plant communities preferences of terrestrial crustaceans (isopoda: oniscidea) in a protected coastal...
TRANSCRIPT

Biologia 69/3: 354—362, 2014Section ZoologyDOI: 10.2478/s11756-013-0321-0
Plant communities preferences of terrestrial crustaceans (Isopoda:Oniscidea) in a protected coastal area of southeastern Sicily (Italy)
Giuseppina Messina, Giuseppe Montesanto, Elisa Pezzino, Saverio Sciandrello,Domenico Caruso & Bianca Maria Lombardo*
University of Catania, Department of Biological, Geological and Environmental Sciences, via Androne 81, I-95124 Catania,Italy; e-mail: [email protected]
Abstract: We studied the plant communities preferences of oniscidean isopods in a protected coastal area in southeasternSicily that is characterized by ponds, dunes and rocky coast. In total, five sites were sampled between March 2003 andFebruary 2004 using pitfall traps. A total of 7,326 specimens were found, belonging to 23 species. The vegetation analysis,using the phytosociological method, recognized a total of 11 types of plant communities, with specific ecological features.Measuring the species richness, diversity and annual activity density (aAD) values of the terrestrial isopod communitiesfrom each sampling site revealed differences among the various plant communities. PCA analysis confirmed the correlationbetween isopods species and specific plant communities. Moreover our results show that the high species richness is associatedwith a high vegetation cover and number of plants.
Key words: terrestrial isopods; pond; diversity; activity density; PCA
Introduction
Terrestrial isopods represent a very important compo-nent of the soil macrofauna. In temperate regions, theyare the dominant decomposing arthropods, and as such,are primarily responsible for regulating decompositionand recycling of nutrients in the ecosystems (Sutton1980; Zimmer 2002).The distribution of isopods species depends on sev-
eral environmental parameters, such as soil properties,air humidity and temperature (Hornung & Warburg1995). The isopods indeed are grouped into differentecological categories, based on the habitat that theycolonize (Vandel 1960; Schmalfuss 1984).Human activities (e.g., agriculture) contribute to
the change of natural ecosystems with consequent lossof habitat and decrease of biological richness (Paoletti1987, 1988; Paoletti & Hassal 1999; Fraja et al. 2010).Changed species equilibrium is observed in agriculturalregions where pesticides and fertilizers have been ap-plied, indicating a relationship between the manage-ment of the territory and the functioning of the ecosys-tem (Paoletti & Hassall 1999). Protected areas are well-preserved environments in which human activities aregreatly reduced or absent and generally represent a fun-damental reference point in terms of high diversity ofterrestrial isopods in the ecosystem.Literature provides many studies about isopods
diversity in the Mediterranean coastal wetland (e.g.,Achouri et al. 2008; Hamaıed-Melki et al. 2011; Ke-maissia et al. 2012; Messina et al. 2012); habitat pref-erence (Vilisics et al. 2007); but a few scientific studieson their plant communities preferences. Among these,David et al. (1999) evaluated the relationship betweendiplopods and isopods and the vegetation structure insouthern France; Sfenthourakis (1992); Sfenthourakiset al. (2005, 2008) studied the habitat preference innatural areas in Greek mountains; Souty-Grosset et al.(2008) reported the relationships among habitats, plantcomponents and communities of terrestrial isopods inthe Region Poitou-Charentes (France).A study has already been conducted on the di-
versity of isopods in the Natural Reserve of Vendi-cari (Messina et al. 2011) that is characterized by saltycoastal lakes (ponds). In this area 23 species were iden-tified, 3 of which were new to this area: Spelaeoniscussp., Bathytropa sp., Agabiformius lentus. The diversityvalues were higher in spring and autumn than in sum-mer and winter.The aim of this study was to verify the relation-
ships between isopods species and plant communitiesin order to obtain more information about the ecologyof each species. Indeed, knowledge of the relationshipswith the environment is a further important elementto establish the usefulness of species as bioindicators(Brandmayr et al. 2005).
* Corresponding author
c©2013 Institute of Zoology, Slovak Academy of Sciences

Isopoda in a coastal area of Sicily 355
Fig. 1. Map of the study area. Sampling sites and transect positions are indicated.
