pantoea sp. s3w-11 antibacterial activities of strain
TRANSCRIPT

Korean Society for Biotechnology and Bioengineering Journal 35(1): 64-71 (2020)http://dx.doi.org/10.7841/ksbbj.2020.35.1.64 ISSN 1225-7117 / eISSN 2288-8268
Pantoea sp. S3W-11의 항균활성과 배양 최적화
남영호, 최아영, 정유진*
Antibacterial Activities of Strain Pantoea sp. S3W-11 and
Optimization of Culture Conditions
Young Ho Nam, Ahyoung Choi and Eu Jin Chung*
Received: 13 October 2019 / Revised: 7 January 2020 / Accepted: 18 March 2020
© 2020 The Korean Society for Biotechnology and Bioengineering
Abstract: In this study, 46 bacteria strains were isolated from
water and soil collected from Geumjin-ri, Ganggu-ri and
Namseock-ri in Yeongdeok-Gun to screen antibacterial bacte-
ria against various pathogenic bacteria. Among them, strain
S3W-11 showed an excellent growth inhibition against methi-
cillin-resistant Staphylococcus aureus, Bacillus cereus and
Escherichia coli. As a result of the 16S rRNA gene sequence
analysis, strain S3W-11 showed the high similarity with Pan-
toea vagans LMG 24199T, Pantoea agglomerans DSM 3493T,
Pantoea conspicua LMG 24534T 99.64%, 99.58%, 99.18%,
respectively. The effects of culture medium composition, tem-
perature, and initial pH on the cell growth and antibacterial
activity were confirmed for culture optimization of strain S3W-
11. Optimal conditions for growth and antibacterial activity in
strain S3W-11 were found to be: YPD medium, 25oC and pH
6.5. When the strain was cultured in LB, NB, TSB, and R2A
media, the antibacterial activity has not been confirmed. Opti-
mal reaction time was 48 h in YPD medium, 100 rpm and 0.3
vvm in 2 L-scale fed-batch fermentation process for antimi-
crobial activity. Strain S3W-11 can be improved on antibacte-
rial activity in optimal culture conditions (YPD medium, 25oC
and pH 6.5). Therefore, the antibacterial activity of Pantoea
sp. S3W-11 had potential as antibiotics for pathogens includ-
ing MRSA.
Keywords: Antibacterial activity, Pantoea sp. S3W-11, methi-
cillin-resistant Staphylococcus aureus (MRSA)
1. INTRODUCTION
인류 최초의 항생제는 페니실린 (Pennicillin)이다. 1943년 페
니실린이 화학적 합성이 가능하게 되어 인류의 생명을 위협
하는 세균감염의 치료에 널리 사용되게 되었다 [1]. 그러나,
얼마 지나지 않아 1940년에 이미 페니실린에 내성을 가진 포
도알구균 (penicillin-R Staphylococcus)이 등장했다. 그 이후,
1960년 메티실린, 1972년 반코마이신, 1998년 이미페넴,
2003년 뎁토마이신 등 여러 항생제가 개발되었지만, 이들에
대한 내성도 잇따랐다. 2016년 영국정부가 발표한 「항생제
내성 보고서」에는 현재 전 세계적으로 70만 이상이 내성 박
테리아의 감염에 의해 숨지고 있으며, 신세대 항생제가 개발
되지 않는다면 2050년에는 1000만 이상의 사망과 100조 달
러의 경제적 손실이 초래될 것이라고 예측하였다 [2].
항생물질이란 다른 세균이나 미생물의 발육과 번식을 억
제하는 물질을 말하며 이러한 물질로 만든 약을 항생제
(antibiotics)라고 하며, 좀 더 광범위한 의미의 항균제 (antimi-
crobials)라는 용어가 함께 사용되고 있다. 그 표적 대상이 세
균일 때 항세균제, 바이러스일 때 항바이러스제, 곰팡이일
때 항진균제라고 부른다. 일반적으로 세균을 죽이는 항세균
제를 항생제라고 한다 [3]. 미생물은 이러한 항생물질을 생
산하는 많은 종들이 널리 알려져 있다. Burkholderia 속
(genus)은 이차 대사산물을 생산하는 길항미생물로 알려져
있고, 식물생장촉진 효과, 항진균 효과가 있다 [4-7]. 바실러
국립낙동강생물자원관 미생물연구실Microbial Research Department, Nakdonggang National Institute ofBiological Resources (NNIBR), Sangju 37242, KoreaTel: +82-54-530-0870, Fax: +82-54-530-0879,e-mail: [email protected]
Research Paper

Pantoea sp. S3W-11의 항균활성과 배양 최적화 65
스 (Bacillus) 종은 자연계 널리 분포하면서 범위가 넓은 항균
물질을 생성하여 병원성 미생물을 억제하는 데 중요한 역할
을 하는 것으로 알려져 있다. 바실러스가 생산하는 2차 대사
산물은 항균범위가 넓고 강력한 항균효과를 가지고 있다. 또
한, 포자를 형성하여 복잡한 자연환경 조건에 저항력 있게
살아남을 수 있다 [8]. 브레비바실러스 (Brevibacillus)는 1996년
바실러스 종에서 재분류된 독립적인 속으로 chitinase와
gramicin S같은 2차대사산물을 생산하고, 그람양성 및 음성
세균 모두에 활성을 가지고 있는 항균제로 현재도 연구가치
가 높은 종이다 [9-11].
