o'niverslly ofrawafi ubrart...the effect ofthe taro base on the growth of weissella confusa is...
TRANSCRIPT

O'NIVERSllY OFRAWAfI UBRARt
IDENTIFICATION OF SOME MICROORGANISMS ISOLATED
FROM POI AND CHARACTERIZATION OF THE BIOLOGICAL
PROPERTIES OF ONE LACTIC ACID BACTERIA
A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE
UNIVERSITY OF HAWAI'I IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
IN
FOOD SCIENCE
DEC 2003
By
Lijun He
Thesis Committee:
Alvin S. Huang, ChairpersonAurora A. Saulo
C. Alan Titchenal

© Copyright 2003By
Lijun He
111

Abstract
Lactic acid producing microorganisms from poi and the biological
properties of one lactic acid bacteria are studied in this work. Based on
phenotypical analysis, five bacterial strains isolated from fresh poi are identified
as Lactobacillus acidophilus, Lactobacillus delbrueckii, Lactococcus lactis lactis,
Leuconostoc citreum and Leuconostoc lactis individually. The gas-producing
bacterial strain first identified as Lactobacillus acidophilus is finally identified as
Weissella confusa by combining the results of phenotypical analysis and
genotypical analysis. Acid tolerance of Weisselfa confusa is examined by
exposing the bacterial cells to MRS broth at different pH with/without adaptation.
When exposed to acid stress, the bacterial strain can survive at pH 3.70 with a
decreased growth rate. No bacteria are detected at pH 3.0. However, the survival
rate of the bacterial cells at pH 3.0 increases by adapting the cells at pH 4.46.
Results suggest that the bacteria can grow in acidic condition and the acid
tolerance is adaptable. The effect of the taro base on the growth of Weissella
confusa is investigated by examining the growth of the bacteria in four kinds of
taro/sucrose mixture (10% taro, 3% taro, 3% taro plus 1.3% sucrose, 3% taro
plus 3.2 sucrose). All four media supported the fast growth of the bacteria during
the incubation period and the pH of the taro base dropped fast, ranging from 3.39
to 3.73 after three-day incubation. The growth curves of W. confusa in
taro/sucrose base (3% taro plus 1.3%sucrose) and acid/heat treated poi were
established with the bacterial count and the pH of the sample every day. The
v

bacteria grew rapidly in both of the medium. These preliminary results seem to
indicate that a taro-based fermented food is feasibly produced by monoculture
with W. confusa as starter culture. In addition, a different pH change was
observed in these two taro-based media. These results suggest that the
acid/heat treatment and solid content had some effect on the pH change of the
taro medium.
Based on genotypical analysis, the bacterial strain isolated from sour poi
is identified as Lactobacillus p/antarum and the yeast-like microorganism isolated
from sour poi is identified as Candida tropicalis. Candida trapicalis can ferment
glucose, fructose, sucrose, and starch. Their existence may explain the acid
production at the late stage of the fermentation and the consumption of starch.
This work helps to elucidate the fermentation process of poi and improve
the production by identifying pure cultures of starters. In addition, this work
suggests Weisselfa confusa may be a potential bacterial strain in food
fermentation.
vi

Acknowledgements
I would like to thank Dr. Alvin S. Huang for being my advisor and giving
me the opportunity to work on this wonderful project focusing on food
fermentation in poi. I appreciate his valuable guidance and encouragement,
without which I would not have been able to successfully complete my Master of
Science degree.
I am grateful to the rest of my committee members Dr. Aurora A. Saulo
and Dr. C. Alan Titchenal. Their guidance and suggestions have proven
invaluable. I would also like to thank Dr. Halina M. Zaleski and Dr. Michael A.
Dunn for their assistance and advice throughout my study in this department.
I would like to acknowledge the US Army Research Center at Natick for
their generous financial support to this project. I would like to thank Honolulu Poi
Company for providing poi and taro corm in the study. I would like to thank
Silliker, Inc. Research Center for their technical support in the identification of
microorganisms. I also would like to thank all my lab mates for their help and
cooperation during this study.
Finally, my deep gratitude is given to my best friend Lei Zuo who inspires
me to complete my work, and also my family who have given me their
unwavering support and love at all times. I also want to thank all my good friends,
who have kept me calm throughout this project and who are available to help me
whenever I need assistance.
iv

Five bacterial strains from fresh poi are identified as Lactobacillus
acidophilus. Lactobacillus delbrueckii, Lactococcus lactis lactis, Leuconostoc
citreum, and Leuconostoc lactis based on phenotypical analysis. The bacterial
strain first identified as Lactobacillus acidophilus is finally identified as Weissella
confusa by combining the results of phenotypical analysis and genotypical
analysis. Weissella confusa can grow in acidic condition and the acid tolerance is
adaptable. Taro bases are made from taro corm or fresh poi. All the taro bases
support rapid growth of the bacteria, resulting in a rapid decrease in the pH of the
media. Based on genotypical analysis, Lactobacillus plantarum and Candida
tropicalis are isolated and identified from sour poi. This work suggests Weissella
confusa is a major bacterial strain involved in the fermentation of poi and it is
feasible to produce taro-based fermented food with the monoculture of Weissella
confusa.

Table of contents
Acknowledgements iv
Abstract v
List of Tables x
List of Figures xi
List of Appendix xii
Chapter 1: literature Review 1
Food Fermentation 1
Definition " '" " 1
Fermentation methods 1
Benefits 2
Lactic Acid Fermentation 2
Characteristics '" '" 2
Benefits 3
Examples 3
Lactic Acid Bacteria 5
. Characteristics 6
Taxonomy , 7
Identification method...........................•......... , 7
Selection 10
Application , 13
Food fermentation 14
vii

Food preservation 14
Probiotics '" " , 17
Research Objective 18
Chapter 2: Materials and Methods .21
Introduction 21
Preparation of bacteria isolates 21
Phenotypic characterization .22
Genotypic characterization 24
Preparation of taro bases 25
Inoculation and measurement ofthe growth of the bacteria 27
Acid stress test.. 28
Examination on preliminary characteristics of yeast-like colony 29
Chapter 3: Results and Discussion 31
Screening bacterial strains from fresh poi. 31
Phenotypic characterization 31
Genotypic characterization of the first bacterial strains 35
The biological properties of of WeisseJla confusa 39
The effect of pH on the growth of WeisseJla confusa 38
Acid tolerance response .41
Effect of taro based medium on the growth of Weissella
Confusa 44
Growth curves of WeisseJla confusa in taro/sucrose mixture and
acid/heat treated poi. .46
viii

Genotypic characterization of a lactic acid bacterial strain isolated from
sour poL 50
Genotypic characterization of one yeast-like colony isolated from sour
poi 53
Conclusion 55
Appendix: '" 57
Literature cited 69
IX

LIST OF TABLES
Table
1. Fermentation ways of major lactic acid bacteria in
fermented plant products 8
2. Comparison of the biochemical characteristics and
culture identification of the five lactic acid bacteria 32
3. Summary of the results of charbohydrate fermentation of
the five bacteria 34
4. Percent genetic difference between the first bacterial
strain and representative strains 36
5. Fermentation characteristics of the bacteria isolated in this
study compared with representative Weissefla confusa
and Lactobacillus acidophilus ATCC 4356 37
6. Percent genetic difference between the isolated bacterial
strain from sour poi and representative strains 52
7. Percent genetic difference between the yeast-like strain
and representative strains 54
x

LIST OF FIGURES
Figure Page
1. The Effect of pH on the growth of Weissella confusa .40
2. Survival of unadapted and acid-adapted cells of
Weissella confusa strains during pH challenge (pH 3) .43
3. Effect of different taro base on the growth of
Weissella confusa and the pH of the taro base .45
4. Values of pH and logarithm numbers of Weissella confusa
during fermentation in taro base treated with heat.. .47
5. Values of pH and logarithm numbers of Weissella confusa
during fermentation in taro base treated with acid and heat.. ....48
xi

LIST OF APPENDIX
Number
A. The USDA's nutritional makeup of poi. 57
B. The results oOhe first LAB strain isolated from poi. 59
C. The results of the second LAB strain isolated from poi. 61
D. The results of the third LAB strain isolated from poi. 63
E. The results of the fourth LAB strain isolated from poi. 65
F. The results of the fifth LAB strain isolated from poi. 67
xii

CHAPTER 1LITERATURE REVIEW
Food fermentation
Definition
Food fermentation is a process in which raw materials are converted to
fermented foods by the growth and metabolic activities of desirable
microorganisms. Common materials used in fermentation include milk, meat,
fish, vegetables, fruits, cereal grains, seeds, and beans. Generally, fermented
foods are composed of unused components of the raw materials and the
microbial by-products (Ray, 2001a).
Fermentation methods
Foods can be fermented in three different ways: natural fermentation (use
microflora in the raw materials as starter cultures), back slopping (use successful
products as starter cultures), and controlled fermentation (use pure culture as
starter cultures). The disadvantages of natural fermentation and back slopping
include inconsistent characteristics over a long period of time and high chances
of product failure due to the growth of undesirable flora and foodborne diseases
by pathogens. In controlled fermentation, pure cultures of single or mixed strains
or species of microorganisms are used as the starter culture. The products from
controlled fermentation are consistent and predictable. Generally, there is less
chance of product failure and foodborne diseases. However, in some cases, due
1

to the lack of growth of desirable secondary flora, the product may not have
certain delicate flavor characteristics (Ray, 2001 b).
Benefits
Fermentation plays an important role in food processing, including
enrichment of the human dietary through production of different flavors, aromas
and textures in food; preservation of a large amount of food; conversion of food
substrates biologically with vitamins, protein, essential amino acids and essential
fatty acids; detoxification; and low requirement in energy cost (Steinkraus, 2002).
Due to its healthful and natural characteristics, the consumption of
fermented foods has become popular since the 1970's. The fermented foods
available for general consumption include yogurt, buttermilk, fermented
sausages, kefir (a refreshing probiotic cultured-milk beverage), and kumiss (a
Russian and Mongolian fermented milk beverage) (Ray and Daeschel, 1992). My
project will focus on lactic acid fermentations.
Lactic acid fermentation
Characteristics
Lactic acid fermentation is a process in which the fermentable sugars are
converted to lactic acid by microorganisms including Leuconostoc, Lactobacillus,
Lactococcus, Pediocccus, Streptococcus, Bifidobacterium, and so on
(Steinkraus, 2002).
2

Benefits
Lactic acid fermentation probably is the simplest and safest way to
preserve food (Steinkraus, 2002). Fermentation has been widely used by
humans since biblical times. Lactic acid fermented foods enrich the human diet
with a wide variety of flavors, aromas, and textures, and play an important role in
feeding the world's population. Fermentation is also a low-cost and energy
efficient method of food processing (Steinkraus, 1983a).
Lactic acid fermentation is an effective way to preserve food and has a
good safety record. The substrates used in lactic acid fermentation include
vegetables, fruit, milk, meat, cereal, and others. Lactic acid fermented foods are
widely present in different social cultures, such as German sauerkraut (fermented
cabbage), Korean kim-chi (fermented vegetables with a seasoning mixture
mainly consisting of salt, red pepper powder, garlic, ginger, green onion and
radish), Mexican pozol (fermented maize), Nigerian gari (fermented cassava),
and Middle-Eastern yogurt (fermented milk) (Steinkraus, 2002).
Examples
One of the most well known lactic acid fermented foods is yogurt.
Generally, the composition, flavor and texture of fermented products depend
upon the nature of fermenting organisms, the type of substrate and the
manufacturing process. Yogurt is fermented from milk by a mixed starter culture
consisting of Streptococcus thermophilus and Lactobacillus delbrueckii. The two
species grow symbiotically in milk. Streptococcus thermophilus provide an
3

anaerobic condition, formic acid, and CO2 to stimulate the growth of Lactobacillus
delbruekii. The bacteria.metabolize lactose to produce lactic acid. The
fermentation is temperature-controlled so as to reduce the pH to 4.6 or lower,
giving yogurt a sour flavor (Tamime and Deeth, 1980, Ray, 2001c).
Some other lactic acid fermented foods have been studied in detail.
Sourdough bread is an important fermented food from cereal. Lactobacillus
sanfranciscensis, L. reuteri and L. pontis have been isolated from traditional and
modern fermentations of rye and wheat (Vogel et aI., 1999). A classic lactic acid
vegetable fermentation, sauerkraut, is prepared from fresh cabbage that
shredded and mechanically pressed to expel the juice, providing fermentable
sugars and other nutrients for microbial growth. Anaerobic condition is obtained
though an tight packaging. Salt at 2.25% (glmlx100%) is added to stimulate the
growth of certain lactic acid bacteria. Cabbage is fermented by the combination
of Leuconostoc mesenteroides, Lactobacillus brevis and Lactobacillus plantarum
(Ray, 2001d; Ballcock and Azam-AIi, 1998).
The use of combined starters is common in lactic acid fermentation. All
natural fermentation use combined starters. The combined starters can stimulate
the growth of each other and achieve the desired properties quickly. However,
combined starters may Cause problems in managing industrial-scale production,
especially in standardizing the product. Compared with the fermentation with
combined starters, monoculture fermentation has some advantages. Some
research results indicated that monoculture fermentation is cost-effective
(Marshall and Tamine, 1997). Another study showed that a single starter may
4

