pigmentary disorders section 10 melanocyte biology 65
TRANSCRIPT

1075
In the inner ear, particularly in the stria vascularis, melanocytes are thought to play a role in the development of hearing. Aberrant migration or survival of melanocytes within the inner ear, the iris, and midportions of the forehead and extremities explains the presence of congenital deafness, heterochromia irides, and patches of leukoderma, respectively, in patients with Waardenburg syndrome, the classic neurocristopathy. Also, aberrant migration or survival of enteric ganglion cells, another neural crestderived cell population, provides an explanation for the association of aganglionic megacolon (Hirschsprung disease) with Waardenburg syndrome or rarely piebaldism.
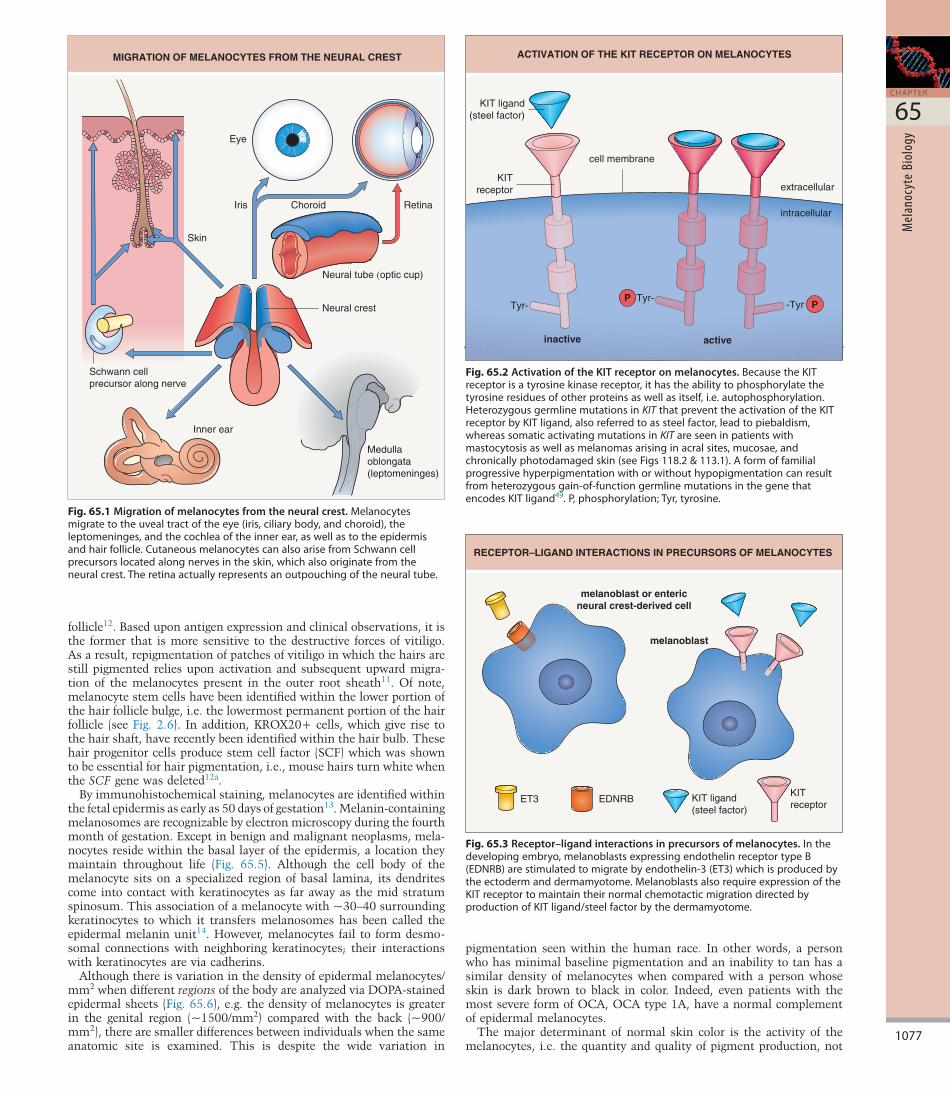
The survival and migration of neural crestderived cells during embryogenesis depends upon interactions between specific receptors on their cell surface and extracellular ligands. For example, KIT ligand (also known as steel factor or stem cell growth factor) binds to the transmembrane KIT receptor on melanocytes and melanocyte precursors (melanoblasts) (Figs 65.2 & 65.3); melanoblasts require expression of the KIT receptor in order to maintain their normal chemotactic migration directed by production of KIT ligand by the dermamyotome. Heterozygous germline mutations in KIT that decrease the ability of the KIT receptor to be activated by KIT ligand are responsible for human piebaldism, whereas in mice, mutations in either kit or steel can lead to white spotting. In the developing embryo, melanoblasts expressing endothelin receptor type B (EDNRB) are stimulated to migrate by endothelin3 (ET3 [EDN3]), which is produced by the ectoderm and dermamyotome. Mutations in one or both copies of EDN3 or EDNRB can result in Waardenburg syndrome plus aganglionic megacolon (see Fig. 65.3).
Transcription factors represent another group of proteins that play an essential role during embryogenesis. Because transcription factors can bind DNA and influence the activity of other genes, they are able to regulate the complex interplay of various sets of genes that is required for embryonic development. Several of the genes that, when mutated, give rise to Waardenburg syndrome encode transcription factors (e.g. PAX3, MITF, SOX10; see Table 66.4). Fig. 65.4 demonstrates some of these interactions (e.g. PAX3 and SOX10 can control expression of MITF)8,9. MITF is sometimes referred to as the master regulator of melanocyte development and function given its modulation of multiple differentiation genes and its early upregulation in neural crest cells that will eventually become melanocytes and emigrate from the dorsal neural tube.
During embryogenesis, melaninproducing melanocytes are found diffusely throughout the dermis. They first appear in the head and neck region at ~10 weeks of gestation. However, by the end of gestation, active dermal melanocytes have “disappeared”, except in three primary anatomic locations – the head and neck, the dorsal aspects of the distal extremities, and the presacral area10. Some of the dermal melanocytes have clearly migrated into the epidermis, but, given the absolute numbers of cells in the two compartments, apoptosis of pigment cells has also occurred. The three sites where active dermal melanocytes are still present at the time of birth coincide with the most common sites for dermal melanocytoses and dermal melanocytomas (blue nevi). Hepatocyte growth factor may play a role in the survival and proliferation of these dermal melanocytes as well as somatic activating mutations in GNA11 and GNAQ, which encode G proteins and are found in blue nevi (see Table 112.3).
As depicted in Fig. 65.1, melanocytes also migrate to the basal layer of the hair matrix and the outer root sheath of hair follicles. Cells that are actively producing melanin are easily recognized in the matrices of pigmented anagen hairs, whereas melanocytes within the outer root sheath are usually amelanotic and more difficult to identify11. It has been hypothesized that there are two populations of melanocytes in the skin, one in the interfollicular epidermis and the second in the hair
INTRODUCTIONIn order to understand the underlying pathophysiology of cutaneous disorders of hypopigmentation and hyperpigmentation, as well as the process of normal physiologic pigment production, an appreciation of the structure and function of the melanocyte is required. A classic example of basic pathogenesis is type 1 oculocutaneous albinism (OCA1), in which pigmentary dilution of the skin, hair, and eyes is due to a reduction or absence of tyrosinase activity secondary to mutations in both copies of the tyrosinase gene (TYR). Within the realm of physiologic pigmentation, melanocytes in individuals with red hair often express variants of the melanocortin 1 receptor (MC1R)1. As a consequence of the altered amino acid sequences of the variant MC1Rs, their cell surface expression and interactions with melanocyte stimulating hormone (MSH) can be affected, leading to an increase in the production of pheomelanin as opposed to eumelanin. Based upon population genetics, genes that are mutated in OCA (e.g. TYR, OCA2, TYRP1, SLC45A2, SLC24A5) also influence normal pigment variation (Table 65.1)2–6.
The major sections in this chapter are:
• the origin and function of the melanocyte
• the formation and function of the melanosome
• regulation of melanin biosynthesis.
ORIGIN AND FUNCTION OF THE MELANOCYTEThe melanocyte is a neural crestderived cell, and during embryogenesis precursor cells (melanoblasts) migrate along a dorsolateral then ventral pathway via the mesenchyme to reach the epidermis and hair follicles of the trunk (see Ch. 2). More recently, it was shown that cutaneous melanocytes can also arise from neural crestderived Schwann cell precursors that migrate along nerves to the skin via a distinct ventral pathway7. Additional sites of melanocyte migration include the uveal tract of the eye (choroid, ciliary body, and iris), the leptomeninges, and the inner ear (cochlea) (Fig. 65.1). Presumably, the death of melanocytes within the leptomeninges, inner ear, and skin is responsible for the aseptic meningitis, auditory symptoms, and areas of vitiligo, respectively, seen in patients with the Vogt–Koyanagi–Harada syndrome (see Ch. 66).
Key features■ Themajordeterminantofnormalskincoloristheactivityof
melanocytes,i.e.thequantityandqualityofpigmentproduction,notthedensityofmelanocytes
■ Melanocytescontainauniqueintracytoplasmicorganelle,themelanosome,whichisthesiteofmelaninsynthesisanddeposition
■ Comparedwithlightlypigmentedskin,darklypigmentedskinhasmelanosomesthatcontainmoremelaninandarelarger;oncetransferredtokeratinocytes,themelanosomesaresinglydispersedanddegradedmoreslowly
■ Tyrosinaseisthekeyenzymeinthemelaninbiosyntheticpathway■ Twomajorformsofmelaninareproducedinmelanocytes:
brown–blackeumelaninandyellow–redpheomelanin■ Theproductionofeumelaninversuspheomelaninisinfluencedby
thebindingofmelanocytestimulatinghormonetothemelanocortin1receptor
65MelanocyteBiologyJean L. Bolognia and Seth J. Orlow
SEC TION 10PIGMENTARY DISORDERS

non-print metadata
1075.e1
C H A P T E R
65
Mel
anoc
yte
Biol
ogy
ABSTRACTThe major determinant of normal skin color is the activity of melanocytes, i.e. the quantity and quality of pigment production, not the density of melanocytes. Melanocytes contain a unique intracytoplasmic organelle, the melanosome, which is the site of melanin synthesis and deposition. Compared with lightly pigmented skin, darkly pigmented skin has melanosomes that contain more melanin and are larger; once transferred to keratinocytes, the melanosomes are singly dispersed and degraded more slowly. Tyrosinase is the key enzyme in the melanin biosynthetic pathway and the two major forms of melanin produced in melanocytes are brown–black eumelanin and yellow–red pheomelanin. The production of eumelanin versus pheomelanin is influenced by the binding of melanocyte stimulating hormone to the melanocortin 1 receptor.
