original article pulmonary immune responses to · pdf filedepartment of plant physiology...
TRANSCRIPT

684 Biomed Environ Sci, 2014; 27(9): 684-694
*This study was supported by the Ministry of Education, Science and Technological Development of the Republic of
Serbia, Grant # 173039.
$MIRKOV Ivana and EL-MUZGHI Amal Atia Mhfuod contributed equally to this work and should be considered as co-first authors.
#Correspondence should be addressed to KATARANOVSKI Milena, PhD, Tel: 381-11-2078375, Fax: 381-11-2761433, E-mail: [email protected]
Biographical notes of the first authors: MIRKOV Ivana, female, born in 1979, PhD, majoring in immunology; EL-MUZGHI Amal Atia Mhfuod, female, born in 1972, PhD candidate, majoring in immunology.
Original Article
Pulmonary Immune Responses to Aspergillus Fumigatus in Rats*
MIRKOV Ivana1,$, EL-MUZGHI Amal Atia Mhfuod1,$, DJOKIC Jelena1, NINKOV Marina1, POPOV ALEKSANDROV Aleksandra1, GLAMOCLIJA Jasmina2, and KATARANOVSKI Milena1,3,#
1. Department of Ecology, Institute for Biological Research ‘Sinisa Stankovic’, University of Belgrade, Belgrade 11000, Serbia; 2. Department of Plant Physiology (Mycology Lab), Institute for Biological Research ‘Sinisa Stankovic’, University of Belgrade, Belgrade 11000, Serbia; 3. Institute of Physiology and Biochemistry, Faculty of Biology, University of Belgrade, Belgrade 11000, Serbia
Abstract
Objective To evaluate immunologic mechanisms underlying Aspergillus fumigatus pulmonary infections in immunocompetent Dark Agouti (DA) and Albino Oxford (AO) rats recognized as being susceptible to some inflammatory diseases in different manners.
Methods Lung fungal burden (quantitative colony forming units, CFU, assay), leukocyte infiltration (histology, cell composition) and their function (phagocytosis, oxidative activity, CD11b adhesion molecule expression) and cytokine interferon-γ (IFN-γ) and interleukin-17 and -4 (IL-17 and IL-4) lung content were evaluated following infection (intratracheally, 1x107 conidia).
Results Slower reduction of fungal burden was observed in AO rats in comparison with that in DA rats, which was coincided with less intense histologically evident lung cell infiltration and leukocyte recovery as well as lower level of most of the their activities including intracellular myeloperoxidase activity, the capacity of nitroblue tetrazolium salt reduction and CD11b adhesion molecule expression (except for phagocytosis of conidia) in these rats. Differential patterns of changes in proinflammatory cytokine levels (unchanged levels of IFN-γ and transient increase of IL-17 in AO rats vs continuous increase of both cytokines in DA rats) and unchanged levels of IL-4 were observed.
Conclusion Genetically-based differences in the pattern of antifungal lung leukocyte activities and cytokine milieu, associated with differential efficiency of fungal elimination might be useful in the future use of rat models in studies of pulmonary aspergillosis.
Key words: A. fumigatus; Rats; Lung immune responses
Biomed Environ Sci, 2014; 27(9): 684-694 doi: 10.3967/bes2014.104 ISSN: 0895-3988
www.besjournal.com (full text) CN: 11-2816/Q Copyright ©2014 by China CDC

Immune response to pulmonary aspergillosis in rats 685
INTRODUCTION
at models of pulmonary aspergillosis are widely used in the assessment of advantageousness of different laboratory
diagnostic methodologies in infection with the fungus[1-7]. Experimental infection of immuno- suppressed rats with Aspergillus fumigatus is used in antifungal drug testing as well[8-12]. Reduction of fungal burden is most frequently used as an indicator of effectiveness of antifungal therapy, but some parameters of immune response are proven to be useful as well. For instance, lung myeloperoxidase (MPO) content was used as a marker of effect of pentraxin administration in a rat model of invasive pulmonary aspergillosis[13]. Decrease of fungal load in cyclophosphamide-immunosuppressed (neutropenic) RP albino rats treated with amphotericin B coincided with lower levels of innate immunity cytokines including IL-6 and chemokines monocyte chemotactic protein (MCP-1) and macrophage inflammatory protein (MIP-2), in lung homogenates[14].
Despite the significant use of rat models of pulmonary infection with A. fumigatus in therapeutic and diagnostic practice, immune-based mechanisms of resistance/sensitivity to the fungus are not well known. Early studies showed granuloma formation in regional lymph nodes of rats administered with A. fumigatus[15]. Histologically evident signs of immune reactivity to fungus (i.e. lung cell infiltration) were demonstrated in several reports of pulmonary infection in rats at the early infection stage (one day post infection)[16-19]. Different data were obtained concerning cytokine involvement in antifungal pulmonary responses. In one report the authors showed that infection with A. fumigatus was not associated with changes in the lung interferon-γ (IFN-γ), tumor necrosis factor-α (TNF-α), interleukin-4 (IL-4) or IL-10 content in neutropenic RP albino rats[14]. In another study, the lack of changes in lung IFN-γ, instead of an increase in lung IL-10 levels, was observed in immunosuppressed (5-fluorouracil-treated) infected Wistar albino rats[20]. Our recent study showed that resistance of immunocompetent (non-suppressed) Dark Agouti (DA) rats to pulmonary aspergillosis was associated with an increase in IFN-γ and proinflammatory cytokine IL-17[21]. Although differences in the antigenic composition of A. fumigatus conidia used in infection or the character of immunosuppression might be responsible for disparate patterns of
cytokine responses to fungus, genetic background (i.e. the rat strain used in above cited studies) might have exerted influence as well. In corroboration, studies in humans and mice showed the relevance of genetic background for susceptibility to pulmonary aspergillosis[22-24].