give the final product a favorable sensory evaluation (Beasley, Tuorila and Saris,
2003). These results suggest that single starter culture is generally a feasible
option for the food industry.
Lactic acid bacteria play an essential role in the preservation and
production of wholesome food. Lactic acid-producing starter cultures have been
isolated and identified iii many traditional fermented foods. For example, lactic
acid bacteria were proved to be the major microorganisms in Mexican pozol
(Ampe et ai, 1999). Lactic acid bacteria have been isolated from the Nigerian
traditional fermented foods such as fufu (fermented yam, plantain, or cassava),
burukutu (fermented sorghum), and ogi-baba (fermented corn or brown millet
grains). Lactic acid bacteria also have been isolated from Congolese retted
cassava and Benin maize sourdough (Sanni at ai, 2002). Lactic acid bacteria are
even the starters of a Ugandan traditional fermented beverage bushera made
from sorghum or millet (Muyanja et ai, 2003).
In order to obtain new starters with beneficial characteristics, intensive
research on the selection and characterization of lactic acid-producing starter
cultures was conducted in the past decades. A brief review of lactic acid bacteria
is presented below.
Lactic acid bacteria
lactic acid bacteria are a diverse group of bacteria united by a
constellation of morphological, metabolic, and physiological characteristics,
5

which produce lactic acid as the major end product during the fermentation of
carbohydrates.
Lactic acid bacteria are widely distributed in nature. The natural reservoir
of lactic acid bacteria is the green plant material. Lactic acid bacteria are
introduced into animals when they consume the green plants. Not only present in
various nutrient-rich food products such as milk, meat and vegetables, these
bacteria also are normal flora of the mouth, intestine, and vagina of mammals
(Salminen and Wright, 1993).
Characteristics
The typical lactic acid bacteria are rods or cocci, short chains or pairs.
They are gram-positive, non-spore forming, catalase-negative, devoid of
cytochromes, facultative anaerobes, nutritionally fastidious, and able to produce
relatively large amounts of lactic acid as the major end product of metabolism
(Salminen and Wright, 1993). There are no functional heme-linked electron
transport systems or cytochromes in the bacteria. Lactic acid bacteria obtain
metabolic energy by substrate-level phosphorylation while oxidizing
carbohydrates and they lack a functional Krebs cycle (Jay, 1996a).
Lactic acid bacteria can be divided into two groups: the homofermenters
and the heterofermenters. Homofermenters produce mainly lactic acid via the
glycolytic (EMP, Embden-Meyerhof pathway) pathway, while heterofermenters
produce lactic acid plus ethanol, acetate and carbon dioxide, via the 6
phosphogluconate/phosphoketolase pathway. The glycolytic pathway is used by
6

all lactic acid bacteria except leuconostocs, group III lactobacilli, oenococci and
weissella (Battcock and Azam-AIi, 1998). Table 1 lists the types of fermentation
produced by major lactic acid bacteria in fermented plant products.
Taxonomv
The classification of lactic acid bacteria is a loosely defined concept.
Generally, lactic acid bacteria include Lactococcus, Leuconostoc, Pediococcus,
streptococcus, Lactobacillus, Enterococcus, Aerococcus, Vagococcus,
Tetragenococcus, Camobacterium and Weissella (Salminen and Wright, 1993;
Jay, 1996a).
Identification methods
Traditional identification methods are based on physiological
characteristics, biochemical characteristics, and some morphology observations.
They are useful and indispensable in leaming the characteristics of the target
bacterium. However, these identification methods are limited in discrimination
and accuracy. Ambiguous results can lead to misidentification.
Many. changes have taken place in the taxonomy of bacteria in the past
decade. Many of the new taxa have been created as a result of the employment
of molecular genetic tools alone, or in combination with some of the more
traditional methods. In addition to the phenotypic characteristics, characteristics
that can be used to identify lactic acid bacteria include DNA homology, base
composition, 23S, 16S, and 58 rRNA sequence similarities, cell wall composition
7
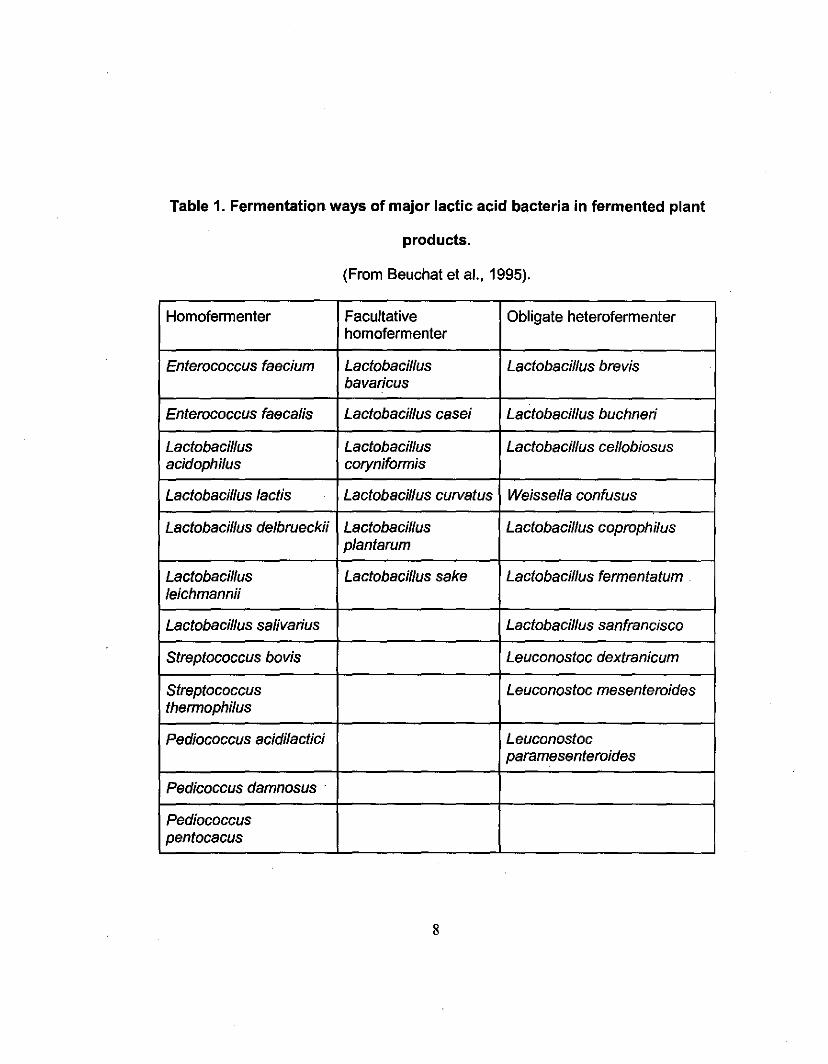
Table 1. Fermentation ways of major lactic acid bacteria in fermented plant
products.
(From Beuchat et aI., 1995).
Homofermenter Facultative Obligate heterofermenterhomofermenter
Enterococcus faecium Lactobacillus Lactobacillus brevisbavaricus
Enterococcus faecalis Lactobacillus casei Lactobacillus buchneri
Lactobacillus Lactobacillus Lactobacillus cellobiosusacidophi/us coryniformis
Lactobacillus lactis Lactobacillus curvatus Weissella confusus
Lactobacillus delbrueckii Lactobacillus Lactobacillus coprophilusplantarum
Lactobacillus Lactobacillus sake Lactobacillus fermentatumleichmannii
Lactobacillus salivarius Lactobacillussanfrancffico
Streptococcus bovis Leuconostoc dextranicum
Streptococcus Leuconostoc mesenteroidesthermophi/us
Pediococcus acidi/actici Leuconostocparamesenteroides
Pedicoccus damnosus .
Pediococcuspentocacus
8

analysis, whole-cell protein fingerprinting, and fatty acid analysis (Jay, 1996b).
The DNA fingerprint technique is the ideal supplementary information in definitive
identification. The genetic fingerprinting techniques used for typing
microorganisms include PFGE (pulsed field gel electrophoresis), Rep-PCR
(repetitive sequence-based polymerase chain reaction), ribotyping, MLST (multi
locus sequencing typing), minisatellites, DNA chips, and AFLP -an acronym for
selective amplification of restriction enzyme fragments (Pouwels and Simons,
2003).
Taxonomic information can be obtained from RNA sequence. The 16S
rRNA subunit is highly conserved and is considered to be an excellent
chronometer of bacteria over time. By use of reverse transcriptase, 168 rRNA
can be sequenced to produce long stretches of genetic code, which allow for the
determination of precise phylogenetic relationships (Jay, 1996b). In all,
comparisons among 16S rRNA sequences are powerful and accurate techniques
for determining the phylogenetic relationships of microorganisms (Woese, 1987).
Some methods were developed based on PCR (polymerase chain
reaction) with species-specific primers derived from the 16S rRNA gene
sequence. PCR amplifications with group-specific or species-specific 16S rONA
primers and ARDRA (Amplified ribosomal DNA and restriction analysis) method
have been used to identify lactobacilli (Roy et aI., 2001). Papadelli et al. (2003)
develop species-specific PCR and DNA hybridization to rapidly detect and
identify Streptococcus macedonicus. This method further confirmed that S.
9

waius, a novel Streptococcus species, actually is phylogenetically identical to S.
macedonicus.
With the development of taxonomic study at the molecular level,
reclassification and new establishment have occurred. For example, the L.
acidophilus complex has been divided into L. acidophilus, L. crispatus, L.
amy/ovorus, L. gallinarum, L. gasser! and L. johnsonii at the species level
(Fujisawa et aI., 1992). With the use of RAPD (randomly amplified polymorphic
DNA) analyses, L. ga/linarum, the genomic sizes of which were estimated by
PFGE (pulse-field gel electrophoresis), was first introduced into characterized
Lactobacillus acidophilus complex, (Roy et aI., 2000). In 1993. a new lactic acid
producing genus was established as Weissella based on the 165 rRNA
sequence analysis (Collins et aI., 1993). Molecular techniques have affected the
taxonomy of lactic acid bacteria as more identification of lactic acid bacteria is
based on the phenotypic analysis and the analysis of 165 rRNA.
Selection
The usage of commercial starter cultures helps to control the
manufacture of fermented food and obtain the desired identity, purity and quality
in the fermented product. The principles of hazard analysis critical control point
(HACCP) are being used in food production to assure stable high quality
production procedures (European food and feed cultures association.
http://www.effca.com).
10

Starter cultures of fermented foods are obtained by both genetic design
and from screening wild strains in nature. Genetic design can make desired
products. Research related to the genetics of lactic acid bacteria provides the
possibility of constructing new powerful starters. However, the time and cost
spent on obtaining legal approval, and subsequently developing a new market,
limit the application of genetically engineered strains (Egon Bech Hansen, 2002).
As a result, traditional screening is still widely used to obtain commercially
valuable strains for food fermentation.
Isolation and identification are the first steps to selecting wild strains with
potentially commercial value. Widely used methods to study microbial
communities in fermented foods are based on the statistical analysis of
information obtained from isolation and identification (Manolopoulou et ai., 2003).
Some other researchers use denaturing gradient gel electrophoresis (DGGE)
and traditional culture-depending methods for examining the bacterial community
of traditional cassava starch fermentation with a better result (Miambi, et ai.,
2003).
The main concerns in food fermentation are spoilage, foodborne
diseases, attractive flavor, taste and appearance, and health benefit. Lactic acid
bacteria contribute much to flavor production and food preservation. Lactic acid
bacteria have a long history of safe use. Some lactic acid bacteria were proved to
be probiolics with health benefit. Consequently, lactic acid bacteria are one of the
most widely studied starter cultures in the food industry.
11

