven by amplification of the housekeeping gene glyceraldehyde-3-phosphatedehydrogenase (GAPDH).
The PCR of the same quantity of total cDNA was performed in atotal volume of 50 �L with 1 U native Taq polymerase (Eppendorf,Hamburg, Germany). PCR was started with a hot start: 10 minutes for94°C to denature DNA, followed by 36 cycles of 1-minute melting at94°C; 1 minute annealing at the respective annealing temperature, and2 minutes extension at 72°C in a thermocycler (Mastercycler Gradient;Eppendorf). After the last cycle, the polymerization step was extendedfor another 10 minutes to complete all strands. Each PCR was repeatedat least twice.
Primers used were as follows: tTgase forward, 5�-CAGAACAG-CAACCTTCTCATCGAG-3�; tTgase reverse, 5�-TTGGACTCCGTAAG-GCAGTCAC-3�; positions, 1054 to 1811; product size, 758 bp; anneal-ing temperature, 59.7°C (Metabion, Munich, Germany); GAPDH(forward, 5�-CCTGCTTCACCACCTTCTTG-3�; GAPDH reverse, 5�-CAT-CATCTCTGCCCCCTCTG-3�; positions, 416 to 852; product size, 437bp; annealing temperature, 59.7°C (Metabion); fibronectin forward,5�-ATTGGTCCAGACACCATGCG-3�; fibronectin reverse, 5�-AACTTC-CAGGTCCCTCGGAACATC-3�; positions, 3752 to 4288; product size,537 bp; annealing temperature, 56.8°C (MWG-Biotech, Ebersberg, Ger-many).44 The specificity of the PCR product was analyzed by auto-mated DNA sequencing (Sequiserve, Vaterstetten, Germany). PCR per-
formed on each sample of RNA that had not been reverse transcribedto cDNA was used as negative control and showed no amplifiedproduct.
For semiquantitative PCR, the number of cycles was optimized bychecking amplification after each cycle from cycles 25 to 36 for tTgaseand from 20 to 34 for GAPDH. This showed that the 29th cycle was inthe geometric phase for tTgase and GAPDH. PCR amplification prod-ucts were separated by agarose gel electrophoresis and stained withethidium bromide for visualization (LAS-1000 imager workstation; Ray-Test, Pforzheim, Germany). In addition, band intensity was measured(LAS-1000 Imager workstation; RayTest). Quantification was per-formed with the AIDA software package (RayTest). The final amount ofPCR product was expressed as the ratio of the tTgase gene amplified tothat of the GAPDH gene. Experiments were repeated three times.
Human Tenon Fibroblast Isolation and CellCulture Conditions
Human Tenon samples were obtained from tissue explants of threewhite patients (ages 40, 50, and 60 years) who underwent routinebuckling surgery and did not have any topical eye treatment before-hand. Informed consent was obtained from the subjects after explana-tion of the nature and possible consequences of the procedure. HTFswere cultured as previously described45,46 and were maintained inDulbecco modified Eagle medium (DMEM) supplemented with 10%FCS, 2 mM L-glutamine, 100 IU/mL penicillin, 100 �g/mL streptomycin,50 mg/mL gentamicin, and 0.25 mg/mL amphotericin B (all from GibcoLife Technologies) at 37°C with 5% CO2 in air.
For growth factor experiments, second- to fifth-passage HTFs weregrown to confluence. HTFs were then washed three times with TBS,pH 7.2 to 7.4, incubated overnight in serum-free medium, and subse-quently incubated in serum-free DMEM supplemented with 2.0 ng/mLTGF-�2 (R&D Systems) for 24 hours. To prevent the adhesion ofTGF-�2, BSA (Roche) was added during the activation process. Con-trols were incubated under identical conditions without growth factorin the medium. The guidelines of the Declaration of Helsinki werefollowed, and institutional human experimentation committee ap-proval was granted.
Immunohistochemistry of Cell Cultures
For the detection of extracellular tTgase, confluent HTFs grown infour-well plastic chamber slides were incubated for 2.5 hours withserum-free DMEM (Biochrom) containing 0.75 �g/mL monoclonal an-tibody to tTgase (Quartett). Cells were then washed in PBS and fixedin 4% paraformaldehyde in PBS. After blocking in 3% BSA, the cellswere incubated for 15 hours at 4°C with rabbit anti–fibronectin anti-body (Sigma) diluted 1:100 in TBS containing 3% BSA. Samples werethen washed with PBS and incubated with goat anti–mouse IgG Cy-2(Dianova) and swine anti–rabbit IgG Cy-3 (Dianova) diluted 1:100 inblocking buffer for 2 hours at RT.
Northern Blot Analysis
Total RNA was isolated from confluent HTF cultures in 10-cm Petridishes using the guanidinium thiocyanate-phenol-chloroform extrac-tion method (RNA isolation kit; Stratagene, Heidelberg, Germany).Total RNA (3 �g per lane) was denatured and size-fractionated by gelelectrophoresis in 1% agarose gels containing 2.2 M formaldehyde. TheRNA was then vacuum blotted onto a nylon membrane (Roche) andcross-linked (1600 �J; Stratalinker; Stratagene). To assess the amountand quality of the RNA, the membrane was stained with methyleneblue and images were taken (LAS-1000 Imager; RayTest). Prehybridiza-tion, hybridization, and chemiluminescence detection of the digoxige-nin riboprobe were performed as described previously.10 In brief, afterhybridization, the membrane was washed twice with 2 � SSC, 0.1%sodium dodecyl sulfate (SDS) at RT, followed by two washes in 0.1 �SSC, 0.1% SDS for 15 minutes at 68°C. After hybridization and posthy-bridization washes, the membrane was washed for 5 minutes in wash-ing buffer (100 mM maleic acid, 150 mM NaCl; pH 7.5; 0.3% Tween 20)
IOVS, September 2006, Vol. 47, No. 9 Tissue Transglutaminase after Filtering Surgery 3837
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018

and incubated for 60 minutes in blocking solution. Blocking solutioncontained 100 mM maleic acid (pH 7.5), 150 mM NaCl, and 1%blocking reagent (Roche). Alkaline phosphatase–conjugated sheep an-ti–digoxigenin IgG (Roche, Mannheim, Germany) was diluted 1:10,000in blocking solution and used to incubate the membrane for 30 min-utes. The membrane was then washed four times (15 minutes eachtime) in washing buffer and was equilibrated in detection buffer (100mM Tris-HCl; 100 mM NaCl; pH 9.5) for 10 minutes. For chemilumi-nescence detection, a 1:100 dilution (CDP-star; Roche) was made indetection buffer and was used to incubate the filter for 5 minutes at RT.After air drying the semi-dry membrane was sealed in a plastic bag.Chemiluminescence was detected (LAS-1000 Imager workstation; Ray-Test) with exposure times ranging from 10 minutes to 1 hour. Chemi-luminescence signal quantification was performed with a softwarepackage (AIDA; RayTest). All experiments were repeated at least threetimes with HTF cells from different cell lines.
