original article narasin, a novel antiviral compound that ... · background: dengue virus (denv) is...
TRANSCRIPT

©2011 International Medical Press 1359-6535 (print) 2040-2058 (online) 1203
Antiviral Therapy 2011; 16:1203–1218 (doi: 10.3851/IMP1884)
Background: Dengue virus (DENV) is a mosquito-borne virus that causes a spectrum of human diseases ranging from mild dengue fever to dengue haemorrhagic fever and dengue shock syndrome in severe cases. Currently, there is no effective antiviral therapy or vaccine against DENV infection.Methods: In order to identify potential antiviral agents against DENV, we performed high-throughput cell-based screening on a highly purified natural products library. Among the screening hits, selected compounds which displayed 50–75% inhibition against DENV2 were vali-dated using secondary assays. Time-of-addition studies, dose-dependent assays, real time quantitative reverse transcriptase (RT)-PCR, Western blot and ultrastructural imaging were conducted in an attempt to elucidate the potential antiviral mechanisms of narasin.Results: In this study, an ionophore, narasin was selected for detailed analysis due to its strong inhibitory profile
against DENV infection with minimal cytotoxicity (50% cytotoxic concentration >1,000 µM). A dose- dependent study revealed narasin to have an 50% inhibitory con-centration of less than 1 µM against all four sero-types of DENV. Time-of-addition studies of narasin-treated, DENV2- infected Huh-7 cells suggested narasin to be involved in inhibiting the post-entry stages of viral replication during DENV infection. Proteomic and ultrastructural analyses revealed the antiviral mechanism of narasin as likely to be associated with the disruption of viral protein synthesis. In addition, quantitative RT-PCR studies showed no differences in viral RNA levels between narasin- treated and control DENV2-infected cells.Conclusions: Narasin was identified and characterized as a novel agent that inhibits DENV replication in vitro through non-cytotoxic mechanisms, thus indicating its potential to be further developed as a therapeutic anti-DENV agent.
Dengue virus (DENV) is a mosquito-borne virus belonging to the genus Flavivirus in the Flaviviridae and can be further classified into four distinct serotypes (DENV1, DENV2, DENV3 and DENV4). Dengue viruses are transmitted to humans through the bites of female mosquitoes of Aedes aegypti, Aedes albopictus or Aedes polynesiensis. Some other medically impor-tant arthropod-borne viruses that belong to the same family include West Nile virus, yellow fever virus and tick-borne encephalitis virus. DENV infection can be asymptomatic or result in dengue fever that is char-acterised by mild symptoms like fever, nausea, myal-gia, arthralgia and rashes. In severe manifestations of DENV infection, the clinical symptoms may include haemorrhage, hypotension and thrombocytopenia in a condition described as dengue haemorrhagic fever
(DHF). DHF may also progress into the potentially fatal dengue shock syndrome (DSS) when systemic fluid leaks into the interstitial spaces and results in shock [1]. DHF and DSS have been associated with sequential infection by different serotypes of DENV.
DENV has grown into a major public health concern in recent years. According to the WHO, approximately 2.5 billion people are at risk of DENV infection in the tropics and subtropics [2]. Mortality caused by severe DENV disease (that is, DHF and DSS) is worrying as all four serotypes of dengue are currently endemic in the Americas and Asia. Despite the growing threat from the spread of DENV, there is still no approved antivi-ral therapy or vaccine for DENV infection. Some stud-ies reported anti-DENV activities in vitro for several compounds such as mycophenolic acid, ribavirin and
Original article
Narasin, a novel antiviral compound that blocks dengue virus protein expression
June Su Yin Low1†, Kan Xing Wu1†, Karen Caiyun Chen1, Mary Mah-Lee Ng1, Justin Jang Hann Chu1*
1Department of Microbiology, Yong Loo Lin School of Medicine, National University Health System, National University of Singapore, Singapore
*Corresponding author e-mail: [email protected]†These authors contributed equally to this work
Introduction
AVT-11-OA-1998_Low.indd 1203 24/11/2011 17:29:20

©2011 International Medical Press1204
JSY Low et al.
castanospermine [3–5]. However, none of these com-pounds have been approved for antiviral treatment of DENV infection.
In the search for inhibitors of DENV infection, an understanding of the different stages of viral replica-tion can be useful for identifying potential targets for the development of antiviral compounds. DENV is a single-stranded, positive-sense, enveloped RNA virus with a viral genome of approximately 11 kb. The RNA genome of DENV encodes for three structural viral proteins, capsid (C), premembrane (prM) and envelope (E) proteins, and seven non-structural proteins, NS1, NS2A, NS2B, NS3, NS4A, NS4B and NS5. Important events in the replication cycle of DENV include the attachment of virus to the host cell, entry into host cell via endocytosis, uncoating of viral genome, translation and replication of viral RNA, virus particle assembly within the endoplasmic reticulum, and the release of infectious virus into the host cell by exocytosis [6].
In this paper, an immunofluorescence cell-based assay was set-up to screen a compound library of highly puri-fied compounds derived from natural products for spe-cific inhibitors of DENV replication. Several ‘hit’ com-pounds that showed significant inhibition of DENV replication were identified. Among these ‘hit’ com-pounds that were identified, narasin, an ionophore, was picked as a promising antiviral candidate for further evaluations due to its strong inhibitory profile against DENV replication and minimal cytotoxicity. Narasin is a polyether antibiotic and antibacterial agent used in the treatment of chicken coccidia and has been proven to be effective against fungi and anaerobic bacteria with minimal cytotoxicity [7]. In this study, we took several approaches to investigate the possible mechanisms of anti-DENV activity of narasin. Results indicated that narasin acts on the post-entry stages of the DENV rep-lication cycle and strongly interferes with the process of viral protein synthesis.
Methods
Cell culture, virus and antibodiesIn this study, three cell lines were used, that is, Huh-7 human hepatoma cells, baby hamster kidney cells (BHK21) and C6/36 mosquito cells isolated from Aedes albopictus embryonic tissue. Huh-7 hepatoma cells were cultured in DMEM medium and BHK21 cells were cultured in RPMI 1640 with 10% fetal calf serum (FCS) at 37°C with 5% CO2, respectively. The C6/36 cells were maintained in L-15 medium contain-ing 10% FCS at 28°C. Human peripheral blood mono-nuclear cells (PBMC) were obtained under the National University of Singapore IRB-approved protocol and prepared using the Ficoll-Paque Plus isolation solu-tion (Amersham Pharmacia Biotech, Uppsala, Sweden).
The purified PBMC were washed three times with phosphate-buffered saline (PBS), and then resuspended in RPMI 1640 (Gibco/BRL; Life Technologies, Carls-bad, CA, USA) containing 25% FCS. The four sero-types of dengue virus (DENV1, Singapore isolate S144; DENV2, New Guinea C strain; DENV3, Singapore isolate Eden 130/05; and DENV4, Singapore isolate S8976) were propagated in C6/36 cells in this study. The BHK21 cell line was used mainly for quantifica-tion of virus titre via plaque assay. The antibodies used in the immunofluorescence assay for DENV detection were the monoclonal mouse anti-DENV type 2 virus (US Biologicals, Salem, MA, USA) targeting the viral envelope (E) protein and the monoclonal mouse anti-DENV virus complex clone D3-2H2-9-21 (Millipore, Billerica, MA, USA) that reacts with all four serotypes of DENV.
Natural product libraryThe 502-compound Natural Product Library (BML-2865, Biomol; Enzo Life Sciences Inc., Farmingdale, NY, USA) was screened for antiviral activity against DENV infection. The 502 compounds include the fol-lowing classes of natural products; terpenoids, pep-tolides, flavones, coumarins, alkaloids, macrolides, isoflavones and synthetic derivatives etc. All these com-pounds are of low molecular weight and known struc-ture. The compounds are highly purified. No extracts or mixtures are included in this library. The library compounds were dissolved in 100% dimethyl sulfoxide (DMSO) to give a final concentration of 10 mM.
Primary screeningHuh-7 hepatoma cells were seeded at 5,000 cells per well in 384-well plates (Corning Inc., Corning, NY, USA) and incubated at 37°C with 5% CO2. The cells were infected with DENV2 at a multiplicity of infection (MOI) of 1 for 1 h at 37°C, 5% CO2 with gentle rocking every 15 min. After 1 h of viral adsorption, the cells were washed twice with PBS to remove the remaining unbound virus. The library compounds were diluted from the 10 mM stock to a final concentration of 10 µM prior to being introduced to the cells. DENV2-infected Huh-7 cells treated with 0.1% DMSO were used as a control. After addition of the compounds, the DENV2-infected cells were incubated for 72 h before processing for immuno-fluorescence assay. The cells were fixed in cold absolute methanol (Sinopharm Chemical, Shanghai, China) for 15 min and were then rehydrated in PBS before incuba-tion with anti-DENV type 2 antibody (US Biologicals) diluted at a concentration of 1:1,000 for 1 h at 37°C. The cells were washed three times with PBS using the EMBLA 384 Cell Plate Washer (Skatron Instruments, Lier, Norway) and incubated for another 1 h at 37°C with the secondary antibody, fluorescein-conjugated
AVT-11-OA-1998_Low.indd 1204 24/11/2011 17:29:20

Antiviral activity of narasin against dengue virus infection
Antiviral Therapy 16.8 1205
goat anti-mouse immunoglobulin G (IgG) (H+L) anti-body (Chemicon) diluted at a concentration of 1:500. DAPI (Invitrogen, Carlsbad, CA, USA) was used to stain cell nuclei for 15 min.