Material and methods
Study areaThe study area is part of the Natural Reserve of Vendi-cari (Fig. 1), which is registered as a Special ProtectionArea (SPA) and a Site of Community Importance (SIC,ITA090002). The reserve is formed by a coastal land be-tween the towns of Noto and Pachino (Syracuse province)in southeastern Sicily (WGS: 36◦48′13′′ N, 15◦05′54′′ E); itconsists of a plain with moderate height differences (up to12 m a.s.l.) that is characterized by sand dunes, rocky coast,and three ponds along the shore line. These ponds are veryclose to each other, and each one is approximately 100–130m from the coastline. From north to south, are called Pan-tano Piccolo (0.2 km2); Pantano Grande (0.38 km2), whichis sometimes continuous with Pantano Piccolo, dependingon water level; and Pantano Roveto (1.24 km2). They areall separated from the sea, although periods of particularlyheavy rain can increase the level of Pantano Roveto suchthat excess water pours into the sea. Each pond has an av-erage depth of 30 cm and a maximum depth of 1–2 m. Thesalinities are of a potentially wide range, depending on theseasonal rainfall; the salinity of Pantano Grande ranges from18 to 280‰, while that of Pantano Roveto ranges from 35to 290‰ (Moscatello & Belmonte 2009).
SamplingSampling was conducted by pitfall trapping between March2003 and February 2004. The traps were constructed ofpolyethylene cylinders (height 20 cm, diameter 7.5 cm) andthey were three-quarters full of a water/sodium chloride sat-urated solution. Pitfall trapping is generally considered agood sampling method because it is simple and easy to ap-ply (Greenslade 1964), however, low-mobility species and/orjuveniles may be under-sampled (Brandmayr et al. 2005).
Five sampling sites were selected, with transects of pit-fall traps traced at each site. The transects were drawn allorthogonally to the sea and where there was an ecologi-cal succession of plant communities. Each trap was usuallyplaced at the same distance to the proximal trap. Differ-ent numbers of traps were placed in correspondence to eachplant community in proportion to its wideness. The sam-pling sites, the type of substrate, the plant communitiesand the number of traps located at each site are shown in
Table 1. Six traps were placed at site 1 (S1), along a 110 mtransect that was traced between the coastline and Pan-tano Roveto. Six traps were placed at site 2 (S2), along a120 m transect that was traced inland from the PantanoRoveto. Seven traps were placed at site 3 (S3), along a 100m transect traced between the coastline and Pantano Pic-colo. Seven traps were placed at site 4 (S4), along a 100 mtransect traced inland from Pantano Piccolo. Three trapswere placed at site 5 (S5), along a 40 m transect tracedbetween the coastline and Pantano Grande.
The numbers assigned to the traps, the distances be-tween the traps, and the orientation of each transect areindicated in Messina et al. (2011). The traps were emptiedmonthly, and the collected material was transferred to 70%alcohol solutions. Species identifications were performed inthe laboratory.
Plant communities of the study areaTo analyze and classify various plant communities, differentphytosociological surveys were performed using the Braun-Blanquet method (Braun-Blanquet 1964). According thismethod in each releve the complete list of vascular plantspecies was recorded and for each species the cover value(percentage of soil surface) was assessed (cover scale +:<1%cover; 1: 1–5% cover; 2: 5–25% cover; 3: 25–50% cover; 4: 50–75% cover; 5: >75% cover). The syntaxonomical framing ofthe vegetation units followed the methods of Rivas-Martinezet al. (2001) and Brullo et al. (2002).
Data analysisThe ecological indices used to assess the isopods diversityin each plant community were: Shannon-Wiener diversityindex (H’), Pielou evenness index (J’), Berger-Parker domi-nance index (B = Nmax / N, where Nmax is the number ofindividuals of the most abundant species) (Magurran 1988).Similarities among plant communities were calculated us-ing Jaccard’s index. Statistical analysis was performed usingPAST version 2.15.
Annual activity density (aAD) (Brandmayr & BrunelloZanitti 1982) values were calculated for each species col-lected in each plant community.
Moreover, in detail, the obtained aAD values were di-vided into five classes (<0.1, 0.1–0.5, 0.5–2, 2–10, and >10).

356 G. Messina et al.
Table 1. Type of substrate, plant communities and their labels and the number of traps at the sampling sites.
Sampling site Substrate Plant communities Label N◦ traps
S1 Sandy Coridothymo-Helichrysetum stoechadis CHs 2S1 Silty-clayey Inulo-Juncetum maritimi IJm 2S1 Silty-clayey Arthrocnemo-Juncetum subulati AJs 5S2 Rocky Myrto communis-Pistacietum lentisci McPl 4S2 Pebbly Hyparrhenietum hirto-pubescentis Hhp 4S3 Rocky Limonietum syracusani Ls 1S3 Rocky Chamaeropo-Sarcopoterietum spinosi CSs 4S4 Silty-clayey Phragmitetum communis Pc 3S4 Pebbly Bromus fasciculatus Bs 1S5 Sandy Ephedro-Juniperetum macrocarpae EJm 1S5 Silty-clayey Agropyro scirpei-Inuletum crithmoidis AsIc 2
Fig. 2. Annual density activity (aAD) for the species from sampling site S1. Labels of the plant communities as reported in Table 1.p – pond; s – sea. Generic names are given by abbreviations. Complete species names are reported in Table 2.
