overexpression nanog activates pluripotent genes in porcine fetal fibroblasts and nuclear transfer...
TRANSCRIPT

THE ANATOMICAL RECORD 294:1809–1817 (2011)
Overexpression Nanog ActivatesPluripotent Genes in Porcine FetalFibroblasts and Nuclear Transfer
EmbryosLI ZHANG, YI-BO LUO, GERELCHIMEG BOU, QING-RAN KONG,
YAN-JUN HUAN, JIANG ZHU, JIAN-YU WANG, HUI LI, FENG WANG,YONG-QIAN SHI, YAN-CHANG WEI, AND ZHONG-HUA LIU*
Department of Life Science, Northeast Agriculture University, Heilongjiang Province, China
ABSTRACTNanog as an important transcription factor plays a pivotal role in
maintaining pluripotency and in reprogramming the epigenome of so-matic cells. Its ability to function on committed somatic cells and embryoshas been well defined in mouse and human, but rarely in pig. To betterunderstand Nanog’s function on reprogramming in porcine fetal fibroblast(PFF) and nuclear transfer (NT) embryo, we cloned porcine Nanog CDSand constructed pcDNA3.1 (þ)/Nanog and pEGFP-C1/Nanog overexpres-sion vectors and transfected them into PFFs. We studied the cell biologi-cal changes and the expression of Nanog, Oct4, Sox2, Klf4, C-myc, andSall4 in transfected PFFs. We also detected the development potential ofthe cloned embryos harboring Nanog stably overexpressed fibroblasts andthe expression of Oct4, Sox2, and both endogenous and exogenous Nanogin these embryos. The results showed that transient overexpressionNanog in PFF could activate the expression of Oct4 (5-fold), C-myc (2-fold), and Sall4 (5-fold) in somatic cells, but they could not be maintainedduring G418 selection. In NT embryos, although Nanog overexpressiondid not have a significant effect on blastocyst development rate and blas-tocyst cell number, it could significantly activate the expression of endoge-nous Nanog, Oct4, Sox2 to 160-fold, 93-fold, and 182-fold, respectively (P< 0.05). Our results demonstrate that Nanog could interact with and acti-vate other pluripotent genes both in PFFs and embryos. Anat Rec,294:1809–1817, 2011. VVC 2011 Wiley-Liss, Inc.
Keywords: pig; Nanog; overexpression; pluripotent genes;activation
INTRODUCTION
Nanog is a novel pluripotent gene that plays a crucial rolein maintaining the undifferentiated state of mouse embry-onic stem cells (mESCs) and inner cell mass (Chamberset al., 2003; Mitsui et al., 2003). We all know there are threeways to erase the developmental programming of differenti-ated cell nuclei, nuclear transfer (NT) into enucleatedoocytes, cell fusion with pluripotent stem cells or introduc-tion of combination defined transcriptional factors into so-matic cells. In generating induced pluripotent stem cells(iPSCs), Nanog is not included in the classic four factors(Oct4, Sox2, Klf4, and C-myc), but it plays important role inreprogramming by interacting with these pluripotent-associ-
Additional Supporting Information may be found in theonline version of this article.
Li Zhang and Yi-Bo Luo contributed equally to this work.
Grant sponsor: The National Basic Research Program of China;Grant number: 2009CB94100; Grant sponsor: National NaturalScience Foundation of China; Grant number: 30871431.
*Correspondence to: Zhong-Hua Liu, Department of Life Sci-ence, Northeast Agriculture University, Heilongjiang Province,China. Fax: þ86045 155191747. E-mail: [email protected]
Received 18 April 2011; Accepted 12 June 2011
DOI 10.1002/ar.21457Published online 3 October 2011 in Wiley Online Library(wileyonlinelibrary.com).
VVC 2011 WILEY-LISS, INC.

ated genes. Actually, Nanog with Lin28 could replace Klf4and C-myc then directly participate in reprogramminghuman somatic cells (Yu et al., 2007; Choi et al., 2009;Hanna et al., 2009). The activation of endogenous Nanog isindispensable for generating iPSCs that had the ability tocontribute to adult chimeras (Takahashi and Yamanaka,2006; Okita et al., 2007). Besides, Nanog is essential fortransforming the dedifferentiated intermediates to groundstate pluripotency in reprogramming process (Silva et al.,2008). Overexpression Nanog in ESCs could enhance thetransfer of pluripotency in fusion experiment and overex-pression Nanog is adequate to convert epiblast stem cells toground state pluripotency (Silva et al., 2009).
Pig is considered as a useful and meaningful animalmodel in therapeutic and biomedical research, such asbioreactor and xenotransplantation. ESCs have been con-sidered as a useful tool for generating transgenic pig.However, the research of these potential applications pro-gressed slowly, because no authentic porcine ESCs isavailable to date (Li et al., 2004; Brevini et al., 2007; Kimet al., 2007; Hall, 2008; Kim et al., 2010). Although threegroups announced recently they had established iPSCs inpig by introduced human or mouse derived transcrip-tional factors (Oct4, Sox2, Klf4, and C-myc), none of themsuccessfully applied the iPSCs in generating chimeras,which is the basic standard to define pluripotency (Este-ban et al., 2009; Ezashi et al., 2009; Wu et al., 2009). Onlyone report showed that porcine mesenchymal stem cells(MSCs) transduced with six human transcription factors(Nanog,Oct4, Sox2, Klf4, C-myc, and Lin28) could gener-ate chimeras and suggested the insufficient endogenousNanog expression may be responsible for the failure ofgenerating chimeras (West et al., 2010). A recent reportalso mentioned that pig iPSCs generated by six genes hadhigher percentage of undifferentiated colonies during theearly passages (Wu et al., 2009). All the work reminded usto pay attention to the Nanog’s function on reprogram-ming in pig, especially on the interaction with other pluri-potent genes. Most of studies were merely confined toNanog expression patterns by immunofluorescence andreal time-PCR in porcine embryos and somatic stem cells,but the function of its role in cellular reprogramming andearly embryo development by cooperating with othergenes is still not defined. (Blomberg et al., 2008; Goelet al., 2008; Magnani and Cabot, 2008; Hall et al., 2009).