KEYWORDS:melanocyte, melanosome, tyrosinase, eumelanin, pheomelanin, melanocortin 1 receptor, MC1R, melanocyte stimulating hormone, MSH, pigmentation, agouti, melanin biosynthetic pathway
1076
S E C T I O N
10
PIGM
ENTA
RY D
ISOR
DERS
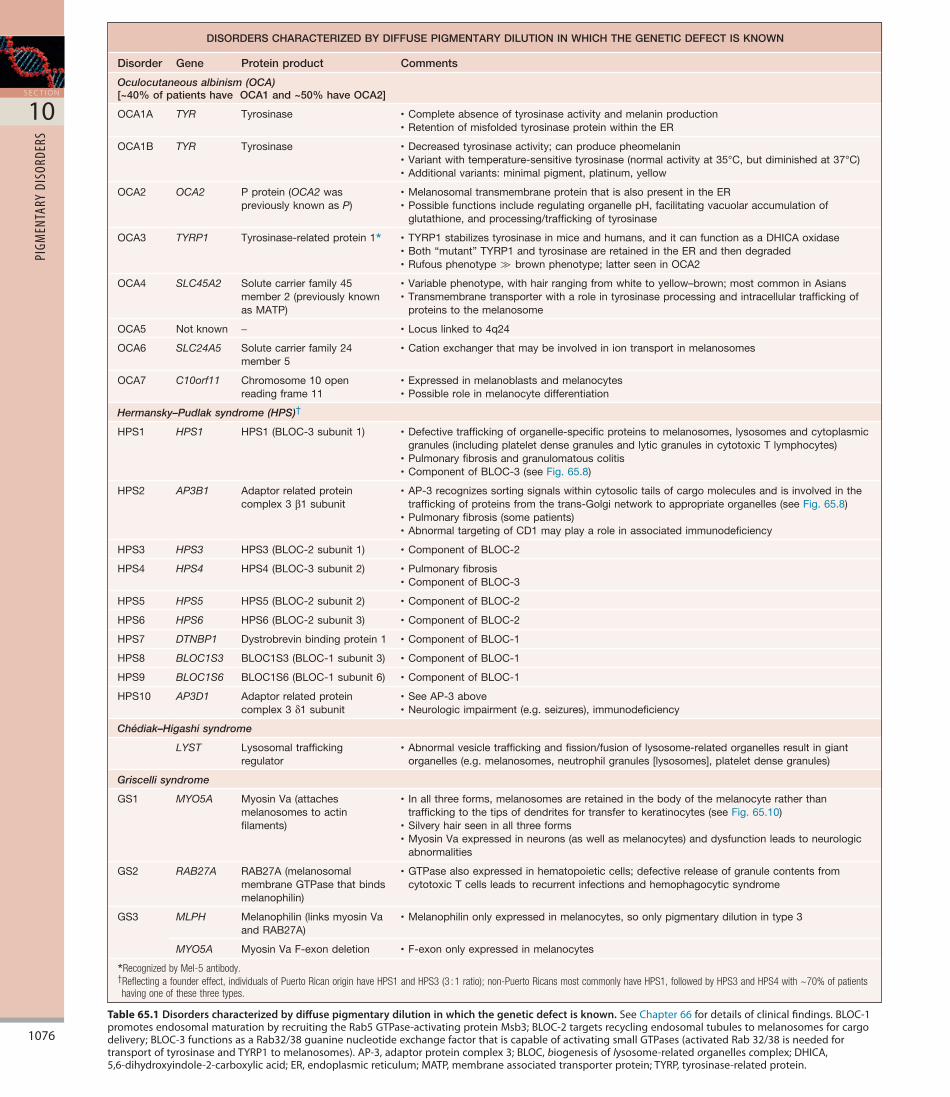
Table 65.1 Disorders characterized by diffuse pigmentary dilution in which the genetic defect is known.SeeChapter66fordetailsofclinicalfindings.BLOC-1promotesendosomalmaturationbyrecruitingtheRab5GTPase-activatingproteinMsb3;BLOC-2targetsrecyclingendosomaltubulestomelanosomesforcargodelivery;BLOC-3functionsasaRab32/38guaninenucleotideexchangefactorthatiscapableofactivatingsmallGTPases(activatedRab32/38isneededfortransportoftyrosinaseandTYRP1tomelanosomes).AP-3,adaptorproteincomplex3;BLOC,biogenesisoflysosome-relatedorganellescomplex;DHICA,5,6-dihydroxyindole-2-carboxylicacid;ER,endoplasmicreticulum;MATP,membraneassociatedtransporterprotein;TYRP,tyrosinase-relatedprotein.
DISORDERS CHARACTERIZED BY DIFFUSE PIGMENTARY DILUTION IN WHICH THE GENETIC DEFECT IS KNOWN
Disorder Gene Protein product Comments
Oculocutaneous albinism (OCA)[~40% of patients have OCA1 and ~50% have OCA2]
OCA1A TYR Tyrosinase •Completeabsenceoftyrosinaseactivityandmelaninproduction•RetentionofmisfoldedtyrosinaseproteinwithintheER
OCA1B TYR Tyrosinase •Decreasedtyrosinaseactivity;canproducepheomelanin•Variantwithtemperature-sensitivetyrosinase(normalactivityat35°C,butdiminishedat37°C)•Additionalvariants:minimalpigment,platinum,yellow
OCA2 OCA2 Pprotein(OCA2waspreviouslyknownasP)
•MelanosomaltransmembraneproteinthatisalsopresentintheER•PossiblefunctionsincluderegulatingorganellepH,facilitatingvacuolaraccumulationof
glutathione,andprocessing/traffickingoftyrosinase
OCA3 TYRP1 Tyrosinase-relatedprotein1* •TYRP1stabilizestyrosinaseinmiceandhumans,anditcanfunctionasaDHICAoxidase•Both“mutant”TYRP1andtyrosinaseareretainedintheERandthendegraded•Rufousphenotype≫brownphenotype;latterseeninOCA2
OCA4 SLC45A2 Solutecarrierfamily45member2(previouslyknownasMATP)
•Variablephenotype,withhairrangingfromwhitetoyellow–brown;mostcommoninAsians•Transmembranetransporterwitharoleintyrosinaseprocessingandintracellulartraffickingof
proteinstothemelanosome
OCA5 Notknown − •Locuslinkedto4q24
OCA6 SLC24A5 Solutecarrierfamily24member5
•Cationexchangerthatmaybeinvolvediniontransportinmelanosomes
OCA7 C10orf11 Chromosome10openreadingframe11
•Expressedinmelanoblastsandmelanocytes•Possibleroleinmelanocytedifferentiation
Hermansky–Pudlak syndrome (HPS)†
HPS1 HPS1 HPS1(BLOC-3subunit1) •Defectivetraffickingoforganelle-specificproteinstomelanosomes,lysosomesandcytoplasmicgranules(includingplateletdensegranulesandlyticgranulesincytotoxicTlymphocytes)
•Pulmonaryfibrosisandgranulomatouscolitis•ComponentofBLOC-3(seeFig.65.8)
HPS2 AP3B1 Adaptorrelatedproteincomplex3β1subunit
•AP-3recognizessortingsignalswithincytosolictailsofcargomoleculesandisinvolvedinthetraffickingofproteinsfromthetrans-Golginetworktoappropriateorganelles(seeFig.65.8)
•Pulmonaryfibrosis(somepatients)•AbnormaltargetingofCD1mayplayaroleinassociatedimmunodeficiency
HPS3 HPS3 HPS3(BLOC-2subunit1) •ComponentofBLOC-2
HPS4 HPS4 HPS4(BLOC-3subunit2) •Pulmonaryfibrosis•ComponentofBLOC-3
HPS5 HPS5 HPS5(BLOC-2subunit2) •ComponentofBLOC-2
HPS6 HPS6 HPS6(BLOC-2subunit3) •ComponentofBLOC-2
HPS7 DTNBP1 Dystrobrevinbindingprotein1 •ComponentofBLOC-1
HPS8 BLOC1S3 BLOC1S3(BLOC-1subunit3) •ComponentofBLOC-1
HPS9 BLOC1S6 BLOC1S6(BLOC-1subunit6) •ComponentofBLOC-1
HPS10 AP3D1 Adaptorrelatedproteincomplex3δ1subunit
•SeeAP-3above•Neurologicimpairment(e.g.seizures),immunodeficiency
Chédiak–Higashi syndrome
LYST Lysosomaltraffickingregulator
•Abnormalvesicletraffickingandfission/fusionoflysosome-relatedorganellesresultingiantorganelles(e.g.melanosomes,neutrophilgranules[lysosomes],plateletdensegranules)
Griscelli syndrome
GS1 MYO5A MyosinVa(attachesmelanosomestoactinfilaments)
• Inallthreeforms,melanosomesareretainedinthebodyofthemelanocyteratherthantraffickingtothetipsofdendritesfortransfertokeratinocytes(seeFig.65.10)
•Silveryhairseeninallthreeforms•MyosinVaexpressedinneurons(aswellasmelanocytes)anddysfunctionleadstoneurologic
abnormalities
GS2 RAB27A RAB27A(melanosomalmembraneGTPasethatbindsmelanophilin)
•GTPasealsoexpressedinhematopoieticcells;defectivereleaseofgranulecontentsfromcytotoxicTcellsleadstorecurrentinfectionsandhemophagocyticsyndrome
GS3 MLPH Melanophilin(linksmyosinVaandRAB27A)
•Melanophilinonlyexpressedinmelanocytes,soonlypigmentarydilutionintype3
MYO5A MyosinVaF-exondeletion •F-exononlyexpressedinmelanocytes
*Recognized by Mel-5 antibody.†Reflecting a founder effect, individuals of Puerto Rican origin have HPS1 and HPS3 (3 : 1 ratio); non-Puerto Ricans most commonly have HPS1, followed by HPS3 and HPS4 with ~70% of patients having one of these three types.
1077
C H A P T E R
65
Mel
anoc
yte
Biol
ogy
pigmentation seen within the human race. In other words, a person who has minimal baseline pigmentation and an inability to tan has a similar density of melanocytes when compared with a person whose skin is dark brown to black in color. Indeed, even patients with the most severe form of OCA, OCA type 1A, have a normal complement of epidermal melanocytes.
The major determinant of normal skin color is the activity of the melanocytes, i.e. the quantity and quality of pigment production, not
follicle12. Based upon antigen expression and clinical observations, it is the former that is more sensitive to the destructive forces of vitiligo. As a result, repigmentation of patches of vitiligo in which the hairs are still pigmented relies upon activation and subsequent upward migration of the melanocytes present in the outer root sheath11. Of note, melanocyte stem cells have been identified within the lower portion of the hair follicle bulge, i.e. the lowermost permanent portion of the hair follicle (see Fig. 2.6). In addition, KROX20+ cells, which give rise to the hair shaft, have recently been identified within the hair bulb. These hair progenitor cells produce stem cell factor (SCF) which was shown to be essential for hair pigmentation, i.e., mouse hairs turn white when the SCF gene was deleted12a.
By immunohistochemical staining, melanocytes are identified within the fetal epidermis as early as 50 days of gestation13. Melanincontaining melanosomes are recognizable by electron microscopy during the fourth month of gestation. Except in benign and malignant neoplasms, melanocytes reside within the basal layer of the epidermis, a location they maintain throughout life (Fig. 65.5). Although the cell body of the melanocyte sits on a specialized region of basal lamina, its dendrites come into contact with keratinocytes as far away as the mid stratum spinosum. This association of a melanocyte with ~30–40 surrounding keratinocytes to which it transfers melanosomes has been called the epidermal melanin unit14. However, melanocytes fail to form desmosomal connections with neighboring keratinocytes; their interactions with keratinocytes are via cadherins.
Although there is variation in the density of epidermal melanocytes/mm2 when different regions of the body are analyzed via DOPAstained epidermal sheets (Fig. 65.6), e.g. the density of melanocytes is greater in the genital region (~1500/mm2) compared with the back (~900/mm2), there are smaller differences between individuals when the same anatomic site is examined. This is despite the wide variation in
Fig. 65.1 Migration of melanocytes from the neural crest.Melanocytesmigratetotheuvealtractoftheeye(iris,ciliarybody,andchoroid),theleptomeninges,andthecochleaoftheinnerear,aswellastotheepidermisandhairfollicle.CutaneousmelanocytescanalsoarisefromSchwanncellprecursorslocatedalongnervesintheskin,whichalsooriginatefromtheneuralcrest.Theretinaactuallyrepresentsanoutpouchingoftheneuraltube.