The usage of bacteriocin-producing strains as starter cultures or
protective co- cultures is one of the possible ways to improve food safety. The
target bacteriocins usually have a wide antimicrobial spectrum. They are heat
stable, pH stable within a wide pH range, and sensitive to proteolytic enzymes.
Some bacteriocin-producing strains have been identified to have potential to
improve the safety of the fermented product and could be potential commercial
strains. These identified bacteria strains include human isolate LactobacuJlus
acidophilus LF221 (Bogovic-Matijasic et aI., 1998; Rogelj et aI., 2002), nisin
producing Lactococcus lactic WNC 20 strain isolated from nham - a traditional
Thai fermented sausage (Noonpakdee et aI., 2003).
The study on the survival ability and mechanisms of starter cultures is
one of the hot topics in the food industry. Starter bacteria are constantly
challenged by environmental stresses. Only the strains that survive against these
stresses can greatly improve the process of food fermentation. Tolerance against
acidic stress is one of the standards to screen useful starter cultures. Lactic acid
bacteria can produce lactic acid during fermentation, which means that they are
frequently challenged by acid stress. Lactic acid bacteria must develop some
mechanisms to survive and exert their effect on food. For example, the lactic acid
bacteria that carry out the malolactic fermentation play an important role in
modern commercial wine production. The bacteria must overcome the stress of
low pH, high ethanol content and the low temperature of the cellars during
fermentation. The acid-resistant mutant Leuconostoc oenos LoV8413 was
isolated and identified, which may be used for development of acid-resistant
12

starter cultures in wine (Drici-Cachon et aI., 1996). Another example of stress
response is related to Lactobacillus acidophilus LA1-1 , which is capable of
displaying adaptive response to stress and was suggested to have commercial
value (Kim et aI., 2001).
Another consideration in screening lactic acid bacterial strains is their
potential to be used as probiotics. Probiotics are viable bacterial cell preparation
or foods containing viable bacterial culture or components of bacterial cells that
have beneficial effects on the health of the host. The widely known
microorganisms used as probiotics include bifidobacterium and lactobacillus (Lee
et aI., 1999). The bacteria which can colonize in the gastrointestinal tract (GI)
and adjust the ecology of GI system can be seen as probiotics. Recent studies
on Lactobacillus acidophilus CRL 639 have shown that the strain is low-pH
inducible acid tolerant and has an antagonistic effect on Helicobacter pylori in
Vitro (Lorca et al.. 2001; Lorca and Font de Valdez, 2001). These studies
suggested the potential usage of this bacteria strain as a probiotic. In conclusion.
bacteriocins, stress tolerance, and probiotics are the properties commonly used
to select commercial bacteria strains.
Application
Lactic acid fermentation is one of the oldest forms of biopreservation.
Lactic acid bacteria are one kind of the starter cultures that are responsible for
the fermentation process. Now lactic acid bacteria have been used extensively in
13

the food industry. Generally, lactic acid bacteria can be used in food
fermentation, food preservation and probiotics.
Food fermentation
Fermentation plays an important role in enriching the diet of humans with
desirable flavor, aroma, and texture. Fermentation process involves the
production of preservative products, such as organic acids, alcohol and carbon
dioxide. Fermentation also is responsible for the unique identity and the sensory
attributes of fermented food. Some desirable products include the flavor
compounds diacetyl, acetaldehyde, as well as compounds which may have
positive health implications such as vitamins, antioxidants and bioactive peptides
(Ross et aI., 2002).
In modern large-scale production, defined lactic acid bacterial strains can
be used to produce desirable foods that have dramatically improved the quality
and consistency of the raw materials. The commonly used lactic acid producing
genera include Lactococcus, Lactobacillus, Leuconostoc, Pediococcus, and
Streptococcus.
Since lactic acid fermentation has been discussed in the former section,
now we will focus on food preservation and probiotics.
Food Preservation
The most ancient lactic acid fermentation probably was the product of
fermented sour milk, in which the low pH inhibited other undesirable
14

microorganisms. Other foods widely preserved by lactic acid fermentation around
the world include fruit, vegetable and vegetable! fish! shrimp mixtures.
Some research suggests that lactic acid bacteria were able to improve
the safety of minimally processed fruit and vegetables (Breidt and Fleming,
1997). Also, some lactic acid bacteria strains were able to preserve the ready-to
use vegetables because of their inhibition against Aeromonas hydrophila, Listeria
monocytogenes, Salmonella typhimurium and Staphylococcus (Vescovo et aI.,
1996). These studies indicate the potential application of lactic acid bacteria as
biopreservatives.
The study on the mechanisms of antimicrobial activity revealed that lactic
acid bacteria preserved foods by the production of organic acids, hydrogen
peroxide, diacetyle, and bacteriocins.
Organic acids can reduce the pH of the environment, in which the growth
of many pathogenic and spoilage microorganisms is inhibited. The antimicrobial
mechanisms include interfering with the maintenance of cell membrane potential,
inhibiting active transport, reducing intracellular pH and inhibiting metabolic
functions (Ross et aI., 2002).
Hydrogen peroxide is produced by many of the lactic acid
microorganisms. This chemical was first used in combination with heat to
produce sterile milk. The antimicrobial effect is based on its reaction with other
components to form inhibitory substances.
Diacetyl (2,3-butanedione, biacetyl) is the compound responsible for the
characteristic aroma and flavor of butter. It is also an antimicrobial compound.
15

The antimicrobial activity depends on dosage. One study suggested that the
inhibitory level for yeasts and gram-negative bacteria is 200 I-l9/ml, while the
inhibitory level for non-lactic gram-positive bacteria is 300 I-l9/ml. The
concentration of less than 350 I-l9!ml can inhibit lactic acid bacteria (Daeschel,
1989; Vandenbergh, 1993).
Bacteriocins are proteins or peptides that are inhibitory to other bacteria,
encompassing metabolic end products, antibiotic-like substances, and
bactericidal proteins. Jack et al. (1995) extended the concept of bacteriocins to
encompass extracellularly released primary or modified products of bacterial
ribosomal synthesis. In all, bacteriocins are heterogeneous antagonists varying in
molecular weight, biochemical properties, sensitive host range, and mode of
action. Many bacteriocins produced by lactic acid bacteria are active against
food-borne pathogens such as Listeria monocytogenes, Clostridium perfringens,
Bacillus cereus, and Staphylococcus aureus. The inhibitory spectrum of
bacteriocins can be narrow or wide (Klaenhammer, 1988). Most lactic acid
bacteria are able to produce bacteriocins. The bacteriocin-producing lactic acid
bacteria include Lactobacillus, Streptococcus, Leuconostocs, Pediococcus, and
others (Klaenhammer, 1988). Recently, more bacteriocin-producing lactic acid
bacterial strains, such as Lactobacillus acidophilus LF221 (Bogovic-Matijasic et
aI., 1998), Lactococcus lactis FS92 (Mao et aI., 2001), Lactococcus lactis NK24
(Lee and Paik, 2001), were isolated and identified.
Since lactic acid bacteria have antibacterial activity and are GRAS
(generally regarded as safe) bacteria, the possible usage of the broad-spectrum
16

bacteriocins from laclic acid bacteria or direct usage of laclic acid bacterial
cultures as natural food preservatives is of increasing interest in the food
industry. Nisin, the well-characterized bacteriocin produced by Lactococcus lactis
lactis, has been accepted and approved in more than 50 countries including the
US and the EC (European Community) countries. Nisin has now been widely
used in the production of cheese, milk, desserts, yogurt, fermented beverages,
meat, fish, and canned foods (Vandenbergh, 1993; Soomro et aI., 2002). With
natural, safe, and antimicrobial characteristics, lactic acid bacteria or some
metabolic end products are good options for food preservation.
Probiotics
Animals, including humans, evolved with the normal flora of
microorganisms that exist in the intestinal tract. The normal flora protects us
against the invasion of other microorganisms. However, this protection can be
destroyed by unbalanced diet, antibiotic therapy and stress (Mattila-Sandholm et
al., 1999).
Probiotics are viable bacterial cell preparations or foods containing viable
bacterial cultures or components of bacterial cells that have beneficial effects on
the health of the host. The most thoroughly studied probiolics are bifidobacterium
and lactobacillus (Lee et aI., 1999).
Probiotics have a number of potential benefits, including antimicrobial
properties. antimutagenic properties, anticarcinogenic properties, immune
system stimulation, reduction in serum cholesterol concentration, reduction in
17

allergy, aid in lactose digestion, and adjuvant function (Tannock, 1999; Shah,
2001).
Probiotics are probably safe even at high dose level. With the injection of
probiotic LAB strains Lactobaillus rhamnosus HN001 (DR20), Lb. acidophifus
HN017, and Bifidobacterium factis HN019 (DR10), no toxic effect was detected in
mice. Consequently, such probiotics are likely to be safe for human use (Zhou et
al.,2000).
The effect of probiotics can be affected by administration. Oral
administration of probiotic Lactobacilli may be therapeutic. One study reported
the ability of Lactobacillus pfantarum 299v to colonize in the intestinal tract of
children with HIV and to elicit specific systemic immune response after oral
supplementation (Cunningham-Rundles et aI., 2000).
In general, studies of probiotics focus on the beneficial effect of probiotics
on human health. Lactobacillus is one of the most Widely studied genera with
beneficial properties. A large number of Lactobacillus strains are classified as
probiotics. One well-known example is Lactobacillus acidophilus NCFM, which
has proved to be probiotics and has industrial applications (Sanders and
Klaenhammer, 2001).
Research objectives
Poi is a traditional staple food in Hawaii. II tastes somewhat like fresh,
pure yogurt if it is left in a cool place for 2-3 days before consumption (Begley et
aI., 1981).
18

Traditionally, the marvelous physical development of the Hawaiian is
attributed to poi (Allen and Allen, 1933). It is an excellent carbohydrate source
with 27 grams of carbohydrates based on 100 grams portion of edible sample
(USDA, 2001). The starch in poi is composed of small granules and is extremely
digestible. In Hawaii, poi commonly was used for infants suffering from
malnutrition and food allergies. The effect of poi on the prevention of allergic
disease in potentially allergic infants has been demonstrated (Roth et aI., 1967).
Some people claimed that poi actually promotes gastrointestinal healing
(http://www.thepoicompany.com). However, there is no peer-reviewed research
to support the gastrointestinal healing function of poi.
The purposes of the study of natural food fermentations are to improve
food safety, improve nutritional value, improve production methods and reduce
production costs (Steinkraus, 1983b). Poi is a naturally fermented food. Its sour
taste results from metabolism of lactic acid bacteria, such as Lactobacillus and
streptococcus (Allen and Allen, 1933; Huang et aI., 1994). However, little is
known about the definite starter cultures of poi and their characteristics.
This study is an attempt to screen potentially industrial strains that can be
used in the fermentation of taro base by investigating the starter cultures of poi
and exploring the characteristics of starter cultures.
The objectives of this study are to
1. Isolate and identify microorganisms in poi.
2. Prepare taro base for inoculation.
19

3. Examine the biological properties of the indigenous bacteria, including
growth in the taro base. and acid tolerance.
The eventual purpose of this project is to screen for bacterial strains and
improve the standardization of poi production and food safety by switching the
natural fermentation process to the controlled inoculation process.
20

