oxygen isotope variation in bone phosphate
TRANSCRIPT

Applied Geochemistry, Vol. 4, pp, 317-323, 1989 0883-2927/89 $3.00 + .00 Printed in Great Britain Pergamon Press plc
Oxygen isotope variation in bone phosphate
BOAZ L u z and YEHOSHUA KOLODNY Department of Geology, Institute of Earth Sciences, The Hebrew University of Jerusalem,
Jerusalem 91904, Israel
Abstract--Temperature dependent ffactionation of O isotopes occurs between water and phosphate of fish bones, mammal bones and the phosphatic fraction of mollusc skeletons. In all these cases the fractionation follows the paleotemperature equation of LONGINELLI and NUTI. In molluscs and in fish the 61sO of the skeleton (dp) depends on the 6180 of the environmental water (dw) and on ambient temperature. Thus, as in other isotopic paleothermometers, estimation of dw is necessary for temperature calculation. Mammal 6p, which is not affected by changes in ambient temperature, is useful for estimating dw. Simple linear relations exist between dp of experimental rats, their body water isotopic composition (d~w) and d w of their drinking water. Body water composition depends on 6w of intake water as well as on the rate of metabolism. When the metabolic rate is high compared to the rate of drinking, bone phosphate will not be very sensitive to changes in 6w of the environmental water. In free ranging deer a significant amount of the intake water is water contained in leaves that the deer eat. In arid environments leaf water becomes highly enriched in lSO due to isotopic effects of evaporation. This enrichment is reflected in the bone phosphate. Modern people consume significant amounts of water from sources other than those in their immediate living environment. Consequently, their bone phosphate is not useful for estimating the isotopic composition of environmental water. However, this was not true for ancestral and more recent man, and therefore the dp of skeletons might be useful for tracing past human migrations.
INTRODUCTION
THE ANALYSIS of variations in the abundance of stable isotopes has led to major advances in various fields of the earth sciences. Reconstruction of earth paleocli- mates over geological ages was possible by measure- ments of O isotope abundances in carbonates and silicates, as well as phosphatic rocks.
UREY (1947) reported calculated equilibrium con- stants for O isotopic exchange reactions between water and various oxyanions, including carbonate and phosphate. Because these constants depend on temperature it was soon realized that they are useful for determination of past temperatures. Following UREY, remarkable advances took place in the development of carbonate paleotemperature equa- tions (McCREA, 1950; EPSTEIN et al., 1951, 1953) and their application to reconstruction of paleoclimates, especially those of the Pleistocene (EMILIANI, 1955; SHACKLETON and OPDYKE, 1973). The reconstruction of paleoclimates requires measurement of the iso- topic ratios of O in an authigenic solid phase (classi- cally CaCO3) and knowledge (or assumption) of the isotopic composition of the aqueous phase with which the solid equilibrated. In pre-Pleistocene paleother- mometry the isotopic composition of sea water is assumed (following EPSTEIN etal. , 1951, 1953) nearly constant. In the Pleistocene, variations in the isotopic ratio of sea water due to changes in global ice volume is the dominant factor affecting shifts in 6180 of carbonate shells between glacial and interglacial periods (SHACKLETON, 1967).
The reconstruction of continental paleoenviron- ments is more complicated. Whereas the assumption that the isotopic composition of sea water (d18Ow) was constant in time can be justified, that is not the case for the isotopic composition of meteoric water which is highly variable both in space and in time (DANSGAARD, 1964; YURTSEVER and GAT, 1981) even at the present. In addition, poor preservation of original mineralogy, especially of carbonates, makes it difficult to derive meaningful conclusions about past environments. Oxygen isotopic variations of bone phosphate have potential for overcoming these problems.
TUDGE (1960) has demonstrated that isotopic exchange between phosphate and water is negligible in inorganic reactions. Phosphates are thus not likely to change isotopically by dissolution-precipitation processes of diagenetic environments. On the other hand, biogenically (enzymatically catalyzed) precipi- tated phosphates in shells and bones are closely related to the isotopic composition and temperature of the solutions in which mineralization takes place (LoNGINELLI, 1965; LONGINELLI and NUTI, 1973a,b; KOLODNY et al., 1983). It thus seems that while bone phosphate records the conditions of its formation, it is likely to resist subsequent post depositional altera- tions. The analysis of the isotopic composition of O of mammal bones has a further advantage in that in warm blooded animals it is not affected by ambient temperatures (LoNGINELLI, 1984) and thus has poten- tial for deriving estimates of composition of meteoric water. Because 6w of meteoric water is related to air
317
318 Boaz Luz and Yehoshua Kolodny
temperature (YuRTSEVER and GAT, 1981), mammal bone phosphate should be useful for estimating paleotemperatures.
