photocontrol of stem elongation in plant … · photocontrol of stem elongation in plant...
TRANSCRIPT

Plant, Cell and Environment (1991) 14, 57-65
Photocontrol of stem elongation in plant neighbourhoods:effects of photon fluence rate under natural conditions ofradiation
C. L. BALLARE, A. L. SCOPEL & R. A. S A N C H E Z Departtnento de Ecologta, Facultad de Agronomia,Universidad de Buenos Aires, 1417 Buenos Aires, Argentina
Received 27 November 1989; received in revised form 16 April 1990; accepted for publication 5 July 1990
Abstract. We investigated the effects of photon fluencerate on internode elongation in fully de-etiolated plantsgrowing under sunlight. Our goal was to find outwhether perception by the stetns of fluence rate changesrelated to canopy density tnay be involved in themodulation of internode growth in canopies formed byplants of sitnilar stature (e.g. crop stands). Using Datutaferox L. and Sinapis alba L. seedlings growing undernatural radiation, we found that internode elongation ispromoted by localized shading. This effect was observedwith internodes receiving light with a high (>0.9) or alow (0.3) red (R) to far-red (ER) photon ratio. Selectiveremoval of the different wavebands ftotn the lightimpinging on the intetnodes showed that part of theresponse to lluence rate is due to photons in the R-I-FRrange. The blue (B) component, most likely actingthrough a specific photoreceptor, also inhibited elonga-tion. However, changes in the fiuenee rate of B light didnot have detectable effects on the response of theinternodes to R:FR ratio. Fibre-optic studies andmeasurements with integrating-cylitider sensors in even-aged populations of seedlings showed that both thequality and quantity of tadiation received by the stetnsare profoundly influenced by changes in canopy detisity.When density is very low (leaf area index = LAI ^ 1)only the R:ER ratio is reduced, due to FR reflected fromnearby leaves. In the LAI tange of 1 to 2, though a largeproportion of the leaf area is still receiving full sunlight,the photon fluence rate at the stem level drops dratnati-cally. These results suggest that in even-aged popu-lations of LAI > 1 elongation growth is promoted by thelow R:ER ratio and the reduced fiuenee rate. Perceptionof these two factors at the stem level may elicitmorphological adaptations in the canopy before theonset of severe competition among neighbours for theresource of light.
Kev-words: blue light; cryptochrornc; Datura ferox L.; light quality;phytochromc; photoriiorphogcncsi.s; plant canopies; Sinapis alba L.;stem elongation.
Abbreyiatiotis: B. blue; FR. tar-rod; G. green; LAI. leal'arca index (rii-leaf rn - soil); t'AR. photosynthctically active radiation (4()t)-7t)tlnm);Pfr. far-red absorbing form of phytochromc; R, red; SOX, low pressuresodium lamp; W. whire.
Correspondence: Carlos L. Ballarc', Department of Forest Science,Oregon State University. Peavy Hall 154. Corvallis, OR 97331. U.S.A.
Introduction
Seedlings of shade-intolerant species with orthotropicstetns typically react to the presence of neighbouringplants increasing the rate of internode elongation. It iswidely accepted that tnodifications introduced by neigh-bours in the light environtnent of the plant play aptotninent role in triggering this 'escapist' reaction.These tnodifications of the light climate include areduction in the R:FR ratio and, depending on canopydensity, structure, and orientation of the photoreceptiveotgans, a drop in photon fluence tate. The itnportance ofthe R:ER balance in the modulation of stem growth incanopies is well established (Smith, 1982, 1986; Ballareet al., 1987, 1988; Ballare, Scopel & Sanchez, 1989,1990). In contrast, the role of changes of photon fluencerate remains controversial.
Hypocotyl elongation in briefly de-etiolated seedlingsis known to be very sensitive to changes in the fiueneerate.of W light (e.g. Meijer, 1959), but the case appearstnote cotnplex with the epicotyls of fully de-etiolatedplants. There are reports of inhibitory effects of W lighton internode elongation (Garrison & Briggs, 1972;Lecharny & Jacques, 1980; Corre, 1983) and studiesshowing that total plant height in some crop speciesincreases when plants are grown under reduced irra-diances (Tarila, Ortnrod & Adedipe, 1977; Warrington,Edge & Green, 1978). However, in a large number ofother experiments, changes in total fluence rate provedto exert very little influence on elongation growth(Smith, 1981; Lecharny & Jacques, 1982; Child &Smith, 1987). Even a slight promotion of internodeextension has been found in some cases as a conse-quence of increased irradiances (Child & Smith, 1987;Casal & Smith, 1988, 1989). These latter results havelent support to the view that fluence rate drops caused bythe presence of neighbours aie likely to play only atninor part in the density-related promotion of internodeelongation in vegetation canopies (Bjorn, 1986; Child& Stnith, 1987).
