of 263, 1535-1541, 1988 1988 and biology, inc. u. a. human transforming growth factor ... · ·...
TRANSCRIPT

THE JOURNAL 0 1988 by The American Society for Biochemistry
OF BIOLOGICAL CHEMISTRY and Moleculal ' Biology, Inc.
Vol. 263, NO. 3, Issue of January pp. 1535-1541, 1988 Printed in U. S. A.
Human Transforming Growth Factor /3. cw2-Macroglobulin Complex Is a Latent Form of Transforming Growth Factor /3*
(Received for publication, March 13, 1987)
Shuan Shian Huang, Pauline O'Grady, and Jung San HuangS From the E. A. Doisy Department of Biochemistry, St. Louis University School of Medicine, St. Louis, Missouri 63104
lZ6I-Labeled human platelet-derived transforming growth factor B ('"'I-TGF-B) and human az-macroglob- ulin (aZM) formed a complex as demonstrated by 5% native polyacrylamide gel electrophoresis. The "'I- TGF-B. azM complex migrated at a position identical to that of the fast migrating form of aZM. Most of the "'I- TGF-6. azM complex could be dissociated by acid or urea treatment. When lZ6I-TGF-B was incubated with serum, the high molecular weight form of '261-TGF-B could be immunoprecipitated by anti-human azM anti- sera as demonstrated by sodium dodecyl sulfate-poly- acrylamide gel electrophoresis. azM purified from platelet-rich plasma also showed the latent transform- ing growth factor activity and immunoreactivity of TGF-8. These results suggest that TGF-B.azM complex is a latent form of TGF-8.
Human platelet-derived transforming growth factor (TGF-P)' is a 25-kDa protein composed of two identical polypeptide chains linked by disulfide bridges (1-4). TGF-P stimulates colony formation of normal rat kidney fibroblasts (NRK cells) and AKR-2B cells in soft agar (5,6). TGF-P also inhibits cell growth of some normal and transformed cells (7, 8). TGF-P appears to stimulate the synthesis of collagen and fibronectin in a variety of normal and transformed cells (9, 10). Normal bronchial epithelial cells undergo squamous dif- ferentiation following stimulation by TGF-/3 (11). The phys- iological function of TGF-P is unknown. Since TGF-P and other a-granule proteins are released from platelets at injury sites, it is believed that TGF-P plays an important role in the process of wound healing (2).
TGF-/3 is released from platelets in the form of a high molecular weight complex with low growth-promoting activity for NRK cells in soft agar (12). Treatment of this high molecular weight complex with acid or urea liberates the active form of TGF-P with an apparent molecular weight of -20,000 (12). The physiological significance of acid or urea activation of the latent form of TGF-P is not clear. It is very likely that the in vivo activation of the latent form of TGF-P is mediated by other mechanism(s). To characterize the in uiuo activation process of the latent form of TGF-P, we wished
* This work was supported by National Institutes of Health Grant CA 38808 (to J. S. H.) and by a grant-in-aid, American Heart Association, Missouri Affiliate (to S. S. H.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Recipient of a faculty research award of the American Cancer Society.
The abbreviations used are: TGF-(3, transforming growth factor (3; PDGF, human platelet-derived growth factor; azM, human az- macroglobulin; HEPES, 4-(2-hydroxyethyl)-l-piperazineethanesul- fonic acid; SDS, sodium dodecyl sulfate.
to identify the binding protein for TGF-P in serum or plasma. In this communication, we report that the major binding protein of TGF-P in serum or plasma is a,-macroglobulin (a,M) and that the TGF-P.azM complex is a latent form of TGF-P.
EXPERIMENTAL PROCEDURES
Materiak-Na1251 (17 Ci/mg) was purchased from Du Pont-New England Nuclear. Soybean trypsin inhibitor, N"-benzoyl-DL-arginine- p-nitroanilide HCl, methylamine HCl, N-ethylmaleimide, chloramine T, human serum albumin, iodoacetic acid, N-acetyl tyrosine, 5,5'- dithiobis-(2-nitrobenzoic acid),p-chloromercuribenzoic acid, and pro- tein markers (high and low molecular weight) were obtained from Sigma. Polyclonal rabbit anti-human azM antisera were obtained from Behring Diagnostics and Sigma. These antisera were further tested using immunoblot analysis. Bovine trypsin was obtained from Worthington. IgGsorb was obtained from the Enzyme Center. Ultro- gel AcA 34 and Ultrogel AcA 22 gels were obtained from LKB. Anti- azM IgG-Sepharose 4B gel (1 mg/ml) was prepared by the cyanogen bromide procedure. Human serum and platelet-poor plasma were prepared as previously described (13). Outdated human platelet-rich plasma was obtained from the American Red Cross, St. Louis, MO. Rabbit anti-human TGF-(3 IgG (0.1 mg/ml) was kindly provided by Drs. Jorma Keski-Oja and Harold L. Moses, Department of Cell Biology, Vanderbilt University School of Medicine, TN. Human fibronectin was kindly provided by Dr. John A. McDonald, Washing- ton University School of Medicine, St. Louis, MO.
Preparation and Iz5Z Labeling of TGF-@-Human platelet-derived TGF-@ was prepared according to Assoian et al. (2) and kindly provided by Dr. Anita B. Roberts, National Cancer Institute. TGF-(3 (4 pg) was iodinated with 2 mCi of Na'T in the presence of chlora- mine T as described by Frolik et al. (14). lZ5I-TGF-(3 and free lZ6I were separated on a column of Sephadex G-25 in 4 mM HCl containing 0.1% human serum albumin. The specific radioactivity of 1251-TGF-(3 was -2 x lo5 cpm/ng. An anchorage-independent cell growth-pro- moting activity assay (15) indicated no loss of the biological activity resulted from the iodination procedure. lZ5I-TGF-@ was stored at -20 'C before use.
