phytoremediation of metals using transgenic...
TRANSCRIPT

439
Critical Reviews in Plant Sciences, 21(5):439–456 (2002)
0735-2689/02/$.50© 2002 by CRC Press LLC
Phytoremediation of Metals Using TransgenicPlants
Elizabeth Pilon-Smits and Marinus PilonColorado State University, Biology Department, A/Z Building, Fort Collins, CO 80523 [email protected]: 970-491-0649
Referee: Dr. Clayton Rugh, Michigan State University, Dept. of Crop and Soil Sciences, 516 Plant and Soil Science Bldg. East
Lansing, MI 48824
ABSTRACT: An ideal plant for environmental cleanup can be envisioned as one with high biomass production,combined with superior capacity for pollutant tolerance, accumulation, and/or degradation, depending on the typeof pollutant and the phytoremediation technology of choice. With the use of genetic engineering, it is feasible tomanipulate a plant’s capacity to tolerate, accumulate, and/or metabolize pollutants, and thus to create the idealplant for environmental cleanup. In this review, we focus on the design and creation of transgenic plants forphytoremediation of metals.
Plant properties important for metal phytoremediation are metal tolerance and accumulation, which aredetermined by metal uptake, root-shoot translocation, intracellular sequestration, chemical modification, andgeneral stress resistance.
If we know which molecular mechanisms are involved in these tolerance and accumulation processes, andwhich genes control these mechanisms, we can manipulate them to our advantage. This review aims to give asuccinct overview of plant metal tolerance and accumulation mechanisms, and to identify possible strategies forgenetic engineering of plants for metal phytoremediation. An overview is presented of what has been achieved sofar regarding the manipulation of plant metal metabolism. In fact, both enhanced metal tolerance and accumulationhave been achieved by overproducing metal chelating molecules (citrate, phytochelatins, metallothioneins,phytosiderophores, ferritin) or by the overexpression of metal transporter proteins. Mercury volatilization andtolerance was achieved by introduction of a bacterial pathway. The typical increase in metal accumulation as theresult of these genetic engineering approaches is 2- to 3-fold more metal per plant, which could potentially enhancephytoremediation efficiency by the same factor. As for the applicability of these transgenics for environmentalcleanup, results from lab and greenhouse studies look promising for several of these transgenics, but field studieswill be the ultimate test to establish their phytoremediation potential, their competitiveness, and risks associatedwith their use.
KEY WORDS: environmental cleanup, genetic engineering, biotechnology, cadmium, copper, iron, zinc, nickel.
TABLE OF CONTENTS
I. Introduction .................................................................................................................. 440A. The Problem with Metals .................................................................................... 440B. Phytoremediation Approaches for Metals .......................................................... 440C. How to Make Metal Phytoremediation More Efficient? ................................... 441
II. Genetic Engineering of Plants — What and How ...................................................... 441III. Genetic Engineering of Plants for Metal Phytoremediation: Species to Use,
Processes to Target ....................................................................................................... 442

440
IV. Molecular Mechanisms of Metal Accumulation and Tolerance ................................. 443V. Genetic Manipulation of Metal Accumulation and Tolerance — State of the
Science .......................................................................................................................... 447A. Metal-Binding Molecules: Metallothioneins, Phytochelatins, Organic
Acids, Phytosiderophores, Ferritin ...................................................................... 447B. Membrane Transporters ...................................................................................... 448C. Metal Metabolism ............................................................................................... 449D. General Stress-Resistance Mechanisms .............................................................. 449
VI. Putting Transgenics to the Test — Phytoremediation Case Studies .......................... 450VII. Risk Assessment Considerations ................................................................................. 450VIII. Perspectives ................................................................................................................... 451
I. INTRODUCTION
A. The Problem with Metals
Metals are present naturally in the Earth’s crustat various levels (Angelone and Bini, 1992). Min-ing, industry, and agriculture lead to acceleratedrelease of metals into ecosystems, causing seriousenvironmental problems and posing a threat to hu-man health (Lantzy and Mackenzie, 1979; Nriagu,1979; Ross, 1994). Although many metals are es-sential for cells (e.g. Cu, Fe, Mn, Ni, Zn), all metalsare toxic at higher concentrations (Marschner, 1995).One reason metals may become toxic is becausethey may cause oxidative stress. Especially redoxactive transition metals, which can take up or giveoff an electron (e.g. Fe2+/3+, Cu+/2+) can give rise tofree radicals that cause damage (Jones et al., 1991;Li and Trush, 1993), but other metals can causeoxidative stress as well (Weckx and Clijsters, 1997;Baccouch et al., 1998; Cho and Park, 2000). An-other reason why metals may be toxic is becausethey can replace other essential metals in pigmentsor enzymes, disrupting the function of these mol-ecules (Rivetta et al., 1997; van Assche and Clijsters,1986). Some metal ions (e.g. Hg+ and Cu+) are veryreactive to thiol groups and can interfere with pro-tein structure and function. When released into theenvironment, some metals occur as free cations(e.g. Zn2+), while others form cations that are boundto organics (e.g. Cu2+) and yet others form oxyanions(e.g. CrO4
3–, MoO42–, WO4
3–). Some metals occur inthe environment as radioactive isotopes, posing anadditional health risk (e.g. 238U, 137Cs, 239Pt, 90Sr).
More than 50,000 metal-contaminated sitesawait remediation in the U.S. alone (Ensley,2000). Approximately 80% of U.S. Superfundsites (designated by the U.S. EnvironmentalProtection Agency as priority sites for cleanup)contain heavy metals, often mixed with organicpollutants (Ensley, 2000). Conventionalremediation methods for metal-contaminatedsubstrates include soil washing, excavation, andreburial of soil, and pump and treat systems forwater (Glass, 1999). The present costs con-cerned with U.S. remediation are $7 to 8 billionper year, of which ~35% involves metalsremediation (Glass, 1999, 2000). The use ofplants for remediation of metals offers an at-tractive alternative, because it is solar drivenand can be carried out in situ, minimizing costand human exposure (Salt et al., 1995b, 1998).Phytoremediation is also aesthetically pleasingand can be used in conjunction with othercleanup methods. The U.S. phytoremediationmarket for metals is expected to grow from~$20 million in 2000 to ~$150 million in 2005(Glass, 1999, 2000).
B. Phytoremediation Approaches forMetals
Unlike organic contaminants, metals cannotbe degraded. Instead, phytoremediation strategiesfor metals are based on stabilization, accumula-tion, and in some cases volatilization (EPA, 1998).Phytostabilization of metals may simply involve

441
the prevention of leaching through the upwardwater flow created by plant transpiration, reducedrunoff due to above-ground vegetation, and re-duced soil erosion via stabilization of soil byplant roots (Berti and Cunningham, 2000). In somecases of phytostabilization, metals may be trans-formed to less bioavailable and therefore less toxicforms. For instance, many wetland plants reducemetals to insoluble precipitates on their root sur-face (Horne, 2000). Thus, in phytostabilization,mobilization of metals is prevented; although metalconcentrations are not reduced, the metal becomesless of a risk to the environment.
Accumulation of metals by roots in a hy-droponic setup, followed by harvesting of theplant biomass is termed rhizofil tration(Dushenkov and Kapulnik, 2000). The accu-mulation of metals in shoot tissue, followedby harvesting of shoot biomass, is calledphytoextraction (Blaylock and Huang, 2000).After harvesting the root and/or shoot biom-ass, the plant material may be ashed, followedby recycling of the metals if economically fea-sible (Chaney et al., 2000), or the disposal ofthe ashes in a landfill. Alternatively, the plantmaterial may be used for non-food purposes,for example, cardboard or wood products.
Certain metal(loid)s can be converted by plantsinto a gaseous form and emitted into the atmosphere(Hansen et al., 1998). The use of plants for volatil-ization of contaminants is called phytovolatilization.
Each of these metal phytoremediation tech-nologies has already been shown to be effective.To give a few examples: a rhizofiltration systemuses sunflowers to remove radioactive U fromcontaminated wastewater to levels below regula-tory limits (95% removal in 24 h, Dushenkov andKapulnik, 2000); chelator-assisted phytoextractionusing Brassica juncea lowered soil Pb levels from2055 mg.kg–1 to 960 mg.kg–1 in three crops(Blaylock, 2000); constructed wetlands routinelyremove over 90% of metals from various waste-water streams (Horne, 2000).
Phytoextraction, phytovolatilization, phytosta-bilization, and rhizofiltration are not exclusive tech-nologies. For instance, in a constructed wetland,phytoextraction, phytostabilization, and phyto-vola-tilization may be used simultaneously (Hansen etal., 1998).
C. How to Make Metal PhytoremediationMore Efficient?
Several approaches may be used to furtherenhance the efficiency of metal phytoremediation.First, a screening study may be performed to iden-tify the most suitable plant species or varieties forremediation of a certain metal. Second, agronomicpractices may be optimized for a selected speciesto maximize biomass production and metal uptake.For instance, planting density and fertilization canbe optimized to enhance plant productivity (Chaneyet al., 2000), and amendments such as organicacids or synthetic chelators may be added to thesoil to enhance metal uptake (Salt et al., 1998;Blaylock and Huang, 2000). Different plant spe-cies may also be combined, spatially or succes-sively, for maximal phytoremediation efficiency(Horne, 2000). Agronomic practices such fertiliza-tion and plant clipping may also affect plant metaluptake by influencing microbial density and com-position in the root zone.
The selected species or variety can be bredfurther for the desired property, either throughclassic breeding or via genetic engineering. Theadvantage of genetic engineering is that it can bemuch faster than conventional breeding, and it ispossible to introduce genes from other species.Thus, it is possible to introduce properties intoplants that could not be introduced via conven-tional breeding. This review focuses on the sec-ond approach: the use of genetic engineering toenhance plants’ phytoremediation potential formetals.
II. GENETIC ENGINEERING OF PLANTS— WHAT AND HOW
In genetic engineering of plants, a foreignpiece of DNA is stably inserted into the genomeof a cell, which is regenerated into a maturetransgenic plant. The piece of DNA can comefrom any organism, from bacteria to mammals.The foreign DNA usually contains two genes, onea resistance gene used for selection after transfor-mation, the other the gene of interest. Each geneis coupled to a plant promoter, ensuring the for-mation of the gene product (usually a protein) in