Based on the above described background concerning paucity of data on mechanisms of pulmonary responses to A. fumigatus in rats and with the knowledge of relevance of genetically-based variations for the susceptibility to aspergillosis in mice and humans, immune responses to sublethal pulmonary infection with A. fumigatus in rats were examined comparatively in individuals of two rat strains, Albino Oxford (AO), and Dark Agouti (DA). These two strains were shown to differ in responsiveness to inflammatory stimuli in various tissues including kidneys[25], nervous system[26], skin[27], and at systemic level as well[28-29]. Reduction of lung fungal burden was monitored in order to get gross insight into antifungal defense. Immune-based activities of lung leukocytes which are relevant for antifungal defense (oxidative activity as most efficient means of destruction of fungus in particular) [30-32], as well as lung cytokine milieu shown to be relevant in infection of DA rats[21], were monitored early (day 1 post infection, p.i.) and late in infection (day 7 p.i.). Immunocompetent rats were used in the study, as antifungal agents might exert immune modulating activity[33]. The knowledge of genetically-based immune-relevant differences in mechanisms of defense against A. fumigatus in rats might be helpful in the future use of rat models of pulmonary aspergillosis in immunotherapeutic antifungal studies.
MATERIALS AND METHODS
Chemicals
Nitroblue tetrazolium (NBT), hexadecyltrimeth- ylammonium bromide (HTAB), o-dianisidine dihydrochloride, myeloperoxidase (MPO), deoxyribonuclease I (DNase I) and bovine serum albumin (BSA) (all purchased from Sigma, Sigma Aldrich., St. Louis, MO, USA), type IV collagenase (Worthington Biochemical Corporation, Lakewood, USA), paraformaldehyde and NaN3 purchased from Serva (Heidelberg, Germany), hydrogen peroxide (H2O2) from Zorka Farma, Sabac (Serbia), were used in experiments. Collagenase and DNase were dissolved in culture medium. NBT was dissolved in
R

686 Biomed Environ Sci, 2014; 27(9): 684-694
phosphate buffered saline (PBS, pH 7.2). These solutions were sterile filtered (Minisart, pore size 0.20 µm; Sartorius Stedim biotech, Goettingen, Germany) before use. Culture medium RPMI-1640 supplemented with 2 mmol/L glutamine (PAA laboratories, Austria), 20 μg/mL gentamycine (Galenika a.d., Serbia), 5% (v/v) heat inactivated fetal calf serum (PAA laboratories, Austria) were used in cell culture experiments. Lysis buffer (eBioscience Inc., San Diego, CA, USA) was used for lysis of red blood cells in lung cell digest preparations. Phycoerythrin (PE)-labeled mouse monoclonal antibody to rat NKG2D (marker of NK cells), fluorescein isothyocyanate (FITC)-conjugated mouse anti-rat CD3 and mouse anti-rat CD4, and PE-conjugated mouse anti-rat CD8 were all purchased from eBioscience Inc. (San Diego, CA, USA) Monoclonal antibody OX-42 (mouse anti-rat CD11b/CD11c) and FITC-conjugated F(ab')2 goat anti-mouse IgG were purchased from Serotec Ltd (Bicester, UK).
Rats
Male Dark Agouti (DA) and Albino Oxford (AO) rats were bred and conventionally housed at the Institute for Biological Research ‘Sinisa Stankovic’ (Belgrade, Serbia) in this study. Animals were ten to twelve weeks old and 4-6 individuals were assigned per group. All animal procedures were in compliance with the EEC Directive (86/609/EEC) on the protection of animals used for experimental and other scientific purposes, and were approved by the Ethical Committee for the Use of Laboratory Animals of the Institute for Biological Research ‘Sinisa Stankovic’, University of Belgrade (No 2-23/10).
Fungal Culture Conditions and Infection
Aspergillus fumigatus (human isolate obtained from the Institute of Public Health of Serbia ‘Dr Milan Jovanovic-Batut’) was grown on Sabouraud maltose agar (SMA, Torlak, Belgrade, Serbia) for seven days[34]. Conidia suspension obtained by flooding the surface of agar slants with pyrogen-free sterile physiological saline was adjusted to a concentration of 5×107 conidia/mL and injected into the trachea of anesthetized rats (Ketamidor, Richter Pharma, Wels, Austria) by tracheostomy (107 conidia of A. fumigatus in 0.2 mL of saline). Control group (sham infected rats) received saline only. Rats were euthanized at 1, 7, and 14 d p.i. for fungal burden measurements and functional analysis.
Lung Homogenates
Lungs were removed aseptically, and wet mass was measured by weighing using a precision balance (+/-0.01 g). Tissue (whole lungs) was homogenized on ice using an IKA T18 basic homogenizer (IKA Works INC., Wilmington NC) in 10 mL of sterile saline.
Fungal Burden Measurements
Quantitative colony forming units (CFU)[35] assay was used for fungal burden measurements in lung homogenates. Primary homogenate dilutions were quantitatively cultured by serial dilution, plated on SMA plates supplemented with streptomycin sulfate (ICN-Galenika, Belgrade, Serbia), and incubated at 37 °C for 24 to 48 h. After the period of incubation, the number of CFU was counted and expressed as CFU per gram of tissue.
Lung Histology
Middle lobe of the right lung was used for processing for histology. Lobes were excised and immediately placed in 4% formaldehyde (pH 6.9) for fixation. After processing, tissue samples were embedded in paraffin wax for sectioning at 5 µm. Hematoxylin and eosin (H&E)-stained histological slides were analyzed subsequently using Coolscope digital light microscope (Nikon Co, Tokyo, Japan). Lung specimens were scored for the intensity of leukocyte infiltration. Semi-quantitative numeric values were assigned for these scores according to the scale: 0-no infiltrates, 1-small, 2-moderately large, and 3-large infiltrates.