Preparation and lZ51 Labeling of PDGF-PDGF (M, -28,000) was purified from outdated human platelet-rich plasma as previously described (16). The homogeneity of PDGF was established by analyt- ical SDS-polyacrylamide gel electrophoresis and isoelectric focusing. PDGF was labeled with Na'T using the IODO-GEN method (16). The specific radioactivity of Y-PDGF was -1-2 X lo4 cpm/ng. lZ5I- PDGF was stored at -20 "C before use.
Preparation of Human aZM-Human azM was prepared from outdated human platelet-poor plasma by zinc chelate chromatography (17) and Barrett's (18) method with slight modifications. In the procedure of Barrett, outdated human plasma (100 ml) was precipi- tated at 5.5-12.5s (w/v) of polyethylene glycol 6000. The precipitates were dissolved in 20 ml of 0.1 M sodium phosphate buffer, pH 7.0, and then applied to a column (5.0 X 72 cm) of Ultrogel AcA 34/ Ultrogel AcA 22 (2:1, v/v) and eluted with the same phosphate buffer. The flow rate and fractional volume were 20 ml/h and 3 ml, respec- tively. The first protein peak was identified as azM by protease inhibitor activity assay and by immunoblot analysis. The protease inhibitor activity of azM was assayed by the soybean trypsin inhibi- tor-resistant bound trypsin activity assay (19). The purified azM preparations showed a single band in reducing and nonreducing SDS- polyacrylamide gel electrophoresis (5 and 7.5% gels) followed by silver
1535
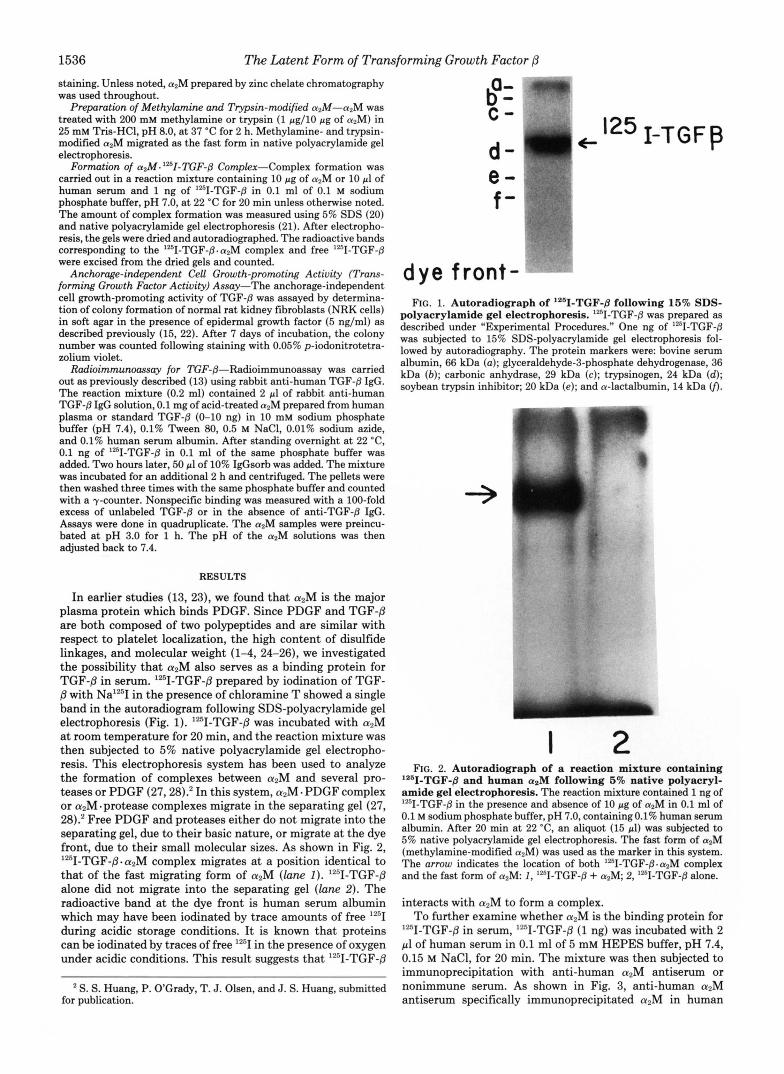
1536 The Latent Form of Transforming Growth Factor /3 staining. Unless noted, a2M prepared by zinc chelate chromatography was used throughout.
Preparation of Methylamine and Tmsin-modified aa--azM was treated with 200 mM methylamine or trypsin (1 pg/lO pg of azM) in 25 mM Tris-HC1, pH 8.0, at 37 "C for 2 h. Methylamine- and trypsin- modified azM migrated as the fast form in native polyacrylamide gel electrophoresis.
Formation of a2M.126Z-TGF-8 Complex-Complex formation was carried out in a reaction mixture containing 10 pg of aZM or 10 pl of human serum and 1 ng of 'z51-TGF-@ in 0.1 ml of 0.1 M sodium phosphate buffer, pH 7.0, a t 22 "C for 20 min unless otherwise noted. The amount of complex formation was measured using 5% SDS (20) and native polyacrylamide gel electrophoresis (21). After electropho- resis, the gels were dried and autoradiographed. The radioactive bands corresponding to the lz5I-TGF-8.a2M complex and free '2sI-TGF-8 were excised from the dried gels and counted.
Anchorage-independent Cell Growth-promoting Activity (Trans- forming Growth Factor Activity) Assay-The anchorage-independent cell growth-promoting activity of TGF-8 was assayed by determina- tion of colony formation of normal rat kidney fibroblasts (NRK cells) in soft agar in the presence of epidermal growth factor (5 ng/ml) as described previously (15, 22). After 7 days of incubation, the colony number was counted following staining with 0.05% p-iodonitrotetra- zolium violet.
