peripheral and central vascular smooth muscle cells from rat lung exhibit different cytoskeletal...
TRANSCRIPT

JOURNAL OF CELLULAR PHYSIOLOGY 159399406 (1994)
Peripheral and Central Vascular Smooth Muscle Cells From Rat Lung Exhibit Different
Cytoskeletal Protein Profiles but Similar Growth Factor Requirements
PAUL DAVIES* AND WAYNE PATTON Department of Pharmacology, University of Pittsburgh School of Medicine,
Pittsburgh, Pennsylvania 1526 1 (P. D.) and Millipore Corporation, Bedford, Massachusetts 0 7 730 (W.P.)
In pulmonary vascular remodelling, the lining smooth muscle cells undergo vari- ous forms of growth involving cellular hypertrophy and hyperplasia. Differences in the growth pattern between central and peripheral regions suggested that cells from both should be obtained when investigating the cellular basis for the remod- elling. Accordingly, we have obtained two smooth muscle cell types in culture: a cell from the central pulmonary artery (CC) and a cell morphologically similar to a pericyte (PC), from the periphery of the lung. Both cell types gave positive imrnu- nostaining for a-smooth muscle isoactin. In vivo, the a-isoactin was immunolo- calized in the extracapillary vasculature. Quantitative two-dimensional gel elec- trophoresis of cell extracts showed that PC express more vimentin and gelsolin than CC. Despite the differences between PC and CC in the expression of cytoskel- eta1 proteins, their response to growth factors was similar. Both cell types in- creased DNA synthesis when stimulated by exogenous PDGF-AB. This occurred in the absence of exogenous progression factors, but depended on a post-compe- tence, suramin-sensitive mechanism that probably represents an autocrine pro- gression factor. The cells were also stimulated by IGF-1 alone, in the absence of exogenous competence factors. At an IGF-1 concentration of 1 ns/ml, this re- sponse appeared specific for the IGF-1 receptor and was sensitive to pretreatment with pertussis toxin, thus implicating a role for a G protein. 0 1994 Wiley-Liss, Inc.
The growth of pulmonary vascular smooth muscle is a feature of normal lung development and a number of diseases. In pulmonary hypertension (PHI, for example, hypertrophy and hyperplasia of smooth muscle occur as part of the vascular remodelling that characterizes the disease (Reid, 1990). These changes are also manifested in animal models of PH (Davies et al., 1987a). The timing and the degree of change, however, are different a t central and peripheral levels. This is particularly evident in the chronic hypoxic rat model: by autorad- iography, thymidine uptake increases early in the so- called precursor smooth muscle cells (intermediate cells and pericytes [PC]) of the intra-acinar arteries. In the smooth muscle cells of the central pulmonary artery (CC), the index rises much later and then only rather moderately (Meyrick and Reid, 1979). These findings suggest that smooth muscle cells in peripheral vessels exhibit a different growth response from cells in central vessels either because of primary differences between the cells or because conditions are different a t the two levels. To test the first of these possibilities, we have made efforts to obtain cultures of smooth muscle cells representative of peripheral and central populations. A method for establishing cultures of PCs from the bovine retinal microvasculature (Gitlin and D’Amore, 1979) Q 1994 WILEY-LISS, INC
was adapted and used to obtain from the lung a popula- tion of cells morphologically similar to PCS (Davies et al., 1987b). In the absence of a specific marker for pul- monary microvascular cells in culture, the cells ob- tained from the lung periphery have hitherto lacked identifiable characteristics that distinguish them from central vascular smooth muscle cells.
We now compare these two populations further with respect to their expression of cytoskeletal proteins and response to growth factors. Because smooth muscle cells in the peripheral lung are sparsely distributed and enter growth phase from a condition of stasis, we chose to perform the studies in subconfluent monolayers, fol- lowing a period of growth arrest. This also allowed us to determine the sequential requirements with respect to competence and progression factors.
Received February 11,1992; accepted December 14,1993. *To whom reprint requests/correspondence should be addressed. Dr. Patton’s present address is Molecular Biology Department, Sloan-Kettering Institute, New York, NY 10021. Address correspondence to Paul Davies, Ph.D., Department of Pharmacology, Biomedical Science Tower, E1351, University of Pittsburgh School of Medicine, Pittsburgh, PA 15261.

400 DAVIES AND PATTON
MATERIALS AND METHODS Cell culture
Cell cultures were established using procedures sim- ilar to those published previously (Davies et al., 1987b). Adult male Sprague Dawley rats (Charles River Breed- ing Laboratories, Wilmington, MA) weighing 125-150 g were anesthetized. For peripheral (PC) cultures, we conventionally used four rats. The lungs were excised and placed in phosphate buffered saline (PBS). The dis- tal third of each lobe was removed and minced manu- ally with a single-edged razor. The minced tissue was pooled and digested with collagenase (Worthington type IV) at a concentration of 4 U/ml in Hanks' bal- anced salt solution (HBSS) at 37°C for 30 min. After washing in HBSS, the digested tissue was filtered through a 110 pm nylon mesh. The filtrate consisted of single cells o r small clumps of cells. These were resus- pended in medium consisting of a 1:l mixture of Dul- becco's modified eagle's medium (DMEM) and Ham's F12 containing 4 mM glutamine, 100 U penicillin/ streptomycin, and supplemented with 10% fetal bovine serum (FBS). The cells were plated into 35 mm wells and incubated at 37°C in an incubator with 5% CO,. After 3 hr the medium was changed.