Isolation of Lung Leukocytes
Leukocytes were obtained by digestion of lung tissue from sham infected and infected rats with 1 mg/mL of collagenase and 30 μg/mL DNase I for half an hour at 37 °C[36]. Following erythrocyte lysis, cells were resuspended in RPMI-1640 culture medium (PAA Laboratories, Pasching, Austria) supplemented with 2 mmol/L glutamine, 20 μg/mL gentamicin (Galenika a.d., Belgrade, Serbia), 5% (v/v) heat-inactivated fetal calf serum (PAA Laboratories, Pasching, Austria). Viability was determined by Trypan blue exclusion and always exceeded 90%. Cell composition was determined by differentiating at least 300 cells from cytospin preparations stained with May Grünwald-Giemsa[37] (Alkaloid AD, Skopje, Macedonia).

Immune response to pulmonary aspergillosis in rats 687
Flow Cytometry
Expression of cell surface molecules in question was analyzed by flow cytometry analysis of lung leukocytes obtained by enzymatic digestion and following lysis of erythrocytes with commercial lysis buffer. Cells were incubated on ice for 30 min with antibodies characteristic for lineage or activation/ function markers for rat leukocytes including T cell marker (CD3), NK cell marker (NKG2D) and adhesion molecules CD11b. All antibodies were conjugated with fluorochromes (see Materials and Methods). For CD11b expression measurements, lung cells were incubated with mouse anti-rat CD11b/CD11c monoclonal antibody OX-42, followed by FITC-conjugated F(ab')2 goat anti-mouse IgG for 30 min. After washing in PBS-0.01% NaN3, cells were fixed in 1% paraformaldehyde and assayed for fluorescence intensity on CyFLOW SPACE (Partec, Munich, Germany).
Lung Leukocyte Assays
For estimation of phagocytosis, lung cells (106 in 0.5 mL of culture medium) were placed on 24 mm × 24 mm sterile glass coverslips in 6-well plates and incubated with 106 viable A. fumigatus conidia at 37 °C for 1 h. After incubation, coverslips were washed and adherent cells were stained by May Grünwald-Giemsa protocol. Cells with internalized conidia were identified and counted by immersion light microscopy. Phagocytic activity was expressed as percent of cells with ingested conidia of the 300 cells counted.
Reduction of nitroblue tetrazolium (NBT) salt assay was employed to assess spontaneous lung cell-derived superoxide formation as described by Shen et al[38]. Briefly, NBT (10 μL, 5 mg/mL) was added to the lung cells (1×105 cells in 100 μL of culture medium) and incubated for 20 min at 37 °C followed by 10 min incubation at room temperature. Enumeration of cells with intracellular dots of formazan (NBT-positive cells) was performed under an optical microscopy on cytospin preparations of lung cells stained by May Grünwald-Giemsa.
Lung cell myeloperoxidase activity was assessed on the basis of the oxidation of o-dianisidine dihydrochloride by cells[39]. Briefly, 33 μL of cell (5×104) lysate supernatant was added to 966 μL of substrate solution (0.167 mg/mL o-dianisidine dihydrochloride and 0.0005% H2O2 in 50 mmol/L potassium phosphate buffer, pH 6.0). Absorbance was read at 450 nm at three-minute intervals during ten-minute measurement against the standard of
myeloperoxidase. Values were expressed as MPO units per 106 cells.
ELISA
Cytokine content in lung homogenates was determined using commercially available ELISA sets for rat IFN-γ and IL-4 (R&D Systems, Minneapolis, USA) and mouse IL-17 cross-reactive with rat IL-17 (eBioscience Inc., San Diego, CA, USA). Tests were done according to manufacturer’s recommendations. Cytokine titer was calculated by a reference to a standard curve constructed using the known amounts of recombinant IFN-γ, IL-4, or IL-17.
Data Presentation and Statistics
Results were pooled from two independent experiments (4-6 animals per group in each of the experiments) and expressed as mean values±SD. Statistical analysis of data was performed using a Mann-Whitney U test. P-values less than 0.05 were considered significant.
RESULTS
Lung Fungal Burden Following Administration of A. fumigatus Conidia to AO and DA Rats
Animals were infected and monitored for 14 d. There were no signs of disease or deaths during observation period. No change was observed in wet lung mass of infected AO rats (2.7±0.7 g and 3.0± 0.9 g at days 1 and 7, respectively), compared to sham infected AO rats at respective time points (2.8±0.7 g and 2.6±0.6 g). Similarly, there were no differences between masses of the lungs of infected individuals of DA strain (1.9±0.4 g and 2.0±0.4 g at days 1 and 7, respectively) in comparison to 1.7±0.4 g and 1.9±0.6 g in sham infected rats at respective time points. Progressive decrease in the lung fungal burden was observed in infected individuals of both strains, but it was more efficient in DA rats, which eliminated infection (CFU/g tissue=0.0±0.0) by the day 14 p.i. (Figure 1).
Lung Histology
Histological examination of lung tissue conducted at days 1 and 7 p.i. revealed signs of inflammatory response characterized by perivascular leukocyte infiltration and interstitial widening at both time points following infection (Figure 2). Semi-quantitative analysis of lung tissue specimens revealed no strain differences in the intensity of cell

688 Biomed Environ Sci, 2014; 27(9): 684-694
Figure 1. Time course of fungal burden reduction in lungs of AO and DA rats following intratracheal administration of A. fumigatus conidia. Results are expressed as mean±SD of individual samples from at least two independent experiments with 4-6 animals for each post infection time point per experiment. Not detectable (n.d.). Significance at P<0.05 (#) vs DA rats.