Data acquisitionThe images for the immunofluorescence assay were obtained using an automated Cellomics ArrayScan VTI HCS System and the auto-focusing parameters were preset from Cellomics Arrayscan Instrument using module ‘Target Activation Bio-Application Version 3’. Each well of the 384-plate was captured in four dif-ferent regions via the automated image capturing func-tion. The images were acquired using both DAPI and FITC channel with a resolution of 512×512 pixels. The brightest plane of the image will act as a refer-ence for the image acquisition to differentiate the DAPI and FITC stains above the background staining. Total numbers of cells in the population were determined by DAPI-stained nuclei and the total numbers of DENV-infected cells were determined by the FITC-stained cytoplasm. The average number of DENV-infected cells per well were then calculated and compared between the mock-infected Huh-7 cells in 0.1% DMSO and DENV-infected Huh-7 cells in 0.1% DMSO. Wells that have low total cell counts of less than 500 cells were not included in the data analyses.
Z′ factor, a statistical measurement of the distance between the standard deviations for the signal versus the noise of an assay, was established to measure the robustness of our screening assay. Experiments to deter-mine Z′ factor were conducted in a 384-well plate using 100 wells of DENV2-infected Huh-7 cells treated with 6 µg/ml MPA and 100 wells of DENV2-infected Huh-7 cells treated with 1% DMSO. The Z′ factor was then computed using Equation 1:
1 − (3 × sd DMSO + 3 × sd MPA)/ (mean DMSO - mean MPA) (1)
The data acquired from the primary screen were then analysed to determine the percentage of inhibition of DENV for each compound when compared to the 0.1% DMSO-treated control. The positive hits from the primary screen were defined as compounds displaying 40% or more inhibition of DENV replication relative to 0.1% DMSO-treated controls. Selected putative hits were then evaluated in secondary assays to validate their anti-DENV activities.
Validation of primary screening hitsDose-dependent experiments were performed to validate the antiviral activities of selected hits. Virus- infection was detected through immunofluorescence while infectious virus production was quantified via plaque assay. In brief,
Huh-7 cells were seeded into 96-well plates at a density of 2×105 cells per well. The cells were then infected with DENV2 at MOI of 1 for 1 h (viral adsorption) before the cells were washed with PBS twice to remove residual virus. The cells were then treated with a range of con-centrations of compounds (0.01 µM, 0.1 µM, 1 µM, 5 µM and 10 µM). The cells were incubated at 37°C, 5% CO2, with the compounds for 72 h before being stained for immunofluorescence assay. The immunofluorescence assay was performed in the same way as described above in the section of immunofluorescence assay for DENV primary screening. The supernatant of each well was also harvested for quantitation of infectious virus via plaque assay. Plaque assay was conducted on monolayers of BHK21 cells in 24-well plates. The DENV-infected sam-ples were diluted in ten-fold serial dilution in DMEM with 2% FCS and added in triplicate for each dilution onto 24-well plate. The cells were incubated for 1 h at 37°C with rocking every 15 min. The cells were washed twice and overlaid with 1% carboxymethyl-cellulose in RPMI with 2% FCS after 1 h of adsorption. After 4 days of incubation in a 37°C, 5% CO2, humidified incubator, the cells were fixed and stained with 10% paraformal-dehyde/1% crystal violet (Sigma–Aldrich Chemical, St Louis, MO, USA) solution for plaque visualization and counting. Quantification of virus titre was expressed as plaque forming unit per millilitre (pfu/ml).
Cell viability assayThe cell viability profiles of the selected compounds were determined using the alamarBlue reagent (Invitro-gen) as recommended by the manufacturer’s protocol. Cell viability assay was conducted in a similar way to drug treatment with the same duration except that the cells were not infected over the course of experiments. In brief, Huh-7 cells or PBMC were incubated with the selected compounds at a concentration range of 1 µM to 1,000 µM in DMEM with 2% FCS for 72 h. Alamar-Blue reagent was then added to the cells for 2 h at 37°C and fluorescence measurements were taken using the Infinite™ 200 series microplate reader (Tecan, Man-nerdorf, Switzerland). The measurements were taken at excitation wavelength of 530 nm and emission wave-length of 585 nm. Sodium azide, a selective inhibitor of cytochrome c oxidase activity and apoptosis-inducing agent, was used as a positive control in this study [8]. The measurements from compound-treated cells were compared to cells treated with 0.1% DMSO (negative control) and 1% sodium azide (assay control) to assess for the percentage of cell viability.
Antiviral activity of narasin against other DENV serotypesHuh-7 cells were seeded onto 96-well plates at a den-sity of 2×105 cells per well. The cells were then infected
AVT-11-OA-1998_Low.indd 1205 24/11/2011 17:29:20

JSY Low et al.
©2011 International Medical Press1206
with different DENV serotypes, that is, DENV1, DENV3 and DENV4 at MOI of 1 for 1 h. After 1 h of the virus adsorption step, the cells were washed twice with PBS and treated with a range of concentra-tions of narasin at 0.01 µM, 0.1 µM, 1 µM and 5 µM. After 72 h of incubation at 37°C, 5% CO2, the DENV-infected cells and narasin-treated DENV-infected cells were then processed for immunofluorescence detec-tion using anti-dengue virus complex antibody, clone D3-2H2-9-21 (Chemicon).
Time-of-addition study of narasin on DENV2-infected cellsTime-of-addition study was performed for narasin (Sigma-Aldrich Chemical) on DENV2-infected Huh-7 cells in 96-well plates. For the pre-treatment assay, Huh-7 cells were pre-treated for 2 h with 1 µM nara-sin. The cells were then washed twice with PBS before DENV2 infection at MOI of 1. Cells were maintained in DMEM with 2% FCS for the pre-treatment study until the experimental end point of 72 hours post infec-tion (hpi).
In the post-treatment assay, Huh-7 cells were first exposed to DENV2 for 1 h (viral adsorption) at a MOI of 1. After the 1 h adsorption step, cells were washed twice with PBS to remove residual virus and mainte-nance media of DMEM with 2% FCS was added to the cells. Narasin solution was then added to the mainte-nance media to give a final concentration of 1 µM of narasin at the time-points 0, 2, 4, 8, 12, 24 and 48 hpi. DENV2-infected cells treated with 0.1% DMSO were used as control in these time-of-addition studies.
In the cotreatment assay, viruses were treated with 1 or 10 µM narasin or 0.1% DMSO for 30 min at 37°C before purification via centrifugal filtration in 100,000 molecular weight (MW) centrifugal filter units (Mil-lipore, Darmstadt, Germany) at 4,000 revolutions per minute (rpm) for 15 min at 4°C. The viruses were then resuspended with PBS and filtered through the same fil-ter units. To ensure thorough removal of any residual narasin solution, the purification process was repeated twice before the ‘treated’-viruses were resuspended in suitable amounts of DMEM with 2% FCS for infecting Huh-7 monolayers at a MOI of 1.
Samples and controls for pre-, post- and cotreatment were all processed for immunofluorescence detection of DENV infection at 72 hpi after incubation in a 37°C, 5% CO2, humidified incubator. The supernatants for each sample were also collected for titration of virus pfu using plaque assay.
Viral RNA transfection into Huh-7 cellsViral RNA was first isolated and purified from DENV2 viral supernatants (9×108 pfu) using the PureLink Viral RNA extraction kit (Invitrogen) and eluted in
RNAse-free water. For the transfection of DENV2 viral RNA into cells, Huh-7 cells were first plated (1×104 cells/well) in a 24-well plate, pre-treated or post-treated with 1 µM and 5 µM of narasin at 37°C upon transfec-tion of the DENV2 RNA. DENV2 RNA was prepared according to the manufacturer’s instructions of TransIT RNA transfection kit (Mirus Bio LLC, Madison, WI, USA) and transfected into Huh-7 cells. The cells and supernatants were harvested 72 hpi for viral antigen detection by immunofluorescence assay.