이러한, 항균물질을 생산할 수 있는 미생물은 항생제, 천연
식물보호제, 사료첨가제 등으로 이용이 가능하며, 미생물 산
업이 각광을 받으면서 미생물들의 유용물질 생산성을 높일
수 있는 배양조건, 정제공정 등의 최적화 연구의 중요성이
대두되었다. 이전의 연구결과에 따르면 환경요인, pH, 온도,
공기량 등의 배양조건이 미생물의 세포생장성과 항생물질
생산에 영향을 줄 수 있다 [12,13].
특히, 본 연구에서 분리한 S3W-11균주와 근연종인 Pantoea
vagans와 Pantoea agglomerans는 Erwinia amylovora에 의해
발생하는 배나무화상병을 제어하는 생물학적 방제제 (bio-
control agent)의 미생물 소재로 사용된다. P. vagans C9-1은
미국과 캐나다에 등록된 상용화된 약제인 BlightBan C9-1S
(NuFarms America)로 개발되었고, 두 종의 P. agglomerans
(E325, P10c)도 배나무화상병 방제제로 상용화되었다. P.
vagans C9-1는 이차대사산물로 pantocin A를 생산하며 관련
유전자 그룹이 이전연구에서 밝혀진 바 있다 [14]. 이러한 바
이오 소재 개발 시 가장 중요한 요건은 미생물을 활용하였을
때 그 효과와 경제성 확보에 있다. 미생물의 상용배지 선정
과정을 통해 미생물이 항균효과를 극대화 할 수 있는 상용배
지를 탐색 한 뒤 선정된 배지 조성 정보를 바탕으로 저렴한
대량배양용 배지 제작으로 경제성 확보에 기여할 수 있다.
또한, 미생물의 배양온도, 시간 등의 배양 최적화 과정을 통
하여 대량배양 용량 대비 효과 증대로 미생물 제제 제조시에
도 효율적인 면에서 긍정적인 역할을 할 수 있다. 이에 본 연
구에서는 유해미생물에 항균 활성을 보유한 미생물 Pantoea
sp. S3W-11을 확보하였다. 이 균주의 배지, 배양 온도, 초기
pH별 생장과 활성 평가를 통해 최적 배양조건을 선정하고자
하였다. 유용미생물의 배양최적화를 통해 항균 활성 물질의
생산을 극대화하여 이 미생물을 이용한 활용도를 증대시키
고자 하였다.
2. MATERIALS AND METHODS
2.1. 미생물 분리 및 동정
본 실험을 위하여 경북 영덕군 금진리, 강구리, 남석리 인근
3지점에서 물시료 및 토양시료를 채취하였다. 수질환경은
멀티수질측정기인 YSI (556MPS)를 이용하여 측정하였다
(Table 1). 채취한 시료는 0.85%(w/v) NaCl 용액에 연속 희석
하여 Reasoner’s2A agar (R2A, Difco) 고체배지에 도말한 후,
25oC에서 3일간 배양하였다. 균체의 다양한 형태적 특성에
따라 각각 분리하고, 계대배양 과정을 거쳐 순수 분리하였다.
유전자 분석서비스는 Macrogen에 의뢰하여 진행하였다. 미
생물의 핵산은 Qiagen DNeasy Tissue Kit를 이용하여 추출하
였고, 16S rRNA 유전자의 증폭은 universal primer (27F, 5’-
AGAGTTTGATCMTGGCTCAG-3’와 1492R, 5’-TACGGYTA
CCTTGTTACGACTT-3’)를 사용하였다. EzBioCloud [15]를
사용하여 연관성이 높은 유전자 서열을 획득, 비교 및 분석
을 진행하였다.
