oume 9 ume 3 ie i e usa (iss eoiyie oweguaes iic oie a umo ...ila.ilsl.br/pdfs/v69n3a07.pdf ·...
TRANSCRIPT

Volume 69. Number 3Printed in the U.S.A.
(ISSN 0148-916X)
INTERNATIONAL JOURNAI. OI , LEPROSY
Pentoxifylline Downregulates Nitric Oxide and TumorNecrosis Factor-a Induced by Mycobacterial
Lipoarabinomannan in a Macrophage Cell Line'Eunkyue Park, Georgia Schuller-Levis, Seung Yong Park,
Jun Hua Jia, and William IR. Levis 2
Pentoxifylline (PTX) is a phosphodi-esterase inhibitor that is FDA approved forthe treatment of peripheral vascular disease(38), There has been enthusiasm for the useof PTX as a potential anti-inflammatoryagent because of its ability to downregulatethe powerful proinllammatory molecule tu-mor necrosis factor-alpha (TNF-a) ( 19,24 33).
Reports in mice ( 29 ' 30 . 31 ) and humans (30) in-dicate possible efficacy in the treatment ofallergic contact dermatitis, rheumatoid arth-ritis and Mycobacterium avium exacerba-tion in late-stage HIV infection ( 5 ' 21 ). Inaddition, two promising reports have ap-peared in the treatment of leprosy reactions( 26.227 ' 35 ) where TNF-a has been identified asa major proinflammatory molecule (19.25).Lipoarabinomannan (LAM), the major car-bohydrate antigen in mycobacterial cellwalls, has been shown to induce TNF-a andnitric oxide (NO) in mouse macrophages,including the mouse macrophage cell lineRAW 264.7 ( 28). These findings provide uswith an in vitro model to further explore theeffects of PTX on the macrophage at thecellular and molecular level.
'Received for publication on I May 2000. Ac-cepted for publication in revised form on 2 July 2001.
'E. Park, Ph.D.; G. Schuller-Levis, Ph.D.; J. H. Jia;C. Kim. Ph.D., Department of Immunology, New YorkState Institute for Basic Research in DevelopmentalDisabilities, 1050 Forest Hill Road. Staten Island, NewYork 103 14, U.S.A. S. Y. Park, D.V.M.. Ph.D., Depart-ment of Veterinary Medicine, Konkuk University,Seoul. Korea 143-701. W. R. Levis, M.D., Departmentof Immunology, New York State Institute for BasicResearch in Developmental Disabilities, New Yorkand Department of Dermatology, Staten Island Uni-versity Hospital. Staten Island. New York, U.S.A.
Reprint requests to G. Schuller-Levis, Ph.D., at theaddress above or FAX 1-718-494-4884; c-mail:[email protected]
MATERIALS AND METHODS
Reagents. Li popolysaccharide (LPS) W(Escherichia co/i 01 11:84) was purchasedfrom Difco Laboratories, Detroit, Michi-gan, U.S.A. Arabinose lipoarabinomannan(ara-LAM), a rapidly growing Mycobacte-rium sp. or M. iirberculosis H37Ra, was agift from Dr. John Belisle, University ofColorado, Fort Collins, Colorado, U.S.A.Murine iNOS cDNA probe and murineTNF-a probe were graciously provided byDr. Carl Nathan, Cornell University, NewYork, New York. U.S.A. and Chiron Corpo-ration, Emeryville, California, U.S.A., re-spectively. RPM' 1640, murine recombi-nant interferon-gamma (rIFN-y), fetal calfserum (FCS), penicillin and streptomycinand RNA size markers were obtained fromGibco-BRL, Grand Island, New York,U.S.A. Nylon membranes (GenescreenPlus) for Northern blot analyses were pur-chased from Dupont, Boston, Massachu-setts, U.S.A., and Tri-Reagent was pur-chased from Molecular Research Center,Cincinnati, Ohio, U.S.A. Agarose was ob-tained from FMC Bioproducts, Rockland,Maine, U.S.A. Herring sperm DNA waspurchased from Roche Molecular Biochem-icals, Indianapolis, Indiana, U.S.A. andDenhardt reagent and 32P-dCTP was fromAmersham Pharmacia, Piscataway, NewJersey, U.S.A. Acrylamide/bis solution(30%), molecular standards of protein andglycine were purchased from Bio-Rad Lab-oratories, Hercules, California, U.S.A. Ni-trocellulose membrane for Western blotanalysis was obtained from Schlecher-Schuell, Keene, New Hampshire, U.S.A.Rabbit anti-mouse iNOS was from BDTransduction Laboratories, San Diego, Cal-ifornia, U.S.A., and alkaline phosphatase
225

226 International Journal of Lepmsy 2001
labeled sheep anti-rabbit antibody was fromRoche Molecular Biochemicals. PTX,formaldehyde, formamide and morpholino-propanesulfonic acid were obtained fromSigma Chemical Company, St. Louis, Mis-souri, U.S.A. Murine recombinant TNF -
alpha (rTNF-a) and monoclonal banisteranti - murine TNF-a were purchased fromR&D Systems, Minneapolis, Minnesota,U.S.A., and polyclonal rabbit anti-murineTNF-a was purchased from Endogen,Cambridge, Massachusetts, U.S.A.