Cultures of central smooth muscle cells (CC) were obtained from explants of the central (intrapulmonary) artery of the left lung. Under a binocular microscope, the artery was dissected out of the surrounding tissue and placed in sterile PBS. The intimal surface was ex- posed by cutting open the artery lengthwise with a pair of iridectomy scissors, then removed by gentle scraping with fine forceps. The artery was cut into explants ap- proximately 5 mm square. Each explant was placed medial side downwards in a T25 flask with a small volume of the same medium used for PC, and left undis- turbed in the incubator for 3 days. At the end of this time, smooth muscle cells could usually be seen migrat- ing from the explant on to the plastic. After a few days, the explant was removed and 4 ml of medium added to the flask.
Cultures of both cell types were fed on alternate days and passaged by trypsinization with 0.05% trypsid 0.02% EDTA using a split ratio of 1:2 or 1:3. They were never used beyond passage 6.
Immunostaining Cells were plated into two- or four-chamber Lab-Tek
(Nunc, Naperville, IL) slides and grown to subconflu- ence. They were fixed in freshly prepared, ice-cold 4% formaldehyde in PBS for 5 min and permeabilized in 10% dimethylsulfoxide (DMSO). After a 30-min incuba- tion in blocking serum, primary antibody against a-smooth muscle isoactin (monoclonal mouse IgG, Sigma, St. Louis, MO) was applied. Signal was detected by incubating with an anti-mouse IgG secondary anti- body conjugated with rhodamine (Sigma). In controls, the primary antibody was replaced by nonimmune mouse serum at the same dilution.
To localize a-smooth muscle isoactin in vivo, fresh frozen sections 10 pm thick were cut on a Reichert 2800 E cryostat and mounted on poly-lysine-coated glass slides. After blocking antibody, they were incubated overnight with monoclonal primary antibody. Signal was detected in the same way as in the cell monolayers.
Fluorescence staining in cells and tissue sections was viewed and photographed in a Nikon photomicroscope.
Two-dimensional (2-D) sodiumdodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE)
Cells of each type were grown to confluence in T75 flasks. They were washed three times in ice-cold Dul- becco's PBS while being kept on ice. Wash buffer was removed and the cells were treated with boiling sample buffer consisting of SDS (10 mM), P-mercaptoethanol (50 mM) in Tris buf€er, pH 8.8. Cells were scraped &om the dish with a rubber policeman, transferred to an eppendorf tube, and heated at 100°C for 5 min. The resulting lysate was then snap frozen in liquid N,. Prior to electrophoresis, samples were treated with 100 pg/ml DNase I, 25 pg/ml RNase A for 5 min, precipi- tated in 80% cold acetone, and resuspended in 4 mg/ml in 9 M urea, 4% Nonidet P-40 (NP-401, 2.2% carrier ampholytes, 100 mM dithiothreitol (DTT).
High resolution 2-D gel electrophoresis was per- formed according to the method of OFarrell(1975) with modifications (Patton et al., 1989, 1990a). Isoelectric focusing (IEF) was performed in a 4% T, 2.6% C poly- acrylamide tube gel (18 cm long and 1 mm internal diameter) prepared from a 30.8% T, 2.6% C (30% w/v acrylamide, 0.8% w/v N,N'-methylene bisacrylamide) stock solution. The IEF gel also contained 9.5 M urea, 2.0% v/v NP-40, and 2.0% v/v Millipore 2-D optimized carrier ampholytes (Patton et al., 1990a). Approxi- mately 20 kg of protein were applied per gel. IEF gels were prefocused to 1,500 volts over a 2-hr period with current limiting to 110 pNgel. Focusing was conducted for a total of 18,000 volt-hr (17 hr a t 1,000 volts, 30 min at 2,000 volts).
The standard second dimension electrophoretic sepa- ration was performed in 10% T, 2.6% C high tensile strength acrylamide-based matrix (DuracrylT*', Milli- pore Corp., Bedford, MA). The polyacrylamide gels con- tained 0.375 M Tris-base/Tris-HC1, 0.1% SDS, pH 8.8, with conductivity of 4 mS/cm, and an electrode buffer of 0.025 M Tris-base, 0.192 M glycine, 0.1% SDS, pH 8.3 (Laemmli, 1970).
The gels (1 mm thick, 22 cm wide, 22 cm long) were run without stacking gels a t a constant power of 20 W per gel as previously described (Patton et al., 1990a). After completion of the electrophoretic separation, gels were fixed for 2-12 hr in 50% methanol, 10% acetic acid. Silver staining was performed according to the method of Morrissey (1981) except that gels were not fixed in 10% glutaraldehyde and the development of the silver stain was stopped using 3.0% v/v acetic acid instead of 2.3 M citric acid. The principal cytoskeletal proteins have been identified in our laboratory by 2-D gel electrophoresis, Western blotting with monoclonal antibodies, and comigration with purified proteins (Chung-Welch et al., 1989; Patton et al., 1990b).
Protein electroblotting and visualization of gelsolin
One-dimensional gel electrophoresis was used to de- tect gelsolin in smooth muscle and PC cell lysates. To- tal protein amount was determined using a Bio-Rad (Hercules, CAI colorimetric protein assay kit, and visu- ally verified using electroblots stained with Amido

40 1 CENTRAL AND PERIPHERAL PULMONARY SMOOTH MUSCLE CELLS
Black. Duplicate lanes of each cell lysate were run us- ing the method of Laemmli (1970) and proteins were transferred to nitrocellulose sheets by the method of Towbin et al. (1979). Prior to electroblotting, the gels were allowed to pre-equilibrate at room temperature in electroblot bufTer (50 mM Tris-base, 95 mM glycine, 0.005% SDS) for 3045 min. The transfer to nitrocellu- lose paper was performed for 3 hr at 250 mA constant current and 40-50 volts. The blots were incubated over- night with a 1:250 dilution of monoclonal anti-gelsolin antibody (clone no. GS = 2C4). The blots were then incubated with alkaline phosphatase-conjugated rabbit anti-mouse IgG or rabbit anti-guinea pig IgG diluted 1:1,000, and visualized using a nitroblue tetrazolium chloride/5-bromo-4-chloro-3-indoylphosphate p-toluid- ine chromogen system. All antibodies and chromogens were purchased from Sigma.
