of metatheria - proceedings of the national academy of sciences
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 91, pp. 12569-12573, December 1994Evolution
New Cretaceous marsupial from Mongolia and the early radiationof Metatheria
(vertebrate paleontology/therian mammals/biogeography)
BORIS A. TROFIMOV* AND FREDERICK S. SZALAYt**Paleontological Institute, Russian Academy of Sciences, Moscow, Russia; and tHunter College, City University of New York, 695 Park Avenue, New York,NY 10021
Communicated by F. Clark Howell, September 8, 1994
ABSTRACT A marsupial mammal from Mongolia repre-senting a previously unreported group, Asiadelphia, offersunequivocal evidence that metatherians were represented onthe continent of Asia during the Late Cretaceous (Campanian),even in the northern arid interior. Asiatherium is known byskull, left and right mandibles, and most of its postcranialskeleton. Comparisons of the dentition, ear region, and aspectsof the postcranium not only allow its diagnosis as an Asiangroup of metatherians but also permit a diagnosis of theMetatheria based on the unequivocally apomorphous conditionof the dental formula and probable replacement pattern of theprotometatherian, in contrast to the more primitive theriandental formula, represented by the few known Aegialodontidaeand the first dental eutherians. It appears an inescapableconclusion that the first metatherians had a more deriveddental formula (and probably replacement pattern) than theearliest dental eutherians, regardless of what specific deriva-tion from a therian ancestry is contemplated. Such a diagnosisalso supports the metatherian status of other Cretaceous Asianand American taxa such as the Deltatheroida and KokopeUia.The hypothesis that Metatheria originated in North America islargely dependent on the preponderance of Cretaceous formsfrom North America and negative evidence from Asia (i.e., onthe lack of lower latitude Early Cretaceous tropical faunas).Conversely, the relative Cretaceous paucity of placentals fromNorth America and their greater abundance in Asia suggeststhe earliest flowering of the Eutheria in an unspecified regionof the Old World. The concept of a holophyletic Theria(=Tribosphenida) employed here, based on the biologicallysignificant apomorphy of the molar form-function of the firsttherian, entails only the tribosphenic mammals (infraclassesTribotheria, Metatheria, and Eutheria) but not the sundryquasi-triangular-toothed mammals and their derivatives in theMesozoic (Eupantotheria, Peramura, Monotremata, etc.). It issuggested that the Asiadelphia and Deltatheroida, the pro-tospecies of both, with the apomorphic therian dental formula,are ancient lineages of Metatheria independent from theirNorth American relatives probably since the early part of theLate Cretaceous.
SYSTEMATICS
Order Asiadelphia, nov.
"Diagnosis" and briefdiscussion. Because unequivocallyderived attributes of the protometatherian or proto-asiadelphian are nearly impossible to separate at present, this"diagnosis" by necessity also includes traits consideredapomorphic for the first metatherian; some traits were prob-
ably also present in the specific tribotherian ancestry as well:small Cretaceous therians with alisphenoid component tobulla and with basicranial attributes characteristic of met-atherians, such as transversely elongated and ellipticalfenestra rotunda and round fenestra ovale; postcanine dentalformula premolars P1,2,3; molars M1,2,3,4, with a sharpbreak between the ultimate premolar and first molar (bothconditions derived therian ones, primitive for all Metatheria);poorly developed stylar cusps (may represent primitive con-dition for Metatheria); protocone, in spite of its large appear-ance due to the large conules, ofa quite small size comparableto the size of the occluding (lingual portion) hypoconid;carpus differing from known primitive Ameridelphia (lastcommon ancestor of living Didelphimorphia and Paucituber-culata) by lacking the prominent distolateral process of thescaphoid of the former; proximal fibula apomorphously slen-der (distinguishing it from other primitive metatherians)-acondition of fibula that is more derived than that of thePaucituberculata (the only comparable condition in ameri-delphians and almost certainly independently evolved), but inarticulation with the femur as in nontherian (i.e., atri-bosphenic) primitive mammals and primitive Theria, with aclear suggestion of terrestrial locomotor adaptation.Included taxa. Family Asiatheriidae, nov.Type genus. Asiatherium gen. nov.Type and only species. Asiatherium reshetovi, sp. nov.Etymology. For the memory of Yuri V. Reshetov, recently
deceased member of the Paleontological Institute of theRussian Academy of Sciences, for his pioneering field workin Mongolia and other parts of Central Asia and for hisseminal contributions to the understanding of early ungulateevolution.