In this study, we cloned Nanog CDS and subcloned it topcDNA3.1 (þ) and pEGFP-C1 vector. After transfecting thevectors into porcine fetal fibroblast (PFF), we systematicallydetected the biological characteristics of these cells and theexpression of Oct4, Sox2, Nanog, Klf4, C-myc, and Sall4both in transient and stable transfected PFF. We also con-structed NT embryos derived from Nanog overexpressedPFF and analyzed the development potential in vitro. Theexpression of the pluripotency master genes, Oct4, Sox2,and Nanog in different stages of early embryos was alsodetected by quantitative Real time-PCR. This research couldhelp us to better understand the role of Nanog in cellularreprogramming and early embryos development in pig.
MATERIALS AND METHODSGene Cloning and Vector Construction
The Nanog gene was amplified by reverse transcrip-tase-polymerase chain reaction (RT-PCR) using totalRNA extracted from pools of MII stage oocytes using Tri-
zol (Invitrogen). And the total RNA was reverse tran-script using High Capacity cDNA Reverse TranscriptionKit (ABI). The PCR amplification was carried out for onecycle with denaturing at 94�C for 4 min, and 35 subse-quent cycles with denaturing at 95�C for 30 s, annealingat 56.5�C for 30 s, extension at 72�C for 30 s, and a finalextension at 72�C for 10 min. The sense primers 50-ATGAGTGTGGATCCAGCTTGTC-30 and antisense primer50-TCACATATCTTCAGGCTGTATG-30 are used forNanog cloning. To construct the pcDNA3.1 (þ)/Nanogplasmid, the Nanog cDNA was insert into pcDNA3.1 (þ)(Invitrogen) between HindIII and XhoI. Nanog cDNAwas also inserted into pEGFP-C1 (Invitrogen) betweenBglII and XhoI to generate pEGFP-C1/Nanog thatexpresses GFP-Nanog fusion protein.
Cell Culture and Transfection
Pig Fetal Fibroblasts (PFF) were isolated from 35-days-old male fetus, which were dissected by scalpel blade inPBS buffer and then treated in collagenase/DNAse I at37�C for 45 min. The solution was mixed with PFF culturemedium: DMEM (high glucose) supplemented with 10%FBS and penicillin/streptomycin (all the reagents arefrom Gibco), centrifuged at 1,000 rpm for 5 min and sus-pended in PFF culture medium. The cells were passagedin PFF culture medium without any antibiotics.
For transfection experiment, cells were plated in a3.5-cm plate to achieve 80–90% confluence within 24 hin PFF culture medium without penicillin/streptomycin.Cells were transfected with Lipofactamine 2000 (Invitro-gen) according the instruction with a ratio of 3:1 trans-fection reagent (ml): DNA (mg). Cells were passaged 24 hafter transfection. Selection medium DMEM (high glu-cose supplemented with 10% FBS and 500 ug/mL G418)replaced the PFF culture medium after cells adhesion.Cells for stable selection were rendered by G418 selec-tion for 20 days, when the untransfected cells died com-pletely, while cells for transient transfection wereharvested at 24 h later after transfection.
Immunofluorescence
Porcine fibroblasts were seeded and cultured for up to 3days before analysis. Growth medium was removed, andthe cells were fixed with 4% Para formaldehyde/PBS (pH7.4) for at least 40 min, followed by permeabilization with1% TritonX-100 at room temperature for 30 min, blockedin 1% BSA supplemented PBS for 1 h and then incubatedwith rabbit anti-Nanog antibody (Santa Cruz; 1:200) over-night at 4�C. After three washes with PBS containing0.1% Tween 20 and 0.01% Triton X-100 for 5 min each,fibroblast cells were labeled with 1:200 FITC-conjugatedIgG for 1 h at room temperature. After washing in PBScontaining 0.1% Tween 20 and 0.01% Triton X-100, thecells were co-stained with Hoechst33342 (10 mg/mL inPBS) and imaged on a Nikon Eclipse 80i microscope.
Oocytes In-Vitro Maturation
Pig (Sus scrofa) ovaries from prepubertal gilts were col-lected at a local slaughterhouse and transported to thelaboratory in an insulated container at 37�C. Antral fol-licles between 5 and 8 mm in diameter were aspiratedmanually with a disposable 10-cc syringe and an 18-gauge
1810 ZHANG ET AL.

needle. Follicular fluid was pooled and allowed to settle bygravity. Cumulus-oocytes complexes (COCs) were resus-pended in Hepes-buffered medium containing 0.01% poly-vinyl alcohol (PVA). Under a dissecting microscope, COCswith multiple layers of intact cumulus cells were selectedfor the experiments. Around 50–75 COCs were placed in500 mL of tissue culture medium 199 (TCM-199; GibcoBRL, Grand Island, NY) containing 0.14% PVA, 10-ng/mLepidermal growth factor, 0.57-mM cysteine, 0.5-IU/mL pigFSH, and 0.5-IU/mL ovine LH. COCs were matured for42–44 h at 39�C and 5% CO2 in air, 100% humidity. Allchemicals were obtained from Sigma Chemical Company(St. Louis, MO) unless stated otherwise.