Western Blot of tTgase
HTFs grown in 35-mm tissue culture dishes were washed twice withPBS, collected, and lysed in NP-40 (150 mM NaCl, 50 mM Tris, pH 8.0,1% NP-40) cell lysis buffer. Samples for gel analysis were boiled for 5minutes, and protein content was measured with the use of BCAprotein assay reagent (Pierce, Rockford, IL). Proteins were loaded (2�g/lane) and separated by electrophoresis with the use of 5% SDS-PAGE stacking gel and 8% SDS-PAGE separating gel.47 After gel elec-trophoresis, the proteins were transferred with semi-dry blotting ontoa PVDF membrane (Roche). The membrane was incubated with PBScontaining 0.1% Tween 20 (PBST, pH 7.2) and 5% BSA for 1 hour. Theprimary antibody (tTgase 1:2000, Cub7402; Quartett) was then addedand allowed to react overnight at RT. After the membrane was washedthree times in PBST, it was incubated with an alkaline phosphatase–conjugated swine-anti–mouse antibody (Dianova; diluted 1:20,000) for30 minutes. The membrane was then washed four times (15 minuteseach) in washing buffer (100 mM maleic acid, 150 mM NaCl, pH 7.5,0.3% Tween 20). The membrane was equilibrated in detection buffer(100 mM Tris-HCl; 100 mM NaCl; pH 9.5) for 10 minutes. Visualizationof the alkaline phosphatase was achieved using chemiluminescence.CDP-star (Roche) was diluted 1:100 in detection buffer, and the filterwas incubated for 5 minutes at RT. After air drying, the semi-drymembrane was sealed in a plastic bag. Chemiluminescence was de-tected (LAS-1000 Imager workstation; RayTest) with exposure timesfrom 1 minute to 5 minutes. Quantification of chemiluminescence wasperformed with software (AIDA; RayTest). All experiments were re-peated at least three times using HTF cells derived from differentdonors.
tTgase Activity of Cultured HTFs
tTgase activity was measured by the incorporation of biotinylatedcadaverine into fibronectin.48 For this assay, 96-well plates were pre-coated with plasma fibronectin (5 �g/mL; Sigma) and were incubatedovernight at 4°C. Twenty-four hours before seeding, HTFs were treatedwith 2.0 ng/mL TGF-�2 (R&D Systems). Untreated and cytokine-treatedHTFs were then plated at a density of 2 � 105 cells/mL in 100 �Lserum-free DMEM (Biochrom) in the presence of 0.1 mM biotinylatedcadaverine (Mobi-Tec, Gottingen, Germany). HTFs were allowed toincubate on the fibronectin-coated plates for different time periods (0,5, 10, 20, 40, 60, 90, or 120 minutes) at 37°C and were then washedtwice with PBS containing 3 mM EDTA. As a negative control, fibronec-tin-coated 96-well plates were incubated with 100 �L serum-freeDMEM containing 0.1 mM biotinylated cadaverine alone. The deter-gent solution (100 �L) consisting of 0.1% deoxycholate in PBS con-taining 3 mM EDTA was then added to each well and was incubatedwith gentle shaking for 20 minutes. The supernatant was discarded,and the remaining fibronectin layer was washed three times with 0.1 MTris-HCl, pH 7.4. Reaction was then blocked with 3% BSA in Tris-HClbuffer for 30 minutes at 37°C and was washed three times withTris-HCl buffer. The incorporated biotinylated cadaverine was revealed
with a 1:5000 dilution of peroxidase conjugate (Extravidin; Sigma)incubated for 1 hour at 37°C. After three washes with Tris-HCl, thefibronectin layer was incubated for 20 minutes at RT in 200 �Lsubstrate solution (one tablet of tetramethylbenzidine was dissolved in1 mL dimethylsulfoxide and added to 9 mL 0.05 M phosphate-citratebuffer, pH 5.0; 2 �L fresh 30% hydrogen peroxide was added per 10mL substrate buffer solution). Color development was stopped byadding 50 �L stop solution (25% H2SO4) to each well. Optical densitywas determined with the use of an ELISA reader (Molecular Devices,Garching, Germany) set at 450 nm. Experiments were performed ninetimes.
Statistical Analysis
Statistical analysis was based on Wilcoxon matched-pair signed-ranktest, and P � 0.05 was considered statistically significant.
RESULTS
Scar Tissue of Failed Blebs
tTgase mRNA and Protein in Failed Blebs. Eight samplesof failed blebs were obtained from patients undergoing trab-eculectomy revision. Six (75%) of those patients had primaryopen angle glaucoma, and the others had pseudoexfoliationglaucoma and congenital glaucoma, respectively (Table 1).Mean age was 59.6 (�12.3) years, mean follow-up time aftertrabeculectomy was 11.5 (�7.1) months, and mean duration oflocal therapy was 12 (�6.3) years.
With the RT-PCR technique, we could demonstrate thepresence of tTgase mRNA in all investigated scar tissue samplesof failed blebs (Fig. 2). The actual size of the PCR product oftTgase was found to be close to the theoretically expectedvalue (on the basis of the primer position 758 bp) and showedthe expected sequence (data not shown). SemiquantitativeRT-PCR analysis revealed expression of tTgase in scar tissue offailed blebs three times higher than in normal conjunctiva (Fig.2). Results of PCR performed on negative controls, when the
FIGURE 2. tTgase expression in failed blebs compared with normalconjunctiva. Agarose gel electrophoresis and ethidium bromide stain-ing of PCR products amplified from human conjunctiva (lane 1) andfrom scar tissue of failed blebs (lane 2). mRNA expression of tTgasewas measured by using semiquantitative RT-PCR. Typical results areshown for experiments repeated three times. The number below eachband shows the ratio of the optical density of the tTgase PCR productnormalized to the GAPDH amplicon of the same cDNA. MW: DNAstandard lane is shown at the left of the gel.
3838 Priglinger et al. IOVS, September 2006, Vol. 47, No. 9
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018
reverse transcriptase step was omitted, were negative (data notshown).