Figure 2. Histological appearance of lungs following intratracheal A. fumigatus administration. A. Leukocyte infiltrations in lungs of infected AO rats at day 1 p.i. B. Histological appearance of lungs of DA rats at day 1 p.i. C. Perivascular leukocyte infiltrations in lungs of DA rats at day 7 p.i. Insert: Histological appearance of lungs of infected AO rats at day 7 p.i.
infiltration at day 1 p.i. judging by similar leukocyte infiltration score values (1.8±1.3 and 2.3±1.0 in AO and DA rats, respectively). The intensity of cell infiltration was more intense in DA rats at day 7 p.i. (1.8±0.8 in DA vs 0.3±0.4 in AO rats, P=0.0043).
Lung Leukocyte Recovery and Cell Composition
Higher numbers of leukocytes were recovered from lungs of infected individuals compared to sham infected individuals of both strains, though only numerically (P=0.113) at day 7 post infection (p.i.) in AO rats (Figure 3A). The yield of lung leukocytes in AO rats at day 7 p.i. was similar (P=0.72) to the one recovered at day 1 p.i., while it was significantly higher compared to that at day 1 p.i. in DA rats. No differences were observed in numbers of lymphocytes in lung digest cell preparations of infected and sham infected individuals of both strains at both time points (Figure 3B). Similar numbers of macrophages were found at day 1 p.i in both strains in sham infected and infected rats (Figure 3C). Numbers of macrophages rose at day 7 p.i. in comparison to those at day 1 p.i. in both sham infected as well as in infected rats of both strains, but they did not exceed those in sham infected rats. Increased numbers of neutrophils were obtained from lungs of infected animals of both strains, but it was evident only as a tendency (P=0.067) of an increase in numbers at day 7 p.i. in AO rats, in contrast to significantly higher numbers at both time points in DA rats (Figure 3D). Numbers of neutrophils recovered from lungs of infected AO rats at this time point were significantly lower compared to DA rats.
As NK cells and T lymphocytes are involved in antifungal responses in mice, flow cytometry analysis of presence of these cells in lung digest leukocyte preparation was determined (Table 1). Increased numbers of cells expressing marker of NK cells were noted in infected animals of both rat strains at day 1 p.i., with a tendency (P=0.06) of lower numbers of positive cells in AO compared to those in DA rats. Return to values close to controls was observed later in infection (day 7 p.i.). Similar numbers of leukocytes expressing CD3 molecules (T cells) were noted at day 1 p.i., with an increase at 7 p.i. No differences were observed in basic (CD4 and CD8) T cell subsets (not shown).
Lung Cell Activity Following Administration of A. fumigatus Conidia to AO and DA Rats
Increased percentages of cells with the capacity of conidia phagocytosis were observed at day 1 p.i.
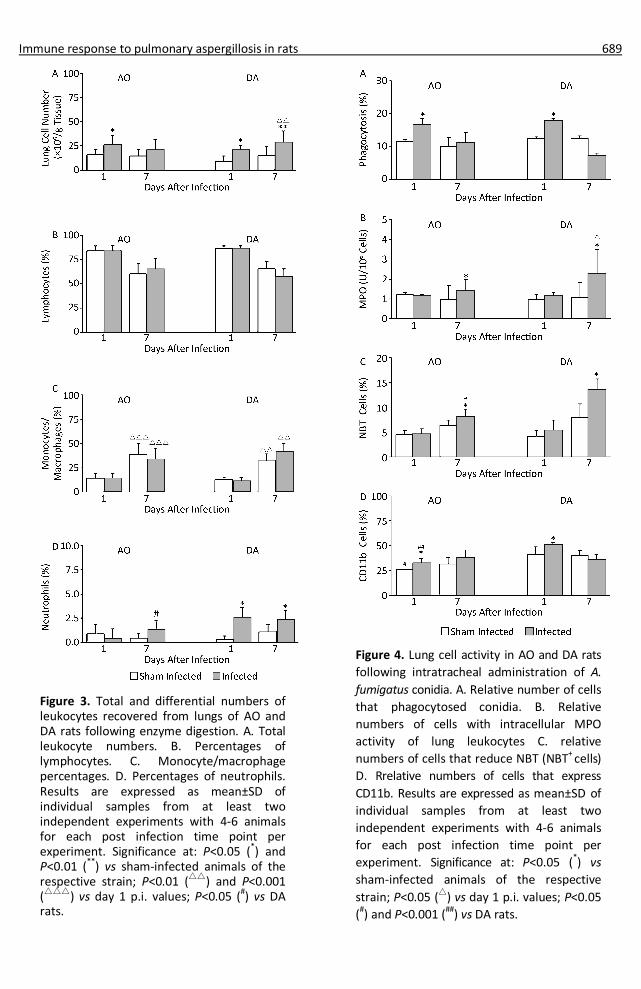
Immune response to pulmonary aspergillosis in rats 689
Figure 3. Total and differential numbers of leukocytes recovered from lungs of AO and DA rats following enzyme digestion. A. Total leukocyte numbers. B. Percentages of lymphocytes. C. Monocyte/macrophage percentages. D. Percentages of neutrophils. Results are expressed as mean±SD of individual samples from at least two independent experiments with 4-6 animals for each post infection time point per experiment. Significance at: P<0.05 (*) and P<0.01 (**) vs sham-infected animals of the respective strain; P<0.01 (△△) and P<0.001 (△△△) vs day 1 p.i. values; P<0.05 (#) vs DA rats.
Figure 4. Lung cell activity in AO and DA rats following intratracheal administration of A. fumigatus conidia. A. Relative number of cells that phagocytosed conidia. B. Relative numbers of cells with intracellular MPO activity of lung leukocytes C. relative numbers of cells that reduce NBT (NBT+ cells) D. Rrelative numbers of cells that express CD11b. Results are expressed as mean±SD of individual samples from at least two independent experiments with 4-6 animals for each post infection time point per experiment. Significance at: P<0.05 (*) vs sham-infected animals of the respective strain; P<0.05 (△) vs day 1 p.i. values; P<0.05 (#) and P<0.001 (##) vs DA rats.