In Vitro Fertilization, NT and Embryos Culture
Following short time incubation at 39.5�C, theextended semen was resuspended with DPBS (0.1%BSA), and washed 2 times with DPBS (0.1% BSA) bycentrifugation at 2,000 rpm for 4 min. The spermatozoaconcentration was measured using a hemocytometer andthe proportion of motile sperm was determined. Thespermatozoa were diluted with mTBM to optimal con-centration. Cumulus-free matured oocytes were washedthree times in mTBM IVF medium. Approximately, 30–35 oocytes were transferred into 50-mL droplets of IVFmedium covering with mineral oil that had been equili-brated for 30 min at 39.5�C in 5% CO2 in air before add-ing the sperm. 50-mL sperm sample was added to thefertilization droplets containing oocytes, giving a finalsperm concentration of 6 � 104 cells/mL. Oocytes werecoincubated with sperm in mTBM for 5 h. Then theoocytes were washed and cultured in PZM-3.
The in-vitro mature oocytes were enucleated by aspi-rating the first polar body and the adjacent cytoplasm inmanipulation droplet (TCM-199-HEPES enriched with0.3% BSA). Nanog stably overexpressed PFF were usedas donor cells and were placed under the zona pellucidaof enucleated oocytes. Fusion/activation was induced bysuccessive DC impulse of 1.2 kv/cm for 30 ls. Con-structed embryos were cultured in PZM-3 at 39�C under5% CO2 in air. The blastocysts were collected at 156 h,and then were stained in DPBS enriched with 10 mg/LHoechst33342 in dark. After staining, the blastocystswere transferred to slides with 7-uL glycerol on it, cover-ing with cover glass and mounting by nail polish. Thewavelength of 488 nm was used to detect whether theblastocysts were GFP positive.
RNA Isolation and Reverse Transcription
Total RNA was extracted from pools of MII oocytes(100–200) using the Absolutely RNA Microprep Kit(Stratagene). Total RNA was extracted from pools ofembryos (40–80) at the following stages: 2-cell, 4-cell, 8-cell, morula and blastocyst. Oocytes and embryos werewashed three times in PVA-PBS (nuclease free) andlysed in 100-lL lysis buffer (Stratagene), and thenstored at �80�C until total RNA isolation. High-CapacitycDNA Reverse Transcription Kit (ABI) was used forgene cloning. PrimeScriptV
R
RT reagent Kit was used toperform Reverse transcription for Real time-PCR accord-ing to manufacturer’s instructions (TaKaRa).
Quantitative Real-Time PolymeraseChain Reaction
Real-time PCR was performed using the SYBR PremixEx TaqTM (TaKaRa) and the 7300 Real-Time PCR Sys-tem (Applied Biosystems), with the following parame-ters: 95�C for 10 s, followed by 40 cycles at 95�C for 5 sand at 61�C for 34 s. Negative controls in the PCR assayincluding an RT reaction are completed by the same pro-cedure with no templates. For each cDNA sample, bothtarget and reference genes were amplified independentlyon the same plate and in the same experimental run intriplicate.
Statistical Analysis
The levels of gene expression among three groupswere analyzed using analysis of variance (ANOVA) andt-test by Stat View (SAS Institute, Cary, NC); P values<0.05 were considered significantly different.
RESULTSCloning of Porcine Nanog CDS andConstruction of pcDNA3.1 (1)/Nanog andpEGFP-C1/Nanog Vector
The complete cDNA of Nanog was cloned from RNA ofMII stage oocytes by RT-PCR. Confirmed by DNAsequencing, CDS of Nanog was composed of 915 nucleo-tides, and shared 99.8% identity with porcine Nanog inNCBI (GenBank accession no.FJ882402.1), and thereare two nucleotide substitutions at 562 (G to A) and 572(A to G) site. The alignment showed that the similaritiesof Nanog CDS sequence of pig with human, mouse, rat,bovine, goat and monkey were 80.3, 72.4, 72.2, 86.0,86.7, and 79.5%, and the similarities of amino acidsequence were 72.9, 60.5, 61.0, 81.9, 83.3, and 72.6%,respectively. The homeodomain shared 100% identitywith porcine Nanog in NCBI, 98% identity with bovineand goat, and the lowest identity with mouse and rat.The amplified PCR product was subcloned into thepcDNA3.1 (þ) and pEGFP-C1 to produce the pcDNA3.1(þ)/Nanog (PC-Nanog), pEGFP-C1/Nanog (PG-Nanog)vectors. The Blast results of CDS and encoded aminoacids from different species were present in SupportingInformation Figure 1.