2.2. 항균활성보유 균주 탐색
순수분리한 미생물 중 항균활성 보유 미생물을 탐색하기 위
해 3종의 표적미생물을 사용하였고, 항균실험을 위한 각 미생
물들은 한국미생물보존센터 (KCCM), 생물자원센터 (KCTC)
에서 methicillin-resistant Staphylococcus aureus subsp. aureus
KCCM 40510, Bacillus cereus KCTC 3624, Escherichia coli
KCTC 2443을 분양받았다. 표적 미생물을 30oC, 24시간 액체
배양 한 뒤, 배양액을 분광광도계 (Novaspec spectrophotometer,
Biochrom, Cambridge, UK)를 이용하여 OD600값 0.4~0.5로 희
석 후 100 µl를 고체배지에 도말하였다. 그 후 순수분리한 균
주를 도말배지 위에 streaking 한 후 28oC, 24시간 정치배양
후 주변의 억제지대 (clear zone) 관찰을 통해 1차로 항균활성
Table 1. Location information
Location
Water
Temperature
(℃)
Electrical
conductivity
(µS/cm)
DO
(%)
DO
(mg/L)pH
Sal
(%)Latitude/Longitude
S1
Geumjin-ri,
Ganggu-myeon,
Yeongdeok-gun
11.4 490.99 126.9 11.34 8.0 31.97 36°22'04.1"N 129°23'42.3"E
S2
Ganggu-ri,
Ganggu-myeon,
Yeongdeok-gun
12.6 181.0 104.3 11.02 6.88 0.92 36°21'44.1"N 129°22'56.1"E
S3
Namseok-ri,
Yeongdeok-eup,
Yeongdeok-gun
12.7 159.2 108.5 11.50 7.0 0.08 36°24'41.3"N 129°21'51.0"E

66 Korean Society for Biotechnology and Bioengineering Journal 35(1): 64-71 (2020)
보유 미생물을 확인하였다. 선발된 미생물을 30oC, 48시간,
130 rpm으로 진탕배양한 후, 배양상등액을 필터 (0.22 µm 크
기)로 여과하였다. 배양여액 400 µl를 종이디스크 (paper
disc)에 올려 완전건조 후 표적미생물이 도말된 고체배지에
올려 28oC, 24시간 정치배양 후 억제지대 (inhibition zone)를
확인하였다.
2.3. 계통도 작성
항균활성 탐색 후, 최종 선정된 미생물의 계통도를 작성하였
다. Multiple alignment는 CLUSTAL X program을 사용하여
수행하였다 [16]. 그 이후에 BioEdit program [17]을 사용하
여 염기서열들을 정렬하고, 계통수는 MEGA 7 program [18]
에서 제공하는 neighbor-joining [19], maximum-likelihood
Table 2. Strains obtained from Yeongdeok-gun province
No. Strains Isolation source Closest type strain (accession number, similarity) Location
1 S1S-1 Soil Bacillus funiculus (AB049195, 99.93%)
Geumjin-ri
2 S1S-2 Soil Rouxiella silvae (KX784908, 99.56%)
3 S1S-3 Soil Bacillus megaterium (CP009920.1, 99.93%)
4 S1S-4 Soil Bacillus altitudinis (ASJC01000029, 100%)
5 S1S-5 Soil Bacillus velezensis (AY603658, 98.8%)
6 S1S-7 Soil Bacillus thuringiensis (NR_024928.1, 99.93%)
7 S1S-8 Soil Bacillus amyloliquefaciens (FN597644.1, 99.8%)
8 S1S-10 Soil Bacillus mobilis (KJ812449, 100%)
9 S1S-11 Soil Paenarthrobacter nitroguajacolicus (AJ512504, 99.93%)
10 S2S-1 Soil Bacillus asahii (NR_024817.1, 99.84%)
Ganggu-ri
11 S2S-2 Soil Psychrobacter piscatorii (NR_112807.1, 100%)
12 S2S-3 Soil Shewanella baltica (NR_025267.1, 98.7%)
13 S2S-4 Soil Paenibacillus lupini (KF769449, 99.39%)
14 S2S-5 Soil Streptomyces lannensis (NR_113181.1, 100%)
15 S2S-6 Soil Oerskovia turbata (NR_029279.1, 99.17%)
16 S2S-7 Soil Bacillus wiedmannii (KU198626.1, 99.93%)
17 S2S-8 Soil Raoultella ornithinolytica (NR_114736.1, 100%)
18 S2S-9 Soil Exiguobacterium sibiricum (NR_075006.1, 99.66%)
19 S2S-10 Soil Acinetobacter harbinensis (KC843488.1, 99.77%)
20 S2S-11 Soil Pseudomonas frederiksbergensis (NR_117177.1, 99.52%)
21 S2S-12 Soil Pseudomonas poae (AJ492829.1, 99.86%)
22 S2S-13 Soil Pedobacter koreensis (NR_113980.1, 99.93%)