442
the plant. When the transformed plant is propa-gated, the foreign gene is inherited by its off-spring.
The foreign stretch of DNA may be transferredto the plant either via a particle gun, for which theDNA is coated onto metal particles and shot intothe plant tissue, or via Agrobacterium, a soil bac-terium that makes a living by inserting part of itsDNA (called T-DNA) into a plant cell and feedingoff of the gene products produced by the plant. TheAgrobacterium T-DNA genes can be replaced bygenes of interest, which are then inserted into theplant by Agrobacterium infection. (Hoekema etal., 1983). For some plant species (e.g. Arabidopsisthaliana, the model plant for plant molecular biol-ogy), transformation simply involves dipping theflowers in an Agrobacterium suspension: some ofthe resulting seeds will be transgenic (Bechtold etal., 1998). Most plants need to be grown as undif-ferentiated callus tissue culture in order to be trans-formed. After the transformation, mature plants areregenerated from the tissue culture using shoot-inducing plant hormones (Horsch et al., 1985).
The gene product can be targeted to certaincellular compartments (e.g. chloroplast, vacuole,mitochondrion, or apoplast) by adding specifictargeting information in the gene construct. Of-ten, constitutive promoters such as the 35S-cauli-flower mosaic virus promoter are used, that directexpression in all tissues and at all times. How-ever, the expression pattern of the gene may alsobe programmed to be only in certain tissue types(e.g. roots, vascular tissue, shoot), or under cer-tain environmental conditions (stress-induced,light-induced), by means of different promoters(Kasuga et al., 1999; Su et al., 1998). Besidesoverexpressing a gene, it is also possible to re-press the expression of an endogenous gene, byinserting a copy of that gene in reverse orientation(antisense technology).
III. GENETIC ENGINEERING OF PLANTSFOR METAL PHYTOREMEDIATION
A. Species to Use
The ideal plant species to engineer forphytoremediation purposes is one with a high bio-
mass production, sufficiently hardy and competi-tive in the climate where it is to be used, and witha good phytoremediation capacity to start with.Ideally, there should be an existing transformationprotocol for the species, because the developmentof such a protocol may take a year or more. Someexamples of suitable species are Indian mustard(Brassica juncea), poplar (Populus spp.), yellowpoplar (Liriodendron tulipifera), and cordgrass(Spartina spp.). In addition to, or prior to, trans-forming these large biomass phytoremediation spe-cies, the same gene construct may be transformedto the model plant species, A. thaliana. This smallplant with its short generation time and high seedproduction is very suitable to test in a short timewhether a biotechnological approach works.
B. Processes to Target
To breed plants with superior phytoremediationpotential, one possible strategy is to enhance thebiomass productivity of species that are good accu-mulators; another strategy is to enhance metal tol-erance and/or accumulation in high biomass spe-cies. General plant productivity is controlled bymany genes and difficult to promote by single geneinsertion. Therefore, this is difficult to achieve inthe short term by means of genetic engineering. Toenhance plant metal tolerance and/or accumula-tion, an existing plant process that is limiting forremediation may be accelerated, for example, ageneral plant mechanism involved in metal accu-mulation or tolerance; alternatively, a new path-way may be introduced into the plant from anyother organism, for example, a bacterial pathwayfor metal detoxification.
To accelerate existing plant processes thatlimit phytoremediation of an element (e.g. rootuptake, root-shoot translocation, sequestration inspecific tissues or cell compartments, biotransfor-mation, plant-microbe interactions), we need toknow which pathways and genes are involved.The overexpression of a gene encoding a rate-limiting gene product is expected to lead to afaster overall rate of the pathway and thus to moreefficient phytoremediation. Conversely, antisenserepression of a rate-limiting gene product willlead to a slower flux through a pathway. Table 1

443
gives an overview of the factors that are impor-tant and may be limiting for different metalphytoremediation applications.
Molecules involved in the processes listedabove include metal transporter membrane pro-teins, metal chelating molecules of various sorts,metal-modifying enzymes, enzymes involved inrepair of metal damage (e.g. oxidative stress), andregulatory proteins. All of these molecules areproduced as the result of the presence and expres-sion of genes. Any (combination) of these genesmay be suitable targets for plant genetic engineer-ing for phytoremediation. In the next section, ashort overview is given of molecules and genesinvolved in metal uptake, translocation, seques-tration, and tolerance.
IV. MOLECULAR MECHANISMS OFMETAL ACCUMULATION ANDTOLERANCE
A. Uptake
For metal ions to be taken up by plants, theyhave to be bioavailable. Roots compete with soilparticle cation/anion exchange sites for ions (Fig-ure 1). Especially in soils with high clay andorganic matter content metal bioavailability islow (Ross, 1994; Marschner, 1995). Plants haveevolved mechanisms to make micronutrient met-als more bioavailable. For instance, many plantsexcrete organic acids (e.g. malate, citrate) that actas metal chelators and decrease the rhizospherepH, making metal cations more bioavailable (Ross,1994). Organic acids have been reported to facili-tate metal uptake, for example, citrate enhances Uuptake in Brassica (Huang et al., 1998). On theother hand, organic acids can also inhibit metaluptake by forming a complex with it outside theroot that is not taken up, for example, citrateinhibits Al uptake in various species (de la Fuenteet al., 1997; Moffat, 1999; Pineros and Kochian,2001; Papernik et al., 2001); a similar mechanismappears to be responsible for Cu tolerance inArabidopsis (Murphy et al., 1999).
Plants can also affect their rhizosphere pH viaproton pumps in the root cell membrane. Forinstance, one proton pump, an H+-ATPase en-
coded by gene AHA2, is upregulated under irondeficiency, leading to enhanced proton efflux fromthe root (Fox and Guerinot, 1998). Also, anArabidopsis mutant was found to have an in-creased rhizosphere pH, caused by enhanced pro-ton influx (Moffat, 1999). This resulted in theprecipitation of Al hydroxides, and therefore re-duced Al uptake and higher Al tolerance.
Another type of exudate produced by grassesare phytosiderophores, which bind Fe and facilitateits uptake. Phytosiderophores are biosynthesizedfrom nicotianamine, which is composed of threemethionines coupled via non-peptide bonds (Higuchiet al., 1994). Rhizosphere microbes can also affectplant uptake of metals: bacteria have been reportedto enhance uptake of Se and Hg (de Souza et al.,1999), and mycorrhizae were reported to reducemetal uptake, leading to enhanced tolerance (Frey etal., 2000; Rufyikiri et al., 2000). Likely, certainplant genes are involved in these plant-microbe in-teractions, for example, in production of signalmolecules.
The uptake of metals requires transport acrossthe root cell membrane into the symplast. Thisprocess involves specific membrane transporterproteins. Membrane transport of cations has beenthe subject of several recent reviews (Fox andGuerinot, 1988, Williams et al., 2000; Mäser etal., 2001; Axelsen and Palmgren 2001). The ge-nome of the model species A. thaliana encodesfor over 150 different cation transporters in atleast nine different families. The abundance ofgenes implied in metal transport in A. thaliana aswell as all other organisms underscores the needfor metal homeostasis in which organisms mustmaintain a fine balance between having enoughessential metals available for metabolic functionsand at the same time avoiding deficiency or tox-icity. Membranes serve to separate compartmentsin which metal concentrations can be regulatedwith the aid of transporters (Nelson, 1999). Theuptake of metal ions into the cell can be driven bythe electrochemical gradient (proton gradient)across the plasma membrane, but the energeticsof transport are not yet fully understood in allcases (Mäser et al., 2001). Often more than onetransport system exists for one metal. For ex-ample, A. thaliana has several transporters of theNRAMP family capable of transporting iron into
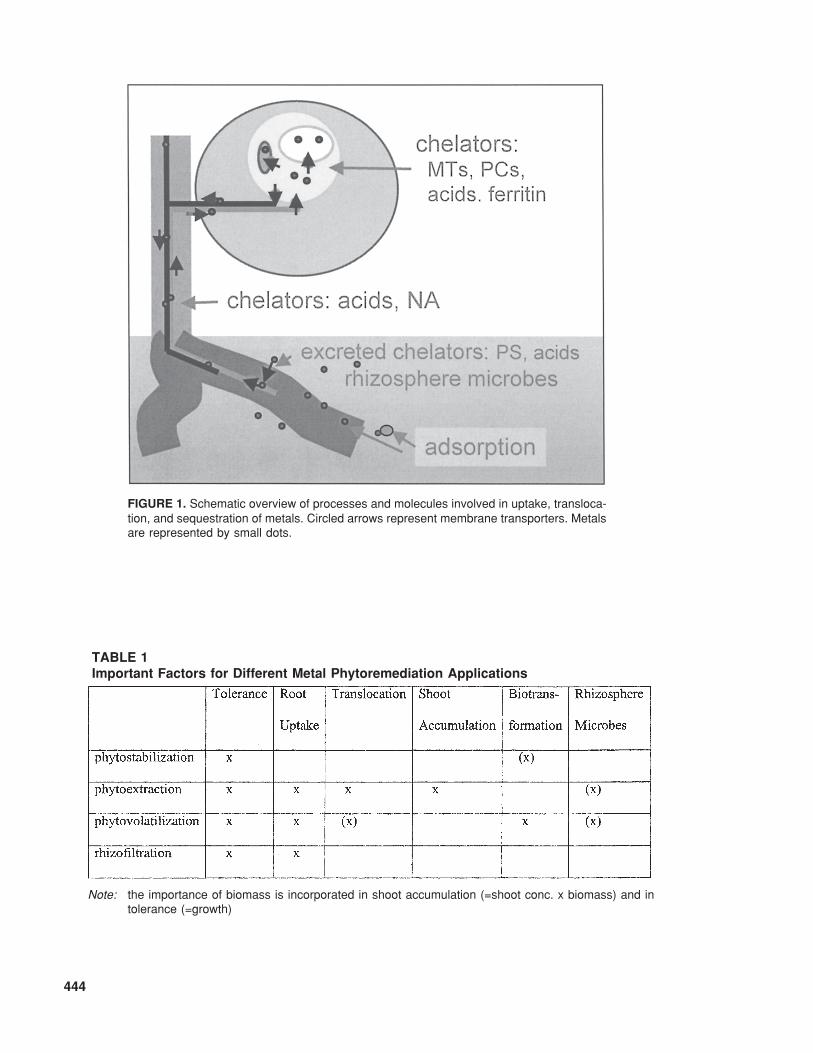
444
TABLE 1Important Factors for Different Metal Phytoremediation Applications
FIGURE 1. Schematic overview of processes and molecules involved in uptake, transloca-tion, and sequestration of metals. Circled arrows represent membrane transporters. Metalsare represented by small dots.
Note: the importance of biomass is incorporated in shoot accumulation (=shoot conc. x biomass) and intolerance (=growth)