THE PHOSPHATE PALEOTHERMOMETER
As mentioned above, O isotope exchange between water and phosphate in inorganic reactions is very slow. For this reason an empirical relation for isotopic fractionation in the system H20-PO 4 at low tempera- tures has not been derived yet. Only recently have preliminary experiments made it possible to calibrate a paleotemperature equation for high temperatures (SHEMESH e t al. , 1988). In the low temperature range exchange takes place when catalyzed by certain enzymes. For example, in vitro experiments of FAL- LER and ELGAVISH (1984) prove that complete exchange of all oxygens of PO4 are catalyzed by gastric ATPase. Although the exact mechanism of phosphate biomineralization is not clear (DEGENS, 1976) similar reactions are expected to effect exchange of O between water and phosphate in bones and shells. We will now demonstrate that isotopic fractionation in various groups of vertebrates and invertebrates seem to obey the same rules.
The PO4 temperature equation was derived by LONGINELLI (1965), and was later revised by LONaI- NELLI and NuxI (1973a).
t = 111.4 - 4.3 (dip - diw). (1)
Calibration of Eqn (1) was done indirectly by com- paring 61sO of phosphate (dip) and carbonate (die) of marine molluscs (mostly bivalves) and 6180 of the sea water (diw) from the site of collection.
It was assumed that both carbonate and phosphate in the skeleton formed at the same conditions of
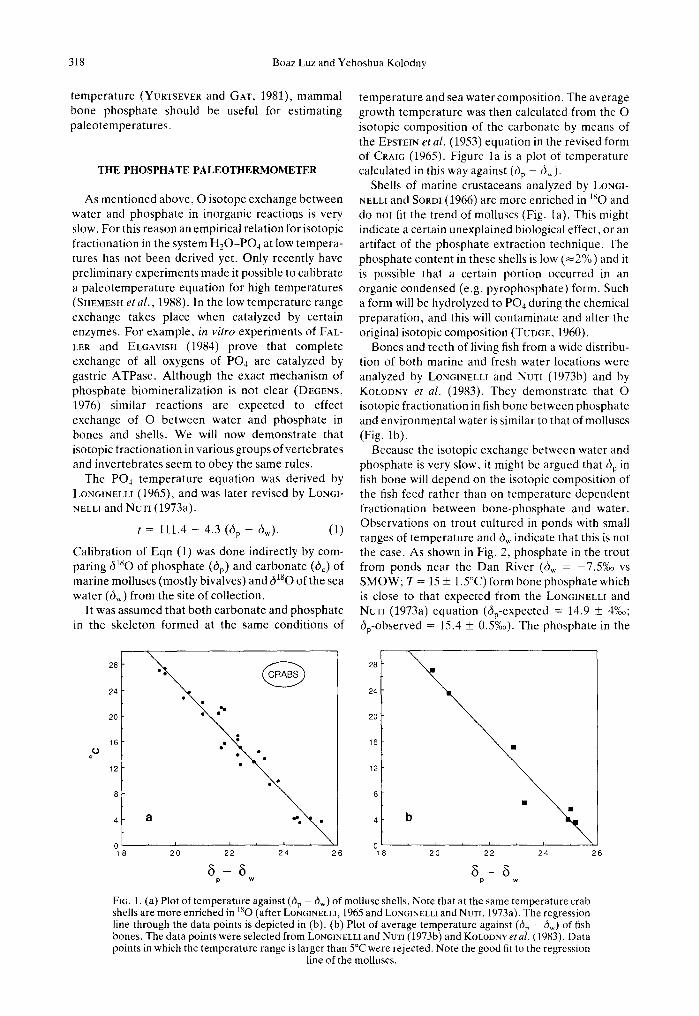
temperature and sea water composition. The average growth temperature was then calculated from the O isotopic composition of the carbonate by means of the EVSTEIN et al. (1953) equation in the revised form of CRAm (1965). Figure la is a plot of temperature calculated in this way against (6p - di,,).
Shells of marine crustaceans analyzed by LONCl- NELLI and SORDI (1966) are more enriched in l S o and do not fit the trend of molluscs (Fig. 1 a). This might indicate a certain unexplained biological effect, or an artifact of the phosphate extraction technique. The phosphate content in these shells is low (~2%) and it is possible that a certain portion occurred in an organic condensed (e.g. pyrophosphate) form. Such a form will be hydrolyzed to PO 4 during the chemical preparation, and this will contaminate and alter the original isotopic composition (TUDGE, 1960).
Bones and teeth of living fish from a wide distribu- tion of both marine and fresh water locations were analyzed by LONGINELL! and NvxI (1973b) and by KOLODNY et al. (1983). They demonstrate that O isotopic fractionation in fish bone between phosphate and environmental water is similar to that of molluscs (Fig. lb).