Cell culture and stimulation. RAW264.7 cells, an Abelson leukemia virus -transformed murinene macrophage cell line(American Type Culture Collection, Rock-ville, Maryland, U.S.A.) were cultured inRPMI 1640 containing 10% heat - inactivatedfetal calf serum (FCS) and antibiotics (com-plete medium). Cells were grown in com-plete RPMI 1640 for maintenance and incomplete DMEM without phenol red for ni-trite and TNF-oc measurements. Cells (2 x105/well) were added to a 96-well plate andadhered overnight for functional assays.Various amounts of PTX (0.05 mg/ml, 0.1mg/ml, 0.5 mg/ml and 1.0 mg/ml) and acti-vators (LPS 1 pg/ml or LAM 1 pg/m1 con-taining IFN-y 50 U/ml) were added and in-cubated for 24 hr. One -hundred Ill wereused for nitrite determination and 501.11 forTNF-a measurement. For Western andNorthern analyses, 1 x 10 7 cells were cul-tured in Petri dishes. Cell viability, moni-tored by both trypan blue exclusion andMTT viability assays, was unaffected by theconcentrations of PTX used in these studies.
Nitrite determination. Nitrite was mea-sured by the method described previously( 2 . ' 7 ). Briefly, 100 pl of the supernatantfrom a 96-well plate culture was mixedwith an equal volume of Griess reagent (I%sulfanilamide, 0.1% naphthylethylene di-amine dihydrochloride and 2.5% phosphoricacid) and incubated at room temperature for10 min. The optical density (OD) of eachsample was read at 550 nm by a microplatereader (Cambridge Technology, Inc., Cam-bridge, Massachusetts, U.S.A.). The calibra-tion curve was prepared using sodium nitriteas a standard.
TNF-a measurement by ELISA. TNF-asecretion was measured by ELISA as de-scribed previously ( 17 . 34). Briefly, 50 pl ofhamster anti-mouse TNF-a monoclonal
antibody (R&D Systems), diluted 1:500 incarbonate buffer (pH 9.6), was incubated in96-well ELISA plate (Dynatech Laborato-ries, Chantilly, Virginia, U.S.A.) overnightat 4°C. After washing the plate, 2% BSA inPBS-Tween 20 (T-PBS, 0.1 M PBS with0.05% Tween) was added, and the plate wasincubated at 37°C for 2 hr. The plate waswashed with T PBS and 501.11 of either vari-ous concentrations of standard murinerTNF-a or diluted samples were added tothe plates. After incubation at 4°C overnightand washing with T-PBS, 50 IA of rabbitanti-mouse TNF-a polyclonal antibody, di-luted 1:500 in T-PBS, was added and theplate was incubated at 37°C for 1.5 hr. Afterwashing, the plate was incubated with 50 p1of goat anti-rabbit immunoglobulin Ghorseradish perox i dase conjugate (ZymedLaboratories, San Francisco, California,U.S.A.) at a 1:5000 dilution for 1 hr. Thecolor was developed with 0-phenylenedi-amine dihydrochloride and H 2 O,, in citratebuffer (pH 5.0). The plate was read at 490nm by an MR 600 microplate reader (Dy-natech Laboratories). The concentration ofTNF-a released into the medium was cal-culated by using murine rTNF-a as a stan-dard.