Quantitative RT-PCRFor the quantitation of the positive and negative-sense of DENV2 RNA in Huh-7 cells, a DENV2 RNA stand-ard was prepared according to the previously published protocol by Wang et al. [9]. The DENV2 RNA standard was first obtained from the Vivid Colors™ pcDNA™ 6.2/C-EmGFP-GW/TOPO® Mammalian Expression Vector containing the DENV prM region being cloned into the vector via TA cloning (Invitrogen). Positive-sense RNA was then generated using MAXIscript® In Vitro Transcription Kit (Applied Biosystems, Oslo Area, Norway) via a T7 promoter. The concentration of DENV2 RNA standard was then measured and its molecular weight was determined for calculation of the positive-sense RNA. The samples were prepared by extracting the total RNA from DENV2-infected Huh-7 cells treated with 1 µM narasin at the time-points, 12, 24 and 48 hpi at 72 hpi. Extraction was carried out using Total RNA Mini Kit (Geneaid, Taoyuan, Taiwan). Total RNA extracted from 0.1% DMSO-treated, mock-infected Huh-7 cells and 0.1% DMSO-treated, DENV2-infected Huh-7 cells were used as controls in this study. cDNA copies of the positive and negative-sense DENV2 RNA were generated for all samples with priming at the DENV NS5 gene as described in previously published protocol [10]. The cDNA copies were then assayed by quantitative real-time PCR using the Maxima™ SYBR Green/ROX qPCR Master Mix (2X) kit (Fermentas, Hanover, MD, USA) containing Maxima™ Hot Start Taq DNA Polymerase, dNTP and SYBR green supple-mented with ROX passive reference dye. Reactions were carried out in ABI 7000 Sequence Detection System (Applied Biosystems) with thermal cycling conditions of denaturation at 95°C for 10 min, followed by 40 ampli-fication cycles of 95°C for 15 s each, and 60°C for 60 s for fluorescence measurement during amplification. The copy number of positive-sense and negative-sense DENV2 RNA were then derived from the cycle thresh-old (CT) value of the amplification plot by using the generated DENV2 RNA standard curve as a reference.
SDS-PAGE and western blotHuh-7 cells were infected with DENV2 at MOI of 1 for 72 h. The cells were either treated with DMEM with
AVT-11-OA-1998_Low.indd 1206 24/11/2011 17:29:20

Antiviral activity of narasin against dengue virus infection
Antiviral Therapy 16.8 1207
2% FCS or DMEM with 2% FCS and 1 µM of narasin at 12 h, 24 h and 48 h after DENV infection. Cells were then washed with PBS and cells lysates were collected at day 3 post infection using 2% sodium dodecyl sulfate (SDS). The proteins in the samples were then separated in a 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) at 80 volts for 3 h. PageR-uler™ Prestained Protein Ladder (Fermentas) was used as a reference for the size standard. After 3 h, the gel was removed and equilibrated in Towbin buffer (0.025 M Tris, 0.192 M glycine 20% methanol) for 10 min. The gel was transferred to nitrocellulose membrane using BioRad semi-dry transfer system at 0.3 A for 45 min (Bio-Rad, San Francisco, CA, USA). The membrane was blocked with 5% skim milk overnight at 4°C. The blot was then washed three times with Tris-buffered saline with Tween-20 and incubated for overnight at 37°C with primary monoclonal anti-DENV E (3H5) antibody and polyclonal anti-dengue NS5 antibody at a dilution of 1:250. The blot was then washed with Tris-buffered saline with Tween-20 and probed with second-ary antibody, ImmunoPure goat anti-mouse IgG (H+L) peroxidase (Thermo Scientific, Hudson, NH, USA) to detect for DENV E or ImmunoPure goat anti-rabbit IgG (H+L) peroxidase (Thermo Scientific) to detect for DENV NS5 at a dilution of 1:2,500 for 1 h at 37°C. The blot was then washed three times prior to development. The membrane was developed by Enhanced Chemi-luminescence (ECL) method using Supersignal® West Pico Chemiluminescent Substrate (Thermo Scientific). The membrane was blocked again with 5% skim milk overnight at 4°C for re-probing of β-actin on the same blot as loading control. The blot was incubated with anti-actin IgG fraction of antiserum (Sigma–Aldrich Chemical) for 1 h at room temperature at a dilution of 1:500. This was followed by incubation of secondary antibody of ImmunoPure goat anti-rabbit IgG (H+L) peroxidise antibody (Thermo Scientific) at a dilution of 1:2,500 for 1 h at room temperature. Membrane was then washed and developed accordingly.
Ultrastructural study using transmission electron microscopy Narasin-treated DENV2-infected cells and DENV2-infected cells were washed twice with PBS. The cell monolayers were then fixed with a primary fixative of 2.5% glutaraldehyde (Agar Scientific, Stansted, UK) at 4°C overnight. After primary fixation, the cell monolayers were washed and scraped off from the cell culture flask with a cell scraper. The cells were then post-fixed with 1% osmium tetroxide (Ted Pella, Redding, CA, USA) for 2 h. A few grains of potas-sium ferrocyanide were added to enhance the contrast of the membranous structure within cells. After 2 h, the cell pellets were washed and dehydrated through
a series of concentrations of ethanol in ascending order, that is, 25%, 50%, 75%, 95% and 100%. The dehydration step was enhanced by another two rounds of absolute acetone treatment for 10 min each. The dehydrated cell pellets were then infiltrated with increasing ratio of araldite 502 (Ted Pella, Red-ding, CA, USA) to acetone at increasing temperature before embedded in fresh araldite for 24 h at 60°C. The embedded samples were then trimmed with an ultramicrotome (Reichert-Jung, Buffalo, NY, USA) to the size of approximately 50–70 nm. The cut sections were then placed onto a 200 mesh copper grid before being stained with 2% uranyl acetate and post-fixed with lead citrate. The stained sections were viewed under the transmission electron microscope Philip EM 208 and captured digitally with a dual view digi-tal camera (Gatan, Werrendale, PA, USA).
Results
Establishment of immunofluorescence screening assayIn order to screen for inhibitors of DENV infection using highly purified compounds from the natural product library (Biomol), we established an immu-nofluorescence assay to detect expression of DENV E protein as an indication of successful DENV infection and replication. Mycophenolic acid (MPA) was shown to inhibit DENV viral RNA replication and was selected as the positive control to inhibit DENV repli-cation and to assess on the robustness of the assay [3]. As shown in Figure 1, MPA exhibited dose-dependent inhibition of DENV2 infection when compared to the 0.1% DMSO control. The 50% inhibitory concen-tration (IC50) value of MPA for DENV2 inhibition in Huh-7 cells was also determined to be 0.39 µM. These results are consistent with the reported study on MPA inhibition of DENV infection [4]. Mock-infected cells and isotype control were also included as negative controls to check for possible background and specificity of antibodies (Figure 1). In addition, a Z′ factor of 0.75 with signal-to-background ratio of 8:1 was obtained for our assay format (JSYL et al., data not shown), satisfying the statistical criteria for a robust assay [11]. Thus, the current dengue virus screening assay via immunofluorescence assay method was found to be suitable and sufficiently robust for the high-throughput screening of compound libraries.
Primary screening assay502 highly-purified compounds with known structures were screened and analysed in the primary screen at a concentration of 10 µM via immunofluorescence assay for potential inhibitors against DENV infection. Using the criteria of ≥40% inhibition to identify positive screening hits, a total of 30 compounds were identified
AVT-11-OA-1998_Low.indd 1207 24/11/2011 17:29:21

JSY Low et al.
©2011 International Medical Press1208
as positive hits from our primary screen (Table 1). In order to validate the reliability of our primary screen-ing data, four compounds were selected for validation of anti-DENV activity in secondary assays. These com-pounds are anisomycin, lycorine HCl, strophanthidin acetate and narasin, with chemical structures shown in Figure 2. These compounds were selected to include a mixture of well-known virus inhibitors, for example, anisomycin and lycorine HCl, and potentially novel compounds with no previously reported anti-flaviviral activities for example, narasin.
