osmotic regulation of transcription: induction of the prou betaine
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Nov. 1986, p. 805-814 Vol. 168, No. 20021-9193/86/110805-10$02.00/0Copyright © 1986, American Society for Microbiology
Osmotic Regulation of Transcription: Induction of the proU BetaineTransport Gene Is Dependent on Accumulation of
Intracellular PotassiumLESLEY SUTHERLAND,1 JOHN CAIRNEY,1 MICHAEL J. ELMORE,2 IAN R. BOOTH,2
AND CHRISTOPHER F. HIGGINS'*Molecular Genetics Laboratory, Department of Biochemistry, University of Dundee, Dundee DDJ 4HN,2 and Department
of Microbiology, Marischal College, University ofAberdeen, Aberdeen AB9 lAS,1 Scotland
Received 17 March 1986/Accepted 4 August 1986
The proU locus, which encodes a high-affinity betaine transport system, and the kdp operon, which encodesa potassium transport system, are the principal osmoresponsive genes in Escherichia coli and Salmonellatyphimurium. The kdp operon is known to be induced in response to changes in cell turgor. We haveinvestigated the control of proU expression and shown that it differs from that of kdp in a number offundamental ways. Rather than responding to changes in turgor, proU expression is principally determined bythe intracellular accumulation of potassium ions. Potassium and betaine were shown to play distinctosmoprotective roles. Potassium serves as the principal osmoprotectant and is accumulated in response tolow-level osmotic stress to restore turgor. As external osmolarity is increased to a level at which thecorresponding increase in internal potassium concentrations is potentially deleterious to enzyme function,betaine (when available) is accumulated in preference to potassium. The different mechanisms ofproU and kdpregulation reflect the different physiological roles of these two osmoprotectants.
Bacterial cells are frequently subjected to fluctuations inthe osmolarity of the medium in which they are growing.Most organisms respond to osmotic stress in a similarmanner, by accumulating high intracellular concentrations ofcompatible solutes (30, 31). These solutes not only balanceexternal osmolytes such that cell turgor is maintained butmust also be compatible with the function of intracellularenzymes. In Escherichia coli and Salmonella typhimuriumthe accumulation of K+ ions plays a primary role in main-taining the osmotic balance of the cell (6; W. Epstein, FEMSMicrobiol. Lett., in press). An ionic balance is normallymaintained by the intracellular synthesis of equivalentamounts of glutamate (41). Potassium uptake is mediated bytwo independent transport systems, a low-affinity system,Trk, and a high-affinity system, Kdp (36). The Kdp systemcontributes to the accumulation of K+ whenever the Trksystem is not sufficiently active to maintain turgor; theexpression of the kdp operon is specifically induced undersuch conditions (26).
It is well established that in the absence of exogenoussolutes such as proline or betaine the intracellular concen-tration of K+ ions is essentially proportional to the osmolar-ity of the external medium (16). However, in complex mediathe situation is less simple. Certain organic compounds(compatible solutes) can serve as osmoprotectants and, ifaccumulated by the cell, will significantly enhance growthrates at high osmolarity. The most universally adoptedcompatible solutes are proline and betaine (N,N,N-trimethylglycine). Betaine is accumulated during osmoticstress by such diverse organisms as halotolerant plant spe-cies (44), marine animals (4), halophilic bacteria (22), andcyanobacteria (35). Both proline and betaine can serve asosmoprotectants for the gram-negative bacteria E. coli andS. typhimurium. Thus, increased synthesis of proline (13) orbetaine (27, 40) or the uptake of these compounds when
* Corresponding author.
supplied exogenously (6, 7, 11, 14, 29, 34) can markedlyenhance growth rates at high osmolarity. There are twodistinct transport systems for betaine in S. typhimurium (6,7). The proP gene encodes a low-affinity betaine uptakesystem which also transports proline (6), whereas the proUgene, originally defined as a proline transport system (14,15), encodes a high-affinity transport system with betaine asits principal substrate (7). ProU is a periplasmic bindingprotein-dependent transport system (unpublished results),and expression of the proU gene is induced under conditionsof osmotic stress (7, 15, 17). The proP and proU genes havealso been identified in E. coli and probably serve analogousroles (17, 18).Osmotic stress affects the expression of only a very
limited number of bacterial genes (7, 12). The relativeexpression of the porins OmpC and OmpF is affected by theosmolarity of the medium (19, 20). However, many otherfactors also influence porin expression (42), and the preciserole of osmolarity is obscure and may be indirect. Expres-sion ofphoA and malB is also affected by medium osmolarity(43), but again these affects are not major and may well be anindirect effect on the cellular concentration of their relevantinducers (5). In E. coli the enzymes involved in the conver-sion of choline to betaine are only synthesized in osmoticallystressed cells (40). The genes encoding these enzymes arelocated near the lac operon and are absent in S. typhimuriumand many E. coli strains (40). It seems likely that theirregulation is at the level of transcription, but this has not yetbeen demonstrated. Unlike any of the above-mentioned,osmotically influenced genes, kdp and proU are fully re-pressed at low osmolarity, and expression is induced over100-fold by osmotic stress. Thus, kdp and proU are the onlytwo known genes which are directly induced at the transcrip-tional level in response to osmotic stress.The kdp and proU genes encode high-affinity potassium
and betaine transport systems, respectively. The relativeroles of K+ and betaine in restoring turgor and facilitating
805
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 26
07:9
880:
1807
:feb
9:b1
9f:f
5c6:
58a:
a718
.

806 SUTHERLAND ET AL.
TABLE 1. Bacterial strains
Strain Genotype Source/Construction
S. typhimuriumCH272 oppA305::Mu dl galE503 Abio-561 (23)CH322 putP202: Mu dl gal-503 Abio-561 (8)CH776 AoppBC250 tppB84: Mu dl-8 (24)CH946 proUl 702: Mu dl-8 (7)CH1301 proUl 705: :Mu dJ(Kanr lac) This paperLT2 Wild type B. N. AmesTT10288 hisD9953:: Mu dJa his-9944:: Mu dl J. Roth
E. coliCH1366 MC4100 proUl706: :Mu d11681 (Kanr) This paperCH1407 MC4100 proUl706: :Mu dI1681 (Kan9 kdp: :TnJO This paperEMG2 Wild type CGSCbMC4100 araD139 rpsL thi A(argF-lac)U169 (9)MJF1 relA araFJ39 araE thi gltSo A(his-gnd) A(argF-lac)U169 rpsL flaB deo (38)MJF261 MJF1 kdpA5:: Mu dIX This paperMJF266 MJF1 kdp: :TnlO W. EpsteinPB13 cysE recA srl: :TnlO M. Jones-MortimerP011681 Mu d11681 (Kan9 ara: :(Mu cts)3 A(proAB-argF-lac)XIII rpsL (10)
a Originally called Mu d11734 (10).b E. coli Genetic Stock Center, Yale University, New Haven, Conn.
growth at high osmolarity are presently obscure. In thispaper we show that these two osmoprotectants play distinctphysiological roles. It has been demonstrated that the kdpgene is primarily regulated in response to changes in turgorpressure (26). Initial studies showed that, like kdp expres-sion, proU expression is also induced by osmotic stress (7,15, 17). We show here that there are a number of fundamen-tal differences in the responses of kdp and proU to osmoticstress. These differences are reconciled by the demonstra-tion that induction of proU expression is not directly con-trolled by changes in cell turgor, but is apparently dependentupon the intracellular concentration of potassium ions.Thus, when a cell is osmotically stressed the consequentchanges in cell turgor activate K+ transport, leading to anincrease in intracellular K+ concentrations which inducesproU expression. The different mechanisms of induction ofproUand kdp are shown to reflect distinct physiological rolesfor betaine and potassium as osmoprotectants.