The classes indicate the variation of activities of the speciesfound in the plant communities along the transect.
Relationship among species, types of plant communityand vegetation cover were examined by Principal Compo-nent Analysis (PCA), using PC-ORD, version 4.34. In par-ticular, the PCA takes into account different quantitativedata such as altitude (Altit.), slope, number of plant species,vegetation cover and isopods abundance.
Results and discussions
Plant communities of the study areaA total of 11 types of plant communities with specificecological features were recognized (Table 1), growingmainly on four types of substrate: rocky, sandy, silty-clayey, pebbly.On the rocky substrate, the following plant com-
munities were observed: Limonietum syracusani Bar-tolo, Brullo & Marceno, 1982 (Ls), Myrto communis-Pistacietum lentisci Rivas-Martinez, 1975 (McPl), andChamaeropo-Sarcopoterietum spinosi Barbagallo, Brul-lo & Fagotto, 1979 (CSs). On the sandy substrate thefollowing plant communities were observed: Ephedro-
Juniperetum macrocarpae Bartolo, Brullo & Marceno,1982 (EJm), and Coridothymo-Helichrysetum stoecha-dis Barbagallo, 1983 (CHs). Surveys of the silty-clayey substrate evidenced the presence of the plantcommunities: Arthrocnemo-Juncetum subulati Brullo& Furnari, 1976 (AJs), Agropyro scirpei-Inuletumcrithmoidis Brullo, 1988 (AsIc), Inulo-Juncetum mar-itimi Brullo, 1988 (IJm), and Phragmitetum communisSchmale, 1939 (Pc). Surveys of the pebbly substraterevealed the plant communities: Hyparrhenietum hirto-pubescentis A. O. Bolos & Br.-Bl., 1950 (Hhp), andBromus fasciculatus C. Presl community (Bs).Figures 2–6 show ecological successions of plant
communities along each transect.
Principal Component Analysis (PCA) and SimilarityAnalysisThe results obtained by PCA analyses are shown inFigure 7. Eigenvalues: axis 1: 3.734; axis 2: 2.734; 44.5%cumulative variance accounted for the first two axes.The axis 1 shows a principal gradient of plants speciesrichness and slope; axis 2 shows a gradient of altitude,isopods abundance and percentage of vegetation cover.

Isopoda in a coastal area of Sicily 357
Fig. 3. Annual density activity (aAD) for the species from sampling site S2. Labels of the plant communities as reported in Table 1.p – pond. Generic names are given by abbreviations. Complete species names are reported in Table 2.
Fig. 4. Annual density activities (aAD) for the species from sampling site S3. Labels of the plant communities as reported in Table 1.p – pond; s – sea. Generic names are given by abbreviations. Complete species names are reported in Table 2.

358 G. Messina et al.
Fig. 5. Annual density activities (aAD) for the species from sampling site S4. Labels of the plant communities as reported in Table 1.p – pond. Generic names are given by abbreviations. Complete species names are reported in Table 2.
Fig. 6. Annual density activities (aAD) for the species from sampling site S5. Labels of the plant communities as reported in Table 1.p – pond; r – road; s – sea. Generic names are given by abbreviations. Complete species names are reported in Table 2.
PCA analysis confirmed the correlation betweenisopods species and plant communities (Fig. 7). Fur-thermore, the species richness is high where there is agreater presence of plants and high percentage of veg-etation cover. Same species are obviously associatedwith a specific plant community. Porcellionides prui-nosus, Agabiformius obtusus, Leptotrichus naupliensisand Armadillidium nasatum are associated with McPl,Halophiloscia couchii and Halophiloscia hirsuta withAsIc, Porcellionides sexfasciatus with EJm.Figure 8 shows the results of similarities analysis;
two clusters were distinguished. The faunal component
was more similar between CHs and IJm (J = 0.75).The similarity values are high among contiguous plantcommunities. A high value of similarity was also foundbetween Pc and AJs (J = 0.71); they are both halophilicplant communities. Low similarity values were foundbetween Ls and other plant communities. These resultsshow that the studied plant communities are separatedin two clusters: the plant communities growing on rockysubstrate and those growing on sandy-clay substrate.Consequently isopods are distributed differently in thetwo types of substrate. As matter of fact Leptotrichuspanzerii, species psammophila, is dominant in CHs, IJm

Isopoda in a coastal area of Sicily 359
Fig. 7. Principal Component Analysis (PCA) of isopod species and plant communities. Labels of the plant communities as reportedin Table 1. S – isopods species; labels of names as reported in Table 2.