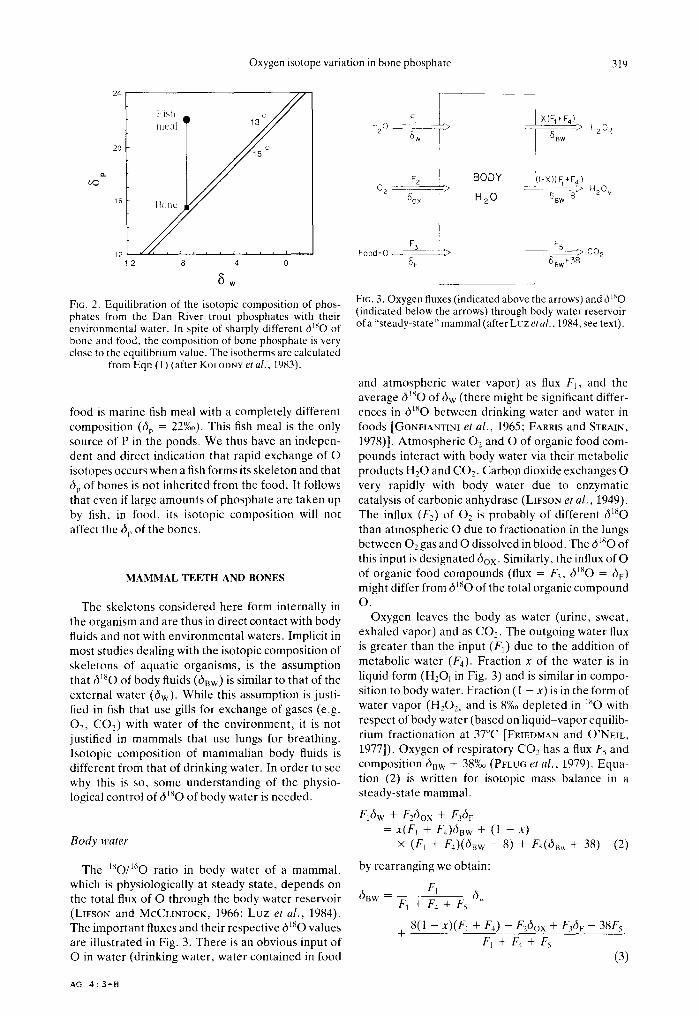
Because the isotopic exchange between water and phosphate is very slow, it might be argued that dip in fish bone will depend on the isotopic composition of the fish feed rather than on temperature dependent fractionation between bone-phosphate and water. Observations on trout cultured in ponds with small ranges of temperature and di,,, indicate that this is not the case. As shown in Fig. 2, phosphate in the trout from ponds near the Dan River (diw = -7.5%0 vs SMOW; T = 15 _+ 1.5°C) form bone phosphate which is close to that expected from the LONGINELLI and NUT~ (1973a) equation (alp-expected = 14.9 _+ 4%o; dip-observed = 15.4 + 0.5%o). The phosphate in the
0 o
28
24
20
16
12
8
4
0 18 20 22 24 26
5 - ~ 5 p w
28
24
20
16
12
8
4
0 ] 8 20 22 24 26
5 - 5 p w
FIG. 1. (a) Plot of temperature against (6p - dw) of mollusc shells. Note that at the same temperature crab shells are more enriched in lsO (after LOn6INELLI, 1965 and LONGINELLI and NuT~, 1973a). The regression line through the data points is depicted in (b). (b) Plot of average temperature against (6p - 6w) of fish bones. The data points were selected from LON6IYELLI and NuxI (1973b) and KOLODNV et al. (1983). Data points in which the temperature range is larger than 5°C were rejected. Note the good fit to the regression
line of the molluscs.
Oxygen isotope variation in bone phosphate 319
cO
24
Vish meal
20
16 BOlle
1 - 8
1/ /o , , i , , i
- 4 0
6w FIG. 2. Equilibration of the isotopic composition of phos- phates from the Dan River trout phosphates with their environmental water. In spite of sharply different dmO of bone and food, the composition of bone phosphate is very close to the equilibrium value. The isotherms are calculated
from Eqn (1) (after KOLODNY et al., 1983).
food is marine fish meal with a completely different composition (6~, = 22%0). This fish meal is the only source of P in the ponds. We thus have an indepen- dent and direct indication that rapid exchange of O isotopes occurs when a fish forms its skeleton and that 6p of bones is not inherited from the food. It follows that even if large amounts of phosphate are taken up by fish, in food, its isotopic composition will not affect the 6p of the bones.
M A M M A L T E E T H A N D BONES
The skeletons considered here form internally in the organism and are thus in direct contact with body fluids and not with environmental waters. Implicit in most studies dealing with the isotopic composition of skeletons of aquatic organisms, is the assumption that dl80 of body fluids (6BW) is similar to that of the external water (dw). While this assumption is justi- fied in fish that use gills for exchange of gases (e.g. O2, CO2) with water of the environment, it is not justified in mammals that use lungs for breathing. Isotopic composition of mammalian body fluids is different from that of drinking water. In order to see why this is so, some understanding of the physio- logical control of dtsO of body water is needed.