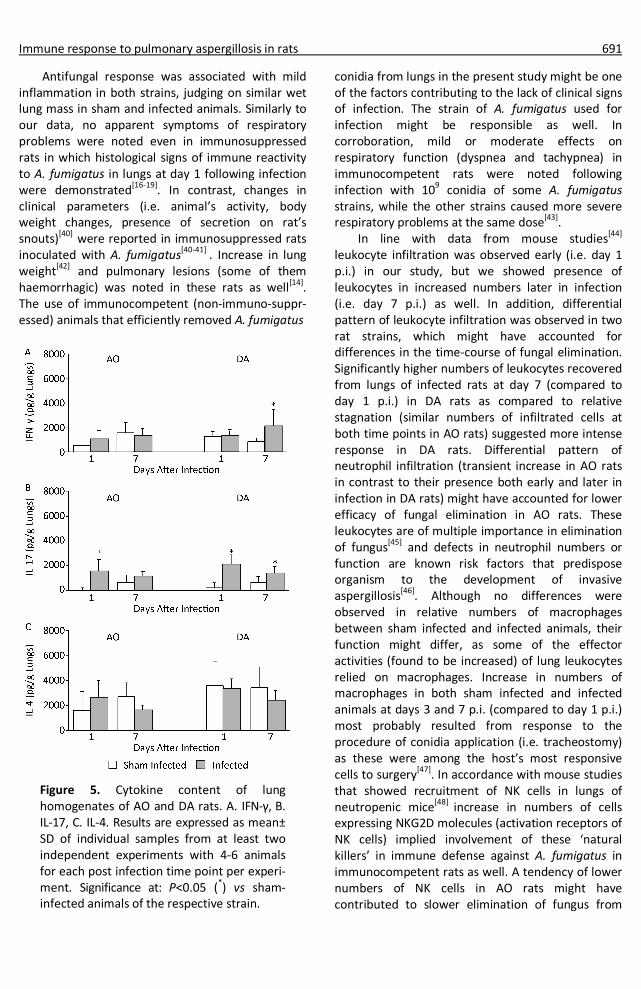
Immune response to pulmonary aspergillosis in rats 691
Antifungal response was associated with mild inflammation in both strains, judging on similar wet lung mass in sham and infected animals. Similarly to our data, no apparent symptoms of respiratory problems were noted even in immunosuppressed rats in which histological signs of immune reactivity to A. fumigatus in lungs at day 1 following infection were demonstrated[16-19]. In contrast, changes in clinical parameters (i.e. animal’s activity, body weight changes, presence of secretion on rat’s snouts)[40] were reported in immunosuppressed rats inoculated with A. fumigatus[40-41] . Increase in lung weight[42] and pulmonary lesions (some of them haemorrhagic) was noted in these rats as well[14]. The use of immunocompetent (non-immuno-suppr- essed) animals that efficiently removed A. fumigatus
Figure 5. Cytokine content of lung homogenates of AO and DA rats. A. IFN-γ, B. IL-17, C. IL-4. Results are expressed as mean± SD of individual samples from at least two independent experiments with 4-6 animals for each post infection time point per experi- ment. Significance at: P<0.05 (*) vs sham- infected animals of the respective strain.
conidia from lungs in the present study might be one of the factors contributing to the lack of clinical signs of infection. The strain of A. fumigatus used for infection might be responsible as well. In corroboration, mild or moderate effects on respiratory function (dyspnea and tachypnea) in immunocompetent rats were noted following infection with 109 conidia of some A. fumigatus strains, while the other strains caused more severe respiratory problems at the same dose[43].
In line with data from mouse studies[44] leukocyte infiltration was observed early (i.e. day 1 p.i.) in our study, but we showed presence of leukocytes in increased numbers later in infection (i.e. day 7 p.i.) as well. In addition, differential pattern of leukocyte infiltration was observed in two rat strains, which might have accounted for differences in the time-course of fungal elimination. Significantly higher numbers of leukocytes recovered from lungs of infected rats at day 7 (compared to day 1 p.i.) in DA rats as compared to relative stagnation (similar numbers of infiltrated cells at both time points in AO rats) suggested more intense response in DA rats. Differential pattern of neutrophil infiltration (transient increase in AO rats in contrast to their presence both early and later in infection in DA rats) might have accounted for lower efficacy of fungal elimination in AO rats. These leukocytes are of multiple importance in elimination of fungus[45] and defects in neutrophil numbers or function are known risk factors that predispose organism to the development of invasive aspergillosis[46]. Although no differences were observed in relative numbers of macrophages between sham infected and infected animals, their function might differ, as some of the effector activities (found to be increased) of lung leukocytes relied on macrophages. Increase in numbers of macrophages in both sham infected and infected animals at days 3 and 7 p.i. (compared to day 1 p.i.) most probably resulted from response to the procedure of conidia application (i.e. tracheostomy) as these were among the host’s most responsive cells to surgery[47]. In accordance with mouse studies that showed recruitment of NK cells in lungs of neutropenic mice[48] increase in numbers of cells expressing NKG2D molecules (activation receptors of NK cells) implied involvement of these ‘natural killers’ in immune defense against A. fumigatus in immunocompetent rats as well. A tendency of lower numbers of NK cells in AO rats might have contributed to slower elimination of fungus from

692 Biomed Environ Sci, 2014; 27(9): 684-694
lungs of these rats, as NK cell contribution was recognized in the early innate immune defense[49]. Late (day 7 p.i.) increase in numbers of CD3+ (T cells) implied existence of adaptive immune responses. T cells are of utmost importance in antifungal responses as producers of cytokines, mediators and regulators of immune responses[44,50].