Holotype. Paleontological Institute of the Russian Acad-emy of Sciences (PIN) no. 3907; crushed skull and bothmandibles, nearly complete, with dentition represented fromP1 to M4; skeleton lacking sternum, left hand, and both feet.
Stratigraphic provenance and geographic range. LateCretaceous Udan Sayr locality, 85 kmNW of Bulgan Somon,Umuni Gobi Aimak (southern region), Mongolia; from bedsof similar lithology and age to the Barun Goyot Formation (1).Udan Sayr, along with Shara Tsav, Khara Khutul, andBambu Khuduk, among others, have been considered ofBarungoyotian Age (2), approximately middle Campanianequivalent, perhaps between 80 and 76 million years beforepresent (MYBP).Genus diagnosis. While dental formula is similar to other
metatherians, the following differences diagnose this species(ofthe monotypic genus) from various Archimetatheria (3, 4),known primarily by dental remains from North America andfrom Asian Deltatheroida (5). P3 (lower premolar 3) notsignificantly taller than M1 (lower first molar) in contrast tothe condition seen in the Albian metatherian Kokopellia (6)and in North American Cretaceous Metatheria (7). Paraconid
tTo whom reprint requests should be addressed.
12569
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

12570 Evolution: Trofimov and Szalay
is the strongest lingual cusp on M4 only, metaconid taller thanparaconid on M1_3, as in Kokopellia, but differs from theNorth American genus in having a deep U-shaped (ratherthan V-shaped) talonid notch (that between metaconid andentoconid), a relatively mesiodistally shorter talonid com-pared with trigonid, taller talonid cusps, but relatively lesstall trigonid cusps compared with the talonid. The buccal anddistal cristids of the talonid (the cristid obliqua and theposthypoconid cristid) form a more acute, smaller angle inAsiatherium than that in Kokopellia. The entoconid andhypoconulid are tall and acute cusps and are slightly moreclosely twinned than in Kokopellia, similar to those inProtalphadon (8) and Alphadon (9), but unlike those indeltatheroidans or known Cretaceous dental eutherians (7,10-13). Asiatherium differs from other Cretaceous NorthAmerican dentally primitive metatherians such as Anchisto-delphys (9), Iqualadelphis (8), Protalphadon (8), or primitiveand early Asian eutherians such as the otlestid Prokennal-estes (10) in having its entoconid and hypoconulid taller andmore acute and having a relatively narrow stylar shelf on theupper molars (or even none on M/1), in addition to havinglost the connection of the preparacrista with the stylocone(cusp B). Unlike in some of the former genera, the stylarcusps are very faint as in Deltatheroida or Pediomys, andonly their traces (incipient or reduced) are visible. As inDeltatheroida [e.g., Sulestes (5)], the metacone is relativelylarge compared with the paracone, larger than that in dentallyprimitive Eutheria, but it is nearly comparable in size to theparacone in other early metatherians. Upper molars havewell-developed pre- and postcingulae. Unlike in all the for-mer taxa listed, the conules and pre- and postprotocristae arestrongly developed. A preliminary examination suggests thatthe enamel contains tubules, typical of most metatherians.