MATERIALS AND METHODSBacteeial strains and growth media. All strains are deriva-
tives of S. typhimurium LT2 or of E. coli K-12. Theirgenotypes and sources are detailed in Table 1. Transductionsin S. typhimurium were carried out with a high-transducingderivative of phage P22 int4 as described by Roth (37). E.coli transductions were with phage P1 vir (32). Aftertransduction of any transposon or Mu derivative, the correctlocation of the element and the presence of only a singlecopy of that element in the transductant were checked bymarker rescue. Cells were grown aerobically at 37°C unlessotherwise stated. For strain constructions cells were grownin LB medium (37). For all other experiments the growthmedium used was LOM (low-osmolarity medium [6]), whichcontains 0.4 mM MgSO4, 6 ,uM (NH4)2SO4. FeSO4, 20 mM(NH4)2HP04, 10 mM bistrispropane, 5 mM NaCl, 10 mMglucose, and 1 mM KCl. For certain experiments the potas-sium concentration was varied by adding extra KCl asappropriate. For potassium limitation KCl was omitted;flame photometry showed the potassium concentration inthis "potassium-free" medium to be <8 1xM.
Isolation of a stable proU-lacZ fusion in S. typhimurium.Although we have previously constructed pro U-lacZ operon
fusions with the Mu derivatives Mu dl and Mu dl-8 (7), amini-Mu derivative (Mu dJ, originally called Mu dI1734;[10]) has recently been described which lacks all transposi-tion functions and is therefore completely stable. A MudJ-mediated proU-lacZ fusion was isolated by transducingLT2 to Kanr with a P22 lysate of TT10288. TT10288 containsa his::Mu dJ insertion and, in addition, a very closely linkedMu dl helper phage that provides the genes encoding trans-position functions and that can be copackaged with the MudJ in P22 transducing particles. About 104 Kanr colonies,each of which contained an independent Mu dJ insertion intothe chromosome, were pooled, washed twice in minimalmedium, and plated on LOM-agar plates containing 0.3 MNaCl and 40 ,ug of 5-bromo-4-chloro-3-indolyl-3-D-galactoside per ml. Deep blue (Lac') colonies were pickedand screened on LOM-agar plates containing 5-bromo-4-chloro-3-indolyl-,-D-galactoside but lacking the addedNaCl. Any colonies which expressed 3-galactosidase on theplates containing 0.3 M NaCl but remained white (Lac-) onlow-salt plates were purified and characterized further. Onthe basis of previous results (7) we anticipated that most ofthese isolates would contain proU-lacZ fusions. One suchderivative, CH1301, was shown to contain a single Mu dJinsertion, and the insertion was shown to be in proU bytransductional mapping and marker rescue as describedpreviously (7). Expression of ,B-galactosidase from this fu-sion was regulated in a manner identical to that reportedpreviously for Mu dl-8-mediated proU-lacZ fusions (7).proU-lacZ fusions in E. coli. A mini-Mu-mediated proU-
lacZ operon fusion was selected from a collection of randomMu dI1681 insertions. These random mini-Mu insertionswere constructed by infection of MC4100 (9) with a temper-ature-induced lysate of strain POI1681, which contains boththe mini-Mu and a Mu helper phage (10). pro'U-lacZ fusionswere selected from this pool as described above for the S.typhimurium fusions, except that MacConkey medium-lactose was used as an indicator rather than 5-bromo-4-chloro-3-indolyl-p-D-galactoside. As for S. typhimurium, allosmotically induced lacZ fusions selected in this mannermapped to a single locus which was shown to be proU bycotransduction with a srl::TnJO insertion. kdp-lacZ fusionswere not selected by this procedure since MacConkey
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 26
07:9
880:
1807
:feb
9:b1
9f:f
5c6:
58a:
a718
.

OSMOTIC REGULATION OF GENE EXPRESSION 807
medium-lactose plates contain sufficient K+ to fully represskdp expression. As discussed below, proU-lacZ fusionsisolated in E. coli were found to be regulated in a matineridentical to that of the more completely characterized pro U-lacZ fusions in S. typhimurium.
Stabilized kdp-lacZ fusion. The E. coli kdpAS::Mu dl(Aprlac) fusion (26) was stabilized by selecting temperature-resistant colonies at 42°C. These temperature-resistant de-rivatives were screened for the absence of Mu dl transposi-tion when transduced into a Mu-free strain. One suchstabilized kdp-lacZ fusion was transduced into MJF1 (38);the resultant strain, MJF261, was shown to contain a singleMu dl insertion, and the insertion was shown to be in kdp,by marker rescue and genetic mapping. This stabilizedfusion was used for all experiments with kdp and is referredto as kdpAS::Mu dlX.Growth of cells under potassium limitation. Cells grown
overnight in LOM were washed twice in LOM lacking KCland resuspended in the same K+-free medium at a 1:50dilution. Cells stopped growing at an optical density at 600nm of about 0.3 (see Fig. 3). Since growth could be rapidlyrestored to normal rates by the addition of 1 mM KCl, we
concluded that growth was indeed limited by potassium.Osmotic stress. The two osmolytes used routinely were 0.3
M NaCl and 0.44 M sucrose. These are isoosmotic concen-trations of the two solutes. Cells were osmotically stressedin one of two ways. Steady-state adaptation to osmoticstress was achieved by growing cells overnight in the pres-
ence of the appropriate osmolyte, subculturing the cell intothe same medium at a 1:50 dilution, And growing to themidexponential phase before assaying. Alternatively, a rapidosmotic shock was achieved by growing cells to midexpo-nential growth in LOM medium before adding sucrose to0.44 M or NaCl to 0.3 M.
Pulse-labeling. Samples (1.0 ml) of cells grown as indicatedbelow were pulsed for 2 min with [35S]methionine (800 Cimmol-1; 5 ,uCi) followed by a 2-min chase with unlabeledmethionine (100 ,ug ml-'). The cells were then rapidlysedimented by centrifugation, resuspended in 50 ,u1 ofLaemnli sample buffer (25), boiled for 2 min, and separatedon a 9% sodium dodecyl sulphate-polyacrylamide gel (2, 25).Labeled proteins were visualized by autoradiography afterthe gel was dried.
,-Galactosidase assays. ,-Galactosidase was assayed bythe procedure of Miller (32). Cells were permeabilized by thesodium dodecyl sulfate-chloroform procedure. All assayswere performed in triplicate and repeated on at least threeindependent occasions.Measurement of intracellular KV. Cells were grown to the
midexponential phase in LOM containing NaCl, KCl, andbetaine as appropriate. Cells from 1 ml of culture wererapidly harvested by centrifugation for 30 s in an Eppendorf
TAB3LE 2. Differences in expression of proU and kdpa
Medium additives 1-Galactosidase activity (U)
Osmolyte K+ S. typhimurium E. coli E. coliproU-lac proU-lac kdp-lac
<8 ,uM 5 23 87850 mM 9 55 11
0.3 M NaCI 1 mM 364 1,293 1,3590.3 M NaCl 50 mM 468 1,201 2
a Cells of S. typhimurium CH1301 (proU-lacZ), E. coli MJF261 (kdp-lacZ),and E. coli CH1366 (proU-lacZ) were grown to the midexponential phase inLOM, with osmolytes and K+ added as indicated, and assayed for ,-galactosidase activity.