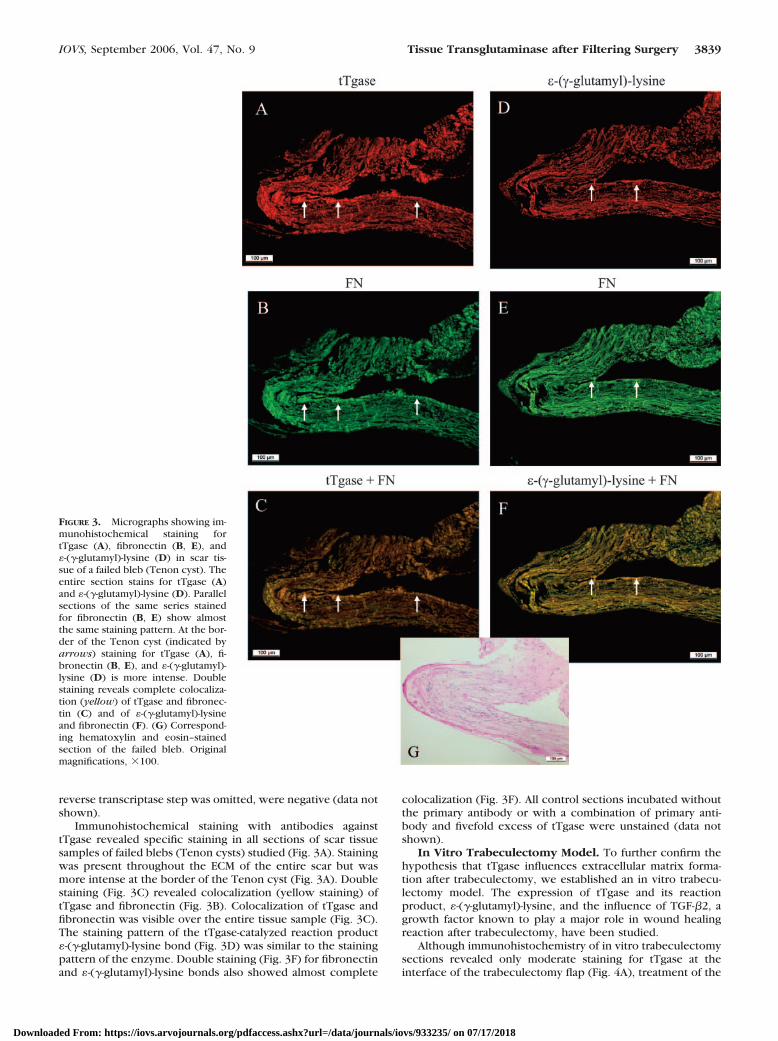
Immunohistochemical staining with antibodies againsttTgase revealed specific staining in all sections of scar tissuesamples of failed blebs (Tenon cysts) studied (Fig. 3A). Stainingwas present throughout the ECM of the entire scar but wasmore intense at the border of the Tenon cyst (Fig. 3A). Doublestaining (Fig. 3C) revealed colocalization (yellow staining) oftTgase and fibronectin (Fig. 3B). Colocalization of tTgase andfibronectin was visible over the entire tissue sample (Fig. 3C).The staining pattern of the tTgase-catalyzed reaction product�-(�-glutamyl)-lysine bond (Fig. 3D) was similar to the stainingpattern of the enzyme. Double staining (Fig. 3F) for fibronectinand �-(�-glutamyl)-lysine bonds also showed almost complete
colocalization (Fig. 3F). All control sections incubated withoutthe primary antibody or with a combination of primary anti-body and fivefold excess of tTgase were unstained (data notshown).
In Vitro Trabeculectomy Model. To further confirm thehypothesis that tTgase influences extracellular matrix forma-tion after trabeculectomy, we established an in vitro trabecu-lectomy model. The expression of tTgase and its reactionproduct, �-(�-glutamyl)-lysine, and the influence of TGF-�2, agrowth factor known to play a major role in wound healingreaction after trabeculectomy, have been studied.
Although immunohistochemistry of in vitro trabeculectomysections revealed only moderate staining for tTgase at theinterface of the trabeculectomy flap (Fig. 4A), treatment of the
FIGURE 3. Micrographs showing im-munohistochemical staining fortTgase (A), fibronectin (B, E), and�-(�-glutamyl)-lysine (D) in scar tis-sue of a failed bleb (Tenon cyst). Theentire section stains for tTgase (A)and �-(�-glutamyl)-lysine (D). Parallelsections of the same series stainedfor fibronectin (B, E) show almostthe same staining pattern. At the bor-der of the Tenon cyst (indicated byarrows) staining for tTgase (A), fi-bronectin (B, E), and �-(�-glutamyl)-lysine (D) is more intense. Doublestaining reveals complete colocaliza-tion (yellow) of tTgase and fibronec-tin (C) and of �-(�-glutamyl)-lysineand fibronectin (F). (G) Correspond-ing hematoxylin and eosin–stainedsection of the failed bleb. Originalmagnifications, �100.
IOVS, September 2006, Vol. 47, No. 9 Tissue Transglutaminase after Filtering Surgery 3839
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018

in vitro trabeculectomy with TGF-�2 led to higher intensity oftTgase staining (Fig. 4E). Staining with antibodies against thetTgase-catalyzed reaction product �-(�-glutamyl)-lysine bondshowed almost the same patterns. Although little specific stain-ing for �-(�-glutamyl)-lysine bond was seen in untreated in vitrotrabeculectomy samples (Fig. 4I), TGF-�2 treatment led todistinct staining along the trabeculectomy flap interface (Fig.4M). Immunohistochemistry of the sections from the sameseries also showed pronounced staining at the trabeculectomyflap interface for fibronectin (Fig. 4B, 4J). Treatment withTGF-�2 further enhanced fibronectin expression (Fig. 4F, 4N).Double staining for tTgase and fibronectin and for �-(�-glu-
tamyl)-lysine bonds and fibronectin, respectively, revealed co-localization along the interface of the trabeculectomy flaps(Fig. 4C, 4G, 4K, 4O). All investigated blebs showed similarstaining patterns.