Successful Expression of ExogenousNanog in PFF
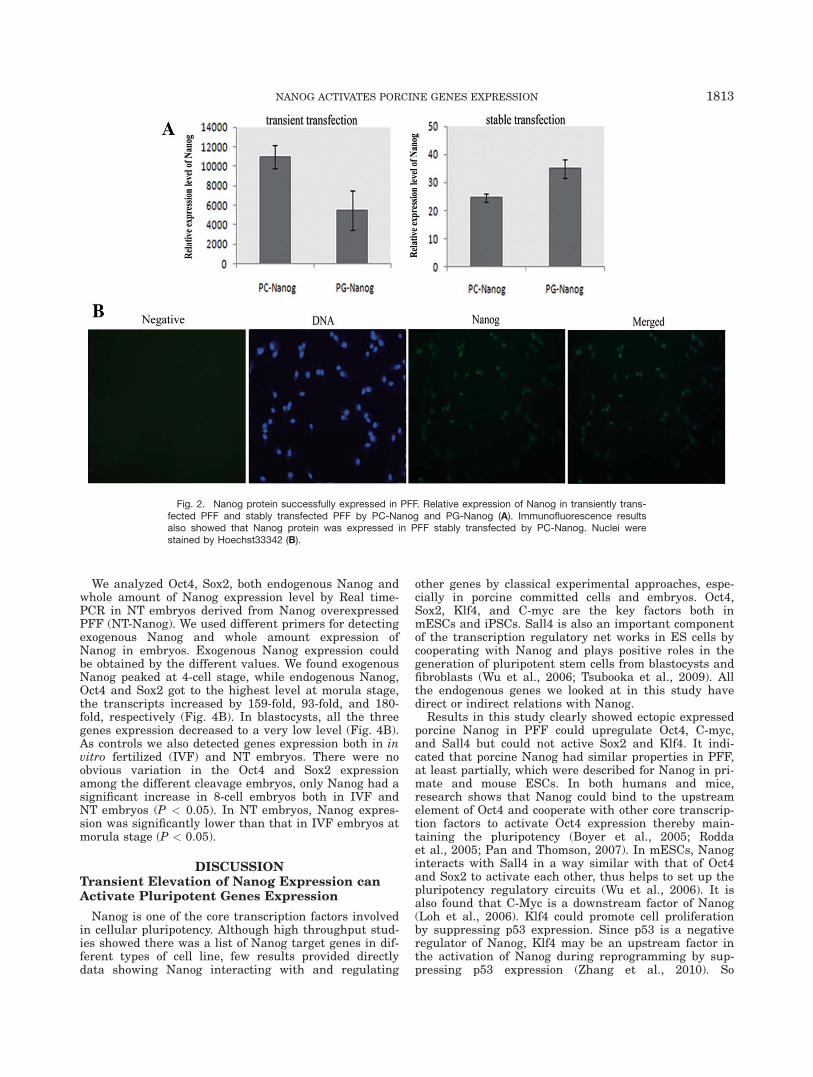
We used PG-Nanog to express GFP-NANOG fusionprotein to confirm the expression and the localization ofexogenous Nanog. After transient transfection, PFFwere transformed from the normal phenotype into prolif-erative phenotype that the shape of cells became roundand small (Fig. 1B). We found the GFP-Nanog was local-ized in the nuclei of both stably and transiently trans-fected PFF (Fig. 1C,D). Real time-PCR resultsdemonstrated that Nanog mRNA was successfullyexpressed in transient and stable transfected PFF (Fig.2A). Immunofluorescence analysis also showed thatNanog protein was expressed in PFF stably transfectedby PC-Nanog (Fig. 2B).
NANOG ACTIVATES PORCINE GENES EXPRESSION 1811

Activation of Pluripotent Genes by NanogOverexpression in PFF
To clarify if Nanog can interact with other pluripotentgenes in PFF, we detected the expression level of Nanogat 1, 5, 10, and 15 days post-transfection without G418selection by Real-time PCR. Compared with the Nanogexpression level in PFF, Nanog expression after tran-sient transfection were 2664, 123, 64, 9-fold at 1, 5, 10,and 15 days, respectively. Nanog peaked at Day 1 anddecreased gradually (Supporting Information Figure 2).Based on these data in our preliminary experiment, weexamined the expression level of pluripotent genes Oct4,Sox2, C-myc, Klf4, and Sall4 at Day 1 after transfectionwhen Nanog expression was at the peak. The expressionlevel of Oct4, Sox2, Klf4, C-myc, and Sall4 were 4.7- and5.4-fold, 1.4- and 0.9-fold, 1.4- and 1.3-fold, 2.2 and 1.6-fold, 4.9- and 1.9-fold in PFF transfected by PC-Nanogand PG-Nanog, respectively. The expression level ofOct4, Sall4, and C-myc increased significantly (P <
0.05), compared with the control group (Fig. 3A). Wethen analyzed the expression level of these pluripotentgenes in transfected PFF after G418 selection for 20days (Fig. 3B).Compared with the control group, Nanogexpression increased by 20-fold (Fig. 2B). But theexpression of Oct4, Sox2, C-myc declined significantly (P< 0.05). The expression of Klf4 and Sall4 changedinsignificantly.
Activation of Endogenous Oct4, Sox2, andNanog in NT Embryos Derived from NanogOverexpressed PFF
We used pcDNA3.1 (þ) empty vector and pcDNA3.1(þ)/Nanog vector stably transfected cells as donor cellsfor SCNT. All reconstructed embryos were in vitro cul-tured at 6.5 days, and there were no statistic differencesin the fusion rate, cleavage rate, and the blastocyst rateand the blastocyst cell numbers among the three groups.
Fig. 1. The morphology of transfected PFF and expression of GFP-Nanog fusion protein. The morphology of pcDNA3.1 (þ) transientlytransfected PFF (A) and pcDNA3.1 (þ)/Nanog (PC-Nanog) transientlytransfected PFF (B). GFP-Nanog fusion protein was localized in the
nuclei of pEGFP-C1/Nanog (PG-Nanog) stably transfected PFF (C)and transiently transfected PFF (D). GFP expression was visualizedunder blue light. Both of cells in (A) and (B) were observed at 24 h af-ter transfection without G418 selection.