Fig. 8. Clustering of plant communities based on number ofspecies of isopods. Labels of the plant communities as reportedin Table 1.
and AsIc growing on sandy, silty-clayey substrate.Furthermore the same species gradient was ob-
served between the sea and the ponds and between theponds and the inland.
Abundance of isopods in relation to plant communitiesSampling revealed the presence of 7,326 specimens ofisopods belonging to 23 species (Table 2); 63% of thespecimens were collected on the silty-clayey substrate,while only 3% were found on sandy substrate.
Among the 11 plant communities recorded, themost isopod-rich plants (containing 36% of collectedspecimens) were the halophilous low shrubs of Arthro-cnemo-Juncetum subulati, a type of vegetation thatgrows on periodically submerged. Isopods were alsoabundantly collected from the halophilous vegeta-tion of Agropyro scirpei-Inuletum crithmoidis (18% ofspecimens) and from the evergreen shrub of Myrtocommunis-Pistacietum lentisci (15% of specimens).Lower abundances of isopods (percentages of speci-mens) were found in the sub-halophilous vegetation ofInulo-Juncetum maritimi (3%), which prefers sand soilsthat are submitted to long periods of submersion; inthe psammophilous shrub vegetation of Coridothymo-Helichrysetum stoechadis (2%); in the coastal maquis ofEphedro-Juniperetum macrocarpae (1%); in the rockycoastal vegetation of Limonietum syracusani (<1%);and finally in the uncultivated Bromus fasciculatus(<1%).
Species richness and diversity in relation to the plantcommunitiesTable 2 shows the aAD values for the 23 isopod speciesfound in the 11 plant communities. The variation ofaAD each species along transects is shown in Figs 5–9. The aAD values ranged from 0.03 to 18.67. Somespecies are dominant in one plant community. Shan-non index values vary from 0.74 to 1.57 (Table 2). Highvalues diversity were shown in McPl (H = 1.56) andin EJm (H = 1.57). This latter was the most balancedcommunity (H’ = 0.81; B = 0.34). The lowest values di-versity (H = 0.74) and the highest value of dominance
360 G. Messina et al.
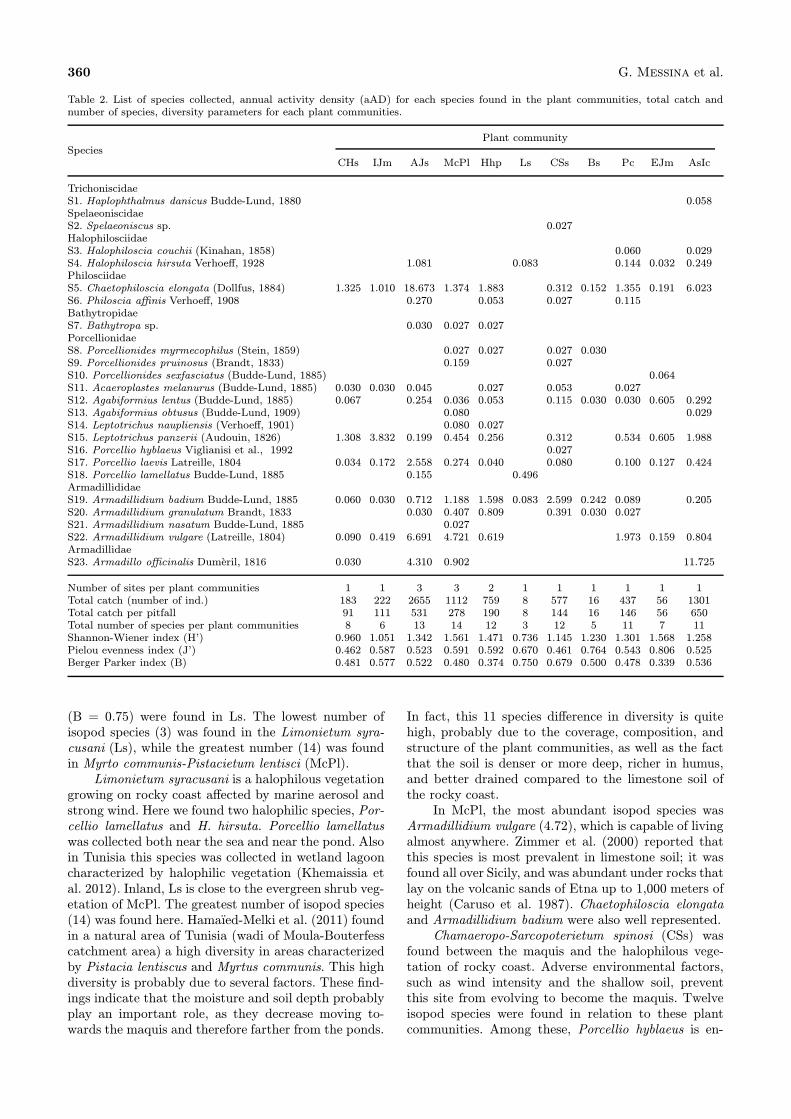
Table 2. List of species collected, annual activity density (aAD) for each species found in the plant communities, total catch andnumber of species, diversity parameters for each plant communities.
