nybelinia queenslandensis sp. n. (cestoda: … · 295 nybelinia queenslandensis sp. n. (cestoda:...
TRANSCRIPT

295
Nybelinia queenslandensis sp. n. (Cestoda: Trypanorhyncha) parasitic in Carcharhinus melanopterus, from Australia, with observations on the fine structure of the scolex including the rhyncheal system
Malcolm K. Jones1and Ian Beveridge2
1University of Queensland, St Lucia, Queensland 4072, Australia; 2University of Melbourne, Parkville, Victoria 3052, Australia
Key words: Cestoda, Trypanorhyncha, Tentaculariidae, Nybelinia, morphology, scolex, ultrastructure, tentacles, taxonomy
Abstract. A new species of Nybelinia Poche, 1926, N. queenslandensis sp. n. (Cestoda: Trypanorhyncha) is described from sharks, Carcharhinus melanopterus (Quoy et Gaimard, 1824) from the coast of northeastern Queensland, Australia. Morphological features of the 46 known species of Nybelinia Poche, 1926 are tabulated and the new species is differentiated from all known taxa that are adequately described on the basis of having a homeomorphous armature, metabasal hooks 20-25 µm long, tentacles 0.07-0.09 mm in diameter, short bulbs (0.38-0.45 mm) and craspedote segments with the testes encircling the female genital complex. The fine structure of the scolex microtriches, frontal and rhynchodeal glands, tentacles and hooks, sheath and retractor muscle is described and compared with that of other trypanorhynchs.
The trypanorhynch cestode genus Nybelinia Poche, 1926 is a large, cosmopolitan genus with most species restricted to a specific geographic region of the world (Dollfus 1942, 1960), although recent collections suggest that some species may be more widely distributed than previously thought (Palm et al. 1998).
The occurrence of this genus in the Australian region has not been investigated in detail, with records being limited to N. thyrsites Korotaeva, 1971, a species apparently limited to southern Australia and New Zealand coastal waters (Beveridge and Campbell 1996) and N. sphyrnae Yamaguti, 1952, a species previously known only from Sphyrna zygaena (Linnaeus, 1758) from Japan (Yamaguti 1952) having recently been reported from the same host species off the southern coast of Australia (Beveridge and Campbell 1996).
Recent examinations of sharks from the northeastern coast of Queensland revealed the presence of a species of Nybelinia which apparently cannot be accommodated within any of the existing taxa. The new species is described in this paper together with observations on its fine structure.
Transmission electron microscopic studies of the ultrastructural features of trypanorhynch cestodes are few and are largely restricted to investigations of the rhyncheal system (Beveridge and Smith 1988), musculature, including that of the rhyncheal system (Ward et al. 1986), tegument (Palm et al. 1998) and glands occurring in the scolex (Davydov and Biserova 1985). Scanning electron microscopic studies of the
tegument and microtriches are more extensive and include the examination of numerous genera (for example see Richmond and Caira 1991, Palm 1995). The opportunity is therefore taken to provide additional information on the fine structure of the tegument, scolex glands and rhyncheal system of Nybelinia. This paper contains the first transmission electron microscopic study of a tentaculariid trypanorhynch.
MATERIALS AND METHODS
Fixation Cestodes collected from the intestines of C. melanopterus
were subdivided into several groups. Some specimens were relaxed in tap water then fixed in AFA solution (Pritchard and Kruse 1982). Other specimens, including some still attached to the gut wall were fixed rapidly in 10% neutral buffered formalin or 3% glutaraldehyde in cacodylate buffer. For scanning electron microscopy (SEM), scoleces were fixed in 3% glutaraldehyde in 0.1M cacodylate buffer. For transmis-sion electron microscopy (TEM), specimens were fixed either detached or in situ with 3% glutaraldehyde in 0.1M cacodylate buffer at pH 7.2.
Light Microscopy One scolex was mounted in Berlese fluid to allow
examination of the hooks. Whole mounts were prepared by staining in Celestine blue, dehydrating in ethanol, clearing in methyl salicylate and mounting in Canada balsam. Serial sections of scoleces and mature segments in transverse and longitudinal planes were prepared by embedding in paraffin, cutting at a thickness of 5 µm and staining the sections in
Address for correspondence: M.K. Jones, Centre for Microscopy and Microanalysis, The University of Queensland, Queensland 4072, Australia. Phone: ++61 7 3365 3249; Fax: ++61 7 3365 2199; E-mail: [email protected]
FOLIA PARASITOLOGICA 45: 295-311, 1998