Increase in numbers of leukocytes having the capacity to ingest conidia observed in this study is in line with data from pulmonary aspergillosis in mice that showed increased phagocytic capacity of alveolar macrophages early (day 1) in post-infection period[30]. Higher numbers of NBT+ cells in infected lungs of both strains reflect the need for such an activity. As production of reactive oxygen species is one of the main effector activity of alveolar macrophages in murine aspergillosis[30], NBT reducing activity of lung leukocytes might be completely ascribed to macrophages. However, neutrophils could have accounted to this aspect of oxidative activity as well[31]. Although neutrophils are traditional sources of MPO[51], macrophages can also increase their MPO content when activated[52]. Lower MPO activity as well as numbers of NBT+ cells in lung leukocyte population of infected AO as compared to DA rats suggest lower propensity of leukocytes from this strain to respond to inflammatory stimuli. In corroboration, we showed lower expression of oxidative activity in leukocytes from peripheral blood of AO rats in settings of systemic inflammation[28]. Lower magnitude of an increase in numbers of NBT+ leukocytes as well as their lower MPO activity in AO compared to DA rats might be responsible for lower efficiency of fungal removal in AO rats, as conjoint antifungal activities of alveolar macrophages[30] and neutrophils[32] are necessary for killing of A. fumigatus.
Increased numbers of lung leukocytes that express CD11b integrin molecules (α subunit of β2 integrin receptors CD11b/CD18) in infected rats of both rat strains (compared to sham infected individuals) early (day 1 p.i.) following infection suggest changes in lung leukocyte activity. These integrin receptors are normally present on macrophages and neutrophils and on small numbers of lymphocytes and are important for their extravasation and migration to peripheral tissues as well as for functional activation of these cells[53]. Increased density of expression on lung leukocyte surface implies their activation, as upregulation of CD11b molecules was noted following cell activation[54]. Although lower numbers of CD11b+
cells in infected AO compared to infected DA rats at day 1 p.i. might rely on lower numbers in sham infected animals, this difference might have exerted influence on the efficacy of elimination of fungus.
Fungal elimination in lungs of AO rats took place in milieu enriched in inflammatory cytokine content, but with unchanged IL-4. Given the fact that T cells are the main source of IFN-γ late in infection,
increase in CD3+ might be responsible for higher lung IFN-γ content in DA compared to AO rats[55]. However, increase in CD3+ cells was similar in both strains, which implied other sources of this cytokine. In this regard, IFN-γ production by neutrophils was reported in some infections in mice[56] and increased numbers of these cells were found at day 7 p.i in DA rats (unchanged in AO rats). Differences in the numbers of activated (NBT+) lung cells in AO and DA rats might possibly be ascribed to different levels of IFN-γ in lung homogenates, as this cytokine is necessary for oxidative activities of phagocytes that are required for killing of ingested A. fumigatus conidia and hyphal damage[31,57]. Lack of increase in IFN-γ in AO rats is in line with data that showed no changes in this cytokine in other studies with rats[14,20]. However, in this study increased levels of IFN-γ in homogenates of lungs of AO rats were found three days following infection (2352±923 pg/mL compared to 1202±701 pg/mL in sham infected AO controls, P<0.05), though higher values (P<0.05) of IFN-γ were observed in infected DA rats (4326±1405 pg/mL) at that time point. This, along with data that showed transient increase in IL-17 in AO as compared to DA rats, suggests that not only the intensity, but the dynamics of cytokine response might be involved in differences in efficiency of antifungal responses.
In conclusion, data presented in this study have shown for the first time that resistance to A. fumigatus in immunocompetent rats of two different strains is associated with immune responses that differ in the type, intensity, as well as in the time course. The knowledge of genetically-based immune-relevant differences in mechanisms of defense against A. fumigatus in rats might be helpful in the future use of rat models of pulmonary aspergillosis in developing and testing immunotherapeutic interventions for fungal diseases.
ACKNOWLEDGEMENTS
The authors would like to thank Dr Lidija

Immune response to pulmonary aspergillosis in rats 693
Zolotarevski for his help with histology.
CONFLICT OF INTEREST
The authors declare no conflict of interest. Received: September 24, 2013; Accepted: December 10, 2013
REFERENCES
1. Yu B, Niki Y, Armstrong D. Use of immunoblotting to detect Aspergillus fumigatus antigen in sera and urines of rats with experimental invasive aspergillosis. J Clin Microbiol, 1990; 28, 1575-9.
2. Becker MJ, de Marie S, Willemse D, et al. Quantitative galactomannan detection is superior to PCR in diagnosing and monitoring invasive pulmonary aspergillosis in an experimental rat model. J Clin Microbiol, 2000; 38, 1434-8.
3. Scotter JM, Chambers ST. Comparison of galactomannan detection, PCR-enzyme-linked immunosorbent assay, and real-time PCR for diagnosis of invasive aspergillosis in a neutropenic rat model and effect of caspofungin acetate. Clin Diagn Lab Immunol, 2005; 12, 1322-7.
4. Khan ZU, Ahmad S, Theyyathel AM. Detection of Aspergillus fumigatus-specific DNA, (1-3)-beta-D-glucan and galacto- mannan in serum and bronchoalveolar lavage specimens of experimentally infected rats. Mycoses, 2008; 51, 129-35.
5. Aydogan S, Kustimur S, Kalkanci A. Comparison of glucan and galactomannan tests with real-time PCR for diagnosis of invasive aspergillosis in a neutropenic rat model. Mikrobiyol Bul, 2010; 44, 441-52.