Plant communitySpecies
CHs IJm AJs McPl Hhp Ls CSs Bs Pc EJm AsIc
TrichoniscidaeS1. Haplophthalmus danicus Budde-Lund, 1880 0.058SpelaeoniscidaeS2. Spelaeoniscus sp. 0.027HalophilosciidaeS3. Halophiloscia couchii (Kinahan, 1858) 0.060 0.029S4. Halophiloscia hirsuta Verhoeff, 1928 1.081 0.083 0.144 0.032 0.249PhilosciidaeS5. Chaetophiloscia elongata (Dollfus, 1884) 1.325 1.010 18.673 1.374 1.883 0.312 0.152 1.355 0.191 6.023S6. Philoscia affinis Verhoeff, 1908 0.270 0.053 0.027 0.115BathytropidaeS7. Bathytropa sp. 0.030 0.027 0.027PorcellionidaeS8. Porcellionides myrmecophilus (Stein, 1859) 0.027 0.027 0.027 0.030S9. Porcellionides pruinosus (Brandt, 1833) 0.159 0.027S10. Porcellionides sexfasciatus (Budde-Lund, 1885) 0.064S11. Acaeroplastes melanurus (Budde-Lund, 1885) 0.030 0.030 0.045 0.027 0.053 0.027S12. Agabiformius lentus (Budde-Lund, 1885) 0.067 0.254 0.036 0.053 0.115 0.030 0.030 0.605 0.292S13. Agabiformius obtusus (Budde-Lund, 1909) 0.080 0.029S14. Leptotrichus naupliensis (Verhoeff, 1901) 0.080 0.027S15. Leptotrichus panzerii (Audouin, 1826) 1.308 3.832 0.199 0.454 0.256 0.312 0.534 0.605 1.988S16. Porcellio hyblaeus Viglianisi et al., 1992 0.027S17. Porcellio laevis Latreille, 1804 0.034 0.172 2.558 0.274 0.040 0.080 0.100 0.127 0.424S18. Porcellio lamellatus Budde-Lund, 1885 0.155 0.496ArmadillididaeS19. Armadillidium badium Budde-Lund, 1885 0.060 0.030 0.712 1.188 1.598 0.083 2.599 0.242 0.089 0.205S20. Armadillidium granulatum Brandt, 1833 0.030 0.407 0.809 0.391 0.030 0.027S21. Armadillidium nasatum Budde-Lund, 1885 0.027S22. Armadillidium vulgare (Latreille, 1804) 0.090 0.419 6.691 4.721 0.619 1.973 0.159 0.804ArmadillidaeS23. Armadillo officinalis Dumeril, 1816 0.030 4.310 0.902 11.725
Number of sites per plant communities 1 1 3 3 2 1 1 1 1 1 1Total catch (number of ind.) 183 222 2655 1112 759 8 577 16 437 56 1301Total catch per pitfall 91 111 531 278 190 8 144 16 146 56 650Total number of species per plant communities 8 6 13 14 12 3 12 5 11 7 11Shannon-Wiener index (H’) 0.960 1.051 1.342 1.561 1.471 0.736 1.145 1.230 1.301 1.568 1.258Pielou evenness index (J’) 0.462 0.587 0.523 0.591 0.592 0.670 0.461 0.764 0.543 0.806 0.525Berger Parker index (B) 0.481 0.577 0.522 0.480 0.374 0.750 0.679 0.500 0.478 0.339 0.536
(B = 0.75) were found in Ls. The lowest number ofisopod species (3) was found in the Limonietum syra-cusani (Ls), while the greatest number (14) was foundin Myrto communis-Pistacietum lentisci (McPl).Limonietum syracusani is a halophilous vegetation
growing on rocky coast affected by marine aerosol andstrong wind. Here we found two halophilic species, Por-cellio lamellatus and H. hirsuta. Porcellio lamellatuswas collected both near the sea and near the pond. Alsoin Tunisia this species was collected in wetland lagooncharacterized by halophilic vegetation (Khemaissia etal. 2012). Inland, Ls is close to the evergreen shrub veg-etation of McPl. The greatest number of isopod species(14) was found here. Hamaıed-Melki et al. (2011) foundin a natural area of Tunisia (wadi of Moula-Bouterfesscatchment area) a high diversity in areas characterizedby Pistacia lentiscus and Myrtus communis. This highdiversity is probably due to several factors. These find-ings indicate that the moisture and soil depth probablyplay an important role, as they decrease moving to-wards the maquis and therefore farther from the ponds.