CHAPTER 2MATERIALS AND METHODS
Introduction
MRS agar (Man, Rogosa and Sharpe agar) was chosen to isolate lactic
acid bacteria from poi in this study because the agar is a selective medium which
contains growth factors, such as polysorbate, acetate, magnesium, and
manganese, for lactobacillus species (http://www.merck.de). The isolated lactic
acid bacteria were first identified with traditional phenotypic tests, including
morphological observation, biochemical tests, and physiological tests. However,
phenotypic tests alone are insufficient to identify microorganisms accurately. The
sequence of 16S rRNA is one of the most effective indexes to define the
taxonomic status. The genotypic test by comparing 16S rRNA sequence was
introduced to positively identify the microorganisms in this project.
Acid tolerance and growth properties of the fermentation process also
were investigated to examine the possible commercial usage of the identified
bacterial strains in this project.
Preparation of bacterial isolates
Fresh poi samples were collected directly from the processing line of a
poi factory (Honolulu Poi Company, Honolulu, Hawaii) and immediately brought
back to the campus of University of Hawaii at Manoa. The unrefrigerated fresh
poi samples were homogenized and serially diluted into appropriate
concentrations with sterilized water at a ratio of 1:10. The MRS agar (Difco) was
21

used for inoculation. Indigenous lactic acid bacteria in poi were screened with the
spread-plate technique. A diluted sample of 0.1 ml was spread over the surface of
MRS agar with a sterilized, L-shaped bent rod. Lactic acid microorganisms were
then incubated at 300e for 24 hours.
The isolated bacteria were purified by randomly sampling single colonies.
A single bacterial colony was mixed with 1 ml of sterilized water and the resultant
bacterial suspension was streaked on MRS agar plates. The plates were
incubated at 300e for 24 hours. After incubation, the cultures could be used for
further testing, or be shipped, or be maintained at 4°e until next usage. The
bacterial isolates were subcultured once a week following the procedure of
culture transfer techniques. A loopful of culture was inoculated on MRS agar
plate by drawing the culture lightly over the surface in a straight or zigzag line.
The plates were incubated at 300e for 24 hours.
Phenotypic characterization
The bacterial isolates were examined by phenotypic tests, including
morphological observations, Gram-stain, biochemical tests, and physiological
tests.
Macroscopic and microscopic methods were used to observe the
morphological characteristics. The colony morphology was observed with
macroscopic methods. The colony characteristics, including size, shape, color,
surface texture, and elevation, were recorded. The colony was mixed with 0.85%
saline on the slide. The 100x phase objective microscope was used to observe
22

cell morphology. Gram stain was performed with 40x phase objective to focus
and 100x phase oil immersion objective.
Biochemical tests used in this project were the catalase test and the
oxidase test. Catalase is the enzyme that degrades hydrogen peroxide into water
and oxygen. The catalase test was performed with a 2% hydrogen peroxide
solution. The result was recorded as positive if there were bubbles in the mixture.
Oxidase plays an important role in the action of the electron transport system
during aerobic respiration. Oxidase is responsible for the formation of H20 or
H20 2 from oxygen (02), Oxidase reagent, N,N,N,N-Tetramethyl-p
phenylenediamine -2HCL was used in the oxidase test. The isolated
microorganism was mixed with the oxidase reagent. The production of a dark
purple color within 30 seconds was recorded as positive.
API Rapid CH Strip system (API System, Plainfield, NY) was used to
investigate other physiological characteristics of the isolated bacteria. The
characteristics are the fermentability in various carbohydrate sources, the ability
to grow at high temperature, the hemolytic properties, and gas production. The
bacterial suspension was centrifuged at 8,000-10,000rpm for 20 minutes after
being incubated in MRS broth for 2 days at 30°C. The supernatant was discarded
and the pellet was resuspended in 10ml of sterile 0.85% saline. The bacterial
suspension was centrifuged in the same condition as stated above and the
supematant was discarded. The pellet was resuspended in 2ml of sterile 0.85%
saline. A total of one milliliter of this saline solution was divided into three
sections to inoculate MRS broth, MRS agar plate and TSA (Trypticase soy agar)
23

/5% blood agar plates individually. The bacteria-inoculated MRS agar plate was
incubated at 45°C for 48 hours to examine the bacteria's ability to grow at high
temperature. The bacteria-inoculated MRS broth was overlayed with paraffin wax
to test the growth and gas production of the isolated bacteria. The TSAl5% blood
agar plate was used to test the hemolytic properties of the bacteria. Both MRS
broth and TSAl5% blood plate were incubated at 30°C for 48 hours. Another 1ml
of resultant saline solution was used for the CH (carbohydrate) strip test. The test
was performed at 30°C for 4 days. The results were analyzed by the Vitek ATB
Expression System (Version 4.0). The Vitek ATB Expression System was
established based on "Bergey's Manual of Systematic Bacteriology" Volume 2,
1986 (Sneath, 1986).
Genotypic characterization
The genotypic characterization was performed by comparing 16S rRNA
sequence. Genomic DNA was isolated from purified bacterial colonies. The 16S
rRNA gene was amplified by PCR from the DNA sequence. The DNA sequences
corresponding to E. coli positions 005 and 1540, and 005 and 531 were used as
full-length package primer and 500bp package primer individually. Amplification
products were purified with Microcon 100 (Millipore) molecular weight cut-off
membranes. Running a portion of the products on an agarose gel was used to
check the quality and quantity of the purified product. Cycle sequencing of the
168 rRNA amplification products was performed with AmpliTaq FS DNA
polymerase and dRhodamine dye terminators. A 8ephadex G-50 spin column
24

was used to remove excess dye-labeled terminators from the reaction. The
resultant samples were ready for further test.
The samples were electrophoresed on an ABI Prism 377 DNA
Sequencer. Applied Biosystems DNA editing and assembly software were used
to analyze data. Sequence comparisons were carried out with the MicroSeq
software (Applied Biosystems)
The samples could be maintained for further usage after they were
centrifuged, dried under vacuum and frozen at -20°C. Once resuspended in a
solution of formamide/blue dextran/EDTA and denatured, the samples were
ready to follow the procedure stated above for further study.
Preparation of taro bases
The taro base treated with only heat was prepared from taro corms. Taro
corms were taken from Honolulu Poi Company. After being washed and stript
thoroughly, the outside layer of the taro corm was cut off carefully. The inside of
the corm was weighed and mixed with sterilized water at the ratio of 1:1 in the
blender. The taro mixture was sampled to measure the solid concentration. The
procedure used to exam the solid concentration was described above. The rest
of the taro mixture was heated until the boiling point while being stirred
continuously. The taro mixture was cooled to room temperature and used as taro
base.
The taro base was diluted with sterile water at the ratio of 1:2. The diluted
taro base was separated into three groups. Sucrose was supplemented into the
25

taro base at the concentration of 0, 1.3g/100g, and 3.2g/100g individually. The
taro bases were heated until the boiling point while continuously being stirred.
After cooled to room temperature, the resultant taro base was ready for further
usage.
The acid/heat treated taro base was prepared with commercial poi
obtained from the Honolulu Poi Company. The solid concentration was examined
to determine further dilution factors. The obtained fresh poi was sampled and
heated in an oven at 100°C overnight. The solid concentration was obtained by
calculating the weight difference between the samples before and after heating.
The acid treatment was performed with a combination of sorbic acid and lactic
acid. Sorbic acid was added to fresh poi until the final concentration reached 0.1 g
/100g (weight ratio). A lactic acid solution of 85% concentration subsequently
was added to the resultant taro paste until the pH decreased to approximately
3.0. Sterilized water was added into the treated taro paste to reach a final solid
concentration of 15%. After an overnight acidic treatment, the taro paste was
placed into water bath at BOaC for 2 hours and then cooled to room temperature.
A 6N NaOH solution was used to adjust the pH of the taro paste to be higher
than 6,0. The resultant taro paste was used as taro base.
The resultant taro bases were sampled and tested for the viability of
microorganisms with MRS agar and Simplate system. The taro base was
sampled and diluted at the ratio of 1:1O. A sample suspension of 0.1 ml was
inoculated and spread on the MRS agar. After a 24-hour incubation at 37°C, the
colonies were counted. Using the Simplate system, a single vial of Butterfield's
26

Buffer Concentrate was poured into a stomacher bag and 223mI of sterile
distilled water was added. After the bag was agitated for about 30 seconds, the
resultant buffer was mix€d completely with 25mg of sample. The sample
suspension was then added into the vial with a different medium for the
examination of TPC (total plate count), Coliform & E. coli, and Yeast & Mold
individually. The entire contents of a single test medium vial were poured on the
center platform of the base plates. Air bubbles were removed and the excess
medium was poured off. The Simplate used to detect the TPC was kepI in an
inverted position (transparent lid down) in the incubator at 35°C for 24 hours. The
wells that changed color from blue to pink after 24-hour incubation were recorded
as positive. The Simplate to detect Coliforms &E. coli was kept in an inverted
position (transparent lid down) in the incubator at 35°C for 24 hours. The wells
with a color different from the original yellow were recorded as positive in the
total coliform count. The wells with fluorescence under U.V. light were recorded
as positive in the E. coli count. The Simplate used to detect the Yeast &Mold
was kept (transparent lid up) in the incubator at 30°C for 48 hours. The wells with
fluoresce under the U.V. light were recorded as positive.
The taro base with negative results was used as the medium for further
inoculation of the bacteria isolated from poi.
Inoculation and Measurement of the growth of the bacteria
The parameters used to monitor the growth of bacteria were bacterial
counts and the pH change of the taro base. The bacteria used for inoculation
27

were grown for 24 hours on MRS agar. The bacterial cultures were inoculated
into a prepared taro base and the initial concentration of bacteria reached a
range of 104 to 106 CFU/ml. Then the cultures were incubated at room
temperature for a period of 5 to 6 days. Two samples were taken out each day:
one for a pH measure and another for a bacteria count. The pH was measured
directly with Orion 230 Aplus pH meter (Orion Research, MA, USA). The samples
used for bacterial counts were appropriately diluted so that the plate count would
be in the readable range of less than 103 CFU/ml. 0.1 ml of the diluted sample
was spread onto MRS agar. The plates were incubated at 30°C for 24 hours and
the numbers of bacteria colonies were counted.
Acid stress test
The acid stress test was conducted in a series of MRS broth, the pH of
which was adjusted to 6.10, 4.46, 3.70, 3.00, and 2.16 respectively. The pH of
the MRS broth was adjusted with the addition of 6.0N hydrochloric acid solution.
The bacterial culture used in acid test was grown on MRS agar at 30°C for 24
hours. A small aliquot of the prepared culture was added to 50ml MRS broth at
various pH values with an initial concentration of 105 CFU/ml. After incubation at
different acidity levels for 24 hours, the bacterial counts were measured on MRS
agar plates following the procedure described in section of the "Inoculation and
measurement of growth of the bacteria". Based on the results, the sublethal pH
and lethal pH were determined.
28

The acid adaptability of the bacteria was examined by challenging
adapted bacterial cells with the MRS broth at lethal pH. The protocol was as
follows. The bacteria were adapted in the MRS broth at sublethal pH and
incubated at 30°C for 24 hours. The adapted bacterial cells were diluted to
approximate concentration. 0.1 ml of bacteria suspension was inoculated into a
fresh MRS broth at lethal pH. The initial inoculation concentration of the bacteria
in the taro base was at the level of 105 CFU/ml. After a 24-hour incubation, the
sample was diluted appropriately and bacterial colonies were counted. The
survival rate was calculated by NINo (N is the CFU/ml after 24-hour incubation
and No is the CFUlml at zero time). The nonadapted bacteria cells were used in
the control group. The nonadapted bacteria cells were challenged directly with
the MRS broth at lethal pH. The challenging procedure is the same as above.
Examination of preliminary characteristics of a yeast-like colony
A yeast-like colony was first isolated from fresh poi after incubation on
MRS agar at 30°C for 24 hours. The cells were sub-cultured on potato dextrose
agar (PDA) at 25°C for 24 hours. The colony morphology and cell morphology
were examined. The methods have been described in the section on "Phenotypic
characterization" in this study.
Fructose and sucrose fermentation were investigated in this test. The
isolate was inOculated into assimilation tubes containing fructose or sucrose
broth. The culture was incubated at 2S"C for 24 hours.
29

The method used to identify the yeast-like colony has been described in
the section on "Genotypic characterization" in this study.
30

CHAPTER 3RESULTS and DISCUSSION
Screening Bacterial Strains from Fresh Poi
Starter cultures can be obtained by genetically designing new bacterial
strains with useful biological characteristics.or screening from wild strains in
nature. The traditional screening by isolating and identifying microorganisms from
fermented food was used in this study. Several microorganism strains were
isolated and identified from the poi obtained from Honolulu Poi Company.
Phenotypic identification
Phenotypic characterization was employed to offer preliminary
identification of microorganisms. A total of five lactic acid bacteria were identified
from the samples isolated form fresh poi. The five lactic acid bacteria were
Lactobacillus acidophilus, Lactobacillus delbruckii, Lactococcus lactis lactis,
Leuconostoc citreum, and Leuconostoc tactis. The results used for identification
were listed in the Table 2 and Table 3. Table 2 shows the biochemical
characteristics and the culture identifications of the five lactic acid bacteria. All
the cells of the five bacterial strains were cocccobacilli, gram-positive, catalase
negative, and oxidase negative, which all are the typical characteristics of lactic
acid bacteria. In addition, the five bacterial strains have their own unique
characteristics. The bacterial strain identified as Leuconostoc citreum showed no
lysis of blood cells. All the other four bacterial strains could imcompletely lyse
blood cells. The bacterial strain identified as Lactobacillus acidophilus was the
31