6. Zhao Y, Park S, Warn P, et al. Detection of Aspergillus fumigatus in a rat model of invasive pulmonary aspergillosis by real-time nucleic acid sequence-based amplification. J Clin Microbiol, 2010; 48, 1378-83.
7. McCulloch E, Ramage G, Rajendran R, et al. Antifungal treatment affects the laboratory diagnosis of invasive aspergillosis. J Clin Pathol, 2012; 65, 83-6.
8. Miyazaki HM, Kohno S, Miyazaki Y, et al. Efficacy of intravenous itraconazole against experimental pulmonary aspergillosis. Antimicrob Agents Chemother, 1993; 37, 2762-5.
9. Kurtz MB, Bernard EM, Edwards FF, et al. Aerosol and parenteral pneumocandins are effective in a rat model of pulmonary aspergillosis. Antimicrob Agents Chemother, 1995; 39, 1784-9.
10. Cicogna CE, White MH, Bernard EM, et al. Efficacy of prophylactic aerosol amphotericin B lipid complex in a rat model of pulmonary aspergillosis. Antimicrob Agents Chemother, 1997; 41, 259-61.
11.Murphy M, Bernard EM, Ishimaru T, et al. Activity of voriconazole (UK-109,496) against clinical isolates of Aspergillus species and its effectiveness in an experimental model of invasive pulmonary aspergillosis. Antimicrob Agents Chemother, 1997; 41, 696-8.
12.Van de Sande WW, Mathot RA, ten Kate MT, et al. Combination therapy of advanced invasive pulmonary aspergillosis in transiently neutropenic rats using human pharmacokinetic equivalent doses of voriconazole and anidulafungin. Antimicrob Agents Chemother, 2009; 53, 2005-13.
13.Lo Giudice P, Campo S, Verdoliva A, et al. Efficacy of PTX3 in a rat model of invasive aspergillosis. Antimicrob Agents Chemother, 2010; 54, 4513-5.
14.Becker MJ, de Marie S, Fens MHAM, et al. Effect of amphotericin B treatment on kinetics of cytokines and parameters of fungal load in neutropenic rats with invasive pulmonary aspergillosis. J Antimicrob Chemother, 2003; 52, 428-34.
15.Turner KJ, Hackshaw R, Papadimitriou J, et al. Experimental aspergillosis in rats infected via intraperitoneal and subcutaneous routes. Immunology, 1975; 29, 55-66.
16.Turner KJ, Hackshaw R, Papadimitriou J, et al. The pathogenesis of experimental pulmonary aspergillosis in normal and cortisone-treated rats. J Pathol, 1976; 118, 65-73.
17.Niki Y, Bernard EM, Edwards FF, et al. Model of recurrent pulmonary aspergillosis in rats. J Clin Microbiol, 1991; 29, 1317-22.
18.Shibuya K, Takaoka M, Uchida K, et al. Histopathology of experimental invasive pulmonary aspergillosis in rats: pathological comparison of pulmonary lesions induced by specific virulent factor deficient mutants. Microb Pathog, 1999; 27, 123-31.
19.Mahmoud YAG, Al-Ghamdi AY, El-Zaher EHFA. A protective mechanism in lungs of rats experimentally infected with Aspergillus fumigatus. Mycobiology, 2011; 39, 40-4.
20.Caglar K, Kalkanci A, Findan I, et al. Investigation of interleukin-10, tumor necrosis factor-alpha and interferon-gamma expression in experimental model of pulmonary aspergillosis. Mikrobiyol Bul, 2011; 45, 344-52.
21.El-Muzghi A, Mirkov I, Djokic J, et al. Regional cytokine responses to pulmonary aspergillosis in immunocompetent rats. Immunobiology, 2013; doi: 10.1016/j.imbio.2013.05.007.
22.Ok M, Einsele H, Loeffler J. Genetic susceptibility to Aspergillus fumigatus infections. Int J Med Microbiol, 2011; 301, 445-52.
23.Zaas AK, Liao G, Chien JW, et al. Plasminogen alleles influence susceptibility to invasive aspergillosis. PLoS Genet, 2008; 4, e1000101.
24.Svirshchevskaya EV, Shevchenko MA, Huet D, et al. Susceptibility of mice to invasive aspergillosis correlates with delayed cell influx into the lungs. Int J Immunogenet, 2009; 36, 289-99.
25.Mijatovic S, Ejdus L, Pravica V, et al. Strain-dependent induction and modulation of autoimmunity by mercuric chloride in two strain of rats. In Immunoregulation in Health and Disease. Experimental and Clinical Aspects (ML Lukic, M Colic, M Mostarica-Stojkovic, K Cuperlovic, Eds.), 1997; pp. 181-8. Academic Press, London.
26.Miljković Đ, Stanojević Ž, Momčilović M, et al. CXCL12 expression within the CNS contributes to the resistance against experimental autoimmune encephalomyelitis in Albino Oxford rats. Immunobiology, 2011; 216, 979-87.
27.Popov A, Mirkov I, Miljković Đ, et al. Contact allergic response to dinitrochlorobenzene (DNCB) in rats: insight from sensitization phase. Immunobiology, 2011; 216, 763-70.
28.Belij S, Popov A, Zolotarevski L, et al. Systemic immunomodulatory effects of topical dinitrochlorobenzene (DNCB) in rats. Activity of peripheral blood polymorphonuclear cells. Environ Toxicol Pharmacol, 2012; 33, 168-80.
29.Djokic J, Ninkov M, Popov Aleksandrov A, et al. Toxicity of Vitamin K antagonist warfarin in rats: the impact of genetic background. J Serb Chem Soc, 2013; 78, 381-94.
30.Philippe B, Ibrahim-Granet O, Prevost MC, et al. Killing of

694 Biomed Environ Sci, 2014; 27(9): 684-694
Aspergillus fumigatus by alveolar macrophages is mediated by reactive oxidant intermediates. Infect Immun, 2003; 71, 3034-42.