In fact, this 11 species difference in diversity is quitehigh, probably due to the coverage, composition, andstructure of the plant communities, as well as the factthat the soil is denser or more deep, richer in humus,and better drained compared to the limestone soil ofthe rocky coast.In McPl, the most abundant isopod species was
Armadillidium vulgare (4.72), which is capable of livingalmost anywhere. Zimmer et al. (2000) reported thatthis species is most prevalent in limestone soil; it wasfound all over Sicily, and was abundant under rocks thatlay on the volcanic sands of Etna up to 1,000 meters ofheight (Caruso et al. 1987). Chaetophiloscia elongataand Armadillidium badium were also well represented.Chamaeropo-Sarcopoterietum spinosi (CSs) was
found between the maquis and the halophilous vege-tation of rocky coast. Adverse environmental factors,such as wind intensity and the shallow soil, preventthis site from evolving to become the maquis. Twelveisopod species were found in relation to these plantcommunities. Among these, Porcellio hyblaeus is en-

Isopoda in a coastal area of Sicily 361
demic to eastern Sicily (Viglianisi et al. 1992). Ar-madillidium badium was the most common (2.60). InCSs we found Spelaeoniscus sp. which is very sensi-tive to dryness and is very rarely captured by pit-fall trap because of its low mobility and scarce su-perficial activity. High species richness was found inArthrocnemo-Juncetum subulati (AJs), Phragmitetumcommunis (Pc), and Agropyro scirpei-Inuletum crith-moidis (AsIc), which grows on salt clay soils humidfor long time (Tomaselli et al. 2011; Sciandrello &Tomaselli 2014). Arthrocnemo-Juncetum subulati (AJs)was the richest with 13 species, two of which, H. hir-suta and P. lamellatus, are purely halophilous species(see also Montesanto et al. 2007) and one, C. elongata,is strongly hydrophilous and also has the highest aADvalue (18.67). This species is typical of areas at highhumidity (Caruso et al. 1987). In the coastal Sicilianwetland and in the wetland of Tunisia it was collectedin areas characterized by halophilic vegetation (Khe-maissia et al. 2012).High species richness was also present on the peb-
bly substrate, which are destroyed by fire every year.This community is characterized by dominance of Hy-parrhenia hirta and several plant of Lygeo-Stipetea class(Hyparrhenietum hirto-pubescentis, Hhp). Here, 12 iso-pod species were found, with C. elongata (1.88) and A.badium (1.60) being the most common.The isopod species Haplophthalmus danicus, Spe-
laeoniscus sp., P. sexfasciatus, Porcellio hyblaeus, andA. nasatum were present with very low aAD values inonly one community each: AsIc, EJm, CSs, and McPl,respectively.Porcellionides sexfasciatus is able to tolerate a
certain amount of salinity and is normally consideredcoastal species, but not halophilic; it was found nearthe beach in sandy substrate (EJm).Finally, our results show that within the study
area, generally the species show preference for a plantcommunity. Furthermore, a high isopod richness is as-sociated with a high vegetation cover, a high floristicrichness and the type of soil. Indeed, on sandy soils,that are dry and fast draining, we have found a fewernumber of isopod species while on clay soils, that aremoist also in the summer, we have found a greater num-ber of isopod species.
Acknowledgements
The authors would like to thank the “Azienda RegionaleForeste Demaniali di Siracusa” for permission to work inthe study area.
References
Achouri M.S., Hamaied S. & Charfi-Cheikhrouha F. 2008. Thediversity of terrestrial isopoda in the Berkoukech area,Kroumire, Tunisia. Crustaceana 81 (8): 917–929. DOI:10.1163/156854008X354948
Brandmayr P. & Brunello Zanitti C. 1982. Le comunita a Coleot-teri Carabidi di alcuni Querco-Carpineti della bassa pianura
del Friuli. Quaderni sulla “Struttura delle Zoocenosi Ter-restri” 4. I boschi della Pianura Padano-Veneta, pp. 69–124.