Table 2. Comparison of the biochemical characteristics and culture
identification of the five lactic acid bacteria
"+"; Positive; "-"; Negative
Lactobacillus Lactobacillus Lactococeus Leuconostoc Leuconostocacidonhiius delbrueckii lactis lactis citreum lactis
Colony Round, Smooth, Smooth, Smooth, Smooth,morphology raised, shiny, tiny, tiny, pinpoint, pinpoint,
and white translucent, translucent, and andand creamy and creamy translucent translucentwhite white
Cell Cocccobacilli Coccobacilli coccobacil1i coccobacilli coccobacillimornholoov-Gram + + + + +reactionCatalase - - - - -Oxidase - - - - -Hemolysis a a a I'Y a.Growth at + + - + +45°CGas + - - - -
I nroduction
32

only one that produced gas. Except the bacterial strain identified as Lactococcus
lactis lactis, all the other four bacterial strains could grow at 45°C. Table 3
summarizes the carbohydrate fermentability of these five bacteria. All these
species can make use of glucose, fructose, sucrose, D-mannose, and maltose.
None of them could use starch as the carbohydrate source.
Four of the five identified lactic acid bacteria could grow at the
temperature of 45°C, indicating that most of the starter cultures could grow at
relatively high temperature. The existence of Lactococcus lactis lactis, which
cannot grow at 45°C, suggests that the taro corms probably are not heated
evenly during poi production. These results are helpful in understanding the
source of starter cultures in natural fermentation process of poi.
Starch is the major component in taro. Since none of isolated bacteria
were able to consume starch, there must be some other mechanisms in poi that
can degrade starch. All the identified bacterial strains utilize glucose, fructose,
and sucrose as the carbon sources. These results partly explain the fast
consumption of sucrose and the change in content of fructose and glucose in poi
production observed in the previous study (Huang et aI., 1994), suggesting that
these sugars are the major carbon sources for growth. These results give some
clues in choosing useful starter cultures and preparation of taro base in the future
application.
Poi fermentation is a process to produce gas. Out of the five identified
strains, L. acidophilus did produce gas and therefore could be one of the most
prevailing strains in poi.
33

Table 3. Summary of the results of charbohydrate fermentation of the five
bacteria
Lactobacillus Lactobacillus Lactococcus Leuconostoc Leuconostocacidophilus delbrueckii lactis lactis citreum lactis
Control - - - - -N- + - + + -AcetvlalucosamineAmygdalin . - - + -L-Arabinose . - - + -Arbutin - - + + -Cellobiose - - + + -Esculin - - + + -D-Fructose + + + + +Galactose + - + - -B-Gentiobiose - - + + -D-Glucose + + + + +Lactose - - - - +Maltose + + + + +Mannitol - - + - -D-Mannose + + + + +Melibiose - - - - +a-Methyl D- - - - + -olucosideD-Raffinose - - - - +Ribose - - + - -Salicin - - + + -Starch - - - - -Sucrose + + + + +Trehalose - - + + -D-Turanose - - - + +D-Xylose + - + + -
34

Based on the results of phenotypic analysis, the bacterial strain first
identified as Lactobacillus acidophilus was further identified with genotypic
identification.
Genotypic identification of the taraet bacterial strain
Lactobacillus species are commonly present in human fecal samples.
Some lactobacillus species have received considerable attention due to their
putative healthy properties. Lactobacillus acidophilus is one of these well-known
probiotics.
The interesting bacterial strain identified as Lactobacillus acidophilus by
phenotypic characterization was further identified with the genotypic
characterization. Based on the analysis of 16S rRNA, the bacterial strain first
identified as Lactobacillus acidophilus was later identified as Weissella confusa.
The results are present in Table 4, which lists the first ten percentage of genetic
differences in 16S rRNA sequence between the tested bacterial strain and the
representative bacterial strains in the MICROSEQTM database. The percentage
of 16S rRNA sequence difference ranges from 0 to 13.88%. Based on this
comparison, the tested bacterial sample was identified as a 100% match to
Weissella confusa.
In Table 5, the carbohydrate fermentation pattern of the isolated bacterial
strain was compared to that of the Weissella confusa representative strain
(Facklam, et ai, 1989; Olano, et aI., 2001) and that of Lactobacillus acidophilus
35

Table 4. Percent genetic difference between the first bacterial strain and the
representative strains
Percent genetic difference from sample SDecies0.00 Weissella confusa3.02 Weissella viridescens4.09 Weissella minor5.86 Weissella kandleri6.25 Weissella Daramesenteroides8.17 Weissella halotolerans13.10 Lactobacillus vaainalis13.10 Lactobacillus reuteri13.64 Lactobacillus oris13.88 Lactobacillus suebicus
36

Table 5. Fermentation characteristics of the bacteria isolated in this study
compared with representative Weissella confusa and Lactobacillus
acidophilus ATCC 4356
+, growth; ., no growth.
Fermentation Weissella confuse Lactobacillus Our resultcharacteristics (from literature) acidophilus ATCC
4356 "(from literature)Arabinose - N/A -Fructose + N/A +Galactose N/A + +Glucose + N/A +Lactose - + -Maltose + + +Mannitol - . -Rhamnose . - -Salicin + + -Sorbitol N/A - -Sucrose + + +Trehalose - + -
37

ATCC 4356 (Sanders and Klaenhammer, 2001). Based on the comparison, the
carbohydrate fermentation pattems of these three bacteria were similar.
The API rapid CH system relies heavily on carbohydrate fermentation
patterns. The analysis system of the API Rapid CHL Strip system used in this
study was established based on "Bergey's Manual of Systematic Bacteriology"
volume 2, 1986 (Sneath, 1986). However, Weisella were established as one
genus of lactic acid bacteria in 1993 based on the 16S rRNA sequence data
(Collins et ai., 1993). Therefore the database of the analysis system doesn't
contain the genus Weissella.
In addition, the results of the characterization also show that the bacterial
strain isolated from poi is heterofermentative with the gas production. However,
Lactobacillus acidophiJus was homofermenter and WeisseJla confusa was
obligate heterofermenter (Beuchat, L. R., 1995). This comparison further
suggested that the isolated bacteria most possibly are Weissella confusa.
The 16S rRNA subunit is highly conserved and is considered to be an
excellent chronometer of bacteria over time. Therefore the information of 16S
rRNA allows for the determination of precise phynogenetic relationships (Jay,
1996b). Based on all the analysis, the isolated bacterial strain finally is identified
as Weissella confusa.
There have been nine species in the genus Weissella, including
WeisseJla confusa. Weissella haJotoJerans, Weissella hellenica, Weissella
kandJeri, Weisse/Ja minor, Weisse/Ja paramesenteroides, Weisse/Ja
thailandensis. Weissella viridescens, and Weissella cibaria sp. nov (Bjorkroth et
38

aI., 2002). WeisselJa strains are distributed in nature and in a variety of fermented
food. Compared with other WeisseJla species, WeisseJla confusa was more
frequently isolated and identified from fermented food. WeisseJla confusa had
been isolated from boza (Hanicoglu and Karapinar, 1997), som-fak (Paludan
Muller et aI., 1999), tapai (Bjorkroth et aI., 2002), togwa (Mugula et aI., 2003) and
bushera (Muyanja et aI., 2003). Till now, there is no further study on WeisseJla
confusa, such as its growth, its response to acidity, and its significance in
fermented food and human health (Facklam et aI., 1989; Collins et aI., 1993;
Christine et aI., 1999; Arrel et al., 2001, K. Johanna Bjorkroth et ai, 2002;). The
purification and identification of the bacterial strain WeisseJla confusa in this
project provide the possibility to study the biological properties of the bacteria
and its significance in food fermentation.
The Biological Properties of Weissella confusa
The effect of pH on the growth of Weissella confusa
The effect of pH on the growth of WeisseJla confusa was examined in the
MRS broth acidified with 6.0N hydrochloric acid. After being adjusted to a specific
pH, the broth was inoculated by W. confusa at a concentration of 1.8x1 05 CFU/ml
and then incubated at 30°C for 24 hours. Fig. 1 shows the effect of pH on the
growth of W. confusa after incubation. The bacteria grew rapidly at pH 6.10 and
the population increased from 105CFU/ml to 1010 CFU/ml after 24 hours. The
39

Fig. 1. The Effect of pH on the growth of Weissella confusa
12
o~..-6.1 4.46 3.7
pH
40
3 2.16

growth rate decreased when the pH of the broth was either 4.47 or 3.7. At the pH
level of 3.0 or 2.16, the bacterial count dropped to zero after the incubation.
These results indicate that these bacteria would be able to grow in sour poi,
which generally have a pH ranges between 3.9 and 4.5.
Lactic acid bacteria tend to be able to grow in the relatively acidic
condition due to a continuous accumulation of laelic acid produced during
growth. However, there are different lower pH limits for growth because of the
various capability of acid tolerance. For example, the known probiotic genus
Bifidobacterium usually doesn't grow at pH below 4.5 and one lactic acid baeleria
Pediococcus acidilactici can grow at pH 3.8 (Ray, 2000e). In comparison with
these known lactic acid bacteria, the isolated W. confusa is able to grow in a
more acidic media. Although there are a number of recent studies on the
mechanism of acid tolerance response of bacteria, the mechanism of acid
tolerance response of W. confusa has not been studied. The acid adaptability of
W confusa was explored in the next study on its acid tolerance response.
Acid tolerance response
In response to stressful stimulation, many microorganisms develop an
adaptive stress response which increases their ability to survive an extreme
condition, such as a high acid environment (Foster, 1999). Acid tolerance
response (ATR) is the process that a brief exposure of cells to mild acidic
41

environment enables them to survive subsequent exposure to a more acidic
environment (Ray, 2000f).
In this acid tolerance response the pH value of 4.46 was chosen as the
sublethal pH and the pH value of 3.0 was chosen as the lethal pH. Both were
based on the previous results. The sublethal pH is an acidic condition that the
tested cells can grow although the growth rate decreases. The lethal pH is the
one that the tested cells can't grow without adaptation.
The results of the survival rate of Weissella confusa with or without
adaptation are displayed in Fig. 2. With an initial concentration of 5.3 x 105
CFU/ml, the adapted cells had a survival count of 3.7x104• For the control group
without adaptation, the cells have only 10 viable cells detected with ihe initial
concentration of 3.2 x 105 CFU/ml. The survival rate of the isolated bacteria was
increased by adaptation when the cells were exposed to the challenge of pH 3.0.
The results indicated that the bacteria could be adapted by the sublethal
environment and became more acid resistant.
Acid tolerance response has been studied in a number of lactic acid
bacteria. There are different identified mechanisms. Some studies show that the
acid tolerance is consistent with the production of acid shock protein (Drici
Cachon et aI., 1996; Lorca et aI., 2001). Arginine catabolism was proved to
contribute to acid tolerance in some other lactic acid bacteria (Rollan et aI.,
2003).
This is the first exploration about the acid tolerance of Weissellaconfusa.
The result suggests that the Weissella confusa strain may be a valuable starter
42

Fig. 2. Survival of unadapted and acid-adapted cells of Weissella confusa
strains during pH challenge (pH 3.00)
o
1-1-
.flfiii -2>.~
:IIII
'0 -3E
~..~-4
-5nonadapted cells adapted cells
_logarithm of survival rate
43

culture with the characteristics of acid tolerance and may play an important role
in food fermentation.
Effect of taro based medium on the growth of Weissella confusa
The effect of taro medium on the growth of W. confusa was studied.
There are two taro solid content levels (10% and 3%) used for comparison.
Sucrose was used as the complementary component in the 3% solid content taro
paste. The levels of sucrose are 0%, 1.3%, and 3.2% respectively. The cell
population and the change in pH were used to measure the growth of W.
confusa. In Figure 3, the bacterial counts and the pH of the medium after
incubation for three days are compared. With the initial pH of 5.60, the pH of
medium decreased fast and the pH values of the four taro mediums were
practically identical, ranging from 3.39 to 3.73 after the incubation. However, the
pH generally dropped from 6.40 to pH 4.13 after 3 days in a natural fermentation
of poi (Huang et ai., 1994). Also the solid concentration of naturally fermented poi
is around 20%, greater than the 3% and 10% used in this study.
All four media supported the fast growth of the bacteria during the
incubation period. The bacterial counts in the media with the solid concentration
of 10% increased from approximately 1x 105 to above 1x 109• At the solid
concentration of 3%, the medium with the addition of sucrose at the level of
1.3g/1 DOg gave the best result and the bacterial count reached 1x 1011• As
indicated in Huang's study (1994), sucrose in taro may be the preferred
carbohydrate source and therefore a proper amount of sucrose would be needed
44