Two separate cultures of each cell type were evalu- ated and two samples from each culture were run on separate gels and transferred to nitrocellulose. Anti- body binding was quantified densitometrically using the Millipore Bioimage 110 computerized gel analysis system (Patton et al., 1990a). All gels were normalized to one another with respect to total protein load using the “RATIO’ and “NORMALIZE” options in the pro- gram.
Growth assays Cells were trypsinized, resuspended in DMEM sup-
plemented with 0.5% FBS, and plated into 16 mm wells at a density of approximately 2 x lo4 cells/well con- firmed by hemocytometer counts. They were then growth arrested by replacing the medium with a de- fined, serum-free medium originally described by Weinstein et al. (1981) as modified by Nemecek et al. (1986) containing DMEM, glutamine (6 mM), ovalbu- min (0.1%), ascorbic acid (25 bg/ml), sodium selenite (25 ng/ml), transferrin (10 pg/ml), and hydrocortisone (50 pM). In some experiments, Cohn Fraction IV (a preparation principally of a and p globulins) was added to 10 Fg/ml, but was not found to alter the responses. They remained in serum-free medium for 2 days. Growth arrest was confirmed by determining 3H-thy- midine (6.7 Ci/mM) uptake into trichloroacetic acid (TC&precipitable DNA (Raines and Ross, 1982) fol- lowing a 4-hr pulse a t a concentration of 2 pCi/ml. The following growth factors were added either singly or in sequence as explained in the text: human platelet de- rived growth factor (PDGF), epidermal growth factor (EGF) (both from Collaborative Research, Bedford, MA), and human recombinant insulin-like growth fac- tor (1GF)-1 (Imcera, Terre Haute, IN). Suramin was obtained from Mobay Corporation (New York, NY). The effects on growth were determined after 48 hr by measuring 3H-thymidine uptake following a 24-hr pulse at a concentration of 2 pCi/ml.
Statistical analysis In analyzing the densitometric levels of cytoskeletal
proteins on 2-D gels, differences between the two cell types were determined by an unpaired “t” test with n = 4 for each cell type (two separate cultures, two replicate gels).
Data on cell growth were analyzed by one-way analy- sis of variance followed by a Newman-Keuls multiple comparison test. In the text, figures, and tables, only the most relevant differences are indicated, but their significance values are based on analyses in which all conditions were included for comparison.
Throughout the text, a P value less than 0.05 is con- sidered significant.
RESULTS As described in a previous study (Davies et al.,
1987b), the technique to obtain cells from the periph- eral lung gives two principal cell types in primary cul- ture. One is the endothelial cell, which initially pre- dominates, the other (PC) overgrows the endothelial cells in the primary culture itself or following passage. This can be demonstrated by incubation with di-acety- lated low density lipoprotein (LDL), which is taken up to a marked degree by endothelial cells, but much less by the PC. Also, the endothelial cells stain for von Willebrand factor, PC do not. Cells from the CC provide a morphologically homogenous population in primary culture. PC tend to spread more widely on the plastic substratum than do CC (Davies et al., 1987b).
Immunostaining for cytoskeletal proteins In PC in culture, positive fluorescence for a-smooth
muscle actin was localized along the complete length of stress fibers prominent in this cell type (Fig. 1). In CC, the staining was usually more discrete, localized along shorter lengths of less prominent fibers. Some cells, usually a t low density, exhibited a staining pattern like that of PC. Negative or nonimmune serum controls were consistently negative or had only a faint back- ground fluorescence.
Since the a-isoactin identified by antibody binding is a marker for smooth muscle and was consistently and strongly expressed, we wished to confirm that cells in vivo at sites of smooth muscle were correspondingly positive. We therefore performed immunostaining on unfixed, frozen sections of rat lung. The localization of a-smooth muscle actin was excellent with bright areas of discrete fluorescence and a low level of background staining. The fluorescence was localized in the medial layer of “muscular” pulmonary arteries and in the subendothelial PC within the elastic lamina of “non- muscular” vessels, both pre- and post-capillary (Fig. 2). Smooth muscle cells in the walls of bronchioli and in the septa1 knobs lining the alveolar ducts were also positive. No staining was evident within the intersti- tium of the lung, within endothelium, or in the suben- dothelium of capillaries. Nonimmune serum controls were negative.