Radioimmunoassay for TGF-8-Radioimmunoassay was carried out as previously described (13) using rabbit anti-human TGF-8 IgG. The reaction mixture (0.2 ml) contained 2 pl of rabbit anti-human TGF-8 IgG solution, 0.1 mg of acid-treated azM prepared from human plasma or standard TGF-8 (0-10 ng) in 10 mM sodium phosphate buffer (pH 7.4), 0.1% Tween 80, 0.5 M NaCI, 0.01% sodium azide, and 0.1% human serum albumin. After standing overnight a t 22 "C, 0.1 ng of '251-TGF-@ in 0.1 ml of the same phosphate buffer was added. T w o hours later, 50 pl of 10% IgGsorh was added. The mixture was incubated for an additional 2 h and centrifuged. The pellets were then washed three times with the Same phosphate buffer and counted with a y-counter. Nonspecific binding was measured with a 100-fold excess of unlabeled TGF-8 or in the absence of anti-TGF-8 IgG. Assays were done in quadruplicate. The azM samples were preincu- bated at pH 3.0 for 1 h. The pH of the azM solutions was then adjusted back to 7.4.
RESULTS
In earlier studies (13, 23), we found that aZM is the major plasma protein which binds PDGF. Since PDGF and TGF-8 are both composed of two polypeptides and are similar with respect to platelet localization, the high content of disulfide linkages, and molecular weight (1-4, 24-26), we investigated the possibility that aZM also serves as a binding protein for TGF-/3 in serum. lZ5I-TGF-/3 prepared by iodination of TGF-
with Na'T in the presence of chloramine T showed a single band in the autoradiogram following SDS-polyacrylamide gel electrophoresis (Fig. 1). lz51-TGF-/3 was incubated with aZM at room temperature for 20 min, and the reaction mixture was then subjected to 5% native polyacrylamide gel electropho- resis. This electrophoresis system has been used to analyze the formation of complexes between azM and several pro- teases or PDGF (27,28).* In this system, azM - PDGF complex or aZM-protease complexes migrate in the separating gel (27, 28).' Free PDGF and proteases either do not migrate into the separating gel, due to their basic nature, or migrate at the dye front, due to their small molecular sizes. As shown in Fig. 2, '2sI-TGF-/3-a2M complex migrates at a position identical to that of the fast migrating form of aZM (lam I). "'1-TGF-8 alone did not migrate into the separating gel ( l a n e 2). The radioactive band at the dye front is human serum albumin which may have been iodinated by trace amounts of free '"I during acidic storage conditions. It is known that proteins can be iodinated by traces of free IZ5I in the presence of oxygen under acidic conditions. This result suggests that Iz5I-TGF-8
* S. S. Huang, P. O'Grady, T. J. Olsen, and J. S. Huang, submitted for publication.
C -
d- I-TGFf3
e - f -
dye front- FIG. 1. Autoradiograph of '261-TGF-8 following 16% SDS-
polyacrylamide gel electrophoresis. lZI-TGF-8 was prepared as described under "Experimental Procedures." One ng of '251-TGF-8 was subjected to 15% SDS-polyacrylamide gel electrophoresis fol- lowed by autoradiography. The protein markers were: bovine serum albumin, 66 kDa (a); glyceraldehyde-3-phosphate dehydrogenase, 36 kDa (b) ; carbonic anhydrase, 29 kDa (c); trypsinogen, 24 kDa (d); soybean trypsin inhibitor; 20 kDa (e); and a-lactalbumin, 14 kDa 0.
I 2 FIG. 2. Autoradiograph of a reaction mixture containing
'261-TGF-~ and human a2M following 5% native polyacryl- amide gel electrophoresis. The reaction mixture contained 1 ng of '2sI-TGF-8 in the presence and absence of 10 pg of a2M in 0.1 ml of 0.1 M sodium phosphate buffer, pH 7.0, containingO.l% human serum albumin. After 20 min at 22 'C, an aliquot (15 pl) was subjected to 5% native polyacrylamide gel electrophoresis. The fast form of a2M (methylamine-modified azM) was used as the marker in this system. The arrow indicates the location of both '[email protected] complex and the fast form of azM: 1, "'1-TGF-8 + a2M 2, "I-TGF-8 alone.
interacts with aZM to form a complex. To further examine whether aZM is the binding protein for
"'I-TGF-/3 in serum, "'1-TGF-8 (1 ng) was incubated with 2 pl of human serum in 0.1 ml of 5 mM HEPES buffer, pH 7.4, 0.15 M NaCl, for 20 min. The mixture was then subjected to immunoprecipitation with anti-human aZM antiserum or nonimmune serum. As shown in Fig. 3, anti-human aZM antiserum specifically immunoprecipitated aZM in human

The Latent Form of Transforming Growth Factor 1537
A. B.
q2M+ s' " +1251 -TGFp- q2M complex
-205 kDa-
dye front -- a b a b
FIG. 3. Coomassie Brilliant Blue staining pattern (A) and autoradiograph (B) of immunoprecipitates of 'z61-TGF-@. azM complex in human serum with a polyclonal rabbit anti-human a z M antiserum. Two pl of human serum was incubated with 1 ng of "'I-TGF-@ in 100 pl of 5 mM HEPES buffer, pH 7.4, 0.15 M NaCI. After 30 min at 22 'C, the reaction mixture was incubated with 5 pl of rabbit anti-human azM antiserum (lanes a ) or non-immune serum (lanes b). Eighteen hours later, the immunocomplexes were absorbed by 50 pl of 10% IgGsorb and subjected to 5% SDS-polyacrylamide gel electrophoresis, followed by Coomassie Brilliant Blue staining (A ) and autoradiography (B). The arrows indicate the location of azM (A) and 12'[email protected] complex ( B ) .
serum, as demonstrated by Coomassie Brilliant Blue staining (Fig. 3A, lane a). The anti-a2M antiserum also immunopre- cipitated a high molecular weight form of '2'I-TGF-/3 as dem- onstrated by autoradiography (Fig. 3B, lane a). In contrast, neither a2M nor the high molecular weight form of "'1-TGF- /3 was seen in the immunoprecipitate using nonimmune serum (Fig. 3, A and B, lane b ) . This suggests that '2'I-TGF-/3 and mzM form a complex which is immunoprecipitated by the anti- azM antiserum. The fact that at least a portion of the complex (-10%) did not dissociate during SDS-polyacrylamide gel electrophoresis indicates a covalent interaction of the com- ponents. The specific interaction of '"II-TGF-/3 with a,M in human serum was further supported by the observation that '''I-TGF-/3 was unable to form a complex in human serum in which azM had been depleted by anti-apM IgG-Sepharose gels (data not shown).