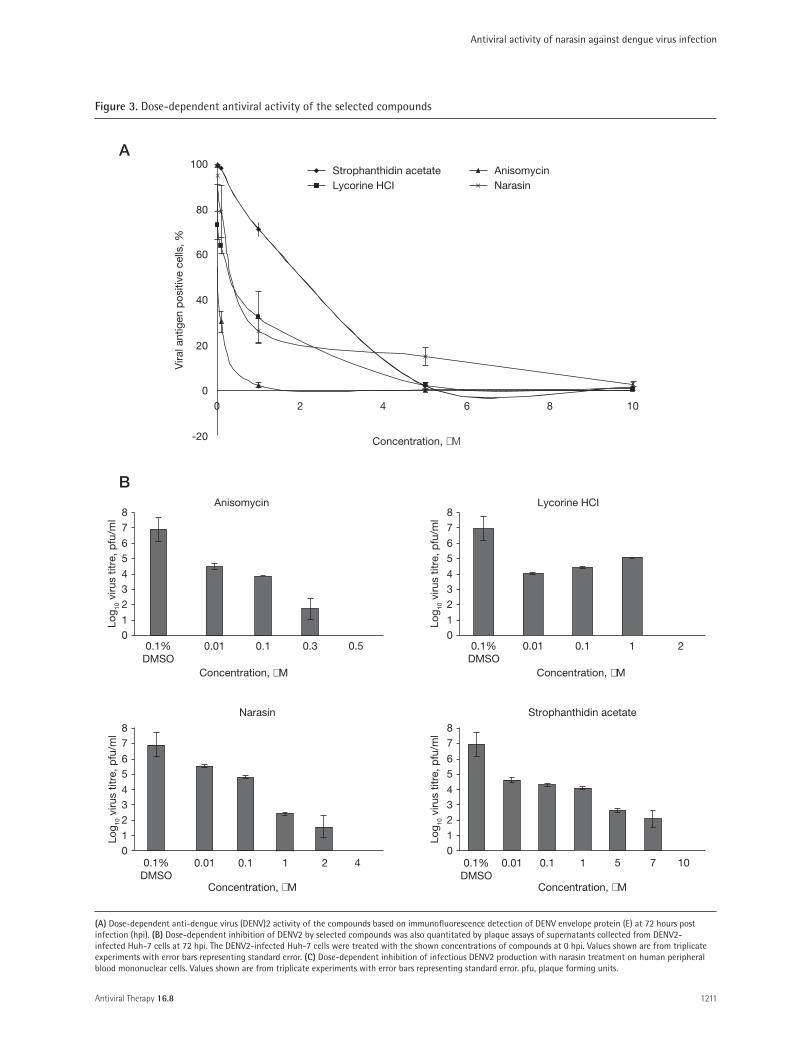
These four compounds were evaluated for dose- dependent anti-DENV activities via immunofluorescence and plaque assays as well as their effects on cellular via-bility. As seen in Figure 3A, all four compounds were val-idated to be true positive hits, displaying dose-dependent inhibition of DENV2 infection with IC50 of 0.05 µM (anisomycin), 0.3 µM (lycorine HCl), 0.39 µM (nara-sin) and 2.0 µM (strophanthidin acetate; Table 2). The reduction in immunofluorescence-detectable DENV2
replication for all 4 compounds were also matched with dose- dependent reductions in infectious virus released from the cells as determined by plaque assays (Figure 3B).
To ensure that the inhibition of DENV replication was not due to the cytotoxic effects from the com-pounds, cell viability assay were conducted using alamarBlue and the 50% cellular cytotoxicity (CC50) of all four compounds were determined (Table 2). From these results, narasin was selected for further investiga-tions into its anti-DENV activity due to its novel status as an antiviral and its relatively high selectivity index (SI) of ≥2,500.
In addition, we have also evaluated the inhibitory effect of narasin on PBMC (mononuclear phagocytic primary cells) that are thought to be the principal den-gue target cells in vivo. PBMC from a healthy donor were infected with the DENV2 at MOI of 1 and treated with a range of concentrations of narasin (0.01 µM, 0.1 µM, 1 µM, 2 µM and 4 µM) for plaque assay quantification at 72 hpi. Similarly, a dose-dependent
100
80
60
40
20
00 2 4 6 8 10
Concentration, µM
Vira
l ant
igen
pos
itive
cel
ls, %
Mock-infected Isotype control 0.1% DMSO+DENV 5 µM MPA+DENV
Figure 1. Development of immunofluorescence assay for primary screening
Immunofluorescence detection of dengue virus (DENV) envelope protein (E) was used as an indication of DENV-infection. DENV2-infection of Huh-7 cells was compared between MPA-treated cells and non-treated cells. Mock-infected cells and isotype control were used as negative controls. Cell nuclei were stained with DAPI (lower intensity) and DENV2 infection was indicated by FITC staining (higher intensity). Mock-infected, isotype control and 5 µM MPA+DENV showed mostly DAPI staining, whereas 0.1% DMSO+DENV showed a mixture of DAPI and FITC staining. Dose-dependent inhibition of DENV2-infection by MPA in Huh-7 cells was also demonstrated with values showing means of triplicate experiments and error bars representing standard errors of the means.
AVT-11-OA-1998_Low.indd 1208 24/11/2011 17:29:21

Antiviral activity of narasin against dengue virus infection
Antiviral Therapy 16.8 1209
inhibition of DENV was observed from PBMC treated with narasin (IC50=0.32 µM; Figure 3C). Minimal cel-lular cytotoxicity was noted for PBMC treated with the different concentrations of narasin in the absence of DENV infection (JSYL et al., data not shown).
Antiviral activity of narasin against other DENV serotypesIn order to determine whether the inhibitory effects of narasin observed against DENV2 was also effective against other serotypes of DENV, Huh-7 cells were infected with DENV serotype 1, 3 and 4 and subjected to narasin treatment at the concentration of 0.01 µM to 5 µM. Dose-dependent inhibition of all the DENV sero-types were observed as shown in Figure 4, suggesting that antiviral activity of narasin was indeed effective against all four serotypes of DENV. The IC50 of narasin for all four DENV serotypes were determined as fol-lows: DENV1: 0.65 µM, DENV2: 0.39 µM, DENV3: 0.44 µM and DENV4: 0.05 µM. One-way ANOVA was performed for the IC50 of all four serotypes and no significant difference was found between the IC50 of DENV serotypes (P=0.064).
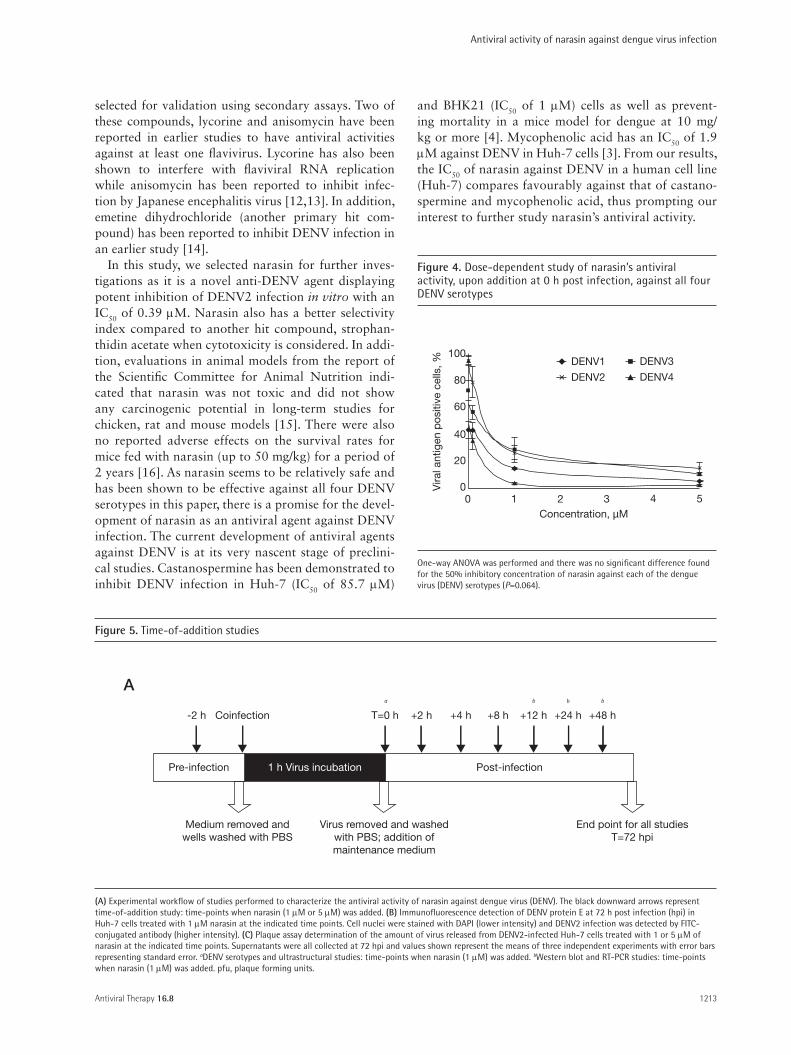
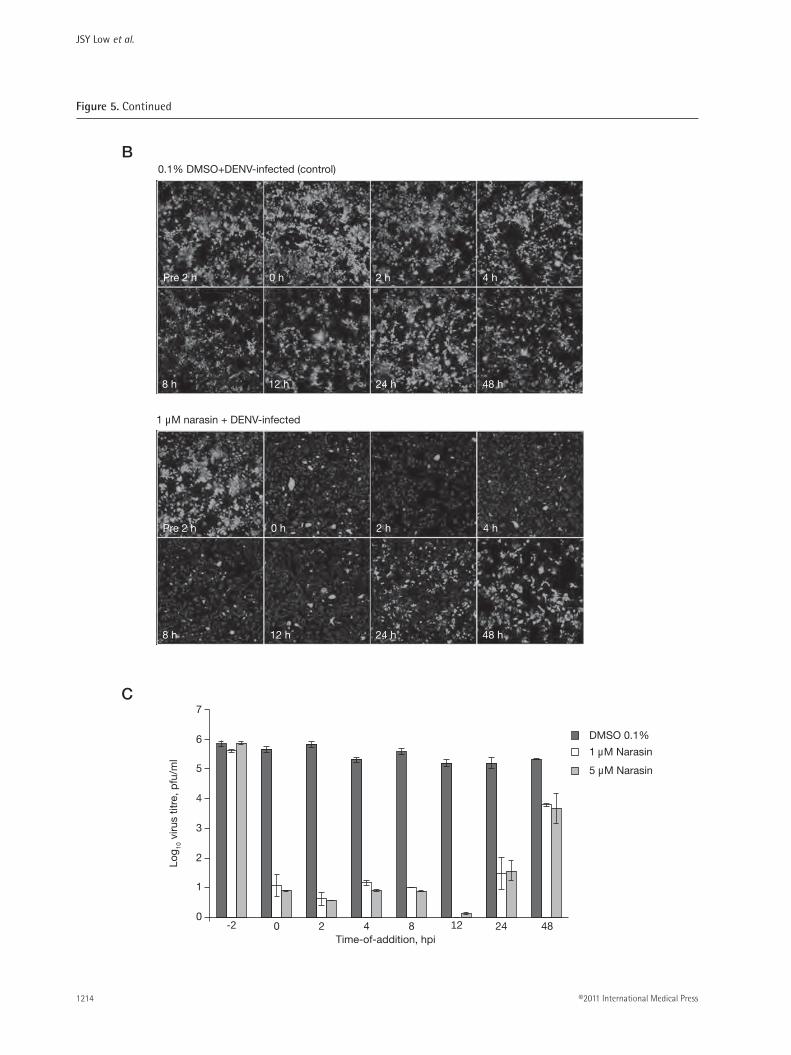