296
haematoxylin and eosin. Drawings were made with a drawing tube attached to an Olympus BH-2 microscope fitted with differential interference contrast optics. Measurements were made with an ocular micrometer and, unless otherwise stated, are presented in millimetres as the range, followed by the mean of five measurements in parentheses. All whole-mounted specimens have been deposited in the South Australian Museum, Adelaide (SAM). Host nomenclature follows Last and Stevens (1994).
Electron Microscopy Specimens for SEM were rinsed in buffer, postfixed in 1%
aqueous osmium tetroxide, dehydrated in ethanol and dried overnight in hexamethyldisilasane (ProSciTech, Townsville, Australia). Dried samples were mounted on aluminium stubs, sputter coated with gold and examined in a JSM 6300F at an accelerating voltage of 5 kV. Specimens for TEM were rinsed in buffer, post-fixed in 1% aqueous osmium tetroxide, dehydrated in ethanol and acetone, and embedded in Spurr resin (ProSciTech, Townsville, Australia). Semithin (1-2 µm) sections were cut with glass knives, mounted on slides and stained with 1% toluidine blue in 1% aqueous sodium tetraborate. Ultrathin sections were stained with uranyl acetate and lead citrate and examined with a JEM-1210 or JEM-1010 at an accelerating voltage of 80 kV.
RESULTS
Nybelinia queenslandensis sp. n. Figs. 1-13 Material studied: From Carcharhinus melanopterus
(Quoy et Gaimard, 1824): two specimens, Mackay, Queensland, coll. B. G. Robertson, 27. viii. 1985, holotype SAM 28016, paratype SAM 28017; 10 specimens, 1 mounted in Berlese fluid (SAM 28018-9); 4 stained (SAM 28020-5), 5 scoleces sectioned (SAM 28026) from Heron Island, Queensland, coll. M. Jones and I. Beveridge, 1. x. 1993.
Description. Small cestodes, largest specimen 28 long, maximum width 0.90. Scolex craspedote 0.97-1.20 (1.10) long. Pars bothridialis 0.59-0.75 (0.68), width across bothridia 0.64-0.80 (0.74); 4 elongate bothridia, margins thick, covered with prominent microtriches, refractile in whole mounts; centre of bothridia slender, cuneiform; bothridia extend almost to mid-region of bulbs. Pars vaginalis 0.38-0.61 (0.54), shorter than pars bothridialis; tentacular orifices at apex of each bothridium; anterior regions of sheaths straight, posterior regions slightly sinuous; bulbs short, 0.34-0.58 (0.45) long by 0.09-0.13 (0.11) wide; retractor muscle originates at base of bulb; prebulbar organ and gland cells within bulb absent; pars post-bulbosa prominent, about 0.06 long; robust bundles of external muscles radiate from bases of bulbs to posterior end of pedunculus scolecis; velum prominent, 0.12-0.22 (0.18) long. Frontal glands prominent in whole mounts as large, single cells scattered in four groups between bothridia on dorsal, ventral and lateral aspects of medulla; greatest concentrations of frontal glands anterior, restricted to anterior region of pars bothridialis.
Rhynchodeal glands prominent, medial to sheaths, immediately anterior to bulbs; ducts of rhynchodeal glands extend anteriorly on medial aspects of sheaths to level of tentacle orifices, openings to surface not detected.
Tentacles short, 0.27 when fully everted, basal swelling absent, 0.07-0.08 (0.08) in diameter at base, not diminishing in diameter in distal tentacular region, 0.08-0.09 (0.08). Hooks arranged in about 25 rows. Armature homeomorphous, homeoacanthous; hooks solid. Hooks arranged in continuous ascending spirals; approximately 14 files of hooks on tentacle. Hooks broadly falcate with rounded bases, bases highly refractile; hook length in mid-tentacular region 0.020-0.025 (0.023), base 0.012-0.015 (0.013). Hooks smaller at base increasing in size towards metabasal region; individual hook lengths (L) and lengths of bases (B) for hooks on single tentacle: row 1, L = 0.010, B = 0.005; row 3, L = 0.012, B = 0.009; row 5, L = 0.013, B = 0.012; row 7, L = 0.014, B = 0.012; row 9, L = 0.015, B = 0.012; row 11, L = 0.016, B = 0.014; row 13, L = 0.018, B = 0.015; row 15, L = 0.020, B = 0.015. At base, on bothridial, antibothridial and internal surfaces of tentacle, hooks extend to very base of tentacle; external surface of base with hooks forming small inverted V-shaped area free of hooks at very base of tentacle.
Mature segments craspedote; velum difficult to detect in first formed segments; not detectable until about segment 12; fully developed velum present by about segment 20. Mature segments wider than long, 0.30-0.51 (0.38) long by 0.67-0.90 (0.80) wide. Genital atrium inconspicuous, ventro-submarginal, in anterior half of segment; genital pores alternate irregularly. Genital ducts cross osmoregulatory canals ventrally. Cirrus sac elongate, directed anteromedially from genital atrium, sac thin-walled; cirrus unarmed coiled within sac, proximal coils of greater diameter, but not forming distinct internal seminal vesicle; external seminal vesicle absent; vas deferens coils medially to mid-line then posteriorly towards female genital complex. Vasa efferentia not seen. Testes ovoid, 0.06-0.09 (0.07) in diameter, arranged in single layer in medulla; testes number 74-80 (77) per proglottis; testes encircle female genital complex, occupy entire medulla except for region of female genital complex and immediately anterior to it, and in vicinity of cirrus sac. Vagina tubiform, lies posterior and dorsal to cirrus sac; diminutive vaginal sphincter detectable in transverse sections, not visible in whole mounts; terminal region of vagina dilated; vagina follows arcuate course medially to female genital complex without dilating to form seminal receptacle. Ovary almost centrally situated, 4 lobed, each lobe c. 0.15 by 0.10; Mehlis’ gland posterior to ovary, c. 0.10 in diameter. Uterine duct coils anteriorly from Mehlis’ gland to enter uterus;