Body water
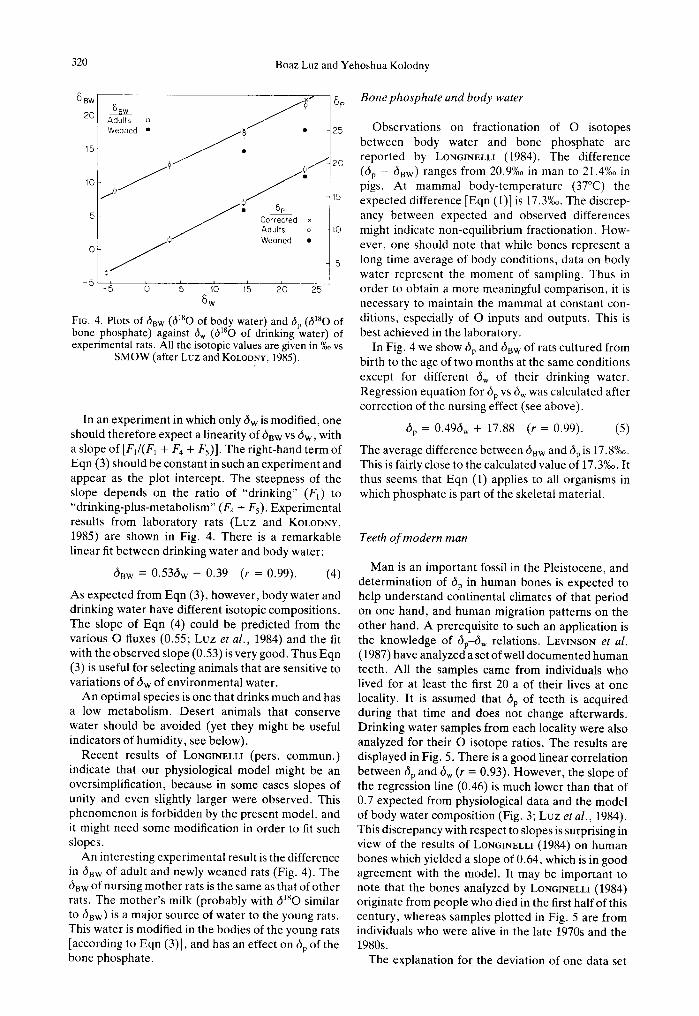
The 180/~60 ratio in body water of a mammal, which is physiologically at steady state, depends on the total flux of O through the body water reservoir (LIFSON and McCLINTOCK, 1966; Luz et al., 1984). The important fluxes and their respective 6~80 values are illustrated in Fig. 3. There is an obvious input of O in water (drinking water, water contained in food
i ! I
% . . . . F~ i
"t Food-O ~F
: X (F1 + F4)~ H23~
6BW
BODY (~-x)(E, F~) . . . . . ~> H2(] v
H20 8sw -8
F 5 CO~
6Bw~38
Fro. 3. Oxygen fluxes (indicated above the arrows) and 6180 (indicated below the arrows) through body water reservoir of a "steady-state" mammal (after Luz et al.. 1984, see text).
and atmospheric water vapor) as flux Ft, and the average 6~sO of dw (there might be significant differ- ences in 6taO between drinking water and water in foods [GONFIANTINI et al., 1965; FARRB and STRAIN, 1978)]. Atmospheric O z and O of organic food com- pounds interact with body water via their metabolic products H20 and CO2. Carbon dioxide exchanges O very rapidly with body water due to enzymatic catalysis of carbonic anhydrase (LIFSON et al., 1949). The influx ( ~ ) of 02 is probably of different 6180 than atmospheric O due to fractionation in the lungs between 02 gas and O dissolved in blood. The dtsO of this input is designated 6ox. Similarly, the influx of O of organic food compounds (flux = F~, 6 m O = 6F) might differ from dlSO of the total organic compound O.
Oxygen leaves the body as water (urine, sweat, exhaled vapor) and as CO2. The outgoing water flux is greater than the input (FI) due to the addition of metabolic water (F4). Fraction x of the water is in liquid form (H2Ol in Fig. 3) and is similar in compo- sition to body water. Fraction (1 - x) is in the form of water vapor ( H z O 2 v and is 8%0 depleted in 180 with respect of body water (based on liquid-vapor equilib- rium fractionation at 37°C ]FRIEDMAN and O'NEIL, 1977]). Oxygen of respiratory CO2 has a flux F5 and composition 6Bw + 38%0 (PFcuO et al., 1979). Equa- tion (2) is written for isotopic mass balance in a steady-state mammal.