2-D gel electrophoresis In each cell type, the duplicate gels from each culture
gave quantitatively similar results and the two cul- tures were also similar. However, differences were evi- dent between cell types (Figs. 3-51. Both cell types ex- pressed similar levels of a-smooth muscle actin (the most acidic isoform in the actin cluster, labelled “a” in Fig. 3). PC exhibited slightly higher levels of nonmus- cle actin isoforms than CC, but the difference failed to reach significance. However, another cytoskeletal pro-
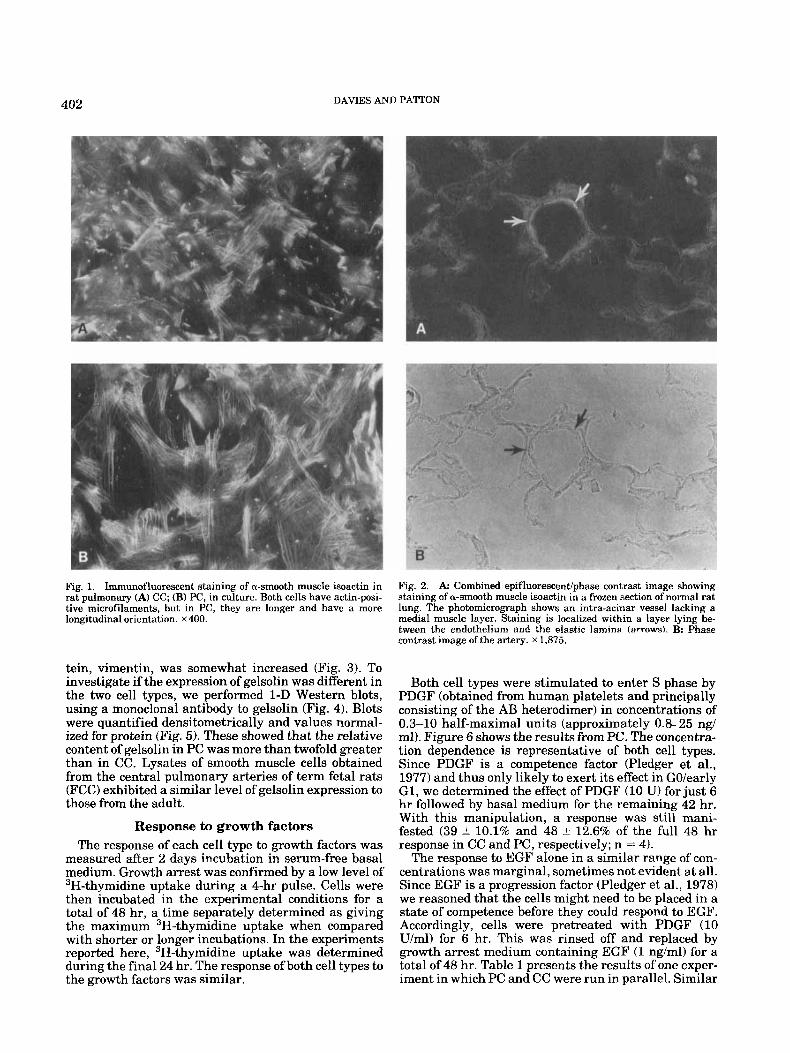
402 DAVIES AND PATTON
Fig. 1. Immunofluorescent staining of a-smooth muscle isoactin in rat pulmonary (A) CC; (B) PC, in culture. Both cells have actin-posi- tive microfilaments, but in PC, they are longer and have a more longitudinal orientation. x400.
Fig. 2. A Combined epifluorescent/phase contrast image showing staining of a-smooth muscle isoactin in a frozen section of normal rat lung. The photomicrograph show8 an intra-acinar vessel lacking a medial muscle layer. Staining is localized within a layer lying be- tween the endothelium and the elastic lamina (arrows\. B: Phase contrast image of the artery. x 1,875.
tein, vimentin, was somewhat increased (Fig. 3). To investigate if the expression of gelsolin was different in the two cell types, we performed 1-D Western blots, using a monoclonal antibody to gelsolin (Fig. 4). Blots were quantified densitometrically and values normal- ized for protein (Fig. 5). These showed that the relative content of gelsolin in PC was more than twofold greater than in CC. Lysates of smooth muscle cells obtained from the central pulmonary arteries of term fetal rats (FCC) exhibited a similar level of gelsolin expression to those from the adult.
Response to growth factors The response of each cell type to growth factors was
measured after 2 days incubation in serum-free basal medium. Growth arrest was confirmed by a low level of 3H-thymidine uptake during a 4-hr pulse. Cells were then incubated in the experimental conditions for a total of 48 hr, a time separately determined as giving the maximum 3H-thymidine uptake when compared with shorter or longer incubations. In the experiments reported here, 3H-thymidine uptake was determined during the final 24 hr. The response of both cell types to the growth factors was similar.
Both cell types were stimulated to enter S phase by PDGF (obtained from human platelets and principally consisting of the AB heterodimer) in concentrations of 0.3-10 half-maximal units (approximately 0.8-25 ngl ml). Figure 6 shows the results from PC. The concentra- tion dependence is representative of both cell types. Since PDGF is a competence factor (Pledger et al., 1977) and thus only likely to exert its effect in GO/early G1, we determined the effect of PDGF (10 U) for just 6 hr followed by basal medium for the remaining 42 hr. With this manipulation, a response was still mani- fested (39 k 10.1% and 48 ? 12.6% of the full 48 hr response in CC and PC, respectively; n = 4).
The response to EGF alone in a similar range of con- centrations was marginal, sometimes not evident at all. Since EGF is a progression factor (Pledger et al., 1978) we reasoned that the cells might need to be placed in a state of competence before they could respond to EGF. Accordingly, cells were pretreated with PDGF (10 U/ml) for 6 hr. This was rinsed off and replaced by growth arrest medium containing EGF (1 ng/ml) for a total of 48 hr. Table 1 presents the results of one exper- iment in which PC and CC were run in parallel. Similar
CENTRAL AND PERIPHERAL PULMONARY SMOOTH MUSCLE CELLS
15000 - 3H-TdR uptake loooo- (CPm)
5Ooo-
403
> 2.5-
B 2-t c bi
? i CELL TYPE
Fig. 3. Two-dimensional gel electrophoresis demonstrating the cy- toskeletal proteins in each cell type. A: CC; B PC. a, actin; a-a, a-ac- tin; t, tubulin; tm, tropomyosin; v, vimentin.