445
cells; in addition, the ZIP family member IRT andperhaps several members of the 8-member YSLfamily are also implied in iron uptake into cells(Mäser et al., 2001; Curie et al., 2001). The pres-ence of several transporters permits having up-take systems with different affinities and capaci-ties. In addition, transporters are present in internalmembranes to allow to regulate the storage ofmetals in organelles such as vacuoles. Transport-ers may be specific for a certain cell type. Ofinterest to phytoremediation is the observationthat some metal transporters can transport morethan one metal ion. For instance, the IRT proteinmediates iron uptake in roots but also transportsCd.
B. Translocation
For root-shoot translocation of metals, metaltransporters export metal ions out of the root sym-plast into the xylem apoplast (Marschner, 1995).Translocation in the xylem is probably transpira-tion driven (Salt et al., 1995a). Different chelatorsmay be involved in translocation of metal cationsthrough the xylem (Figure 1), such as organicacid chelators (e.g. malate, citrate, histidine, Saltet al., 1995a; von Wiren et al., 1999), ornicotianamine (Stephan et al., 1996; von Wiren etal., 1999). Uptake of metal ions from the xylemapoplast into the shoot symplast is mediated bymetal transporters in the shoot cell membrane.
For translocation of metals in the phloem,nicotianamine may function as a chelator (vonWiren et al., 1999).
C. Sequestration
Once inside the shoot cells, essential metals willbe translocated to their final destination, which mayinvolve membrane metal transporters, and metal-bind-ing proteins. One class of metal chelating moleculesthat may play a role in sequestration — they areupregulated under conditions of high metal availabil-ity— are the metallothioneins (MTs). Metallothioneinsare small (~7 kDa) cysteine-rich metal-binding pro-teins that occur in all organisms. Although the exactrole of MTs is still not clear, they likely play a role in
homeostasis of essential metals and perhaps also intolerance to nonessential metals (Goldsbrough, 2000;Cobbett and Goldsbrough, 2000). Metal chaperonesare a different class of proteins that bring metals tospecific targets in the cell. An example is the ATXprotein, which is upregulated under Cu deficiency(Himelblau et al., 1998).
Toxic levels of essential or nonessential met-als are stored in a location where the metal can dothe least harm to vital cellular processes. Thismay involve storage in special cellular compart-ments such as the vacuole by means of special-ized transporters such as ZAT1, a CDF-type trans-porter for zinc (van der Zaal et al., 1999).Sequestration may also be in the apoplast, or inspecialized cell types, such as epidermal cells andtrichomes (Heath et al., 1997; Küpper et al., 1999;Salt and Krämer, 2000; Hale et al., 2001).
For storage in the vacuole, certain metals may becomplexed by phytochelatins (PCs). Phytochelatinsare small cysteine-rich metal-binding peptides (5 to23 amino acids) that occur in all plants tested so far(Rauser, 1995; Zenk, 1996; Cobbett, 2000), as well asin some fungi and animals (Vatamaniuk et al., 2001).Phytochelatins are induced only under metal stressand are thought to mainly function in tolerance totoxic metals (Goldsbrough, 2000; Cobbett andGoldsbrough, 2000). They are synthesized enzymati-cally from glutathione. Complexes of metals boundby glutathione or phytochelatins are shuttled to thevacuole by an ABC-type transporter protein in thetonoplast (Lu et al., 1997). The same type of trans-porter is involved in shuttling glutathione-conjugatedanthocyanins to the vacuole (Marrs, 1996). In fact,anthocyanins can also bind metals (Kondo et al.,1992; Everest and Hall, 1921; Takeda et al., 1985),and recently have been suggested to play a role inmetal sequestration (Hale et al., 2001). Other metal-binding molecules that are involved in metal com-plexation in the vacuole are organic acids (Krämer etal., 2000). Excess iron, in contrast to other metals, isstored in chloroplasts, bound to the protein ferritin(Theil, 1987).
D. Chemical Modification
Metal-modifying enzymes may be involvedin assimilation of metals into organic molecules

446
(e.g. selenate is metabolized to dimethylselenide;de Souza et al., 2000), or in changing the oxida-tion state of metals (e.g. toxic Cr(VI) is reducedto nontoxic Cr(III), Lytle et al., 1998, and indicots Fe (and possibly also Cu) is reduced by areductase at the root cell membrane before up-take; Robinson et al., 1999).
E. Hyperaccumulation Mechanisms
Metal hyperaccumulators are commonly definedas plant species that accumulate ~100-fold highermetal levels than nonaccumulator species (Brooks,1998), for example, 1% of DW for Mn and Zn, 0.1%of DW for Cu and Ni, and 0.01% of DW for Cd(Baker et al., 2000). Metal hyperaccumulation hasbeen found in over 500 species from over 75 fami-lies (Baker et al., 2000) and has probably evolvedindependently in these different taxa (Wu, 1990;Pollard et al., 2000). The evolutionary selectionpressure for hyperaccumulation may be protectionagainst herbivory (Pollard and Baker, 1997) andpathogens (Boyd et al., 1994). Hyperaccumulatorsare usually slow growing, low biomass species. Theyhyperaccumulate metals even from low externalmetal concentrations, and most of the metal is trans-located to the shoot (Salt and Krämer, 2000). For acomprehensive review of hyperaccumulation mecha-nisms see Salt and Krämer (2000). Following is asummary of findings to date.
At the root membrane level, metal uptake isunusually high in hyperaccumulators. This maybe due to constitutive high expression of a metaltransporter in the plasma membrane, as was foundfor the Zn and Cd hyperaccumulator Thlaspicaerulescens (Pence et al., 2000; Lombi et al.,2001). The uptake of metals in hyperaccumulatorsmay be enhanced further by the excretion ofmetal chelators like histidine (Krämer et al.,1996), and/or by rhizosphere microbes capableof mobilizing nonlabile soil metals (Whiting etal., 2001). Thlaspi caerulescens shows reducedmetal accumulation in root vacuoles, enhancedroot-shoot translocation, enhanced uptake intoleaf cells, and higher metal tolerance (Lasat etal., 2000). The high metal tolerance may inpart be due to highly efficient intracellular com-partmentalization. The Ni hyperaccumulator
T. goesingense was shown recently to have avacuolar metal transporter, TgMTP1, which isconstitutively highly expressed compared toorthologues in nonaccumulators (Persans et al.,2001). Efficient chelation is likely to be anotherkey factor for metal tolerance and accumulationin hyperaccumulators. Different chelator mol-ecules may function in different plant parts. Inthe Ni hyperaccumulator Alyssum lesbiacum,Ni chelation by histidine appears to play an im-portant role in Ni translocation in the xylem(Krämer et al., 1996). In the Ni hyperaccumulatorT. goesingense, histidine does not seem to be akey compound for Ni accumulation (Persans etal., 1999); Ni appears to be predominantly local-ized in the vacuole as a Ni-organic acid complexin this species (Krämer et al., 2000). In the Znhyperaccumulator T. caerulescens, zinc chela-tion appears to be mainly by histidine in roots,by organic acids — or no chelator — in thexylem, and by organic acids in shoots (Salt etal., 1999).
F. General Stress ResistanceMechanisms
One reason why metals are toxic is becausethey cause oxidative stress. Metal stress activatesantioxidative systems, composed of free radicalscavenging molecules such as ascorbate and glu-tathione, and the enzymes involved in their biosyn-thesis and reduction (Noctor and Foyer, 1998).Other molecules involved in preventing oxidativestress are the superoxide dismutase enzymes, whichthemselves require either Cu/Zn, Mn, or Fe ascofactors (Bowler et al., 1994). The overproduc-tion of any of these components may lead to highermetal stress tolerance. Alternatively, if regulatorygenes can be identified that orchestrate the activa-tion of many metal-induced genes, overexpressionof such a regulatory gene may be the most efficientway to enhance metal tolerance. Recently, an iron-dependent cis-regulatory element was identified inmaize that mediates repression of ferritin genesunder low iron conditions (Petit et al., 2001). Also,transcription factors that mediate salt, drought, andfreezing tolerance have been identified (Su et al.,1998; Kasuga et al., 1999).