TABLE 3. Effects of a kdp mutation on proU expressiona
Medium additives 1-Galactosidase activity (U)
Osmolyte K+ proU-lacZ proU-lacZkdp: :TnlO
100 ,uM 23 24350 F.M 24 22
1 mM 23 2410 mM 23 2350 mM 55 26
0.3 M NaCI 100 ,uM 1,570 640.3 M NaCI 350 ,uM 1,606 4750.3 M NaCl 1 mM 1,293 8300.3 M NaCl 10 mM 1,060 8660.3 M NaCI 50 mM 1,201 923
a Cells of E. coli strains CH1366 (proU-lacZ) and CH1407 (proU-lacZkdp:: TnlO) were grown to the midexponential phase in LOM, with osmolytesand K+ added as indicated, and 3-galactosidase activity was assayed.
centrifuge, all of the supernatant was removed, and thepellet was suspended in 1 ml of deionized water. Thesuspension was boiled for I min to release k' from the cell,and the K+ concentration was determined by flame photom-etry. Each data point is an average of at least three separatedeterminations that did not vary by more than -+-10%.Results obtained with this inexpensive and rapid procedurewere very similar to those obtained with 42K (see below).Cell volumes were determined by the distribution of 3H20and [14C]sucrose by the method of Stock et al. (39) asdescribed previously (1). Values obtained for cell volumeswere consistent with previously reported results (34, 39).Increasing the osmolarity of LOM by adding 0.3 M NaClcaused a decrease in cell volume of approximately 35%(from 3.0 to 1.95 ,ul mg-').Measurement of relative intracellular K+ and betaine con-
centrations. Cells were grown to saturation in LOM contain-ing 0.3 M NaCl, harvested by centrifugation, and suspendedat 10 mg of protein per ml in LOM containing chloramphen-icol (50 ,ug ml-') and NaCl as appropriate. 42KHCO3 (150mCi mmol-1) and [14C]betaine (10 mCi mmol-1) were addedto a final concentration of 1 mM, and the cells were incu-bated at 20°C for 90 min. Preliminary experiments showedthis to be sufficient time for equilibration of the internal andexternal pools. Cells were then collected by filtrationthrough a 0.45-p.m membrane filter (Millipore Corp.) andwashed in the same medium in which they had been incu-bated but lacking the radioisotopes, and the 42K' and[14C]betaine accumulated by the cells were determined byscintillation counting. Due to the very short half-life of 42Kand the overlap of the scintillation windows, the 14C wascounted after allowing sufficient time for 42K decay.
RESULTS
Because direct assays for proU and kdp function aredifficult to perform, expression of these genes was assessedby assaying 0-galactosidase expression from isolates withkdp-lacZ or proU-lacZ operon fusions. Assays were per-formed during steady-state growth in the appropriate me-dium, rather than after sudden osmotic shock, unless other-wise indicated.The proU gene and the ProU transport system are consid-
erably better characterized in S. typhimurium than they arein E. coli. However, strains with kdp-lacZ fusions havenever been isolated in S. typhimurium; despite severalattempts with various selections, we have so far been unable
VOL. 168, 1986
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 26
07:9
880:
1807
:feb
9:b1
9f:f
5c6:
58a:
a718
.

808 SUTHERLAND ET AL.
TABLE 4. Intracellular potassium concentrationsa
Medium additives IntracellularStrain Relevant K + concn
genotype Osmolyte KCI Proline-betaine (Kimol ml-,)
S. typhimurium CH1301 proU-lacZ 1 mM 86S. typhimurium CH1301 proU-lacZ 50 mM 93S. typhimurium CH1301 proU-lacZ 0.3 M NaCl 1 mM 386S. typhimurium CH1301 proU-lacZ 0.3 M NaCl 10 mM 395S. typhimurium CH1301 proU-lacZ 0.3 M NaCl 50 mM 410S. typhimurium CH1301 proU-lacZ 0.44 M sucrose 50 mM 398S. typhimurium CH1301 proU-lacZ 0.05 M NaCl 1 mM 139S. typhimurium CH1301 proU-lacZ 0.1 M NaCl 1 mM 206S. typhimurium CH1301 proU-lacZ 0.2 M NaCl 1 mM 278S. typhimurium CH1301 proU-lacZ 0.3 M NaCl 1 mM 1 mM proline 304S. typhimurium CH1301 proU-lacZ 0.3 M NaCl 1 mM 1 mM betaine 288
E. coli CH1366 proU-lacZ 1 mM 98E. coli CH1366 proU-lacZ 50 mM 109E. coli CH1366 proU-lacZ 0.3 M NaCl 100 ,uM 380E. coli CH1366 proU-lacZ 0.3 M NaCl 350 ,uM 423E. coli CH1366 proU-lacZ 0.3 M NaCl 1 mM 400E. coli CH1366 proU-lacZ 0.3 M NaCl 10 mM 380E. coli CH1366 proU-lacZ 0.3 M NaCI 50 mM 426
E. coli CH1407 proU-lacZ 0.3 M NaCl 100 ,uM 159kdp: :TnlO
E. coli CH1407 proU-lacZ 0.3 M NaCl 350 ,uM 271kdp: :TnlO
E. coli CH1407 proU-lacZ 0.3 M NaCl 1 mM 332kdp: :TnJO
E. coli CH1407 proU-lacZ 0.3 M NaCl 10 mM 395kdp: :TnlO
E. coli CH1407 proU-lacZ 0.3 M NaCl 50 mM 416kdp: :TnlO
a Cells were grown in the appropriate medium to midexponential growth, and the intracellular K+ concentrations were determined as described in Materialsand Methods.
to isolate strains with such fusions in this species. Thissuggests there may be some differences in the Kdp systembetween these two species. However, any difference is likelyto be minor, for example, in the affinity of the transportsystem for its substrate, rather than a major difference in theactual mechanism of kdp regulation. It is known that S.typhimurium possesses a Kdp transport system, the geneshave been cloned by virtue of their sequence homology tothe E. coli kdp genes (W. Epstein, personal communication),and, like in E. coli, the Kdp proteins of S. typhimurium areinduced by potassium starvation (3; unpublished results).We have therefore studied the regulation of kdp expressionin E. coli MJF261 and the regulation of proU expressionprimarily in S. typhimurium CH1301. However, to excludethe possibility that the differences in regulation we observebetween proU and kdp (see below) are due to a speciesdifference, and to show that these differences are not a
consequence of any species difference in Kdp, we alsoisolated aproU-lacZ fusion in E. coli (CH1366) and repeatedsome experiments with this species. As described below, no
significant difference in the regulation of proU expressionwas seen between E. coli and S. typhimurium.
Potassium limitation induces kdp but not proU. In minimal-salts medium, K+ accumulation provides the principalmeans of maintaining cell turgor (16). When cells are grown
under potassium limitation, potassium cannot be accumu-lated to sufficient intracellular concentrations to maintaincell turgor, and any turgor-sensitive gene would be expectedto be induced. Strains containing either a proU-lacZ or a
kdp-lacZ fusion were grown under potassium limitation (<8
,uM), and P-galactosidase activity was assayed (Table 2).Our results with kdp-lacZ fusions were in full agreementwith those of Laimins et al. (26); expression of kdp at lowosmolarity was strongly induced by potassium limitation (<8,uM K+). However, under identical growth conditions noinduction of proU expression was observed in either E. colior S. typhimurium. This indicates an important differencefrom the regulation of kdp and, in addition, implies that theexpression ofproU cannot be simply in response to changesin cell turgor.
Excess potassium represses kdp but not proU. When suffi-cient potassium is present in the growth medium it isaccumulated by the cell to restore turgor. It is well estab-lished that intracellular potassium concentrations increase inmore or less direct proportion to external osmolarity, both to
TABLE 5. K+ requirement for proU inductiona
P-GalactosidaseStrain Fusion activity (U)
+ K+ -K+
CH946 proU-lacZ 436 181CH272 oppA-lacZ 766 716CH776 tppB-lacZ 248 281CH322 putP-lacZ 70 65
a Cells were grown under potassium limitation (50 ,uM) or excess (1 mM) inmedium containing 0.3 M NaCl, and ,B-galactosidase activity was assayedduring midexponential growth. Leucine (1 mM) was included in the growthmedium for CH776 to induce expression of tppB (23). oppA (23) and putP (8)are both constitutively expressed under these growth conditions.
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 26
07:9
880:
1807
:feb
9:b1
9f:f
5c6:
58a:
a718
.