Western blot analyses. Western blotanalysis was performed by the method de-scribed previously ( 18 ). Briefly, activatedRAW 264.7 cells were washed with Hanksbalanced salt solution (HBSS). The cellswere lysed with HBSS containing 1.0%Nonidet P-40, aprotinin (10 pg/ml), anti-pain (5 pg/ml), and PMSF (0.2 mM). Aftercentrifugation, cell lysates (50 pg protein)were diluted in SDS-containing samplepreparation buffer, electrophoresed on 7.5%SDS polyacrylamide gels, and transferredovernight to nitrocellulose filters. The nitro-cellulose filters were blocked with a solu-tion containing 2% BSA and 2% normalsheep serum for 1 hr at room temperatureand washed with 0.2% Tween 20 in PBSonce for 5 min. The blot was incubated withrabbit inducible nitric oxide synthase(iNOS) antiserum (1/5000 dilution) as a pri-mary antibody for 2 hr at 37°C, and washedthree additional tines. After incubationwith alkaline phosphatase - labeled sheepanti-rabbit antibody for 1.5 hr at 37°C theblot was again washed three times. The re-action product was visualized using nitro-
Pentoxifylline conc.. mg/ml Pentoxifylline conc., mg/ml
0 0.05 0.1 0.5 1.0
6
5 -
0 0.05 0.1 0.5 1.0
20
35
30 - __L
20 -
15 -
10 -
I ^0
A
4
3
2
0 0.05 0.1 0.5 1.0
B
5 -- IL
3 —
2 —
0.05 0.1 0.5o o 1.0
0
6 B
69, 3 Park, et al.: Pentoxifylline and Macrophages 227
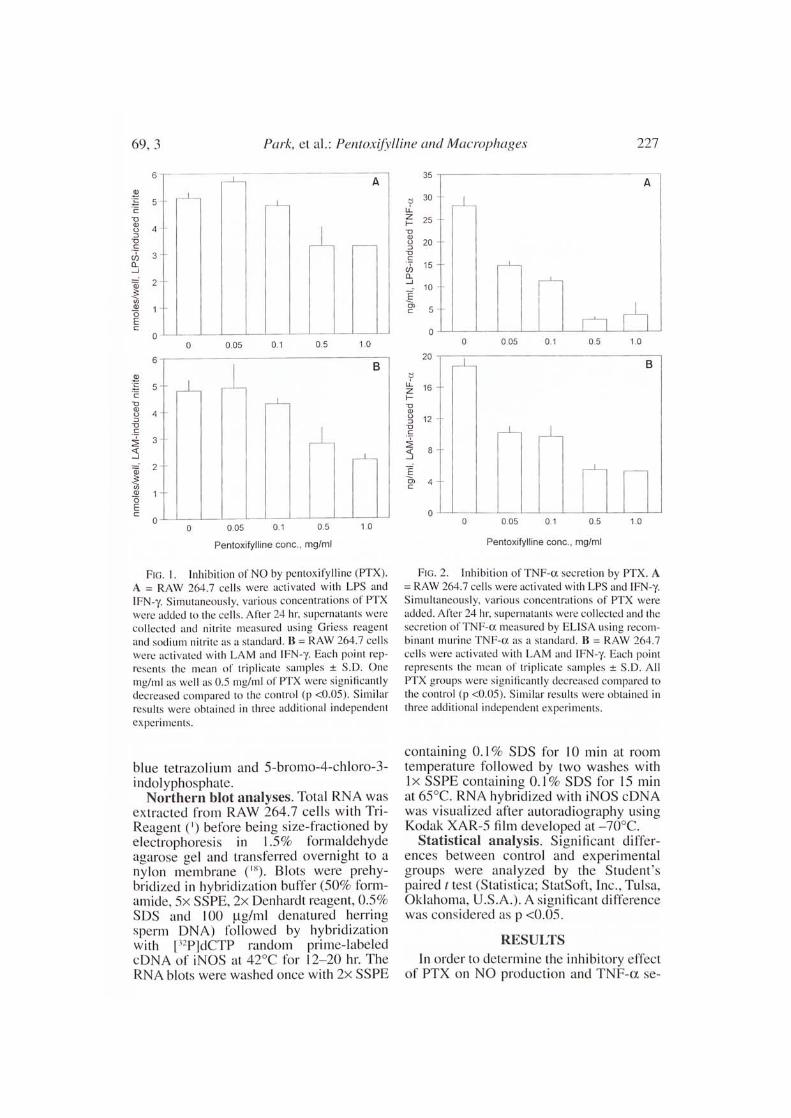
FIG. I. Inhibition of NO by pentoxifylline (PTX).A = RAW 264.7 cells were activated with LPS andIFN-y. Simutaneously, various concentrations of PTXwere added to the cells. After 24 hr, supernatants werecollected and nitrite measured using Griess reagentand sodium nitrite as a standard. B = RAW 264.7 cellswere activated with LAM and IFN-y. Each point rep-resents the mean of triplicate samples ± S.D. Onemg/ml as well as 0.5 mg/n11 of PTX were significantlydecreased compared to the control (p <0.05). Similarresults were obtained in three additional independent
experiments.
blue tetrazolium and 5-bromo-4-chloro-3-Indolyphosphate.
Northern blot analyses. Total RNA wasextracted from RAW 264.7 cells with Tri-Reagent (') before being size-fractioned byelectrophoresis in 1.5% formaldehydeagarose gel and transferred overnight to anylon membrane ('Y). Blots were prehy-bridized in hybridization buffer (50% form-amide, 5x SSPE, 2x Denhardt reagent, 0.5%SDS and 100 lg/ml denatured herringsperm DNA) followed by hybridizationwith I 32PIdCTP random prime-labeledcDNA of iNOS at 42°C for 12-20 hr. TheRNA blots were washed once with 2x SSPE
FIG. 2. Inhibition of TNF-a secretion by PTX. A= RAW 264.7 cells were activated with LPS and IFN-y.Simultaneously, various concentrations of PTX wereadded. After 24 hr, supernatants were collected and thesecretion of TNF-a measured by ELISA using recom-
binant murinc TNF-a as a standard. B = RAW 264.7cells were activated with LAM and IFN-y. Each pointrepresents the mean of triplicate samples ± S.D. AllPTX groups were significantly decreased compared tothe control (p <0.05). Similar results were obtained inthree additional independent experiments.
containing 0.1% SDS for 10 min at roomtemperature followed by two washes withlx SSPE containing 0.1% SDS for 15 minat 65°C. RNA hybridized with iNOS cDNAwas visualized after autoradiography usingKodak XAR-5 film developed at —70°C.