The inhibition of complex formation between '"I-PDGF and azM by sulfhydryl agents and the dissociation of "'1- PDGF . a2M complex by reducing agents led us to suggest that '"I-PDGF forms a covalent complex with azM through a disulfide-sulfhydryl exchange reaction (23). In order to inves- tigate whether '2'I-TGF-/3 forms a covalent complex with azM through a similar mechanism, we examined the effects of sulfhydryl-reactive agents on complex formation between '%I- TGF-/3 and a2M. No inhibition of complex formation was observed in the presence of 5 mM sulfhydryl-reactive agents such as iodoacetamide, p-chloromercuribenzoate, N-ethyl- maleimide, and 5,5'-dithiobis-(2-nitrobenzoic acid) (data not shown). These results suggest that the nature of the covalent interaction between "'I-TGF-/3 and asM may be different from that between "'I-PDGF and azM.
We recently found that modification of aZM with methyl- amine or trypsin diminishes the binding of azM with 9 - PDGF.2 To further compare the interactions of a2M with '9- TGF-8 and '"I-PDGF, we investigated the effects of trypsin or methylamine modification of azM on complex formation with 12'I-TGF-/3. As in the case of the '2'I-PDGF-a2M inter- action, modification of a2M with trypsin diminished the abil- ity of azM to bind '''I-TCF-/3 (Fig. 4, lane 2). Methylamine modification of a2M did not inhibit its ability to form a complex with '2'I-TGF-/3 but rather enhanced it as shown by native polyacrylamide gel electrophoresis (Fig. 5A) and SDS-
1251-TGFp-+ I q 2M complex - - d y e front
1 2 3 4 FIG. 4. Effects of trypsin modification on complex forma-
tion by a z M with 1261-TGF-fl. Ten pg of aZM was treated with or without trypsin (1 pg) in the presence and absence of soybean trypsin inhibitor (1 pg) in 0.1 ml of 0.1 M sodium phosphate buffer, pH 7.0. After 1 h at 37 "C, 1 gg of soybean trypsin inhibitor was added to stop the reaction. One ng of "'I-TGF-@ was then added to the reaction mixtures. Twenty min later, an aliquot was then subjected to 5% native polyacrylamide gel electrophoresis. The fast form of azM was used as the marker. 1 , azM + "'I-TGF-@; 2, aZM + trypsin + soybean trypsin inhibitor + 1261-TGF-@; the factors were added sequentially; 3, aZM + soybean trypsin inhibitor + trypsin + 12sI-TGF-@, soybean trypsin inhibitor was added at zero time; 4, azM + soybean trypsin inhibitor + '*'I-TGF-@.
a b a b
FIG. 5. Effect of methylamine modification on complex for- mation of azM with '261-TGF-@. Ten pg of azM was incubated with or without methylamine (200 mM) in 0.1 ml of 25 mM Tris-HCI, pH 8.0, for 1 h at 37 "C. One ng of "'I-TGF-@ was then added to the reaction mixtures. Twenty min later a t 22 "C, an aliquot was subjected to 5% native polyacrylamide gel electrophoresis (A) and 5% SDS- polyacrylamide gel electrophoresis (B) , followed by autoradiography. The arrows indicate the location of the 12'[email protected] complex. The locations of the slow and fast forms of azM in native polyacryl- amide gel electrophoresis are indicated by s and j , respectively. a, methylamine-modified azM + "'I-TGF-@; b, azM + 1251-TGF-@.
polyacrylamide gel electrophoresis (Fig. 5B). Methylamine not only increased the binding 3-4-fold, as demonstrated by native polyacrylamide gel electrophoresis (Fig. 5A, lane a), but also increased the amount of the covalent complex (2-3- fold), as shown by SDS-polyacrylamide gel electrophoresis (Fig. 5B, lane a). A portion of the complex of '2'I-TGF-/3 and azM appeared to be resistant to reduction with mercaptoeth- anol. When complexes of '''I-TGF-/3 with either azM or a2M modified by methylamine were reduced at 100 "C and sub- jected to SDS-polyacrylamide gel electrophoresis and auto- radiography, an M , -195,000 radioactive band was observed (Fig. 6). This M, -195,000 '2sI-labeled protein is presumably the conjugate of the subunit (Mr -12,500) of '2'I-TGF-/3 and azM monomer (MI -180,000). These results clearly indicate that the covalent bond in the '2sII-TGF-/3.a2M complex is distinct from the disulfide linkage observed in the '"I-PDGF. a2M complex (23).
The '2sI-TGF-/3-azM dimer complex had an apparent mo- lecular weight of 310,000 in nonreducing SDS-polyacrylamide

1538
205 kDa - l l6kDa - 97kDa - 66kDa -
The Latent Form of Transforming Growth Factor j3
c ca 2M complex -
I 2 3 FIG. 6. AutoradioaraDhs of '*%TGF-B - aaM comdex fol-
lowing reducing SD&&yacrylamide gel elktrophoresis. An aliquot of reaction mixtures containing azM or methylamine-modified azM and '2sI-TGF-p, which were prepared as described in the legend of Fig. 5, was subjected to 5% reducing SDS-polyacrylamide gel electrophoresis, followed by autoradiography. 1, methylamine-modi- fied azM + L251-TGF-p; 2, a2M + "'I-TGF-@; 3, '2sI-TGF-@ alone.