FC C c c P C Fig. 4. Western blot of cell lysates fractionated by 1-D gel electro- phoresis, using duplicate lanes for each cell type. The blots were stained with an antibody to gelsolin followed by an alkaline phos- phatase-conjugated secondary antibody.
results were obtained in a second experiment not shown. The sequential treatment of PDGF (6 hr) fol- lowed by EGF gave a thymidine uptake value signifi- cantly greater than to PDGF alone for 6 hr. Thus PDGF
Fig. 5. Histogram showing the relative optical densities of the gelso- lin band identified in Western blot. The values were normalized to total protein.
I . a
I
” / I I I 1
Basal .3 3 10 Medium
[PDGF] in units/ml
Fig. 6. 3H-thymidine (TdR) uptake of rat PC in response to stimula- tion with PDGF-A€? (mean 2 SEM of triplicate determinations in a representative experiment).
TABLE 1. Response to PDGF and EGF. alone and in combination’
CPm Treatment cc PC
PDGF 13,577 5 1,212* 9,318 2 602* PDGF (6 hr)+BM 1,936 f 178* 1,269 2 156* EGF 540?3 400 2 154 PDGF + EGF 15,221= -+ 1,275* 6,178 ? 584* PDGF (6 hr)--EGF 6,575 ? 369** 5,419 2 329** FBS (10%) 16,179 k 808* 19,739 -+ 181* BM alone 68 2 6 97 2 4
‘Mean 2 SEM. PDGF was added at a concentration of 10 U/ml for 24 hr alone or with EGF, or for 6 hr, and then replaced by other conditions as indicated hy an arrow; EGF was at 1 ngiml; BM, basal medium. *P<0.05, comparison with BM alone, * * P < 0.05, comparison with PDGF (6 hr) EGF. Newman-Keuls test.
primed the cells to respond to EGF. If PDGF and EGF were both present for the total 48-hr period, thymidine uptake was similar to that given by PDGF alone for 48 hr.
Although PDGF may act as a competence factor in these cells, it is clearly capable alone of stimulating passage through G1 into S. To determine if this action requires autocrine activity, we blocked receptor bind-

DAVIES AND PATTON 404
TABLE 2. Response to PDGF effect of suramin’ TABLE 3. Response to IGF-1: receptor specificity and the effect of IT’
CPm Treatment cc PC
PDGF (6 hr)--BM 14,943 f 1,741 9,708 t 426 PDGF (6 hr)-tBM + suramin 191 t 19* 774 5 127* PDGF (6 hr)+
BM + suramin + 10% FBS ND2 17,349 f 529** BM + suramin alone 312 k 184 718 ? 79 BM alone 178 k 16* 1,591 t 146* BM + 10% FBS 46,137 f 2,180* 40,717 t 648%
‘Mean i- SEM of triplicate determinations. PDGF was added at a concentration of 10 U / d , suramin at 100 F M BM, basal medium. * P i 0.05, comparison with PDGF (6 br)-CBM, ‘*P < 0.05, comparison with BM t suramin alone, Newman-Keuls test. 4 n another experiment (not presented because of an unrepresentatively poor response to PDGF), BM + suramin + 10% FBS gave 3H-TdR uptake values 36% those of BM + 10% FBS.
3000 4000 1 . ‘OF-’ i 1 0 / I I I
Basal 0.1 1 .o 10 Medium
[IGF-11 in ng/ml
Fig. 7. 3H-thymidine uptake of rat PC in response to stimulation with IGF-1 (mean % SEM of triplicate determinations in one experi- ment). As explained in the text, the response was sometimes maximal at 1 ng/ml.
ing of a putative factor with the drug suramin (Table 2). Cells were incubated with PDGF (10 U/ml) for 6 hr, rinsed thoroughly, and incubated in growth arrest me- dium for the remaining 42 hr. While a 6-hr incubation with PDGF stimulated entry into S phase, suramin (100 FM) blocked this, indicating that an additional autocrine factor is required for PDGF to stimulate growth. This effect was noted for CC (n = 4) and PC (n = 3). Suramin was not cytotoxic: cultures incubated with the drug plus 10% FBS underwent DNA synthesis, although counts were not as high as with 10% FBS alone (43% in Table 2).
IGF-1, like EGF, is also a progression factor and is required for Balb/c 3T3 cells to pass through later stages of G1(26), but in both of the rat pulmonary cells it stimulated thymidine uptake in the absence of serum competence factors (Fig. 7). The degree of response var- ied, however, and was sometimes maximal a t 1 ngiml. Since IGF-1 binds to the insulin receptor, albeit with lower affinity (Sara and Hall, 1990), the receptor speci- ficity was determined by adding monoclonal antibody against the IGF-1 receptor. At 10 ng/ml, it blocked IGF-1 stimulation (Table 3); a t 100 ng/ml some stimu- lation was occasionally noted. To identify a transduc- tion mechanism previously linked to IGF-receptor
cum Treatment cc PC
IGF-1 1,023 f 3 1,077 2 63 IGF-1 + R.Ab 229 k 31* 439 k 117* PT+ IGF-1 313 * 58* 478 t 22* BM alone 261 _i 38* 395 5 6* BM + R.Ab alone 163 2 21* 347 t 16* BM + PT alone 150 2 35* 256 k 10*
’Mean 2 SEM of triplicate determinations. IGF-1 was added at a concentration of 1 ngiml; R.Ab, the IGF-1 receptor antibody, was at 10 ngiml; PT was incubated with the cells for 3 hr at 10 ngiml before adding IGF-1; BM, basal medium. *P < 0.05, comparison with IGF-1, Newman-Keuls test.
binding, we pretreated growth-arrested cells with per- tussis toxin (PT) for 3 hr before addition of the growth factor. PT blocked IGF-1-stimulated DNA synthesis.