Most of the above tnentioned experiments werecarried out under low PAR levels with the light treat-ments applied to the whole shoot. Therefore, theobserved lack of response to lowering the fluence ratemight be (1) a true (phototnorphogenetic) property ofthe internodes of green seedlings (e.g. Wall & Johnson,1981), (2) a consequence of limitations posed by thereduction in the rate of photosynthesis, or (3) the result
57

58 C, L, BALLARE, A, L, SCOPEL & R, A, SANCHEZ
of eomplex interactions among photoreceptive organs(e,g, Casal & Smith, 1988), Distinguishing between thefirst possibility and the other two becomes importantwhen the goal is to understand the mechanisms wherebyelongation growth is modulated in canopies formed byplants of similar stature (e,g, crop stands). In thesepopulations, where elongation plasticity may be crucialfor individual success (Grime & Jeffrey, 1965; Ballare etal., 1988), neighbouring plants may cause localizeddrops in fluence rate at the level of the internodes whenthe leaf area index of the canopy is still small, and thus,even when most of the leaves are likely to be exposed tounaltered sunlight. For example, working with even-aged monocultures of Datura ferox, we have found that,depending on height above the ground and solar eleva-tion, the presence of neighbouring seedlings may reducethe flux of R light that impinges laterally on the stems by17-60% when the LAI of the canopy is about 1,2, Atthese LAI values, less than 15% of the leaf area isshaded by neighbouring plants (Ballare et al., 1987),The observation of these uneven patterns of shading inpopulations formed by seedlings of similar heightprompted the following questions; (1) Do the internodesof fully de-etiolated plants respond to local depressionsof fluence rate, and if so, by which photoreceptor(s)? (2)Are the responses to light quality (R;FR ratio) affectedby the fluence rate sensed by the internodes? (3) Dolocalized changes in fluence rate play any role in thecontrol of elongation growth in vegetation canopies?
These questions were addressed in the work reportedhere combining physiological experiments under naturalradiation with a detailed description of the effects ofcanopy density on the light conditions sensed by plantsin even-aged populations.
Materials and methods
Plant culture
The experiments were eondueted in a greenhouse at theFaculty of Agronomy (UBA), Buenos Aires, Argentina(34° 35' S; 58° 29' W) under natural radiation withseedlings of two speeies from open habitats, Sinapis albaL. (white mustard) was used as experimental materialfrom early autumn to early spring, and Datura ferox L.,an annual weed of summer erops, was used in late springand summer experiments. Seeds of D. ferox wereobtained from plants invading soybean fields in Lobos(Buenos Aires) in 1988, Seeds were germinated in thelabortory and seedlings were planted in individual800-em'' pots filled with a light-textured soil. Seeds of 5,alba (Compant'a Fanacoa, Martinez, Buenos Aires,Argentina) were sown directly in the pots; other culturalpractices were as described previously (Ballare et al.,1989), The treatment period typically began 2 weeksafter sowing, when the first internode was between 0,5and 2cm long and the primary leaves were activelyexpanding. Elongation rates in Table 1 and Figs 4-7were obtained from length measurements taken at the
beginning and at the end of 1-week treatment periods. Inall the experiments involving manipulations ofthe lightconditions of the stems, the cotyledons were severed tofacilitate positioning of the filters and to restrict the lighttreatment to the first internode. During the experimentswith D, ferox the fluence rate of PAR at plant level wasca, 1500 /Limol m "̂̂ s"' at midday (Li 190 SB sensor,Li-Cor Inc , NE, U,S,A,); photoperiod varied between11,3 and 14,4h d~'. The temperature regitne was closeto the field situation with minima and maxima around18°C and 33°C, respectively. Typical environmentalconditions during the experiments with S. alba were asfollows; peak PAR > 1000 /xmol m^^ s"'; photoperiodbetween 9,9 and 12,9h d"'; temperature between 15°Cand25°C,
Eilters and mirrors
Figure la-c shows the three types of annular filters thatwere used to modify the light conditions of the firstinternode. Black plastic fabric (0,5-mm mesh. Fig, la)was used to reduce the fluence rate of W light at the firstinternode level without affecting the spectral photondistribution between 400 and 800nm. Cellulose-acetatefilters (Fig, lb) were used in some experiments to modifythe level of B light reeeived by the first internode.Cuvettes eonstructed with transparent polystyrene (Fig,lc) were designed to contain water solutions of organicdyes or inorganic salts, and were used to producedifferent fluence rates of B, G, R and R-I-FR radiation.In sotne cases, one of the inner walls of the cuvettes waswrapped with coloured acetates (all from La Casa delCeluloide, Buenos Aires, Argentina) in order to obtainthe desired spectral distribution (see 'if in Fig, lc),Tattrazine-OR (Mallinckrodt Chem, Works, NY,U,S,A,) solutions allowed filtering-out of the Beomponent without affeeting longer-wavelength bands,Phenolphtaleine (Cienttfica Central, Buenos Aires,Argentina) dissolved in 100 mol tn~^ Tris (pH 9) wasused to cut the G waveband. Water solutions of iodinegreen (Sleaveg SRL, Buenos Aires, Atgentina), a
Table 1, Comparison of the effects of reducing B or G light on theelongation of the lirst internode of ,SV;iu/;/.vrt//«/seedlings. In both cases,
the drop of PAR at Ihe internode level is 4(1% "
Effeet on B (450nm)light (%)
Elongation rate ± SE(mm d"') , n = 38
Promotion (%)
Control
0
1.61±0,11
—
Treatment
- B
^ 9 5
2.76±0,13
71.4
- G
- 2 1
1.93±0,12
19.9
*Cuvettc litters were used. Control liltcrs were lillccl witti distilledwater. Low B was obtained with an internal filter of orange acetate no.531, as in Fig. 5. LowG wasobtaincd with a solution of phenolphtaleine(56gm •', pH 9, see spectrum in Fig. lc). The filter had no elTect on Ihecalculated phytochrome photoequilibrium and reduced the ealeulatedrate of cycling by less than 6%.