FlOw + F26ox + F3,6F = x(F~ + F 4 ) a B w + (1 - x )
x (F, + F4)(aBw - 8) + Fs(6B~, + 38) (2)
by rearranging we obtain:
0my - Ft 6, , F~ + Fa + I(~
8(1 - x)(F, + F4) - Fz60x + F3O v - 38Fs + FI + Fa + F5
(3)
AG 4:3-H
320 Boaz Luz and Yehoshua Kolodny
20 ~ w
Weaned ~ ~ ? * 25
15
5 ~ CorrOded ,, Adults o IO
~ [ ) ~ " Weoned • ° / 5
-s -i & ; ,b ,; 2'o ~ 6w
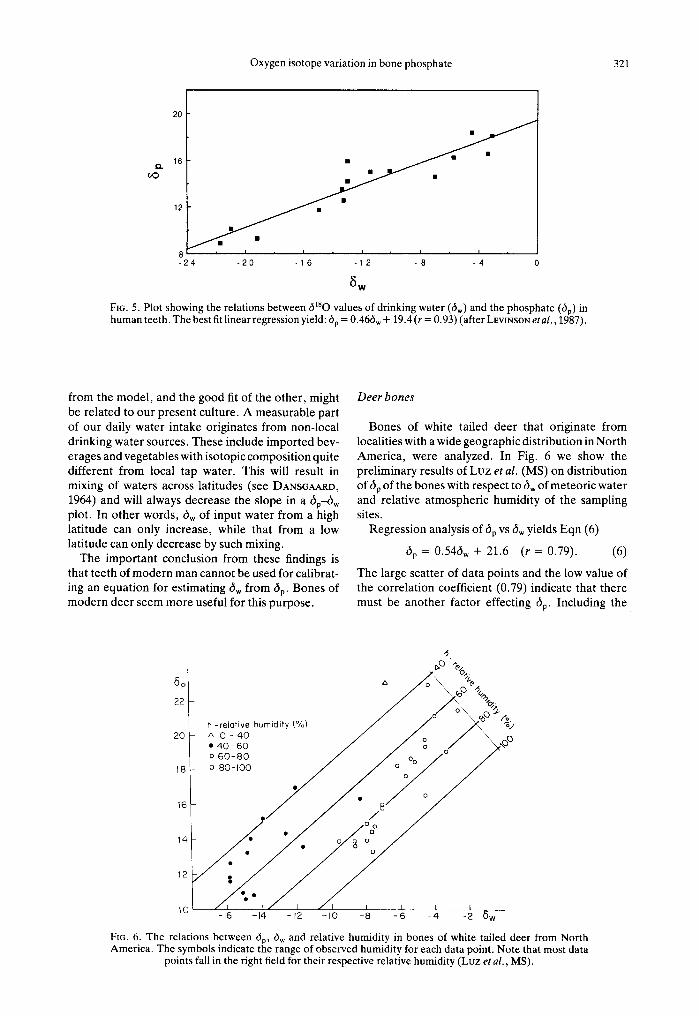
FIG. 4. Plots of 6Bw (6180 of body water) and 6p 6180 of bone phosphate) against 6w (6180 of drinking water) of experimental rats. All the isotopic values are given in %0 vs
SMOW (after Luz and KOLOBNY, 1985).
In an experiment in which only 6w is modified, one should therefore expect a linearity of 6BW VS 6w, with a slope of [FJ(F1 + F4 + Fs)]. The right-hand term of Eqn (3) should be constant in such an experiment and appear as the plot intercept. The steepness of the slope depends on the ratio of "drinking" (F1) to "drinking-plus-metabolism" (F4 + Fs). Experimental results from laboratory rats (Lvz and KOLODNY, 1985) are shown in Fig. 4. There is a remarkable linear fit between drinking water and body water:
6BW = 0.53dw -- 0.39 (r = 0.99). (4)
As expected from Eqn (3), however, body water and drinking water have different isotopic compositions. The slope of Eqn (4) could be predicted from the various O fluxes (0.55; Luz et al., 1984) and the fit with the observed slope (0.53) is very good. Thus Eqn (3) is useful for selecting animals that are sensitive to variations of dw of environmental water.
An optimal species is one that drinks much and has a low metabolism. Desert animals that conserve water should be avoided (yet they might be useful indicators of humidity, see below).
Recent results of LONOINELLI (pers. commun.) indicate that our physiological model might be an oversimplification, because in some cases slopes of unity and even slightly larger were observed. This phenomenon is forbidden by the present model, and it might need some modification in order to fit such slopes.
An interesting experimental result is the difference in 6Bw of adult and newly weaned rats (Fig. 4). The 6BW of nursing mother rats is the same as that of other rats. The mother 's milk (probably with 6180 similar to dBw) is a major source of water to the young rats. This water is modified in the bodies of the young rats [according to Eqn (3)], and has an effect on dp of the bone phosphate.
Bone phosphate and body water
Observations on fractionation of O isotopes between body water and bone phosphate are reported by LONGINELLI (1984). The difference (6p - 6Bw) ranges from 20.9%0 in man to 21.4%o in pigs. At mammal body-temperature (37°C) the expected difference [Eqn (1)] is 17.3%o. The discrep- ancy between expected and observed differences might indicate non-equilibrium fractionation. How- ever, one should note that while bones represent a long time average of body conditions, data on body water represent the moment of sampling. Thus in order to obtain a more meaningful comparison, it is necessary to maintain the mammal at constant con- ditions, especially of O inputs and outputs. This is best achieved in the laboratory.