1812 ZHANG ET AL.
We analyzed Oct4, Sox2, both endogenous Nanog andwhole amount of Nanog expression level by Real time-PCR in NT embryos derived from Nanog overexpressedPFF (NT-Nanog). We used different primers for detectingexogenous Nanog and whole amount expression ofNanog in embryos. Exogenous Nanog expression couldbe obtained by the different values. We found exogenousNanog peaked at 4-cell stage, while endogenous Nanog,Oct4 and Sox2 got to the highest level at morula stage,the transcripts increased by 159-fold, 93-fold, and 180-fold, respectively (Fig. 4B). In blastocysts, all the threegenes expression decreased to a very low level (Fig. 4B).As controls we also detected genes expression both in invitro fertilized (IVF) and NT embryos. There were noobvious variation in the Oct4 and Sox2 expressionamong the different cleavage embryos, only Nanog had asignificant increase in 8-cell embryos both in IVF andNT embryos (P < 0.05). In NT embryos, Nanog expres-sion was significantly lower than that in IVF embryos atmorula stage (P < 0.05).
DISCUSSIONTransient Elevation of Nanog Expression canActivate Pluripotent Genes Expression
Nanog is one of the core transcription factors involvedin cellular pluripotency. Although high throughput stud-ies showed there was a list of Nanog target genes in dif-ferent types of cell line, few results provided directlydata showing Nanog interacting with and regulating
other genes by classical experimental approaches, espe-cially in porcine committed cells and embryos. Oct4,Sox2, Klf4, and C-myc are the key factors both inmESCs and iPSCs. Sall4 is also an important componentof the transcription regulatory net works in ES cells bycooperating with Nanog and plays positive roles in thegeneration of pluripotent stem cells from blastocysts andfibroblasts (Wu et al., 2006; Tsubooka et al., 2009). Allthe endogenous genes we looked at in this study havedirect or indirect relations with Nanog.
Results in this study clearly showed ectopic expressedporcine Nanog in PFF could upregulate Oct4, C-myc,and Sall4 but could not active Sox2 and Klf4. It indi-cated that porcine Nanog had similar properties in PFF,at least partially, which were described for Nanog in pri-mate and mouse ESCs. In both humans and mice,research shows that Nanog could bind to the upstreamelement of Oct4 and cooperate with other core transcrip-tion factors to activate Oct4 expression thereby main-taining the pluripotency (Boyer et al., 2005; Roddaet al., 2005; Pan and Thomson, 2007). In mESCs, Nanoginteracts with Sall4 in a way similar with that of Oct4and Sox2 to activate each other, thus helps to set up thepluripotency regulatory circuits (Wu et al., 2006). It isalso found that C-Myc is a downstream factor of Nanog(Loh et al., 2006). Klf4 could promote cell proliferationby suppressing p53 expression. Since p53 is a negativeregulator of Nanog, Klf4 may be an upstream factor inthe activation of Nanog during reprogramming by sup-pressing p53 expression (Zhang et al., 2010). So
Fig. 2. Nanog protein successfully expressed in PFF. Relative expression of Nanog in transiently trans-fected PFF and stably transfected PFF by PC-Nanog and PG-Nanog (A). Immunofluorescence resultsalso showed that Nanog protein was expressed in PFF stably transfected by PC-Nanog. Nuclei werestained by Hoechst33342 (B).
NANOG ACTIVATES PORCINE GENES EXPRESSION 1813

transient transfection of Nanog in our study could notactivate Klf4 directly. However, it is unexpected thatNanog could activate Sox2 in NT embryos but could notupregulate Sox2 expression in PFF. One possibility isthat Nanog shares different targets between porcineembryos and fibroblasts. Microarray analysis confirmsthat there is very little overlap of Nanog’s target genesin cell lines derived from different tissues, suggestingthat Nanog’s target genes are variable in a tissue spe-cific manner (Piestun et al., 2006). The genome is deme-thylated in ESCs and embryos, so Nanog is prone tooccupy its target sites in many promoters (Boyer et al.,2005). Another possible explanation is that Nanog is notenough to activate Sox2 in PFF, but along with addi-tional transcription factors existed in embryos to acti-vate Sox2 successfully.
Pluripotent Genes Expression Cannot beMaintained During G418 Selection
In our experiment, after 20 days’ G418 selection, theexpression of Oct4, C-myc and Sall4 gradually silenced,although Nanog could still be maintained at compara-tively high level. We speculate that transient high Nanogexpression can activate part of the pluripotency relatedgenes in PFF, but it is not enough for inducing PFF to afully pluripotent state, thus cannot set up the stable regu-latory circuit. Pan et al. also indicate that the key pluripo-tent factors always work together to control the whole setof pluripotency machinery rather than work individually(Pan and Thomson, 2007). So in a partially transformedPFF, these activated pluripotent genes can be easilysilenced in many ways including DNA highly methylat-ing. And C-myc as the target transcriptional factors ofNanog has the function in loosing chromosome structure,which makes many transcriptional factors prone to
Fig. 3. Relative expression of pluripotent-related genes after tran-sient and stable transfection by PC-Nanog and PG-Nanog in PFF.Genes expression in transiently transfected PFF was detected at 24 hafter transfection and in stably transfected PFF with G418 selectionfor 20 days. Fold difference was calculated with respect to the PFFtransfected by pcDNA3.1 (þ) empty vector. Bars represent the means� standard errors of three independent experiments.