447
V. GENETIC MANIPULATION OF METALACCUMULATION AND TOLERANCE –STATE OF THE SCIENCE
As is clear from the above, many genes areinvolved in metal uptake, translocation, seques-tration, chemical modification, and tolerance. Theoverexpression of any (combination) of thesegenes is a possible strategy for genetic engineer-ing. Depending on which phytoremediation ap-plication is to be used, the genetic engineeringstrategy may strive to create plants that accumu-late more metals in harvestable plant parts(phytoextraction), or adsorb more metals at theirroot surface (rhizofiltration, phytostabilization).A plant property essential for all phytoremediationapplications is plant tolerance (Table 1), so en-hancing plant metal tolerance is an obvious av-enue for genetic engineering approaches. En-hanced tolerance to metals may be achieved byreducing metal uptake, by more efficient seques-tration of metals in plant storage compartments,overproduction of metal chelating molecules, orincreasing activity of enzymes involved in gen-eral (oxidative) stress resistance.
The overexpression of metal transporter genesmay lead to enhanced metal uptake, translocation,and/or sequestration, depending on the tissueswhere the gene is expressed (root, shoot, vasculartissue, or all), and on the intracellular targeting(e.g. cell membrane, vacuolar membrane). Theoverexpression of genes involved in synthesis ofmetal chelators may lead to enhanced or reducedmetal uptake, as well as enhanced metal translo-cation and/or sequestration, depending on the typeof chelator and its location.
Unless regulatory genes are identified thatsimultaneously induce many metal-related genes,it is feasible that more than one gene will need tobe upregulated in order to substantially enhancemetal phytoremediation capacity. To our knowl-edge, this has not been done so far (in one trans-formation event) in the context of manipulatingplant metal metabolism. Encouraging fortransgenic approaches, classic genetic studies indi-cate that there are usually very few genes (1 to 3)responsible for metal tolerance (MacNair et al.,2000). Also, metal accumulation, tolerance, andplant productivity are not necessarily correlated
(Wu, 1990; MacNair et al., 2000). Therefore, itshould be possible to breed or genetically engi-neer a plant with high metal tolerance and metalaccumulation as well as high productivity. Thiswould be the ideal plant for metal phytoextraction.
In the next section we give an overview ofwhat has been achieved so far with respect togenetic manipulation of plant metal tolerance andaccumulation.
A. Metal-Binding Molecules:Metallothioneins, Phytochelatins,Organic Acids, Phytosiderophores,Ferritin
Overproduction of various metal chelator mol-ecules has been shown to affect plant metal toler-ance and accumulation. Several research groups haveoverexpressed the metal-chelating proteinsmetallothioneins (MTs). The expression of the hu-man MT2 gene in tobacco or oil seed rape resultedin higher Cd tolerance at the seedling level (Misraand Gedamu, 1989). Similarly, the expression of themouse MT1 gene in tobacco led to enhanced Cdtolerance at the seedling level (Pan et al., 1994). Theoverexpression of a pea MT in A. thaliana resultedin a severalfold higher Cu accumulation (Evans etal., 1992). The most pronounced effect of MToverexpression was observed by Hasegawa et al.(1997), who overexpressed the yeast gene CUP1 incauliflower, leading to a 16-fold higher Cd toler-ance, as well as higher Cd accumulation. Thus, itappears that the overexpression of MTs is a prom-ising approach to enhance Cd/Cu tolerance and ac-cumulation.
In a different approach to enhance metaltolerance and accumulation, the metal-bindingpeptides phytochelatins (PCs) were overproducedvia expression of enzymes involved in their bio-synthesis. Transgenic mustard (Brassica juncea)plants with higher levels of glutathione andphytochelatins were created through theoverexpression of either of two glutathione syn-thesizing enzymes,— γ-glutamylcysteine syn-thetase (γECS) or glutathione synthetase (GS).Both types of transgenics showed enhanced Cdtolerance and accumulation (Zhu et al., 1999a,b),illustrating the importance of these metal-bind-

448
ing peptides for metal tolerance and accumula-tion. In a related study, γECS was overex-pressed or knocked out (antisense approach) inArabidopsis, leading to increased or decreasedGSH levels (Xiang et al., 2001). Transgenicswith decreased GSH levels showed reduced Cdtolerance, confirming the importance of GSHand PCs for Cd tolerance. However, plants withincreased GSH levels did not show enhanced Cdtolerance, suggesting that GSH production is notlimiting for PC production and Cd tolerance inthis species. Harada et al. (2001) also createdtransgenic plants with enhanced phytochelatinlevels, through overexpression of cysteine syn-thase. The resulting transgenics displayed en-hanced Cd tolerance but lower Cd concentra-tions.
To our knowledge, there are no publishedrecords concerned with the overexpression of ei-ther phytochelatin synthase or a tonoplast PC-metal transporter. However, the overexpressionof a tobacco glutathione-S-transferase gene (parB)in Arabidopsis was reported to lead to enhancedCu, Al, and Na tolerance (Ezaki et al., 2000).Glutathione-S-transferases mediate glutathioneconjugation, followed by transport of the result-ing complex to the vacuole (Marrs, 1996).
Enhanced production of the metal chelatorcitric acid was achieved by the overexpression ofcitrate synthase (de la Fuente et al., 1997). Theresulting CS transgenics were shown to have en-hanced Al tolerance, apparently via extracellularcomplexation of Al by citrate after excretion fromroot cells. The same CS transgenics take up morephosphorus (Lopez-Bucio et al., 2000) and aremore resistant to iron deficiency (Guerinot, 2001),illustrating that citrate excretion can affect theuptake of different elements in different ways. Ascitrate amendment has been shown to enhance Uuptake (Huang et al., 1998), it would be interest-ing to test these CS transgenics for U uptake.
The overproduction of the iron-chelatordeoxymugineic acid (phytosiderophores) wasachieved through the overexpression ofnicotianamine aminotransferase (NAAT) in rice(Takahashi et al., 2001). The resulting plantsreleased more phytosiderophores and grew bet-ter on iron-deficient soils. Iron levels in theplants were not determined.
The overexpression of the iron-binding pro-tein ferritin was shown to lead to a 1.3-fold higheriron level in tobacco leaves (Goto et al., 1998)and a three-fold higher level in rice seeds (Goto etal., 1999).
B. Membrane Transporters
The genetic manipulation of several metaltransporters has been shown to result in alteredmetal tolerance and/or accumulation. Theoverexpression of the Zn transporter ZAT (alsoknown as AtMTP1) in A. thaliana gave rise toplants with enhanced Zn resistance and two-foldhigher root Zn accumulation (van der Zaal et al.,1999). ZAT is a putative vacuolar transporter andof the same gene family as the TgMTP1 isolatedfrom the hyperaccumulator T. goesingense(Persans et al., 2001).
The overexpression of the calcium vacuolartransporter CAX2 from A. thaliana in tobaccoresulted in enhanced accumulation of Ca, Cd, andMn, and to higher Mn tolerance (Hirschi et al.,2000). Another vacuolar transporter, AtMHX, wasoverexpressed in tobacco (Shaul et al., 1999).The resulting plants showed reduced tolerance toMg and Zn, but it did not show altered accumu-lation of these elements.
Another putative metal transporter gene fromtobacco (NtCBP4), encoding a calmodulin-bind-ing protein, when overexpressed resulted in en-hanced Ni tolerance and reduced Ni accumula-tion, as well as reduced Pb tolerance and enhancedPb accumulation (Arazi et al., 1999). When atruncated form of the protein was overexpressed,however, from which the calmodulin-binding partwas removed, the resulting transgenics showedenhanced Pb tolerance and attenuated accumula-tion (Sunkar et al., 2000).
In order to enhance iron uptake by plants, twoyeast genes encoding ferric reductase (FRE1 andFRE2, involved in iron uptake) were overexpressedin tobacco (Samuelsen et al., 1998). Iron content inthe shoot of the transgenics was 1.5-fold highercompared with wild-type plants. Earlier, enhancedaccumulation of various metals (Fe, Cu, Mn, Zn,Mg) was already observed in an Arabidopsis mu-tant with enhanced ferric-chelate reductase activity

449
(Delhaize, 1996). The affected gene in theArabidopsis mutant meanwhile has been identifiedas FRO2 and isolated (Robinson et al., 1999); itwill be interesting to see what effect itsoverexpression has on plant metal uptake.
The overexpression of another metal trans-porter, AtNramp1, resulted in an increase in Fetolerance (Curie et al., 2000), while theoverexpression of AtNramp3 led to reduced Cdtolerance but no difference in Cd accumulation(Thomine et al., 2000).
In addition to overexpressing metal transport-ers, it is also possible to alter their metal specific-ity. For instance, while IRT1, the Arabidopsisiron transporter, can transport Fe, Zn, Mn, andCd, the substitution of one amino acid was shownto result in loss of either Fe and Mn transportcapacity, or Zn transport capacity (Rogers et al.,2000). With the overexpression of such engineeredtransporters, it may be possible to tailor transgenicplants to accumulate specific metals.
C. Metal Metabolism
Rather than accelerating existing processes inplants, an alternative approach is to introduce anentirely new pathway from another organism. Thisapproach was taken by Richard Meagher and co-workers, who introduced two bacterial genes inplants that together convert methylmercury to vola-tile elemental mercury. MerB encodes organomer-curial lyase, which converts methylmercury to ionicmercury or Hg(II); MerA encodes mercuric reduc-tase, which reduces ionic mercury to elementalmercury or Hg(0) (Summers, 1986). TransgenicMerA A. thaliana plants showed significantly highertolerance to Hg(II) and volatilized elemental mer-cury (Rugh et al., 1996). Transgenic MerBA. thaliana plants were significantly more tolerantto methylmercury and other organomercurials(Bizily et al., 1999). The MerB plants were shownto convert methylmercury to ionic mercury, a formthat is ~100-fold less toxic to plants.
MerA-MerB double-transgenics, obtained bycrossing MerA and MerB transgenics, were com-pared with their MerA, MerB, and wild-type counter-parts with respect to tolerance to organic mercury(Bizily et al., 2000). While MerB plants were 10-foldmore tolerant to organic mercury than wild-type plants,
MerA-MerB plants were 50-fold more tolerant. Whensupplied with organic mercury, MerA-MerB doubletransgenics volatilized elemental mercury, whereassingle transgenics and wild-type plants did not; thus,MerA-MerB plants were able to convert organicmercury all the way to elemental mercury, which wasreleased in volatile form.
The same MerA/MerB gene constructs wereused to create mercury-volatilizing plants fromother species. Transgenic MerA and MerB to-bacco and yellow poplar also showed enhancedmercury tolerance (Rugh et al., 2000). In an ini-tial experiment to analyze the potential of theseplants for phytoremediation, MerA tobacco plantsremoved 3- to 4-fold more mercury from hydro-ponic medium than untransformed controls(Meagher et al., 2000).
To our knowledge, no reports have been pub-lished at this point involving the expression ofmetal hyperaccumulator genes in nonaccumulatorspecies. However, an alternative approach hasbeen used to transfer hyperaccumulation capacityto a nonaccumulator high biomass species. Breweret al. (1999) used somatic hybridization (proto-plast electrofusion) to create a hybrid betweenThlaspi caerulescens and Brassica napus. Someof the hybrids showed high biomass combinedwith high metal tolerance and accumulation, mak-ing them attractive for metal phytoextraction.
A different way of using genetic engineeringto study metal metabolism is by creating hairyroot cultures of plants using Agrobacteriumrhizogenes. The resulting fast growing root cul-ture can be grown in vitro indefinitely. Hairy rootculture of Thlaspi caerulescens was shown to bemore tolerant to Cd, and accumulated 1.5- to 1.7-fold more Cd than hairy roots of nonaccumulatorspecies (Nedelkoska and Doran, 2000).
Agrobacterium rhizogenes infection may alsobe used to bring about root proliferation, and thusto increase the root surface area of a plant. Theuse of such plants may be attractive forrhizofiltration applications.
D. General Oxidative Stress ResistanceMechanisms
Overexpression of enzymes involved in gen-eral stress resistance mechanisms present an alter-