23 S2S-14 Soil Pseudomonas poae (AJ492829.1, 99.86%)
24 S2W-1 Surface water Serratia glossinae (FJ790328, 99.86%)
25 S2W-2 Surface water Pseudomonas lurida (AJ581999, 99.66%)
26 S2W-3 Surface water Sphingomonas kyungheensis (NR_118263.1, 99.86%)
27 S2W-4 Surface water Pseudomonas peli (NR_042451.1, 99.93%)
28 S3S-3 Surface water Bacillus megaterium (CP009920.1, 99.93%)
Namseock-ri
29 S3S-4 Surface water Pseudomonas moorei (AM293566, 98.82%)
30 S3S-5 Surface water Bacillus mobilis (KJ812449, 99.93%)
31 S3S-6 Surface water Pseudomonas azotoformans (LT629702.1, 99.79%)
32 S3S-7 Surface water Pseudomonas frederiksbergensis (NR_117177.1, 99.52%)
33 S3S-8 Surface water Bacillus toyonensis (NR_121761.1, 99.93%)
34 S3S-9 Surface water Pantoea vagans (EF688012, 99.57%)
35 S3W-1 Surface water Pseudomonas baetica (FM201274, 99.59%)
36 S3W-2 Surface water Pseudomonas protegens (CP003190.1, 99%)
37 S3W-3 Surface water Aeromonas sobria (NR_037012.2, 99.86%)
38 S3W-4 Surface water Leptothrix cholodnii (ASJC01000029, 99.93%)
39 S3W-5 Surface water Exiguobacterium artemiae (AM072763, 99.86%)
40 S3W-6 Surface water Aeromonas salmonicida subsp. salmonicida (NR_043324.1, 99.86%)
41 S3W-7 Surface water Exiguobacterium undae (NR_043477.1, 99.8%)
42 S3W-8 Surface water Bacillus marisflavi (NR_025240.1, 100%)
43 S3W-9 Surface water Xanthomonas cynarae (AF208315, 100%)
44 S3W-10 Surface water Caulobacter flavus (AF208315, 99.15%)
45 S3W-11 Surface water Pantoea vagans (LMG 24199, 99.64%)
46 S3W-12 Surface water Micrococcus yunnanensis (FJ214355, 99.79%)

Pantoea sp. S3W-11의 항균활성과 배양 최적화 67
[20] 그리고 maximum-parsimony [21] algorithms을 이용하여
작성하였다. 순수 분리된 미생물의 16S rRNA 유전자의 상동
성은 EzBioCloud를 통해 계산하였다. Bootstrap values는
1,000 replicates를 만들어 계산하였다 [22].
2.4. 생장과 항균활성을 위한 배양조건 최적화
최종 선발한 항균미생물의 최적 배양조건 설정을 위해 배지,
온도, 초기 pH별 생육 및 항균활성을 조사하였다. 초기 배양
조건은 선발된 항균미생물을 종균 (seed culture)으로 3%(v/
v)로 접종하여, 28oC, 100 rpm으로 2일간 배양하였다. 배지는
LB, NA, TSB, YPD, R2A, Bennett`s의 상용화된 배지를 사용
하여 배지종류별 생육 및 항균활성을 확인하였다. 온도는
20oC, 25oC, 30oC, 35oC, 40oC에서 온도별로 배양하였으며,
pH는 5.5~8.5까지 0.5간격으로 배지의 초기 pH를 조절한 후
각각의 최종 배양물의 생육 및 항균활성을 확인하였다. 위의
배양조건 결과를 바탕으로 5 L fermenter를 이용한 시간대별
배양액의 항균활성 확인하여 가장 효율적으로 항균물질을
생산할 수 있는 배양시간을 선정하였다.
3. RESULTS AND DISCUSSION
3.1. 미생물 분리
본 연구를 통하여 2018년 3월 13일 영덕군 금진리, 강구리,
남석리 3개 지점에서 물과 주변 흙시료를 채취하였다. 금진
리, 강구리, 남석리의 염농도는 각각 31.92%, 0.92%, 0.08%
로 해수, 기수역, 일반하천 (경북 영덕군 강구면 오십천) 샘
플이었다. 3개 지점에서 총 46균주를 분리하였고, 16S rRNA
유전자 염기서열을 분석하여 동정하였다. 금진리의 흙 시료
에서 총 9주, 강구리에서는 총 18주, 남석리에서는 총 19주를
분리하였다 (Table 2).
3.2. 항균활성 보유 균주 탐색
순수분리된 46종의 균주를 대상으로 B. cereus와 methicillin-
resistant S. aureus에 대한 항균활성 보유 미생물을 탐색한 결
과, 획선평판법과 배양여액을 이용한 paper disc법 에서 활성
을 보유한 균주들을 확인하였다. S1S-2, S3W-2, S3W-11 총 3
종의 균주에서 항균활성을 확인하였고, 가장 억제거리가 큰
S3W-11균주를 최종적으로 선택하였다 (Fig. 1). 항균활성을
보유하고 Rouxiella silvae 와 99.56% 유사도를 보였던 S1S-2
균주는 배양 최적화 및 생리적 특성연구를 진행 중이다.