MIGRATION OF MELANOCYTES FROM THE NEURAL CREST
Neural crest
Neural tube (optic cup)
Skin
Iris Choroid
Inner ear
Schwann cellprecursor along nerve
Eye
Medullaoblongata(leptomeninges)
Retina
Fig. 65.2 Activation of the KIT receptor on melanocytes.BecausetheKITreceptorisatyrosinekinasereceptor,ithastheabilitytophosphorylatethetyrosineresiduesofotherproteinsaswellasitself,i.e.autophosphorylation.HeterozygousgermlinemutationsinKITthatpreventtheactivationoftheKITreceptorbyKITligand,alsoreferredtoassteelfactor,leadtopiebaldism,whereassomaticactivatingmutationsinKITareseeninpatientswithmastocytosisaswellasmelanomasarisinginacralsites,mucosae,andchronicallyphotodamagedskin(seeFigs118.2&113.1).Aformoffamilialprogressivehyperpigmentationwithorwithouthypopigmentationcanresultfromheterozygousgain-of-functiongermlinemutationsinthegenethatencodesKITligand49.P,phosphorylation;Tyr,tyrosine.
KIT ligand(steel factor)
inactive active
Tyr-Tyr-
-Tyr
cell membrane
extracellular
intracellular
KITreceptor
ACTIVATION OF THE KIT RECEPTOR ON MELANOCYTES
PP
Fig. 65.3 Receptor–ligand interactions in precursors of melanocytes.Inthedevelopingembryo,melanoblastsexpressingendothelinreceptortypeB(EDNRB)arestimulatedtomigratebyendothelin-3(ET3)whichisproducedbytheectodermanddermamyotome.MelanoblastsalsorequireexpressionoftheKITreceptortomaintaintheirnormalchemotacticmigrationdirectedbyproductionofKITligand/steelfactorbythedermamyotome.
ET3
melanoblast or entericneural crest-derived cell
melanoblast
EDNRB KIT ligand(steel factor)
KITreceptor
RECEPTOR–LIGAND INTERACTIONS IN PRECURSORS OF MELANOCYTES

1078
S E C T I O N
10
PIGM
ENTA
RY D
ISOR
DERS
FORMATION AND FUNCTION OF THE MELANOSOMEWithin the cytoplasm of melanocytes is a unique organelle known as the melanosome, in which melanin pigments are synthesized, deposited, and transported. The melanosome is most closely related to the lysosome16. Through compartmentalization, both organelles provide protection for the remainder of the cell – lysosomes protect against
the density of melanocytes15. Several factors play a role in determining the level of melanocyte activity; they include specific characteristics of the individual melanosomes (e.g. their dimensions) as well as both baseline (constitutive) and stimulated (facultative) levels and activity of the enzymes involved in the melanin biosynthetic pathway. The latter are influenced by receptormediated interactions with extracellular ligands such as MSH which then influence the expression of transcription factors including MITF (see Fig. 65.4).
Fig. 65.4 Signal transduction pathways and transcription factors that contribute to melanocyte differentiation.MITFexpressionisactivatedearlyonduringthetransitionfrompluripotentneuralcrestcellstomelanoblastsandisrequiredformelanoblastsurvival8;mutationsinMITFleadtoWaardenburgsyndrome,aclassicneurocristopathy.MITFalsoregulatestheexpressionofmultiplepigmentgenesincludingthosethatencodetyrosinase,TYRP1,TYRP2,PMEL/PMEL17/gp100,andMART-1/Melan-A.AdditionaltranscriptionaltargetsareCDK2,CDKN2A,andBCL-2(whoseproteinproductisaninhibitorofapoptosis)9.SmallmoleculeinhibitorsofSIK(salt-induciblekinase)canupregulateMITF,andapplicationoftheseinhibitorstonormalhumanskinledtoanincreaseinpigmentation54.Inmice,Wntsignalinginmelanocytestemcellsiscriticalforhairpigmentation50.DetailsofhowactivationofG-protein-coupledreceptorsleadstoanincreaseinintracellularcAMPisshowninFig.65.15.Ofnote,EDNRBinteractswiththeGproteinsGNAQandGNA11,andactivatingmutationsinthegenesthatencodetheselattertwoproteinscanleadtoblueneviandphakomatosispigmentovascularis.cAMP,cyclicadenosinemonophosphate;CREB,cAMPresponse-elementbindingprotein;ET3,endothelin-3;EDNRB,endothelinreceptortypeB;LEF1,lymphoidenhancerbindingfactor1;MC1R,melanocortin1receptor;MITF,microphthalmia-associatedtranscriptionfactor;MSH,melanoctyestimulatinghormone;P,phosphorylation;PKA,proteinkinaseA;SOX10,SRY-boxcontaininggene10.
SIGNAL TRANSDUCTION PATHWAYS AND TRANSCRIPTION FACTORSTHAT CONTRIBUTE TO MELANOCYTE DIFFERENTIATION
α-MSH
β-Catenin
SOX10 LEF1 PAX3CREB
cAMP
PKA
Melanogenesis
Melanocyte differentiation
Melanocyte proliferation
Melanocyte survival
MITF promoter
RAS/RAF/MEK/ERK
MITF
MITF(activated)
Activation and Degradation
P
P
MC1R
KIT ligand
WNT
KIT receptor
ET3
EDNRB
Fig. 65.5 A melanocyte residing in the basal layer of the epidermis.Innormalskin,approximatelyeverytenthcellinthebasallayerisamelanocyte.Melanosomesaretransferredfromthedendritesofthemelanocyteintoneighboringkeratinocytesoftheepidermis,hairmatricesandmucousmembranes;notransferoccursinthepigmentepitheliumoftheretina.Theepidermalmelaninunitreferstotheassociationofamelanocytewith~30–40surroundingkeratinocytestowhichittransfersmelanosomes.
A MELANOCYTE RESIDING IN THE BASAL LAYER OF THE EPIDERMIS
phagocytosisof tip
stratumcorneum
stratumgranulosum
epidermis
dermis
stratumspinosum
basallayer
melanocyte
fusion ofmembranes
melanosomes
keratinocyte
dendrite

1079
C H A P T E R
65
Mel
anoc
yte
Biol
ogy
Fig. 65.7). In patients with OCA1 and OCA3, misfolded tyrosinase or aberrant tyrosinaserelated protein 1 (TYRP1) plus tyrosinase polypeptides, respectively, accumulate within the endoplasmic reticulum (ER)17. This leads to ER stress and activation of the unfolded protein response (UPR)18. As a result of the UPR, which acts as a form of quality control, these proteins become targets for destruction by proteasomes and therefore are not incorporated into melanosomes (see Fig. 65.7 inset).
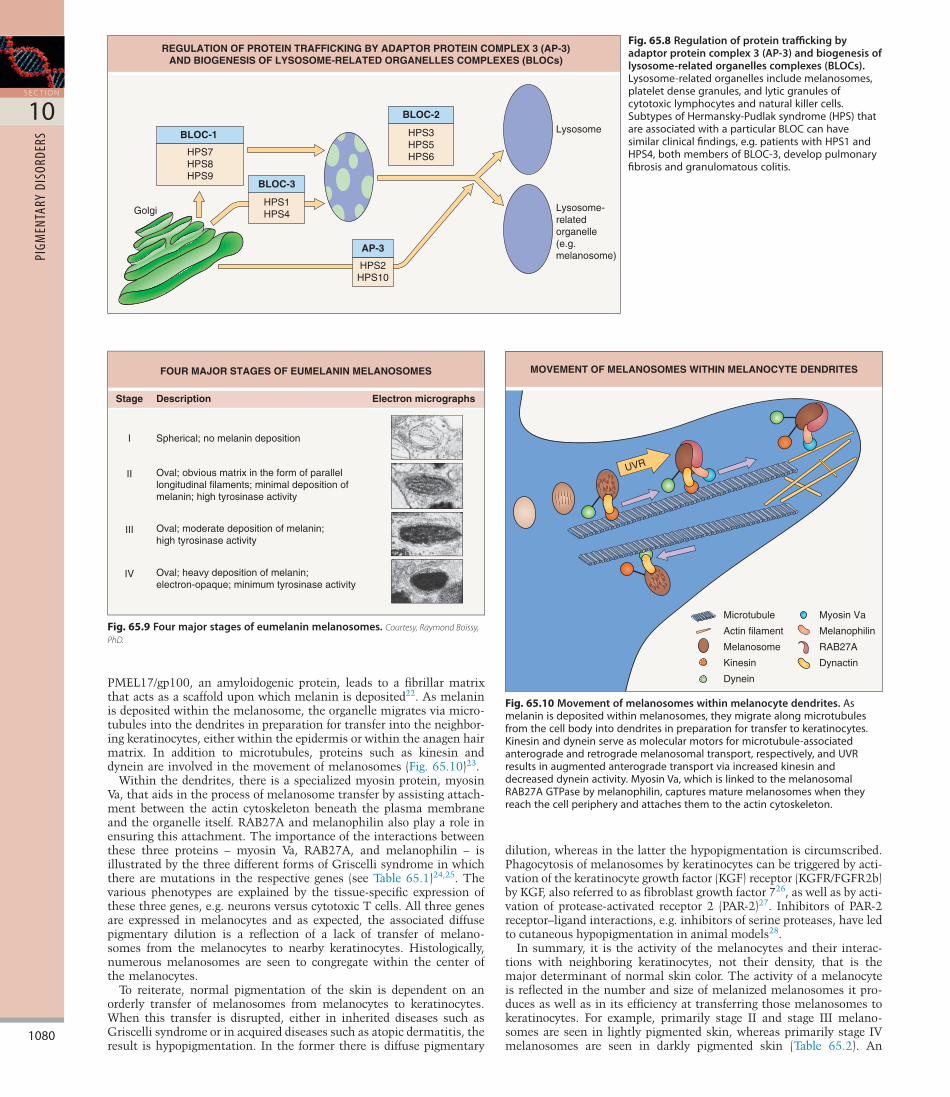
Several of the enzymes in the melanin biosynthetic pathway are glycoproteins that require the attachment of sugars in order to gain full function. For this reason, they undergo posttranslational modification in the ER and the Golgi apparatus and then join up with the matrix proteins (e.g. PMEL/PMEL17/gp100) to initiate melanogenesis. The targeting of proteins to the plasma membrane versus intracytoplasmic organelles and the targeting of specific proteins to the correct type of organelle are complicated processes. This triaging requires the equivalent of traffic police within the cell, an example of which is adaptor protein complex 3 (AP3). Mutations in the gene that encodes the β3A subunit of AP3 are responsible for a subset of patients with Hermansky–Pudlak syndrome (HPS2)19. Other forms of HPS are due to dysfunction of the proteins that serve as components of biogenesis of lysosomerelated organelles complexes (BLOCs; Fig. 65.8)20. It then follows that these patients have dysfunction of more than one intracytoplasmic organelle, i.e. not just melanosomes but other lysosomerelated organelles such as NK cell granules and platelet dense granules (see Table 65.1). The associated hypopigmentation can be explained by the failure to efficiently deliver melanogenic proteins, e.g. tyrosinase, TYRP1, to the melanosome.
The progression of a melanosome from an organelle that lacks melanin to one that is fully melanized is arbitrarily divided, for convenience, into four stages (Fig. 65.9). Proteins that play a key role in the biogenesis of earlystage melanosomes are PMEL/PMEL17/gp100, MART1/MelanA, and the ocular albinism type 1 (OA1) Gproteincoupled receptor21. Cleavage and processing of PMEL/
Fig. 65.6 Dihydroxyphenylalanine (DOPA)-stained epidermal sheet.Followingincubationwith5mML-DOPAfor4–5hours,theepidermalmelanocytesturnblackbecausetheycontaintheenzymetyrosinase,whichconvertsDOPAtoblackDOPA-melanin.Notethemultipledendritesandregularspacingofthemelanocytes.