450
native approach to bring about metal tolerance.Several studies using this approach have led topromising results. Ezaki et al. (2000) reported thatthe overexpression of several genes involved inoxidative stress response (glutathione-S-transferase,peroxidase) resulted in enhanced Al tolerance.Oberschall et al. (2000) overexpressed an aldose/aldehyde reductase responsible for detoxifying alipid peroxide degradation product and found thatthe transgenics were more metal tolerant. Theoverexpression of glutathione reductase resulted inreduced Cd accumulation and enhanced Cd toler-ance, as judged from chlorophyll content and chlo-rophyll fluorescence measurements (Pilon-Smitset al., 2000). Grichko et al. (2000) found that theoverexpression of 1-aminocyclopropane-1-carboxy-lic acid (ACC) deaminase led to an enhanced accu-mulation of a variety of metals, as well as highermetal tolerance. ACC is the precursor for ethylene,the plant hormone involved in senescence.
VI. PUTTING TRANSGENICS TO THETEST — PHYTOREMEDIATION CASESTUDIES
As listed above, several different strate-gies have been used successfully to createtransgenics that show promising propertiesfor phytoremediation, as judged from lab stud-ies involving hydroponic systems or agarmedia . The next s tep is to tes t thesetransgenics on more realistic contaminatedsubstrates, collected from the environment.So far, to our knowledge, no results fromsuch studies have been published. Recently,a greenhouse study was performed to testtransgenic phytochelatin-overproducing mus-tard plants (ECS/GS plants, see above) fortheir phytoextraction capacity, using metal-contaminated soil from Leadville, Colorado(Bennett et al., 2002). Both types of transgenicsaccumulated significantly higher levels of Cdand Zn in their shoots than untransformedplants. These results are encouraging becauseit suggests that metal accumulation resultsobtained from hydroponic studies are a valu-able indication of transgenics’ metal accu-mulation potential from environmental soilscontaining metal mixtures.
VII. RISK ASSESSMENTCONSIDERATIONS
Transgenic plants with altered metal toler-ance, accumulation, or transformation propertiesare valuable for various reasons. They shed newlight on basic biological mechanisms involved inthese processes: which pathways are involved andwhich enzymes are rate limiting. Plants with al-tered metal accumulation properties may also beapplicable, not only for phytoremediation but alsoto enhance crop productivity in areas with subop-timal soil metal levels, or as “fortified foods” forhumans or livestock (Guerinot and Salt, 2001).
When genetically engineered plants are usedfor any of these applications, a thorough riskassessment study should be performed in eachcase (Wolfenbarger and Phifer, 2000). Some ofthe possible risks involved are biological trans-formation of metals into forms that are morebioavailable, enhanced exposure of wildlife andhumans to metals (in the case of enhanced metalaccumulation in palatable plant parts, or volatil-ization), uncontrolled spread of the transgenicplants due to higher fitness (e.g. metal tolerance)or general weedy nature, and/or uncontrolledspread of the transgene by interbreeding withpopulations of wild relatives (for a comprehen-sive report on this topic, see Glass, 1997). Theserisks will have to be assessed on a case-by-casebasis and weighed against the benefits, and againstthe risks of doing nothing or using alternativetechnologies.
To our knowledge, no transgenics are used inthe field for phytoremediation at this point. There-fore, the actual risks involved with the use oftransgenic plants for phytoremediation have neverbeen tested. However, theoretical calculations ofrisks associated with the use of mercury-volatilizating plants have been done by Meagherand coworkers (Meagher et al., 2000; Rugh et al.,2000). According to their calculations, the mer-cury emitted by these plants would pose no sig-nificant threat to the environment and would benegligible compared with other sources of mer-cury, such as burning of fossil fuels and medicalwaste. Even if the level of volatile mercury at thephytoremediation site is 400-fold higher thanbackground levels, that would still be 25 timesbelow regulatory limits. In addition, the retention

451
time of elemental mercury in the atmosphere,before precipitation, is 1 to 2 years during whichthe mercury is diluted to nontoxic levels.
Norman Terry and coworkers have done asimilar theoretical analysis of the risk of volatileSe emitted by plants (see Berken et al., this issue,and Lin et al., 2000), and came to the conclusionthat the volatile Se will likely be beneficial ratherthan toxic, as it is likely to precipitate in Se-deficient areas.
Metal accumulation in plant shoots bringsalong the risk of wildlife ingestion, and any in-crease in metal accumulation via biotechnologywill lead to a proportional increase of this risk. Onthe other hand, if a site can be cleaned in a shortertime, the duration of exposure may be reducedwhen using transgenics. The risk of metal inges-tion by wildlife may be minimized by fencing offthe area, using deterrents such as periodic noise,and the use of less palatable plant species.
The risk of transgenic plants or their genes“escaping” is not considered a significant problemby Meagher et al. (2000), because they generallyoffer little or no advantage over untransformedplants, either in a pristine or a contaminated envi-ronment. However, before using specific transgenicsfor phytoremediation in the field, this could beverified by a greenhouse or pilot field experiment,analyzing transgenic gene frequency over a num-ber of generations, on polluted and uncontami-nated soil. To further minimize the risk of out-crossing to wild relatives, transgenic plant speciesmay be chosen that have no compatible wild rela-tives, male-sterile transgenics may be bred, and/orthe plants may be harvested before flowering.
Government regulation should not be a sig-nificant obstacle to the use of transgenic plants inphytoremediation in the U.S. (Glass, 1997). Per-mit applications should be filed to the U.S.D.A.approximately 4 months before the planned start-ing date, but in most jurisdictions approval forresearch field tests should be routine.
VIII. PERSPECTIVES
It has been shown in multiple studies thatplant trace element metabolism can be geneticallymanipulated, leading to plants with altered metal
tolerance, accumulation, and/or biotransformationcapacity. When natural plant processes were ac-celerated by genetic engineering, the typical in-crease in metal accumulation per plant was 2- to3-fold. This would potentially reduce the cost ofphytoremediation to the same extent, if the sameresults hold true in the field. Furthermore, theintroduction of a new pathway has led to plantsthat can detoxify Hg in ways that other plantscannot — this is potentially even more valuable.In the coming years some of these newly avail-able transgenics will likely be put to the test in amore realistic phytoremediation setting.
As more metal-related genes are discovered,facilitated by the genome sequencing projects,many new possibilities will open up for the cre-ation of new transgenics with favorable proper-ties for phytoremediation. In addition to constitu-tive overexpression of one gene, several genesmay be overexpressed simultaneously, and theoverexpression may be fine-tuned in specific tis-sues, under specific conditions, or in specific cel-lular compartments.
Some promising strategies may be (1) the manynewly discovered metal transporters, including theones from hyperaccumulator plants (ZNT1,TgMTP1), may be overexpressed in high biomassplant species, targeted to different tissues and intra-cellular locations; (2) nicotianamine overproduc-tion may be an interesting avenue to manipulatemetal translocation and tolerance, as well as ironuptake in cereals, NA being the precursor ofphytosiderophores. Overproduction of NA is fea-sible via overexpression of enzymes from the NAbiosynthesis pathway, the genes for which havebeen cloned (Herbik et al., 1999; Higuchi et al.,1999; Ling et al., 1999; Takahashi et al., 1999); (3)overexpression of phytochelatin synthase (PS), theenzyme mediating PC synthesis from GSH, mayfurther enhance metal tolerance and accumulation.The overexpression of PS is possible, because genesencoding PS have been cloned (Clemens et al.,1999; Ha et al., 1999; Vatamaniuk et al., 1999).The overexpression of the vacuolar transporter re-sponsible for shuttling the PC-metal complex intothe vacuole may also enhance metal tolerance andaccumulation; this too is possible because the A.thaliana gene encoding this transporter has beencloned (Lu et al., 1997); (4) overproduction of