PHOTOCONTROL OF STEM ELONGATION 59
400 600 800Wavelength (nm)
400 600 800Wavelength (nm)
400 600 800Wavelength (nm)
Figure 1. Filters and mirrors used in the experiments; (a) Neutral lilter,eonsisting of a collar of black plastic fabric (0,5-nim mesh), (b)Cellulose-acetate filters used to reduee the fluenee rate of B light, theinset shows the transmittanee curve of orange acetate no. 531 (La Casadel Celuluoide, Buenos Aires, Argentina); a transparent acetate wasused to obtain the reference spectrum, (c) Annular cuvettes used tomodify the lluence rate of B, G, R or R+FR radiation. Cuvettes werefilled with distilled water, aqueous solutions of organie dyes or CUSO4that, in eonibination with an internal lilter of cellulose aeetate,produced the different light qualities; the inset shows typieal trans-mittanee curves obtained with tartrazine (. . . , minus B), phenolph-taleine at pH 9 (—, minus G), iodine green (—, minus R), or withCUSO4 solutions and internal filters of blue aeetate (—, minus R + FR);the referenee spectrum was obtained with cuvettes filled with distilledwater, (d) Seleetive mirrors used to FR enrieh the light reeeived by thestems. Mirrors were used in combination with acetate filters in order totest the effeet of FR on stems receiving different fluenee rates of B light.Letter codes; a, cellulose-acetate lilter; e, eap; h, hypocotyl; i, firstinternode; if, internal filter of cellulose aeetate; n, black-plastic mesh;N, north; p, polystyrene euvette; the scale bars indicate 10mm,
the R:FR ratio. Some of the organic dyes tend to fade toleuco-compounds at very high fluenee rates. This wasprevented in our experiments using the appropriatebuffers and keeping the cuvettes hermetically sealed.The solutions were replaced every week in experimentsof long duration (e.g. Fig. 3), To avoid overheating ofthe internodes caused by the cuvettes, cool and humidair was putnped into the stem-filter interspace at a rate of40ctn-^s"' using3,6-mmPVCtubes. Figure ldshowsthedisposition of selective mirrors (type FrM, Ballare etal., 1987) that were used to add FR to the radiationreceived by seedlings of D. ferox. Mirrors were re-orien-ted to follow the sun during the central hours of the dayand they did not affect the light environment of the stemduring the last l,5h of the photoperiod, when thegreenhouse received diffuse sunlight only (see upperpanels in Fig, 7),
Light measurements
The effects of annular filters on the light environment ofthe first internode were evaluated with an integratingcylinder probe (Ballare et al., 1987) introduced withinthe filters in place of the stem and attached to an ISCOSR spectroradiotneter. The integrating cylinder collectsthe light flux parallel to the ground (i,e, perpendicular tothe long axis of the stem) which tnakes an importantcontribution to the photons scattered inside the inter-nodes (Rich, Whitelatn & Smith, 1987; Ballare et al.,1989), However, it is likely that, in the experimentsusing wide filtets (Fig, la & b), the actual effect oftreattnents on the light sensed by the stems had beenslightly less pronounced than what the measurementswith the integrating cylinder indicate.
The effects of canopy density on the light received bythe stems were investigated in even-aged stands of D.ferox and S. alba seedlings. The canopies covered asquare area of 0,25m', and the different LAI valueswere generated with different combinations of seedlingdensity and size. To do the measurements the integrat-ing cylinder probe was positioned in place of the centralseedling at the height of the first internode. The effectsof canopy density on the spectral photon distributioninside the stems were evaluated with a fibre-optic probeinserted into the first internode of D. ferox and S. albaseedlings and attached to a LI-1800 spectroradiometer(Li-Cor Inc,, NE, U,S, A,), A detailed description of themeasuring procedure was given in a previous paper(Ballare et al., 1989), Sensitivity of the system in the4()0-550-nm range is poor; therefore, only the resultsobtained in the R-FR waveband are presented, PARintetception per plant was measured with a cosine-corrected detector positioned at the same angle ofdisplay as the leaves (Ballare et al., 1988),
R-absorbing dye, were used to lower the R;FR ratio.Dilute solutions of CUSO4 (high absorption beyond700 nm) were cotnbined with intertial filters of blueacetate in order to reduee the lluence rate of R-hFRradiation without affecting the fluenee rate of B light or
Results and discussion
Effects of fluenee rate
The effect of neutral shading on stem elongation was

60 C. L. BALLARE, A. L. SCOPEL & R. A. SANCHEZ
4 6 8 10 12Days of treatment
14
Figure 2. Effeet of loealized shading on the elongation ot the firstmternodc o( Sinapis alba seedlings when the stems reeeived light with ahigh R:FR ratio (1.06, eontrols or l . l l , shade). Neutral shade wasobtained with blaek fabrie mesh (see Fig. la). The insel shows the effeetof the neutral filter on the fluenee rates of B (450nm), R (650nm), andFR (725nm) light (100% = no lilter). 'Vertieal bars represent ±1 SE {n= 18).
tested by fitting filters of black fabric mesh around thefirst internode of isolated seedlings of S. alba growingunder natural radiation (Eig. 1 a). This local reduction ofphoton fluence rate promoted internode elongation(Eig. 2). A similar response was observed when aR-absorbing filter was interposed between the neutralnet and the stems in order to reduce the R:ER ratio (Eig.3). These experiments, in which the light environment ofthe leaves was not altered by treatments, demonstratethat a fluence rate dependent control of elongationcontinues to operate on the internodes of fully de-etiolated plants (cf. Wall & Johnson, 1981). Our resultsagree with those of Garrison & Briggs (1972) andLecharny & Jacques (1980), who found that localtreatments of darkness stimulated internode elongationin light-grown seedlings of Helianthus and Vigna,respectively.