Jones, Beveridge: Nybelinia queenslandensis sp n..
297
Fig. 1-4. Nybelinia queenslandensis sp. n. Scolex and tentacles. Fig. 1. Scolex, dorso-ventral view. Fig. 2. Tentacle, bothridial surface, external face on left hand side. Fig. 3. Basal region of tentacle, external surface. Fig. 4. Profiles of individual hooks belonging to a single file on the bothridial surface of the tentacle showing change in shape from row 1, at the base, to row 23 at the tip of the tentacle. Scale bars = 0.1mm (Fig. 1) and 0.01mm (Figs. 2-4).
uterus initially sacciform with two lateral branches; branches fill with eggs, spread posterolaterally, eventually converge and unite posterior to Mehlis’ gland. Gravid segments c. 0.70 by 1.75. No fully formed eggs present. Vitelline follicles circum-medullary, individual follicles 0.02-0.03 (0.03) in diameter. Longitudinal musculature weakly developed, arranged in approximately 30 bundles. Ventral osmoregulatory canals paired, c. 0.02 in diameter, at lateral extremities of medulla; 6 dorsal canals present, c.
0.005 in diameter, spread evenly across dorsal surface of medulla.
Ultrastructure Tegument. The scolex of N. queenslandensis is lined
by a thin syncytial tegument (Figs. 14, 15, 18-21). Filamentous microtriches, 1.7 µm in length are evident across the entire surface of the scolex (Figs. 20-22). Along the bothridial margins, however, the filamentous microtriches are interspersed among a distinct band of stout microtriches (Figs. 15, 18, 21-23, 35-38). The