452
histidine can be done because the genes involvedin His biosynthesis have been cloned (Persans etal., 1999). In fact, preliminary data suggest thathistidine overproducing plants have enhanced Nitolerance (Krämer and Chardonnens, 2001); (5) aresearch area that may render a wealth of newinformation in the coming years is molecular biol-ogy of the rhizosphere. Manipulation of the qualityand quantity of root-released compounds offer apromising alternative strategy to affect metal up-take or exclusion. Together, these new develop-ments likely will give rise to much new informa-tion about metal metabolism in plants in the nearfuture and may lead to the fruitful applications inenvironmental cleanup, nutrition, and crop produc-tivity.
ACKNOWLEDGMENTS
The authors’ work is supported by NationalScience Foundation Grant MCB9982432 (E.P.S.),Environmental Protection Agency Grant G8A11586(E.P.S.), and National Science Foundation GrantMCB0091163 (M.P.).
REFERENCES
Angelone, M. and Bini, C. 1992. Trace element concentra-tions in soils and plants of Western Europe. In: Bio-geochemistry of Trace Metals. pp. 19–60. Adriano,D.C. Ed., Lewis, Boca Raton, Florida.
Arazi, T., Sunkar, R., Kaplan, B., and Fromm, H. 1999. Atobacco plasma membrane calmodulin-binding trans-porter confers Ni2+ tolerance and Pb2+ hypersensi-tivity in transgenic plants. Plant J. 20: 171–182.
Axelsen, K.B., and Palmgren, M.G. 2001. Inventory of thesuperfamily of P-type ion pumps in Arabidopsis. PlantPhysiol. 126: 696–706.
Baccouch, S., Chaoui, A., and El Ferjani, E. 1998. Nickel-induced oxidative damage and antioxidant responsesin Zea mays shoots. Plant Physiol. Biochem. 36: 689–694.
Baker, A.J.M., McGrath, S.P., Reeves, R.D., and Smith,J.A.C. 2000. Metal hyperaccumulator plants: a reviewof the ecology and physiology of a biological resourcefor phytoremediation of metal-polluted soils. In:Phytoremediation of contaminated soil and water. pp.85–108. Terry, N., and Bañuelos, G., Eds., Lewis,Boca Raton, Florida.
Bechtold, N. and Pelletier, G. 1998. In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana
plants by vacuum infiltration. In: Arabidopsis proto-cols. pp. 259–266. Martinez-Zapater, J.M., and Sali-nas, J. Eds. Humana Press, Totowa, New Jersey.
Bennett, L.E., Burkhead, J.L., Hale, K.L., Terry, N., Pilon,M., and Pilon-Smis, E.A.H. 2002. Analysis oftransgenic indian mustard plants for phytoremediationof metal-contaminated mine tailings. J. Environ. Qual.,in press.
Berti, W.R. and Cunningham, S.D. 2000. Phytostabilizationof metals. In: Phytoremediation of Toxic Metals —Using Plants to Clean up the Environment. pp. 71–88.Raskin, I. and Ensley, B.D., Eds., Wiley, New York.
Bizily, S.P., Rugh, C.L., Summers, A.O., and Meagher, R.B.(1999) Phytoremediation of methylmercury pollution:merB expression in Arabidopsis thaliana confers re-sistance to organomercurials. Proc. Natl. Acad. Sci.U.S.A. 96: 6808–6813.
Bizily, S.P., Rugh, C.L., and Meagher, R.B. (2000)Phytodetoxification of hazardous organomercurials bygenetically engineered plants. Nature Biotechnol. 18:213–217.
Blaylock, M.J. and Huang, J.W. 2000. Phytoextraction ofmetals. In: Phytoremediation of Toxic Metals — Us-ing Plants to Clean Up the Environment. pp. 53–70.Raskin, I. and Ensley, B.D., Eds., Wiley, New York.
Bowler, C., van Camp, W., van Montagu, M., and Inze, D.1994. Superoxide dismutase in plants. Crit. Rev. PlantSci. 13: 199–218.
Boyd, R.S., Shaw, J.J., and Martens, S.N. 1994. Nickelhyperaccumulation defends Streptanthus polygaloides(Brassicaceae) against pathogens. Am. J. Bot. 81: 294–300.
Brewer, E.P., Saunders, J.A., Angle, J.S., Chaney, R.L., andMcIntosh, M.S. 1999. Somatic hybridization betweenthe zinc accumulator Thlaspi caerulescens and Bras-sica napus. Theor. Appl. Genet. 99: 761–771.
Brooks, R.R. 1998. Plants That Hyperaccumulate HeavyMetals. CAB International, Wallingford, U.K.
Chaney, R.L., Li, Y.M., Brown, S.L., Homer, F.A., Malik, M.,Angle, J.S., Baker, A.J.M., Reeves, R.D., and Chin, M.2000. Improving metal hyperaccumulator wild plantsto develop commercial phytoextraction systems: ap-proaches and progress. In: Phytoremediation of Con-taminated Soil and Water. pp. 129–158. Terry, N. andBañuelos, G., Eds., Lewis, Boca Raton, Florida.
Cho, U.H. and Park, J.O. 2000. Mercury-induced oxidativestress in tomato seedlings. Plant Sci. 156: 1–9.
Clemens, S., Kim, E.J., Neumann, D., and Schroeder, J.I.1999. Tolerance to toxic metals by a gene family ofphytochelatin synthases from plants and yeast. EMBOJ. 18: 3325–3333.
Cobbett, C.S. 2000. Phytochelatins and their roles in heavymetal detoxification. Plant Physiol. 123: 825–832.
Cobbett, C.S. and Goldsbrough, P.B. 2000. Mechanisms ofmetal resistance: phytochelatins and metallothioneins.In: Phytoremediation of Toxic Metals — Using Plantsto Clean up the Environment. pp. 247–271. Raskin, I.and Ensley, B.D., Eds., Wiley, New York.

453
Curie, C., Alonso, J.M., Le Jean, M., Ecker, J.R., and Briat,J.F. 2000. Involvement of Nramp1 from Arabidopsisthaliana in iron transport. Biochem. J. 347: 749–755.
Curie, C., Panaviene, Z., Loulergue, C., Dellaporta, S.L.,Briat, J.F., and Walker, E.L. 2001. Maize yellow stripe1encodes a membrane protein directly involved inFe(III) uptake. Nature 409: 346–349.
De la Fuente, J.M., Ramírez-Rodríguez, V., Cabrera-Ponce,J.L., and Herrera-Estrella, L. 1997. Aluminum toler-ance in transgenic plants by alteration of citrate syn-thesis. Science 276: 1566–1568.
Delhaize, E. 1996. A metal accumulator mutant of Arabidopsisthaliana. Plant Physiol. 111: 849–855.
De Souza, M.P., Huang, C.P.A., Chee, N., and Terry, N.1999. Rhizosphere bacteria enhance the accumulationof selenium and mercury in wetland plants. Planta209: 259–263.
De Souza, M.P., Pilon-Smits, E.A.H., and Terry, N. 2000.In: Phytoremediation of Toxic Metals — Using Plantsto Clean up the Environment. pp. 171–190. Raskin, I.and Ensley, B.D., Eds., Wiley, New York.
Dushenkov, S., and Kapulnik, Y. 2000. Phytofiltration ofmetals. In: Phytoremediation of Toxic Metals — Us-ing Plants to Clean Up the Environment. pp. 89–106.Raskin, I. and Ensley, B.D., Eds., Wiley, New York.
Ensley, B.D. 2000. Rationale for use of phytoremediation.In: Phytoremediation of Toxic Metals — Using Plantsto Clean up the Environment. pp. 1–12. Raskin, I. andEnsley, B.D., Eds., Wiley, New York.
EPA publication 542–F-98–011. 1998. A Citizen’s Guide toPhytoremediation.
Evans, K.M., Gatehouse, J.A., Lindsay, W.P., Shi, J.,Tommey, A.M., and Robinson, N.J. 1992. Expressionof the pea metallothionein-like gene PsMTA in Es-cherichia coli and Arabidopsis thaliana and analysisof trace metal ion accumulation: implications for genePsMTA function. Plant Mol. Biol. 20: 1019–1028.
Everest, A.E. and Hall, A.J. 1921. Anthocyanins andanthocyanidins. IV. Observations on (a) anthocyancolours in flowers and (b) the formation of anthocyansin plants. Proc. R. Soc. B. 92: 150–162.
Ezaki, B., Gardner, R.C., Ezaki, Y., and Matsumoto, H.2000. Expression of aluminum-induced genes intransgenic Arabidopsis plants can ameliorate alumi-num stress and/or oxidative stress. Plant Physiol. 122:657–665.
Fox, T.C. and Guerinot, M.L. 1998. Molecular biology ofcation transport in plants. Annu. Rev. Plant Physiol.Plant Mol. Biol. 49: 669–696.
Frey, B., Zierold, K., and Brunner, I. 2000. Extracellularcomplexation of Cd in the Hartig net and cytosolic Znsequestration in the fungal mantle of Picea abies –Hebeloma crustuliniforme ectomycorrhizas. Plant CellEnviron. 23: 1257–1265.
Glass, D.J. 1997. Prospects for use and regulation of transgenicplants in phytoremediation. In: In Situ and On-Sitebioremediation, Vol. 4. pp. 51–56. Alleman, B.C. andLeeson, A., Eds., Battelle Press, Columbus, Ohio.
Glass, D.J. 1999. U.S. and international markets forphytoremediation, 1999–2000. Report, D. Glass Assoc.Inc., Needham, Massachusetts.
Glass, D.J. 2000. Economic potential of phytoremediation.In: Phytoremediation of Toxic Metals — Using Plantsto Clean Up the Environment. pp. 15–33. Raskin, I.and Ensley, B.D., Eds., Wiley, New York.
Goldsbrough, P. 2000. Metal tolerance in plants: the role ofphytochelatins and metallothioneins. In: Phytoremediationof Contaminated Soil and Water. pp. 221–234. Terry, N.and Bañuelos, G., Eds., Lewis, Boca Raton, Florida.
Goto, F., Yoshihara, T., and Saiki, H. 1998. Iron accumula-tion in tobacco plants expressing soyabean ferritingene. Transgenic Res. 7: 173–180.
Goto, F., Yoshihara, T., Shigemoto, N., Toki, S., and Takaiwa,F. 1999. Iron fortification of rice seed by the soybeanferritin gene. Nature Biotechnol. 17: 282–286.
Grichko, V.P., Filby, B., and Glick, B.R. 2000. Increasedability of transgenic plants expressing the bacterialenzyme ACC deaminase to accumulate Cd, Co, Cu,Ni, Pb, and Zn. J. Biotechnol. 81: 45–53.
Guerinot, M.L. 2001. Improving rice yields — ironing outthe details. Nature Biotechnol. 19: 417–418.
Guerinot, M.L. and Salt, D.E. 2001. Fortified foods andphytoremediation. Two sides of the same coin. PlantPhysiol. 125: 164–167.
Ha, S.B., Smith, A.P., Howden, R., Dietrich, W.M., Bugg,S., O’Connell, M.J., Goldsbrough, P.B., and Cobbett,C.S. 1999. Phytochelatin synthase genes fromArabidopsis and the yeast Schizosaccharomycespombe. Plant Cell 11: 1153–1164.
Hale, K.L., McGrath, S., Lombi, E., Stack, S., Terry, N.,Pickering, I.J., George, G.N., and Pilon-Smits, E.A.H.2001. Molybdenum sequestration in Brassica: a rolefor anthocyanins? Plant Physiol. 126: 1391–1402.
Hansen, D., Duda, P.J., Zayed, A., and Terry, N. 1998.Selenium removal by constructed wetlands: role ofbiological volatilization. Environ. Sci. Technol. 32:591–97.
Hasegawa, I., Terada, E., Sunairi, M., Wakita, H., Shinmachi,F., Noguchi, A., Nakajima, M., and Yazaki, J. 1997.Genetic improvement of heavy metal tolerance inplants by transfer of the yeast metallothionein gene(CUP1). Plant Soil. 196: 277–281.
Harada, E., Choi, Y.E., Tsuchisaka, A., Obata, H., and Sano,H. 2001. Transgenic tobacco plants expressing a ricecysteine synthase gene are tolerant to toxic levels ofcadmium. J. Plant Physiol. 158: 655–661.
Heath, S.M., Southworthe, D., and D’Allura, J.A. 1997.Localization of nickel in epidermal subsidiarycel ls of leaves of Thlaspi montanum varsikiyouense (Brassicaceae) using energy-disper-sive X-ray microanalysis. Int. J. Plant Sci. 158:184–188.
Herbik, A., Koch, G., Mock, H.P., Dushkov, D., Czihal, A.,Thielmann, J., Stephan, U.W., and Bäumlein, H. 1999.Isolation, characterization and cDNA cloning ofnicotianamine synthase from barley — a key enzyme