Fig. 65.7 Formation of a melanosome and pathophysiology of oculocutaneous albinism type 1 (OCA1).Afterglycosylationandprocessingwithintheendoplasmicreticulum(ER)andGolgiapparatus,severaloftheenzymesinvolvedintheformationofmelanin(includingtyrosinase)arepackagedinvesiclesandthencombinewiththematrixproteins(e.g.PMEL/PMEL17/gp100).Asmoremelaninisdepositedwithinthemelanosomes,theymigrateintothedendritesinpreparationfortheirtransferintoneighboringkeratinocytes.InpatientswithOCA1andOCA3(triangularinsert),tyrosinasepolypeptidesareretainedtoolongwithinthelumenoftheRERandthenbecometargetsfordestructionwithinproteasomes.Thus,theyarenotincorporatedintomelanosomes.TheprimarymatrixproteinisPMEL/PMEL17/gp100(recognizedbyHMB45antibody);formationofthematrixfibersrequirescleavageofthisproteinbyaproproteinconvertase51.Sortingofproteinstothecorrectorganellesisacomplexprocessanditrequiresregulatorssuchasadaptorproteincomplex3(AP-3;seeFig.65.8);inthecaseoftyrosinase,specificdileucineresiduesinitscytoplasmictailaidinthesorting.M,matrixproteins;RER,roughendoplasmicreticulum;SER,smoothendoplasmicreticulum;T,tyrosinase.
RER
SER
nucleus
Golgiapparatus
proteasome
abnormal/misfolded nascentprotein(e.g. T in OCA1)
ribosome
dendrite
late endosome myosin Va
T
T
M
M
T
plasmamembrane
actincytoskeleton
FORMATION OF A MELANOSOME AND PATHOPHYSIOLOGY OF OCA1
proenzymes such as proteases and melanosomes protect against melanin precursors (e.g. phenols, quinones) that can oxidize lipid membranes.
The melanosome contains both matrix proteins (chiefly PMEL/PMEL17/gp100), which form a scaffolding upon which the melanin is deposited, and enzymes such as tyrosinase that regulate the biosynthesis of melanin. Following their formation via ribosomes, all these proteins are found within the rough endoplasmic reticulum (RER;
1080
S E C T I O N
10
PIGM
ENTA
RY D
ISOR
DERS
dilution, whereas in the latter the hypopigmentation is circumscribed. Phagocytosis of melanosomes by keratinocytes can be triggered by activation of the keratinocyte growth factor (KGF) receptor (KGFR/FGFR2b) by KGF, also referred to as fibroblast growth factor 726, as well as by activation of proteaseactivated receptor 2 (PAR2)27. Inhibitors of PAR2 receptor–ligand interactions, e.g. inhibitors of serine proteases, have led to cutaneous hypopigmentation in animal models28.
In summary, it is the activity of the melanocytes and their interactions with neighboring keratinocytes, not their density, that is the major determinant of normal skin color. The activity of a melanocyte is reflected in the number and size of melanized melanosomes it produces as well as in its efficiency at transferring those melanosomes to keratinocytes. For example, primarily stage II and stage III melanosomes are seen in lightly pigmented skin, whereas primarily stage IV melanosomes are seen in darkly pigmented skin (Table 65.2). An
PMEL17/gp100, an amyloidogenic protein, leads to a fibrillar matrix that acts as a scaffold upon which melanin is deposited22. As melanin is deposited within the melanosome, the organelle migrates via microtubules into the dendrites in preparation for transfer into the neighboring keratinocytes, either within the epidermis or within the anagen hair matrix. In addition to microtubules, proteins such as kinesin and dynein are involved in the movement of melanosomes (Fig. 65.10)23.
Within the dendrites, there is a specialized myosin protein, myosin Va, that aids in the process of melanosome transfer by assisting attachment between the actin cytoskeleton beneath the plasma membrane and the organelle itself. RAB27A and melanophilin also play a role in ensuring this attachment. The importance of the interactions between these three proteins – myosin Va, RAB27A, and melanophilin – is illustrated by the three different forms of Griscelli syndrome in which there are mutations in the respective genes (see Table 65.1)24,25. The various phenotypes are explained by the tissuespecific expression of these three genes, e.g. neurons versus cytotoxic T cells. All three genes are expressed in melanocytes and as expected, the associated diffuse pigmentary dilution is a reflection of a lack of transfer of melanosomes from the melanocytes to nearby keratinocytes. Histologically, numerous melanosomes are seen to congregate within the center of the melanocytes.
To reiterate, normal pigmentation of the skin is dependent on an orderly transfer of melanosomes from melanocytes to keratinocytes. When this transfer is disrupted, either in inherited diseases such as Griscelli syndrome or in acquired diseases such as atopic dermatitis, the result is hypopigmentation. In the former there is diffuse pigmentary
Fig. 65.8 Regulation of protein trafficking by adaptor protein complex 3 (AP-3) and biogenesis of lysosome-related organelles complexes (BLOCs).Lysosome-relatedorganellesincludemelanosomes,plateletdensegranules,andlyticgranulesofcytotoxiclymphocytesandnaturalkillercells.SubtypesofHermansky-Pudlaksyndrome(HPS)thatareassociatedwithaparticularBLOCcanhavesimilarclinicalfindings,e.g.patientswithHPS1andHPS4,bothmembersofBLOC-3,developpulmonaryfibrosisandgranulomatouscolitis.
REGULATION OF PROTEIN TRAFFICKING BY ADAPTOR PROTEIN COMPLEX 3 (AP-3)AND BIOGENESIS OF LYSOSOME-RELATED ORGANELLES COMPLEXES (BLOCs)
Lysosome
Lysosome-relatedorganelle (e.g. melanosome)
BLOC-1
HPS7HPS8HPS9
AP-3
HPS2HPS10
Golgi
BLOC-2
HPS3HPS5HPS6
BLOC-3
HPS1HPS4
Fig. 65.9 Four major stages of eumelanin melanosomes.Courtesy, Raymond Boissy, PhD.
Stage Description Electron micrographs
I Spherical; no melanin deposition
Oval; obvious matrix in the form of parallellongitudinal filaments; minimal deposition ofmelanin; high tyrosinase activity
Oval; moderate deposition of melanin;high tyrosinase activity
Oval; heavy deposition of melanin;electron-opaque; minimum tyrosinase activity
II
III
IV
FOUR MAJOR STAGES OF EUMELANIN MELANOSOMES
Fig. 65.10 Movement of melanosomes within melanocyte dendrites.Asmelaninisdepositedwithinmelanosomes,theymigratealongmicrotubulesfromthecellbodyintodendritesinpreparationfortransfertokeratinocytes.Kinesinanddyneinserveasmolecularmotorsformicrotubule-associatedanterogradeandretrogrademelanosomaltransport,respectively,andUVRresultsinaugmentedanterogradetransportviaincreasedkinesinanddecreaseddyneinactivity.MyosinVa,whichislinkedtothemelanosomalRAB27AGTPasebymelanophilin,capturesmaturemelanosomeswhentheyreachthecellperipheryandattachesthemtotheactincytoskeleton.
MOVEMENT OF MELANOSOMES WITHIN MELANOCYTE DENDRITES
Microtubule
Actin filament
Melanosome
Kinesin
Dynein
UVR
Myosin Va
Melanophilin
RAB27A
Dynactin

1081
C H A P T E R
65
Mel
anoc
yte
Biol
ogy
additional factor is the rate of degradation of the melanosomes once they are transferred to the surrounding keratinocytes and this is related in part to the size of the individual melanosomes. The smaller melanosomes of lightly pigmented skin are clustered in groups of two to ten within secondary lysosomes in the keratinocytes and are degraded by the mid stratum spinosum (Table 65.3)15. In darkly pigmented skin, the melanosomes are larger and singly dispersed within lysosomes of the keratinocytes; they are degraded more slowly, such that melanin granules can still be found in the stratum corneum.
REGULATION OF MELANIN BIOSYNTHESISThis section will begin with a review of the melanin biosynthetic pathway and then examine the factors, both external and internal, that can influence the level of melanin production. The “starting material” for the production of melanin, both the brown–black eumelanin and the yellow–red pheomelanin, is the amino acid tyrosine. The key regulatory enzyme in the pathway is tyrosinase, which controls the initial biochemical reactions in this pathway (Fig. 65.11). It should then come as no surprise that the initial investigations into the molecular basis of OCA focused on the gene that encodes tyrosinase.
In OCA1A, the form of OCA where mutations in both copies of the tyrosinase gene lead to complete loss of enzyme activity, no melanin is found in the hair, skin, or eyes (see Table 65.1). However, in OCA1B, where there is decreased enzyme activity, pheomelanin is produced, especially in the hair as the patient ages. The formation of pheomelanin requires less tyrosinase activity than does the formation of eumelanin (see Fig. 65.11) and therefore the formation of pheomelanin can be thought of as a default pathway.
The activity of tyrosinase is enhanced by DOPA and is stabilized by tyrosinaserelated protein 1 (TYRP1) (see below). Competitive inhibitors of tyrosinase activity include hydroquinone, which is used to treat disorders of hyperpigmentation such as melasma, and Lphenylalanine. In patients with phenylketonuria (PKU), there is a diffuse pigmentary
Fig. 65.11 The melanin biosynthetic pathway.ThepathwayincludesthesitesofdysfunctioninOCA1(tyrosinase)andOCA3(TYRP1).Thetwomajorformsofmelaninintheskinandhairarebrown–blackeumelaninandyellow–redpheomelanin.Theenzymesaretransmembraneproteinslocatedwithinthemelanosome.DHI,5,6-dihydroxyindole;DHICA,5,6-dihydroxyindole-2-carboxylicacid;DOPA,dihydroxyphenylalanine;MW,molecularweight;TYRP,tyrosinase-relatedprotein.Adapted from Hearing VJ. Determination of melanin synthetic pathways. J Invest Dermatol. 2011;131:E8–E11.
HO HO
HO
NH2 NH2
CO2H CO2HHO
Dopachrome tautomerase(DCT)/TYRP2
Tyrosinase or TYRP1
Tyrosinase
Cysteine
Tyrosine DOPAquinone CycloDOPA then DOPA DOPAquinone
DQ
DOPA
DQ
DOPA
DHICA
DHICA-melaninbrown, slightly soluble,
intermediate MW
Pheomelaninyellow/red, alkali-soluble, low MW
DHI-melaninblack, insoluble,
high MW
Eumelanin
*Or 2-S-cysteinylDOPA
DOPAchrome DHI
5-S-cysteinylDOPA*
THE MELANIN BIOSYNTHETIC PATHWAY
HO NH CO2H
HO
NH2
CO2H
+ CO2
O
ONH2
CO2HO
O
O
HONH2
NH2
CO2H
CO2H
CD-quinones
o-Quinoneimine
1, o-Benzothiazine intermediates
HO
HO NH
HO
N CO2H
S
O2O2
Tyrosinase
Table 65.2 Variation of predominant melanosomal stages with level of cutaneous pigmentation.
VARIATION OF PREDOMINANT MELANOSOMAL STAGES WITH LEVEL OF CUTANEOUS PIGMENTATION
Pigmentation of skin
Predominant melanosomal stages
Melanocytes Keratinocytes
Fair II,III OccasionalIII
Medium II,III,IV III,IV
Dark IV>III IV
Table 65.3 Melanosomes in lightly pigmented versus darkly pigmented skin.