Statistical analysis. Significant differ-ences between control and experimentalgroups were analyzed by the Student'spaired t test (Statistica; StatSoft, Inc., Tulsa,Oklahoma, U.S.A.). A signilicant differencewas considered as p <0.05.
RESULTSIn order to determine the inhibitory effect
of PTX on NO production and TNF-a se-

228 International .I0c1r7ral of Leprosy 2001
A
2O8-
13.3-
8.7-
B1 2 3 4 5 1 2 3 4 5
20.8-
13.3-
8.7-
LPS+I - + + + + LAM+I - + + + +
PTX - - 0.1 0.5 1.0 PTX - - 0.1 0.5 1.0(mg/ml) (mg/ml)
FIG. 3. Western blot analyses of iNOS. A = RAW 264.7 cells were activated with LPS and IFN-y. Lanes: 1 =unactivated cells, 2 = LPS + IFN-y only. 3 = LPS + IFN-y + PTX 0.1 mg/m1, 4 = LPS + IFN-y + PTX 0.5 mg/ml,5 = LPS + IFN-y + PTX 1.0 mg/ml. B = RAW 264.7 cells were activated with LAM and IFN-y. Lanes: 1 = unac-tivated cells, 2 = LAM + IFN-y only. 3 = LAM + IFN-y + PTX 0.1 mg/ml, 4 = LAM + IFN-y + PTX 0.5 mg/ml, 5= LAM + IFN-y + PTX 1.0 mg/ml. Similar results were obtained in two additional independent experiments.
cretion in LAM activated RAW 264.7 cells,various amounts of PTX (0.05 mg/ml, 0.1mg/ml, 0.5 mg/ml and 1.0 mg/m1) wereused simultaneously when the cells wereactivated with LFN-y (50 U/ml) and eitherLPS (1 µg/m1) or LAM (1 1.1g/m1). PTX in-hibited NO induced by LPS (Fig. IA) ( b0).
PTX inhibited the production of NO in-duced by LAM in a dose-dependent manner(Fig. 1B). Even though 0.05 mg/ml and 0.1mg/ml of PTX was not effective, 0.5 mg/m1and 1.0 mg/ml significantly inhibited NOproduction (p <0.01). The inhibition ofTNF-a was greater than that of NO by PTXwhen the cells were activated with eitherLPS and IFN-y or LAM and IFN-y (Figs.2A and 2B). Even as little as 0.05 mg/ml ofPTX inhibited the secretion of TNF-a sig-nificantly (p <0.001). In contrast. NO wasnot significantly inhibited at either 0.05mg/ml or 0.1 m g/m 1 PTX .
Since PTX inhibited the production ofNO induced by LAM or LPS in RAW 264.7cells, iNOS protein was examined by West-
ern blot analyses. RAW 264.7 cells weretreated with PTX (0.1, 0.5 and 1.0 mg/m1)in the presence of either LPS or LAM withIFN-y and separated by SDS-PAGE aftercell lysis. iNOS proteins induced by LPS orLAM with IFN-y were inhibited by PTX(Figs. 3A and 3B). At 0.1 mg/ml of PTX,NOS protein induced by LPS and IFN-ywas actually increased. iNOS protein in-duced by LAM and IFN-y was decreased ina dose-dependent manner. Untreated cellsdid not show an iNOS protein band.
Since iNOS and TNF-a proteins were in-hibited by PTX, Northern blot analyseswere performed to determine whether PTXcould inhibit transcription of mRNA ofiNOS and TNF-a. After extraction of totalRNA and separation of total RNA byagarose gel, specific 32 P-labeled cDNAprobes were used to detect the niRNA ofiNOS and TNF-a, using a-actin as a con-trol. All cell preparations for Northern blotanalyses were activated with LPS or LAM,including IFN-y, for 16 hr, which gives

a-actin
LPS+1 -PTX
(mg/ml)
R.O.D. 0
+ +
0.5 1.0
5.7 5.7
NOS NOS
a-actin
LAM+1 - + + + +PTX
(mg/mI)- 0.1 0.5 1.0
R.O.D. 0 3.6 3.5 3.6 3.6
69, 3 Park, et al.: Pentoxi/yl/ine and Macrophages 229
A 1 2 3 4 5 B 1 2 3 4 5
Ftc. 4. Northern blot analyses of iNOS. A = RAW 264.7 cells were activated with LPS and IFN-y. Lanes: 1= unactivated cells, 2 = LPS + IFN-y only, 3 = LPS + IFN-y + PTX 0.1 mg/ml, 4 = LPS + IFN-y + PTX 0.5mg/ml, 5 = LPS + IFN-y+ PTX 1.0 mg/m1. B = RAW 264.7 cells were activated with LAM and IFN-y. Lanes: 1= unactivated cells, 2 = LAM + IFN-y only, 3 = LAM + IFN-y + PTX 0.1 mg/ml, 4 = LAM + IFN-y + PTX 0.5mg/ml, 5 = LAM + IFN-y+ PTX 1.0 mg/ml. Similar results were obtained in two additional independent exper-iments. Relative optical density (ROD) was performed using a-actin as a control.
maximum expression of iNOS mRNA ( 18 ).When the data were normalized using rela-tive optical density (ROD), PTX failed toinhibit the transcription of iNOS induced byLPS at 0.5 mg/ml or 1.0 mg/ml (Fig. 4A).Similarly, when the data were normalizedusing ROD, iNOS mRNA induced by LAMwas not inhibited at any dose tested (Fig.4B). PTX inhibited the transcription ofTNF-u. induced by LPS as well as by LAM(Figs. 5A and 5B).