454
for iron homeostasis in plants. Eur. J. Biochem. 265:231–239.
Higuchi, K., Kanazawa, K, Nishizawa, N.K., Chino, M., andMori, S. 1994. Purification and characterization ofnicotianamine synthase from Fe-deficient barley roots.Plant Soil 165: 173–179.
Higuchi, K., Suzuki, K., Nakanishi, H., Yamaguchi, H.,Nishizawa, N.K., and Mori, S. 1999. Cloning ofnicotianamine synthase genes, novel genes involvedin the biosynthesis of phytosiderophores. Plant Physiol.119: 471–479.
Himelblau, E., Mira, H., Lin, S., Cullota, V., Penarrubia, L.,and Amasino, R.M. 1998. Identification of a func-tional homolog of the yeast copper homeostasis geneATX1 from Arabidopsis. Plant Physiol. 117: 1227–1234.
Hirschi, K.D., Korenkov, V.D., Wilganowski, N.L., andWagner, G.J. 2000. Expression of Arabidopsis CAX2in tobacco. Altered metal accumulation and increasedmanganese tolerance. Plant Physiol. 124: 125–133.
Hoekema, A., Hirsch, R.R., Hooykaas, P., and Schilperoort,R.A. (1983). A binary vector strategy based on sepa-ration of vir and T-region of the Agrobacteriumtumefaciens Ti-plasmid. Nature 303: 179–180.
Horne, A.J. 2000. Phytoremediation by constructed wet-lands. In: Phytoremediation of Contaminated Soil andWater. pp. 13–40. Terry, N. and Bañuelos, G., Eds.,Lewis, Boca Raton, Florida.
Horsch, R.B., Fry, J.E., Fry, J.E., Hoffman, N.L., Eichholtz,D., Rogers, S.G., and Fraley, R.T. (1985). A simpleand general method for transferring genes into plants.Science 227: 1129–1232
Huang, J.W., Blaylock, M.J., Kapulnik, Y., and Ensley, B.D.1998. Phytoremediation of uranium-contaminatedsoils: role of organic acids in triggering uraniumhyperaccumulation in plants. Environ. Sci. Technol.32: 2004–2008.
Jones, P., Kortenkamp, A., O’Brien, P., Wang, G., and Yang,G. 1991. Evidence for the generation of hydroxylradicals from a chromium(V) intermediate isolatedfrom the reaction of chromate with glutathione.Biochim. Biophys. Acta 286: 652–655
Kasuga, M., Liu, Q., Miura, S., Yamaguchi-Shinozaki, K., Shinozaki,K. 1999. Improving plant drought, salt, and freezing toler-ance by gene transfer of a single stress-inducible transcrip-tion factor. Nature Biotechnol. 17: 287–291.
Kondo, T., Yoshida, K., Nakagawa, A., Kawai, T., Tamura,H., and Goto, T. 1992 Structural basis of blue-colourdevelopment in flower petals from Commelina com-munis. Nature 358: 515–518.
Krämer, U. and Chardonnens, A.N. 2001. The use oftransgenic plants in the bioremediation of soils con-taminated with trace elements. Appl. Microbiol.Biotechnol. 55: 661–672.
Krämer, U., Cotter-Howells, J.D., Charnock, J.M., Baker,A.J.M., and Smith, J.A.C. 1996. Free histidine as ametal chelator in plants that accumulate nickel. Na-ture 379: 635–638.
Krämer, U., Pickering, I.J., Prince, R.C., Raskin, I., and Salt,D.E. 2000. Subcellular localization and speculation ofnickel in hyperaccumulator and non-accumulatorThlaspi species. Plant Physiol. 122: 1343–1353.
Küpper, H., Zhao, F., and McGrath, S.P. 1999. Cellularcompartmentation of zinc in leaves of the hyperaccumulatorThlaspi caerulescens. Plant Physiol. 119: 305–311.
Lantzy, R.J. and Mackenzie, F.T. 1979. Atmospheric tracemetals: global cycles and assessment of man’s impact.Geochim. Cosmochim. Acta 43: 511–525.
Lasat, M.M., Pence, N.S., Garvin, D.F., Ebbs, S.D., andKochian, L.V. 2000. Molecular physiology of zinctransport in the Zn hyperaccumulator Thlaspicaerulescens. J. Exp. Bot. 51: 71–79.
Li, Y. and Trush, M.A. 1993. DNA damage resulting fromthe oxidation of hydroquinone by copper: role for aCu(II)/Cu(I) redox cycle and reactive oxygen genera-tion. Carcinogenes 7: 1303–1311.
Lin, Z-Q., Schemenauer, R.S., Cervinka, V., Zayed, A., Lee,A., and Terry, N. 2000. Selenium volatilization fromthe soil — Salicornia bigelovii Torr. treatment systemfor the remediation of contaminated water and soil inthe San Joaquin Valley. J. Environ. Qual. 29: 1048–1056.
Ling, H.Q., Koch, G., Bäumlein, H., and Ganal, M.W. 1999.Map-based cloning of chloronerva, a gene involved iniron uptake of higher plants encoding nicotianaminesynthase. Proc. Natl. Acad. Sci. U.S.A. 96: 7098–7103.
Lombi, E., Zhao, F.J., McGrath, S.P., Young, S.D., andSacchi, G.A. 2001. Physiological evidence for a high-affinity cadmium transporter highly expressed in aThlaspi caerulescens ecotype. New Phytol. 149: 53–60.
López-Bucio, J., Martinez de la Vega, O., Guevara-García,A., and Herrera-Estrella, L. 2000. Enhanced phospho-rus uptake in transgenic tobacco plants that overpro-duce citrate. Nature Biotechnol. 18: 450–453.
Lu, Y.P., Li, Z.S., and Rea, P.A. 1997. AtMRP1 gene ofArabidopsis encodes a glutathione S-conjugate pump:isolation and functional definition of a plant ATP-binding cassette transporter gene. Proc. Natl. Acad.Sci. U.S.A. 94: 8243–8248.
Lytle, C.M., Lytle, F.W., Yang, N., Qian, J.H., Hansen, D.,Zayed, A., and Terry, N. 1998. Reduction of Cr(VI) toCr(III) by wetland plants: potential for in situ heavymetal detoxification. Environ. Sci. Technol. 32: 3087–3093.
MacNair, M.R., Tilstone, G.H., and Smith, S.E. 2000. Thegenetics of metal tolerance and accumulation in higherplants. In: Phytoremediation of Contaminated Soiland Water. pp. 235–250. Terry, N. and Bañuelos, G.,Eds., Lewis, Boca Raton, Florida.
Marrs, K.A. 1996. The functions and regulation of glu-tathione-S-transferases in plants. Annu. Rev. PlantPhysiol. Plant Mol. Biol. 47: 127–158.
Marschner, H. 1995. Mineral Nutrition of Higher Plants.Academic. Press, London.

455
Mäser, P., Thomine, S., Schroeder, J.I., Ward, J.M., Hirschi,K., Sze, H., Talke, I.N., Amtmann, A., Maathuis,F.J.M., Sanders, D., Harper, J.F., Tchieu, J., Gribskov,M., Persans, M.W., Salt, D.E., Kim, S.A., and Guerinot,M.L. 2001. Phylogenetic relationships within cationtransporter families of Arabidopsis. Plant Physiol.126: 1646–1667.
Meagher, R.B., Rugh, C.L., Kandasamy, M.K., Gragson, G.,and Wang, N.J. (2000) Engineered phytoremediationof mercury pollution in soil and water using bacterialgenes. In: Phytoremediation of Contaminated Soiland Water. pp. 201–221. Terry, N. and Bañuelos, G.,Eds., Lewis, Boca Raton, Florida.
Misra, S. and Gedamu, L. 1989. Heavy metal toleranttransgenic Brassica napus L. and Nicotiana tabacumL. plants. Theor. Appl. Genet. 78: 161–168.
Moffat, A.S. 1999. Engineering plants to cope with metals.Science 285: 369–370.
Murphy, A.S., Eisenger, W.R., Shaff, J.E., Kochian, L.V.,and Taiz, L. 1999. Early copper-induced leakage ofK+ from Arabidopsis seedlings is mediated by ionchannels and coupled to citrate efflux. Plant Physiol.121: 1375–1382.
Nedelkoska, T.V. and Doran, P.M. 2000. Hyperaccumulationof cadmium by hairy roots of Thlaspi caerulescens.Biotechnol. Bioeng. 67: 607–615.
Nelson, N. 1999. Metal ion transporters and homeostasis.EMBO J. 18:4361–4371.
Noctor, G. and Foyer, C.H. 1998. Ascorbate and glutatione:keeping active oxygen under control. Annu. Rev. PlantPhysiol. Plant Mol. Biol. 49: 249–279.
Nriagu, J.O. 1979. Global inventory of natural and anthropo-genic emissions of trace metals to the atmosphere.Nature 279: 409–411.
Oberschall, A., Deak, M., Torok, K., Sass, L., Vass, I.,Kovacs, I., Feher, A., Dudits, D., and Horvath, G.V.2000. A novel aldose/aldehyde reductase protectstransgenic plants against lipid peroxidation underchemical and drought stress. Plant J. 24: 437–446.
Pan, A., Yang, M., Tie, F., Li, L., Chen, Z., and Ru, B. 1994.Expression of mouse metallothionein-I gene conferscadmium resistance in transgenic tobacco plants. PlantMol. Biol. 24: 341–351.
Papernik, L.A., Bethea, A.S., Singleton, T.E., Magalhaes,J.V., Garvin, D.F., and Kochian, L.V. 2001. Physi-ological basis of reduced Al tolerance in ditelosomiclines of Chinese Spring wheat. Planta 212: 829–834.
Pence, N.S., Larsen, P.B., Ebbs, S.D., Letham, D.L.D., Lasat,M.M., Garvin, D.F., Eide, D.E., and Kochian, L.V.2000. The molecular physiology of heavy metal trans-port in the Zn/Cd hyperaccumulator Thlaspicaerulescens. Proc. Natl. Acad. Sci. U.S.A. 97: 4956–4960.
Persans, M.W., Yan, X., Patnoe, J.M.L., Krämer, U., andSalt, D.E. 1999. Molecular dissection of the role ofhistidine in nickel hyperaccumulation in Thlaspigoesingense (Hálácsy). Plant Physiol. 121, 1117–1126.
Persans, M.W., Nieman, K, and Salt, D.E. 2001. Functionalactivity and role of cation-efflux family members inNi hyperaccumulation in Thlaspi goesingense. Proc.Natl. Acad. Sci. U.S.A. 98: 9995–10000.
Petit, J.M., van Wuytswinkel, O., Briat, J.F., and Lobreaux,S. 2001. Characterization of an iron-dependent regu-latory sequence involved in the transcriptional controlof AtFer1 and ZmFer1 plant ferritin genes by iron. J.Biol. Chem. 276: 5584–5590.
Pilon-Smits, E.A.H., Zhu, Y.L., Sears, T., and Terry, N.2000. Overexpression of glutathione reductase in Bras-sica juncea: effects on cadmium accumulation andtolerance. Physiol. Plant. 110: 455–460.
Pineros, M.A. and Kochian, L.V. 2001. A patch-clamp studyon the physiology of aluminum toxicity and alumi-num tolerance in maize. Identification and character-ization of Al3+-induced anion channels. Plant Physiol.125: 292–305.
Pollard, A.J., Baker, A.J.M. 1997. Deterrence of herbivoryby zinc hyperaccumulation in Thlaspi caerulescens(Brassicaceae). New Phytol. 135: 655–658.
Pollard, A.J., Dandridge, K.L., and Jhee, E.M. 2000. Eco-logical genetics and the evolution of trace elementhyperaccumulation in plants. In: Phytoremediation ofContaminated Soil and Water. pp. 251–264. Terry, N.and Bañuelos, G., Eds., Lewis, Boca Raton, Florida.
Rauser, W.E. 1995. Phytochelatins and related peptides. PlantPhysiol. 109: 1141–1149.
Rivetta, A., Negrini, N., and Cocucci, M. 1997. Involvementof Ca2+-calmodulin in Cd2+ toxicity during the earlyphases of radish (Raphanus sativus L.) seed germina-tion. Plant Cell Environ. 20, 600–608.
Robinson, N.J., Procter, C.M., Connolly, E.L., and Guerinot,M.L. 1999. A ferric-chelate reductase for iron uptakefrom soils. Nature 397: 694–697.
Rogers, E.E., Eide, D.J., and Guerinot, M.L. 2000. Alteredselectivity in an Arabidopsis metal transporter. Proc.Natl. Acad. Sci. U.S.A. 97: 12356–12360.
Ross, S.M. 1994. Toxic Metals in Soil-Plant Systems. Wiley,Chichester, U.K.
Rufyikiri, G., Declerck, S., Dufey, J.E., and Delvaux, B.2000. Arbuscular mycorrhizal fungi might alleviatealuminum toxicity in banana plants. New Phytol. 148:343–352.
Rugh, C.L., Wilde, H.D., Stack, N.M., Thompson, D.M.,Summers, A.O., and Meagher, R.B. 1996. Mercuricion reduction and resistance in transgenic Arabidopsisthaliana plants expressing a modified bacterial merAgene. Proc. Natl. Acad. Sci. U.S.A. 93: 3182–3187.
Rugh, C.L., Bizily, S.P., and Meagher, R.B. (2000)Phytoreduction of environmental mercury pollution.In: Phytoremediation of Toxic Metals — Using Plantsto Clean up the Environment. pp. 151–171. Raskin, I.and Ensley, B.D., Eds., Wiley, New York.
Salt, D.E., Prince, R.C., Pickering, I.J., and Raskin, I. 1995a.Mechanisms of cadmium mobility and accumulationin Indian mustard. Plant Physiol. 109: 1427–1433.