MELANOSOMES IN LIGHTLY PIGMENTED VERSUS DARKLY PIGMENTED SKIN
Lightly pigmented skin
Darkly pigmented skin
Melanization StagesII,III StageIV
Size(diameter) 0.3–0.5microns 0.5–0.8microns
Numberpercell <20 >200
Distributionofmelanosomeswithinthelysosomesofkeratinocytes
Groupsof2–10 Single
Degradation Fast Slow

1082
S E C T I O N
10
PIGM
ENTA
RY D
ISOR
DERS
Among the multiple protein products of the proopiomelanocortin (POMC) gene are adrenocorticotropic hormone (ACTH), βendorphin, and the three forms of MSH (α, β, and γ) (Fig. 65.13); in humans, αMSH represents the major biologically active form of MSH. The primary site of expression of POMC is the pituitary gland; however, other sites of expression include the testis, the endothelium, and, of particular importance, epidermal keratinocytes. Although MSH is classically associated with the pigmentary system, this peptide has a wide range of biologic properties, including suppression of inflammation and regulation of body weight. For example, mutations in POMC can result in severe earlyonset obesity, adrenal insufficiency, and red hair33. It then follows that mutations in the genes that encode the melanocortin receptors to which the protein products of POMC bind can lead to similar clinical findings; for example, mutations in MC4R are associated with morbid obesity and variant alleles in MC1R are associated with red hair (see below). Of note, subcutaneous afamelanotide (4norleucyl7phenylalanineαMSH), which has enhanced binding to MC1R compared to αMSH, can lead to cutaneous hyperpigmentation. When subcutaneous implants of afamelanotide were administered monthly, patients with erythropoietic protoporphyria experienced fewer phototoxic reactions and could tolerate more direct sunlight without pain while patients with vitiligo had more rapid and extensive repigmentation in response to treatment with UVB.
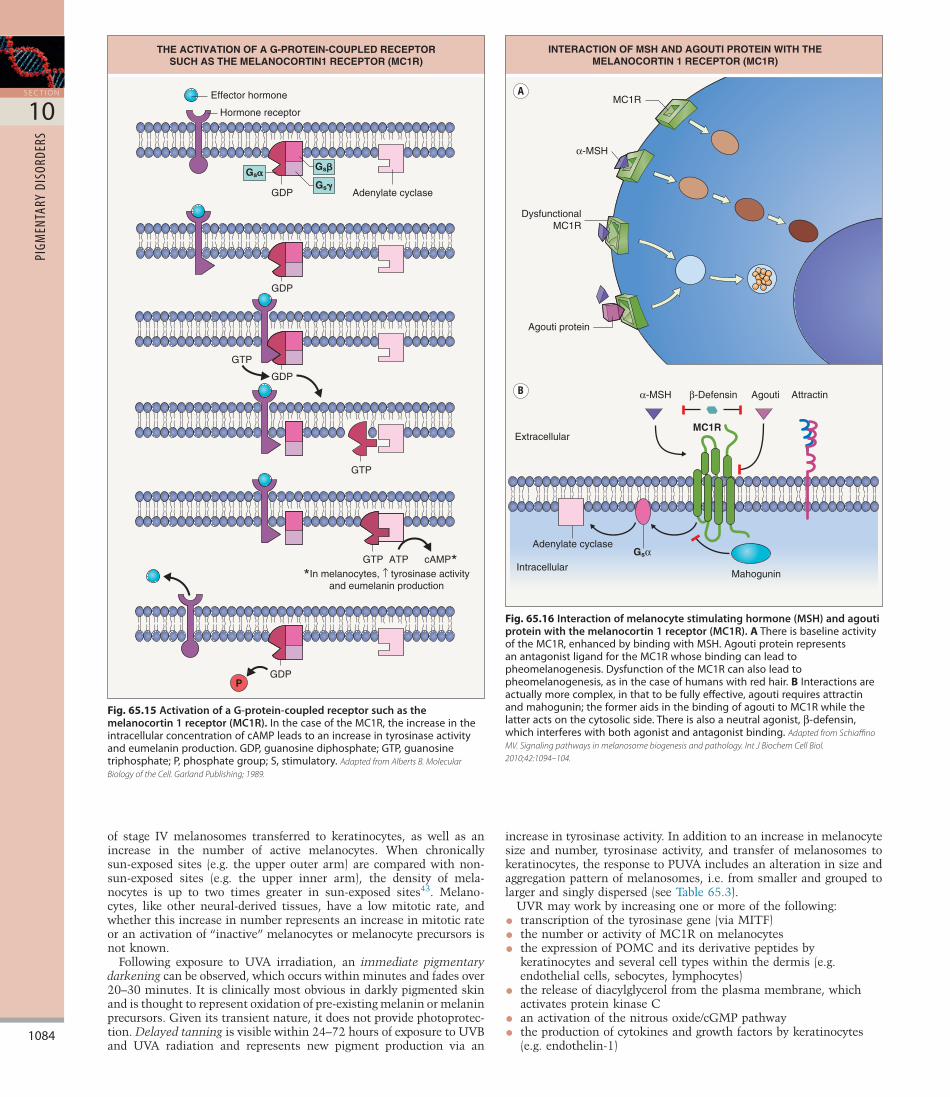
As outlined in Table 65.4, there are five major melanocortin receptors, all of which have seven transmembrane domains (Fig. 65.14). Although the MC1R is present on a variety of cells within the skin, from endothelial cells to fibroblasts, the highest density of this receptor is found on melanocytes34. As in the case of the βadrenergic receptor, the MC1R is a Gproteincoupled receptor, i.e. it uses proteins that bind guanosine triphosphate (GTP) and guanosine diphosphate (GDP) as intermediary messengers (Fig. 65.15). Following the binding of MSH to the MC1R, the protein pair interacts with a complex of G proteins. The GTPGsα subunit then activates adenylate cyclase, leading to increased production of cyclic adenosine monophosphate (cAMP) within the melanocyte. An increase in the intracellular concentration of cAMP leads to an increase in tyrosinase activity and eumelanin production via MITF (see Fig. 65.4). If the MC1R is dysfunctional and fails to initiate a significant rise in the intracellular level of cAMP, then pheomelanogenesis is favored (Fig. 65.16). Of note, the majority of individuals with red hair are compound heterozygotes or homozygotes for a variant R allele in the gene that encodes the MC1R1,35 (see Fig. 65.14).
MC1R also interacts with a protein known as the agouti protein (mouse) or agouti signaling protein (ASIP; human). Agouti is a term
dilution due to elevated levels of Lphenylalanine resulting from a deficiency in the enzyme Lphenylalanine hydroxylase that converts Lphenylalanine to Ltyrosine15. The characteristic blonde hair of PKU can undergo darkening when the patient is on a lowphenylalanine diet. Of note, tyrosinase is a copperrequiring enzyme and it has two copperbinding sites. Rare cases of copper deficiency can lead to diffuse cutaneous pigmentary dilution, and in patients with Menkes disease, where a transmembrane Cu2+transporting ATPase that delivers copper to the transGolgi network and melanosomes is dysfunctional29, the kinky hair is hypopigmented.
In a test tube, LDOPA can spontaneously oxidize to form melanin, an insoluble biopolymer. For this reason, it was originally thought that tyrosinase was the sole enzyme involved in melanin biosynthesis. However, by the late 1970s, it was becoming clear that there were additional control points in the pathway (see Fig. 65.11). For example, dopachrome tautomerase, also known as tyrosinaserelated protein 2 (TYRP2), which like TYRP1 shares similarities in its amino acid sequence with tyrosinase, converts DOPAchrome to 5,6dihydroxyindole2carboxylic acid (DHICA). In mice and humans, TYRP1 stabilizes tyrosinase30 and mutations in both copies of TYRP1 lead to OCA3 (see Table 65.1).
Decreased function of yet another transmembrane protein, the P protein, leads to OCA231 (Fig. 65.12). Based upon its amino acid sequence, a prediction was made that the P protein was involved in the transport of small molecules across the membrane of the melanosome. Tyrosine, the initial precursor in the melanin biosynthetic pathway, was considered the most likely candidate for transmembrane transport. However, the nature of what might be transported by the P protein remains unclear. Data suggest that the P protein regulates processing and trafficking of tyrosinase, possibly via control of pH or glutathione content within intracellular compartments32.
Melanins represent a group of complex polymers whose functions vary from camouflage to the quenching of oxidative free radicals generated via exposure to UV radiation (UVR). The level and type of melanin production is a complex interplay of the activity of the various enzymes involved in the biosynthetic pathway as well as the activity of proteins such as the P protein and those that stabilize the activity of tyrosinase (e.g. TYRP1). Several factors are known to influence the activity of these key proteins of melanogenesis, and they include αMSH, basic fibroblast growth factor (bFGF), endothelin1, KIT ligand, and UVR (see below).
Fig. 65.12 The P protein within the lipid bilayer.Thepolypeptidehas12putativetransmembranedomains.Thesolutecarrierfamily45member2(previouslyknownasmembrane-associatedtransporterprotein),whichisdysfunctionalinOCA4,hasasimilarconfigurationwithintheplasmamembrane.Adapted from Rinchik EM, Bultman SJ, Horsthemke B, et al. A gene for the mouse pink-eyed dilution locus and for human type II oculocutaneous albinism. Nature. 1993;361:72–6.
THE P PROTEIN WITHIN THE LIPID BILAYER
NH3+
− −
−
+
1 471 08
1 2 3 4 5 6 7 8 9 10 11 12
3 253 74
3 253 45
4 914 42
6 716 12
7 247 73
6 076 46
5 335 03
6 676 08
7 957 16
8 73
Cytoplasmic
Luminal
COO–
8 43
4 704 10
1 691 99
3 133 92
3 073 27
4 044 94
6 736 84
7 127 81
6 846 54
5 415 30
6 697 10
7 777 08
8 818 21
3 583 28
Fig. 65.13 Post-translational processing of the POMC polypeptide.PituitaryhypersecretionofACTHand/orα-MSHcanleadtogeneralizedhyperpigmentationinpatientswithAddisondisease.ac,acetylated;ACTH,adrenocorticotropichormone;des,desacetyl;END,endorphin;JP,joiningpeptide;LPH,lipotropichormone;MSH,melanocytestimulatinghormone;PC,prohormone-convertingenzyme;POMC,proopiomelanocortin.
POST-TRANSLATIONAL PROCESSING OF THE POMC POLYPEPTIDE
POMC1 241
ACTH β-LPHJPPC1
PC2
Pro γ-MSH
β-ENDACTH1-17
des α-MSH
ac α-MSH

1083
C H A P T E R
65
Mel
anoc
yte
Biol
ogy
Fig. 65.14 Melanocortin 1 receptor (MC1R) within the plasma membrane of a melanocyte.MC1RhasseventransmembranedomainsandisaG-coupledreceptor(seeFig.65.15).TherearenumerousgeneticvariantsoftheMC1R.ThenullorhypomorphicRallelesD84E,R151C,R160W,andD294Hhaveredhairoddsratiosof62,118,50,and94,respectively;thelowpenetrancerallelesofV60L,V92M,andR163Qhaveredhairoddsratiosof6,5,and2,respectively52.Ingeneral,R/Rindividualshaveredhairandlightskin.
Cytoplasm
MELANOCORTIN-1 RECEPTOR (MC1R) WITHIN THE PLASMA MEMBRANE OF A MELANOCYTE
NH2
Major R alleles: D84ER151CR160WD294H
COOH
Plasma membrane
Table 65.4 Major forms of the melanocortin receptor (MCR).ACTH,adrenocorticotropichormone;MSH,melanocytestimulatinghormone.