Time-of-addition studies of narasin on DENV infectionTime-of-addition studies were carried out to character-ize the window of narasin’s antiviral activity in relation to the replication cycle of DENV. Narasin (two differ-ent concentrations of 1 µM and 5 µM) was added at the indicated time-points starting from pre-treatment of 2 h, cotreatment to post-treatment of 48 h (Figure 5A). All culture supernatants were collected for infectious virus quantitation using plaque assays and cells were fixed and stained for immunodetection of DENV at 72 hpi. From the images shown in Figure 5B, 0.1% DMSO treatment of cells at any time points did not result in reduction of DENV2 infection. The pre-treatment of cells with 1 µM of narasin for 2 h before exposure of DENV also showed no inhibitory effects against viral infection. In contrast, 1 µM of narasin reduced DENV infection by more than 90% when added from 0 hpi to 12 hpi (Figure 5B). This inhibitory effect was gradually diminished when narasin was introduced later at 24 hpi with only approximately 30% inhibition of DENV replication. This loss of inhibition by narasin was com-plete when narasin was added at 48 hpi, with immuno-fluorescence detection showing similar infection rates as 0.1% DMSO-treated cells. Plaque assay determina-tion of infectious virus titres from each samples also revealed no inhibition when cells were pre-treated with either 1 µM or 5 µM of narasin for 2 h and a similar trend of diminishing inhibition of DENV replication when narasin (1 µM or 5 µM) was added at 24 hpi and beyond (Figure 5C). These experiments were car-ried out with two different concentrations of narasin to
ensure the specificity of narasin in inhibiting the repli-cation of DENV in this time-of-addition study.
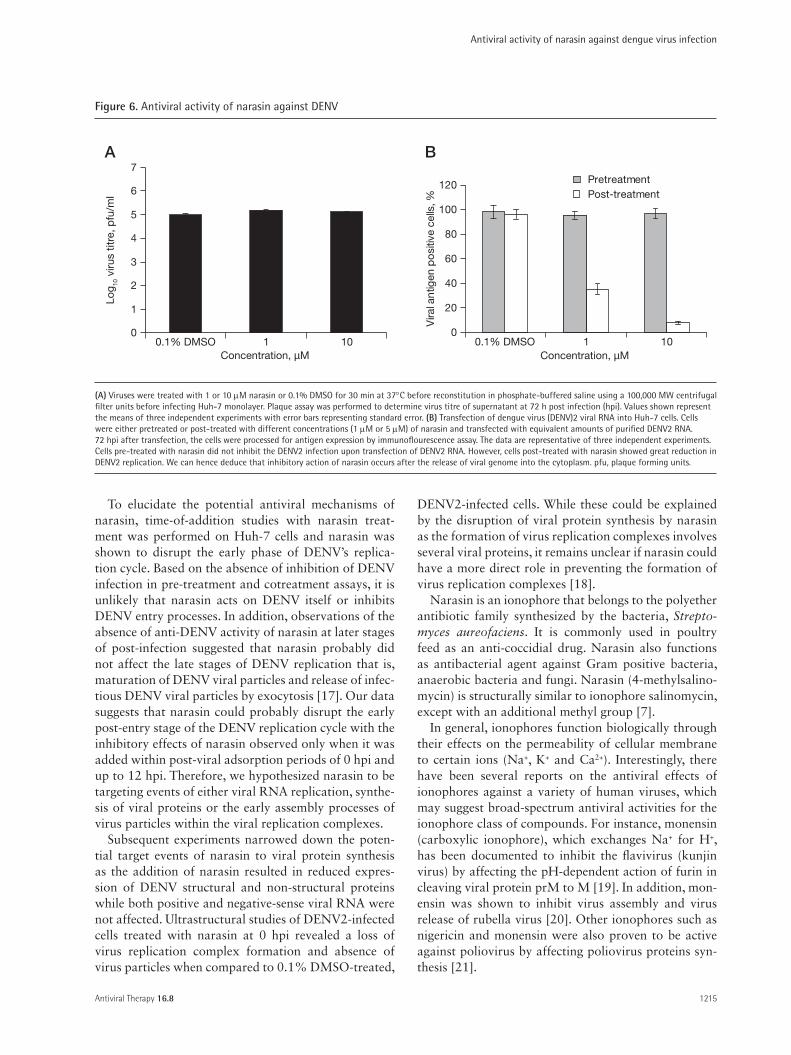
The results from the pretreatment assay of the time-of-addition studies may suggest that narasin is not affecting the early steps (attachment, entry, nucle-ocapsid escape and uncoating) of the DENV replica-tion cycle. To affirm that the early or entry process of DENV replication is not affected by narasin, treat-ment of DENV directly with 1 or 10 µM of narasin in the cotreatment study failed to reproduce any of the inhibitory effects of narasin when it was added after the 1 h virus adsorption period as no plaque reduction was observed relative to 0.1% DMSO-treated viruses (Figure 6A). Furthermore, this was confirmed by uti-lizing an indirect assay whereby naked dengue viral genomic RNA was transfected into Huh-7 cells that were either pre-treated or post-treated with 1 µM and 5 µM of narasin. Pre-treatment of cells with narasin did not inhibit the replication of DENV2 upon trans-fection of the dengue viral RNA as revealed by the sig-nificant levels of viral proteins accumulating within the
Inhibition of dengueCompounds virus infection, %
Lapachone, α 98.84Mycophenolic acid 92.38Lycorine HCl 86.86Ivermectin 84.62C6 Ceramide 84.06Anisomycin 82.49Shikonin 78.94Cycloheximide 78.71Chelerythrine Cl 73.36Skimmianine 61.96Actinomycin D 59.60Narasin 55.89Rapamycin 55.12Tunicamycin B 55.01Acivicin 54.32Cotinine, (-)- 52.71Cantharidin 51.87Epigallocatechin gallate 50.92Gitoxigenin 49.01Kainic acid 48.87Brefeldin A 47.12Dicoumarol 45.99Diacetoxyscirpenol 45.55Austricin 45.01Valinomycin 43.34Thapsigargin 42.90Strophanthidin acetate 41.96Trimethylpsoralen, 4,5’,8- 41.86Echinomycin 41.52Chromomycin A3 40.87
Table 1. Positive ‘hit’ compounds from primary screen, ≥40% inhibition
AVT-11-OA-1998_Low.indd 1209 24/11/2011 17:29:21

JSY Low et al.
©2011 International Medical Press1210
cells (Figure 6B). In contrast, post-treatment of Huh-7 cells with the different concentrations of narasin upon transfection of the purified dengue viral RNA virtu-ally abolished the replication of DENV2 (Figure 6B). Hence, these data may indicate that the antiviral action of narasin may occur at a step after the release of the viral genome into the cytoplasm for virus replication.