Fig. 3. Effect of different taro base on the growth of Weissella confusa and
the pH of the taro base
12
10
8
6
4
2
0
v§i"
-tJ0
...~rJ"o
• cell population (Log CFUlg
45
.pH values

in a taro-based medium for a sustainable fermentation. The results of three-day
fermentation seem to indicate that there is an optimal amount of sucrose addition
and 1.3g/100g is the best level among the tested three levels.
Growth curves of Weissella confusa in taro/sucrose mixture and acid/heat
treated poi
The growth curves of W. confusa in taro/sucrose base and acid/heat
treated poi were established. The taro/sucrose mixture with 3% solid content of
taro and 1.3% added sucrose was chosen to test the growth curve of W. confusa.
Fig. 4 exhibits the growth curve of the bacteria along with its pH change in the
taro/sucrose mixture during a five-day period. With an initial concentration of
1.7x1 05 CFUlml, W. confusa grew rapidly in the first three days, increasing to
1011 CFU/ml and the pH dropped to 3.39 at the third day. At the end of the fifth
day, the bacterial number reached to 1010 CFU/ml. Fig. 5 shows the growth curve
of W. confusa and its pH change in the acid/heat treated poi. The initial
concentration of bacterial cells was 9x1 03 CFU/g. The bacterial cells also grew
fast and reached above 1011 CFUlg after three-day incubation. At the same time
the pH level decreased to around 5.25.
The above results are the first attempts to inoculate the lactic acid
bacteria W. confusa in taro/sucrose mixture or acid/heat treated poi. W. confusa
used in the study was isolated from naturally fermented poi. The bacteria grew
vigorously in both taro bases. The relatively high bacterial counts were
maintained for up to 7 days in the acid/heat treated poi. These preliminary results
46

Fig. 4. Values of pH and logarithm numbers of Weissella confusa during
fermentation in taro-base treated with heat
12 6
10 5-m-::::)II.CJ 8 4m0
...I-c 3%0 Q.;;
C'll'5Q.
4 20Q.
a;CJ
2 1
0 0
0 1 2 3 4 5
Days
-logarithm numbers of bacteria count
47

Fig. 5. Values of pH and logarithm numbers of Weissella confusa during
fermentation in taro-base treated with acid and heat
14 8
-!!J 12:J - 7LL(J 10en0 6.J 8-c J:0 a... 6.!! - 5~a.
40a.- - 4CD 2(J
o- - 3
0 1 2 3 4 5 6 7
days
-+-IogarithmnulTbers c:A bacteria count pH
48

seem to indicate that the isolated W. confusa would be a feasible starter culture
in making a taro-based fermented food with controlled fermentation. Although
commercial fermented food production usually uses a combination of more than
one bacterial strain in order to maximize the synergies among the strains,
Marshall and Tamine (1997) pointed out that monoculture is cost-effective in
managing industrial-scale production. In addition, Beasley et al. (2003) have
recently used a monoculture of Lactococcus /actis to produce a fermented
soymilk with promising sensory characteristics. The results here seem to indicate
that the monoculture of W. confusa in a taro base is rather feasible.
The solid content in the acid/heat treated poi was about 15% at the point
of inoculation, while the solid content in the taro/sucrose mixture was about
4.3%. The fact that both media supported the fast growth of W. confusa indicated
that there were sufficient nutrients even in the taro-based medium with low level
of solid content. This suggests that there would be a wide range of acceptable
taro-based medium to be used for fermentation with the lactic acid bacteria. The
flexibility would be important in formulating a taro-based fermented product, in
which taste, mouthfeel, and costs are all factors to be considered.
One noticeable difference between the taro/sucrose mixture and the
acid/heat treated poi was the changes of pH. After three days, the pH was 5.25 in
the acid/heat treated poi and 3.39 in taro/sucrose individually. The pH in the
acid/heat treated poi was much higher than the pH value around 4.10 in most
three-day sour poi products. The fresh poi was sterilized by a combination of acid
and heat treatment. The acidlheat treatment may have some effect on the pH
49

change of the taro medium. In addition, the solid concentration of the acid/heat
poi was 15% comparing with that of 4.3% in the taro/sucrose. The solid content
may also contribute to the buffer capacity of the acid/heat treated poi and explain
the difference between these two media.
Fresh poi is hard to handle. The sticky texture of poi renders poor heat
transfer so that sterilization could not be accomplished by heat alone. Lactic acid
alone is not enough to kill all the microorganisms in the poi. Therefore, fresh poi
was treated by a combination of acid and heat, which indicated a long time of
preparation. By comparison, the preparation of taro/sucrose mixture needed heat
alone and cost shorter time. The fast drop of pH in taro/sucrose mixture helps to
reduce the rate of contamination. The appropriate addition of sucrose would be
beneficial for improving the taste and promoting the growth. Given to all these
factors, the taro/sucrose mixture gives beiter result than acid/heat treated poi.
Some improvement to the taro/sucrose mixture is needed to obtain the desired
final product. Further study should be focused on the optimization of the taro
based medium, based on the growth of the bacteria, pH change, effect in taste,
and cost factors.
Genotypic Characterization of a Lactic Acid Bacterial Strain
Isolated from Sour Poi
Some effort was given to the isolation and identification of
microorganisms from sour poi. One strain of bacteria was isolated from sour poi
50

at pH 3.84. Based on the preliminary observation, the bacteria are single or
paired cocobacilli, gram positive, catalase negative, and oxidase negative. The
colony is flat and translucent.
Table 6 shows the percentage of genetic differences in the 16S rRNA
sequence between the tested bacteria and the first ten representative strains in
the MICROSEQTM database. The percentages of 16S rRNA sequence difference
ranged from 0.27 to 10.96%. Based on this comparison, the tested bacterial
strains showed a 99.73% match to Lactobacillus plantarum. The bacterial strain
was most probably identified as Lactobacillus plantarum.
Lactobacilus plantarum is facultative homofermenter. Its growth will end at
a pH of 3.6 to 4.0 (Sneath et aI., 1986). Therefore, Lactobacilus plantarum
usually dominate at the late stages of fermentation (Oyewole and Odunfa, 1990;
Hounhouigan et aI., 1993). These studies may elucidate the reason why L.
pantarum can be isolated from sour poi.
Lactobacilus plantarum is a valuable inhabitant of the intestinal tract of.
human beings (Lund et aI., 2000). Lactobacilus plantarum is also widely
distributed in the fermenting plant materials, such as most vegetable
fermentations and natural cereal fermentations (Oyewole and Odunfa, 1990;
Olasupo er aI., 1997). Lactobacilus plantarum may be responsible for the acid
production, which inhibited the growth of some undesirable microorganisms. The
existence of Lactobacilus plantarum may suggest part of the healthy benefits of
poi.
51

Table 6 Percent genetic difference between the isolated bacterial strain
from sour poi and representative strains
Percent genetic difference from sample Soecies0.27 Lactobacillus Dlantarum0.36 Lactobacillus Dentosus8.89 Lactobacillus aJimentarius9.36 Lactobacillus collinoides9.51 Lactobacillus brevis10.07 Lactobacillus farciminis10.24 Lactobacillus hi/Clardii10.42 Lactobacillus Darabuchneri10.87 Lactobacillus rhamnosus10.96 Pediococcus Darvulus
52

Genotypic Characterization of One Yeast-like Colony Isolated
from Sour Poi
There was a preliminary study on a yeast-like colony isolated from a sour
poi product. Yeasts have been frequently observed in sour poi. Allen & Allen
(1933) has identified two yeast-like colonies, Mycoderma cerevisiac and Oidium
/actis, as starter cultures of traditional poi. In this study, the cells of the isolated
yeast-like colony are large, round and oval. The colonies on PDA are white,
opaque, round and raised. The yeast-like culture was able to ferment fructose
and sucrose.
Table 7 shows that the percentage of genetic difference in 168 rRNA
sequence between the sample and the first ten representative strains in the
MICR08EQTM database. The percentage of 168 rRNA sequence difference
ranges from 0 to 10.94%. Based on the comparison, the tested samples showed
a 100% match to Candida tropicalis.
Candida tropicalis is found among the normal human mucocutaneous flora
and in nutrient-rich environment. Candida tropicalis is one of the opportunistic
mycoses. It is known to cause infection, especially in immunocompromised or
predisposed patients. It is a major cause of endocarditis among patients with
heart damages or intravenous drug abuse. Also it leads to systemic disease in
patients with cancer, diabetes, or chronic alcoholism (Land & McCracken, 1991).
Generally, Candida tropica/is is gas production positive and acid
production positive (Larose et aI., 1976). However, it doesn't produce acetic acid.
53

Table 7. Percent genetic difference between the yeast-like strain and
representative strains
Percent genetic difference from sample Species
0.00 Candida tropicalis2.81 Candida maltosa4.38 Lodderomvces elonqisporus4.84 Candida lodderae5.94 Candida paraDsiJosis5.84 Candida osornensis7.19 Candida albicans10.00 Candida erqatensis10.66 Candida shehatae shehatae10.94 Candida sake
54

Candida tropicalis can make use of carbohydrates for growth, such as starch,
glucose, galactose, maltose, succinic acid, and others. (Barnett et ai., 2000)
The existence of Candida tropicafis partly explains the usage of starch.
Based on my study, the lactic acid bacteria isolated from poi cannot make use of
starch as a carbon source. However, starch is the major component of taro.
There must be some other microorganisms that degrade starch to provide
enough carbon sources for the growth of species in poi. Candida tropicalis may
be one of the starter cultures which are able to make use of starch.
As Candida tropicalis maybe cause opportunistic mycosis under some
condition, poi should not be suggested as a dietary aid to immunosuppressive
patients or immunodeficient patients.
Finally, the existence of Candida tropicalis shows that the present
processing method of poi potentially could cause some diseases if the conditions
are not well controlled. Based on these results, it is necessary to study the
microflora in poi and improve the safety of poi by changing the production from
natural fermentation to controlled fermentation.
Conclusion
Lactic acid bacteria are responsible for the production of poi. Base on the
phenotypic analysis, the lactic acid bacteria isolated from fresh poi in this study
are Lactobacillus acidophilus. Lactobacillus delbrucki/, Lactococcus factis factis,
Leuconostoc citreum, and Leuconostoc factis. One lactic acid bacteria isolated
55

from sour poi is Lactobacilus plantarum and one yeast-like colony isolated from
poi is Candida tfOpicalis based on genotypic analysis.
The lactic acid bacteria first identified as Lactobacillus acidophilus is
identified as Weissella confusa based on phenotypic analysis and genotypic
analysis. Weissella confusa can grow in the acidic condition and its acid
tolerance is adaptable. Sucrose doesn't inhibit the growth of the bacteria and the
taro/sucrose mixture supports the fast growth of the bacteria. The result suggests
that the bacteria would be a feasible starter culture in making a taro based
fermented food with controlled fermentation and it is feasible to produce taro
based fermented food with the monoculture of W. confusa.
Based on these results, More characterizations of the biological
properties of Weissella confusa are necessary.
56

APPEDIXA
The USDA's nutritional makeup of Poi
NOB No' 11349.Value per
Sample Std.Nutrient Units 100 grams of
edible portionCount Error
Proxlmates·Water g 71.64 2Energy kcal 112 0Energy kj 469 0Protein Il 0.38 2
Total lipid (fat) g 0.14 1Carbohvdrate, by difference g 27.23 0
Fiber, total dietary g 0.4 0Ash g 0.61 1
MineralsCalcium, Ca mg 16 1
Iron, Fe mg 0.88 1Magnesium, Mg mg 24 1Phosphorus, P mg 39 1Potassium, K mg 183 2Sodium, Na mg 12 2
Zinc, Zn mg 0.22 0Copper, Cu mil 0.166 0
Manganese, Mn mg 0.370 0Selenium, Se mcg 0.7 0
VitaminsVitamin C, total ascorbic acid mg 4.0 0
Thiamin mg 0.130 0Riboflavin mg 0.040 0
Niacin mg 1.100 0Pantothenic acid mg 0.293 0
Vitamin B-6 mg 0.273 0Folate, total mcg 21 0
Folic acid mcg 0 0Folate, food mcg 21 0Folate, DFE MC!l-DFE 21 0
Vitamin B-12 mcg 0.00 0Vitamin A, IU IU 20 0Vitamin A, RE mC!l-RE 2 0
57

Vitamin E mg ATE I 0.180 I 0 ILipids
Fatty acids, total saturated g 0.029 04:0 g 0.000 06:0 g 0.000 08:0 g 0.000 010:0 g 0.000 012:0 g 0.000 014:0 g 0.000 016:0 g 0.025 018:0 g 0.004 0
Fatty acids, total monounsaturated g 0.011 016:1 undifferentiated g 0.000 018:1 undifferentiated g 0.011 0
20:1 g 0.000 022: 1 undifferentiated g 0.000 0
Fatty acids, total polyunsaturated g 0.058 018:2 undifferentiated g 0.040 018:3 undifferentiated g 0.018 0
18:4 g 0.000 020:4 undifferentiated l! 0.000 0
20:5 n-3 g 0.000 022:5 n-3 l! 0.000 022:6 n-3 g 0.000 0
Cholesterol mg 0 0USDA Nutrient Database for Standard Reference, Release 14 (July 2001)
58