OSMOTIC REGULATION OF GENE EXPRESSION 809
TABLE 6. K+ requirement for proU inductiona
P-Galactosidase activityStrain Fusion Inducer (U) Induction point
+ K+ -K+
CH1301 proU-lacZ 0.44 M Sucrose 190 39 1CH1301 proU-lacZ 0.44 M Sucrose 204 27 2CH1301 proU-lacZ 0.44 M Sucrose 143 34 3CH776 tppB-lacZ 1 mM Leucine 276 218 3EMG2 lacZ 4 mg of IPTGb per ml and 5 mM cAMP 1,416 1,434 3
a Cells were grown to potassium starvation, and the inducer was added, with or without 1 mM K+, at the points on the growth curve indicated in Fig. 3.13-Galactosidase activity was assayed 30 min after the inducer was added. For the lacZ experiment with EMG2, glycerol rather than glucose was used as carbonsource.
b IPTG, Isopropyl-,-D-thiogalactopyranoside.
maintain an osmotic balance and to restore turgor (16; seebelow). Thus, during steady-state growth at high osmolarityin the presence of excess potassium (50 mM), sufficient K+can be accumulated to partially or completely restore turgor,and expression of turgor-sensitive genes would be expectedto be repressed. Consistent with this, and with the view thatkdp is turgor regulated, we found that kdp was not expressedduring steady-state growth in medium of high osmolarity aslong as sufficient K+ (50 mM) was present (Table 2). How-ever, at lower K+ concentrations (1 mM), turgor could notbe maintained in these Kdp- strains, and kdp expression wasinduced. In marked contrast to kdp, expression ofproU wasmaintained at a fully induced level during steady-stategrowth at high osmolarity, even in the presence of 50 mMK+ (Table 2). Again, there was no obvious difference in theregulation of proU-lacZ fusions in E. coli and S. typhi-murium. These results illustrate another clear differencebetween kdp and proU and again imply that induction ofproU expression cannot be a direct response to changes incell turgor.
Effect of kdp mutations on proU expression. Because theproU-lacZ strains are Kdp+, whereas the kdp-lacZ strain weused is phenotypically Kdp-, it is important to ascertain thatthe differences in regulation between proU and kdp are notsimply a result of the presence or absence of the Kdptransport system. Thus, a kdp::TnlO insertion was intro-
03
3
0~2. 2
'p
0iO1
1 2 3 4 5 6 7Time (hours)
FIG. 1. Growth of cells under potassium limitation. An overnightculture of S. typhimurium CH1301 (proU-lacZ) grown in LOMcontaining 1 mM KCl was washed twice in LOM lacking K+ (<8 ,uMK+) and resuspended at a 1:50 dilution in the same medium. Growthwas followed by monitoring the optical density of the culture at 600nm (O.D.6m). After about 5 h, growth ceased due to K+ limitation;growth could be rapidly restored by adding 1 mM KCl. The arrowsindicate the points at which samples were taken for osmotic shock(see text and Table 6).
duced into an E. coli proU-lacZ fusion strain, and theexpression of proU was compared with that of its Kdp+parent (Table 3). The differences in the regulation of proUand kdp cannot be attributed to the presence or absence of afunctional Kdp system. As expected (16, 26), growth of theKdp- derivative was limited by potassium even at extracel-lular concentrations as high as 350 ,uM (data not shown).This is because the remaining K+ transport system, Trk, hasa low affinity for K+ (K, = 1 mM) and is unable toaccumulate K+ at sufficient rates to maintain optimal intra-cellular concentrations (see below). In contrast, the Kdp+derivative grew normally at both 100 and 350 ,uM K+. proUexpression was not induced by K+ limitation, even whensuch limitation was made more severe by the introduction ofa kdp mutation (Table 3). Similarly, expression ofproU wasnot repressed by excess K+ (50 mM) in either the Kdp- orthe Kdp+ derivative. The only effect of introducing a kdplesion was that proU expression at high osmolarity wasreduced when extracellular K+ concentrations were lowenough (100 or 350 j,M) to be limiting for K+ accumulation.This is in contrast to the Kdp+ derivative, where K+ doesnot become limiting at these extracellular concentrations(see below) and where proU expression remains relativelyconstant even at low extracellular K+ concentrations. Thus,when the cell cannot accumulate sufficient K+ to restoreturgor and maintain optimum growth rates, osmotic stress isincreased, yet proU expression is actually reduced. Signifi-cantly, these results are completely opposite to those foundfor kdp, which is induced rather than repressed as K+becomes limiting (Table 2), and confirm that proU and kdpare induced by very different signals.These observations suggest that intracellular potassium
concentrations may be important in regulating proU expres-sion. proU is induced under conditions where intracellularpotassium concentrations would be expected to be high andis repressed when intracellular potassium concentrationswould be expected to be low. To confirm these interpreta-tions, we measured intracellular potassium concentrationsunder the various growth conditions (Table 4). In Kdp+strains, intracellular K+ concentrations were found not tovary significantly with various extracellular concentrationsbetween 100 puM and 50 mM but to increase with increasingmedium osmolarity. This is in excellent agreement with thedata of Epstein and Schultz (16). In contrast, in a Kdp-strain intracellular potassium levels are reduced at extracel-lular potassium levels below the K, of Trk (100 and 350 ,uM).Again, these data are consistent with published results (26);in addition, the reduced intracellular K+ concentrationscorrelate well with reduced growth rates under the sameconditions (data not shown). In all cases, the measuredintracellular concentrations of K+ ions, altered by varying
VOL. 168, 1986
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 26
07:9
880:
1807
:feb
9:b1
9f:f
5c6:
58a:
a718
.

810 SUTHERLAND ET AL.
1- +
2_+
3_+
FIG. 2. Protein synthesis during potassium starvation. Cells ofS. typhimurium CH1301 were grown in potassium-free medium asdescribed in the legend to Fig. 1. At the appropriate points on thegrowth curve (Fig. 1), samples were osmotically shocked by addingsucrose to 0.44 M; 1 mM K' was added together with the sucrosewhere indicated. At 30 min after the osmotic shock cells werepulse-labeled with [35S]methionine, and the proteins were separatedby electrophoresis on a 9%1 sodium dodecyl sulfate-polyacrylamidegel.
the extracellular K+ concentrations or medium osmolarity orby introducing kdp mutations, are compatible with the viewthat induction ofproU is dependent upon the accumulationof intracellular potassium.