Effect of narasin on DENV RNA synthesisTo investigate if narasin affects the early post-entry stages of DENV replication, quantitative RT-PCR was conducted on narasin-treated and untreated DENV2-infected Huh-7 cell to assess the effect of narasin on DENV RNA synthesis. The dengue virus genome con-sists of single positive-sense, single-stranded RNA that is translated to form the viral polyprotein. For the ini-tiation of the viral RNA synthesis, the positive-sense RNA of the dengue genome will serve as template for the synthesis of the complementary, minus-sense RNA genome. Subsequently, the complementary minus-sense
RNA genome will serve as the template strand for the synthesis of more positive-sense viral RNA genomes.
Upon treatment with 1 µM narasin on DENV2- infected cells from 12 h to 48 hpi, the amount of positive-sense and negative-sense DENV RNA were noted to be relatively similar to that of the non-treated DENV2-infected cells (Figure 7A). cDNA from mock-infected cells and non-template control were used as controls in this study and only minimal signals were observed. As such, these data suggested that narasin did not affect DENV RNA replication.
Narasin inhibits DENV viral protein synthesisWestern blot analyses were performed to investigate whether narasin affects the expression of newly synthe-sized DENV viral protein by 72 hpi. From Figure 7B, viral structural protein of DENV E (size ≈54 kDa) and non-structural protein, DENV NS5 (size ≈104 kDa) were detected strongly in DENV-infected cells in the absence of narasin treatment. In contrast, these viral
N
O
O
HOO
O N
OH
H
HO
H
O
O
OH
O
OH
H
HO O
Anisomycin Lycorine HCI
Strophanthidin acetate
Narasin
O
O
OO
HO O
HOH
HOHO
O
H
OH
Figure 2. Structures of compounds
Anisomycin, lycorine HCl, narasin and strophanthidin acetate. These compounds were selected from primary screening hits for further evaluations of dose-dependent antiviral activity and effects on cell viability.
AVT-11-OA-1998_Low.indd 1210 24/11/2011 17:29:22
Antiviral activity of narasin against dengue virus infection
Antiviral Therapy 16.8 1211
100
80
60
40
20
00
-20
8 87
65
432
10
87
65
432
10
87
65
432
10
7
65
432
10
0.1%DMSO
0.5 0.01 0.1 1 2
42 5 7 1010.10.010.1%DMSO
2 4
Concentration, µM
Vira
l ant
igen
pos
itive
cel
ls, %
6 8 10
A
BAnisomycin Lycorine HCI
Log 10
viru
s tit
re, p
fu/m
lLo
g 10 v
irus
titre
, pfu
/ml
Log 10
viru
s tit
re, p
fu/m
lLo
g 10 v
irus
titre
, pfu
/ml
Concentration, µM
Concentration, µMConcentration, µM
Concentration, µM
Narasin Strophanthidin acetate
0.1%DMSO
0.1 1
0.01 0.1
Strophanthidin acetateLycorine HCI
AnisomycinNarasin
0.3
0.1%DMSO
0.01
Figure 3. Dose-dependent antiviral activity of the selected compounds
(A) Dose-dependent anti-dengue virus (DENV)2 activity of the compounds based on immunofluorescence detection of DENV envelope protein (E) at 72 hours post infection (hpi). (B) Dose-dependent inhibition of DENV2 by selected compounds was also quantitated by plaque assays of supernatants collected from DENV2-infected Huh-7 cells at 72 hpi. The DENV2-infected Huh-7 cells were treated with the shown concentrations of compounds at 0 hpi. Values shown are from triplicate experiments with error bars representing standard error. (C) Dose-dependent inhibition of infectious DENV2 production with narasin treatment on human peripheral blood mononuclear cells. Values shown are from triplicate experiments with error bars representing standard error. pfu, plaque forming units.
AVT-11-OA-1998_Low.indd 1211 24/11/2011 17:29:23

JSY Low et al.
©2011 International Medical Press1212
proteins were barely detectable in DENV-infected cells when narasin was added at 12 hpi after DENV infec-tion. These data indicated that the addition of narasin at the early time point of 12 hpi resulted in a reduced expression of DENV E and NS5 viral proteins. How-ever, this inhibitory effect by narasin was gradually lost when narasin was added to the cells at the later time-points of 24 hpi and 48 hpi. Narasin added at 48 hpi no longer has any effects on inhibiting the expression of DENV viral proteins. β-actin served as a loading control in the experiment as well as to indicate that narasin did not affect the synthesis and expression of host cellular proteins. These observations reflect the loss of the inhib-itory effects of narasin when it was added at 48 hpi in the time-of-addition studies, suggesting an association between the observed inhibition of DENV replication in the time-of-addition studies and the failure to detect viral proteins in DENV-exposed cells upon early (not later than 12 hpi) post-entry treatment with narasin.
Ultrastructural analyses of DENV-infected and narasin-treated cellsTo visualize the effects of narasin on DENV replica-tion at the ultrastructural level, transmission electron
microscopy was performed on 1 µM narasin-treated DENV-infected cells and 0.1% DMSO treated DENV-infected cells (virus infection - positive control) at 72 hpi. Typical DENV-infected cells exhibited double-layered membranous replication complex (RC) as shown by the arrows (Figure 8A). Large numbers of electron-dense virus particles can also be observed within or in close association with the RCs as indicated by the arrowheads (Figure 8A). In contrast, upon narasin treatment, there was proliferation of ER membrane but minimal accu-mulation of DENV-induced RCs was observed. In addi-tion, there was a distinct absence of virus particles in the narasin-treated cells (Figure 8B). The mock-infected cells that were treated with narasin (negative control; Figure 8C) were observed to exhibit the typical ultras-tructural morphology of healthy, uninfected cells. These observations suggested a lack of adverse effects on cel-lular morphology arising from narasin-treatment.
Discussion
In an attempt to address the need for antiviral therapy in DENV infections, we developed a high-throughput screening platform using immunofluorescence assay to screen for novel inhibitors of DENV from a compound library consisting of highly purified natural products. The assay has been validated statistically and met the criteria for a robust assay, with a Z′ factor of 0.75 and signal-to-background ratio of 8:1 [11]. These data indi-cated the reliability of our screening assay for use in high-throughput screening. The use of high-throughput screening can facilitate the rapid identification of com-pounds with potential antiviral therapy applications as demonstrated by our study.
From our primary screen, we successfully identified at least 30 positive hits (Additional file 1). Notably, mycophenolic acid, which is also one of the 502 com-pounds screened, appeared as one of the top hits. This independent identification of mycophenolic acid as a screening ‘hit’ indicated the reliability of our screening assay in identifying compounds with anti-DENV activ-ity. Of the hit compounds, four compounds, lycorine, anisomycin, strophanthidin acetate and narasin were
0.01 0.1
Concentration, µM
Log 10
viru
s tit
re, p
fu/m
l
1 2 4
7
6
5
4
3
2
1
00.1%
DMSO
C
Figure 3. Continued
Compounds IC50a, µM CC50
b, µM Selectivity index (CC50/IC50)
Anisomycin 0.05 >1,000 >20,000Lycorine HCL 0.3 >1,000 >3,333Narasin 0.39 >1,000 >2,500Strophanthidin acetate 2.0 77.18 38.6
Table 2. Anti-DENV2 activity of selected compounds from primary screening
aInihibitory activity against dengue virus (DENV)2 infection in Huh-7 cells. Values represent the means from 3 independent experiments. Inhibitory activity was determined by comparing the percentage of viral antigen-positive cells in compound-treated DENV2-infected cells to 0.1% DMSO-treated DENV2-infected cells at 72 hours post infection. bCell viability of compound-treated cells was measured in relation to 0.1% DMSO-treated cells using alamarBlue assay after 72 h incubation. CC50, 50% compound concentration; IC50, 50% inhibitory concentration.
AVT-11-OA-1998_Low.indd 1212 24/11/2011 17:29:23
Antiviral activity of narasin against dengue virus infection
Antiviral Therapy 16.8 1213
selected for validation using secondary assays. Two of these compounds, lycorine and anisomycin have been reported in earlier studies to have antiviral activities against at least one flavivirus. Lycorine has also been shown to interfere with flaviviral RNA replication while anisomycin has been reported to inhibit infec-tion by Japanese encephalitis virus [12,13]. In addition, emetine dihydrochloride (another primary hit com-pound) has been reported to inhibit DENV infection in an earlier study [14].
In this study, we selected narasin for further inves-tigations as it is a novel anti-DENV agent displaying potent inhibition of DENV2 infection in vitro with an IC50 of 0.39 µM. Narasin also has a better selectivity index compared to another hit compound, strophan-thidin acetate when cytotoxicity is considered. In addi-tion, evaluations in animal models from the report of the Scientific Committee for Animal Nutrition indi-cated that narasin was not toxic and did not show any carcinogenic potential in long-term studies for chicken, rat and mouse models [15]. There were also no reported adverse effects on the survival rates for mice fed with narasin (up to 50 mg/kg) for a period of 2 years [16]. As narasin seems to be relatively safe and has been shown to be effective against all four DENV serotypes in this paper, there is a promise for the devel-opment of narasin as an antiviral agent against DENV infection. The current development of antiviral agents against DENV is at its very nascent stage of preclini-cal studies. Castanospermine has been demonstrated to inhibit DENV infection in Huh-7 (IC50 of 85.7 µM)
and BHK21 (IC50 of 1 µM) cells as well as prevent-ing mortality in a mice model for dengue at 10 mg/kg or more [4]. Mycophenolic acid has an IC50 of 1.9 µM against DENV in Huh-7 cells [3]. From our results, the IC50 of narasin against DENV in a human cell line (Huh-7) compares favourably against that of castano-spermine and mycophenolic acid, thus prompting our interest to further study narasin’s antiviral activity.