A. B.
IZ5I-PDGF-q2M+ WM c/ '251-lGFg-a2M cornplea complex I
. 1251-TGFp-a2M complex
'251-PDGF-hL e o m d e x
1 2
FIG. 7. Autoradiographs of '*'I-TGF-fl-apM complex and '*%PDGF. alM complex following native polyacrylamide gel electrophoresis. One ng of lZ5I-TGF-p or 30 ng of '"I-PDGF was incubated with 10 pg of human a2M (A) or with 10 pl of human serum ( B ) in 0.1 M sodium phosphate buffer, pH 7.0. After 20 min at 22 "C, an aliquot of the reaction mixture was then subjected to 5% native polyacrylamide gel electrophoresis, followed by autoradiography. A: 1, T - P D G F + azM; 2, '2sI-TGF-@ + aZM. B 1, '"I-PDGF + human serum; 2, "'I-TGF-@ + human serum. The arrows indicate the h a - tion of the '"I-PDGF-azM complex and the '"I-TGF-p. azM com- plex.
gel electrophoresis (Figs. 3B and 5B, lane a). Under reducing conditions, the molecular weight of the complex was 195,000 (Fig. 6, lanes 1 and 2). Under similar nonreducing and reduc- ing conditions, azM shows apparent molecular weights of 290,000 and 180,000, respectively (23). Based on the apparent molecular weights of '2'I-TGF-/3~a2M dimer (MI -310,000), a2M dimer (Mr -290,000), and TGF-/3 (M, -25,000), we estimated that the '251-TGF-/3. azM binary complex consists of equimolar "51-TGF-/3 and azM dimer. The 195-kDa lz5I- TGF-8. a2M complex seen under reducing conditions presum- ably is composed of a 'z'I-TGF-/3 subunit (M. -12,500) and the a2M monomer (Mr -180,000).
As previously noted, the '*'II-TGF-/3. a2M complex migrates at a position identical to that of the fast form of azM in native polyacrylamide gel electrophoresis. The PDGF - a2M complex migrates at a position intermediate between the slow and fast forms of a2M.' In addition, methylamine modification in- creases the ability of a2M to bind '"II-TGF-/3 while it decreases its ability to bind PDGF.' These results suggest that the reactive form of azM for "51-TGF-#? binding may be different from that for '"I-PDGF binding. To further define the migra- tion difference, '251-TGF-/3- azM and ""I-PDGF - azM com- plexes were simultaneously run in native polyacrylamide gel electrophoresis. As shown in Fig. 7, the ""I-TGF-/3- azM com- plex migrated a little faster than the ""I-PDGF a azM complex when complexes were formed with purified azM (Fig. 7 A ) or
"
1 2 3 4 5 FIG. 8. Dissociation of '*"I-TGF-8.azM complex by 0.5 M
acetic acid and 4 M urea. The reaction mixture contained 10 pg of azM and 1 ng of '251-TGF-p in 50 pl of 0.1 M sodium phosphate buffer, pH 7.0. After 20 min at 22 "C, an equal volume of 1 M acetic acid ( l a n e 2), 8 M urea (lane 3), or 0.1 M sodium phosphate buffer, pH 7.0 ( l a n e 4 ) was added to the reaction mixture. Thirty min later, the reaction mixture was dialyzed against 0.1 M sodium phosphate buffer, pH 7.0, to remove acetic acid and urea. An aliquot of the reaction mixture was then subjected to 5% native polyacrylamide gel electro- phoresis and autoradiography. Lane 1 represents a2M incubated at 22 "C with '=I-TGF-p, but not subjected to dialysis. Lane 5 represents '2"I-TGF-j3 alone. The arrom indicates the location of the "'1-TGF- @.azM complex and the fast form of azM. The bar indicates the location of azM dimer detected by Coomassie Brilliant Blue staining. atM tetramer was irreversibly converted to azM dimer by acid or urea treatment.
with a2M in human serum (Fig. 7B). These results suggest that the product of the '2'I-TGF-/3 and a2M interaction is different from that of the "'I-PDGF and azM interaction.
Pircher et al. (12) recently demonstrated that the high molecular weight latent form of TGF-/3 can be converted to the active form of TGF-/3 with an apparent molecular weight of -20,000 by acid or urea treatment. If the latent form of TGF-/3 is a TGF-/3 - a2M complex, the "'I-TGF-/3 - azM com- plex should show similar sensitivity to acid and urea treat- ment. To test this, we incubated "'I-TGF-/3 with a2M to form a complex and then subjected the complex to acid and urea treatment. As shown in Fig. 8, most of the '251-TGF-/3.azM complex could be dissociated by acid or urea treatment while -10% remained associated, migrating a t a position identical to that of azM dimer in native polyacrylamide gel electropho- resis. The azM tetramer was irreversibly converted to the dimer by acid or urea treatment. The radioactivity at the top of the separating gel (Fig. 8) came from free '2'I-TGF-/3 which stayed in the stacking gel throughout electrophoresis, due to its slightly basic nature. These results are consistent with the [email protected] complex being the latent form of TGF-/3 in serum.