MAJOR FORMS OF THE MELANOCORTIN RECEPTOR
Receptor Distribution, major (minor) Ligands
MC1R* Melanocytes(keratinocytes,fibroblasts,endothelialcells,antigen-presentingcells)
α-MSH,ACTH>β-MSH
MC2R Adrenalcortex(adipocytes) ACTH
MC3R† Brain(gut,placenta) α-,β-,γ–MSH,ACTH
MC4R† Brain α-,β-MSH,ACTH
MC5R Peripheraltissues,e.g.sebaceousglands,fibroblasts,adipocytes
α-,β-MSH,ACTH
*Antagonistic ligands – agouti protein (mouse) and agouti signaling protein (ASIP; human).†Antagonistic ligand – agouti-related protein (AGRP).
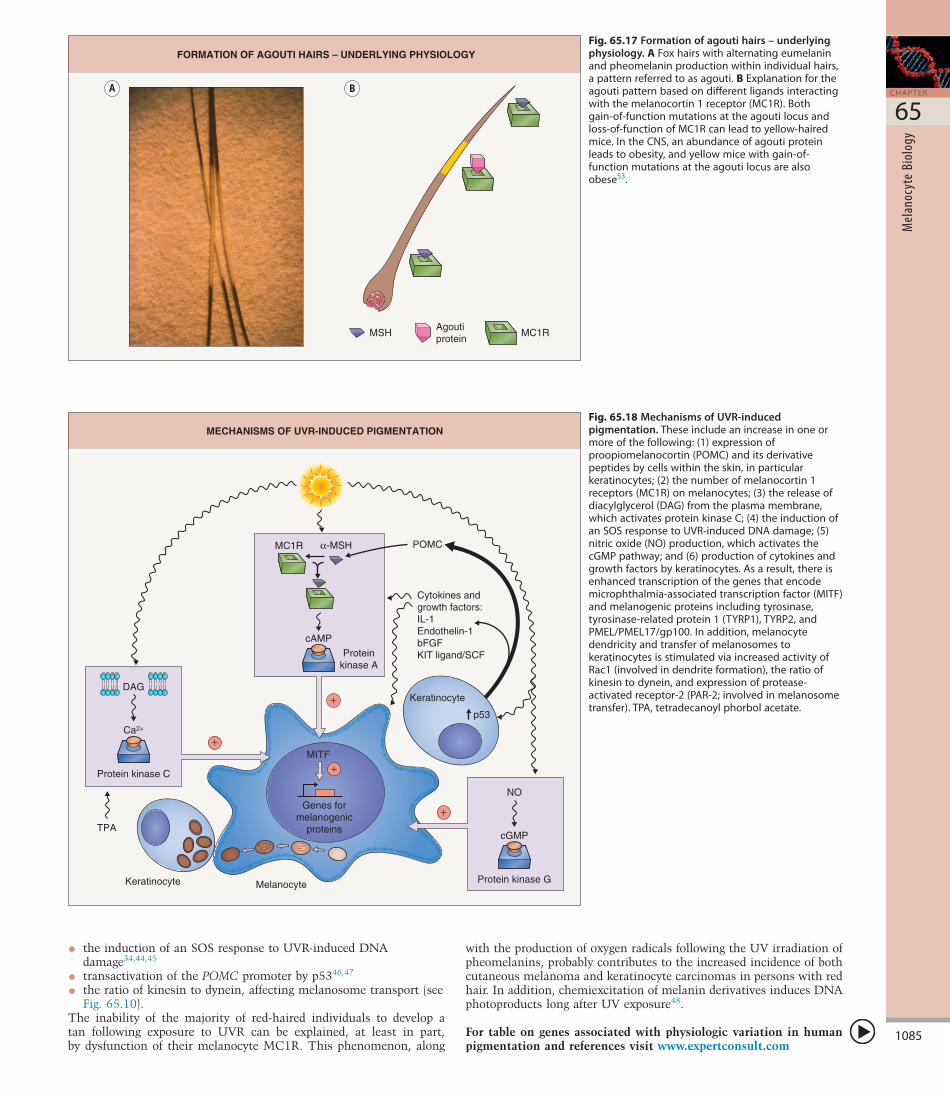
used to describe the banding of hairs seen in some mammals, including dogs, foxes and mice, that is due to alternating production of eumelanin and pheomelanin (Fig. 65.17). The production of agouti protein by the cells in the hair follicle papillae is cyclic, and, when the agouti protein is present, it effectively competes with MSH and the formation of pheomelanin within pheomelanosomes is favored36,37 (see Fig. 65.16). Compared to eumelanosomes, pheomelanosomes are characterized by a more spherical shape and the presence of an unstructured matrix with vesicular bodies.
An enhancement of pigment production can be seen following exposure of melanocytes to agents that increase intracytoplasmic levels of cAMP such as cholera toxin, forskolin38, dibutyryl cAMP, and MSH (see Fig. 65.15). The activation of protein kinase A (PKA) by cAMP leads to the phosphorylation of various proteins, which can result in their activation. One of the proteins that is phosphorylated by PKA is the cAMP responseelement binding protein (CREB), which functions as a transcription factor, regulating the expression of other genes, including MITF (see Fig. 65.4). Patients with the McCune–Albright syndrome (polyostotic fibrous dysplasia) are mosaics for an activating mutation in the gene that encodes the G protein Gsα (see Fig. 65.15). As a result, the cAMP cascade is permanently “turned on” and, with continued transcription of the CREBcontrolled genes, there is hyperplasia of the bones and endocrine organs, albeit in a mosaic pattern. Presumably within the melanocytes of the segmental caféaulait macules typical
of this disorder, the increase in intracellular cAMP leads to increased tyrosinase activity and eumelanin production.
Exposure of melanocytes to phorbol esters, e.g. tetradecanoyl phorbol acetate (TPA), can lead to increased melanin formation via the activation of protein kinase C (PKC) (Fig. 65.18). Growth factors, including bFGF and KIT ligand, can also lead to an increase in the pigment content within melanocytes whereas KIT receptor inhibitors (e.g. imatinib) can lead to hypopigmentation. Endothelin1, a small peptide originally isolated from endothelial cells, is produced by keratinocytes and can lead to an increase in tyrosinase activity followed by an increase in melanin production.
Multiple genes have been implicated as playing a role in normal pigment variation in humans, including: (1) MC1R and ASIP; (2) several genes that play a role in OCA, e.g. TYR, OCA2, TYRP1, SLC45A2, SLC24A5 (see Table 65.1); (3) KITLG which encodes KIT ligand; (4) TPCN2 which encodes an ion channel transporter; and (5) IRF4 which encodes interferon regulatory factor 42–4. The presence of a variant (rather than the African ancestral) allele of SLC24A5, whose protein product is a putative cation exchanger in the melanosomal membrane, correlates with lighter skin color5. In addition, variants in OCA2 are thought, at least in part, to determine the normal phenotypic variation in human eye color2. A haplotype with several polymorphisms in ASIP (and presumably a gain of ASIP function) has been associated with red or blonde hair, freckling and a tendency to burn, while variants that lead to destabilized ASIP mRNA have been associated with darker skin phototypes3,39,40. A polymorphism in the regulatory region of KITLG that reduces its responsiveness to the WNTactivated transcription factor LEF1 is associated with blonde hair41, and in mice, decreased lef1 leads to lightcolored hair. Of note, some lossoffunction mutations in MC1R confer a risk for developing cutaneous melanoma, independent of pigmentary phenotype35.
In humans, graying or whitening of hair is a normal aging phenomenon. There is evidence that reactive oxygen species accumulate within affected hair follicles and lead to oxidative damage of hair follicle melanocytes. Both hair bulb melanocytes and precursor cells in the bulge region can be affected42. In addition, melanocytes that disappear fail to express TYRP2 and SOX10. The role of SCF production by KROX20+ cells within the hair bulb was discussed previously.
Ultraviolet Radiation (UVR)Following a single exposure to UVR, an increase in the size of melanocytes can be observed, along with an increase in tyrosinase activity15. Repeated exposures to UVR lead to an increase in the number
1084
S E C T I O N
10
PIGM
ENTA
RY D
ISOR
DERS
increase in tyrosinase activity. In addition to an increase in melanocyte size and number, tyrosinase activity, and transfer of melanosomes to keratinocytes, the response to PUVA includes an alteration in size and aggregation pattern of melanosomes, i.e. from smaller and grouped to larger and singly dispersed (see Table 65.3).
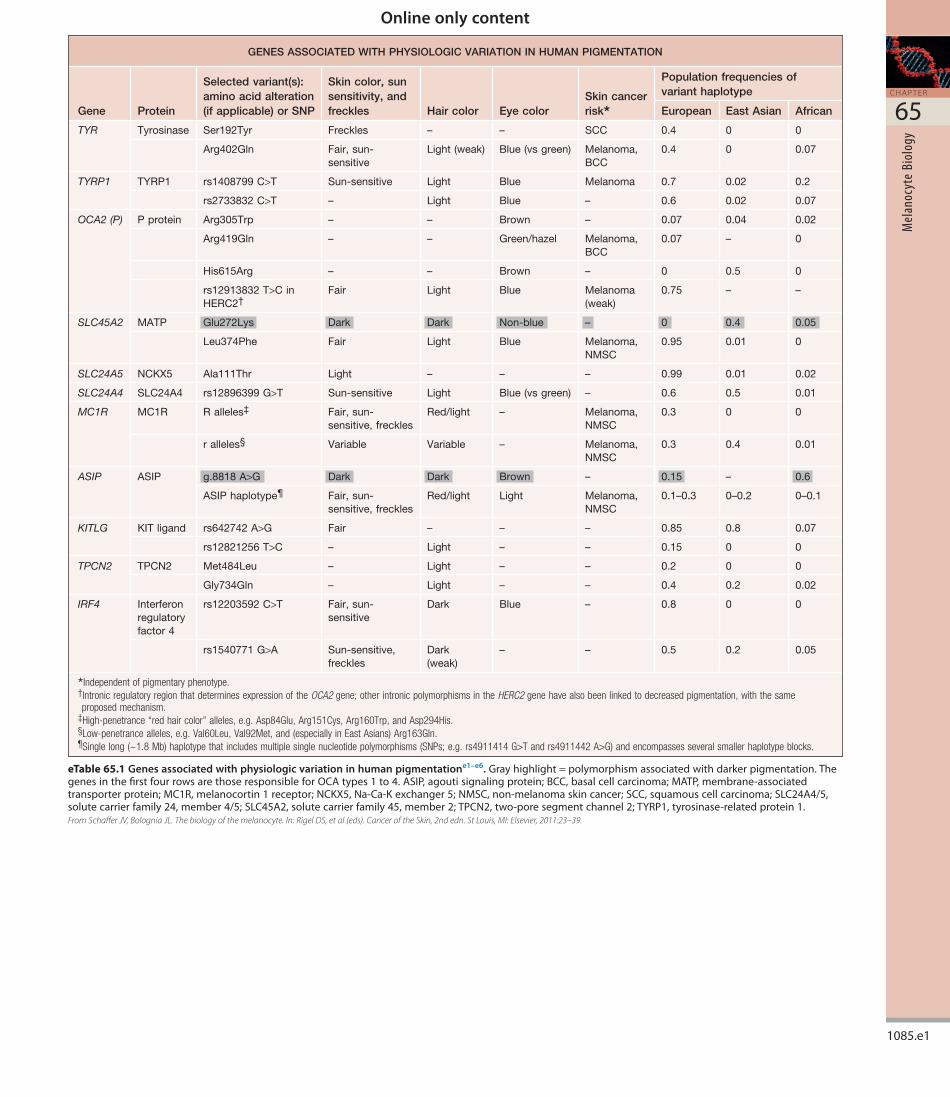
UVR may work by increasing one or more of the following:
• transcription of the tyrosinase gene (via MITF)
• the number or activity of MC1R on melanocytes
• the expression of POMC and its derivative peptides by keratinocytes and several cell types within the dermis (e.g. endothelial cells, sebocytes, lymphocytes)
• the release of diacylglycerol from the plasma membrane, which activates protein kinase C
• an activation of the nitrous oxide/cGMP pathway
• the production of cytokines and growth factors by keratinocytes (e.g. endothelin1)
of stage IV melanosomes transferred to keratinocytes, as well as an increase in the number of active melanocytes. When chronically sunexposed sites (e.g. the upper outer arm) are compared with non sunexposed sites (e.g. the upper inner arm), the density of melanocytes is up to two times greater in sunexposed sites43. Melanocytes, like other neuralderived tissues, have a low mitotic rate, and whether this increase in number represents an increase in mitotic rate or an activation of “inactive” melanocytes or melanocyte precursors is not known.