0
20
40
60
80
100
0 1 2 3 4 5Concentration, µM
Vira
l ant
igen
pos
itive
cel
ls, %
DENV4
DENV1 DENV3
DENV2
Figure 4. Dose-dependent study of narasin’s antiviral activity, upon addition at 0 h post infection, against all four DENV serotypes
One-way ANOVA was performed and there was no significant difference found for the 50% inhibitory concentration of narasin against each of the dengue virus (DENV) serotypes (P=0.064).
-2 h Coinfection
1 h Virus incubation
Medium removed andwells washed with PBS
Virus removed and washedwith PBS; addition ofmaintenance medium
End point for all studiesT=72 hpi
T=0 h +2 h +4 h +8 h +12 h +24 h +48 h
a b b b
Post-infectionPre-infection
A
Figure 5. Time-of-addition studies
(A) Experimental workflow of studies performed to characterize the antiviral activity of narasin against dengue virus (DENV). The black downward arrows represent time-of-addition study: time-points when narasin (1 µM or 5 µM) was added. (B) Immunofluorescence detection of DENV protein E at 72 h post infection (hpi) in Huh-7 cells treated with 1 µM narasin at the indicated time points. Cell nuclei were stained with DAPI (lower intensity) and DENV2 infection was detected by FITC-conjugated antibody (higher intensity). (C) Plaque assay determination of the amount of virus released from DENV2-infected Huh-7 cells treated with 1 or 5 µM of narasin at the indicated time points. Supernatants were all collected at 72 hpi and values shown represent the means of three independent experiments with error bars representing standard error. aDENV serotypes and ultrastructural studies: time-points when narasin (1 µM) was added. bWestern blot and RT-PCR studies: time-points when narasin (1 µM) was added. pfu, plaque forming units.
AVT-11-OA-1998_Low.indd 1213 24/11/2011 17:29:24
JSY Low et al.
©2011 International Medical Press1214
B
C
0.1% DMSO+DENV-infected (control)
Pre 2 h 0 h 2 h 4 h
8 h 12 h 24 h 48 h
Pre 2 h 0 h 2 h 4 h
8 h 12 h 24 h 48 h
1 µM narasin + DENV-infected
0
1
2
3
4
5
6
7
-2 0 2 4 8 12 24 48Time-of-addition, hpi
Log 10
viru
s tit
re, p
fu/m
l
DMSO 0.1%
1 µM Narasin
5 µM Narasin
Figure 5. Continued
AVT-11-OA-1998_Low.indd 1214 24/11/2011 17:29:26
Antiviral activity of narasin against dengue virus infection
Antiviral Therapy 16.8 1215
To elucidate the potential antiviral mechanisms of narasin, time-of-addition studies with narasin treat-ment was performed on Huh-7 cells and narasin was shown to disrupt the early phase of DENV’s replica-tion cycle. Based on the absence of inhibition of DENV infection in pre-treatment and cotreatment assays, it is unlikely that narasin acts on DENV itself or inhibits DENV entry processes. In addition, observations of the absence of anti-DENV activity of narasin at later stages of post-infection suggested that narasin probably did not affect the late stages of DENV replication that is, maturation of DENV viral particles and release of infec-tious DENV viral particles by exocytosis [17]. Our data suggests that narasin could probably disrupt the early post-entry stage of the DENV replication cycle with the inhibitory effects of narasin observed only when it was added within post-viral adsorption periods of 0 hpi and up to 12 hpi. Therefore, we hypothesized narasin to be targeting events of either viral RNA replication, synthe-sis of viral proteins or the early assembly processes of virus particles within the viral replication complexes.
Subsequent experiments narrowed down the poten-tial target events of narasin to viral protein synthesis as the addition of narasin resulted in reduced expres-sion of DENV structural and non-structural proteins while both positive and negative-sense viral RNA were not affected. Ultrastructural studies of DENV2-infected cells treated with narasin at 0 hpi revealed a loss of virus replication complex formation and absence of virus particles when compared to 0.1% DMSO-treated,
DENV2-infected cells. While these could be explained by the disruption of viral protein synthesis by narasin as the formation of virus replication complexes involves several viral proteins, it remains unclear if narasin could have a more direct role in preventing the formation of virus replication complexes [18].
Narasin is an ionophore that belongs to the polyether antibiotic family synthesized by the bacteria, Strepto-myces aureofaciens. It is commonly used in poultry feed as an anti-coccidial drug. Narasin also functions as antibacterial agent against Gram positive bacteria, anaerobic bacteria and fungi. Narasin (4-methylsalino-mycin) is structurally similar to ionophore salinomycin, except with an additional methyl group [7].
In general, ionophores function biologically through their effects on the permeability of cellular membrane to certain ions (Na+, K+ and Ca2+). Interestingly, there have been several reports on the antiviral effects of ionophores against a variety of human viruses, which may suggest broad-spectrum antiviral activities for the ionophore class of compounds. For instance, monensin (carboxylic ionophore), which exchanges Na+ for H+, has been documented to inhibit the flavivirus (kunjin virus) by affecting the pH-dependent action of furin in cleaving viral protein prM to M [19]. In addition, mon-ensin was shown to inhibit virus assembly and virus release of rubella virus [20]. Other ionophores such as nigericin and monensin were also proven to be active against poliovirus by affecting poliovirus proteins syn-thesis [21].
Concentration, µM Concentration, µM
A
0
1
2
3
4
5
6
7
1 10
Log 10
viru
s tit
re, p
fu/m
l
B
0
20
40
60
80
100
120
1 10
Vira
l ant
igen
po
sitiv
e ce
lls, %
PretreatmentPost-treatment
0.1% DMSO 0.1% DMSO
Figure 6. Antiviral activity of narasin against DENV
(A) Viruses were treated with 1 or 10 µM narasin or 0.1% DMSO for 30 min at 37°C before reconstitution in phosphate-buffered saline using a 100,000 MW centrifugal filter units before infecting Huh-7 monolayer. Plaque assay was performed to determine virus titre of supernatant at 72 h post infection (hpi). Values shown represent the means of three independent experiments with error bars representing standard error. (B) Transfection of dengue virus (DENV)2 viral RNA into Huh-7 cells. Cells were either pretreated or post-treated with different concentrations (1 µM or 5 µM) of narasin and transfected with equivalent amounts of purified DENV2 RNA. 72 hpi after transfection, the cells were processed for antigen expression by immunoflourescence assay. The data are representative of three independent experiments. Cells pre-treated with narasin did not inhibit the DENV2 infection upon transfection of DENV2 RNA. However, cells post-treated with narasin showed great reduction in DENV2 replication. We can hence deduce that inhibitory action of narasin occurs after the release of viral genome into the cytoplasm. pfu, plaque forming units.
AVT-11-OA-1998_Low.indd 1215 24/11/2011 17:29:26

JSY Low et al.
©2011 International Medical Press1216
While our study indicated a characteristic disruption of viral protein synthesis by narasin without affecting viral RNA replication, more studies will be needed to understand the exact molecular mechanisms of nara-sin in the inhibition of DENV protein synthesis and replication.
The ability of narasin to inhibit DENV replication and reduce the infectious virus titre from DENV- infected cells bears strong relevance to the progression
of severe dengue-induced diseases, that is, DHF/DSS, in human patients. During the initial stages of DENV infection, DENV will infect the Langerhans cells and dendritic cells before spreading to the spleen, liver and kidney tissues and eventually resulting in a systemic infection [22]. Studies on the pathogenesis of DENV leading to DHF have found a strong relationship between the plasma viral load and the severity of dis-ease [23]. Higher DENV viral RNA and plasma viral
Time-of-addition, hpi
1×106
A
1×10
1×103
1×109
12 24 48
Vira
l RN
A, c
opie
s p
er m
illio
n ce
lls
B
Positive-sense viral RNA (0.1% DMSO)
Positive-sense viral RNA (1 µM narasin)
Negative-sense viral RNA (0.1% DMSO)
Negative-sense viral RNA (1 µM narasin)
12 h 24 h 48 h 12 h 24 h 48 h
DENV E (54 kDa)
β-Actin loading control (42 kDa)
DENV NS5 (104 kDa)
DENV-infected 1 µM Narasin +DENV-infected
Figure 7. Effects of narasin on DENV RNA synthesis and viral protein expression
(A) Real time quantitative reverse transcriptase (RT)-PCR assay revealed similar levels of positive and negative-sense dengue virus (DENV) RNA in both narasin-treated and 0.1% DMSO-treated DENV2-infected Huh-7 cells. Narasin was introduced at time points of 12, 24 or 48 h post infection (hpi) before total RNA was extracted for all samples at 72 hpi. The data was obtained from triplicate set of experiments. (B) Treatment of DENV2-infected Huh-7 cells appeared to inhibit the synthesis of viral proteins without affecting host protein synthesis (β-actin). Both viral structural protein (E) and non-structural protein (NS5) were barely detectable in cells treated with 1 µM narasin at 12 hpi in contrast to the strong protein bands observed for the control (0.1% DMSO-treated). 1 µM of narasin or 0.1% DMSO was added at 12, 24 or 48 hpi before all samples were collected at 72 hpi for SDS-PAGE and western blot analysis.