Fig. 4. Constructed embryos using PFF, PC and PC-Nanog stablytransfected PFF as donor cells develop to blastocysts in vitro (�40)(A). Relative expression of endogenous Nanog, whole amount ofNanog, Oct4 and Sox2 in porcine IVF, NT and NT-Nanog embryos atdifferent stages (B). Nanog histogram height of green bars representstotal Nanog mRNA relative expression; Histogram height of purple
bars represents endogenous Nanog expression. Exogenous Nanogexpression could be obtained by the different values, which in figure4B are the green parts. Different subscripts represent significant differ-ences among the three kinds of embryos at the same developmentstages after t-test by Stat View (P values <0.05). Fold difference wascalculated with respect to the MII stage oocytes.
1814 ZHANG ET AL.

binding to the genome DNA site. Following with Nanogexpression decreased intensively, C-myc has an extraordi-nary reduction in PFF, which made other pluripotentgenes (Oct4, Sox2, and Nanog) difficult to bind to the tar-get site in condensed chromosomes. As a result, the tran-scription factors are degraded and represent significantreduction. That might be the reason that Oct4, Sox2, andC-myc were inhibited in Nanog stable expressed PFF.
Exogenous Nanog Activates Endogenous Oct4,Sox2, and Nanog Expression in NT Embryos
Nanog is a critical regulator for embryonic develop-ment and plays an important role in lineage differentia-tion by cooperating with Oct4, which has been defined inmouse, but are still not clearly understood in pig(Strumpf et al., 2005; Messerschmidt and Kemler, 2010).Immunofluorescence studies demonstrate that Nanogexpression cannot be detected in in vitro porcine blasto-cysts and Oct4 is not confined to the ICM, which is notidentical with the studies in mouse (Kuijk et al., 2008).In this study, we find that there is expression of Nanog,Oct4, and Sox2 both in IVF and NT porcine embryosfrom 2-cell to morula stage, which keeps a comparativelystable level except there is an elevation of Nanog at 8-cell stage. However, all of these three genes expressionis downregulated at blastocyst stage, which is partiallycoincident with the previous reports (Magnani andCabot, 2008; Xing et al., 2009). Although overexpressionNanog in transgenic NT embryos could activate endoge-nous Nanog, Oct4, and Sox2 expression and the level ismuch higher than that is in IVF and normal NTembryos, these genes expression is still downregulatedat blastocyst stage. Combining with the result that theexpression of all these three genes have no influence on
the embryos’ development in vitro which is assessed byblastocyst rate and blastocyst cell number, we speculatethat Nanog cooperating with Sox2 and Oct4 may func-tion as a key regulating factor in ICM/trophoblast andepiblast /hypoblast differentiation at a relatively latestage of porcine blastocysts compared with mouseembryos. After all, there exists a long spanning blasto-cyst stage and different developmental process in por-cine embryos. And a recent study also demonstratedthat Nanog was detected later in the epiblast and hypo-blast of spherical blastocysts of pig (Wolf et al., 2011).But in early developmental stages before morula, itmust play other regulatory roles different with that inthe later stage, which is still needed to be identified.
Formation of ES-like Colony Observed inNanog Overexpressed PFF
Another interesting phenomenon in our study is thatwe observed the formation of ES-like colonies (Support-ing Information Figure 3). Both sizes and shapes ofthese colonies were not disciplinary with unclear cellboundaries. By removal of these colonies, the rest of thebasal cells could regenerate colonies through passaging.But after 15–20 days, it would not reoccur. To excludethe possibility that transfection and followed prolongedantibiotic selection cause considerable stress on the cul-tured cells, we also ran the control experiments thattransfection with pcDNA3.1(þ) empty vector in PFF, butwe did not find the colonies that we observed in PFFtransfected either by pcDNA3.1(þ)/Nanog or by pEGFP-C1/Nanog. Previous studies point that overexpressionNanog in fibroblasts results in increased cell prolifera-tion and transformation of foci-forming phenotype(Zhang et al., 2005). Although overexpression Nanog in
TABLE 1. Primers for quantitative Real time-PCR
Gene Primer sequences (50 to 30) Product size (bp) GenBank accession no.
Klf4 F:TGGGTGCGGAGGAACTGCTA 153 NM_001005154.1R:GGGACTGACCTTGGTAATGGAGC
C-myc F:CCCCTACCCGCTCAACGACA 133 NM_001005154.1R:AGCCAAGGGTTCGGGACTGC
Oct4 F:GAAGGTGTTCAGCCAAACGAC 185 NM_001113060.1R:CGATACTTGTCCGCTTTC
Sox2 F:AACCAGAAGAACAGCCCAGAC 155 NM_001123197.1R:TCCGACAAAAGTTTCCACTCG
Sal14 F:ATCCACCTCCGCTCCCATACC 166 NM_001114673.1R:CGTTGCCTGCCGTCATCTTGT
Nanog F:AGCCTCCAGCAGATGCAAGAACTCT 181 FJ882402.1R:TTCTGCCACCTCTTACATTTCATTCG
EndoNanog F:AGCCCCAGCTCCAGTTTCAGC 104 FJ882402.1R:AATGATCGTCACATATCTTCAGGCTGTA
18SrRNA F:GCCCGAAGCGTTTACTTTGA 93 NR_002170.3R:CCGCGGTCCTATTCCATTATT
TABLE 2. Development potential of NT embryos produced by different donor cells
Donor cells
No. ofembryosproduced
No. ofembryos fused
(% � SD)
No. of embryoscleaved(% � SD)
No. ofblastocysts(% � SD)
Nuclearnumbers
of blastocysts
pCDNA transfected PEF 242 168(70.2 � 14.9) 67(40.2 � 11.6) 21(12.1 � 5.0) 49 � 4.6pCDNA3.1(þ)/Nanog transfected PEF 243 210(86.4 � 4.3) 83(39.1 � 20.2) 28(14.1 � 3.4) 50 � 5PEF(F-7) 224 168(75.1 � 9.6) 86(43.8 � 22.8) 33(17.8 � 7.9) 67 � 2.6
NANOG ACTIVATES PORCINE GENES EXPRESSION 1815

human ESCs and MSCs can enhance colony formation(Darr et al., 2006; Liu et al., 2009). These results indi-cate that in PFF the porcine Nanog can promote colonyformation as in human ESCs and MSCs, other than justpromote cell proliferation that also happens in other spe-cies fibroblast. Actually, we examined changes in celldoubling time in Nanog transfected cells and the resultsshowed that Nanog could also promote the proliferationof PFF (Supporting Information Figure 4). We presumethis phenomenon caused by Nanog overexpression maybe also associated with LIF/Stat3 signal pathway,because our results showed there was expression eleva-tion of LIF, LIFr, Gp130, and Stat3 in the Nanog tran-sient overexpression PFF (Supporting InformationFigure 5). Primers are shown in Supporting InformationTable 1. All the information strongly suggests thatNanog should be one of the key factors for porcine iPSCsinduction.