Expression of proU depends upon intracellular potassium.The data above show that kdp and proU expression isregulated differently and demonstrate that proU cannotrespond directly to changes in turgor. Other than turgor, themost direct physiological response to osmotic stress is theaccumulation of K+ ions within the cell. In the absence ofosmoprotectants such as betaine, intracellular K+ concen-trations increase in proportion to extracellular osmolarity(16; this paper). It therefore seemed possible that proUexpression might be determined by the intracellular concen-tration of K+ ions. This hypothesis is strongly supported bythe data presented above, which show that proU expressionis induced whenever intracellular K+ concentrations arehigh and is repressed whenever concentrations are low. (i)Iso-osmotic concentrations of different osmolytes whichincrease intracellular K+ to similar extents (16) have similareffects on the induction of proU. (ii) At high osmolarity theintracellular K+ concentration remains high, even when
sufficient extracellular K+ is supplied to permit restorationof turgor (16). proU expression is fully induced under theseconditions even during steady-state growth. (iii) Potassiumlimitation, which reduces intracellular K+ concentrations,does not induce proU expression even though the cell isunder turgor stress. (iv) In a Kdp- strain, proU expression athigh osmolarity is considerably reduced when the externalK+ concentrations are low. These K+ concentrations arebelow the K, for Trk (36), and the ability of the Kdp- strainto accumulate sufficient intracellular K+ becomes limiting.To test this hypothesis, strain CH946 (proU-lacZ) was
grown under K+ limitation (50 ,uM) in medium containing 0.3M NaCl. After 3 h of growth, expression of proU wasreduced to less than 50% of the level in cells grown with 1mM K+. Expression of several control fusions of lacZ toother genes (opp, tpp, and putP) was essentially identicalwhatever the extracellular K+ concentration (Table 5).These results imply that K+ ions play a specific role in proUexpression.To define more precisely the effects of K+ ions on proU
expression, cells from an overnight culture of strain CH1301were washed in LOM lacking K+ (<8 ,uM K+), and resus-pended at a 1:50 dilution in the same medium. Growth wasfollowed by monitoring the optical density of the culture at600 nm (Fig. 1). At an optical density of about 0.3, growthceased due to potassium starvation; growth could be re-stored to normal by the addition ofpotassium to the medium.At various stages during growth (arrows in Fig. 1), cells weretaken and osmotically shocked by adding sucrose to a finalconcentration of0.44 M. IfproU induction were mediated byan increase in internal K+ concentrations, then an osmoticshock under K+ limitation would not be expected to induceexpression. However, if K+ (1 mM) were added at the sametime as the osmotic shock is given, this will be accumulatedand normal induction would be anticipated; this is preciselywhat was observed (Table 6). Expression of proU was notinduced by osmotic shock unless K+ ions were provided.Essentially identical results were obtained with a proU-lacZfusion in E. coli (data not shown). When this experiment wasrepeated with NaCl as the osmolyte, significant induction ofproU was seen even when the osmotic shock was given in
1400O
1200 F
@i 1000v)0
u 800
da 600t.l))
a
4VVr
200
I ,
2 4 6 8 10 50
[K1 mMFIG. 3. Effect of different osmolytes on kdp expression. Cells of
E. coli MJF261 (kdp-lacZ) were grown overnight in LOM containingthe indicated concentrations of KCI and added osmolytes as appro-priate, and ,B-galactosidase activity was assayed. Symbols: x, noadded osmolyte; E, grown in 0.44 M sucrose; 0, grown in 0.3 MNaCl.
J. BACTERIOL.
F
LD
ownl
oade
d fr
om h
ttps:
//jou
rnal
s.as
m.o
rg/jo
urna
l/jb
on 2
1 D
ecem
ber
2021
by
2607
:988
0:18
07:f
eb9:
b19f
:f5c
6:58
a:a7
18.
OSMOTIC REGULATION OF GENE EXPRESSION 811
the absence of added K+. However, this was found to be an
artifact, since 0.3 M NaCl (Analar grade) contains approxi-mately 50 ,uM K+ as a contaminant, sufficient to permit proUinduction in a Kdp+ strain.Because cell growth was reduced and eventually halted by
potassium limitation during these experiments, it was impor-tant to establish that protein synthesis was identical whetheror not potassium was added at the same time as the cellswere given the osmotic shock. Thus, cells were grown andosmotically shocked under conditions identical to thosedescribed above, but instead of assaying ,B-galactosidase wepulse-labeled cells with [35S]methionine. The labeled pro-
teins were separated by sodium dodecyl sulfate-polyacryl-amide gel electrophoresis (Fig. 2). Quite clearly, proteinsynthesis is unaffected by K+ starvation and is essentiallyidentical whether or not K+ is supplied at the same time as
the cells are shocked. Presumably, the internal K+ concen-
tration is maintained at a sufficient level for normal ribosomefunction but at insufficient levels to restore turgor and permitgrowth.As a further control, we wished to show that lacZ expres-
sion from promoters other than proU could be induced underconditions of potassium starvation. Unlike proU-lacZ fu-sions, isopropyl-p-D-thiogalactopyranoside induction ofchromosomally encoded lacZ and the induction of a tppB-lacZ fusion by leucine were little affected by potassiumstarvation (Table 6). Thus, neither protein synthesis nor thespecific synthesis of P-galactosidase was significantly re-
duced by K+ starvation. It must therefore be concluded thatproU expression requires the accumulation of intracellularK+ ions.
Different osmolytes have different effects on kdp induction.The expression of kdp is induced by sudden osmotic shockwith any osmolyte (with the exception of glycerol, which isfreely permeable through the membrane and hence does notcause a change in turgor) (26). In apparent contradiction, ithas been reported (17) that during steady-state growth at
1-4*
12.
10-
06
0.6
04
0-2
100 200 300 400 500 0
Time (min)
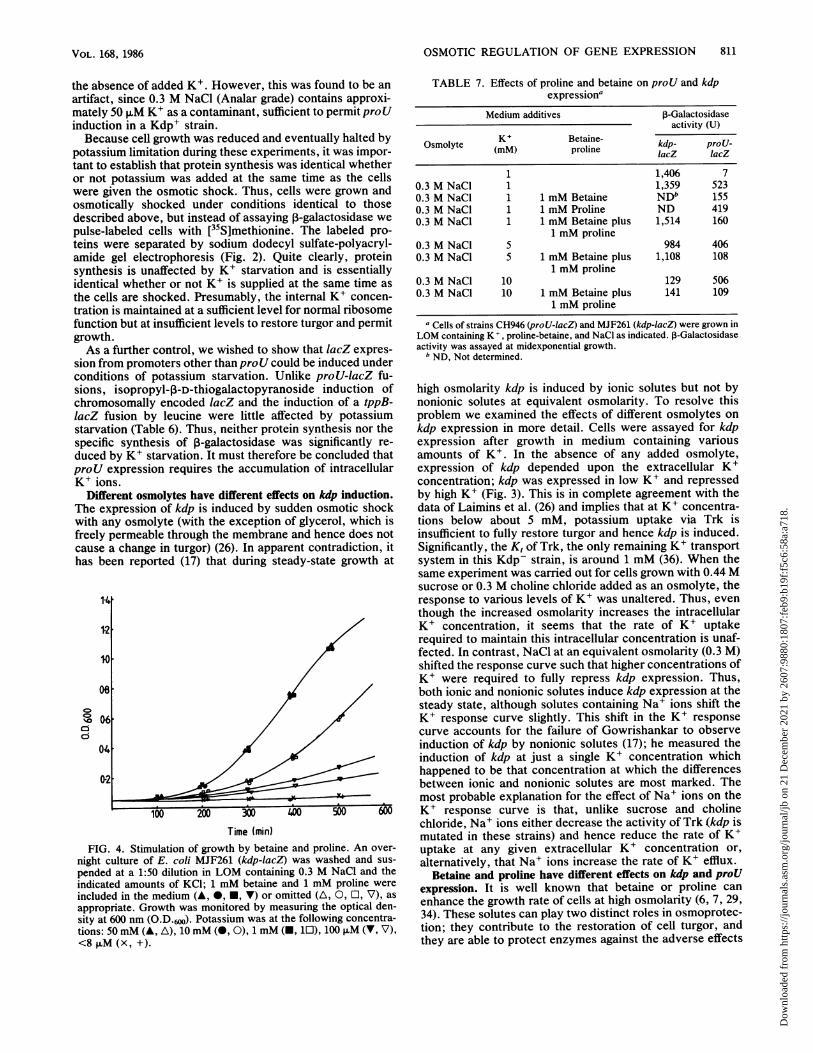
FIG. 4. Stimulation of growth by betaine and proline. An over-
night culture of E. coli MJF261 (kdp-lacZ) was washed and sus-
pended at a 1:50 dilution in LOM containing 0.3 M NaCl and theindicated amounts of KCI; 1 mM betaine and 1 mM proline were
included in the medium (A, *, E, V) or omitted (A, 0, O, V), as
appropriate. Growth was monitored by measuring the optical den-sity at 600 nm (O.D.6w). Potassium was at the following concentra-tions: 50 mM (A, A), 10 mM (0, 0), 1 mM (-, 10), 100 ,uM (V, V),<8 ,uM (x, +).