APPEDIX B
The result of the first LAB strain isolated from poi
CULTURE 10 REPORT
Culture Condition Test ResultDeMan Rogosa Sharpe Colony Morphology Tiny, white, roundAgar 48 hous at 30°C colonies
Cell Morpholoav CoccobacilliCatalase NeaativeOxidase NegativeGram reaction Positive
DeMan Rogosa Sharpe Growth PositiveBroth with agar plug, 4 Gas Production Positivedays at 30°CBlood Agar Plate, 24 Hemolysis Alphahours at 30°CDeMan Rogosa Sharpe Growth at 45°C PositiveAgar, 4 days at 45°C
59

API RAPID CH STRIP RESULTS
Test Result Test ResultControl - Esculin -Glycerol - Salicin -Erythritol - Cellobiose -D-Arabinose - Maltose +L-Arabinose - Lactose -Ribose . Melibiose -D-Xylose + Sucrose +L-Xylose - Trehalose -Adonitol - Inuline -B·Methyl-xyloside - Melezitose -Galactose + D-Raffinose -D·Glucose + Starch -D·Fructose + Glycogen -D-Mannose + Xylitol -L-Sorbose - B-Gentiobiose -Rhamnose - D·Turanose -Dulcitol - D-Lyxose -Inositol . D-Tagatose -Mannitol - D-Fucose -Sorbitol - L-Fucose -a-Methyl D· - D·Arabitol -Mannosidea-Methyl D- - L·Arabitol -glucosideN- + Gluconate -AcetylglucosamineAmygdalin - 2-Keto·gluconate -Arbutin - 5-Keto·gluconate -
IDENTIFICATION
Most probable identification:Lactobacillus acidophilus
60

APPEDIX C
The result of the second LAB strain isolated from poi
CULTURE 10 REPORT
Culture Condition Test ResultDeMan Rogosa Sharpe Colony Morphology Smooth, translucent, tiny,Agar 48 hous at 30°C creamy white colonies
Cell Moroholoav Sinale caired coccobacilliCatalase NegativeOxidase NegativeGram reaction Positive
DeMan Rogosa Sharpe Growth PositiveBroth with agar plug, 4 Gas Production Negativedays at 30°CBlood Agar Plate, 24 Hemolysis Alphahours at 30°CDeMan Rogosa Sharpe Growth at 45°C PositiveAgar, 4 days at 45°C
61

API RAPID CH STRIP RESULTS
Test Result Test ResultControl - Esculin -Glycerol - Salicin - .
Erythritol - Cellobiose -D-Arabinose - Maltose +L-Arabinose - Lactose -Ribose - Melibiose -D-Xylose - Sucrose +L-Xvlose - Trehalose -Adonitol - Inuline -~·Methyl. - Melezitose -xylosideGalactose · D·Raffinose -D·Glucose + Starch -D·Fruciose + Glycogen -D-Mannose + Xylitol -L·Sorbose - B-Gentiobiose -Rhamnose · D·Turanose -Dulcitol - D-Lyxose -Inositol - D·Tagatose -Mannitol - D·Fucose -Sorbitol - L-Fucose -a-Methyl D· · D·Arabitol -Mannosidea-Methyl D· · L·Arabitol -glucosideN- - Gluconate -AcetvlalucosamineAmygdalin - 2·Keto-gluconate -Arbutin - 5-Keto-gluconate -
IDENTIFICATION
Most probable identification:Lactobacillus de/brueckii
62

APPEDIX 0
The result of the third LAB strain isolated from poi
CULTURE 10 REPORT
Culture Condition Test ResultDeMan Rogosa Sharpe Colony Morphology Smooth, translucent, tiny,Agar 48 hous at 30°C creamy white colonies
Cell MoroholoaY Sinale paired coccobacilliCatalase NeaativeOxidase NeaativeGram reaction Positive
DeMan Rogosa Sharpe Growth PositiveBroth with agar plug, 4 Gas Production Negativedays at 30°CBlood Agar Plate, 24 Hemolysis Alphahours at 30°CDeMan Rogosa Sharpe Growth at 45°C NegativeAaar, 4 days at 45°C
63

API RAPID CH STRIP RESULTS
Test Result Test ResultControl - Esculin +Glycerol - Salicin +Erythritol - Cellobiose +D·Arabinose - Maltose +L·Arabinose - Lactose -Ribose + Melibiose -D-Xylose + Sucrose +L·Xvlose - Trehalose +Adonitol - Inuline -B-Methvl-xvloside - Melezitose -Galactose + D-Raffinose -D-Glucose + Starch -D-Fructose + Glvcoaen -D-Mannose + Xylitol -L-Sorbose - 13-Gentiobiose +Rhamnose - D-Turanose -Dulcitol - D-Lyxose -Inositol - D-TaQatose -Mannitol + D-Fucose -Sorbitol - L-Fucose -lX-Methyl D- - D-Arabitol -Mannosidea-Methyl D- - L-Arabitol -glucosideN- + Gluconate -AcetylglucosamlneAmvadalin - 2-Keto-aluconate -Arbutin + 5-Keto-gluconate -
IDENTIFICATION
Most probable identification:Lactococcus lactis lact;s
64

APPEDIX E
The result of the fourth LAB strain isolated from poi
CULTURE 10 REPORT
Culture Condition Test ResultDeMan Rogosa Sharpe Colony Morphology Smooth, pinpoint,Agar 48 hous at 30°C translucent colonies
Cell Morphology Single paired coccobacilliCatalase NeQativeOxidase NeaativeGram reaction Positive
DeMan Rogosa Sharpe Growth PositiveBroth with agar plug, 4 Gas Production Negativedavs at 30°CBlood Agar Plate, 24 Hemolysis Negativehours at 30°CDeMan Rogosa Sharpe Growth at 45°C PositiveAoar, 4 days at 45°C
65

API RAPID CH STRIP RESULTS
Test Result Test ResultControl - Esculin +Glycerol - Salicin +Erythritol - Cellobiose +D-Arabinose - Maltose +L-Arabinose + Lactose -Ribose - Melibiose -D·Xylose + Sucrose +L-Xylose - Trehalose +Adonitol - Inuline -~-Methyl- - Melezitose -xvtosideGalactose - D-Raffinose -D-Glucose + Starch -D-Fructose + Glycogen -D-Mannose + Xvtitol -L-Sorbose - a-Gentiobiose +Rhamnose - D-Turanose +Dulcitol - . D-Lvxose -Inositol - D-Tagatose -Mannitol - D-Fucose -Sorbitol - L-Fucose -a-Methyl 0- - D-Arabitol -Mannosidea-Methyl D- + L-Arabitol -alucosideN- + Gluconate -AcetvlalucosamineAmygdalin + 2-Keto-aluconate -Arbutin + 5-Keto-aluconate -
IDENTIFICATION
Most probable identification:Leuconostoc citreum
66

APPEDIX F
The result of the fifth LAB strain isolated from poi
CULTURE 10 REPORT
Culture Condition Test ResultDeMan Rogosa Sharpe Colony Morphology Smooth, pinpoint,Agar 48 hous at 30°C translucent colonies
Cell MorDholoav Sinale Daired coccobacilliCatalase NeqaliveOxidase NeaativeGram reaction Positive
DeMan Rogosa Sharpe Growth PositiveBroth with agar plug, 4 . Gas Production Negativedays at 30°CBlood Agar Plate, 24 Hemolysis Alphahours at 30°CDeMan Rogosa Sharpe Growth at 45°C PositiveAaar, 4 days at 45°C
67

API RAPID CH STRIP RESULTS
Test Result Test ResultControl - Esculin -Glycerol - Salicin -Erythritol - Cellobiose -D-Arabinose - Maltose +L-Arabinose - Lactose +Ribose - Melibiose +D-Xv\ose - Sucrose +L-Xvlose - Trehalose -Adonitol - Inuline -~-Methyl- - Melezitose -xylosideGalactose - D-Raffinose +D-Glucose + Starch -D-Fructose + Glvcoaen -D-Mannose + Xylitol -L-Sorbose - 6-Gentiobiose -Rhamnose - D-Turanose +Dulcitol - D-Lyxose -Inositol - D-Tagatose -Mannitol - D-Fucose -Sorbitol - L-Fucose -a-Methyl D· - D-Arabitol -Mannosidea-Methyl D· - L-Arabitol -alucosideN- . Gluconate -AcetvlalucosamineAmygdalin - 2-Keto-aluconate -Arbutin - 5-Keto-gluconate -
IDENTIFICATION
Most probable identification:Leuconostoc lactis
68

LITERATURE CITED
Allen, O.N., Allen, EK, 1933. The manufacture of poi from taro in Hawaii: with
special emphasis upon its fermentation. Hawaii, Agri. Exp. Stn., Bull. No. 70.
Ampe, F., Omar, N. B., Moizan, C., Wacher, C and Guyot, J.-P., 1999.
Polyphasic study of the spatial distribution of microorganisms in Mexican
Pozol, a fermented maize dough, demonstrates the need for cultivation
independent methods to investigate traditional fermentations. Applied and
Environmental Microbiology 65 (12),5464-5473.
Barnett, J.A., Payne, RW. &Yarrow, D., 2000. Yeasts: Characteristics and
identification, 3rd Ed., Cambridge University Press, pp. 270-271.
Battcock, M., Azam-AIi, S., 1998. Bacterial fermentation. In: Fermented Fruits
and Vegetables: A Global Perspective.
http://www.fao.org/docrep/x0560e/x0560e10.htm
Beasley, S., Tuorila, H., Saris, P.E.J., 2003. Fermented soymilk with a
monoculture of Lactococcus lactis, International Journal of Food Microbiology
81,159-162.
Begley, BW., Spielmann, H., Vieth, G.R, 1981. Poi consumption: consumption
of a traditional Staple in the Contemporary Era in Honolulu, Hawaii. Hawaii
Institute of Tropical Agriculture and Human Resources, College of Tropical
Agriculture and Human Resources, University of Hawaii, pp. 3-4.
Beuchat, L. R, 1995. Application of biotechnology to fermented foods. Food
Technology 49 (1), 97-99.
69

Bjorkroth, K. J., Schillinger, U., Geisen, R., Weiss, N., Hoste, B., Holzapfel, W.H.,
Korkeala, H.J. and Vandamme. P., 2002. Taxonomic study of Weissella
confusa and description of Weissella cibaria sp. nov, detected in food and
clinical samples. International Journal of Systermatic and Evolutionary
Microbiology 52,141-148.
Bogovic-Matijasic, B., Rogelj, I., Nes, I.F., Holo, H., 1998. Isolation and
characterization of two bacteriocins of Lactobacillus acidophilus LF221.
Applied Microbiology and Biotechnology 49, 606-612.
Breidt, F., Fleming, H. P., 1997. Using lactic acid bacteria to improve the
safety of minimally processed fruits and vegetables. Food Technology 51 (9),
44-49.
Collins, M.D.• Samelis, J., Metaxopoulos, J. and Wallbanks, S., 1993. Taxonomic
studies on some leuconostoc-like organisms from fermented sausages:
description of a new genus Weissella for the Leuconostoc paramesenteroides
group of species. Journal of Applied Bacteriology 75, 595-603.
Cunningham-Rundles, S., Ahrne, S., Bengmark, 5., 2000. Probiotics and
immune response, Am J Gastroenterol 95(1 Suppl), pp. 22-25.
Daeschel, M.A., 1989. Antimicrobial Substances from Lactic Acid Bacteria for
Use as Food Preservatives. Food Technology 42, 164-167.
Drici-Cachon, Z., Guzzo. J., Cavin. J.-F., Divies, C., 1996. Acid tolerance in
Leuconostoc oemos: Isolation and characterization of an acid-resistant
mutant. Applied Microbiology and Biotechnology 44,785-789.
70

European food and feed cultures association, http://W.JIIW.effca.com
Facklam, R., Hollis, D. and Collins, M.D., 1989. Identification of gram-positive
coccal and coccobacillary vancomycin-resistant bacteria. Journal of Clinical
Microbiology 27,724-730.
Foschino, R., Arrigoni, C., Picozzi, C., Mora, D., Galli, A, 2001. Phenotypic and
genotypic aspects of Lactobacillus samfranciscensis strains isolated from
sourdoughs in Italy. Food Microbiology 18, 277-285.
Foster, J.W., 1999. When protons attach: microbial strategies of acid adaptation.
Curro Opin. Microbiol. 2, 170-174.
Fujisawa, T., Benno, Y., Yaeshima, T., Mitsuoka, T., 1992. Taxonomic study of
the Lactobacillus acidophilus group, with recognition of Lactobacillus
gallinarum sp. Nov. and Lactobacillus johnsonii sp. Nov. and synonymy of
Lacobacillus acidophilus group A3 (Johnson et al. 1980) with the type strain of
Lactobacillus amylovorus (Nakamura 1981). Int. J. Syst. Bacteriol. 42, 487
491.
Hanicoglu, 0., Karapinar, M., 1997. Microflora of boza, a traditional fermented
Turkish beverage. International Journal of Food Microbiology 35,271-274
Hansen, E.B., 2002. Commercial bacterial starter cultures for fermented foods of
the future. International Journal of Food Microbiology, 78, 119-131.
Hounhouigan, D.J., Nout, M.J.R., Nago, C.M., Houben, J.H., Rornbouts, F.M.,
1993. Characterization and frequency distribution of species of lactic acid
bacteria involved in the processing of rnawe, a fermented maize dough from
Benin. International Journal of Food Microbiology 18 (4),279·287.
71