ACKNOWLEDGEMENTS
The authors are grateful to Jun-Yu Ma at NortheastAgricultural University for his valuable discussions. Theauthors are also thankful to colleagues in the ‘‘Lab ofEmbryo Biotechnology’’ for their helpful discussions.
LITERATURE CITED
Blomberg LA, Schreier LL, Talbot NC. 2008. Expression analysis ofpluripotency factors in the undifferentiated porcine inner cellmass and epiblast during in vitro culture. Mol Reprod Dev75:450–463.
Boyer LA, Lee TI, Cole MF, Johnstone SE, Levine SS, Zucker JP,Guenther MG, Kumar RM, Murray HL, Jenner RG, Gifford DK,Melton DA, Jaenisch R, Young RA. 2005. Core transcriptional reg-ulatory circuitry in human embryonic stem cells. Cell 122:947–956.
Brevini TA, Antonini S, Cillo F, Crestan M, Gandolfi F. 2007. Por-cine embryonic stem cells: facts, challenges and hopes. Therioge-nology 68(Suppl 1):S206–S213.
Chambers I, Colby D, Robertson M, Nichols J, Lee S, Tweedie S,Smith A. 2003. Functional expression cloning of Nanog, a pluripo-tency sustaining factor in embryonic stem cells. Cell 113:643–655.
Choi KD, Yu J, Smuga-Otto K, Salvagiotto G, Rehrauer W, Vodya-nik M, Thomson J, Slukvin I. 2009. Hematopoietic and endothe-lial differentiation of human induced pluripotent stem cells. StemCells 27:559–567.
Darr H, Mayshar Y, Benvenisty N. 2006. Overexpression of NANOGin human ES cells enables feeder-free growth while inducingprimitive ectoderm features. Development 133:1193–1201.
Esteban MA, Xu J, Yang J, Peng M, Qin D, Li W, Jiang Z, Chen J,Deng K, Zhong M, Cai J, Lai L, Pei D. 2009. Generation ofinduced pluripotent stem cell lines from Tibetan miniature pig. JBiol Chem 284:17634–17640.
Ezashi T, Telugu BP, Alexenko AP, Sachdev S, Sinha S, RobertsRM. 2009. Derivation of induced pluripotent stem cells from pigsomatic cells. Proc Natl Acad Sci USA 106:10993–10998.
Goel S, Fujihara M, Minami N, Yamada M, Imai H. 2008. Expres-sion of NANOG, but not POU5F1, points to the stem cell potentialof primitive germ cells in neonatal pig testis. Reproduction135:785–795.
Hall V. 2008. Porcine embryonic stem cells: a possible source for cellreplacement therapy. Stem Cell Rev 4:275–282.
Hall VJ, Christensen J, Gao Y, Schmidt MH, Hyttel P. 2009. Porcinepluripotency cell signaling develops from the inner cell mass tothe epiblast during early development. Dev Dyn 238:2014–2024.
Hanna J, Saha K, Pando B, van Zon J, Lengner CJ, Creyghton MP,van Oudenaarden A, Jaenisch R. 2009. Direct cell reprogramming
is a stochastic process amenable to acceleration. Nature 462:595–601.
Kim HS, Son HY, Kim S, Lee GS, Park CH, Kang SK, Lee BC,Hwang WS, Lee CK. 2007. Isolation and initial culture of porcineinner cell masses derived from in vitro-produced blastocysts. Zy-gote 15:55–63.
Kim S, Kim JH, Lee E, Jeong YW, Hossein MS, Park SM, Park SW,Lee JY, Jeong YI, Kim HS, Kim YW, Hyun SH, Hwang WS. 2010.Establishment and characterization of embryonic stem-like cellsfrom porcine somatic cell nuclear transfer blastocysts. Zygote18:93–101.
Kuijk EW, Du Puy L, Van Tol HT, Oei CH, Haagsman HP,Colenbrander B, Roelen BA. 2008. Differences in early lineagesegregation between mammals. Dev Dyn 237:918–927.
Li M, Li YH, Hou Y, Sun XF, Sun Q, Wang WH. 2004. Isolation andculture of pluripotent cells from in vitro produced porcineembryos. Zygote 12:43–48.