Photoreceptors
Effects of W light fluence rate on elongation growthhave been reported for hypocotyls of many species (e.g.Meijer, 1959). Experiments with chlorophyll-freeseedlings indicated that this pigment is not essential forthe response (Beggs et al., 1980; Ritter, Wagner &Holmes, 1981; Holmes et al., 1982). Eluence ratedependence was variously attributed to: (1) the action ofB light through cryptochrome (Gaba & Black, 1979;Thomas & Dickinson, 1979; Ritter et al., 1981); (2)changes in phytochrome cycling rate (Jose & Vince-Prue, 1978; Wall & Johnson, 1981; Gaba & Black,1985); and (3) changes in the importance of darkreactions relative to photochemical reactions in the
control of phytochrome status (Ritter el al., 1981;Holmes etal., 1982; Wall & Johnson, 1982).
To investigate the basis of the phenomenon in theepicotyls, a series of experiments was carried out with D.ferox and S. alba seedlings under natural radiation. Theinvolvement of the B component was tested usingannular filters of orange acetate fitted around theseedlings at the height of the first internode (see Eig. lb).Orange filters produced a localized depression of B light(Eig. 4, upper panels) that, in both species, promotedelongation (Eig. 4, bottom). This response can not beattributed to handling of the plants or to some side effectof the acetates, since transparent filters did not influenceinternode elongation. A promotive effect of reducing Blight was also observed when cuvette filters were used toobtain a range of fiuence rates at the first internode level(Eig. 5). The effect was more pronounced in S. alba thanin D. ferox. Laskowski & Briggs (1989) and Warpeha &Kautmaii ^198^) found that B light inhibits epicotylelongation in pea seedlings grown under continuous Rlight and concluded that inhibition was not due tochanges in Pfr concentration during the treatment withB light. It is also very unlikely that phytochrome mayhave played a role in the present experiments. This isbecause the different levels of B light were obtainedwithout affecting the R and ER components, which (1)contribute a large proportion of the photons received bythe stems, and (2), are much more efficient than B lightin driving phytochrome phototransformations (e.g.Pratt & Briggs, 1966).
Calculations, using photoconversion cross-sectionspublished by Mancinelli (1988, Table 4), showed thatthe filter that produced the minimum level of B light in
70
60
50
40
30
20
10
p IOO Lx)w R:FR +neutral shade
4 6 8 10 12Days of treafment
14
Figure 3. Effeel of loealized shading on the elongation of the firstinternode ot Sinapis atba seedlings when the stems reeeived light with alow R:FR ratio (0.30). The diamonds indieale the length of internodessurrounded by euvettes filled wilh distilled water (R:FR = 0.95).Neutral shade was obtained with black fabrie mesh and low R:FR withan R-absorbing solution of iodine green (42g ni ^). The inset summar-izes the effeets of the lilterson the fluenee rates of B, Rand FR light (D.R-absorbing filter; • , R-absorbing + neutral filter; 100% = waterfilter). Vertieal bars represent ±1 SE {n = 36).
PHOTOCONTROL OF STEM ELONGATION 61
(a ) Sinapis (b ) Datura
CDCCCCCC GDCCCCCCL L L L L U L L
CC' CC
Controls I 1Neutral shode (••
" 4
I 3
.2 2
Controls! 1Neutral shade I
0Clear Low blue " Clear
TreatmentsLow blue
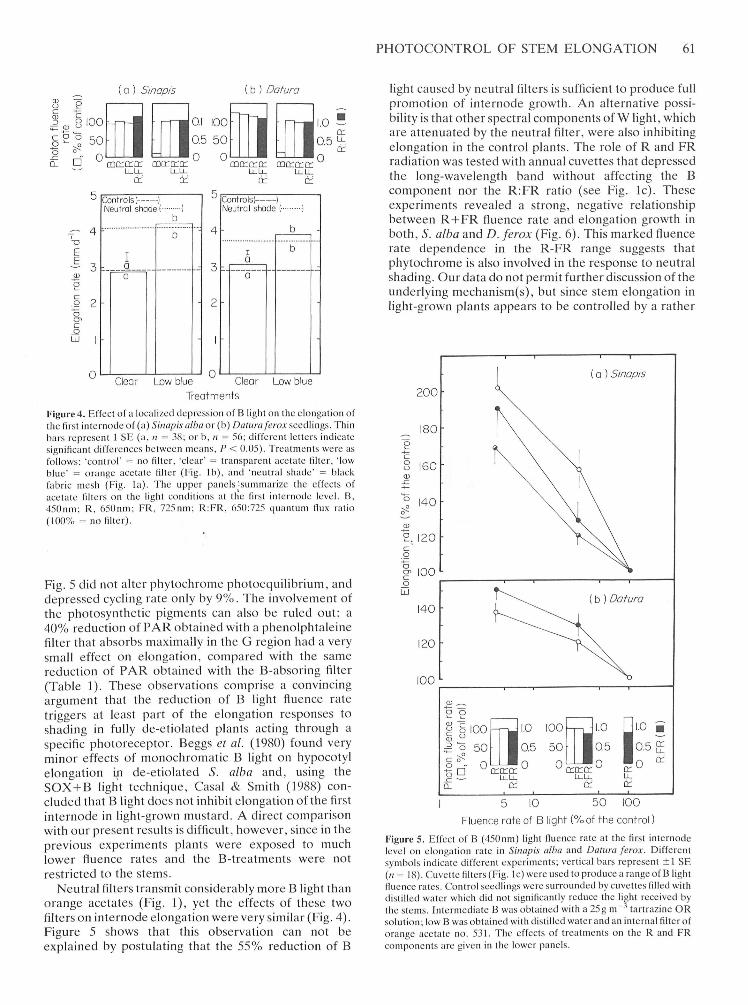