DISCUSSIONThe type specimen ofAsiatherium reshetovi is exceptionallycomplete for a Cretaceous therian mammal, particularly fora metatherian in that it has most of its skeleton preserved(Fig. 1). The crushed skull (Figs. 2 and 3) has been recon-structed and shows, in many respects, what may be consid-ered primitive therian conditions in addition to a suite ofapomorphies. The nasal structure contacts the lacrimal struc-ture, and the jugal structure contacts the glenoid fossa, bothbeing primitive, probably late synapsid traits [see particularly
Sinoconodon (3)]. The palate is fenestrated as in the probableprimitive condition represented for the deltatheroidans by theGurlin Tsav skull (5), but unlike in the ameridelphian bor-hyaenids. The basicranium shows continuity between thealisphenoid and a medially located, partially ossified bulla.The postcanine dental formula is unequivocally metathe-
rian (Fig. 4). While the suggestion (14) that the Owen (15)-Lydekker (16) hypothesis for the homology of the metathe-rian Ml may prove to be correct [i.e., equivalent to theeutherian dP4 (fourth deciduous premolar)], at present wecontinue to use the M1-4 designation. As we and othersdiscuss elsewhere (3, 4, 14), in any probable derivation of themetatherian dental formula, the reduction of the premolarnumber and the (putative) minimal replacement pattern of theantemolar dentition [only dP3 is replaced (14)] is unequivo-cally derived in therians (tribosphenic mammals), and there-fore it represents a significant diagnostic trait (3, 4, 6, 14) thatdesignates the first metatherian in a testable manner.Thorough reviews of molar features of early therians (7,
17-20) have attempted to sort out those topographic at-tributes that maximally postdict whether these rare taxa wereeither tribotherian, metatherian, or eutherian. In spite of anumber of important new forms described during the pastdecade (8-10, 21-27), this has become increasingly difficulton the basis of molar teeth alone. As noted, the distolingualportion of the talonid (entoconid-hypoconulid) in Asiathe-rium is characteristically metatherian (Fig. 4), although thisis not necessarily a diagnostic feature of the protometathe-rian. The near subequality of the metacone to the paraconesignals the nearly ubiquitous metatherian "tendency" tohypertrophy the metacone (tied to entoconid-hypoconulidtwinning functionally). Many living didelphids, Cretaceousand Paleogene archimetatherians, sudameridelphians (3),primitive deltatheroidans (in particular the Gurlin Tsavskull), Asiatherium, and Kokopellia have an increasing sizegradient from M1-4 (except for the slightly reduced M3),where the first upper molar (Ml) is decidedly smaller than M2and the latter is smaller than M3. This is not the conformationof the dentition in the dentally primitive eutherians such asthe Early Cretaceous otlestids, where M1 is larger than M2(10, 11) as in a number of Paleogene groups. By this criterion,in addition to the dental formula discussed below, not onlyasiadelphians but also deltatheroidans and Kokopellia areMetatheria. Protoconid is the tallest cusp in Asiatherium, aprimitive therian condition, and the paraconid is taller than