298
Figs. 5-7. Nybelinia queenslandensis sp. n. Transverse histological sections through scolex. Fig. 5. Section at anterior end of bothridia, through cerebral ganglia, showing frontal glands. Fig. 6. Section near posterior end of pars vaginalis showing rhynchodeal glands. Fig. 7. Section through bulbs at posterior end of bothridia. All sections oriented with dorsal aspect uppermost. Scale bar = 0.1mm. Abbreviations: fg - frontal glands, g - ganglion, n - nerve trunk, o - osmoregulatory canal, rg - rhynchodeal gland, sh - sheath.
bothridial margin, the microtriches appear hook-like (Figs. 36, 37). Distally, the microtriches of the margin are longer and appear spine-like (Figs. 36, 38). Both forms of stout microtriches have an expanded cap consisting of an external plasma membrane enclosing a distinct cortex surrounding a medulla (Figs. 21-23) but lack a distinct basal region and junctional tubule (Figs. 21-23) evident in filamentous microtriches (Fig. 22, and Holy and Oaks 1986). The stout microtriches are anchored in small depressions of the apical cytoplasm (Fig. 21). The medullary regions of the caps are rich in canaliculi, which have an external diameter of 8 nm (Figs. 22, 23). At the tentacular orifice, the apical cytoplasm is characterised by short filamentous microtriches that are pressed against the surface of the tegument (Fig. 27). Membranous microlaminations occupy the space between the tegument and the tentacle (Fig. 27).
The contents of the apical cytoplasm of the tegument varies among different region of the scolex. The prominent vesicles of the bothridial apical cytoplasm are spherical and electron-lucent (Fig. 21). In non-bothridial regions, discoid vesicles with a moderately electron-opaque matrix (Fig. 20) predominate and are interspersed with spherical electron-opaque vesicles (Fig. 20). At the tentacular orifice, the apical cytoplasm is thicker and is rich in ovate electron-lucent vesicles (Fig. 27).
Sub-tegumental musculature is smooth. In non bothridial regions, the sub-tegumental musculature is a thin layer with few myofibrils and myocytons evident (Figs. 15, 20). A basal matrix is evident beneath the apical cytoplasm. The musculature of the bothridia
forms a dense band beneath the apical cytoplasm (Figs. 15, 21). The basal matrix underlying this region, however, is poorly developed.
Scolex glands. Rhynchodeal glands are evident as four strands of minute cells running longitudinally near the inner surfaces of the sheaths (Figs. 1, 6, 16). The plasma membrane of the rhynchodeal glands cells is strongly invaginated. The invaginations pass almost to the nucleus, giving the cells a stellate appearance (Fig. 24). The nucleus is rich in euchromatin and possesses a thin peripheral band of heterochromatin. The cells possess a prominent nucleolus. The cytoplasm is rich in rough endoplasmic reticulum, Golgi bodies and secretory vesicles (Fig. 24). Endoplasmic reticulum is observed frequently near the periphery of cells. Secretory vesicles are spherical to ovate, and contain a coarsely granular electron-opaque material. Openings of rhynchodeal glands were not observed by light or electron microscopy. However, granular vesicles of similar shape and size to those of the rhynchodeal glands were observed in specialized secretory pockets in tentacles (Fig. 33).
The frontal glands are unicellular. Each flask-shaped cell contains a small nucleus rich in euchromatin (Fig. 25). The cells contain peripheral arrays of an expanded rough endoplasmic reticulum and mitochondria. Secretory vesicles are spherical, and contain an electron-opaque material with a homogeneous matrix. Frontal glands open to the surface by neck processes that pass through the tegument (Fig. 26). Neck processes are characterised by a peripheral array of microtubules.