In Fig. 4 we show 6p and 6BW of rats cultured from birth to the age of two months at the same conditions except for different 6w of their drinking water. Regression equation for 6p vs 6 w was calculated after correction of the nursing effect (see above).
dp = 0.49dw + 17.88 (r = 0.99). (5)
The average difference between 6Bw and 6p is 17.8%o. This is fairly close to the calculated value of 17.3%o. It thus seems that Eqn (1) applies to all organisms in which phosphate is part of the skeletal material.
Teeth of modern man
Man is an important fossil in the Pleistocene, and determination of 6p in human bones is expected to help understand continental climates of that period on one hand, and human migration patterns on the other hand. A prerequisite to such an application is the knowledge of 6p-6w relations. LEVINSON et al. (1987) have analyzed a set of well documented human teeth. All the samples came from individuals who lived for at least the first 20 a of their lives at one locality. It is assumed that bp of teeth is acquired during that time and does not change afterwards. Drinking water samples from each locality were also analyzed for their O isotope ratios. The results are displayed in Fig. 5. There is a good linear correlation between 6p and 6w (r = 0.93). However, the slope of the regression line (0.46) is much lower than that of 0.7 expected from physiological data and the model of body water composition (Fig. 3; Lvz et al., 1984). This discrepancy with respect to slopes is surprising in view of the results of LONGINELL! (1984) on human bones which yielded a slope of 0.64, which is in good agreement with the model. It may be important to note that the bones analyzed by LONGINELLI (1984) originate from people who died in the first half of this century, whereas samples plotted in Fig. 5 are from individuals who were alive in the late 1970s and the 1980s.
The explanation for the deviation of one data set
Oxygen isotope variation in bone phosphate 321
20
12 •
8 i I I , I , I i I
- 2 4 - 2 0 - 1 6 - 1 2 - 8 - 4
8w FIG. 5. Plot showing the relations between 6180 values of drinking water (6w) and the phosphate (dp) in human teeth. The best fit linear regression yield: dp= 0.466 w + 19.4 (r = 0.93) (after LEVlNSON et al., 1987).
from the model, and the good fit of the other, might be related to our present culture. A measurable part of our daily water intake originates from non-local drinking water sources. These include imported bev- erages and vegetables with isotopic composition quite different from local tap water. This will result in mixing of waters across latitudes (see DANSGAARD, 1964) and will always decrease the slope in a 6p-6w plot. In other words, 6w of input water from a high latitude can only increase, while that from a low latitude can only decrease by such mixing.
The important conclusion from these findings is that teeth of modern man cannot be used for calibrat- ing an equation for estimating 6w f r o m 6p. Bones of modern deer seem more useful for this purpose.
Deer bones
Bones of white tailed deer that originate from localities with a wide geographic distribution in North America, were analyzed. In Fig. 6 we show the preliminary results of Loz et al. (MS) on distribution of 60 of the bones with respect to 6w of meteoric water and relative atmospheric humidity of the sampling sites.
Regression analysis of 60 vs 6w yields Eqn (6)
6p = 0.546w + 21.6 (r = 0.79). (6)
The large scatter of data points and the low value of the correlation coefficient (0.79) indicate that there must be another factor effecting 6p. Including the
6p
22
20
18
16
14
12
I 0
h -relotive humidity (%1 z~ 0 - 4 0 • 4 0 - 60 o 6 0 - 8 0 a 80-100
-16 - 1 4 - 1 2
4
/ / ", / / / ° / / / ' / Y
¢" I I I t I - I 0 - 8 - 6 - 4 - 2 ~W
FIG. 6. The relations between 6p, 6w and relative humidity in bones of white tailed deer from North America. The symbols indicate the range of observed humidity for each data point. Note that most data
points fall in the right field for their respective relative humidity (Luz et al., MS).

322 Boaz Luz and Yehoshua Kolodny
relative humidity (h, %) in a multiple regression analysis yields Eqn (7)
dp = 0.876w - 0.13h + 32.2 (r = 0.92). (7)
The effect of humidity is to increase 6p in arid locali- ties. This does not result from a direct effect on the deer water-budget (see Luz and KOLODNY, 1985), and the explanation is probably related to the drink- ing and diet of the animals.
As discussed above, linear relations between dp and dw are observed in experimental rats. We expect similar relations in wild mammals, but with the signifi- cant difference that all water inputs have to be con- sidered. In particular, deer obtain a large part of their water supply from eating leaves (RUE, 1978). In turn, leaves contain water which may be highly enriched in 180, especially at low relative humidity (GONFIANTINI et al., 1965; EPSTEIN et al., 1977; BURK and STUIVER, 1981 ; FARRIS and STnArN, 1978). In addition to evap- oration effects in leaves, surface meteoric water also shows evaporative 180 enrichment (GAT, 1981), which will contribute to an increase in bp through drinking.
We have a strong indication that analysis of bones of fossil deer should reflect the combined effects of changes in dw of post precipitation (and thus paleo- temperature) and changes in relative humidity. Other mammals which are less dependent on leaf water for their water needs might be less sensitive to humidity changes. Thus analysis of bones of different species may be useful for resolving changes in both 6w and relative humidity.