DISCUSSIONOur findings with LPS and PTX are in
keeping with prior studies that show adownregulation of TNF-a at both theproduct and transcriptional levels and adownregulation of NO at the product level(4. 10. 12. 36 )
. In addition, we extended these
findings to show similar effects whenmacrophages are activated with LAM, themajor carbohydrate antigen of mycobacte-rial cell walls (2832
) . Our findings also showsome downregulation of iNOS with PTX atthe translational level when activated with
LPS or LAM. In fact, at low concentrationsof PTX there was a slight increase of iNOSat the translational level (Fig. 3A) when thecells were activated with LPS.
Our findings indicate a possible problemwith the use of PTX as an immunosuppres-sive or anti-inflammatory agent in derma-tology. NO as an important proinflamma-tory component of the immune system ( 1422°)is not suppressed by PTX at the transcrip-tional level. Furthermore, even at the prod-uct level, high concentrations (0.5-1.0mg/ml) were required to suppress NO pro-duction. However, TNF-a was inhibited byPTX at 50 ftg/ml. The possible benefit ofPTX as an anti-inflammatory/immunosup-pressive agent has not been limited to der-matology, but has been used in diverse in-flammatory conditions such as cerebralmalaria (') and bacterial meningitis ( 37). Asdiscussed by Van Furth, cal al., some contro-versy exists on the efficacy of PTX on sup-pressing IL-6 and IL-8 ( 37). Neuner, et al.showed a decrease in IL-8 and IL-6 as wellas TNF-u when leukocytes were pretreated

A ., B 1 2 3 4 51 2 3 4 5
230 International Journal of Leprosy 2001
TNF TNF
a -actin a -actin
LPS+1 — + + + +PTX _ — 0.1 0.5 in
LAM+1 —PTX —
(mg/ml) (mg/ml)
R.O.D. 0 2.0 t5 1.1 1.0 R.O.D. 0
+ + + +
— 0.1 0.5 1.0
1.4 1.3 1.1 0.9
FIG. 5. Northern blot analyses of TNF-a. A = RAW 264.7 cells were activated with LPS and IFN-y. Lanes:1 = unactivated cells, 2 = LPS + IFN-y only, 3 = LPS + IFN-y + PTX 0.1 mg/ml, 4 = LPS + IFN-y + PTX 0.5mg/ml, 5 = LPS + IFN-y + PTX 1.0 mg/ml. B = RAW 264.7 cells were activated with LAM and IFN-y. Lanes: I =unactivated cells, 2 = LAM + IFN-y only, 3 = LAM + IFN-y + PTX 0.1 mg/ml, 4 = LAM + IFN-y + PTX 0.5mg/ml, 5 = LAM + IFN-y+ PTX 1.0 mg/ml. Similar results were obtained in two additional independent exper-iments. Relative optical density (ROD) was performed using a-actin as a control.
for 24 hr ('h). However, in keeping withprior studies without 24 hr pretreatment,IL-6 and IL-8 were not downregulated incontrast to TNF-a. In fact, the mechanismof PTX as an anti-inflammatory/immuno-suppressive agent is not as simple as ana-lyzing only product and transcriptional lev-els. Han, et al. have shown that dexametha-sone strongly impedes translationalderepression of NO and TNF-a in sharpcontrast to PTX ( 7 ).
Finally, since LAM plays a role inmacrophage activation, we need to interpretour findings in the mycobacterial model atthe clinical, immunologic and molecularlevels. LAM preparations from nonvirulentspecies of mycobacteria have been shownto induce elevated levels of TNF-a at theproduct level; whereas virulent (e.g., M. tu-berculosis Erdman and M. leprae) inducethe TNF-a message in the absence of prod-uct ( 22). Future studies need to address thestructural (e.g., mannose capping of viru-lent LAMs) and functional relationships to
mycobactericidal activity in IFN-y-primedmacrophages. Leprosy has a spectrum ofimmunologic disease ( 22) from a Th 1 domi-nant tuberculoid form (IFN-y positive) to aTh2 dominant lepromatous form (IFN-ynegative) ( 13 ' 32), including two types of re-actions [type 1 or reversal reactions andtype 2 or erythema nodosum leprosurn(ENL) reactions], both of which are cur-rently best treated with corticosteroids or intype 2 reactions, thalidomide and/or corti-costeroids (11 25.26) . PTX has been suggestedto theoretically benefit ENL reactions (19,24)
and, indeed, there have been two reports ofpossible benefit, although both reports in-cluded patients who required additionalcorticosteroids ( 7 ' x).