When cells were treated with IGF-1 alone (at 1 ngl ml) for 6 hr, rinsed, and placed in basal medium for a total of 48 hr, DNA synthesis was not increased above basal levels (results not shown), suggesting that IGF-1 needs to be present during a post-competence progres- sion phase. However, its action as a progression factor might be potentiated by pretreating the cells with a competence factor like PDGF. Cells were therefore in- cubated in PDGF (10 U/ml) for 6 hr, rinsed, and then treated with IGF-1 (1 ng/ml) for a total of 48 hr. In contrast to the results obtained with PDGF and EGF, this sequential addition of PDGF and IGF-1 did not potentiate the response. Since PDGF pretreatment might speed up the rate of entry into S phase, we com- pared its effects of 24 hr and 48 hr, but again, no in- crease in the response above that given by PDGF alone was found at the earlier time (results not shown).
DISCUSSION The fact that PC stain positively for a-smooth muscle
actin indicates that they exhibit some smooth muscle- specific characteristics (Skalli et al., 1989). This was the conclusion reached in a previous study that showed that PC and CC in culture stain positively for smooth muscle and nonmuscle myosin (Davies et al., 1987b). The distribution of a-smooth muscle actin in the lung also supports this idea since it was confined to the smooth muscle layer of pulmonary vessels and airways and was absent from the interstitium. Kapanci et al. (19901, using an antibody that was also generated against the unique sequence at the amino terminal of the actin molecule, demonstrated a similar distribution in normal rat and human lungs. Interstitial staining, presumably expressed by myofibroblasts, was present only in fibrosis and “post-capillary” PH. Capillary PC are sparse in the rat lung (Weibel, 1974) and could not be detected in our study. The fact that PC are different from CC is indicated by their morphology and pattern of a-isoactin immunostaining. The rat PC resemble bo- vine retinal PC in that actin stress fibers are pro- nounced and in immunostained preparations are typi- cally long with a distinct polar orientation (Patton, personal observation). PC and microvascular smooth muscle cells from other tissues have been reported to stain selectively with a monoclonal antibody (3G5) that specifically recognizes a membrane ganglioside (Nayak et al., 1988). A small proportion of the cells from the rat

405 CENTRAL AND PERIPHERAL. PULMONARY SMOOTH MUSCLE CELLS
lung stain positively in the PC primary culture, but staining is lost in later passages. However, parallel studies on the distribution of 3G5 staining in vivo indi- cate that although it is largely confined to the mi- crovasculature, it is most prevalent in cells that lie outside the elastic lamina in contrast to the a-actin- positive cells that lie between this structural boundary and the endothelium (Davies, personal observations). The reason for this is unknown, but given this distribu- tion, 3G5 would not appear to be an unequivocal marker of PC and intermediate cells in the lung, both of which are intimately associated with the endothelium (Weibel, 1974; Davies et al., 1986). Furthermore, since the 2-D gel electrophoresis demonstrates that rat CC express nonmuscle actin, this characteristic does not provide the discrimination between large vessel and microvascular cells as has been shown by immunohis- tochemical staining to distinguish bovine aortic smooth muscle cells and retinal PC (Herman and D’Amore, 1985). However, since species, tissue of origin, and methodology are all different in the current instance, other explanations are obviously possible. Differences between the two cell types are evident in the higher levels of vimentin and gelsolin expressed by PC. The abundance of the latter protein is likely to have major functional implications. Gelsolin is involved in the po- lymerization and depolymerization of actin (Yin et al., 1980), but it has also been shown to increase the activ- ity of actin-activated myosin ATPase in the presence of tropomyosin (Ebisawa and Nonomura, 1985; Onji et al., 1988). Higher levels may therefore suggest a greater contractile or migratory capability for the PC. Absence of cytokeratins in the PC cultures indicates little or no contamination by pulmonary microvascular endothe- lial cells (Patton et al., 1990b).
The possibility exists that differences between the cells could be due to the different methods used to ob- tain them. Thus, CC migrate from explants of the pul- monary artery whereas PC are obtained by a low con- centration collagenase digestion. The possibility that CC may represent a more migratory cell, however, is at odds with their lower content of gelsolin. However, the fact that they migrate from an area closest to the in- tima may make them rather more similar to PC than cells deeper in the medial layer. It is becoming appar- ent that the medial layer of CC is a heterogenous mix- ture of subpopulations that may have widely different functions and responses (Frid et al., 1993). Certainly, the differences in the preparative technique used in the present study to obtain the two cell types was not re- flected in their response to growth factors. In another study, similarities were also found in the two cell types with respect to their growth response to serotonin, and the expressions of serotonin receptor subtypes and sero- tonin transporter (Pitt et al., 1993).