1 cm
FIG. 1. Asiatherium reshetovi, new genus and species, type specimen. Paleontological Institute ofthe Russian Academy of Sciences no. 3907,Udan Sayr, Mongolia. Nearly complete skeleton, crushed skull, and mandibles (mandibles are not shown).
Proc. Natl. Acad. Sci. USA 91 (1994)

Proc. Natl. Acad. Sci. USA 91 (1994) 12571
FIG. 2. Asiatherium reshetovi, stereophotos in dorsal (Upper) and ventral (Lower) views. (Subdivisions on scale = 1 mm.)
the metaconid only on M4; on the antecedent teeth, it is themetaconid that is the dominant cusp on the lingual side of thetrigonid. The latter is a common didelphid condition, butsignificantly for the analysis of dental taxonomic properties(particularly for Deltatheroida), it was altered probably in-dependently more in carnivorous (i.e., shearing) lineagessuch as Lutreolina, Didelphis, Chironectes, sparassocynids,sparassodontans, and dasyuromorphians.The scapula has a long and attenuated scapular spine, a
primitive therian condition prominently displayed in livingdidelphids. A similarly ancient condition, not a metatheriandiagnostic trait, is the presence of well-developed epipubicbones. The head of the radius is oval in outline, not round asin most didelphids, and the anterior edge of the olecranonprocess is twisted medially, in a manner characteristic ofterrestrial didelphids and caenolestids. The knee is imper-fectly preserved, but a number of attributes are clear, such asthe articular contact ofthe fibula with the distal femur and therelatively reduced proximal fibula compared with didelphids,
in which it is usually wide and flared (4). The derivedcondition (within Metatheria) of the distal femur is alsocharacteristic in a number of terrestrial metatherian lineagessuch as caenolestids and peramelids. The lack of any sign ofa patellar groove on the distal femur is a very strong corrob-oration that the animal is not a eutherian but a metatherian(4).The tarsus has proved to be a singularly reliable and
diagnostic attribute of metatherian higher taxa (3, 4, 28), andthere are preliminary indications that the system of con-straints that appears to govern the changes in the tarsus aresomewhat similarly operational in the hand as well (4). Thereis enough preserved of the right hand of Asiatherium to ruleout specific allocations to other higher taxa within Metathe-ria. The radius and ulna are closely pressed against oneanother, and the distal ends on both bones have relativelyacute styloid processes, suggestive of the terrestrial adapta-tion seen in caenolestids. Only the scaphoid, trapezium, andtrapezoid appear to be in their natural position, whereas the
Evolution: Trofimov and Szalay
12572 Evolution: Trofimov and Szalay
/
B
B
53
C D
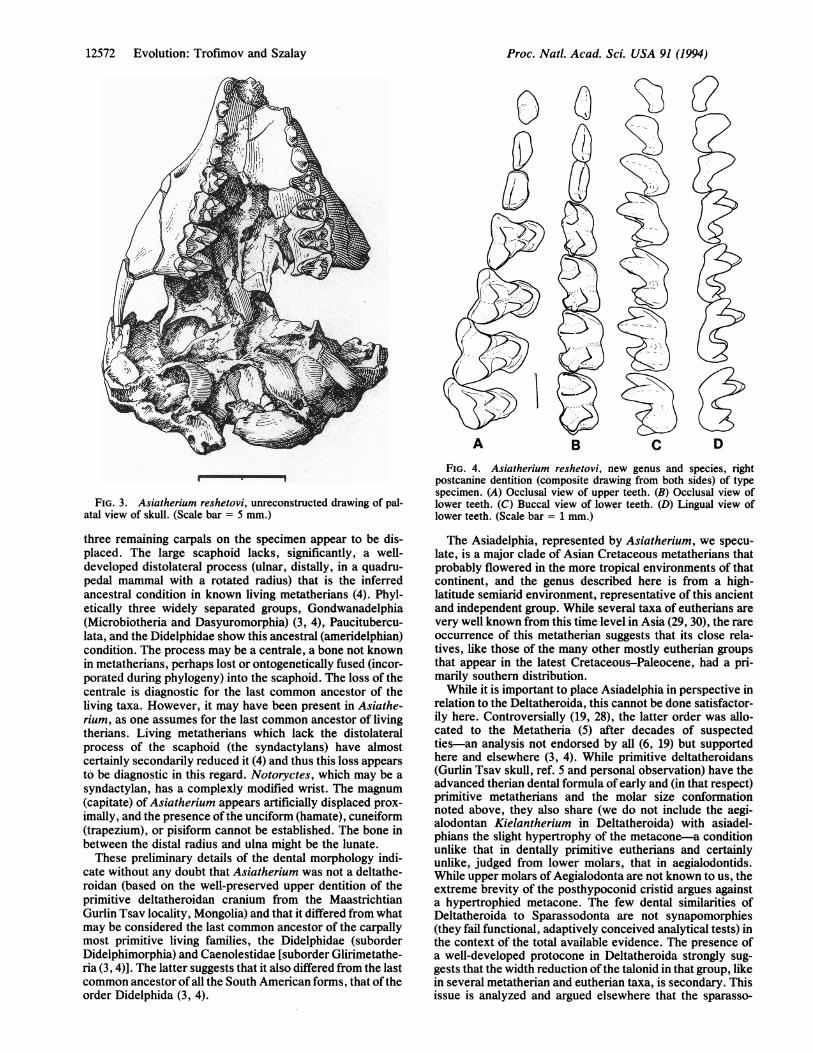
FIG. 3. Asiatherium reshetovi, unreconstructed drawing of pal-atal view of skull. (Scale bar = 5 mm.)