TABLE 7. Effects of proline and betaine on proU and kdpexpressiona
Medium additives P-Galactosidaseactivity (U)
Osmolyte KM Betaine- kdp- proU-(MM) proline lacZ lacZ
1 1,406 70.3 M NaCI 1 1,359 5230.3 M NaCl 1 1 mM Betaine NDb 1550.3 M NaCl 1 1 mM Proline ND 4190.3 M NaCl 1 1 mM Betaine plus 1,514 160
1 mM proline0.3 M NaCl 5 984 4060.3 M NaCl 5 1 mM Betaine plus 1,108 108
1 mM proline0.3 M NaCl 10 129 5060.3 M NaCl 10 1 mM Betaine plus 141 109
1 mM prolinea Cells of strains CH946 (proU-lacZ) and MJF261 (kdp-lacZ) were grown in
LOM containing K +, proline-betaine, and NaCI as indicated. lB-Galactosidaseactivity was assayed at midexponential growth.
b ND, Not determined.
high osmolarity kdp is induced by ionic solutes but not bynonionic solutes at equivalent osmolarity. To resolve thisproblem we examined the effects of different osmolytes onkdp expression in more detail. Cells were assayed for kdpexpression after growth in medium containing variousamounts of K+. In the absence of any added osmolyte,expression of kdp depended upon the extracellular K+concentration; kdp was expressed in low K+ and repressedby high K+ (Fig. 3). This is in complete agreement with thedata of Laimins et al. (26) and implies that at K+ concentra-tions below about 5 mM, potassium uptake via Trk isinsufficient to fully restore turgor and hence kdp is induced.Significantly, the K, of Trk, the only remaining K+ transportsystem in this Kdp- strain, is around 1 mM (36). When thesame experiment was carried out for cells grown with 0.44 Msucrose or 0.3 M choline chloride added as an osmolyte, theresponse to various levels of K+ was unaltered. Thus, eventhough the increased osmolarity increases the intracellularK+ concentration, it seems that the rate of K+ uptakerequired to maintain this intracellular concentration is unaf-fected. In contrast, NaCl at an equivalent osmolarity (0.3 M)shifted the response curve such that higher concentrations ofK+ were required to fully repress kdp expression. Thus,both ionic and nonionic solutes induce kdp expression at thesteady state, although solutes containing Na+ ions shift theK+ response curve slightly. This shift in the K+ responsecurve accounts for the failure of Gowrishankar to observeinduction of kdp by nonionic solutes (17); he measured theinduction of kdp at just a single K+ concentration whichhappened to be that concentration at which the differencesbetween ionic and nonionic solutes are most marked. Themost probable explanation for the effect of Na+ ions on theK+ response curve is that, unlike sucrose and cholinechloride, Na+ ions either decrease the activity of Trk (kdp ismutated in these strains) and hence reduce the rate of K+uptake at any given extracellular K+ concentration or,alternatively, that Na+ ions increase the rate of K+ efflux.
Betaine and proline have different effects on kdp and proUexpression. It is well known that betaine or proline canenhance the growth rate of cells at high osmolarity (6, 7, 29,34). These solutes can play two distinct roles in osmoprotec-tion; they contribute to the restoration of cell turgor, andthey are able to protect enzymes against the adverse effects
VOL. 168, 1986
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 26
07:9
880:
1807
:feb
9:b1
9f:f
5c6:
58a:
a718
.

812 SUTHERLAND ET AL.
100
25
0 0. 1 0.2 0.3 0-4 0.5 0.6 0-7 0.8
NaCl [MlFIG. 5. Relative accumulation of K+ and betaine. Cells were
grown at high osmolarity (0.3 M NaCl) to induce proU, washed, andequilibrated in media containing the indicated concentrations ofNaCl, and the intracellular K+ and betaine concentrations were
determined as described in Materials and Methods. The results areplotted as a percentage of the total pool of K+ plus betaine.Symbols: vJ, betaine; _, K+.
of high ionic strength (21, 28, 33, 45). During growth at highosmolarity in 1 mM K+, which in a kdp strain limits turgorand induces kdp expression (Table 2) (26), betaine markedlyenhances growth rate (Fig. 4). However, even when strainsare grown at high osmolarity in excess K+ (50 mM), which isbelieved to restore turgor and fully represses kdp expres-
sion, growth rates are still significantly enhanced by betaine.It is, of course, possible that turgor is only partially restoredby K+ accumulation, even in the presence of excess extra-cellular K+, and that betaine results in full restoration ofturgor and thereby enhances growth. However, this expla-nation seems improbable because it has been demonstratedthat at these osmolarities sufficient K+ can be accumulatedto restore turgor (16). More probably, the enhancement ofgrowth rates by betaine in excess potassium is not due torestoration of turgor but to protection of enzyme functionfrom high intracellular ionic strength. Interestingly, at lowK+ concentrations (100 p,M) betaine does not enhancegrowth, implying that at these concentrations K+ itself isgrowth limiting.The effects of proline and betaine on proU and kdp
expression were examined (Table 7). Cells were assayedduring steady-state growth in LOM (1 mM K+) at highosmolarity (0.3 M NaCl), with 1 mM betaine or 1 mM prolineor both added as appropriate. As we have reported previ-ously (7), expression of proU in S. typhimurium is consid-erably reduced by added betaine or proline. This effect is notsimply an effect of growth rate on proU expression; alteringthe growth rate in other ways (i.e., amino acid starvation)did not influence proU expression. Interestingly, betainecaused a greater reduction in proU expression than didproline. The significance of this observation is discussedbelow. Similar results were obtained for pro U-lacZ fusionsin E. coli (data not presented). In contrast, the addition ofbetaine or proline to the growth medium had little or no
effect on the expression of kdp. In fact, betaine actuallyincreased kdp expression slightly. The absence of any majoreffect of betaine on kdp expression is in agreement with thegrowth data presented above, which imply that betaine doesnot play a role in restoring cell turgor under these conditions.These data provide yet another distinction between theregulation ofproU and kdp expression. If, as implied by our
data above, proU expression were dependent upon intracel-lular potassium concentrations, we would predict thatbetaine and proline, which reduce proU expression, alsoreduce the intracellular K+ pools. Table 4 shows that this isindeed the case. However, despite their rather differenteffects on proU expression, proline and betaine actuallyreduce the intracellular K+ pools to similar extents. Theimplications of this apparently inconsistent result are dis-cussed below.
It should be noted that Gowrishankar reported that theaddition of proline to the growth medium does not affectproU expression (17), in apparent contradiction to the re-sults reported here. However, his experiments were carriedout at 0.4 M NaCl, at which osmolarity proline uptake iseffectively abolished in a proU strain (6, 7). In addition, weshow below that there is a difference between growth at 0.3and 0.4 M NaCl in terms of the relative roles of K+ andbetaine in restoring cell turgor.
Distinct roles for K+ and betaine at different osmolarities.Because expression of proU is only fully induced at rela-tively high extracellular osmolarities, once a significantintracellular K+ concentration has already been achieved, itseemed possible that K+ and betaine might play ratherdistinct roles in osmoprotection. We therefore measured therelative amounts of K+ and betaine accumulated by cellsgrown at different osmolarities. As the osmolarity wasincreased the relative role of betaine as the intracellularosmolyte became more important (Fig. 5). In media contain-ing 50 mM NaCl, betaine accounted for less than 10% of thetotal pool of the two osmolytes, whereas at 0.8 M NaClbetaine accounted for more than 80% of this pool.