Cell Culture of Human Tenon Fibroblasts
Induction of Fibronectin and tTgase mRNA and Pro-tein by TGF-�2. Northern blot analysis of untreated HTFsshowed a single faint band of 7.7 kb after hybridization with anantisense fibronectin RNA probe (Fig. 5A) and of 3.5 kb afterhybridization with an antisense tTgase RNA probe, respectively
FIGURE 4. TGF-�2 induces tTgase and �-(�-glutamyl)-lysine in an in vitro trabeculectomy model. (A) Immunohistochemical staining for tTgase ina cross-section of an in vitro trabeculectomy. (B) Section from the same series stained for fibronectin (FN). Staining for tTgase (A) and FN (B) ispronounced at the margin of the trabeculectomy flap (arrows). (C) Double staining of tTgase and FN demonstrates partial colocalization (yellow).Treatment of the in vitro trabeculectomy model with TGF-�2 revealed an increase in tTGase (E) and FN (F) expression. (G) Double staining showsbroad colocalization (yellow) of tTgase and FN. (I, M) Immunohistochemical staining of the in vitro trabeculectomy sections for the tTgase reactionproduct �-(�-glutamyl)-lysine bonds. As with the staining for tTgase (A, E) and FN (B, F, J, N), �-(�-glutamyl)-lysine is most pronounced at the marginof the trabeculectomy incision (arrows). Although staining for �-(�-glutamyl)-lysine (I) was only marginal in untreated in vitro trabeculectomy flaps,TGF-�2 treatment led to a distinct increase in �-(�-glutamyl)-lysine staining (M). (K, O) Double staining of parallel sections with FN and�-(�-glutamyl)-lysine reveals broad colocalization (yellow). (D, H, L). Phase-contrast images of the corresponding cryosections. Original magnifi-cations, �100. (P) Illustration demonstrating a cross-section of a trabeculectomy flap with a box marking the region of magnification as shown inthe (immuno) histologic sections (A-O). Representative immunohistochemical images of cryosections obtained from six patients with failed blebsare shown.
3840 Priglinger et al. IOVS, September 2006, Vol. 47, No. 9
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018

(Fig. 6A). Treatment of cultured HTFs with TGF-�2 markedlyincreased the levels of fibronectin (Fig. 5A) and tTgase mRNA(Fig. 6A). The levels of fibronectin mRNA in TGF-�2–treatedHTFs were approximately fourfold higher (4.0 � 0.8; P � 0.05)than those detected in the untreated control cells, and thelevels of tTgase mRNA in TGF-�2 treated HTFs were approxi-mately fivefold higher (4.95 � 0.52; P � 0.05) than thosedetected in the untreated control cells. Representative blotsare shown in Figures 5A and 6A.
To validate that this increase in mRNA transcription trans-lates into increased protein synthesis, whole cellular proteinextracts were analyzed by Western blotting (Figs. 5C, 6C).Immunoblot analysis revealed a single band at the molecularmass of approximately 240 kDa for fibronectin and of approx-imately 80 kDa for tTgase, respectively. Treatment withTGF-�2 led to an increase of fibronectin by a factor of 2 to 3(2.5 � 0.5; P � 0.05) and of tTgase by a factor of 4 to 5 (4.53 �0.9; P � 0.05) in cultured HTF cells. Representative blots ofexperiments that were repeated at least three times are shownin Figures 5C and 6C.
Extracellular tTgase Activity. To demonstrate that tTgaseis functionally active and further influenced by TGF-�, assay
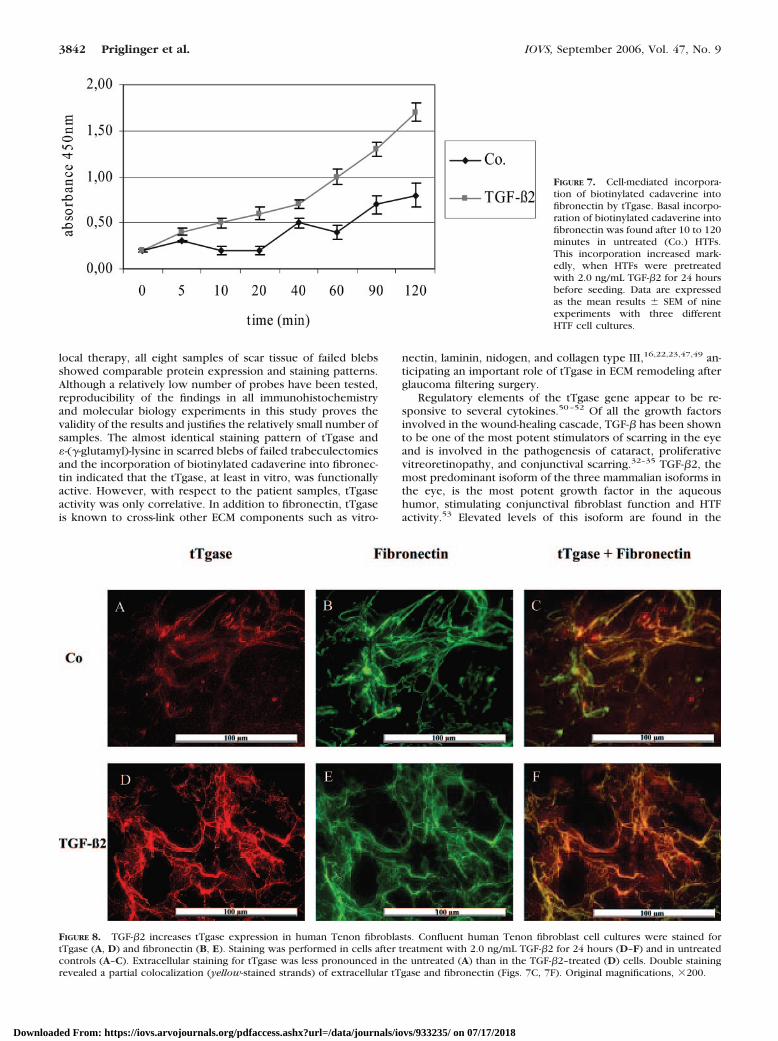
screening for the incorporation of biotinylated cadaverine intofibronectin by tTgase activity was performed. Basal incorpora-tion of biotinylated cadaverine into fibronectin was found after10 to 120 minutes (Fig. 7). This incorporation increased mark-edly, when HTFs were pretreated for 24 hours with 2.0 ng/mLTGF-�2 before seeding (Fig. 7).
Colocalization of tTgase and Fibronectin. To demon-strate extracellular tTgase immunohistochemically, anti–tTgaseantibody was added to the culture medium of unfixed HTFs.Only weak staining of tTgase developed in untreated HTFs (Fig.8A). Treatment with TGF-�2 increased the amount of extracel-lular tTgase (Fig. 8D). Double staining of fibronectin (Figs. 8B,8E) and tTgase revealed partial colocalization of extracellulartTgase and fibronectin (Figs. 8C, 8F).