Figure 4, El'lecl of a localized depression of B light on Ihc elongation ofthe lirst internode of (a) Sinapis alba or (ii) Dattira ferox scctllings. Thinbars represent 1 SE (a, n = 38; or b, n = 56; different letters indicatesignificant differenees between means, P < (),t)5). Treatments were asfollows: 'eontrol' = no filter, 'clear' = transparent acetate filter, 'lowblue' = orange aeetale liltcr (Fig. lb), and 'neutral shade' = blackfabric mesh (Fig, la). The upper panels'summarize the elTeets ofacetate lilters on the light conditions at the lirsl internode level, B,450nm; R, 650nni; PR, 725nm; R:FR, 650:725 quantum llux ratio(100% = no filter).
Fig, 5 did not alter phytoehrome photoequilibrium, anddepressed eyeling rate only by 9%, The involvetnent ofthe photosynthetie pigtnents ean also be ruled out: a40% reduetion of PAR obtained with a phenolphtaleinefiller that absorbs maximally in the G region had a verysmall effeet on elongation, eompared with the samereduetion of PAR obtained with the B-absoring filter(Table 1), These observations eomprise a eonvineingargument that the reduetion of B light fluenee ratetriggers at least part of the elongation responses toshading in fully de-etiolated plants aeting through aspeeifie photoreeeptor, Beggs et cd. (1980) found veryminor effeets of monoehromatie B light on hypoeotylelongation in de-etiolated S. alba and, using theSOX+B light teehnique, Casal & Stnith (1988) eon-eluded that B light does not inhibit elongation ofthe firstinternode in light-grown mustard, A direet eomparisonwith our present results is diflieult, however, sinee in theprevious experitnents plants were exposed to muehlower fluence rates and the B-treatments were notrestricted to the stems.
Neutral filters transmit considerably tnore B light thanorange acetates (Fig, 1), yet the effects of these twofilters on internode elongation were very similar (Fig, 4),Figure 5 shows that this observation can not beexplained by postulating that the 55% reduction of B
light caused by neutral filters is sufficient to produee fullpromotion of internode growth. An alternative possi-bility is that other spectral components of W light, whichare attenuated by the neutral filter, were also inhibitingelongation in the control plants. The role of R and FRradiation was tested with annual cuvettes that depressedthe long-wavelength band without affeeting the Beomponent nor the R:FR ratio (see Fig, lc). Theseexperitnents revealed a strong, negative relationshipbetween R + FR fluenee rate and elongation growth inboth, S. alba and D. ferox (Fig, 6), This marked flueneerate dependence in the R-FR range suggests thatphytochrome is also involved in the response to neutralshading. Our data do not permit further discussion oftheunderlying tneehanism(s), but since stem elongation inlight-grown plants appears to be controlled by a rather
200
180
160
° 140
2, 120
100
LJ
140
120
100
(a ) Sinapts
(b ) Datura
"o 'o
"11u
^=- o
o ° •-
DO
50
0
T i1cccccc
LLU_
ci.
IO
05
0
100
50
0
5 10Fluence rate ot B light
50 100%of the control)
Figure 5, Effeet of B (450nm) tight lluence rate at the first internodelevet on elongation rate in Sinapis alba and Datura ferox. Differentsymbols indicate different experiments; vertical bars represent ± 1 SE(n = 18). Cuvette lilters (Fig. lc) were used to produee a range of B lighttluence rates. Control seedlings were surrounded by cuvettes filled withdistilled water which did not significantly reduce the light received bythe steins. Intermediate B was obtained with a 25g m ' tartrazine ORsolution; low B was obtained with distilled water and an internal filter oforange acetate no. 531. The effects of treatments on the R and FRcomponents are given in the lower panels.