DISCUSSIONIt is now well established that the transcription of two
bacterial genes, kdp and proU, is determined primarily byosmotic stress (7, 15, 17, 26). However, in neither case is themolecular basis of this regulation understood. The elegantstudies of Epstein have led to the conclusion that expressionof the kdp operon is induced in response to a lowering of cellturgor and that induction of kdp is independent of eitherintracellular or extracellular K+ concentrations (26). Thissystem provides a reference point for the investigation of theexpression of other genes under osmotic control. The findingthat expression of proU, like that of kdp, is induced in cellsunder osmotic stress (7, 15, 17) implied that this gene mightalso be sensitive to changes in turgor. However, the datapresented here illustrate a number of important differencesbetween the regulation of proU and kdp expression. (i)Expression of kdp during growth at low osmolarity is in-duced by potassium starvation, whereas proU remains re-pressed. (ii) Expression of kdp during growth at highosmolarity is repressed by added potassium, whereas ex-pression ofproU remains unaffected. (iii) Expression of kdpin cells grown at high osmolarity is unaffected by theaddition of betaine or proline, whereas expression ofproU issignificantly reduced. These data quite clearly demonstrate adifference in the osmotic regulation of the two genes. It isalso clear from these observations that changes in turgorcannot directly influence proU expression. For example,proU is not expressed under potassium limitation, whichreduces turgor. Similarly, proU is expressed during steady-state growth at high osmolarity, even in the presence ofexcess potassium which results in the substantial, if notcomplete, restoration of turgor.We have now obtained strong evidence that proU expres-
sion is induced in response to an increase in intracellular K+
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 26
07:9
880:
1807
:feb
9:b1
9f:f
5c6:
58a:
a718
.

OSMOTIC REGULATION OF GENE EXPRESSION 813
concentrations. We have manipulated the intracellular po-tassium concentrations in a number of ways, by alteringmedium osmolarity, by limiting potassium availability, andby introducing potassium transport mutants. The changes inintracellular K+ concentrations resulting from these proce-dures have been measured and are in excellent agreementwith previously published results (16, 26; Epstein, in press).We find a strong correlation between proU expression andintracellular potassium: (i) increases in external osmolarity,which result in a corresponding increase in intracellular K+,induce proU. (ii) Potassium starvation, which leads to turgorstress but does not increase intracellular K+, does notinduce proU. (iii) At high osmolarity, limitation of K+ (eitherby starvation or by introducing potassium transport lesions)decreases proU expression, despite the fact that this limita-tion increases turgor stress. It should of course be remem-bered that the induction curve for proU expression issigmiodal (7, 17), implying cooperativity, and that conse-quently a precise linear relationship between K+ concentra-tions and proU expression would not be anticipated. Tomore critically demonstrate a requirement for potassium, wehave shown that sudden osmotic shock fails to induce proUwhen potassium is limiting for growth, even though proteinsynthesis and the induction of other genes are unaffected byK+ starvation. Thus, the accumulation of intracellular K+seems to be both required and sufficient for the induction ofproU. The only circumstances in which the correlationbetween K+ concentrations and proU expression breaksdown is when betaine and proline are provided in the growthmedium. However, the lack of correlation here can readilybe explained by roles for betaine and proline other than inturgor regulation (see below).How might intracellular potassium effect the induction of
proU? One possibility is that the interaction of RNA poly-merase with the proU promoter is directly influenced byionic strength. It is well known that ionic strength can alterpromoter function in vitro, and increases in ionic strength ofthe magnitude which occurs under severe osmotic stressmight induce structural transitions in DNA. More probably acytoplasmic regulatory protein is involved in modulatingproU expression. This protein would undergo a confor-mational change in response to increased concentrations ofintracellular potassium and activate proU transcription. Ei-ther a positive or a negative regulatory protein might beinvolved. Such a conformational change could be achievedin one of two ways. The simplest hypothesis is that K+ bindsto a specific site on the protein. This site would have abinding constant for K+ of around 150 mM, the concentra-tion which results in half-maximal expression of proU (7,17). Alternatively, the conformational changes in the puta-tive regulatory protein could simply be the result of sensi-tivity of the protein structure to fluctuations in ionicstrength. Such a model permits an explanation for the findingthat betaine causes a greater reduction in the expression ofproU than does proline, whereas both have similar effects onintracellular K+ concentrations. It is well known that betaineis a better osmoprotectant than proline (6) and that it isbetter able to protect proteins from ionic or other denatur-ation (30, 31, 33, 45). The different effects of betaine andproline on proU expression may therefore reflect theirrelative efficiencies at protecting the putative regulatoryprotein from conformational changes at high internal ionicstrength. We have shown that, under the conditions usedhere, betaine is able to enhance growth rates even underconditions where sufficient potassium is present to restoreturgor and that therefore betaine must be providing another
function such as the protection of proteins against denatur-ation. It is interesting to note that the induction profile forproU expression is sigmoidal with respect to osmolarity (7).This is compatible with a model in which the putative proUregulatory protein is a dimer (or larger oligomer) and thatrenaturation-denaturation by K+ involves a monomer-dimertransition. Betaine has been shown in vitro to facilitate themultimerization of proteins under otherwise dissociatingconditions (21).
Although unlikely, it is of course possible that K+ ions actindirectly. For example, it may be that the glutamate, whichis synthesized in response to K+ accumulation to restore theionic balance (41), provides the actual signal for proUinduction. Alternatively, the signal may simply be an effectof ionic strength within the cell, to which K+ makes themajor contribution. Since the intracellular concentration ofNa+ is maintained at a low level compared with that of K+(16), it is not possible to distinguish in vivo whether K+functions in a specific manner or is simply the primarycontributing ion to an overall increase in intracellular ionicstrength. We are currently testing this possibility in an invitro system. However, even if the effects of K+ are indirect,the accumulation of K+ still remains the primary event thatleads to the induction of proU expression.The differential regulation of proU and kdp expression is
entirely consistent with the different roles played by betaineand potassium in osmoregulation. We have found that at lowosmolarities potassium is the principal osmotic species,whereas at higher osmolarities betaine (when available)becomes more important. Because K+, unlike betaine, isalmost always available to cells it is clearly the most appro-priate primary osmoprotectant and, when cells are subjectedto mild osmotic stress, is accumulated to restore turgor.However, at high external osmolarities the correspondinglyhigher intracellular K+ concentrations are potentially dele-terious to enzyme function and can reduce growth rates.Thus, if it is available, betaine is taken up in preference toK+. The accumulated betaine contributes to the restorationof turgor, reducing the need to accumulate potentially dele-terious levels of K+. In addition, betaine is known to protectproteins from ionic denaturation (4, 45) and may thereforealso serve to protect intracellular enzymes from inhibition bythe K+ ions which are accumulated. Thus, the cell possessesan integrated response to achieve optimum growth at dif-ferent osmolarities. This scheme is best achieved by induc-ing the expression of the potassium uptake system (kdp) inresponse to changes in turgor and of the betaine uptakesystem (proU) in response to intracellular K+ concentra-tions.
ACKNOWLEDGMENTS
We are grateful to Wolf Epstein for stimulating discussions andsuggestions as to the possible role of ionic strength in regulatingproU expression. He also generously provided the kdp-lacZ fusionstrain. We also thank Rob Reed for assistance with the measure-ments of intracellular K+ concentrations and Evert Bakker for manyhelpful discussions.
This work was supported by a research grant from the MedicalResearch Council to C.F.H. and I.R.B. C.F.H. is a Lister InstituteResearch Fellow.
LITERATURE CITED1. Ahmed, S., and I. R. Booth. 1983. The use of valinomycin,
nigericin and tetrachlorosalicylanilide in the control of theprotonmotive force. Biochem. J. 212:105-112.
2. Ames, G. F.-L. 1974. Resolution of bacterial proteins by poly-acrylamide gel electrophoresis on slabs. J. Biol. Chem.
VOL. 168, 1986
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 26
07:9
880:
1807
:feb
9:b1
9f:f
5c6:
58a:
a718
.

814 SUTHERLAND ET AL.
249:634-644.3. Ames, G. F.-L., and K. Nikaido. 1981. Phosphate-containing
proteins of Salmonella typhimurium and Escherichia coli. Eur.J. Biochem. 115:525-531.
4. Bowlus, R. D., and G. N. Somero. 1979. Solute compatibilitywith enzyme function and structure: rationales for the selectionof osmotic agents and end products of anerobic metabolism inmarine invertebrates. J. Exp. Zool. 208:137-152.
5. Bukau, B., M. Ehrmann, and W. Boos. 1986. Osmoregulation ofthe maltose regulon in Escherichia coli. J. Bacteriol. 166:884-891.