Following exposure to UVA irradiation, an immediate pigmentary darkening can be observed, which occurs within minutes and fades over 20–30 minutes. It is clinically most obvious in darkly pigmented skin and is thought to represent oxidation of preexisting melanin or melanin precursors. Given its transient nature, it does not provide photoprotection. Delayed tanning is visible within 24–72 hours of exposure to UVB and UVA radiation and represents new pigment production via an
Fig. 65.15 Activation of a G-protein-coupled receptor such as the melanocortin 1 receptor (MC1R).InthecaseoftheMC1R,theincreaseintheintracellularconcentrationofcAMPleadstoanincreaseintyrosinaseactivityandeumelaninproduction.GDP,guanosinediphosphate;GTP,guanosinetriphosphate;P,phosphategroup;S,stimulatory.Adapted from Alberts B. Molecular Biology of the Cell. Garland Publishing; 1989.
THE ACTIVATION OF A G-PROTEIN-COUPLED RECEPTORSUCH AS THE MELANOCORTIN1 RECEPTOR (MC1R)
Effector hormone
Hormone receptor
Adenylate cyclaseGDP
GDP
GDP
GTP
GDP
GTP ATP cAMP**In melanocytes, ↑ tyrosinase activity
and eumelanin production
GTP
Gs�Gs�
Gs�
P
Fig. 65.16 Interaction of melanocyte stimulating hormone (MSH) and agouti protein with the melanocortin 1 receptor (MC1R).AThereisbaselineactivityoftheMC1R,enhancedbybindingwithMSH.AgoutiproteinrepresentsanantagonistligandfortheMC1Rwhosebindingcanleadtopheomelanogenesis.DysfunctionoftheMC1Rcanalsoleadtopheomelanogenesis,asinthecaseofhumanswithredhair.BInteractionsareactuallymorecomplex,inthattobefullyeffective,agoutirequiresattractinandmahogunin;theformeraidsinthebindingofagoutitoMC1Rwhilethelatteractsonthecytosolicside.Thereisalsoaneutralagonist,β-defensin,whichinterfereswithbothagonistandantagonistbinding.Adapted from Schiaffino MV. Signaling pathways in melanosome biogenesis and pathology. Int J Biochem Cell Biol. 2010;42:1094–104.
INTERACTION OF MSH AND AGOUTI PROTEIN WITH THEMELANOCORTIN 1 RECEPTOR (MC1R)
α-MSH
MC1R
DysfunctionalMC1R
Agouti protein
A
B
Extracellular
Intracellular
α-MSH Agouti Attractinβ-Defensin
Gsα
MC1R
Adenylate cyclase
Mahogunin
1085
C H A P T E R
65
Mel
anoc
yte
Biol
ogy
with the production of oxygen radicals following the UV irradiation of pheomelanins, probably contributes to the increased incidence of both cutaneous melanoma and keratinocyte carcinomas in persons with red hair. In addition, chemiexcitation of melanin derivatives induces DNA photoproducts long after UV exposure48.
For table on genes associated with physiologic variation in human pigmentation and references visit www.expertconsult.com
• the induction of an SOS response to UVRinduced DNA damage34,44,45
• transactivation of the POMC promoter by p5346,47
• the ratio of kinesin to dynein, affecting melanosome transport (see Fig. 65.10).
The inability of the majority of redhaired individuals to develop a tan following exposure to UVR can be explained, at least in part, by dysfunction of their melanocyte MC1R. This phenomenon, along
Fig. 65.17 Formation of agouti hairs – underlying physiology.AFoxhairswithalternatingeumelaninandpheomelaninproductionwithinindividualhairs,apatternreferredtoasagouti.BExplanationfortheagoutipatternbasedondifferentligandsinteractingwiththemelanocortin1receptor(MC1R).Bothgain-of-functionmutationsattheagoutilocusandloss-of-functionofMC1Rcanleadtoyellow-hairedmice.IntheCNS,anabundanceofagoutiproteinleadstoobesity,andyellowmicewithgain-of-functionmutationsattheagoutilocusarealsoobese53.
MSHAgouti protein
MC1R
FORMATION OF AGOUTI HAIRS – UNDERLYING PHYSIOLOGY
A B
Fig. 65.18 Mechanisms of UVR-induced pigmentation.Theseincludeanincreaseinoneormoreofthefollowing:(1)expressionofproopiomelanocortin(POMC)anditsderivativepeptidesbycellswithintheskin,inparticularkeratinocytes;(2)thenumberofmelanocortin1receptors(MC1R)onmelanocytes;(3)thereleaseofdiacylglycerol(DAG)fromtheplasmamembrane,whichactivatesproteinkinaseC;(4)theinductionofanSOSresponsetoUVR-inducedDNAdamage;(5)nitricoxide(NO)production,whichactivatesthecGMPpathway;and(6)productionofcytokinesandgrowthfactorsbykeratinocytes.Asaresult,thereisenhancedtranscriptionofthegenesthatencodemicrophthalmia-associatedtranscriptionfactor(MITF)andmelanogenicproteinsincludingtyrosinase,tyrosinase-relatedprotein1(TYRP1),TYRP2,andPMEL/PMEL17/gp100.Inaddition,melanocytedendricityandtransferofmelanosomestokeratinocytesisstimulatedviaincreasedactivityofRac1(involvedindendriteformation),theratioofkinesintodynein,andexpressionofprotease-activatedreceptor-2(PAR-2;involvedinmelanosometransfer).TPA,tetradecanoylphorbolacetate.
+
+MITF
Keratinocyte Melanocyte
TPA
MC1R
Cytokines andgrowth factors: IL-1Endothelin-1bFGF KIT ligand/SCFProtein
kinase A
POMC
Ca2+
DAG
Protein kinase C
cAMP
α-MSH
Genes formelanogenic
proteins
MECHANISMS OF UVR-INDUCED PIGMENTATION
cGMP
NO
Protein kinase G
Keratinocyte
p53
+
+
Online only content
1085.e1
C H A P T E R
65
Mel
anoc
yte
Biol
ogy
GENES ASSOCIATED WITH PHYSIOLOGIC VARIATION IN HUMAN PIGMENTATION
Gene Protein
Selected variant(s): amino acid alteration (if applicable) or SNP
Skin color, sun sensitivity, and freckles Hair color Eye color
Skin cancer risk*
Population frequencies of variant haplotype
European East Asian African
TYR Tyrosinase Ser192Tyr Freckles – – SCC 0.4 0 0
Arg402Gln Fair, sun-sensitive
Light (weak) Blue (vs green) Melanoma, BCC
0.4 0 0.07
TYRP1 TYRP1 rs1408799 C>T Sun-sensitive Light Blue Melanoma 0.7 0.02 0.2
rs2733832 C>T – Light Blue – 0.6 0.02 0.07
OCA2 (P) P protein Arg305Trp – – Brown – 0.07 0.04 0.02
Arg419Gln – – Green/hazel Melanoma, BCC
0.07 – 0
His615Arg – – Brown – 0 0.5 0
rs12913832 T>C in HERC2†
Fair Light Blue Melanoma (weak)
0.75 – –
SLC45A2 MATP Glu272Lys Dark Dark Non-blue – 0 0.4 0.05
Leu374Phe Fair Light Blue Melanoma, NMSC
0.95 0.01 0
SLC24A5 NCKX5 Ala111Thr Light – – – 0.99 0.01 0.02
SLC24A4 SLC24A4 rs12896399 G>T Sun-sensitive Light Blue (vs green) – 0.6 0.5 0.01
MC1R MC1R R alleles‡ Fair, sun-sensitive, freckles
Red/light – Melanoma, NMSC
0.3 0 0
r alleles§ Variable Variable – Melanoma, NMSC
0.3 0.4 0.01
ASIP ASIP g.8818 A>G Dark Dark Brown – 0.15 – 0.6
ASIP haplotype¶ Fair, sun-sensitive, freckles
Red/light Light Melanoma, NMSC
0.1–0.3 0–0.2 0–0.1
KITLG KIT ligand rs642742 A>G Fair – – – 0.85 0.8 0.07
rs12821256 T>C – Light – – 0.15 0 0
TPCN2 TPCN2 Met484Leu – Light – – 0.2 0 0
Gly734Gln – Light – – 0.4 0.2 0.02
IRF4 Interferon regulatory factor 4
rs12203592 C>T Fair, sun-sensitive
Dark Blue – 0.8 0 0
rs1540771 G>A Sun-sensitive, freckles
Dark (weak)
– – 0.5 0.2 0.05
*Independent of pigmentary phenotype.†Intronic regulatory region that determines expression of the OCA2 gene; other intronic polymorphisms in the HERC2 gene have also been linked to decreased pigmentation, with the same proposed mechanism.
‡High-penetrance “red hair color” alleles, e.g. Asp84Glu, Arg151Cys, Arg160Trp, and Asp294His.§Low-penetrance alleles, e.g. Val60Leu, Val92Met, and (especially in East Asians) Arg163Gln.¶Single long (~1.8 Mb) haplotype that includes multiple single nucleotide polymorphisms (SNPs; e.g. rs4911414 G>T and rs4911442 A>G) and encompasses several smaller haplotype blocks.
eTable 65.1 Genes associated with physiologic variation in human pigmentatione1–e6.Grayhighlight=polymorphismassociatedwithdarkerpigmentation.ThegenesinthefirstfourrowsarethoseresponsibleforOCAtypes1to4.ASIP,agoutisignalingprotein;BCC,basalcellcarcinoma;MATP,membrane-associatedtransporterprotein;MC1R,melanocortin1receptor;NCKX5,Na-Ca-Kexchanger5;NMSC,non-melanomaskincancer;SCC,squamouscellcarcinoma;SLC24A4/5,solutecarrierfamily24,member4/5;SLC45A2,solutecarrierfamily45,member2;TPCN2,two-poresegmentchannel2;TYRP1,tyrosinase-relatedprotein1.From Schaffer JV, Bolognia JL. The biology of the melanocyte. In: Rigel DS, et al (eds). Cancer of the Skin, 2nd edn. St Louis, MI: Elsevier, 2011:23–39.

1086
S E C T I O N
10
PIGM
ENTA
RY D
ISOR
DERS
REFERENCES1. ValverdeP,HealyE,JacksonI,etal.Variantsofthe
melanocyte-stimulatinghormonereceptorgeneareassociatedwithredhairandfairskininhumans.NatGenet1995;11:328–30.