Liu TM, Wu YN, Guo XM, Hui JH, Lee EH, Lim B. 2009. Effects ofectopic Nanog and Oct4 overexpression on mesenchymal stemcells. Stem Cells Dev 18:1013–1022.
Loh YH, Wu Q, Chew JL, Vega VB, Zhang W, Chen X, Bourque G,George J, Leong B, Liu J, Wong KY, Sung KW, Lee CW, Zhao XD,Chiu KP, Lipovich L, Kuznetsov VA, Robson P, Stanton LW, WeiCL, Ruan Y, Lim B, Ng HH. 2006. The Oct4 and Nanog transcrip-tion network regulates pluripotency in mouse embryonic stemcells. Nat Genet 38:431–440.
Magnani L, Cabot RA. 2008. In vitro and in vivo derived porcineembryos possess similar, but not identical, patterns of Oct4,Nanog, and Sox2 mRNA expression during cleavage development.Mol Reprod Dev 75:1726–1735.
Messerschmidt DM, Kemler R. 2010. Nanog is required for primi-tive endoderm formation through a non-cell autonomous mecha-nism. Dev Biol 344:129–137.
Mitsui K, Tokuzawa Y, Itoh H, Segawa K, Murakami M, TakahashiK, Maruyama M, Maeda M, Yamanaka S. 2003. The homeopro-tein Nanog is required for maintenance of pluripotency in mouseepiblast and ES cells. Cell 113:631–642.
Okita K, Ichisaka T, Yamanaka S. 2007. Generation of germline-competent induced pluripotent stem cells. Nature 448:313–317.
Pan G, Thomson JA. 2007. Nanog and transcriptional networks inembryonic stem cell pluripotency. Cell Res 17:42–49.
Piestun D, Kochupurakkal BS, Jacob-Hirsch J, Zeligson S, Koudrit-sky M, Domany E, Amariglio N, Rechavi G, Givol D. 2006. Nanogtransforms NIH3T3 cells and targets cell-type restricted genes.Biochem Biophys Res Commun 343:279–285.
Rodda DJ, Chew JL, Lim LH, Loh YH, Wang B, Ng HH, Robson P.2005. Transcriptional regulation of nanog by OCT4 and SOX2. JBiol Chem 280:24731–24737.
Silva J, Barrandon O, Nichols J, Kawaguchi J, Theunissen TW,Smith A. 2008. Promotion of reprogramming to ground state plu-ripotency by signal inhibition. PLoS Biol 6(10):e253. doi:10.1371/journal.pbio.0060253.
Silva J, Nichols J, Theunissen TW, Guo G, van Oosten AL, Barran-don O, Wray J, Yamanaka S, Chambers I, Smith A. 2009. Nanogis the gateway to the pluripotent ground state. Cell 138:722–737.
Strumpf D, Mao CA, Yamanaka Y, Ralston A, ChawengsaksophakK, Beck F, Rossant J. 2005. Cdx2 is required for correct cell fatespecification and differentiation of trophectoderm in the mouseblastocyst. Development 132:2093–2102.
Takahashi K, Yamanaka S. 2006. Induction of pluripotent stem cellsfrom mouse embryonic and adult fibroblast cultures by definedfactors. Cell 126:663–676.
Tsubooka N, Ichisaka T, Okita K, Takahashi K, Nakagawa M,Yamanaka S. 2009. Roles of Sall4 in the generation of pluripotentstem cells from blastocysts and fibroblasts. Genes Cells 14:683–694.
West FD, Terlouw SL, Kwon DJ, Mumaw JL, Dhara SK, Hasneen K,Dobrinsky JR, Stice SL. 2010. Porcine induced pluripotent stemcells produce chimeric offspring. Stem Cells Dev 19:1211–1220.
Wolf XA, Serup P, Hyttel P. 2011. Three-dimensional localisation ofNANOG, OCT4, and E-CADHERIN in porcine pre- and peri-im-plantation embryos. Dev Dyn 240:204–210.
1816 ZHANG ET AL.

Wu Q, Chen X, Zhang J, Loh YH, Low TY, Zhang W, Sze SK, LimB, Ng HH. 2006. Sall4 interacts with Nanog and co-occupiesNanog genomic sites in embryonic stem cells. J Biol Chem281:24090–24094.
Wu Z, Chen J, Ren J, Bao L, Liao J, Cui C, Rao L, Li H, Gu Y, DaiH, Zhu H, Teng X, Cheng L, Xiao L. 2009. Generation of piginduced pluripotent stem cells with a drug-inducible system. JMol Cell Biol 1:46–54.
Xing X, Magnani L, Lee K, Wang C, Cabot RA, Machaty Z. 2009.Gene expression and development of early pig embryos producedby serial nuclear transfer. Mol Reprod Dev 76:555–563.
Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, FraneJL, Tian S, Nie J, Jonsdottir GA, Ruotti V, Stewart R, Slukvin, II,Thomson JA. 2007. Induced pluripotent stem cell lines derivedfrom human somatic cells. Science 318:1917–1920.
Zhang J, Wang X, Chen B, Suo G, Zhao Y, Duan Z, Dai J. 2005.Expression of Nanog gene promotes NIH3T3 cell proliferation.Biochem Biophys Res Commun 338:1098–1102.
Zhang P, Andrianakos R, Yang Y, Liu C, Lu W. 2010. Kruppel-likefactor 4 (Klf4) prevents embryonic stem (ES) cell differentiationby regulating Nanog gene expression. J Biol Chem 285:9180–9189.
NANOG ACTIVATES PORCINE GENES EXPRESSION 1817