To further prove that the TGF-8. azM complex is the latent form of TGF-/3, we purified azM from freeze-thawed human platelet-rich plasma (40-50-ml volume/unit) which contains both a2M and the latent form of TGF-8 (2,12). If the latent form of TGF-/3 is in fact the TGF-8. aZM complex, we should be able to demonstrate TGF-/3 activity in these a2M prepa- rations following acidification. As shown in Fig. 9A, fractions 94-116 of the gel filtration contained azM as indicated by 5% SDS-polyacrylamide gel electrophoresis followed by Coo- massie Brilliant Blue staining (Fig. 9B) and trypsin inhibitor activity assay (data not shown). Without acidification, these azM-containing fractions did not exhibit the TGF-/3 activity. However, after acidification, these fractions showed signifi- cant TGF-/3 activity. This result suggests that azM prepared from human platelet-rich plasma contains the latent form of TGF-8. This suggestion is further supported by the observa-

The Latent Form of Transforming Growth Factor /3 1539
Frc. 9. Co-chromatography of asM and a latent TGF-8 activity on Ultrogel AcA34/Ultrogel AcA22 (2:1, v/v). Outdated human platelet- rich plasma (3 units) was frozen and thawed. After centrifugation to remove cell debris, 100 ml of the supernatant was precipitated at 5.5-12.5% (w/v) pol- yethylene glycol 6000 according to Bar- rett's method (18). The precipitates were dissolved in 20 ml of 0.1 M sodium phos- phate buffer, pH 7.0, and the solution was applied to a column (5.0 X 72 cm) of Ultrogel AcA34/Ultrogel AcA22 (2:1, v/ v) and eluted with the same buffer. The flow rate and fraction volume were 20 ml/h and 3 ml, respectively. The protein concentrations of fractions were moni- tored by absorbance at 280 nm ( A ) . Frac- tions containing a2M, 94-116, were iden- tified by 5% SDS-polyacrylamide gel electrophoresis followed by Coomassie Brilliant Blue staining ( B ) and trypsin inhibitor activity (data not shown). The purity of azM in these fractions was >98%. The TGF-,!I activity (anchorage- independent cell growth-promoting ac- tivity) of the fractions 86-130 and pooled fractions 134-188 was assayed before and after acidification. In the acidifica- tion, 1 ml of each fraction was adjusted to pH 3.0 with 6 N HCI and kept a t room temperature for 1 h. The pH of the so- lution was then adjusted back to 7.0 with 0.2 N NaOH. Each fraction (with or without acidification) was sterilized by filtration. Ten pl of the sterilized solu- tions was then subjected to TGF-B activ- ity assay as indicated by colony number formation by NRK cells (A) .
1.8 - A
- o..oTransformlng Assay (acidified) ~-~TransformingAssay (non-acidlfied)
1.4 -
1.2 -
10-
0.8 -
0.6 -
0.4 - "0
0.2 -
0 " 1
3'80 too 120 140 160 180 200 Fraction Number
6. 130 120 116 112 108 102 98 94 86
205 k o a - .
116 koa-
97 koa-
66 koa -
tion that about 15 +. 5 ng of TGF-j3 was found in 1 mg of acid-treated a2M prepared by zinc chelate chromatography (17) from human platelet-rich plasma using a radioimmuno- assay for human TGF-j3 (Table I). The standard curve of this radioimmunoassay is shown in Fig. 10. It is noteworthy that no significant quantity of TGF-j3 antigen was found in a2M prepared from platelet-poor plasma, suggesting that TGF-j3 is derived from platelets. All of these observations clearly sug- gest that TGF-8. a2M complex is a latent form of TGF-j3.
DISCUSSION
Using native polyacrylamide gel electrophoresis and SDS- polyacrylamide gel electrophoresis we have demonstrated that purified a2M forms a complex with lZ5I-TGF-j3. We have also shown that when lZ6I-TGF-j3 is incubated with human serum, a complex is formed which may be immunoprecipitated by anti-human a2M antiserum. These results suggest that a2M is a binding protein for TGF-j3 in serum and that a TGF-8. a2M complex may be the latent form of TGF-8 previously described in serum (12). These suggestions have been sup- ported by the following observations. 1) The '2sI-TGF-j3. azM
complex can be dissociated by acid or urea treatment; 2) a2M prepared from platelet-rich plasma shows TGF-j3 activity whereas no TGF-8 activity is found in a2M prepared from platelet-poor plasma; 3) the high molecular weight latent form of TGF-j3 has been previously converted to the active form of TGF-8 by acid or urea treatment (12). Recently, Lyons et al. (30) found an inactive form of TGF-j3 in the conditioned media of NRK-49F and HT-1080 cells. This inactive form (Mr -62,000) of TGF-j3 was converted to the active form of TGF-j3 by treatment with plasmin, but not by acid or urea. It is evident that this inactive form of TGF-j3 is not similar to the latent form of TGF-j3 in human serum or platelets (12).
Assay of the transforming growth factor activity of TGF-j3 is usually performed in the presence of 510% fetal calf serum. Exogenous TGF-j3 added would be expected to form a complex with a2M in the serum. TGF-j3.a2M complex as the latent form of TGF-j3 raises an important question: how does exog- enous TGF-j3 express a potent transforming growth factor activity in the presence of a2M? Active TGF-j3 has been purified under relatively harsh conditions with treatments such as methanol HCl extraction and chromatography in

1540 The Latent Form of Transforming Growth Factor p TABLE I
TGF-8 contents in a,M prepared from human platelet-rich plasma and platelet-poor plasma
TGF-8 content"
w l m g 1. apM prepared from platelet-rich 15 & 5 (n = 5)
plasmab
2. apM prepared from platelet-poor co.1 (n = 3) Dlasmab a The TGF-@ contents in a2M preparations were measured as
described under "Experimental Procedures" using a polyclonal rabbit anti-TGF-8 IgG.
a2M were prepared from freeze-thawed human platelet-rich plasma and platelet-poor plasma by zinc chelate chromatography (17). The homogeneity of purified azM was judged by SDS-polyacryl- amide gel electrophoresis (5 and 7.5% gels) followed by silver staining. The platelet factor 4 contents in these platelet-poor plasmas were less than 50 ng/ml. The volume for 1 unit of platelet-rich plasma was 40-50 ml.