Growth stimulation by PDGF is characteristic of mesenchymal cells, but requires the presence of pro- gression factors present in plasma (Pledger et al., 1977; Clemmons, 1985). Our experiments showed that these were not necessary. The fact that the cells are stimu- lated to enter S phase by PDGF in the absence of exog- enous progression factors suggests that they are sup- plied endogenously. This conclusion is reinforced by the results of experiments utilizing the drug suramin. This
is a polysulfonated naphthylurea used in the therapy of trypanosomiasis and onchocerciasis. It noncompeti- tively inhibits binding of exogenous PDGF (Hosang, 19851, EGF (Coffey et al., 19871, IGF-1 (Pollack and Richard, 19901, and transforming growth factor beta (TGFP; Beauchamp et al., 1992) to their receptors and prevents transformation by v-sis, presumably by inhib- iting binding of the endogenously produced sis protein to its receptor (Betsholtz et al., 1986). While sugges- tions have been made that like other polyanionic com- pounds it inhibits protein kinase C (Powis, 1991), this has not yet been conclusively demonstrated. Its ability in the present study to supress PDGF-stimulated DNA synthesis suggests the presence of an autocrine path- way.
In stimulating entry into S phase, exogenous IGF-1 acts through its own receptor. The fact that its stimula- tory action occurs in the absence of serum suggests that an endogenous competence factor is present or induc- ible. Under quiescent conditions, both cell types ex- press TGFPl mRNA and release TGFp protein (Kubiak et al., 1991). Since the protein is mainly in the latent form, however, it is not clear if it is likely to have autocrine effects under the conditions tested.
The action of PT in the current study is consistent with that in other cells in which it has been shown t o perturb the early effects of IGF-receptor binding (Ko- jima et al., 1990). The simple interpretation of our data is that PT ribosylates a Gp protein linked to the IGF-1 receptor. Alternatively, the GP pTotein may be linked to other receptors that bind autocrine factors necessary to establish competence.
ACKNOWLEDGMENTS The authors wish to acknowledge Donna Kunkel who
was responsible for maintaining cell cultures and Jay Spence who performed the immunostaining. Supported by NHLBI grant HL 41811.
LITERATURE CITED Beauchamp, R.D., Sheng, H.-M., Bascom, C.C., Miller, D.A., Lyons,
R.M., Torre-Amione, G., and Moses, H.L. (1992) Phenotypic alter- ations in fibroblasts and fibrosarcoma cells that overexpress latent transforming growth factor-pl. Endocrinology, 130.24762486.
Betsholtz, C., Johnsson, A,, Heldin, C.-H., and Westermark, B. (1986) Efficient reversion of simian sarcoma virus-transformation and in- hibition of growth factor-induced mitogenesis by suramin. Proc. Natl. Acad. Sci. USA, 83:644&6444.
Chung-Welch, N., Patton, W., Yen-Patton, G., Hechtman, H., and Shepro, D. (1989) Phenotypic comparison between mesothelial and microvascular endothelial cell lineages using conventional endothe- lial cell markers, cytoskeletal protein markers and in vitro assays of angiogenic potential. Differentiation, 42.4453.
Clemmons, D.R. (1985) Variables controlling the secretion of a so- matomedin-like peptide by cultured porcine smooth muscle cells. Circ. Res., 56.41H26.
Coffey, R.J., Leof, E.B., Shipley, G.D., et al. (1987) Suramin inhibition of growth factor receptor binding and mitogenicity in AKR-PB cells. J . Cell. Physiol., 132:143-148.
Davies, P., Burke, B., and Reid, L. (1986) The structure of the wall of the rat intraacinar artery: An electron microscopic study of micro- dissected preparations. Microvasc. Res., 32:50-63.
Davies, P., Reid, L., Jones, R., and Schloo, B. (1987a) Endothelium of the pulmonary vasculature in health and disease. In: Pulmonary Endothelium in Health and Disease. U. Ryan, ed. Marcel Dekker, New York, pp. 375-445.
Davies, P., Smith, B.T., Maddalo, F.B., Lanaleben, D.. Tobias. D.. Fujiwara, K., and Reid, L. (198713) Characteristics of lung pericytes in culture including their growth inhibition by endothelial sub- strate. Microvasc. Res., 33:300-314.

DAVIES AND PATTON 406
Patton, W., Pluskal, M., Skea, W., Beucker, J., Lopez, M., Zimmer- Tion of actin gelation. J . Bgl. Chem., 25539494-9500.
Dodge, A,, Patton, W., Yoon, M., Hechtman, H., and Shepro, D. (1991) Organ and species specific differences in cytoskeletal protein pro- files of cultured microvascular endothelial cells. Comp. Biochem. ...._ .~
Physiol., 98B:461470. Ebisawa. K., and Nonomura, Y. (1985) Enhancement of actin-acti-
vated myosin ATPase by an 84K Mr actin-binding protein in verte- brate smooth muscle. J. Biochim. (Tokyo), 9831127-1130.
Frid, M.G., Mecham, R.P., Moiseeva, E.O., and Stenmark, K.R. (1993) Identification of phenotypically distinct smooth muscle cell popula- tions in mature and developing pulmonary arteries. Am. Rev. Respir. Dis., 174:A493.
Gitlin, J.D., and D'Amore, P.A. (1979) Culture of retinal capillary cells using selective growth media. Microvasc. Res., 26374-80.
Herman, I.M., and DAmore, P.A. (1985) Microvascular pericytes con- tain muscle and non-muscle actins. J . Cell Biol., 101:4%52.
Hosang, M. (1985) Suramin binds to platelet-derived growth factor and inhbits biological activity. J. Cell. Biochem., 29:265-273.
Kapanci, Y., Burgan, S., Pietra, G.G., Conne, B., and Gabbiani, G. (1990) Modulation of actin isoform expression in alveolar myofibro- blasts (contractile interstitial cells) during pulmonary hyperten- sion. Am. J . Pathol., 136:8814389.