This has also been our experience in theNew York regional program, and similarexperience at the National Hansen's Dis-ease Center (personal communication, Dr.Leo Yoder, National Hansen's Disease Cen-ter, Carville, Louisiana, U.S.A.) that somepatients while receiving PTX appear to im-

69, 3 Park, et al.: Pentoxi/vlline and Macrophages 231
prove but then suddenly exacerbate on PTXand require high doses of corticosteroids.These observations may reflect a decreaseof both TNF-a and NO at the product level,but with only partial inhibition of NO.Thus, the possibility of a sudden break-through and leprosy reaction exists withPTX. In contrast, corticosteroids have beenshown to powerfully downregulate allproinflammatory cytokines at both theproduct and transcriptional levels, includingTNF-a and iNOS (3 ' 8 ' 9), which would avoidthe problem of a late (60 days) excerbationwith 20% of patients treated with pentoxi-fylline, as described by Nery, et aI. ( 15 )
SUMMARYPentoxifvlline (PTX), a phosphodi-
esterase inhibitor, is known to downregu-late tumor necrosis factor-alpha (TNF-a)secretion induced by lipopolysacchride(LPS) and gamma interferon (MN-7). Wehave had limited success in treating leprosyreactions, including erythema nodosum lep-rosum (ENL), in which TNF-a has beenidentified as a major proinflammatory cy-tokine. PTX inhibited production of NO(IC50 _ 1.0 mg/ml) and TNF-a (IC500.05 mg/ml) in a dose-dependent fashion.As little as 0.5 mg/ml of PTX decreasedNO production and 0.01 mg/m1 of PTX in-hibited TNF-a production. Western blotanalyses demonstrated that iNOS was sup-pressed by PTX. Northern blot analysesshowed significant reduction of TNF-amRNA. We conclude that PTX is an effectiveinhibitor of lipoarabinomannan (LAM)-induced TNF-a production at both theproduct and transcriptional levels in ourmacrophage cell line. PTX also showedmoderate inhibition of NO at the productlevel as well as translation of iNOS.
RESUMENSe sabe que Ia pentoxililina (PTX). un inhibidor de
fosfodiesterasas. disminuye Ia secreción de factor denecrosis tumoral alfa (TNFa.) inducida porIIpopolisacárido (LPS) e interferón ganona (IFNy).Nosotros hemos tenido un éxito limitado en eItratamiento (Ic reacciones leprosas, incluyendo eritemanodoso leproso, en Ias cuales eI TNFa se ha identifi-cado como la principal citocina proinflamatoria. LaPTX inhihió la producción de óxido nítrico (NO)(1050 = 1.0 mg/m1) y de TNFu IC50 = 0.05 mg/ml) enforma dosis-dependiente. Cantidades de PTX tan pe-
querias conto 0.5 mg/ml y 0.01 mg/ml, inhibieron laproducción de NO y de TNFa, respectivamente. Elanálisis por Western blot detnostró que la PTXsuprimió la producción de oxido-nítrico sintasa in-ducible (iNOS) micntras que el andlisis por Northernblot indicó una reducción significante en la síntesis demRNA para TNFa. Concluimos que en nuestra líneade macrófagos, la PTX es un inhibidor efectivo de laproducción de TNFa inducida por lipoarabinomanana(LAM) tanto al nivel de producto como al nivel tan-scripcional. La PTX también mostró una inhibiciónmoderada de NO tanto a nivel de producto como anivel de la traducción de iNOS.
RÉSUMÉPentoxifylline (PTX), un inhihiteur des phosphodi-
estérases, est capable de diminua la sécrétion du fac-teur alpha de nécrose des tumeurs (TNF-u) induite parles lipopolysaccharides (LPS) et l'interféron gamma(IFN-y). Un succès limité a did observé dans le traite-mcnt des réactions immunopathologiques hanséni-cones, y compris I'érythème noueux ldpreux, chez quiTNF-a est coam comme un acteur majeur parmi lescytokines pro-inllammatoires. PTX a inhibé de façondose-dépendante la production de monoxyde d'azote(NO) (concentration inhibitrice 50% ou IC50 — = 1,0mg/ml) et de TNF-a (IC50 = 0,05 mg/ml). Une quan-tité aussi faible que 0.5 mg/kg et que 0.01 mg/kg dePTX a diminué la production de NO et inhibé Belle deTNF-a, respectivement. Une analyse par Western-Blota montré que la présence de la synthétase inductible dumonoxyde d'azote (iNOS) était réprimée par PTX.Une analyse par Northern blot a montre une réductionsignificative de l'ARNm du TNF-a. Nous concluonsque PTX est un inhibiteur efficace de la production deTNF-a induite par Ic lipoarabinomannane (LAM) à IaPois au Ili Yea' du produit que cclui de la transcriptionsur noun lignée cellulare macrophagique. P"I'X exerceégalement une activité inhibitrice modérée sur la pro-duction dc NO, ainsi que sur la traduction de iNOS.
Acknowledgment. This work was supported by theOffice of Mental Retardation and Developmental Dis-abilities of New York State and Staten Island Univer-sity Hospital.