1 I I I I I 1 1 1 1 1 1 1 I
0.1 I IO
TGFB (ng)
FIG. 10. Standard curve for the radioimmunoassay with rabbit anti-TGF-8 IgG. The procedure is described under "Exper- imental Procedures." Assays were carried out in quadruplicate. Each point represents the mean * S.E. of measurements obtained from six different experiments.
strong acids or urea solutions (2, 31,32). It is reasonable that the ability of TGF-P to bind azM may have been partially inactivated during its purification. This has been supported by the observation that only -15% of '251-TGF-P was able to form a complex with azM even in the presence of >300 molar excess of The -85% free TGF-P should be able to stimulate cell growth in the presence of 10% fetal calf serum.
The lZ51-TGF-P. azM complex migrates at a position iden- tical to that of the fast form of a2M in native polyacrylamide gel electrophoresis. Since azM is a very large molecule (tetra- mer, M, -720,000), complex formation with lZ5I-TGF-P may not alter the electrophoretic migration of azM. It has been shown that azM-protease complexes migrate at the same position as the fast form of azM, regardless of the type of protease or their acidic or basic nature (27, 28). I t is possible that the reactive form of azM which binds lZ5I-TGF-P is the fast form of azM. This possibility was supported by the observation that methylamine modification, which increased the ability of azM to form a complex with lZ51-TGF-P, con- verted a2M to the fast form. However, the fast form produced by modification of aZM with trypsin appears to be inactive in forming a complex with '251-TGF-P. This suggests that the fast forms produced by methylamine and trypsin modification are different with respect to their binding activity for lz5I-
S. S. Huang, and J. S. Huang, unpublished results.
TGF-P. These results also suggest that TGF-@-apM interac- tion is distinct from protease-azM interaction. It has been shown that modification of aZM with methylamine abolishes its binding to proteases (27, 28).
The majority of purified azM migrated as the slow form in native polyacrylamide gel electrophoresis. If the reactive form of aZM for lZ5I-TGF-P is the fast migrating form, this reactive form should be a minor species in purified a2M preparations. Our observations seem to indicate that the reactive form of aZM is the fast migrating form. However, we cannot com- pletely rule out the possibility that the reactive form of azM and the "51-TGF-P. aZM complex migrate at different posi- tions.
A portion of the lZ5I-TGF-P. aZM complex is resistant to heating at 100 "C in the presence of reducing agent followed by 5% SDS-polyacrylamide gel electrophoresis, suggesting that in at least a portion of the complex molecules of lZ5I- TGF-P and azM are covalently linked. This linkage is distinct from the disulfide linkage in the '251-PDGF.azM complex (23). The chemical nature of the covalent linkage in the lZ5I- TGF-P-azM complex is not known. One of the possibilities is that the putative thioester linkage of azM may be involved in the formation of a covalent bond between Iz5I-TGF-P and azM. However, the increase in covalent complex formation between '251-TGF-P and azM following methylamine modifi- cation of the azM strongly argues against this possibility. Methylamine modification is thought to inactivate the puta- tive thioester linkages, resulting in the loss of covalent binding ability of a2M for protease (33-36). It has also been reported that the covalent binding property of lZ5I-epidermal growth factor to its receptor is a useful artifact of chloramine-T treatment (37). Since '251-TGF-(3 was also prepared by the chloramine-T-oxidation procedure, it is likely that the cova- lent binding property of '251-TGF-/3 is generated by the same mechanism. It is noteworthy that both covalent linkages of lZ5I-TGF-@. azM and lZ5I-epidermal growth factor - receptor complexes are resistant to reduction with reducing agents.
The interaction of IZ5I-TGF-P with azM is different from that of lz5I-PDGF with azM in several aspects. 1) 9 - T G F - P. aZM complex migrates faster than the '251-PDGF. a2M com- plex in native polyacrylamide gel electrophoresis. 2) Methyl- amine modification diminishes the ability of azM to bind lz5I- PDGF but enhances its ability to form a complex with lz5I- TGF-P. 3) A portion of '251-TGF-/3-azM complex appears to have a covalent bond which is distinct from the disulfide linkage in the '251-PDGF. azM complex. Taken together, these results suggest different reactive forms of a2M for lZ5I-TGF-P and for lZ5I-PDGF binding.
The physiological relevance of the TGF-P azM complex is not known. We have suggested that complex formation be- tween PDGF and azM may be a mechanism for clearing excess PDGF released from activated platelets during blood clotting and vessel injury (23). Since TGF-P and PDGF are simulta- neously released from activated platelets, azM may also play such a role in the clearance of excess TGF-P. The TGF-P. aZM complex may possibly be cleared from the plasma by the same mechanism as the fast form of azM (38-41).
Acknowledgments-We thank Dr. Anita B. Roberts, Laboratory of Chemoprevention, National Cancer Institute, for the gift of TGF-8; Dr. Jorma Keski-Oja and Dr. Harold L. Moses, Vanderbilt University, for gifts of rabbit anti-TGF-P IgG; Dr. John A. McDonald, Washing- ton University School of Medicine, for gifts of human fibronectin; Dr. William S. Sly and Dr. Catherine M. Nolan for a critical review of the manuscript; Beth Parker for her excellent technical assistance; and Peggy Kerner and Kimberly Short for preparing the manuscript.
Addendum-After this paper was submitted, Fava and McClure

The Latent Form of Tran (29) reported that some but not all commercial preparations of human fibronectin are contaminated with TGF-j3 activity. This observation raises the important question of whether human fibronectin also can serve as a binding protein for TGF-j3. To test this possibility, we have used native gel electrophoresis and gel filtration on Bio-Gel A-1.5m (13) to study the interaction between lZ5I-TGF-j3 and fibronectin. No complex formation was observed at concentrations of fibronectin up to 200 pg/ml. This result is consistent with an interaction of high specificity between '261-TGF-,3 and a2M in serum or plasma.