456
Salt, D.E., Blaylock, M., Kumar, N.P.B.A., Dushenkov, V.,Ensley, B.D., Chet, I., and Raskin, I. 1995b.Phytoremediation: a novel strategy for the removal oftoxic metals from the environment using plants. Bio-technology 13: 468–474.
Salt, D.E., Smith, R.D., and Raskin, I. 1998. Phytoremediation.Annu. Rev. Plant Physiol. Plant Mol. Biol. 49: 643–668.
Salt, D.E., Prince, R.C., Baker, A.J.M., Raskin, I., andPickering, I.J. 1999. Zinc ligands in the metalhyperaccumulator Thlaspi caerulescens as determinedusing X-ray absorption spectroscopy. Environ. Sci.Technol. 33: 713–717.
Salt, D.E. and Krämer, U. 2000. Mechanisms of metalhyperaccumulation in plants. In: Phytoremediation ofToxic Metals — Using Plants to Clean Up the Envi-ronment. pp. 231–246. Raskin, I. and Ensley, B.D.,Eds., Wiley, New York.
Samuelsen, A.I., Martin, R.C., Mok, D.W.S., and Machteld,C.M. 1998. Expression of the yeast FRE genes intransgenic tobacco. Plant Physiol. 118: 51–58.
Shaul, O., Hilgemann, D.W., de Almeida-Engler, J., vanMontagu, M., Inze, D., and Galili, G. 1999. Cloningand characterization of a novel Mg2+/H+ exchanger.EMBO J. 18: 3973–3980.
Stephan, U.W., Schmidke, I., Stephan, V.W., and Scholz, G.1996. The nicotianamine molecule is made-to-mea-sure for complexation of metal micronutrients in plants.Biometals 9: 84–90.
Su, J., Shen, Q., Ho, T.H.D., and Wu, R. 1998. Dehydration-stress-regulated transgenic expression in stably trans-formed rice plants. Plant Physiol. 117: 913–922.
Summers, A.O. 1986. Organization, expression, and evolu-tion of genes for mercury resistance. Annu. Rev.Microbiol. 40: 607–634.
Sunkar, R., Kaplan, B., Bouche, N., Arazi, T., Dolev, D.,Talke, I.N., Maathuis, F.J.M., Sanders, D., Bouchez,D., and Fromm, H. 2000. Plant J. 24: 533–542.
Takahashi, M., Yamaguchi, H., Nakanishi, H., Shioiri, T.,Nishizawa, N.K., and Mori, S. 1999. Cloning twogenes for nicotianamine aminotransferase, a criticalenzyme in iron acquisition (strategy II) in graminaceousplants. Plant Physiol. 121: 947–956.
Takahashi, M., Nakanishi, H., Kawasaki, S., Nishizawa, N.K.,and Mori, S. 2001. Enhanced tolerance of rice to lowiron availability in alkaline soils using barleynicotianamine aminotransferase genes. NatureBiotechnol. 19: 466–469.
Takeda, K., Kariuda, M., and Itoi, H. 1985. Blueing of sepalcolour of Hydrangea macrophylla. Phytochemistry24: 2251–2254.
Theil, E.C. 1987. Ferritin: structure, gene regulation, andcellular function in animals, plants and microorgan-isms. Annu. Rev. Biochem. 56: 289–315.
Thomine, S., Wang, R., Ward, J.M., Crawford, N.M., andSchroeder, J.I. 2000. Cadmium and iron transport bymembers of a plant metal transporter family inArabidopsis with homology to Nramp genes. Proc.Natl. Acad. Sci. U.S.A. 97: 4991–4996.
Van Assche, F. and Clijsters, H. 1986. Inhibition of photo-synthesis in Phaseolus vulgaris by treatment withtoxic concentration of zinc: effect on ribulose-1,5–bisphosphate carboxylase/oxygenase. J. Plant Physiol.125, 355–360.
Van der Zaal, B.J., Neuteboom, L.W., Pinas, J.E.,Chardonnens, A.N., Schat, H., Verkleij, J.A.C., andHooykaas, P.J.J. 1999. Overexpression of a novelArabidopsis gene related to putative zinc-transportergenes from animals can lead to enhanced zinc resis-tance and accumulation. Plant Physiol. 119: 1047–1055.
Vatamaniuk, O.K., Mari, S., Lu, Y.P., and Rea, P.A. 1999.AtPCS1, a phytochelatin synthase from Arabidopsis:isolation and in vitro reconstitution. Proc. Natl. Acad.Sci. U.S.A. 96: 7110–7115.
Vatamaniuk, O.K., Bucher, E.A., Ward, J.T., and Rea, P.A.2001. A new pathway for heavy metal detoxificationin animals. J. Biol. Chem. 276: 20817–20820.
Von Wiren, N., Klair, S., Bansal, S., Briat, J.F., Khodr, H.,Shiori, T., Leigh, R.A., and Hider, R.C. 1999.Nicotianamine chelates both FeIII and FeII. Implica-tions for metal transport in plants. Plant Physiol. 119:1107–1114.
Weckx, J.E.J. and Clijsters, H.M.M. 1996. Oxidative dam-age and defense mechanisms in primary leaves ofPhaseolus vulgaris as a result of root assimilation oftoxic amounts of copper. Physiol. Plant. 96: 506–512.
Whiting, S.N., de Souza, M.P., and Terry, N. 2001. Rhizo-sphere bacteria mobilize Zn for hyperaccumulationby Thlaspi caerulescens. Environ. Sci. Technol. 35:3144–3150.
Williams, L.E., Pittman, J.K., and Hall, J.L. 2000. Emergingmechanisms for heavy metal transport in plants.Biochim. Biophys. Acta 1465: 104–126.
Wolfenbarger, L.L. and Phifer, P.R. 2000. The ecologicalrisks and benefits of genetically engineered plants.Science 290: 2088–2093.
Wu, L. 1990. Colonization and establishment of plants incontaminated sites. In: Heavy Metal Tolerance inPlants: Evolutionary Aspects. pp. 269–284. Shaw,A.J., Ed., CRC Press, Boca Raton, Florida.
Xiang, C., Werner, B.L., Christensen, E.M., and Oliver,D.J. (2001) The biological functions of glu-tathione revisited in Arabidopsis transgenic plantswith altered glutathione levels. Plant Physiol.126: 564–574.
Zenk, M.H. 1996. Heavy metal detoxification in higher plants— a review. Gene 179: 21–30.
Zhu, Y., Pilon-Smits, E.A.H., Jouanin, L., and Terry, N.1999a. Overexpression of glutathione synthetase inBrassica juncea enhances cadmium tolerance andaccumulation. Plant Physiol. 119:73–79.
Zhu, Y., Pilon-Smits, E.A.H., Tarun, A., Weber, S.U., Jouanin,L., and Terry, N. 1999b. Cadmium tolerance and accumu-lation in Indian mustard is enhanced by overexpressingγ-glutamylcysteine synthetase. Plant Physiol. 121: 1169–1177.