Brandmayr P., Zetto T. & Pizzolotto R. 2005. I Coleotteri Cara-bidi per la valutazione ambientale e la conservazione della bio-diversita. APAT, Manuale operativo 34/2005. I.G.E.R. Roma,240 pp. ISBN: 88-448-0152-3
Braun-Blanquet J. 1964. Pflanzensoziologie. Grundzuge derVegetations-kunde. Zweite, umgearbeitete und vermehrteAuflage. Springer Verlag, Wien, 865 pp.
Brullo S., Giusso Del Galdo G., Minissale P., Siracusa G. &Spampinato G. 2002. Considerazioni sintassonomiche e fito-geografiche sulla vegetazione della Sicilia. Boll. Accad. Gioe-nia Sci. Nat. Catania 35 (361): 325–359.
Caruso D., Baglieri C., Di Maio M. & Lombardo B.M. 1987.Isopodi terrestri di Sicilia ed isole circumsiciliane (Crustacea,Isopoda, Oniscoidea). Animalia 14 (Suppl.): 5–211.
David J.F., Devernay S., Loucougaray G. & Le Floch E. 1999.Belowground biodiversity in a Mediterranean landscape: re-lationships between saprophagous macroarthropod commu-nities and vegetation structure. Biodivers. Conserv. 8 (6):753–767. DOI: 10.1023/A:1008842313832
Fraj M., Charfi-Cheikrouha F. & Souty-Grosset C. 2010. Terres-trial isopods diversity related to irrigation and agriculturalpractices in North-East of Tunisia. Anadolu J. Agric. Sci. 25(S-3): 217–223.
Greenslade P.J.M. 1964. Pitfall trapping as a method for study-ing populations of Carabidae (Coleoptera). J. Anim. Ecol. 33(2): 301–310.
Hamaıed-Melki S., Achouri M.S., Aroui O.E., Bohli D. & Charfi-Cheikhrouha F. 2011. Terrestrial isopod diversity in the wadiMoula-Bouterfess catchment area (Kroumire, north-west ofTunisia). Afr. J. Ecol. 49 (1): 31–39. DOI: 10.1111/j.1365-2028.2010.01227.x
Hornung E. & Warburg M.R. 1995. Seasonal changes in the dis-tribution and abundance of isopod species in different habi-tats within the Mediterranean region of northern Israel. ActaOecol. 16 (4): 431–445
Kemaissia H., Souty-Grosset C. & Nasri-Ammar K. 2011. Bio-diversity of terrestrial isopods in coastal wetlands (Tunisia),pp. 99–100. In: Zidar P. & Štrus J. (eds), Proceedings of theEighth International Symposium on Terrestrial Isopod Biol-ogy, Bled (Slovenia), 165 pp. ISBN: 978-961-6822-10-7
Khemaissia H., Touihri M., Jelassi R., Souty-Grosset C. & Nasri-Ammar K. 2012. A preliminary study of terrestrial isopoddiversity in coastal wetlands of Tunisia. Vie et milieu – Lifeand Environment 62 (4): 203–211.
Magurran A.E. 1988. Ecological diversity and its measurement.New Jersey: Princeton University Press, 192 pp. ISBN:9780691084916
Messina G., Montesanto G., Pezzino E., Caruso D. & Lom-bardo B.M. 2011. Diversity of terrestrial isopods in a pro-tected area characterized by salty coastal ponds (Vendi-cari, Sicily). J. Nat. Hist. 45 (35–36): 2145–2158. DOI:10.1080/00222933.2011.587899
Messina G., Pezzino E., Montesanto G., Caruso D. & LombardoB.M. 2012. The diversity of terrestrial isopods in the natu-ral reserve “Saline di Trapani e Paceco” (Crustacea, Isopoda,Oniscidea) in northwestern Sicily. In: Štrus J., Taiti S. & Sfen-thourakis S. (eds), Advances in Terrestrial Isopod Biology.ZooKeys 176: 215–230. DOI: 10.3897/zookeys.176.2367
Montesanto G., Caruso D. & Lombardo B.M. 2007. Taxo-nomic status of the Mediterranean terrestrial isopod Porcelliolamellatus Budde-Lund as inferred from genetic and morpho-logical differentiation (Crustacea, Isopoda, Oniscidea). Crus-taceana 80 (8): 917–938. DOI: 10.1163/156854007781681229
Moscatello S. & Belmonte G. 2009. Egg banks in hypersaline lakesof the South-East Europe. Saline Syst. 5 (1): article:3, 7 pp.DOI: 10.1186/1746-1448-5-3
Paoletti M.G. 1987. Terrestrial isopods in the agroecosystems ofthe low-lying plain in North-eastern Italy, pp. 423–426. In:Striganova B.R. (ed.), Soil Fauna and Soil Fertility, Proc. 9th
Int. Coll. Soil Zoology, Moscow Nauka.Paoletti M.G. 1988. Soil invertebrates in cultivated and unculti-vated soils in north-east Italy. Redia 71: 501–563.