62 C. L. BALLARE, A. L. SCOPEL & R. A. S A N C H E Z
200
180
„ 160
140
120
_o
100
160
140
120
100
(a ) Stnapts
[ b) Dafura
I 5 10 50 100Fluence rate of [R + FR] ligfit (% of fhe confrol)
Figure 6, Effects of R (625nm) + FR (725 nm) lluence rate al the firstinternode level on elongation rate in Sinapis alba and Datura ferox.Different symbols indicate difterent experiments; vertical bars rep-resent ±SE (n = 18). Cuvette lilters (Fig. lc) were used to produce theR-I-FR gradient by combining CuSO^ solutions with internal lilters ofblue acetate as follows: intermediate R-I-FR = 6.2Kg ni"^ CUSO4 -I-acetate no. 502; low R - F F R = 25Kg m"' CUSO4 + acetate no. 501.Control seedlings were surrounded by cuvettes frlled with distilledwater. The effects of treatments on the B (450nni) component and theR:FR ratio are given in the lower panel.
Stable pool of phytochrome (Jabben & Holmes, 1983;Schafer, Ebert & Schweitzer, 1984), it is unlikely thatthe observed fiuenee rate dependence may be explainedon the basis of phytochrome dark reactions.
Effects on B light on the response to R.ER ratio
The effects of daytime changes in the R:FR balance onstem elongation have largely been explored with plantsgrown under modest levels of W light (e.g. Morgan &Smith, 1978; Lecharny & Jacques, 1982). The possibilityexists that, under strong sunlight, inhibition exerted bythe B component (Figs 4 & 5) may reduce the effect oflowering the R:FR ratio. We investigated this possibilitywith D. ferox seedlings growing under natural radiationin the summer. Selective reflecting mirrors (Fig. fd)were used to add FR during daytime to the light received
by stems subjected to different levels of B light. Themirrors had no measurable effect on the light conditionsat the end of the day (Fig. 7, upper panels). The results(Fig, 7, bottom) showed that internode elongation canbe promoted by low R:FR ratios even if they areestablished during the central part of the photoperiod,when plants are exposed to very high fiuenee rates. Thegreatest elongation rates were obtained when inter-nodes were exposed sitnultaneously to low R:FR ratiosand low B light levels, but analysis of variance indicatedthat the interaction 'FR X B' was not significant (f-test;P > 0,05). Thus, in this system, the effect of R:FRappeared to be independent of the fiuenee rate of B lightreceived by the internodes.
Ecological significance
We used a fibre-optic probe inserted into the stetn tissueand an integrating cylinder probe in order to obtain adetailed description of the etfects of canopy density onthe light clitnate of the internodes of D. ferox and S. albaseedlings growing in even-aged tnonocultures. Thesetwo approaches gave very sitnilar results (Fig. 8a & b).
Dayfime Cnd-of-day Doyfime
P P200
100
0 amcrtrcr cQCt:ct:cr
L L _ 1 _ I _ I l l l
Control-I-
FR
^ 4
E
'5
i 2
Elo
nga
3 _
I
bb
a
c
Lowblue
• Confrol -
-
Confrol-I-FR
Low blue-I-FR
Treofnienf
Figure 7, Effeets of FR added during daytime to Datura ferox seedlingson the elongation of the internodes when these were subjeeted todifterent levels of B light. The thin bar represents 1 SE (n = 64);different letters denote significant dilferences between treatmentmeans (P < O.t)5). Internodes were surrounded by transparent acetateniters (control) or orange acetate (low blue); additional FR wasprovided with mirrors (see Fig. Id for details). The upper panelssummarize the light conditions at the tirst internode level.

PHOTOCONTROL OE STEM ELONGATION 63
ooa
( b )
o =RL A = F R
A'l'- "*-'.oOo'^To'^—
° o° °̂ °
10" IOrO.5 10',0 10',0.5 10,-1.5 10'-1.0 10- 0 5 10',0 10'0.5
0 0.05 010 050 1.00 2.00 5.00Leaf area index
0 0.05 O.IO 0.50 1.00 2.00 5.00Leaf area index
Figure 8. Elfects of eanopy density on the light ciiviromnent of Ihe lirst internode; (a) measurements obtained with the integratingcylinder probe positioned in the ecntre of the canopy al the height of the lirsl internode; (b) measurements of the light regime within thefirsl inlernode obtained with a libre-optie probe inserted into the stem tissue. In both panels, a value of 1 In the ordinate represents thephoton lluence rate measured at the corresponding wavelength when the LAI of the eanopy was zero. Results obtained with Daturafeto.x and .Sitiapis alba populations were poolctl as no qualitative diflerenees were found between these two data sets.
and both demonstrated quite clearly that the proximityof neighbouring plants tnay have dramatic effects on thequality fl/7f/ quantity of light received by the stetns beforecausing a severe reduction in the amount of PARintercepted at leaf level (Eig. 9).
The most important consequence of increasing thenumber of neighbours in the very-low LAI range (LAI^ 1) is a marked rise in the fiuence rate of ER (Eig. 8a &b). The resulting drop in the R:ER ratio may besufficient to promote elongation in fully illutninatedseedlings (Eig. 7), and this shift in spectral photondistribution tnay be among the first signals of neighbour-hood density perceived by plants in uniform populations(Ballare et al., 1987, 1989; Smith, Casal & Jackson,1990). This proposal is supported by the observationthat seedlings growing in even-aged stands begin toelongate faster than isolated controls before theybecome shaded by neighbours, and that this responsecan be reduced if the R:ER ratio received by theinternodes is kept constant and high with a ER-absor-bing filter of CuSOa (Ballare e/a/., 1990).