DISCUSSION
To the best of our knowledge, this is the first study to demon-strate the presence of tTgase and its end product, �-(�-glu-tamyl)-lysine, in scarred tissue of failed trabeculectomy blebsand in an in vitro trabeculectomy model. Irrespective of patientage, mean follow-up time after trabeculectomy, and duration of
FIGURE 5. TGF-�2 induces fibronectin expression in human Tenonfibroblasts. (A) Representative Northern blot analysis of fibronectin(FN) mRNA expression in confluent HTF cells 24 hours after treatmentwith 2 ng/mL TGF-�2. Levels of FN mRNA in TGF-�2–treated HTFswere 4.0 (�0.8)-fold higher than those detected in the untreatedcontrol cells. (B) Methylene blue staining of the 28S and 18S rRNAbands to demonstrate relative integrity and loading of RNA (C) Repre-sentative Western blot analysis of FN in HTF cell culture treated asdescribed for Northern blot analysis. The number below each bandshows the chemiluminescence measurement (2.5 [�0.5] ma). All ex-periments were repeated four times with HTF cells from different celllines. Co, control; RDI, relative densitometric intensity (normalized tovalues of untreated controls [�1.0]; differences in 28S rRNA wereconsidered for Northern blot quantification. For Western blot, equalamounts were loaded per lane).
FIGURE 6. TGF-�2 induces tTgase expression in human tenon fibro-blasts. (A) Representative Northern blot analysis of tTgase mRNA inconfluent HTF cells 24 hours after treatment with 2.0 ng/mL TGF-�2.Levels of tTgase mRNA in TGF-�2–treated HTFs were 4.95 (�0.52)-foldhigher than those detected in the untreated control cells. (B) Methyl-ene blue staining of the 28S and 18S rRNA bands to demonstraterelative integrity and loading of RNA (C) Representative Western blotanalysis of tTgase in HTF cell culture treated as described for Northernblot analysis. The number below each band shows the chemilumines-cence measurement (4.53 [�0.9] ma). All experiments were repeatedfour times using HTF cells from different cell lines. Co indicatescontrol; RDI, relative densitometric intensity (normalized to values ofuntreated controls [�1.0]; differences in 28S rRNA were consideredfor Northern blot quantification. For Western blot, equal amounts wereloaded per lane.)
IOVS, September 2006, Vol. 47, No. 9 Tissue Transglutaminase after Filtering Surgery 3841
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018
local therapy, all eight samples of scar tissue of failed blebsshowed comparable protein expression and staining patterns.Although a relatively low number of probes have been tested,reproducibility of the findings in all immunohistochemistryand molecular biology experiments in this study proves thevalidity of the results and justifies the relatively small number ofsamples. The almost identical staining pattern of tTgase and�-(�-glutamyl)-lysine in scarred blebs of failed trabeculectomiesand the incorporation of biotinylated cadaverine into fibronec-tin indicated that the tTgase, at least in vitro, was functionallyactive. However, with respect to the patient samples, tTgaseactivity was only correlative. In addition to fibronectin, tTgaseis known to cross-link other ECM components such as vitro-
nectin, laminin, nidogen, and collagen type III,16,22,23,47,49 an-ticipating an important role of tTgase in ECM remodeling afterglaucoma filtering surgery.
Regulatory elements of the tTgase gene appear to be re-sponsive to several cytokines.50–52 Of all the growth factorsinvolved in the wound-healing cascade, TGF-� has been shownto be one of the most potent stimulators of scarring in the eyeand is involved in the pathogenesis of cataract, proliferativevitreoretinopathy, and conjunctival scarring.32–35 TGF-�2, themost predominant isoform of the three mammalian isoforms inthe eye, is the most potent growth factor in the aqueoushumor, stimulating conjunctival fibroblast function and HTFactivity.53 Elevated levels of this isoform are found in the
FIGURE 7. Cell-mediated incorpora-tion of biotinylated cadaverine intofibronectin by tTgase. Basal incorpo-ration of biotinylated cadaverine intofibronectin was found after 10 to 120minutes in untreated (Co.) HTFs.This incorporation increased mark-edly, when HTFs were pretreatedwith 2.0 ng/mL TGF-�2 for 24 hoursbefore seeding. Data are expressedas the mean results � SEM of nineexperiments with three differentHTF cell cultures.
FIGURE 8. TGF-�2 increases tTgase expression in human Tenon fibroblasts. Confluent human Tenon fibroblast cell cultures were stained fortTgase (A, D) and fibronectin (B, E). Staining was performed in cells after treatment with 2.0 ng/mL TGF-�2 for 24 hours (D–F) and in untreatedcontrols (A–C). Extracellular staining for tTgase was less pronounced in the untreated (A) than in the TGF-�2–treated (D) cells. Double stainingrevealed a partial colocalization (yellow-stained strands) of extracellular tTgase and fibronectin (Figs. 7C, 7F). Original magnifications, �200.
3842 Priglinger et al. IOVS, September 2006, Vol. 47, No. 9
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018

aqueous humor of glaucomatous eyes compared with normaleyes.5,54–58 In a previous study, Picht et al.41 demonstrated atendency for favorable bleb development in patients with nor-mal TGF-�2 levels, indicating that there might be some rela-tionship between bleb formation and TGF-�2 levels, especiallyfor patients with primary open angle glaucoma. Analogous tothis previous study, most (75%) failed blebs in this study wereobtained from patients with primary open angle glaucoma(Table 1). Furthermore, 10-fold higher concentrations ofTGF-�2 were found in the tear fluid.59 Given that TGF-�2 playsa major role in the pathogenesis of glaucoma and conjunctivalscarring treatment of cultured scleral fibroblasts or in vitrotrabeculectomy with TGF-�2 seems to be an ideal model tomimic the in vivo condition after glaucoma surgery.
In our study cultured HTFs and in vitro trabeculectomies,when exposed to TGF-�2, showed increased tTgase expressionand enhanced cross-linking of the ECM component fibronectinthrough �-(�-glutamyl)-lysine bonds. These results suggest thatTGF-�2, in addition to stimulating proliferating HTFs to pro-duce numerous ECM components, induces irreversible cross-links of these ECM components, thereby forming the fibrocel-lular scar tissue of failed blebs. In keeping with this hypothesisin a recent experimental animal study,60 postoperative use ofthe TGF-� antibody CAT-152 significantly improved outcomesafter surgery and reduced subconjunctival scarring.