362 G. Messina et al.
Paoletti G.P. & Hassall M. 1999. Woodlice (Isopoda: Oniscidea):their potential for assessing sustainability and use as bioindi-cators. Agr. Ecosyst. Environ. 74 (1-3): 157–165. DOI:10.1016/S0167-8809(99)00035-3
Rivas Martínez S., Fernández González F., Loidi J., Lousa M. &Penas A. 2001. Syntaxonomical Chek-List of vascular plantcommunities of Spain and Portugal to association level. Itin.Geobot. 14: 5–341.
Schmalfuss H. 1984. Eco-morphological strategies in terrestrialisopods. pp. 49–63 In: Sutton S.L. & Holdich D.M. (eds),The Biology of Terrestrial Isopods, Proceedings of a sympo-sium held at the Zoological Society of London on 7th and 8th
of July 1983, Symp. Zool. Soc. Lond. 53, Clarendon Press,Oxford, 518 pp.
Sciandrello S. & Tomaselli V. 2014. Coastal salt marshes plantcommunities of the Sarcocornietea fruticosae class in Apulia(Italy). Biologia 69 (1): 53–69.
Sfenthourakis S. 1992. Altitudinal effect on species richnessof Oniscidea (Crustacea; Isopoda) on three mountains inGreece. Global Ecology and Biogeography Letters 2 (5): 157–164.
Sfenthourakis S., Anastasiou I. & Strudenschi T. 2005. Altitudi-nal terrestrial Isopod diversity. Eur. J. Soil Biol. 41 (3-4):91–98. DOI: 10.1016/j.ejsobi.2005.09.006
Sfenthourakis S., Orfanou V. & Anastastiou Y. 2008. A compar-ative study of isopod assemblages of elevated habitats on fivemountains of Peloponnisos peninsula (Greece), pp. 13–19. In:Zimmer M., Charfi-Cheikhrouha F. & Taiti S. (eds), Proceed-ings of the International Symposium on Terrestrial IsopodBiology, Aachen, Shaker, 175 pp. ISBN: 978-3-8322-6418-5
Souty-Grosset C., Corre S., Freyssinel G., Aubert V., DonqueV. & Raimond R. 2008. Diversity of terrestrial isopods andhabitats in Poitou-Charentes (western France), pp. 21–37. In:Zimmer M., Charfi-Cheikhrouha F. & Taiti S. (eds), Proceed-ings of the International Symposium on Terrestrial IsopodBiology, Aachen, Shaker, 175 pp. ISBN: 978-3-8322-6418-5
Sutton S.L. 1980. Woodlice. Pergamon Press, Oxford, 144 pp.Tomaselli V., Di Pietro R. & Sciandrello S. 2011. Plant commu-nities structure and composition in three coastal wetlands insouthern Apulia (Italy). Biologia 66 (6): 1027–1043. DOI:10.2478/s11756-011-0113-3
Vandel A. 1960. Faune de France, Vol. 64, Isopodes terrestres(premiere partie). Lechevalier, Paris, 416 pp.
Viglianisi F., Lombardo B.M. & Caruso D. 1992. Differenzia-mento genetico e morfologico di alcune specie siciliane diIsopodi terrestri del genere Porcellio e descrizione di tre nuovespecie (Crustacea, Isopoda, Oniscoidea). Animalia 19 (1/3):235–273.
Vilisics F., Sólymos P., Hornung E. 2007. A preliminary study onhabitat features and associated terrestrial isopod species, pp195–199. In: Tajovsky K., Schlaghamersky J. & Pižl V. (eds),Contributions to soil zoology in Central Europe II: Proceed-ings of the 8th Central European Workshop on Soil Zoology,Ceske Budejovice, 217 pp. ISBN: 978-80-86525-08-2
Zimmer M. 2002. Nutrition in terrestrial isopods (Isopoda: Onis-cidea): an evolutionary-ecological approach. Biol. Rev. 77:455–493. DOI: 10.1017/S1464793102005912
Zimmer M., Brauckmann H., Broll G. & Topp W. 2000.Correspondence analytical evaluation of factors that influ-ence soil macro-arthropod distribution in abandoned grass-land. Pedobiologia 44 (6): 695–704. DOI: 10.1078/S0031-4056(04)70082-7
Received June 25, 2013Accepted November 25, 2013