REFERENCESI . CnoMCZYNSKt, P. A reagent for the single step si-
multaneous isolation of RNA, DNA and proteinsfrom cell and tissue samples. Biotechniques 15(1993) 532-535.
2. DING, A., NATHAN, C. F. and STUEHR, D. J. Re-lease of reactive nitrogen intermediates and reac-tive oxygen intermediates from mouse peritonealmacrophages. J. Immunol. 141 (1988) 2407-2412.
3. DIRosA, M., RADOMSKI, M., CARNUCCto, R. andMON('ADA, S. Glucocorticoids inhibit the inductionof nitric oxide synthase in macrophages. 13iochem.Biophys. Res. Comm. 172 (1990) 1246-1252.
4. DotttiRTY, G., JENSEN, C., ALEXANDER, R.. BLI-RESH, C. and NoRToN, J. A. Pentoxifylline sup-

232 International Journal of Leprosy 2001
pression of tumor necrosis factor gene transcrip-tion. Surgery 110 (I 991 ) 192-198.
5. DUBOST, J. J., SouIumR, M.. RISTORI, J. M., BEAU-JON, G., OUALID, T., BuSsu RE, J. L. and SAUVEZIE,
B. An open study of the anti-TNF alpha agentpentoxifylline in the treatment of rheumatoidarthritis. Rev. Rhum. Engl. Ed. 64 (1997) 789-793.
6. GRAD, G. E., FAJAtt o, L. F., PiGUET, P. F., ALLET,B., LAMBERT. P. H. and VASSALLI, P. Tumornecrosis factor (cachectin) as an essent i a I mediatorin iflUI - ine cerebral nlalaria. Science 237 (1989)1210-1212.
7. HAN, J., THOMPSON, P. and BEUTLER, B. 1)exa-methasone and pentoxifylline inhibit endotoxin-induced cachectin/tumor necrosis factor synthesisat separate points on the signaling pathway. J.Exp. Med. 172 (1996) 391-394.
8. HOWE, L. M. Treatment of endotoxic shock: glu-cocorticoids, lazaroids, nonsteroidals, others. Vet.Clin. North. Am. Small Anim. Pract. 28 (1998)249-267.
9. KUNZ, D., WALKER, G.. EBERHARDT, W. andPFEiLSCHIFTER, J. Molecular mechanisms of dexa-methasone inhibition of nitric oxide synthase ex-pressed in interleukin 1 heta-stimulated mesangialcells: evidence for the involvement of transcrip-tional and post-transcriptional regulation. Proc.Natl. Acad. Sci. U.S.A. 93 (1996) 255-259.
10. LAUTERBACK, R., GRABOWSKA, A. and MARCIN-
KiEWICZ, J. Effect of pentoxifylline on nitric oxidereleased by murine macrophages. Biol. Neonate67 (1995) 72-76.
11. LEVIS, W. R. Treatment of leprosy in the UnitedStates. Bull. NY Acad. Med. 60 (1984) 696-711.
12. LoFTis, L. L., MEALS, E. A. and ENGLISH, B. K. Dif-ferential effects of pentoxifylline and interleukin-10on production of tumor necrosis factor and induciblenitric oxide synthase by murine macrophages. J. in-fect. Dis. 175(4) (1997) 1008-1011.
13. MODLIN, R. L. Th l -Th2 paradigm: insights fromleprosy. J. Invest. Dermatol. 102 (1994) 828-832.
14. NATHAN, C. Nitric oxide as a secretory product ofmammalian cells. FASEB J. 6 (1992) 3051-3064.
15. NERY, J. A., PERISSE, A. R., SALES, A. M., VIEIRA,
L. M., SouZA, R. V., SAMPAIO, E. P. and SARNO, E.N. The use of pentoxifylline in the treatment oftype 2 reactional episodes in leprosy. Indian .1.Lepr. 72 (2000) 457-467
16. NEUNER, P., KLOSNER, G., SCHAUER, E., POURMO-
JIB, W., MACHEINER, W., GRUNWALD, C., KNOBLER.R., SCHWARZ, A., LUGER, T. and SCHWARZ, T.Pentoxifylline in vivo downregulates the release ofIL-1 p. IL-6, IL-8 and tumor necrosis factor-a byhuman peripheral blood mononuclear cells. Im-munology 83 (1994) 262-267.
17. PARK, E., QUINN, M. R., WRIGHT, C. E. andSCHULLER-LEViS G. Taurine chloramine inhibitssynthesis of nitric oxide and the release of tumornecrosis factor in activated RAW 264.7 cells. J.Leukoc. Biol. 54 ( 1993) 1 19-124.
I8. PARK, E., SCHULLER-LL.Vis, G. and QuINN, M.R. Taurine chloraminc inhibits production of ni-tric oxide and TNT-a in activated RAW 264.7cells by mechanisms that involve transcriptionaland translational events. .1.Immunol. 154 (1995)4778-4784.
19. PETERSON, T. C. Therapeutic potential for cy-tokine agonists: thalidomide and pentoxifylline inHansen's Disease. Can. J. Infect. Dis. 6 (1995)30-33.