Huang, A.S., Lam, S.Y., Nakayama, T.M., Lin, H., 1994. Microbiological and
chemical changes in poi stored at 20°C. Journal of Agricultural and Food
Chemistry 42, 45-48.
Jack, R.W., Tagg, J.R., Ray B., 1995. Bacteriocins of gram-positive bacteria.
Microbiological Review 59 (2), 171-200.
Jay, J.M., 1996a. Modern Food Microbiology, Fifth ed. Chapman & Hall Inc., New
York, pp. 132-133.
Jay, J.M., 1996b. Modern Food Microbiology, Fifth ed. Chapman & Hall Inc., New
York, pp. 14-15.
Jobin, M-P, Clavel, T., Carlin, F., Schmitt, P., 2002. Acid tolerance response is
low-pH and late-stationary growth phase inducible in Bacillus cereus TZ415.
International Journal of Food Microbiology 79,65-73.
Kim, W.S., Perl, L., Park, J.H., Tandianus, J.E., Dunn, N.W., 2001. Assessment
of stress response of the probiotic Lactobacillus acidophilus. Current
Microbiology 43, 346-350.
Klaenhammer, T.R. 1988. Bacteriocins of lactic acid bacteria. Biochimie 70,
337-349.
Land, G.A. and McCrachen, A.W., 1991. Fungal infections in the compromised
host. In: Arora, D.K., Ajello, L., Mukerji, K.G. (Ed.), Handbook of Applied
Mycology. Marcel Dekker, New York, pp.94-96
Larone, D.H., 1976. Medically Important Fungi: A guide to identification. Harper
and Row Publisher, Hagerstown, MD, pp. 49.
72

Larone, D.H., 1995. Medically Important Fungi: A guide to identification, 3rd Ed.,
ASM Press, Washington D.C., pp. 65-67.
Lee, N. -K., Paik, H. -D., 2001. Partial characterization of lacticin NK24, a newly
identified bacteriocin of lactococcus lactis NK24 isolated from Jeot-gal.
Food Microbiology 18, 17-24.
Lee, Y. -K., Nomoto, K., Salminen, S., Gorbach, S.L., 1999. Handbook of
Probiotics. Wiley Publisher, New York, pp.1; 19.
Lorca, G.L., Font de Valdez, G., 2001. A low-pH-inducible, stationary-phase acid
tolerance response in Lactobacillus acidophilus CRL 639. Current
Microbiology 42,21-25.
Lorca, G.L., Wadtrom, T., Font de Valdez, G., Ljungh, A., 2001. Lactobacillus
acidophilus autolysins inhibit Helicobacter pylori in Vitro. Current Microbiology
42,39-44.
Lund, B.M., Braid-Parker, T.C., Gould, GW., 1999. Microbiological Safety and
Quality of Food. KLUWER academic pUblishers, Vol I & II.
MRS Agar
http://service.merck.de/microbiologyltedisdata/prods/4973-1106600500.html
Manolopoulou, E, Sarantinopoulos, P., Zoidou, E, Aktypis, A., Moschopoulou,
E, Kandarakis, I. G., Anifantakis, EM., 2003. Evolution of microbial
populations during traditional Feta cheese manufacture and ripening.
International Journal of Food Microbiology 82, 153-161.
73

Mao, Y., Muriana, P. M., Cousin MA, 2001. Purification and transpositional
inactivation of Lacticin FS92, a broad-spectrum bacteriocin produced by
Lactococcus !aclis FS92. Food Microbiology 18, 165-175.
Marshall, V.M., Tamine, A.Y., 1997. Starter cultures employed in the
manufacture of biofermented milks. International Journal of Dairy Technology
50,35-41.
Maltila-Sandholm, T., Blum, S., Collins, J.K., Crittenden, R, deVos, W., Dunne,
C., Fondan, R, Grenov, G., Isolauri, E., Kiely, B., Marteau, P., Morelli, L.,
Ouwehand, A, Reniero, R, Saarela, M., Salminen, S., Saxelin, M., Schiffrin,
E., Shanahan, F., Vaughan, E., von Wright, A., 1999. Probiotics: towards
demonstrating efficacy. Trends Food Sci. Techno!. 10,393-399.
Miambi, E., Guyot, J.-P., Ampe, F., 2003. Identification, isolation and
quantification of representative bacteria from fermented cassava dough using
an integrated approach of culture-dependent and culture-independent
methods. International Journal of Food Microbiology 82 (2), 111-120.
Mugula, J.K., Nnko, SAM., Narvhus, J.A., Sorhaug, T., 2003. Microbiological
and fermentation characteristics of togwa, a Tanzanian fermented food.
International Journal of Food Microbiology 80,187-199.
Muyanja, C. M. B. K, Narvhus, J. A, Treimo, J., Langsrud, T., 2003. Isolation,
characterisation and identification of lactic acid bacteria from bushera: a
Ugandan traditional fermented beverage. International Journal of Food
Microbiology 80(3),201-210.
74

Noonpakdee, W., Santivarangkna, C., Jumriangrit, P., Sonomoto, K., Panyim, S.,
2003. Isolation of nisin-producing Lactococcus factis WNC 20 strain from
nham, a traditional Thai fermented sausage. Intemational Journal of Food
Microbiology 81, 137-145.
Olano, A., Chua, J., Schroeder, S., Minari, A., Salvia, M.L., Hall, G., 2001.
Weissella confusa (Basonym: Lactobacillus confuses) Bacteremia: a
Case Report. Journal of Clinical Microbiology 39 (4), 1604-1607.
Olasupo, NA, Olukoya, D.K., Odunfa, SA, 1997. Identification of Lactobacillus
species associated with selected African fermented foods. Z. Naturforsch 52,
105-108
Oyewole, O.B., Odunfa, SA, 1990. Characterisation of lactic acid bacteria in
cassava fermentation during fufu production. J. Appl. Bacteriol. 68,145-152.
Paludan-Muller, C., Huss, H.H., Gram, L.. 1999. Characterization of lactic acid
bacteria isolated from a thai low-salt fermented fish product and the roe of
garlic as substrate for fermentation. International Journal of Food Microbiology
46,219-229.
Papadelli. M., Manolopoulou, E., Kalantzopoulos, G., Tsakalidou, E., 2003. Rapid
detection and identification of Streptococcus macedonicus by species-specific
PCR and DNA hydridisation. International journal of Food Microbiology 81,
231-239.
Pouwels. D., Simons, G., 2003. Fingerprinting Microorganisms. Food
Technology 57(2}, 36-40.
75

Ralph, W., John, J., Tagg, R, Ray, B., 1995. Bacteriocins of gram-positive
bacteria. Microbiological Reviews 59(2), 171-200.
Ray, B., 2001a. Fundamental Food Microbiology, second ed. CRC Press LLC.
Boca Raton, Florida, pp. 110.
Ray, B., 2001b. Fundamental Food Microbiology, second ed. CRC Press LLC.
Boca Raton, Florida, pp. 165-166.
Ray, B., 2001c. Fundamental Food Microbiology, second ed. CRC Press LLC.
Boca Raton, Florida, pp. 170-171.
Ray, B., 2001d. Fundamental Food Microbiology, second ed. CRC Press LLC.
Boca Raton, Florida, pp. 184-185.
Ray, B., 2001e. Fundamental Food Microbiology, second ed. CRC Press LLC.
Boca Raton, Florida, pp. 111-115.
Ray, B., 2001f. Fundamental Food Microbiology, second ed. CRC Press LLC.
Boca Raton, Florida, pp. 91.
Ray, B., Daeschel, M., 1992. Food biopreservalives of microbial origin. CRC
Press, Inc. Boca Raton, Florida, pp. 17-21.
Rogelj, I. Bogovic-Matijasic, B., Canzek-Majhenic, A., Stojkovic, S., 2002. The
survival and persistence of Lactobacillus acidophilus LF221 in different
ecosystems. International Journal of Food Microbiology 76, 83-91.
Ross, R P., Morgan, S., Hill, C., 2002. Preservation and fermentation: past,
present and future. International Journal of Food Microbiology, 79, 3-16.
Roth, A., Worth, RM., Lichton, I.J., 1967. Use of poi in the prevention of allergic
disease in potentially allergic infants. Annals of Allergy 25, 501·506.
76

Roy, D., Ward, P., Vincent, D., Mondou, F., 2000. Molecular identification of
potentially probiotic lactobacilli. Current Microbiology 40, 40-46.
Roy, D., Sirois, S., Vincent, D., 2001. Molecular Discrimination of Lactobacilli
used as starter and probiotic cultures by amplified ribosomal DNA restriction
analysis. Current Microbiology 42, 282-289.
Salminen, S., Wright, AV., 1993. Lactic Acid Bacteria. Marcel Dekker, Inc.,
New York, pp. 1-4.
Sanders, M.E., Klaenhammer, T.R., 2001. Invited review: the scientific basis of
Lactobacillus acidophilus NCFM functionality as a probiotic. Journal of Dairy
Science 84, 319-331.
Sanni AI., Morton-Guyot J., Guyot J.P., 2002. New efficient amylase-producing
strains of Lactobacillus plantarum and L. fermentum isolated from different
Nigerian traditional fermented foods. International Journal of Food
Microbiology 72(1-2),53-62.
Shah, N.P., 2001. Functional foods from probiotics and prebiotics. Food
Technology 55(11),46-53.
Sneath, P.H.A. (Ed.), 1986. Bergey's Manual of Systematic Bacteriology, Vol. 2.
Williams &Wilkins, Baltimore.
Soomro, AH., Masud, T., Anwaar, K., 2002. Role of lactic acid bacteria (LAB) in
food preservation and human health- a review. Pakistan Journal of Nutrition 1
(1),20-24.
Steinkraus, K. H., 2002. Fermentations in Wortd Food Processing.
Comprehensive Reviews in Food Science and Food Safety 1, 23-32.
77

Steinkraus, K.H., 1983a. Lactic acid fermentation in the production of foods from
vegetables, cereals and legumes. Antonie Van Leeuwenhoek 49 (3), 337-348.
Steinkraus, K.H., 1983b. Handbook of Indigenous Fermented Foods
(Microbiology series, Vol. 9). Marcel Dekker, Inc., New York, pp. 620
Tamime A.Y., Deeth H.C., 1980. Yougurt: Technology and Biochemistry. Journal
of Food Protection 43(12),939-977.
Tannock, G.W., 1999. Probiotics-a critical review. Horizon Scientific Press,
Norfolk, England, pp. 1.
Tetteh, G.L., Beuchat, L.R, 2003. Exposure of Shigella f1exneri to acid stress
and heat shock enhances acid tolerance. Food Microbiology 20 (2), 179-185.
USDA Nutrient Database for Standard Reference.
http://www.nal.usda.gov/fnic/cgi-bin/listnut.pl
Vandenbergh, P.A., 1993. Lactic acid bacteria, their metabolic products and
interference with microbial growth. FEMS Microbiology Reviews 12,
221-238.
Vescovo, M., Torriani, S., Orsi, C., Macchiarolo, F., Scolari, G., 1996. Application
of antimicrobial-producing lactic acid bacteria to control pathogens in ready-to
use vegetables. Journal of Applied Bacteriology 81, 113-119.
Vogel, RF., Knorr, R, Muller, M.R, et aI., 1999. Non- dairy lactic fermentations:
the cereal world, Antonie Van Leeuwenhoek 76 (1-4), 403-411.
Woese, C.R, 1987. Bacterial evolution. Microbiological Review 51, 221-271.
78

Zhou, J.S., Shu, Q., Rutherfurd, K.J., 2000. Safety assessment of potential
probiotic lactic acid bacterial strains Lactobacillus rhamnosus HN001, Lb.
acidophilus HN017, and Bifidobacterium lactis HN019 in BALB/c mice.
International Journal of Food Microbiology 56, 87-96.
79