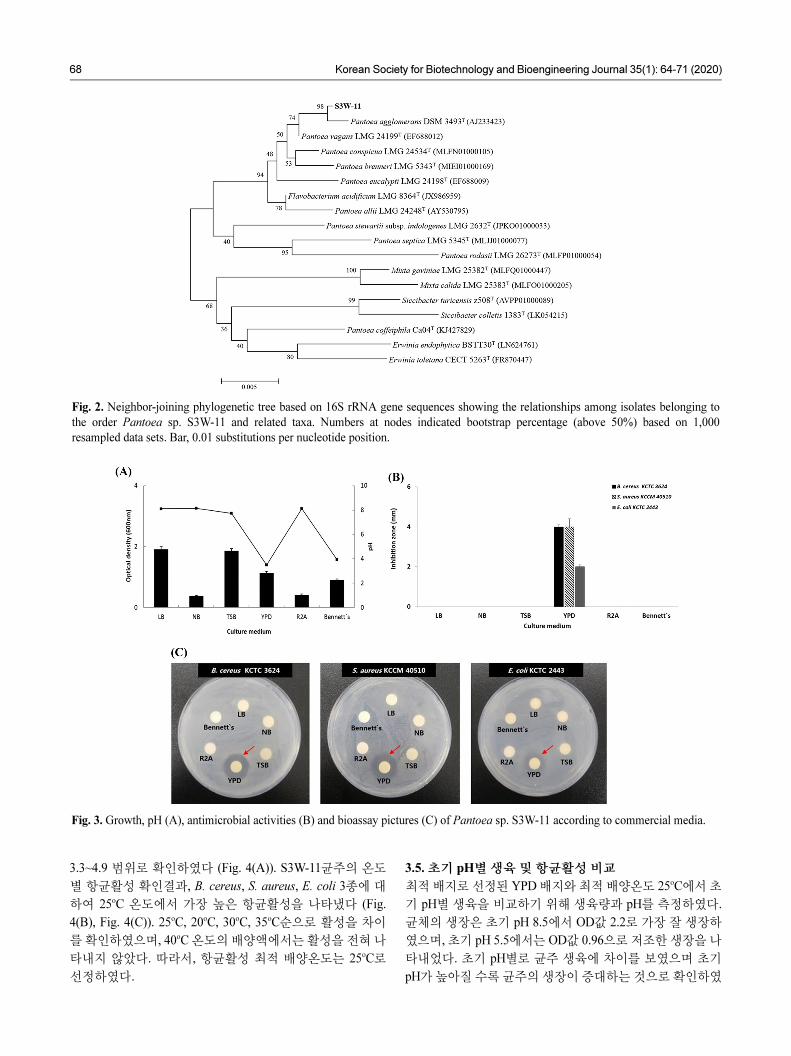
3.3. 계통도 작성
S3W-11균주는 근연종들과 16S rRNA 유전자 염기서열 분석
에 의한 계통분석을 진행하였다. 그 결과, S3W-11균주는
Pantoea vagans LMG 24199T, Pantoea agglomerans DSM
3493T, Pantoea conspicua LMG 24534T와 각각 99.64%,
99.58%, 99.18%의 상동성을 보였고 Pantoea 속 (genus)에 속
하는 종 (species)으로 확인하였다 (Fig. 2).