In the present study we used a human in vitro trabeculec-tomy model to study conjunctival scarring after trabeculec-tomy. The major drawback of this in vitro approach comparedwith in vivo animal models was the lack of blood supply,which might have significantly contributed to the inflamma-tory response. However, as evidenced by several previousstudies, findings obtained from animal models do not necessar-ily translate into the human in vivo condition. For instance,Cordeiro et al.33 demonstrated successful inhibition of con-junctival scarring in a rabbit animal model of aggressive con-junctival scarring with significant improvement of glaucomafiltering surgery by the use of recombinant neutralizing anti-bodies to human TGF-�2 (rhAnti-TGF-�2 mAb). However, invivo application of rhAnti-TGF-�2 mAb could not confirm thedata of the animal model (Khaw PT, et al. IOVS 2005;46:ARVOE-Abstract 87). In view of these experiences, we do think theapplication of a human in vitro trabeculectomy model is anappropriate approach to study conjunctival scarring after tra-beculectomy. However, despite the apparent limitations ofanimal models, future studies in animals will be needed toconfirm the present findings in in vivo conditions.
Current established concepts of therapy to prevent theonset of increased fibrocellular scarring after trabeculectomy,such as intraoperative administration of the antiproliferativeagents 5-fluourouracil (5-FU) and mitomycin C (MMC), haveincreased the success rate of filtration surgery, and these tech-niques have been accepted in clinical practice.45,61–68 How-ever, despite the use of these antiproliferative agents, failurecan occur later in the postoperative period,5,6,66,69,70 and theadministration of MMC and 5-FU, which induce widespreadcell death and apoptosis, can result in corneal erosions, cysticavascular blebs, chronic hypotony with maculopathy, and en-dophthalmitis.65,68,71–73
Therefore, more physiologic antiscarring agents are neededfor the postoperative prevention of bleb failure and increasedintraocular pressure. Although fully speculative at this point,the inhibition of tTgase might offer a future therapeutic ap-proach to prevent irreversible ECM cross-linking of TGF-�–activated HTFs. Possible agents are competitive inhibitors oftransglutaminase such as putrescine, which decreases woundbreaking strength in incisional skin wounds by topical appli-cation,8 and spermidine, which decreases the healing of gastricand duodenal stress ulcers by inhibiting transglutaminase.74
Furthermore, tranglutaminase-mediated effects can be inhib-ited by specific antibodies,75 and some pharmaceutical com-pounds even inhibit transglutaminase not only competitivelybut irreversibly.76 The use of such components may preventincreased scar tissue formation after trabeculectomy and there-fore offer a new, specific therapeutic strategy to prevent blebfailure after filtrating glaucoma surgery.
Acknowledgments
The authors thank Katja Obholzer and Harald Kroehn for experttechnical assistance.
References
1. Addicks EM, Quigley HA, Green WR, Robin AL. Histologic charac-teristics of filtering blebs in glaucomatous eyes. Arch Ophthalmol.1983;101:795–798.
2. Hitchings RA, Grierson I. Clinico pathological correlation in eyeswith failed fistulizing surgery. Trans Ophthalmol Soc UK. 1983;103:84–88.
3. Jay JL. Rational choice of therapy in primary open angle glaucoma.Eye. 1992;6:243–247.
4. Migdal C, Gregory W, Hitchings R. Long-term functional outcomeafter early surgery compared with laser and medicine in open-angle glaucoma. Ophthalmology. 1994;101:1651–1656.
5. Khaw PT, Occleston NL, Schultz G, Grierson I, Sherwood MB,Larkin G. Activation and suppression of fibroblast function. Eye.1994;8:188–195.
6. Occleston NL, Daniels JT, Tarnuzzer RW, et al. Single exposures toantiproliferatives: long-term effects on ocular fibroblast wound-healing behavior. Invest Ophthalmol Vis Sci. 1997;38:1998–2007.
7. Bowness JM, Tarr AH, Wong T. Increased transglutaminase activityduring skin wound healing in rats. Biochim Biophys Acta. 1988;967:234–240.
8. Dolynchuk KN, Bendor-Samuel R, Bowness JM. Effect of pu-trescine on tissue transglutaminase activity in wounds: decreasedbreaking strength and increased matrix fucoprotein solubility.Plast Reconstr Surg. 1994;93:567–573.
9. Haroon ZA, Hettasch JM, Lai TS, Dewhirst MW, Greenberg CS.Tissue transglutaminase is expressed, active, and directly involvedin rat dermal wound healing and angiogenesis. FASEB J. 1999;13:1787–1795.
10. Priglinger SG, May CA, Neubauer AS, et al. Tissue transglutaminaseas a modifying enzyme of the extracellular matrix in PVR mem-branes. Invest Ophthalmol Vis Sci. 2003;44:355–364.
11. Welge-Lussen U, May CA, Lutjen-Drecoll E. Induction of tissuetransglutaminase in the trabecular meshwork by TGF-beta1 andTGF-beta2. Invest Ophthalmol Vis Sci. 2000;41:2229–2238.
12. Raghunath M, Cankay R, Kubitscheck U, et al. Transglutaminaseactivity in the eye: cross-linking in epithelia and connective tissuestructures. Invest Ophthalmol Vis Sci. 1999;40:2780–2787.
13. Folk JE, Finlayson JS. The �-(�-glutamyl) lysine crosslink and thecatalytic role of transglutaminases. Adv Protein Chem. 1977;31:1–133.
14. Folk JE. Transglutaminases. Annu Rev Biochem. 1980;49:517–531.15. Fesus L, Thomazy V, Autuori F, Ceru MP, Tarcsa E, Piacentini M.
Apoptotic hepatocytes become insoluble in detergents and chao-tropic agents as a result of transglutaminase action. FEBS Lett.1989;245:150–154.
16. Fesus L, Davies PJA, Piacentini M. Apoptosis-molecular mecha-nisms in programmed cell death. Eur J Cell Biol. 1991;56:170–177.
17. Aeschlimann D, Paulsson M. Cross-linking of laminin-nidogen com-plexes by tissue transglutaminase: a novel mechanism for base-ment membrane stabilization. J Biol Chem. 1991;266:15308–15317.
18. Aeschlimann D, Wetterwald A, Fleisch H, Paulsson M. Expressionof tissue transglutaminase in skeletal tissues correlates with eventsof terminal differentiation of chondrocytes. J Cell Biol. 1993;120:1461–1470.