31.Roilides E, Uhlig K, Venzon D, et al. Enhancement of oxidative response and damage caused by human neutrophils to Aspergillus fumigatus hyphae by granulocyte colony-stimulating factor and γ-interferon. Infect Immun, 1993; 61, 1185-93.
32.Roilides E, Dimitriadou A, Kadiltsoglou I, et al. IL-10 exerts suppressive and enhancing effects on antifungal activity of mononuclear phagocytes against Aspergillus fumigatus. J Immunol, 1997; 158, 322-9.
33.Safdar A. Immunomodulation therapy for invasive aspergillosis: Discussion on myeloid growth factors, recombinant cytokines and antifungal drug immune modulation. Curr Fungal Infect Rep, 2010; 4, 1-7.
34.Booth C. Fungal culture media. In Methods in Microbiology (JR Norris and DW Ribbons, Eds), 1971; pp. 49-94. Academic Press, London & New York.
35.Sheppard DC, Marr KA, Fredricks DN, et al. Comparison of three methodologies for the determination of pulmonary fungal burden in experimental murine aspergillosis. Clin Microbiol Infect, 2006; 12, 376-80.
36.Harbord M, Novelli M, Canas B, et al. Ym1 is a neutrophile granule protein that crystallizes in p47phox-deficient mice. J Biol Chem, 2002; 277, 5468-75.
37.Bain BJ, Lewis SM. Preparation and staining methods for blood and bone marrow films. In: Dacie and Lewis Practical Haematology (Lewis S, Bain B, Bates I, Eds.), 2006; pp. 25-57. Churcuill Livingstone, Edinburg.
38.Shen K, DeLano FA, Zweifach BW, et al. Circulating leukocyte counts, activation and degranulation in Dahl hypertensive rats. Circ Res, 1995; 76, 276-83.
39.Orciuolo E, Stanzani M, Canestraro M, et al. Effects of Aspergillus fumigatus gliotoxin and methylprednisolone on human neutrophils: implications for the pathogenesis of invasive aspergillosis. J Leukoc Biol, 2007; 82, 839-48.
40.Becker MJ, de Marie S, Bakker-Woudenberg IAJM. Clinical signs predicting imminent death in a rat model of invasive pulmonary aspergillosis. In Human Endopints in Animal Experiments for Biomedical Research (Hendriksen CFM and Morton DB, Eds), 1999; pp. 127-132. Royal Society Medicine Press, London.
41.Becker MJ, de Marie S, Fens MH, et al. Pathophysiology of unilateral pulmonary aspergillosis in an experimental rat model. Med Mycol, 2006; 44, 133-9.
42.Becker MJ, de Marie S, Fens MHAM, et al. Enhanced antifungal
efficacy in experimental invasive pulmonary aspergillosis by combination of AmBisome with Fungizone as assessed by several parameters of antifungal response. J Antimicrob Chemother, 2002, 49, 813-20.
43.Land CJ, Sostaric B, Fuchs R, et al. Intratracheal exposure of rats to Aspergillus fumigatus spores isolated from sawmills in Sweden. Appl Environ Microbiol, 1989; 55, 2856-60.
44.Cenci E, Mencacci A, d’Ostiani CF, et al. Cytokine-and T-helper-dependent immunity in murine aspergillosis. Res Immunol, 1998; 149, 445-54.
45.Balloy V, Chignard M. The innate immune response to Aspergillus fumigatus. Microb Infect, 2009; 11, 919-27.
46.Gerson SL, Talbot GH, Hurwitz S, et al. Prolonged granulocytopenia: the major risk factor for invasive pulmonary aspergillosis in patients with acute leukemia. Ann Intern Med, 1984; 100, 345-51.
47.DiPietro LA. Wound healing: the role of the macrophage and other immune cells. Shock, 1995; 4, 233-40.
48.Morrison BE, Park SJ, Mooney JM, et al. Chemokine-mediated recruitment of NK cells is a critical host defense mechanism in invasive aspergillosis. J Clin Invest, 2003; 112, 1862-70.
49.Park SJ, Hughes MA, Burdick M, et al. Early NK cell-derived IFN-γ is essential to hoste defense in neutropenic invasive aspergillosis. J Immunol, 2009; 182, 4306-12.
50.Hebart H, Bollinger C, Fisch P, et al. Analysis of T-cell responses to Aspergillus fumigatus antigens in healthy individuals and patients with hematologic malignancies. Blood, 2002; 100, 4521-8.
51.Hampton MB, Kettle AJ, Winterbourn CC. Inside the neutrophil phagosome: oxidants, myeloperoxidase, and bacterial killing. Blood, 1998; 92, 3007-17.
52.Rodrigues MR, Rodriguez D, Russo M, et al. Macrophage activation includes high intracellular myeloperoxidase activity. Biochem Biophys Res Commun, 2002; 292, 869-73.
53.Arnaout MA. Structure and function of the leukocyte adhesion molecules CD11/CD18. Blood, 1990; 75, 1037-50.
54.Abram CL, Lowell CA. The ins and outs of leukocyte integrin signaling. Annu Rev Immunol, 2009; 27, 339-62.
55.Rivera A, Ro G, van Epps HL, et al. Innate immune activation and CD4+ T cell priming during respiratory fungal infection. Immunity, 2006; 25, 665-75.
56.Elis TN, Beaman BL. Murine polymorphonuclear neutrophils produce interferon-γ in response to pulmonary infection with Nocardia asteroids. J Leukoc Biol, 2002; 72, 373-81.
57.Shao C, Qu J, He L, et al. Transient overexpression of γ interferon promotes Aspergillus clearance in invasive pulmonary aspergillosis. Clin Exp Immunol, 2005; 142, 233-41.