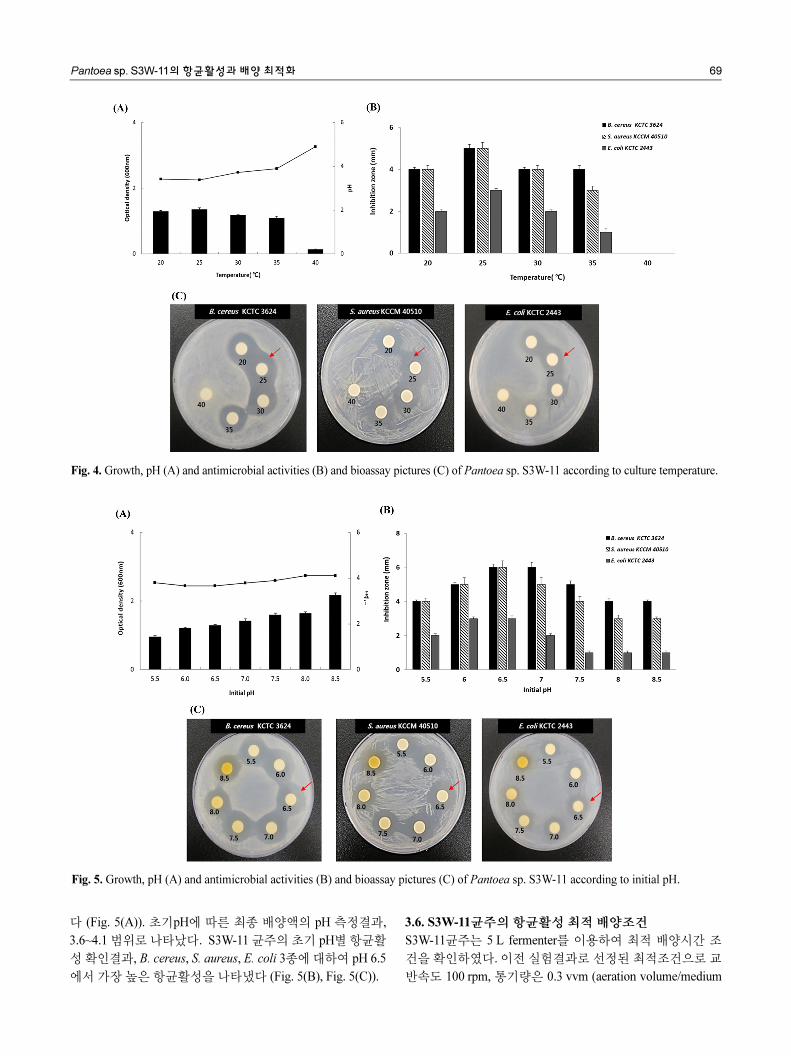
3.4. 상용 배지별 생육 및 항균활성 비교
S3W-11균주의 배양배지 최적화 연구를 위해 상용배지 LB,
NB, TSB, YPD, R2A, Bennett’s 배지에서 균주 생육량과 pH
를 측정하였다. 가장 높은 생육을 보인 배지는 LB, TSB배지
로 OD값이 각각 1.9, 1.8로 확인되었고, NB배지에서는 OD
값이 0.4로 가장 낮은 생육을 보였다 (Fig. 3(A)). S3W-11 균
주는 배지에 따라 생육의 차이가 많이 났으며 상용배지에 따
른 최종 배양액의 pH는 3.5~8.1 범위로 나타났다. S3W-11 균
주의 상용 배지별 항균활성 확인 결과, B. cereus, S. aureus,
E. coli 3종에 대하여 YPD배지에서 각각 4 mm, 4 mm, 2 mm
의 억제거리를 관찰하였다. 다른 상용배지 배양액에서는 활
성이 전혀 나타나지 않아 최적배지로 YPD배지를 선정하였
다 (Fig. 3(B), Fig. 3(C)). 항균활성을 보였던 유일한 배지는
YPD배지로 조성은 dextrose 2%, yeast extract 1%, peptone
2%이다. 이 결과로 S3W-11균주의 항균물질 생산에 적합한
배지의 조성과 함량을 확인하였다. S3W-11균주의 최적배지
는 YPD로 선정하였고 이후 진행되는 온도, 초기 pH별 생육
및 활성비교 실험은 YPD배지에서 배양하여 진행하였다.
3.5. 배양온도별 생육 및 항균활성 비교
최적 배지로 선정된 YPD 배지를 이용하여 온도별 생육을 비
교하기 위해 생육량과 pH를 측정하였다. 균체성장은 20~
35oC 범위에서 고르게 잘 생장하였으며, 각각의 온도별 조건
에서 가장 높은 생육을 보인 온도는 25oC로 OD값이 1.4였다.
배양온도 40oC에서는 가장 저조한 생육을 나타내었으며 OD
값이 0.1로 확인되었다. 온도에 따른 최종 배양액의 pH는
Fig. 1. Antimicrobial activity against B. cereus (A) and S. aureus
(B) of isolated strains from Yeongdeok-gun.
68 Korean Society for Biotechnology and Bioengineering Journal 35(1): 64-71 (2020)
3.3~4.9 범위로 확인하였다 (Fig. 4(A)). S3W-11균주의 온도
별 항균활성 확인결과, B. cereus, S. aureus, E. coli 3종에 대
하여 25oC 온도에서 가장 높은 항균활성을 나타냈다 (Fig.
4(B), Fig. 4(C)). 25oC, 20oC, 30oC, 35oC순으로 활성을 차이
를 확인하였으며, 40oC 온도의 배양액에서는 활성을 전혀 나
타내지 않았다. 따라서, 항균활성 최적 배양온도는 25oC로
선정하였다.
3.5. 초기 pH별 생육 및 항균활성 비교
최적 배지로 선정된 YPD 배지와 최적 배양온도 25oC에서 초
기 pH별 생육을 비교하기 위해 생육량과 pH를 측정하였다.