19. Thomazy V, Fesus L. Differential expression of tissue transglutami-nase in human cells. Cell Tissue Res. 1989;225:215–224.
IOVS, September 2006, Vol. 47, No. 9 Tissue Transglutaminase after Filtering Surgery 3843
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018

20. Cai D, Ben T, De Luca LM. Retinoids induce tissue transglutami-nase in NIH-3T3 cells. Biochem Biophys Res Commun. 1991;175:1119–1124.
21. Verderio EA, Johnson T, Griffin M. Tissue transglutaminase innormal and abnormal wound healing: review article. Amino Acids.2004;26:387–404.
22. Greenberg CS, Birckbichler PJ, Rice RH. Transglutaminases: mul-tifunctional cross-linking enzymes that stabilize tissues. FASEB J.1991;5:3071–3077.
23. Bowness JM, Folk JE, Timpl R. Identification of a substrate site forliver transglutaminase on the aminopropeptide of type III collagen.J Biol Chem. 1987;262:1022–1024.
24. Chen W, Fu X, Sun X, Sun T, Zhao Z, Sheng Z. Analysis ofdifferentially expressed genes in keloids and normal skin withcDNA microarray. J Surg Res. 2003;113:208–216.
25. Zhang LQ, Laato M, Muona P, Kalimo H, Peltonen J. Normal andhypertrophic scars: quantification and localization of messengerRNAs for type I, III and VI collagens. Br J Dermatol. 1994;130:453–459.
26. Skuta GL, Parrish RK 2nd. Wound healing in glaucoma filteringsurgery. Surv Ophthalmol. 1987;32:149–170.
27. Fourman S, Wiley L. Tissue plasminogen activator modifies healingof glaucoma filtering surgery in rabbits. Ophthalmic Surg. 1991;22:718–723.
28. Kottler UB, Junemann AG, Aigner T, Zenkel M, Rummelt C,Schlotzer-Schrehardt U. Comparative effects of TGF-beta 1 andTGF-beta 2 on extracellular matrix production, proliferation, mi-gration, and collagen contraction of human Tenon’s capsule fibro-blasts in pseudoexfoliation and primary open-angle glaucoma. ExpEye Res. 2005;80:121–134.
29. Mietz H, Esser JM, Welsandt G, et al. Latanoprost stimulates secre-tion of matrix metalloproteinases in Tenon fibroblasts both in vitroand in vivo. Invest Ophthalmol Vis Sci. 2003;44:5182–5188.
30. Kawashima Y, Saika S, Yamanaka O, Okada Y, Ohkawa K, OhnishiY. Immunolocalization of matrix metalloproteinases and tissueinhibitors of metalloproteinases in human subconjunctival tissues.Curr Eye Res. 1998;17:445–451.
31. Daniels JT, Cambrey AD, Occleston NL, et al. Matrix metallopro-teinase inhibition modulates fibroblast-mediated matrix contrac-tion and collagen production in vitro. Invest Ophthalmol Vis Sci.2003;44:1104–1110.
32. Cordeiro MF, Reichel MB, Gay JA, D’Esposita F, Alexander RA,Khaw PT. Transforming growth factor-beta1, -beta2, and -beta3 invivo: effects on normal and mitomycin C-modulated conjunctivalscarring. Invest Ophthalmol Vis Sci. 1999;40:1975–1982.
33. Cordeiro MF, Gay JA, Khaw PT. Human anti-transforming growthfactor-beta2 antibody: a new glaucoma anti-scarring agent. InvestOphthalmol Vis Sci. 1999;40:2225–2234.
34. Connor TB, Roberts AB, Sporn MB, et al. Correlation of fibrosis andtransforming growth factor-beta type 2 levels in the eye. J ClinInvest. 1989;83:1661–1666.
35. Hales AM, Chamberlain CG, McAvoy JW. Cataract induction inlenses cultured with transforming growth factor-�. Invest Oph-thalmol Vis Sci. 1995;36:1709–1713.
36. Denk PO, Hoppe J, Hoppe V, Knorr M. Effect of growth factors onthe activation of human Tenon’s capsule fibroblasts. Curr Eye Res.2003;27:35–44.
37. Mietz H, Chevez-Barrios P, Lieberman MW. A mouse model tostudy the wound healing response following filtration surgery.Graefes Arch Clin Exp Ophthalmol. 1998;236:467–475.
38. Bundesartzekammer. Richtlinien zum Fuhren einer Hornhautbank.Dt �rztebl. 2000;97:B1805–B1807.
39. Jones E, Clarke J, Khaw PT. Recent advances in trabeculectomytechnique. Curr Opin Ophthalmol. 2005;16:107–113.
40. Bohnke M. A new system for storing donor corneas. Klin MonatsblAugenheilkd. 1991;198:135–137.
41. Picht G, Welge-Luessen U, Grehn F, Lutjen-Drecoll E. Transforminggrowth factor beta 2 levels in the aqueous humor in different typesof glaucoma and the relation to filtering bleb development.Graefes Arch Clin Exp Ophthalmol. 2001;239:199–207.
42. Priglinger SG, Neubauer AS, May CA, et al. Optical coherencetomography for the detection of laser in situ keratomileusis indonor corneas. Cornea. 2003;22:46–50.
43. Neubauer AS, Priglinger SG, Thiel MJ, et al. Sterile structuralimaging of donor cornea by optical coherence tomography. Cor-nea. 2002;21:490–494.
44. Kornblihtt AR, Vide-Pedersen K, Baralle FE. Isolation and charac-terisation of cDNA clones for human and bovine fibronectins. ProcNatl Acad Sci USA. 1983;80:3218–3222.
45. Khaw PT, Ward S, Porter A, Grierson I, Hitchings RA, Rice NS. Thelong-term effects of 5-fluorouracil and sodium butyrate on humanTenon’s fibroblasts. Invest Ophthalmol Vis Sci. 1992;33:2043–2052.
46. Eibl KH, Banas B, Kook D, et al. Alkylphosphocholines: a newtherapeutic option in glaucoma filtration surgery. Invest Ophthal-mol Vis Sci. 2004;45:2619–2624.
47. Sane DC, Moser TL, Pippen AM, Parker CJ, Achyuthan KE, Green-berg CS. Vitronectin is a substrate for transglutaminases. BiochemBiophys Res Commun. 1988;157:115–120.
48. Laemmli UK. Cleavage of structural proteins during the assemblyof the head of bacteriophage T4. Nature. 1970;227:680–685.