three remaining carpals on the specimen appear to be dis-placed. The large scaphoid lacks, significantly, a well-developed distolateral process (ulnar, distally, in a quadru-pedal mammal with a rotated radius) that is the inferredancestral condition in known living metatherians (4). Phyl-etically three widely separated groups, Gondwanadelphia(Microbiotheria and Dasyuromorphia) (3, 4), Paucitubercu-lata, and the Didelphidae show this ancestral (ameridelphian)condition. The process may be a centrale, a bone not knownin metatherians, perhaps lost or ontogenetically fused (incor-porated during phylogeny) into the scaphoid. The loss of thecentrale is diagnostic for the last common ancestor of theliving taxa. However, it may have been present in Asiathe-rium, as one assumes for the last common ancestor of livingtherians. Living metatherians which lack the distolateralprocess of the scaphoid (the syndactylans) have almostcertainly secondarily reduced it (4) and thus this loss appearsto be diagnostic in this regard. Notoryctes, which may be a
syndactylan, has a complexly modified wrist. The magnum(capitate) of Asiatherium appears artificially displaced prox-imally, and the presence ofthe unciform (hamate), cuneiform(trapezium), or pisiform cannot be established. The bone inbetween the distal radius and ulna might be the lunate.These preliminary details of the dental morphology indi-
cate without any doubt that Asiatherium was not a deltathe-roidan (based on the well-preserved upper dentition of theprimitive deltatheroidan cranium from the MaastrichtianGurlin Tsav locality, Mongolia) and that it differed from whatmay be considered the last common ancestor of the carpallymost primitive living families, the Didelphidae (suborderDidelphimorphia) and Caenolestidae [suborder Glirimetathe-ria (3, 4)]. The latter suggests that it also differed from the lastcommon ancestor of all the South American forms, that oftheorder Didelphida (3, 4).
FIG. 4. Asiatherium reshetovi, new genus and species, rightpostcanine dentition (composite drawing from both sides) of typespecimen. (A) Occlusal view of upper teeth. (B) Occlusal view oflower teeth. (C) Buccal view of lower teeth. (D) Lingual view oflower teeth. (Scale bar = 1 mm.)
The Asiadelphia, represented by Asiatherium, we specu-late, is a major clade of Asian Cretaceous metatherians thatprobably flowered in the more tropical environments of thatcontinent, and the genus described here is from a high-latitude semiarid environment, representative of this ancientand independent group. While several taxa of eutherians arevery well known from this time level in Asia (29, 30), the rareoccurrence of this metatherian suggests that its close rela-tives, like those of the many other mostly eutherian groupsthat appear in the latest Cretaceous-Paleocene, had a pri-marily southern distribution.While it is important to place Asiadelphia in perspective in
relation to the Deltatheroida, this cannot be done satisfactor-ily here. Controversially (19, 28), the latter order was allo-cated to the Metatheria (5) after decades of suspectedties-an analysis not endorsed by all (6, 19) but supportedhere and elsewhere (3, 4). While primitive deltatheroidans(Gurlin Tsav skull, ref. 5 and personal observation) have theadvanced therian dental formula of early and (in that respect)primitive metatherians and the molar size conformationnoted above, they also share (we do not include the aegi-alodontan Kielantherium in Deltatheroida) with asiadel-phians the slight hypertrophy of the metacone-a conditionunlike that in dentally primitive eutherians and certainlyunlike, judged from lower molars, that in aegialodontids.While upper molars of Aegialodonta are not known to us, theextreme brevity of the posthypoconid cristid argues againsta hypertrophied metacone. The few dental similarities ofDeltatheroida to Sparassodonta are not synapomorphies(they fail functional, adaptively conceived analytical tests) inthe context of the total available evidence. The presence ofa well-developed protocone in Deltatheroida strongly sug-gests that the width reduction ofthe talonid in that group, likein several metatherian and eutherian taxa, is secondary. Thisissue is analyzed and argued elsewhere that the sparasso-
A
Proc. Natl. Acad Sci. USA 91 (1994)

Proc. Natl. Acad. Sci. USA 91 (1994) 12573
donts are unequivocally ameridelphians and not relatives ofdeltatheroidans (4).
Figs. 1 and 3 were prepared by V. D. Kolganov. Travel for F.S.S.was supported by the National Geographic Society. Critical readingand constructive criticisms by Richard L. Cifelli, Zofia Kielan-Jaworowska, and Christian de Muizon are greatly appreciated; theirthorough readings have improved the manuscript.