균체의 생장은 초기 pH 8.5에서 OD값 2.2로 가장 잘 생장하
였으며, 초기 pH 5.5에서는 OD값 0.96으로 저조한 생장을 나
타내었다. 초기 pH별로 균주 생육에 차이를 보였으며 초기
pH가 높아질 수록 균주의 생장이 증대하는 것으로 확인하였
Fig. 2. Neighbor-joining phylogenetic tree based on 16S rRNA gene sequences showing the relationships among isolates belonging to
the order Pantoea sp. S3W-11 and related taxa. Numbers at nodes indicated bootstrap percentage (above 50%) based on 1,000
resampled data sets. Bar, 0.01 substitutions per nucleotide position.
Fig. 3. Growth, pH (A), antimicrobial activities (B) and bioassay pictures (C) of Pantoea sp. S3W-11 according to commercial media.
Pantoea sp. S3W-11의 항균활성과 배양 최적화 69
다 (Fig. 5(A)). 초기pH에 따른 최종 배양액의 pH 측정결과,
3.6~4.1 범위로 나타났다. S3W-11 균주의 초기 pH별 항균활
성 확인결과, B. cereus, S. aureus, E. coli 3종에 대하여 pH 6.5
에서 가장 높은 항균활성을 나타냈다 (Fig. 5(B), Fig. 5(C)).
3.6. S3W-11균주의 항균활성 최적 배양조건
S3W-11균주는 5 L fermenter를 이용하여 최적 배양시간 조
건을 확인하였다. 이전 실험결과로 선정된 최적조건으로 교
반속도 100 rpm, 통기량은 0.3 vvm (aeration volume/medium
Fig. 4. Growth, pH (A) and antimicrobial activities (B) and bioassay pictures (C) of Pantoea sp. S3W-11 according to culture temperature.
Fig. 5. Growth, pH (A) and antimicrobial activities (B) and bioassay pictures (C) of Pantoea sp. S3W-11 according to initial pH.

70 Korean Society for Biotechnology and Bioengineering Journal 35(1): 64-71 (2020)
volume/minute) 조건하에 진행하였다. 생육패턴 확인 결과,
접종 후 배양 48시간에 정지기로 들어서는 것으로 확인되었
다 (Fig. 6). 시간별로 배양액을 회수하여 B. cereus, S. aureus,
E. coli 3종을 대상으로 배양시간별 항균활성을 확인하였다.
배양시간별 항균활성 확인결과, 대수기에서 정지기로 들어
선 48시간 후 항균활성이 가장 좋았으며, 배양 120시간까지
항균활성은 더 증대되지 않고 유지되는 것으로 확인하였다
(Fig. 6). 최적 배양시간은 접종 후 48시간까지 배양하는 것이
가장 효율적으로 항균 유용물질을 생산할 수 있는 조건으로
판단된다. 최종적으로 S3W-11 균주의 항균활성 증대를 위한
최적화 배지는 YPD로 선정하였고, YPD 배지를 이용한 일반
배양 및 최적화된 조건하에 48시간 배양 후 활성확인 결과
약 40%의 항균 활성효능이 증대되는 것으로 확인하였다.
S3W-11균주는 항균활성 물질 생산시 5 L fermenter의 최적
배양조건은 YPD배지, 배양온도 25oC, 초기 pH 6.5, 48시간
배양조건으로 결정하였다. 이 조건으로 배양하는 것이 가장
효율적으로 항균물질을 생산할 수 있는 조건으로 판단된다.
항균물질을 순수하게 정제하여 단일 물질로 인해 나타난 것
인지 확인하고, 화학적 특성과 구조를 규명하고자 한다. 아
울러 세포 및 동물 실험을 통해 인체에 대한 독성여부도 파
악하여, 산업용 소재로 개발 가능성을 확인하고자 한다.
Acknowledgements
본 연구는 환경부 산하 국립낙동강생물자원관 연구과제 (‘산
업화유망미생물의 대량배양기술개발’, NNIBR201902113)에
서 지원받았음.
REFERENCES
1. Kang, C. S. (2004) Antibiotic resistance and countermeasures. Pat-
ent 21 54:37-43.
2. KIHAST. (2016) Antibiotics resistance. Global social policy brief.
May, Vol. 20.
3. Shin, E. (2017) Antimicrobials and antimicrobial resistant super-
bacteria. Ewha Med. J. 40(3): 99-103.
4. Tawfik, K. A., P. Jeffs, B. Bray, G. Dubay, J. O. Falkinham, M.
Mesbah, D. Youssef, S. Khalifa, and E. W. Schmidt (2010) Burk-
holdines 1097 and 1229, potent antifungal peptides from Burk-
holderia ambifaria 2.2N. Org. Lett. 12: 664-666.