FUTURE DEVELOPMENTS
In addition to the potential application of O isotope variations in bones for paleoclimate reconstruction, it might be possible to use that method in tracing human migrations. This suggestion is based on the different residence time of phosphate in various parts of the skeleton (HEANEY, 1976). Thus when people migrate and change the isotopic composition of their intake water, different bones might readjust to the change at different rates. Bones with long residence time will not change much, but bones in which phos- phate is more easily replaced will adjust to the new conditions.
We have analyzed human teeth of immigrants who moved to Israel from countries where the climate is colder. The isotopic composition of these teeth is always less enriched in lSO than in teeth of native Israelis. It would be desirable to compare results from other bones of the same individuals. Unfortu- nately, these are not easy to obtain. Experiments with rats in which isotopic composition of intake water was changed for a period of a few weeks, indicate a considerable shift in the isotopic compo- sition of the bone phosphate (Lvz and KOLODNY, 1985). Although the rate of change in human bones is
expected to be slower than in rats, the experimental result suggests that it might change if given sufficient time.
In an attempt to test the possibility of migrations across climatic boundaries, we have analyzed two skeletons from the Byzantine site at Rehovoth, Israel. The site is located at the border between the desert and the marine influence of the Mediterranean Sea. In one skeleton which is poorly preserved, Op has an average value of about 22%0 while in the other (well preserved) dp values range from 19 to 21%o. The results may indicate migration of the individual with the isotopically heterogenous skeleton from a rela- tively humid climate to the desert. After migrating, dp of certain bones was reset while other bones retained the original composition. It is also possible that bone diagenesis affected the original isotopic composition and homogenized the g)p values of vari- ous bones. Clearly, more research is needed before reaching any further conclusions.
CONCLUSIONS
(1) Fractionation of O isotopes between biogenic phosphate in skeletons and water is similar in mol- luscs, fish and mammals. This suggests that the biochemical reactions leading to precipitation of skeletal phosphate is similar in many organisms.
(2) The isotopic composition of fish bones (6p) depends on that of the environmental water (dw) and the ambient temperature. In mammals it depends on the isotopic composition of body water (6Bw) and the constant body temperature.
(3) Linear relations exist between dw of intake water and dBw and between 6w and dp. Thus dp of mammal bones is useful for estimating 6,,..
(4) Leaves constitute a main portion of the diet of wild deer, and thus leaf water amounts to a significant portion of the total water intake. Because in arid environments leaf water becomes highly enriched in 1sO, this enrichment is recorded in the deer bones.
(5) The isotopic composition of teeth of modern people corresponds to the meteoric water of their living environment. However, because a significant portion of the water intake originates from imported vegetables and beverages, the modern human bone phosphate is a poor indicator of the isotopic compo- sition of local meteoric water.
(6) The residence time of phosphate in different parts of the skeleton is not uniform. When people migrate across climatic boundaries, and the isotopic composition of their intake water changes, different bones will respond to the change at different rates. As a result the skeleton will acquire a heterogenous isotopic composition. This might be useful in tracing past human migrations.
Acknowledgements--The first author wishes to thank the organizers of the Oxford Fossil Bone Workshop for their

Oxygen isotope variation in bone phosphate 323
invitation to participate and present this review, and for their hospitality during the meeting. The various studies on which this review is based were supported by two grants from the U.S.A.-Israel Binational Science Foundation. Grant No. 80-002231 on Variations of Oxygen Isotope Ratios in Biogenic Apatites, and grant No. 85-00044 on Variations of Oxygen Isotope Ratios in the Marine Phos- phorus Cycle.
REFERENCES
BURK R. L. and STUIVER M. (1981) Oxygen isotope ratios in trees reflect mean annual temperature and humidity. Science 211, 1417-1419.
CRAX6 H. (1965) The measurement of oxygen isotope paleD- temperatures. In Stable Isotopes in Oceanographic Studies and Paleotemperatures, pp. 1-23, Pisa, Consiglio Nazionale delle Richerche Laboratorio di Geologia Nucleare.
DANSGAARD W. (1964) Stable isotopes in precipitation. Tellus 16,436-468.
DE6ENS E. T. (1976) Molecular mechanisms on carbonate, phosphate, and silica deposition in the living cell. In Topics" in Current Chemistry, pp. 1-112. Springer.
EMILIANI C. (1955) Pleistocene temperatures, J. Geol. 63, 538-578.
EPSTEIN S., BUCHSBAUM R., LOWENSTAM H. A. and UREY H. C. (1951) Carbonate-water isotopic temperature scale. Geol. Soc. Am. Bull. 62,417-426.
EPSTEIN S., BUCHSBAUM R., LOWENSTAM H. A. and UREY H. C. (1953) Revised carbonate-water isotopic tem- perature scale. Geol. Soc. Am. Bull. 64, 1315-1326.
EPSTEIN S., THOMPSON P. and YAPP C. J. (1977) Oxygen and hydrogen isotopic ratios in plant cellulose. Science 198, 1209-1215.