Even if the density of the canopy is very low, theproximity of other plants may reduce the photon fluencerate received by the stems at the extremes of thephotoperiod (Ballare et al., 1987). Previous experi-ments with D. ferox indicated that transient depressionsof this kind are unlikely to have important effects oninternode growth (Ballare et al., 1989). When the LAIof the canopy is increased further, however, there is a
sharp fall in the light flux perpendicular lo the stetn axis(Eig. 8a). which is paralleled by a reduction in theamount of photons scattered within the internodes (Eig.8b). Variability in the light records at LAI 5̂ 1 isconsiderable, reflecting the importance of small changesin the spatial arrangetnent of the leaves standing in theimtnediate proximity of the target internode. Althoughwe do not know how this stnall-scale variability is'integrated', our results (Eigs 2 & 3) strongly suggest thatinternode elongation will be promoted by a local fluencerate drop that, on the average, may be as high as 70% inpopulations of LAI = 2 (Eig. 8). Notice that, in such
IO
0 0.05 01 0.5 1.00 200 5.00Leaf areo index
Figure 9. Effects of increasing LAI in Sinapis alba canopies on PARinterception per plant. Results are presented as pereentage of the meanvalue for isolated seedlings; vertieal bars indicate ±1 SE {tt = 5).

64 C, L, BALLARE, A, L, SCOPEL & R, A, S A N C H E Z
canopies, PAR interception per plant is barely 20-25%lower than that of isolated individuals (Fig, 9),
Conclusions
Our data show that epicotyl elongation in fully de-etiolated plants is affected by the photon fluence ratereceived by the internodes. This response is likely toinvolve the action of both, cryptochrome and phyto-chrome. We propose that the relative contributions oflight quality and quantity to the control of elongationgrowth in eanopies depends on faetors such as canopydensity and vertical organization. Thus the R:FR ratioappears to be the most important eontrolling faetor intwo extreme situations, namely: (1) in uniform stands ofvery low LAI, where FR seattered by adjaeent leavesmay alter the R:FR balanee reeeived by the stems ofunshaded plants (Ballare etat,, 1987, 1989, 1990), and(2), in the ease of plants that experienee extremely lowirradiances under dense eanopies, where elongationresponses to small variations in fluenee rate are likely tobe very minor (Child & Smith, 1987), But for inter-mediate situations, whieh are very frequent in plantcommunities of arable land, the depressions of lightquantity caused by neighbours are almost eertainlyinvolved, together with the R:FR balance, in the controlof internode growth.
Our results also demonstrate that in even-aged plantpopulations the intensity and spectral distribution of thelight received by the stems may be substantially alteredbefore the LAI of the eanopy beeotnes large enough toimply a severe reduetion in PAR intereeption by leaves.Light changes at the stem level may thus represent thefirst indicators of future competition for light, and theirsignifieanee for the morphological adjustment of shade-intolerant species might be substantial.
Acknowledgment
This work was supported finaneially by grants from theConsejo Naeional de Investigaeiones Cientt'fieas y Tee-nicas (PID 307040088) and the University of BuenosAires (AG 062), Argentina,
References
Ballare, CL, , Sanchez, R.A., Seopel, A.L,, Casal, J,J, & Ghersa,CM, (1987) Early detection of neighbour plants by phytoehromepereeption of speetral ehanges in relleeted snntight. Plant, Celt andEnvironment, 10,551-557.
Ballare, CL. , Sanehcz, R.A., Seopel, A,L. & Ghersa, CM. (1988)Morphological responses of Datura ferox seedlings to the preseneeof neighbours: their relationships with eanopy mieroellmate, Oeeo-logia (Berlin), 76, 28H-293.
Ballare, CL, , Seopel, A.L. & Sanehez, R.A, (1989) Photomodu-lation of axis extension in sparse eanopies: role of the stem in thepereeption of light-quality signals of stand density. Plant Physiology,89, 1324-1330,
Ballare, C.L., Seopel, A.L. & Sanehez, R.A. (199(1) Far-red radia-tion refleeted from adjacent leaves: an early signal of competition inplant eanopies. Seienee, 247, 329-332,
Beggs, CJ , , Holmes, M.G., Jabben, M. & Sehafer, E, (1980) Actionspeetra for the inhibition of hypoeotyl growth by continuous
irradiation in tight and dart<-grown Sinapis alba L. seedtings. I'lantPhysiology, 66, 615-618,
Bjorn, L.O. (1986) Introduction. In Photomorphogenesis in Plants(eds R.E. Kendriek & G.H.M, Kronenberg), pp. 3-14, MarthinusNiholf Publishers, Dordrecht.
Casal, J.J. & Smith, H. (1988) The loei of pereeption lor phytoehromeeontrot of internode growth in light-grown mustard: promotion bylow phytochrome photoequilibria in the internode is enhaneed byblue light perceived by the leaves. Planta. 176, 277-282.
Casal, J.J. & Smith, H. (t989)The 'end-of-day' phytoehrome eontrol ofinternode elongation in mustard: kineties, interaction with theprevious fluenee rate and eeologieal implieations. Plant, Cell andEnvironment, 176, 511-520.
Child, R. & Smith, H. (1987) Phytochrome aetion in light-grownmustard: kinetics, fluenee-rate compensation and ecological signifi-cance. Planta. 172,219-229,
Corre, W.J. (1983) Growth and morphogenesis of sun and shadeplants. II. The inlluence of light quality. Acta Botaniea Neerlandiea,32, 185-202.