20. QURESHI, A. A., LERNER. L. H. and LERNER, E. A.Nitric oxide and the cutis. Arch. Dermatol. 132(1996) 889-893.
21. REEVES, G. E., FERGUSON, J. K., DOBSON, P. andBOYLE, M. J. Pentoxifylline to treat Mycobacte-rium avium complex exacerbation in late-stage HIVinfection I letter'. Med. .1. Aust. 166 (1997) 446.
22. RIDLEY, R S. and JOPLIN, W. H. Classification ofleprosy according to immunity. A five-group sys-tem. Int. J. Lepr. 34 (1996) 255-273.
23. ROACH, T. I., BARTON, C. H., CHATTERJEE, D., andBLACKWELL, J. M. Macrophage activation:Iipoarabinomannan from avirulent and virulentstrains of Mycobacterium trrhercirlosi.s . differen-tially induces the early genes cfos, KC, JE, and tu-mor necrosis factor-a. .1. I mni unol . 150 (1993)1886-1896.
24. SAMLASKA, C. R and WiNFIELD, E. A. Pentoxi-fylline. J. Am. Acad. Dermatol. 30 (1994) 603-621.
25. SAMPAIO, E. P., KAPLAN, G., MIRANDA, A., NERY, J.A., MIGUEL, C. P., ViANA, S.M. and SARNO, E. N.The influence of thalidomide on the clinical and im-In unological manifestations of erythema nodosumleprosum. J. Infect. Dis. 168 (1993) 408-414.
26. SAMPAIO, E. P.. MORAES, M. O.. NERY, J. A., SAN-
TOS, A. R., MATOS, H. C. and SARNO, E. N. Pent-
oxifylline decreases in vivo and in vitro tumournecrosis factor-alpha (TNF-alpha) production inlepromatous leprosy patients with erythema no-dosum leprosum (ENL). Clin Exp. Imniunol. 111(1998) 300-308.
27. SARNO, E. N., NERY, J. A., GARCIA, C. C. and SAM-
PAiO, E. P. Is pentoxifylline a viable alternative inthe treatment of ENL? Int. J. Lepr. 63 (1995)570-571.
28. SCHULLER-LEVIS, G. B., LEVIS, W. R., AMMAllA-
LORSO, M., NOSRATI. A. and PARK, E. Mycohacte-rial lipoarabinomannan induces nitric oxide andtumor necrosis factor alpha production in a macro-phage cell line: down-regulation by taurine chlo-rumine. Infect. Immun. 62 (1994) 4671-4674.
29. SCHWARZ, A., KRONE, C., TRAUTINGER, F., AR-
GANE, Y., NEUNER, P.. LUGER, T. and SCHWARZ, T.Pentoxifylline suppresses Irritant and contact hy-persensitivity reactions. J. Invest. Dermatol. 101(1993) 549-552.
30. SCHWARZ, T., SCHWARZ, A., KRONE, C. H. andLUGER, T. A. Pentoxifylline suppresses allergicpatch test reaction in humans. Arch. Dermatol.129 (1993) 513-514.

69, 3 Park, et al.: Pentoai/i'Iline and Macrophages 233
31. Scttw, t z, T., URBANSKI, A., TAUTINGER, F., er,v..Pentoxifylline blocks the elicitation phase of con-tact hypersensitivity and irritant dermatitis. J. In-vest. Derntatol. 98 (1992) 593.
32. SIELING, P. A. and MoDt,tN, R. L. Cytokine pat-terns at the site of mycobacterial infection. Im-munobiology 191 (1994) 378-387.
33. SrRIETEts, R. M., REMICK, D. G., WARD, P. A.,SPENGLER, R. N., LYNCH, J. P., LAIRICK, J. andKUNKEL, S. L. Cellular and molecular regulationof tumor necrosis factor-alpha production by pent-oxifylline. Biochem. Biophys. Res. Comm. 155(1998) 1230-1236.
34. TACHIBANA, K., CHEN, G. J., I IUANG, D. S., Scuotasi,P. and WATSON, R. R. Production of tumor necro-sis factor-a by resident and activated marinemacrophages. J. Leukoc. Biol. 51 (1992) 251-255.
35. TALHARI, S., 0H51, A. T., TALI-IAtn, A. C., SOUZA, F.H. and Fi:nztaRA, L. C. Pentoxifylline may be use-ful in the treatment of type 2 leprosy reaction.Lepr. Rev. 66 (1995) 261-263.
36. TRAJKOVIC, V., BADOVINAC, V., POPADIC, D.,HADzte, O. and SroJKOVtc, M. M. Cell-specific
effects of pentoxifylline on nitric oxide productionand inducible nitric oxide synthase mRNA expres-sion. Immunology 92 (1997) 402-406.
37. VAN FURTH, A. M., ROORD, J. J. and VAN FURTH, R.Roles of proinflammatory and anti-inflammatorycytokines in pathophysiology of bacterial menin-gitis and effect of adjuvant therapy. Infect. lm-man. 64 (1996) 4883-4890.
38. WARD, A. and CLlSSor.O, S. P. Pentoxyfylline.Drugs 34 (I 987) 50.