6. Cairney, J., I. R, Booth, and C. F. Higgins. 1985. Salmonellatyphimurium proP gene encodes a transport system for theosmoprotectant betaine. J. Bacteriol. 164:1218-1223.
7. Cairney, J., I. R. Booth, and C. F. Higgins. 1985. Osmoregula-tion of gene expression in Salmonella typhimurium: proUencodes an osmotically induced betaine transport system. J.Bacteriol. 164:1224-1232.
8. Cairney, J., C. F. Higgins, and I. R. Booth. 1984. Proline uptakethrough the major transport system of Salmonella typhimuriumis coupled to sodium ions. J. Bacteriol. 160:22-27.
9. Casadaban, M. 1976. Transposition and fusion of the lac genesto selected promoters in E. coli using bacteriophage lambda andMu. J. Mol. Biol. 104:541-555.
10. Castilho, B. A., P. Olfson, and M. J. Casadaban. 1984. Plasmidinsertion mutagenesis and lac gene fusion with mini-Mubacteriophage transposons. J. Bacteriol. 158:488-495.
11. Christian, J. H. B. 1955. The influence of nutrition on the waterrelations of Salmonella oranienburg. Aust. J. Biol. Sci. 8:75-82.
12. Clark, D., and J. Parker. 1984. Proteins induced by highosmotic pressure in Escherichia coli. FEMS Microbiol. Lett.25:81-83.
13. Csonka, L. N. 1981. Proline over-production results in enhancedosmotolerance in Salmonella typhimurium. Mol. Gen. Genet.182:82-86.
14. Csonka, L. N. 1982. A third L-proline permease in Salmonellatyphimurium which fuinctions in media of elevated osmoticstrength. J. Bacteriol. 151:1433-1443.
15. Dunlap, V. J., and L. N. Csonka. 1985. Osmotic regulation ofL-proline transport in Salmonella typhimurium. J. Bacteriol.163:296-304.
16. Epstein, W., and S. G. Schultz. 1965. Cation transport inEscherichia coli. V. Regulation of cation content. J. Gen.Physiol. 49:221-234.
17. Gowrishankar, J. 1985. Identification of osmoresponsive genesin Escherichia coli: evidence for participation of potassium andproline transport systems in osmoregulation. J. Bacteriol.164:434-465.
18. Grothe, S., R. L. Krogsrud, D. J. McClellan, J. C. Milner, andJ. M. Wood. 1986. Proline transport and osmotic stress re-sponse in Escherichia coli. J. Bacteriol. 166:253-259.
19. Hall, M. N., and T. J. Silhavy. 1981. The ompB locus and theregulation of the major outer membrane porin proteins ofEscherichia coli K12. J. Mol. Biol. 146:23-43.
20. Hall, M. N., and T. J. Silhavy. 1981. Genetic analysis of theompB locus of Escherichia coli K12. J. Mol. Biol. 151:1-15.
21. Hand, S. C., and G. N. Somero. 1982. Urea and methylamineeffects on rabbit muscle phosphofructokinase. J. Biol. Chem.257:734-741.
22. Imhoff, J. F., and F. Rodriguez-Valera. 1984. Betaine is the maincompatible solute of halophilic eubacteria. J. Bacteriol.160:478-479.
23. Jamieson, D. J., and C. F. Higgins. 1984. Anaerobic andleucine-dependent expression of a peptide transport gene inSalmonella typhimurium. J. Bacteriol. 160:131-136.
24. Jamieson, D. J., and C. F. Higgins. 1986. Two geneticallydistinct pathways for transcriptional regulation of anaerobicgene expression in Salmonella typhimurium. J. Bacteriol.
168:389-404.25. Laemmli, U. K. 1970. Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature (London)227:680-685.
26. Laimins, L. A., D. B. Rhoads, and W. Epstein. 1981. Osmoticcontrol of kdp operon expression in Escherichia coli. Proc.Natl. Acad. Sci. USA 78:464-468.
27. Landfald, B., and A. R. Strom. 1986. Choline-glycine betainepathway confers a high level of osmotic tolerance in Escherichiacoli. J. Bacteriol. 165:849-855.
28. LeRudulier, D., T. Bernard, G. Goas, and J. Hamelin. 1984.Osmoregulation in Klebsiella pneumoniae: enhancement ofanaerobic growth and nitrogen fixation under stress by prolinebetaine, a-butyrobetaine and other related compounds. Can. J.Microbiol. 30:299-305.
29. LeRudulier, D., and L. Bouillard. 1983. Glycine betaine, anosmotic effector in Klebsiella pneumoniae and other membersof the Enterobacteriaceae. Appl. Environ. Microbiol. 46:152-159.
30. LeRudulier, D., A. R. Strom, A. M. Dandekar, L. T. Smith, andR. C. Valentine. 1984. Molecular biology of osmoregulation.Science 224:1064-1068.
31. LeRudulier, D., and R. C. Valentine. 1982. Genetic engineeringin agriculture: osmoregulation. Trends Biochetn. Sci. 7:431-433.
32. Miller, J. H. 1972. Experiments in molecular genetics. ColdSpring Harbor Laboratory, Cold Spring Harbor, N.Y.
33. Paleg, L. G., G. R. Stewart, and R. Starr. 1985. The effects ofcompatible solutes on proteins. Dev. Plant Soil Sci. 17:83-94.
34. Perroud, B., and D. LeRudulier. 1985. Glycine betaine transportin Escherichia coli: osmotic modulation. J. Bacteriol.161:393-401.
35. Reed, R. H., J. A. Chudek, R. Foster, and W. D. P. Stewart.1984. Osmotic adjustment in cyanobacteria from hypersalineenvironments. Arch. Microbiol. 138:333-337.
36. Rhoads, D. B., F. B. Waters, and W. Epstein. 1976. Cationtransport in Escherichia coli. VIII. Potassium transport mu-tants. J. Gen. Physiol. 67:325-341.
37. Roth, J. R. 1970. Genetic techniques in studies of bacterialmetabolism. Methods Enzymol. 17A:3-35.
38. Rowland, G. C., P. M. Gifford, and I. R. Booth. 1985. phs locusof Escherichia coli, a mutation causing lesions in metabolism, isan rpoA allele. J. Bacteriol. 164:972-975.
39. Stock, J. B., B. Rauch, and S. Roseman. 1977. Periplasmic spacein Salmonella typhimurium and Escherichia coli. J. Biol. Chem.252:7850-7861.
40. Styrvold, D. B., P. Falkenberg, B. Landfald, M. W. Eshoo, T.Bjornsen, and A. R. Strom. 1986. Selection, mapping and char-acterization of osmoregulatory mutants of Escherichia coliblocked in the choline-glycine betaine pathway. J. Bacteriol.165:856-863.
41. Tempest, D. W., J. L. Meers, and C. M. Brown. 1970. Influenceof environmnent on the content and composition of microbial freeamino acid pools. J. Gen. Microbiol. 64:171-185.
42. van Alphen, W., and B. Lugtenberg. 1977. Influence of osmolar-ity of the growth medium on the outer membrane protein patternof Escherichia coli. J. Bacteriol. 131:623-630.
43. Villarejo, M., J. L. Davis, and S. Granett. 1983. Osmoregulationof alkaline phosphatase synthesis in Escherichia coli K-12. J.Bacteriol. 156:975-978.
44. Wyn Jones, R. G., and R. Storey. 1981. Betaines, p. 171-204. InL. G. Paleg and D. Aspinall (ed.), The physiology and biochem-istry of drought-resistance in plants. Academic Press, Inc.,Sydney, Australia.
45. Yancey, P. H., M. E. Clark, S. C. Hand, R. Bowlus, and G. N.Somero. 1982. Living with water stress: evolution of osmolytesystems. Science 217:1214-1222.
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
21
Dec
embe
r 20
21 b
y 26
07:9
880:
1807
:feb
9:b1
9f:f
5c6:
58a:
a718
.