REFERENCES
1. Derynck, R., Jarrett, J. A., Chen, E. Y., Eaton, D. H., Bell, J. R., Assoian, R. K., Roberts, A. B., Sporn, M. B., and Goeddel, D. V. (1985) Nature 3 1 6 , 701-705
2. Assoian, R. K., Komoriya, A,, Meyers, C. A., Miller, D. M., and Sporn, M. B. (1983) J. Biol. Chem. 258 , 7155-7160
3. Sporn, M. B., and Roberts, A. B. (1985) Nature 313,745-747 4. Goustin, A. S., Leof, E. B., Shipley, G. D., and Moses, H. L.
(1986) Cancer Res. 46,1015-1029 5. Roberts, A. B., Anzano, M. A., Lamb, L. C., Smith, J. M., and
Sporn, M. B. (1981) Proc. Natl. Acad. Sci. U. S. A. 7 8 , 5339- 5343
6. Moses, H. L., Branum, E. L., Proper, J. A., and Robinson, R. A. (1981) Cancer Res. 4 1,2842-2848
7. Tucker, R. F., Shipley, G. D., Moses, H. L., and Holley, R. W. (1984) Science 2 2 6 , 705-707
8. Roberts, A. B., Anzano, M. A., Lamb, L. C., Smith, J. M., and Sporn, M. B. (1985) Proc. Natl. Acad. Sei. U. S. A. 8 2 , 119- 123
9. Ignotz, R. A., and Massague, J. (1986) J. Bid. Chem. 261,4337- 4345
10. Roberts, A. B., Sporn, M. B., Assoian, R. K., Smith, J. M., Roche, N. S., Wakefield, L. M., Heine, U. I., Liotta, L. A., Falanga, V., Kehrl, J. H., and Fauci, A. S. (1986) Proc. Natl. Acad. Sci. U. S. A. 83,4167-4171
11. Masui, T., Wakefield, L. M., Lechner, J. F., LaVeck, M. A., Sporn, M. B., and Harris, C. C. (1986) Proc. Natl. Acad. Sci. U. S. A. 83,2438-2442
12. Pircher, R., Jullien, P., and Lawrence, D. A. (1986) Biochem. Biophys. Res. Commun. 136,30-37
13. Huang, J. S., Huang, S. S., and Deuel, T. F. (1983) J. Cell Biol.
14. Frolik, C. A,, Wakefield, L. M., Smith, D. M., and Sporn, M. B.
15. DeLarco, J . E., and Todaro, G. J. (1978) Proc. Natl. Acad. Sci. U.
16. Huang, J . S., Huang, S. S., Kennedy, B., and Deuel, T. F. (1982)
97,383-388
(1984) J. Biol. Chem. 2 5 9 , 10995-11000
S. A. 76,4001-4005
sforming Growth Factor /3 1541
J. Biol. Chem. 257,8130-8136
Biochem. 99,415-420 17. Kurecki, T., Kress, L. F., and Laskowski, M., Sr. (1979) Anal.
18. Barrett, A. J. (1981) Methods Enzymol. 9 0 , 737-754 19. Garrott, P. 0. (1966) Clin. Chim. Acta 14,493-501 20. Laemmli, U. K. (1970) Nature 227,680-685 21. Van Leuven, F., Cassiman, J.-J., and Van den Berghe, H. (1981)
22. Huang, S. S., Kuo, M.-D., and Huang, J. S. (1986) Biochem.
23. Huang, J. S., Huang, S. S., and Deuel, T. F. (1984) Proc. Natl.
24. Heldin, C.-H., and Westermark, B. (1984) Cell 37, 9-20 25. Deuel, T. F., and Huang, J. S. (1984) J. Clin. Invest. 74,669-676 26. Ross, R., Raines, E. W., and Bowen-Pope, D. F. (1986) Cell 4 5 ,
27. Gonias, S. L., Reynolds, J. A., and Pizzo, S. V. (1982) Biochim.
28. Straight, D. L., and McKee, P. A. (1982) Biochemistry 21,4550-
29. Fava, R. A., and McClure, D. B. (1987) J. Cell. Physiol. 131 ,
30. Lyons, R. M., Keski-Oja, J., and Moses, H. L. (1986) J. Cell Biol. 103, 443a (abstr.)
31. Roberts, A. B., Anzano, M. A,, Meyers, C. A., Wideman, J., Blacher, R., Pan, Y.-C-E., Stein, S., Lehrman, R., Smith, J. M., Lamb, L. C., and Sporn, M. B. (1983) Biochemistry 2 2 ,
32. Frolik, C. A., Dart, L. L., Meyers, C. A., Smith, D. M., and Sporn,
33. Howard, J. B. (1981) Proc. Natl. Acad. Sci. U. S. A. 78, 2235-
34. Sottrup-Jensen, L., Petersen, T. E., and Magnusson, S. (1980)
35. Van Leuven, T., Cassiman, J.-J., and Van den Berghe, H. (1981)
36. Salvesen, G. S., and Barrett, A. J. (1980) Biochem. J. 187, 695-
37. Comens, P. G., Simmer, R. F., and Baker, J. B. (1982) J. Biol.
38. Imber, M. J., and Pizzo, S. V. (1981) J. Biol. Chem. 256 , 8134-
39. Ohlsson, K. (1971) Acta Physiol. Scad. 81, 269-272 40. Feldman, S. R., Rosenberg, M. R., Ney, K. A., Michalepoulos, G.,
and Pizzo, S. V. (1985) Biochem. Biophys. Res. Commun. 128,
41. Marynen, P., Van Leuven, F., Cassiman, J.-J., and Van den
J. Biol. Chem. 266,9016-9022
Biophys. Res. Commun. 139,619-625
Acad. Sci. U. S. A. 8 1 , 342-346
155-169
Biophys. Acta 704,306-314
4556
184-189
5692-5698
M. B. (1983) Proc. Natl. Acad. Sci. U. S. A. 80, 3676-3680
2239
FEBS Lett. 121,275-279
J. Bid. Chem. 256,9023-9027
701
Chem. 257,42-45
8139
795-802
Berghe, H. (1984) J. Biol. Chem. 2 5 9 , 7075-7079