2. SturmRA.Moleculargeneticsofhumanpigmentationdiversity.HumMolGenet2009;18:R9–17.
3. NanH,KraftP,HunterDJ,HanJ.Geneticvariantsinpigmentationgenes,pigmentaryphenotypes,andriskofskincancerinCaucasians.IntJCancer2009;125:909–17.
4. BinstockM,HafeezF,MetchnikoffC,ArronST.Single-nucleotidepolymorphismsinpigmentgenesandnonmelanomaskincancerpredisposition:asystematicreview.BrJDermatol2014;171:713–21.
5. LamasonRL,MohideenMA,MestJR,etal.SLC24A5,aputativecationexchanger,affectspigmentationinzebrafishandhumans.Science2005;310:1782–6.
6. MontoliuL,GronskovK,WeiA-H,etal.Increasingthecomplexity:newgenesandnewtypesofalbinism.PigmentCellMelRes2013;27:11–18.
7. AdameykoI,LallemendF,AquinoJB,etal.Schwanncellprecursorsfromnerveinnervationsareacellularoriginofmelanocytesinskin.Cell2009;139:366–79.
8. LinJY,FisherDE.Melanocytebiologyandskinpigmentation.Nature2007;445:843–50.
9. CheliY,OhannaM,BallottiR,BertolottoC.Fifteen-yearquestformicrophthalmia-associatedtranscriptionfactortargetgenes.PigmentCellMelanomaRes2010;23:27–40.
10. ZimmermanAA,BeckerSWJr.PrecursorsofepidermalmelanocytesintheNegrofetus.In:GordonM,editor.PigmentCellBiology.NewYork:AcademicPress;1959.p.159–70.
11. HorikawaT,NorrisDA,JohnsonTW,etal.DOPA-negativemelanocytesintheouterrootsheathofhumanhairfolliclesexpresspremelanosomalantigensbutnotamelanosomalantigenorthemelanosome-associatedglycoproteinstyrosinase,TRP-1,andTRP-2.JInvestDermatol1996;106:28–35.
12. TobinDJ,BystrynJC.Differentpopulationsofmelanocytesarepresentinhairfolliclesandepidermis.PigmentCellRes1996;9:304–10.
12a. LiaoC-P,BookerRC,MorrisonSJ,LeLQ.Identificationofhairshaftprogenitorsthatcreateanicheforhairpigmentation.GeneDevelop2017;31:1–13.
13. HolbrookKA,UnderwoodRA,VogelAM,etal.Theappearance,densityanddistributionofmelanocytesinhumanembryonicandfetalskinrevealedbytheantimelanomamonoclonalantibody,HMB-45.AnatEmbryol1989;180:443–55.
14. JimbowK,QuevedoWCJr,FitzpatrickTB,etal.Someaspectsofmelaninbiology:1950–1975.JInvestDermatol1976;67:72–89.
15. BologniaJL,PawelekJM.Biologyofhypopigmentation.JAmAcadDermatol1988;19:217–55.
16. OrlowSJ.Melanosomesarespecializedmembersofthelysosomallineageoforganelles.JInvestDermatol1995;105:3–7.
17. HalabanR,SvedineS,ChengE,etal.Endoplasmicreticulumretentionisacommondefectassociatedwithtyrosinase-negativealbinism.ProcNatlAcadSciUSA2000;97:5889–94.
18. RonD,WalterP.Signalintegrationintheendoplasmicreticulumunfoldedproteinresponse.NatRevMolCellBiol2007;8:519–29.
19. Dell’AngelicaEC,ShotelersukV,AguilarRC,etal.AlteredtraffickingoflysosomalproteinsinHermansky-Pudlaksyndromeduetomutationsinthebeta3AsubunitoftheAP-3adaptor.MolCell1999;3:11–21.
20. WeiML.Hermansky-Pudlaksyndrome:adiseaseofproteintraffickingandorganellefunction.PigmentCellRes2006;19:19–42.
21. GiordanoF,BonettiC,SuraceEM,etal.Theocularalbinismtype1(OA1)G-protein-coupledreceptorfunctionswithMART-1atearlystagesofmelanogenesistocontrolmelanosomeidentityandcomposition.HumMolGenet2009;18:4530–45.
22. McGlincheyRP,ShewmakerF,McPhieP,etal.TherepeatdomainofthemelanosomefibrilproteinPmel17formstheamyloidcorepromotingmelaninsynthesis.ProcNatlAcadSciUSA2009;106:13731–6.
23. HaraM,YaarM,ByersHR,etal.Kinesinparticipatesinmelanosomalmovementalongmelanocytedendrites.JInvestDermatol2000;114:438–43.
24. PasturalE,BarratFJ,Dufourcq-LagelouseR,etal.Griscellidiseasemapstochromosome15q21andisassociatedwithmutationsinthemyosin-Vagene.NatGenet1997;16:289–92.
25. MénashchéG,HoCH,SanalO,etal.Griscellisyndromerestrictedtohypopigmentationresultsfromamelanophilindefect(GS3)oraMYO5AF-exondeletion(GS1).JClinInvest2003;112:450–6.
26. BelleudiF,PurpuraV,ScrofaniC,etal.ExpressionandsignalingofthetyrosinekinaseFGFR2b/KGFRregulatesphagocytosisandmelanosomeuptakeinhumankeratinocytes.FASEBJ2011;25:170–81.
27. ChoiHI,SohnKC,HongDK,etal.Melanosomeuptakeisassociatedwiththeproliferationanddifferentiationofkeratinocytes.ArchDermatolRes2014;306:59–66.
28. SeibergM,PaineC,SharlowE,etal.Inhibitionofmelanosometransferresultsinskinlightening.JInvestDermatol2000;115:162–7.
29. SettySR,TenzaD,SviderskayaEV,etal.Cell-specificATP7Atransportsustainscopper-dependenttyrosinaseactivityinmelanosomes.Nature2008;454:1142–6.
30. BoissyRE,SakaiC,ZhaoH,etal.Humantyrosinaserelatedprotein-1(TRP-1)doesnotfunctionasaDHICAoxidaseactivityincontrasttomurineTRP-1.ExpDermatol1998;7:198–204.
31. RinchikEM,BultmanSJ,HorsthemkeB,etal.Ageneforthemousepink-eyeddilutionlocusandforhumantypeIIoculocutaneousalbinism.Nature1993;361:72–6.
32. PuriN,GardnerJM,BrilliantMH.AberrantpHofmelanosomesinpink-eyeddilution(p)mutantmelanocytes.JInvestDermatol2000;115:607–13.
33. KrudeH,BiebermannH,LuckW,etal.Severeearlyonsetobesity,adrenalinsufficiencyandredhairpigmentationcausedbyPOMCmutationsinhumans.NatGenet1998;19:155–7.
34. LugerTA,ScholzenT,GrabbeS.Theroleofalphamelanocyte-stimulatinghormoneincutaneousbiology.JInvestigDermatolSympProc1997;2:87–93.
35. RaimondiS,SeraF,GandiniS,etal.MC1Rvariants,melanomaandredhaircolorphenotype.IntJCancer2008;122:2753–60.
36. LuD,WillardD,PatelIR,etal.Agoutiproteinisanantagonistofthemelanocyte-stimulating-hormonereceptor.Nature1994;371:799–802.
37. SuzukiI,TadaA,OllmanMM,etal.Agoutisignalingproteininhibitsmelanogenesisandtheresponseofhumanmelanocytestoalpha-melanotropin.JInvestDermatol1997;108:838–42.
38. D’OrazioJA,NobuhisaT,CuiR,etal.TopicaldrugrescuestrategyandskinprotectionbasedontheroleofMc1rinUV-inducedtanning.Nature2006;443:340–4.
39. GudbjartssonDF,SulemP,StaceySN,etal.ASIPandTYRpigmentationvariantsassociatewithcutaneousmelanomaandbasalcellcarcinoma.NatGenet2008;40:886–91.
40. DuffyDL,ZhaoZZ,SturmRA,etal.Multiplepigmentationgenepolymorphismsaccountforasubstantialproportionofriskofcutaneousmalignantmelanoma.JInvestDermatol2010;130:520–8.
41. GuentherCA,TasicB,LuoL,etal.AmolecularbasisforclassicblondhaircolorinEuropeans.NatGenet2014;46:748–52.
42. ShiY,LuoL-F,LiuX-M,etal.Prematuregrayingasaconsequenceofcompromisedantioxidantactivityinhairbulbmelanocytesandtheirprecursors.PLoSONE2014;9:e93589.
43. GilchrestBA,BlogFB,SzaboG.Effectsofagingandchronicsunexposureonmelanocytesinhumanskin.JInvestDermatol1979;73:141–3.
44. GilchrestBA,ParkHY,EllerMS,etal.Mechanismsofultravioletlight-inducedpigmentation.PhotochemPhotobiol1996;63:1–10.
45. LugerTA.ImmunomodulationbyUVlight:roleofneuropeptides.EurJDermatol1998;8:198–9.
46. CuiR,WidlundHR,FeigeE,etal.Centralroleofp53inthesuntanresponseandpathologichyperpigmentation.Cell2007;128:853–64.
47. OrenM,BartekJ.Thesunnysideofp53.Cell2007;128:826–8.
48. PremiS,WallischS,ManoCM,etal.ChemiexcitationofmelaninderivativesinducesDNAphotoproductslongafterUVexposure.Science2015;347:842–7.
49. WangZQ,SiL,TangQ,etal.Gain-of-functionmutationofKITligandonmelaninsynthesiscausesfamilialprogressivehyperpigmentation.AmJHumGenet2009;84:672–7.
50. RabbaniP,TakeoM,ChouWC,etal.CoordinatedactivationofWntinepithelialandmelanocytestemcellsinitiatespigmentedhairregeneration.Cell2011;145:941–55.
51. LeonhardtRM,VigneronN,RahnerC,CresswellP.ProproteinconvertasesprocessPmel17duringsecretion.JBiolChem2011;286:9321–37.
52. DuffyDL,BoxNF,ChenW,etal.InteractiveeffectsofMC1RandOCA2onmelanomariskphenotypes.HumMolGenet2004;13:447–61.
53. BarshG.FromagoutitoPOMC–100yearsoffatblondemice.NatMed1999;5:984–5.
54. MujahidN,LiangY,MurakamiR,etal.AUV-independenttopicalsmall-moleculeapproachformelaninproductioninhumanskin.CellRep2017;19:2177–84.

Online only content
1086.e1
C H A P T E R
65
Mel
anoc
yte
Biol
ogy
E-REFERENCESe1.SturmRA.Moleculargeneticsofhumanpigmentation
diversity.HumMolGenet2009;18:R9–17.e2.GudbjartssonDF,SulemP,StaceySN,etal.ASIPandTYR
pigmentationvariantsassociatewithcutaneousmelanomaandbasalcellcarcinoma.NatGenet2008;40:886–91.
e3.NanH,KraftP,HunterDJ,etal.Geneticvariantsinpigmentationgenes,pigmentaryphenotypes,andrisk
ofskincancerinCaucasians.IntJCancer2009;125:909–17.
e4.DuffyDL,ZhaoZZ,SturmRA,etal.Multiplepigmentationgenepolymorphismsaccountforasubstantialproportionofriskofcutaneousmalignantmelanoma.JInvestDermatol2010;130:520–8.
e5.LandiMT,BauerJ,PfeifferRM,etal.MCIRgermlinevariantsconferriskforBRAF-mutantmelanoma.Science2006;313:521–2.
e6.ChatzinasiouF,LillCM,KypreouK,etal.Comprehensivefieldsynopsisandsystematicmeta-analysesofgeneticassociationstudiesincutaneousmelanoma.JNatlCancerInst2011;103:1127–35.