Kojima, I., Kitaoka, M., and Ogata, E. (1990) Studies on the cell-cycle dependency of the actions of insulin-like growth factor-I in Balb/c 3T3 cells. J . Cell. Physiol., 143:529433.
Kubiak, J.S., Pitt, B.R., and Davies, P. (1991) Synthesis and secretion of transforming growth factor p (TGF-P) by rat pulmonary artery smooth muscle cells in vitro. Am. Rev. Respir. Dis., 143:A374.
Laemmli, U. (1970) Cleavage of structural proteins during the assem- bly of the head of bacteriophage T4. Nature, 227:680-685.
Meyrick, B., and Reid, L. (1979) Hypoxia and incorporation of 'H- thvmidine bv cells of the rat uulmonarv arterv and alveolar wall. A&. J. Pathil., 96:51-70. A
Morrissey, J.H. (1981) Silver stain for proteins in polyacrylamide gels: A modified urocedure with enhanced uniform sensitivity. Anal. Bio- chem., 117:i07310.
Nayak, R.C., Berman, A.B., George, K.L., Eisenbarth, G.S., and King, G.L. (1988) A monoclonal antibody (3G5)-defined ganglioside anti- gen is expressed on the cell surface of microvascular pericytes. J . Exp. Med., 167:1003-1015.
Nemecek, G.M., Coughlin, S.R., Handley, D.A., and Moskowitz, M.A. (1986) Stimulation of aortic smooth muscle cell mitogenesis by sero- tonin. Proc. Natl. Acad. Sci. USA, 83:674-678.
OFarrell, P. (1975) High resolution two dimensional electrophoresis. J. Biol. Chem., 250:40074021.
Onji, T., Takagi, M., and Shibata, N. (1988) Gelsolin is Ca2+-sensitive regulator of actomyosin system in platelet. Biochem. Biophys. Res. Commun., 155:91-99.
Patton, W., Dhanak, M., and Jacobson, B. (1989) Identification of DictyosteZirtrn discoideun plasma membrane proteins by cell surface labeling and quantitative two-dimensional gel electrophoresis. Anal. Biochem.. 179:37-49.
mann, R., Belanger, L., and Hatch, P. (1990a) Development of a dedicated two-dimensional gel electrophoresis system that provides optimal pattern reproducibility and polypeptide resolution. Biotech- niques, 8.518-527.
Patton, W., Yoon, M., Alexander, S., Chung-Welch, N., Hechtman, H., and Shepro, D. (1990b) Expression of simple epithelial cytokeratins in bovine pulmonary microvascular endothelial cells. J . Cell. Phys- iol., 143:140-149.
Pitt, B.R., Weng, W., Steve R., Blakely, R.D., Reynolds, I., and Davies, P. (1993) Serotonin increases DNA synthesis in rat proximal and distal pulmonary vascular smooth muscle cells in culture. Am. J. Physiol. (in press).
Pledger, W.J., Stiles, C.D., Antoniades, H.N., and Scher, C.D. (1977) Induction of DNA synthesis in BALBlc-3T3 cells by serum compo- nents: Reevaluation of the commitment process. Proc. Natl. Acad. Sci. USA, 74:4481485.
Pledger, W.J., Stiles, C.D., Antoniades, H.N., and Scher, C.D. (1978) An ordered sequence of events is required before BALBlc-3T3 cells become committed to DNA synthesis. Proc. Natl. Acad. Sci. USA, 75:283%2843.
Pollack, M., and Richard, M. (1990) Suramin blockade of insulinlike growth factor-I-stimulated proliferation of human osteosarcoma cells. J . Natl. Cancer Inst., 82:1349-1352.
Powis, G. (1991) Signalling targets for anticancer drug development. Trends Pharmacol. Sci., 12:18%194.
Raines, E.W., and Ross, R. (1982) Platelet-derived growth factor. I. High yield purification and evidence for multiple forms. J . Biol. Chem., 257r515P5160.
Reid, L.M. (1990) Vascular remodeling. In: The Pulmonary Circula- tion: Normal and Abnormal. A.P. Fishman, ed. University of Penn- sylvania Press, Philadelphia, pp. 259-282.
Sara, V.K., and Hall, K. (1990) Insulin-like growth factors and their binding proteins. Physiol. Rev., 70:591414.
Skalli, O., Pelte, M.-F., Peclet, M.-C., Gabbiani, G., Gugliotta, P., Bussolai, G., Ravalezza, M., and Orci, L. (1989) Alpha smooth mus- cle actin, a differentiation marker of smooth muscle cells, is present in microfilamentous bundles of pericytes. J. Histochem. Cytochem., 373315321.
Towbin, T., Staehelin, and Gordon, J . (1979) Electrophoretic transfer of protein from polyacrylamide gels to nitrocellulose sheets: Proce- dure and some applications. Roc. Natl. Acad. Sci. USA, 76:435& 4354.
Verhoeven, D., and Buyssens, N. (1988) Desmin immunoreactivity in precapillary cells of rat. J . Histochem. Cytochem., 36t1079-1080.
Weibel, E.R. (1974) On pericytes, particularly their existence on lung capillaries. Microvasc. Res., 8:21%235.
Weinstein, R., Stermerman, M.B., and Maciag, T. (1981) Hormonal requirements for growth of arterial smooth muscle cells in vitro: An endocrine approach to atherosclerosis. Science, 212:818-820.
Yin, H.L., Zaner, K.S., and Stossel, T.P. (1980) Ca" control of actin eelation. Interaction with eelsolin with actin filaments and r e d a -