1. Ivakhnenko, M. F. & Kurzanov, S. M. (1988) Tr. SovmestnayaSov. Mong.-Paleotol. Eksped. 34, 100-105 (in Russian).
2. Jerzykiewicz, T. & Russell, D. A. (1991) Cretaceous Res. 12,345-377.
3. Szalay, F. S. (1993) in Mammalian Phylogeny: Mesozoic Dif-ferentiation, Multituberculates, Monotremes, Early Theriansand Marsupials, eds. Szalay, F. S., Novacek, M. J. & Mc-Kenna, M. C. (Springer, New York), pp. 216-241.
4. Szalay, F. S. (1994) Evolutionary History ofthe Marsupials andan Analysis of Osteological Characters (Cambridge Univ.Press, New York).
5. Kielan-Jaworowska, Z. & Nessov, L. A. (1990) Lethaia 23,1-10.
6. Cifelli, R. L. (1993) Proc. Natl. Acad. Sci. USA 90,9413-9416.7. Clemens, W. A. & Lillegraven, J. A. (1986) Contrib. Geol.
Spec. Pap. 3, 55-85.8. Cifelli, R. L. (1990) J. Vertebr. Paleontol. 10, 295-319.9. Cifelli, R. L. (1990) J. Vertebr. Paleontol. 10, 320-331.
10. Kielan-Jaworowska, Z. & Dashzeveg, D. (1989) Zool. Scr. 18,347-355.
11. Nessov, L. A. & Kielan-Jaworowska, Z. (1991) Contrib. Pal.Mus. Univ. Oslo 364, 51-52.
12. Clemens, W. A. (1979) in Mesozoic Mammals: The First Two-Thirds ofMammalian History, eds. Lillegraven, J. A., Kielan-
Jaworowska, Z. & Clemens, W. A. (Univ. of California Press,Berkeley), pp. 192-220.
13. Fox, R. C. (1984) Carnegie Mus. Nat. Hist. 9, 9-20.14. Luckett, W. P. (1993) in Mammalian Phylogeny: Mesozoic
Differentiation, Multituberculates, Monotremes, Early Theri-ans and Marsupials, eds. Szalay, F. S., Novacek, M. J. &McKenna, M. C. (Springer, New York), pp. 182-204.
15. Owen, R. (1868) On the Anatomy of Vertebrates (Longmans,Green, London), Vol. 3.
16. Lydekker, R. (1887) Catalogue ofthe Fossil Mammalia in theBritish Museum (Natural History) (Trustees of the BritishMuseum, Natural History, London).
17. Patterson, B. (1956) Fieldiana Geol. 13, 1-105.18. Butler, P. M. (1978) Breviora Mus. Comp. Zool. 446, 1-27.19. Cifelli, R. L. (1993) in Mammalian Phylogeny: Mesozoic Dif-
ferentiation, Multituberculates, Monotremes, Early Theriansand Marsupials, eds. Szalay, F. S., Novacek, M. J. & Mc-Kenna, M. C. (Springer, New York), pp. 208-215.
20. Fox, R. C. (1975) Can. J. Earth Sci. 12, 412-442.21. Fox, R. C. (1971) Zool. J. Linn. Soc. 50 (Suppl. 1), 145-164.22. Fox, R. C. (1972) Can. J. Earth Sci. 9, 1479-1494.23. Fox, R. C. (1980) Can. J. Earth Sci. 17, 1489-1498.24. Cifelli, R. L. (1990) J. Vertebr. Paleontol. 10, 330-331.25. Cifelli, R. L. (1990) J. Vertebr. Paleontol. 10, 332-345.26. Cifelli, R. L. (1990) J. Vertebr. Paleontol. 10, 346-360.27. Cifelli, R. L. & Eaton, J. G. (1987) Nature (London) 325,
520-522.28. Szalay, F. S. (1982) in Carnivorous Marsupials, ed. Archer, M.
(Royal Zoological Society of New South Wales, Sydney), pp.621-640.
29. Kielan-Jaworowska, Z. (1981) Paleontol. Pol. 42, 27-78.30. Kielan-Jaworowska, Z. (1984) Paleontol. Pol. 46, 173-183.
Evolution: Trofimov and Szalay