49. Jones RA, Nicholas B, Mian S, Davies PJA, Griffin M. Reducedexpression of tissue transglutaminase in a human endothelial cellline leads to changes in cell spreading, cell adhesion and reducedpolymerisation of fibronectin. J Cell Sci. 1997;110:2461–2472.
50. Vollberg TM, George MD, Nervi C, Jetten AM. Regulation of typeI and type II transglutaminase in normal human bronchial epithe-lial and lung carcinoma cells. Am J Respir Cell Mol Biol. 1992;7:10–18.
51. Ikura K, Shinagawa R, Suto N, Sasaki R. Increase caused by inter-leukin-6 in promoter activity of guinea pig liver transglutaminasegene. Biosci Biotechnol Biochem. 1994;58:1540–1541.
52. Johnson K, Hashimoto S, Lotz M, Pritzker K, Terkeltaub R. Inter-leukin-1 induces pro-mineralizing activity of cartilage tissue trans-glutaminase and factor XIIIa. Am J Pathol. 2001;159:149–163.
53. Cordeiro MF, Bhattacharya SS, Schultz GS, Khaw PT. TGF-beta1,-beta2, and -beta3 in vitro: biphasic effects on Tenon’s fibroblastcontraction, proliferation, and migration. Invest Ophthalmol VisSci. 2000;41:756–763.
54. Lutty GA, Merges C, Threlkeld AB, Crone S, McLeod DS. Hetero-geneity in localization of isoforms of TGF-beta in human retina,vitreous, and choroids. Invest Ophthalmol Vis Sci. 1993;34:477–487.
55. Pasquale LR, Dorman-Pease ME, Lutty GA, Quigley HA, Jampel HD.Immunolocalization of TGF-beta 1, TGF-beta 2, and TGF-beta 3 inthe anterior segment of the human eye. Invest Ophthalmol Vis Sci.1993;34:23–30.
56. Tripathi RC, Chan WF, Li J, Tripathi BJ. Trabecular cells expressthe TGF-beta 2 gene and secrete the cytokine. Exp Eye Res.1994;58:523–528.
57. Tripathi RC, Li J, Chan WF, Tripathi BJ. Aqueous humor in glau-comatous eyes contains an increased level of TGF-beta 2. Exp EyeRes. 1994;59:723–727.
58. Jampel HD, Roche N, Stark WJ, Roberts AB. Transforming growthfactor-beta in human aqueous humor. Curr Eye Res. 1990;9:963–969.
59. Gupta A, Monroy D, Ji Z, Yoshino K, Huang A, Pflugfelder SC.Transforming growth factor beta-1 and beta-2 in human tear fluid.Curr Eye Res. 1996;15:605–614.
60. Mead AL, Wong TT, Cordeiro MF, Anderson IK, Khaw PT. Evalu-ation of anti-TGF-�2 antibody as a new postoperative anti-scarringagent in glaucoma surgery. Invest Ophthalmol Vis Sci. 2003;44:3394–3401.
61. The Fluorouracil Filtering Surgery Study Group. Fluorouracil Fil-tering Surgery Study one-year follow-up. Am J Ophthalmol. 1989;108:625–635.
62. The Fluorouracil Filtering Surgery Study Group. Five-year fol-low-up of the Fluorouracil Filtering Surgery Study. Am J Ophthal-mol. 1996;121:349–366.
63. Goldenfeld M, Krupin T, Ruderman JM, et al. 5-Fluorouracil ininitial trabeculectomy: a prospective, randomized, multicenterstudy. Ophthalmology. 1994;101:1024–1029.
64. Rothmann RF, Liebermann JM, Ritch R. Low-dose 5-fluouracil tra-beculectomy as initial surgery in uncomplicated glaucoma: long-term follow-up. Ophthalmology. 2000;107:1184–1190.
3844 Priglinger et al. IOVS, September 2006, Vol. 47, No. 9
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018

65. Lamping KA, Belkin JK. 5-Fluouracil and mitomycin C in pseu-dophakic patients. Ophthalmology. 1995;102:70–75.
66. Katz GJ, Higginbotham EJ, Lichter PR, et al. Mitomycin C versus5-fluouracil in high-risk glaucoma filtration surgery: extended fol-low-up. Ophthalmology. 1995;102:1263–1269.
67. Doyle JW, Sherwood MB, Khaw PT, McGrory S, Smith MF. Intra-operative 5-fluouracil for filtration surgery in the rabbit. InvestOphthalmol Vis Sci. 1993;34:3313–3319.
68. Khaw PT, Doyle JW, Sherwood MB, Smith MF, McGorray S. Effectsof intraoperative 5-fluouracil or mitomycin C on glaucoma filtra-tion surgery in the rabbit. Ophthalmology. 1993;100:367–372.
69. Shin DH, Juzych MS, Khatana AK, Swendris RP, Parrow KA. Nee-dling revision of failed filtering blebs with adjunctive 5-fluouracil.Ophthalmic Surg. 1993;24:242–248.
70. Allen LE, Manuchehri K, Corridan PG. The treatment of encapsu-lated trabeculectomy blebs in an out-patient setting using a nee-dling technique and subconjunctival 5-fluouracil injection. Eye.1998;12:119–123.
71. Jampel HD, Pasquale LR, Dibernardo C. Hypotony maculopathyfollowing trabeculectomy with mitomycin C. Arch Ophthalmol.1992;110:1049–1050.
72. Parrish R, Minckler D. “Late endophthalmitis”: filtering surgerytime bomb? Ophthalmology. 1996;103:1167–1168.
73. Stamper RL, McMenemy MG, Lieberman MF. Hypotonous macu-lopathy after trabeculectomy with subconjunctival 5-fluouracil.Am J Ophthalmol. 1992;114:544–553.
74. Wang JY, Johnson LR. Role of transglutaminase and protein cross-linking in the repair of mucosal stress erosions. Am J Physiol.1992;262:818–825.
75. Priglinger SG, Alge CS, Neubauer AS, et al. TGF-beta2-induced cellsurface tissue transglutaminase increases adhesion and migrationof RPE cells on fibronectin through the gelatin-binding domain.Invest Ophthalmol Vis Sci. 2004;45:955–963.
76. de Macedo P, Marrano C, Keillor JW. Synthesis of dipeptide-boundepoxides and alpha, beta-unsaturated amides as potential irreversibletransglutaminase inhibitors. Bioorg Med Chem. 2002;10:355–360.
IOVS, September 2006, Vol. 47, No. 9 Tissue Transglutaminase after Filtering Surgery 3845
Downloaded From: https://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933235/ on 07/17/2018