FALLER L. D. and ELGAVlSH G. A. (1984) Catalysis of oxygen-18 exchange between inorganic phosphate and water by the gastric H,K-ATPase. Biochem. 23, 6584- 6590.
FARRIS F. and STRAIN B. R. (1978) The effects of water- stress on leafH 2180 enrichment. Rad. Environ. Biophys. 15, 167-202.
FRIEDMAN I. and O'NEIL J. R. (1977) Compilation of stable isotope fractionation factors of geochemical interest. U.S. Geol. Surv. Prof. Paper 440KK, pp. 1-12.
GAT J. R. (1981) Isotopic fractionation. In Deuterium and Oxygen-18 in the Water Cycle, pp. 203-221. IAEA, Vienna.
GONEIANTINI G., GRATZIU S. and TONGIORGI E. (1965) Oxygen isotopic composition of water in leaves. In Isotopes and Radiation in Soil Plant Nutrition Studies, pp. 405-410. IAEA, Vienna.
HEANEY R. P. (1976) Calcium kinetics in plasma: as they apply to the measurements of bone formation and resorp- tion rates. In The Biochemistry and Physiology o f Bone, pp. 105-133. Academic Press.
KOLODNY Y., Luz B. and NAVON O. (1983) Oxygen isotope variations in phosphate of biogenic apatites, I. Fish bone apatite--rechecking the rules of the game. Earth Planet. Sci. Lett. 64,398-404.
LEVINSON A. A., Luz B. and KOLODNY Y. (1987) Variations in oxygen isotopic compositions of human teeth and urinary stones. Appl. Geochem. 2,367-371.
LIESON N., GORDON G. B., VISSCHER M. B. and NlER A. O. C. (1949) The fate of utilized molecular oxygen and the source of the oxygen of respiratory carbon dioxide, studied with the aid of heavy oxygen. J. biol. Chem. 180,803-811.
LIFSON N. and McCLINTOCK R. (1966) Theory of use of the turnover rates of body water for measuring energy and material balance. J. theor. Biol. 12, 46-74.
LONG1NELLI A. (1965) Oxygen isotopic composition of orthophosphate from shells of living marine organisms. Nature 207,716.
LONG1NELLI A. (1984) Oxygen isotopes in mammal bone phosphate: a new tool for paleohydrological and paleD- climatological research? Geochim. cosmochim. Acta 48, 385-390.
LONGINELEI A. and NUTt S. (1973a) Revised phosphate- water isotopic temperature scale. Earth Planet. Sci. Len. 19,373-376.
LONGINELLI A. and NUTI S. (1973b) Oxygen isotope meas- urements of phosphate from fish teeth and bones. Earth Planet. Sci. Lett. 20,337-340.
LONGINELLI A. and SORDI M. (1966) Oxygen isotopic com- position of phosphate from shells of some living crus- taceans. Nature 211,727.
Lvz B. and KOLODNV Y. (1985) Oxygen isotope variations in phosphate of biogenic apatites. IV. Mammal teeth and bones. Earth Planet. Sci. Lett. 75, 29-36.
Luz B., KOLODNY Y. and HOROWITZ M. (1984) Fraction- ation of oxygen isotopes between mammalian bone- phosphate and environmental drinking water. Geochim. cosmochim. Acta 48, 1689-1693.
MCCREA J. M. (1950) On the isotopic chemistry of car- bonates and a paleotemperature scale. J. chem. Phys. 18, 849- 857.
PFLUG K. P., SCHUSTER K. D., PICHOTKA J. P. and FORSTEL H. (1979) Fractionation effects of oxygen isotopes in mammals. In Stable Isotopes: Proceedings o f the Third International Conference (ed. E. R. KLEIN and P. D. KLEIN) pp. 553--561. Academic Press.
RUE L. L. (1978) The Deer of North America. Crown. SHEMESH A., KOLODNY Y. and Luz B. (1988) Isotope
geochemistry of oxygen and carbon in phosphate and carbonate of phosphorite francolite. Geochim. cosmo- chim. Acta, 52, 2565-2572.
SHACKLETON h. (1967) Oxygen isotope analyses and Pleis- tocene temperatures reassessed. Nature 215, 15-17.
SHACKLETON N. J. and OPDYKE N. D. (1973) Oxygen isotope and paleomagnetic stratigraphy of equatorial Pacific core V28-238: oxygen isotope temperatures and ice volumes on a 105 year and 106 year scale. Quatern. Res. 3, 39-55.
TUDGE A. P. (1960) A method of analysis of oxygen isotopes in orthophosphate--its use in the measurement of paleo- temperatures. Geochim. cosmochim. Acta 18, 81-93.
UREV H. C. (1947) The thermodynamic properties of isotopic substances. J. Chem. Soc., 562-58l.
YURTSEVER Y. and BAT J. R. (1981) Atmospheric waters. In Stable Isotope Hydrology (ed. J. R. GAT and R. GONFIANT1NI) Chap. 6, pp. 103-142. Tech. Report Series, No. 210. IAEA, Vienna.