AVT-11-OA-1998_Low.indd 1216 24/11/2011 17:29:27

Antiviral activity of narasin against dengue virus infection
Antiviral Therapy 16.8 1217
load were detected in life-threatening DHF/DSS patients as compared to patients with dengue fever [23,24]. In our study, narasin was shown to be highly effective in reducing infectious virus production by inhibiting DENV replication at the post-entry early phase. This may suggest that the use of narasin as an antiviral drug could possibly reduce DENV viral load in patients if its inhibitory effects can be reproduced in vivo. Such an antiviral treatment may halt the progression into severe DHF/DSS with vascular leakage and thrombocytope-nia by keeping viral load in check. Currently, work is in progress to determine narasin’s efficacy in reducing DENV replication in a murine model.
In summary, narasin with its potent antiviral activ-ity at low concentrations and minimal cytotoxicity, has potential for further development as an antiviral drug against DENV infection.
Acknowledgements
This project was funded by DSTA-DIRP Grant (POD0713895), NMRC-NIG Grant (Project num-ber NMRC/NIG/0012/2007) and NUS ODPRT Fund (R-182-000-165-133).
Disclosure statement
The manuscript has been read and approved by all authors. The authors declare no competing interests.
Additional file
Additional file 1: A complete list of compounds screened with the primary screening assay against dengue virus infection can be found at http://www.intmedpress.com/uploads/documents/AVT-11-OA-1998_Low_Add_file1.pdf
References1. Halstead SB. Antibody, macrophages, dengue virus
infection, shock, and hemorrhage: a pathogenetic cascade. Rev Infect Dis 1989; 11 Suppl 4:S830–S839.
2. World Health Organization. Dengue haemorrhagic fever: early recognition, diagnosis and hospital management – an audiovisual guide for health-care workers responding to outbreaks. Wkly Epidemiol Rec 2006; 81:362–363.
3. Diamond MS, Zachariah M, Harris E. Mycophenolic acid inhibits dengue virus infection by preventing replication of viral RNA. Virology 2002; 304:211–221.
4. Whitby K, Pierson TC, Geiss B, et al. Castanospermine, a potent inhibitor of dengue virus infection in vitro and in vivo. J Virol 2005; 79:8698–8706.
5. Takhampunya R, Ubol S, Houng HS, Cameron CE, Padmanabhan R. Inhibition of dengue virus replication by mycophenolic acid and ribavirin. J Gen Virol 2006; 87:1947–1952.
6. Qi RF, Zhang L, Chi CW. Biological characteristics of dengue virus and potential targets for drug design. Acta Biochim Biophys Sin (Shanghai) 2008; 40:91–101.
A
B
C
NMNu
rERMitoMito
0.5 µM
0.5 µM
0.5 µM
Figure 8. Ultrastructural analysis of narasin-treated, DENV2-infected Huh-7 cells
Transmission electron microscopy was carried out at 72 h post infection. (A) Dengue virus (DENV)2-infected Huh-7 cells treated with 0.1% DMSO as a control. Cells exhibited extensive proliferation of membranous replication complexes (arrows) which are characteristic of flavivirus DENV infection. DENV2 particles (arrowheads) were localized within the endoplasmic reticulum (ER) lumen in close association with the virus replication complexes (arrows). (B) DENV2-infected Huh-7 cells treated with 1 µM narasin. There were minimal accumulations of DENV-induced membranous replication complexes and absence of virus particles in the ER lumen and membranous structures. However, the proliferation of ER membranes (arrows) was observed within the cytoplasm of the cells. (C) Mock-infected Huh-7 cells treated with 1 µM narasin. These cells exhibited typical ultrastructural morphology of mock-infected Huh-7 cells. Mito, mitochondria; NM, nuclear membrane; Nu, nucleus; rER, rough endoplasmic reticulum.
AVT-11-OA-1998_Low.indd 1217 24/11/2011 17:29:29

JSY Low et al.
©2011 International Medical Press1218
7. Berg DH, Hamill RL. The isolation and characterization of narasin, a new polyether antibiotic. J Antibiot (Tokyo) 1978; 31:1–6.
8. Chen SJ, Bradley ME, Lee TC. Chemical hypoxia triggers apoptosis of cultured neonatal rat cardiac myocytes: modulation by calcium-regulated proteases and protein kinases. Mol Cell Biochem 1998; 178:141–149.
9. Wang WK, Sung TL, Tsai YC, Kao CL, Chang SM, King CC. Detection of dengue virus replication in peripheral blood mononuclear cells from dengue virus type 2-infected patients by a reverse transcription-real-time PCR assay. J Clin Microbiol 2002; 40:4472–4478.
10. Richardson J, Molina-Cruz A, Salazar MI, Black W, IV. Quantitative analysis of dengue-2 virus RNA during the extrinsic incubation period in individual Aedes aegypti. Am J Trop Med Hyg 2006; 74:132–141.
11. Zhang JH, Chung TD, Oldenburg KR. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J Biomol Screen 1999; 4:67–73.
12. Zou G, Puig-Basagoiti F, Zhang B, et al. A single-amino acid substitution in West Nile virus 2K peptide between NS4A and NS4B confers resistance to lycorine, a flavivirus inhibitor. Virology 2009; 384:242–252.
13. Chen CJ, Raung SL, Kuo MD, Wang YM. Suppression of Japanese encephalitis virus infection by non-steroidal anti-inflammatory drugs. J Gen Virol 2002; 83:1897–1905.
14. Low SYJ, Chen CK, Wu KX, Ng ML, Chu JHJ. Antiviral activity of emetine dihydrochloride against dengue virus infection. J Antivir Antiretrovir 2009; 1:62–71.
15. (SCAN) ECSCfAN. Report of the Scientific Committee for Animal Nutrition on the use of Narasin + Nicarbazin in feedingstuffs for chickens 1991. (Accessed 10 January 2011.) Available from http://ec.europa.eu/food/fs/sc/oldcomm6/antibiotics/31_en.pdf
16. World Health Organization. Evaluation of certain veterinary drug residues in food. World Health Organ Tech Rep Ser 2009; 954:1–134.
17. Seema, Jain SK. Molecular mechanism of pathogenesis of dengue virus: entry and fusion with target cell. Indian J Clin Biochem 2005; 20:92–103.
18. Miller S, Romero-Brey I, Bartenschlager R. The dengue virus replication complex. In Hanley KA, Weaver SC (Editors). Frontiers in dengue virus research. Norfolk: Caister Academic Press 2010; chapter 3, pp. 35–53.
19. Mackenzie JM, Westaway EG. Assembly and maturation of the flavivirus Kunjin virus appear to occur in the rough endoplasmic reticulum and along the secretory pathway, respectively. J Virol 2001; 75:10787–10799.
20. Qiu Z, Tufaro F, Gillam S. Brefeldin A and monensin arrest cell surface expression of membrane glycoproteins and release of rubella virus. J Gen Virol 1995; 76:855–863.
21. Irurzun A, Sanchez-Palomino S, Novoa I, Carrasco L. Monensin and nigericin prevent the inhibition of host translation by poliovirus, without affecting p220 cleavage. J Virol 1995; 69:7453–7460.
22. Wu SJ, Grouard-Vogel G, Sun W, et al. Human skin Langerhans cells are targets of dengue virus infection. Nat Med 2000; 6:816–820.
23. Wang WK, Chao DY, Kao CL, et al. High levels of plasma dengue viral load during defervescence in patients with dengue hemorrhagic fever: implications for pathogenesis. Virology 2003; 305:330–338.
24. Thomas L, Verlaeten O, Cabie A, et al. Influence of the dengue serotype, previous dengue infection, and plasma viral load on clinical presentation and outcome during a dengue-2 and dengue-4 co-epidemic. Am J Trop Med Hyg 2008; 78:990–998.
Accepted 1 March 2011; published online 16 August 2011
AVT-11-OA-1998_Low.indd 1218 24/11/2011 17:29:29