Gaba, V. & Blaek, M. (t979) Two separate photoreeeptors eontrothypocotyl growth in green seedlings. Nature, 278, 51-54.
Gaba, V. & Blaek, M. (1985) Photocontrot of hypocotyl elongation intight-grown Cueumis sativus L.: responses to phytoehrome photo-stationary state and tluenee rate. PlatU Physiology, 79, lOt t-t()t4.
Garrison, R. & Briggs, W.R. (1972) Internodal growth in localizeddarkness. Botanieal Gazette. 133, 270-276.
Grime, J.P. & Jeffrey, D.W. (1965) Seedling estabhshment in vertiealgradients of sunlight. Jotirnal of Eeology, S3, 621-642.
Holmes, M.G., Beggs, C.J., Jabben, M. & Sehafer, E. (1982)Hypocotyl growth in Sinapis alba L.: the roles of light quality andquantity. Plant, Cell and Environment. 5, 45-51.
Jabben, M, & Holmes, M,G. (1963) Phytoehrome in light-grownplants. In Encyclopedia of Plant Physiology (New Series), Vol. 16B(eds W. Shropshire Jr & H, Mohr), pp. 704-722. Springer-Verlag,Berlin.
Jose, A.M. & Vince-Prue, D. (1978) Phytochrome action: a reapprai-sal. Photoeheinistry and Photobiology. 11, 209-216.
Laskowski, M.J. & Briggs, W.R. (1989) Regulation of pea epicotylelongation by blue-light: lluenee response retationships and growthdistribution. Plant Physiology. 89, 293-298.
Lecharny, A. & Jacques, R, (198(1) Light inhibition of internodeelongation in green plants. A kinetic study with Vigna sinensis L.Planta. 149, 384-388.
Lecharny, A. & Jaques, R. (1982) Photo-inhibition of internodeelongation rate in light-grown Vigna sinesis L,: control by lightquality. Plant, Cell and Environment, 5, 3l-,36.
Maneinelli, A.L. (1988) Some thoughts about the use of predictedvalues of the state of phytochrome in plant photomorphogenesisresearch. Plant, Cell and Environment, 11,429-439.
Meijer, G. (1959) The speetral dependence of llowering and elonga-tion, Acta Botaniea Neerlandiea, 8, 189-246.
Morgan, D.C & Smith, II. (1978) The relationship between phyto-ehrome photoequilibrium and development in light-grown Cheno-podium album L. Planta, 142, 187-193.
Pratt, L. & Briggs, W.R. (1966) Photochemical and nonphotochcmiealreaetions of phytoehrome /;; vivo. Plant Physiology, 41, 467-474.
Rieh,T,CG., Whitelam, G.C & Smith, H. (1987) Analysis ol growthrates during phototropism: modifications by separate light-growthresponses. Plant, Celt and Environment, 10, 303-311.
Ritter, A,, Wagner, E. & Holmes, M.G. (1981) Light quantity andquality interactions in the eontrol of elongation growth in light-grown Chetiopoditim rubrum L. seedlings. Ptanta. 153, 556-560,
Sehafer, E., Ebert, C & Sehweitzer, M. (1984) Control of hypocotylgrowth in mustard seedlings after light-dark transitions. Photo-ehemistry and Photobiotogy. 39, 95-100.
Smith, H. (1981) Adaptation to shade. In Physiological ProeessesEimiting Plant Produetivity (ed. C.B, Johnson), pp. 159-173.Butterworths, London.
Smith, H. (1982) Light quatity, photoperception, and plant strategy.Annual Review of Plant Physiology. 33, 481-518,
Smith, H, (1966) The pereeption of light quality. In Photomorphogene-sis in Plants (eds R.E, Kendriek & G.H.M. Kronenberg), pp.187-217. Marthinus Nihoff Publishers, Dordreeht.

PHOTOCONTROL OF STEM ELONGATION 65
Smith. H., Casal, J.J. & Jackson. G.M. (lyyO) Rettection signals andthe perception by phytoehrome of the proximity of neighbouringvegetation. Plant. Cell and Envirotitnent, 13, 7.V78.
Tarila. A.G.I., Ormrod. D.P. & Adedipe. N.O. (1977) Effeets ofphosphorus nutrition and light intensity on growth and developtnentof the cowpea (Vigna unguiculata L.). Amtals of Botatiy, 41, 75-83.
Thotnas, B. & Dickinson. hl.G. (t97y) lividenec for two photorecep-tors eontrolling growth in de-etiolated seedlings. Platita. 146,545-550.
Wall. J.K. & Johnson. C.B. (1981) Phytoehtome action in light-grown
plants: the influence of light quality and lluence rate on extensiongrowth in Sittapis alba L. Plattta. 153, 10t-t08.
Wall. J.K. & Johnson, C.B. (1982) The effeet of temperature onphytoehrome controlled hypocotyl extension in Sinapis alba L. NewPhytologist, 91, 405-412.
Warphea. K.M.E. & Kauftnan, L.S. (1989) Blue-light regulation ofepicotyl elongation in Pisitnisativum. Plant Physiology, 89, 544-548.
Warrington. I.J.. Edge, E.A. & Green, L.M. (1978) Plant growthunder high radiant energy fluxes. Annals of Botany, 42, 1305-t313.