5. Mahenthiralingam, E., L. Song, A. Sass, J. White, C. Wilmot, A.
Marchbank, O. Boaisha, J. Paine, D. Knight, and G. L. Challis
(2011) Enacyloxins are products of an unusual hybrid modular
polyketide synthase encoded by a cryptic Burkholderia ambifaria
Genomic Island. Chem. Biol. 18: 665-677.
6. Farh Mel, A., Y. J. Kim, H. Van An, J. Sukweenadhi, P. Singh, M.
A. Huq, and D. C. Yang (2015) Burkholderia ginsengiterrae sp.
nov. and Burkholderia panaciterrae sp. nov., antagonistic bacteria
against root rot pathogen Cylindrocarpon destructans, isolated
from ginseng soil. Arch. Microbiol. 197: 439-447.
7. Nam, Y. H., A. Choi, B. S. Hwang, and E. J. Chung (2018) Anti-
microbial activities of Burkholderia sp. strains and optimization of
culture conditions. Kor. J. Microbiol. 54(4): 428-435.
8. Aouadhi, C., Z. Rouissi, S. Kmiha, S. Mejri, and A. Maaroufi
(2016) Effect of sporulation conditions on the resistance of Bacil-
lus sporothermodurans spores to nisin and heat. Food Microbiol.
54: 6-10.
9. Mogi, T., and K. Kita (2009) Gramicidin S and polymyxins: the
revival of cationic cyclic peptide antibiotics. Cell. Mol. Life Sci.
66: 3821-3826.
10. Shida, O., H. Takagi, K. Kadowaki, and K. Komagata (1996) Pro-
posal for two new genera Brevibacillus gen. nov. and Aneuriniba-
cillus gen. nov. Int. J. Syst. Bacteriol. 46: 939-946.
11. Choi, A., Y. H. Nam, K. Baek, and E. J. Chung (2019) Brevibacil-
lus antibioticus sp. nov., with a broad range of antibacterial activ-
ity, isolated from soil in the Nakdong River. J. Microbiol. 57(11):
991-996.
12. Guerra-Santos, L. H., O. Kappeli, and A. Fiechter (1986) Depen-
dence of Pseudomonas aeruginosa continous culture biosurfactant
production on nutritional and environmental factors. Appl. Micro-
biol. Biotechnol. 24: 443-448.
13. Abdel-Mawgoud, A. M., M. M. Abouwafa, and N. A. Hassouna
(2008) Optimization of surfactin production by Bacillus subtilis
Isolate BS5. Appl. Biochem. Biotechnol. 150: 305-325.
14. Smits Theo, H. M., F. Rezzonico, T. Kamber, J. Blom, A. Goes-
mann, A. Ishimaru Carol, E. Frey Jurg, V. O. Stockwell, and B.
Duffy (2011) Metabolic versatility and antibacterial metabolite
biosynthesis are distinguishing genomic features of the fire blight
antagonist Pantoea vagans C9-1. PLOS one 6(7): e22247.
15. Yoon, S., S. Ha, S. Kwon, J. Lim, Y. Kim, H. Seo and J. Chun
(2017) Introducing EzBioCloud: a taxonomically united database
of 16S rRNA gene sequences and whole-genome assemblies. Int.
J. Syst. Evol. Microbiol. 67: 1613-1617.
16. Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and D.
G. Higgins (1997) The CLUSTAL_X Windows interface: flexible
strategies for multiple sequence alignment aided by quality analy-
Fig. 6. Growth, pH and antimicrobial activities of Pantoea sp.
S3W-11 according to culture time by 5 L fermenter. Cell growth
(�), pH(▲), Inhibition zone(■).

Pantoea sp. S3W-11의 항균활성과 배양 최적화 71
sis tools. Nucleic. Acids Res. 25: 4876-4882.
17. Hall, T. A. (1999) BioEdit: a user-friendly biological sequence
alignment editor and analysis program for Windows 95/98/NT.
Nucleic Acids Symp Ser 41: 95-98.
18. Kumar, S., G. Stecher, and K. Tamura (2016) MEGA7: molecular
evolutionary genetics analysis version 7.0 for bigger datasets. Mol.
Biol. Evol. 33: 1870-1874.
19. Saitou, N., and M. Nei (1987) The neighbor-joining method: a
new method for reconstructing phylogenetic trees. Mol. Biol. Evol.
4: 406-425.
20. Felsenstein, J. (1981) Evolutionary trees from gene frequencies
and quantitative characters: finding maximum likelihood esti-
mates. Evolution 35(6): 1229-1242.
21. Fitch, W. M. (1971) Towards defining the course of evolution:
Minimum change for a specific tree topology. Syst. Biol. 20: 406-
416.
22. Felsenstein, J. (1985) Confidence limits on phylogenies: an approach
using the bootstrap. Evolution 39: 783-791.