Jones, Beveridge: Nybelinia queenslandensis sp n..
299
Figs. 8-13. Nybelinia queenslandensis sp. n. Mature and gravid segments. Fig. 8. Cirrus sac. Fig. 9. Mature segment, dorsal view. Fig. 10. Transverse section of mature segment, dorsal surface uppermost. Fig. 11. Gravid segment. Fig. 12. Sagittal section through gravid segment showing displacement of testes to the dorsal side of the medulla by the developing uterus. Fig. 13. Junctions of segments of same strobila showing increase in size of velum. a - junction of segments 11 and 12, b - junction of segments 13 and 14, c - junction of segments 16 and 17, d - junction of segments 20 and 21. (Scale bars = 0.1mm). Abbreviations: do - dorsal osmoregulatory canals, s - sphincter, t - testis, u - uterus, v - vagina, vi - vitelline follicle, vo - ventral osmoregulatory canal.

300
Figs. 14-19. Nybelinia queenslandensis sp. n., semithin sections, light microscopy. Fig. 14. Scolex, transverse section in situ. Fig. 15. Scolex, transverse section, in situ. Stout microtriches (arrowhead) are evident along the margins of bothridia. Fig. 16. Tentacular sheaths and rhynchodeal glands. Note the retractor muscle (asterisk). Fig. 17. Everted tentacle in situ. Fig. 18. Bothridium, transverse section, surface in situ. Fig. 19. Non-bothridial surface, transverse section, in situ. [Scale bars = 0.1 mm (Fig. 14) and 0.01 mm (Figs. 15-19)]. Abbreviations: B - bothridium, H - host tissue, N - nerve, R - rhynchodeal gland, S - sheath.

Jones, Beveridge: Nybelinia queenslandensis sp n..
301
Figs. 20-23. Nybelinia queenslandensis sp. n. Scolex tegument, TEM. Fig. 20. Nonbothridial surface. Fig. 21. Bothridial surface. Stout microtriches emerge from slight invaginations of the apical cytoplasm (arrowhead). Fig. 22. Microtriches of bothridial margin. A junctional tubule in a filamentous microthrix is indicated (arrowhead). Fig. 23. Cap of stout microthrix of margins. Scale bars = 1 µm (Figs. 20, 21) and 500 nm (Figs. 22, 23). Abbreviations: A - apical cytoplasm, b - base of filamentous microthrix, BM - basal matrix, C - cortex of microthrix cap, F - filamentous microtriches, M - myofibrils, Me - medulla of microthrix cap, SM - stout microthrix.

302
Figs. 24-25. Nybelinia queenslandensis sp. n., scolex glands, TEM. Fig. 24. Rhynchodeal gland cells. Note Golgi body, peripheral endoplasmic reticulum (arrowheads) and invaginations of the cytoplasm (asterisk). Fig. 25. Frontal glands. Scale bars = 100 nm (Fig. 24) and 1 µm (Fig. 25). Abbreviations: G - Golgi body.

Jones, Beveridge: Nybelinia queenslandensis sp n..
303
Figs. 26-29. Nybelinia queenslandensis sp. n., wall of tentacles and sheaths, TEM. Fig. 26. Tentacular orifice. Note the zone of homogenous fibrils (arrowheads) and elaborated membranes of the tentacular epithelium (asterisk). Fig. 27. Tentacular orifice. Flattened microtriches (arrowheads) are evident along the apical cytoplasm of the tegument. The trilaminar cortex lines the tentacle externally (asterisk). Fig. 28. Tentacular hook. Fibrils radiate from the base of the hook (arrowhead). The trilaminar cortex (asterisk) does not continue around the hook. Fig. 29. Sheath wall. The sheath wall merges with the parenchyma through a loose array of fibrils (arrowheads). Scale bars = 10µm (Fig. 26) and 500 nm (Figs. 27-29). Abbreviations: A - apical cytoplasm, B - blade of hook, E - epithelium of tentacles and sheath, H - tentacular hook, Fg - frontal gland, Fl - tentacular fluid, M - myofibrils, ML - microlaminations, R - retractor muscle, S - sheath wall, Te - tentacular wall.
304
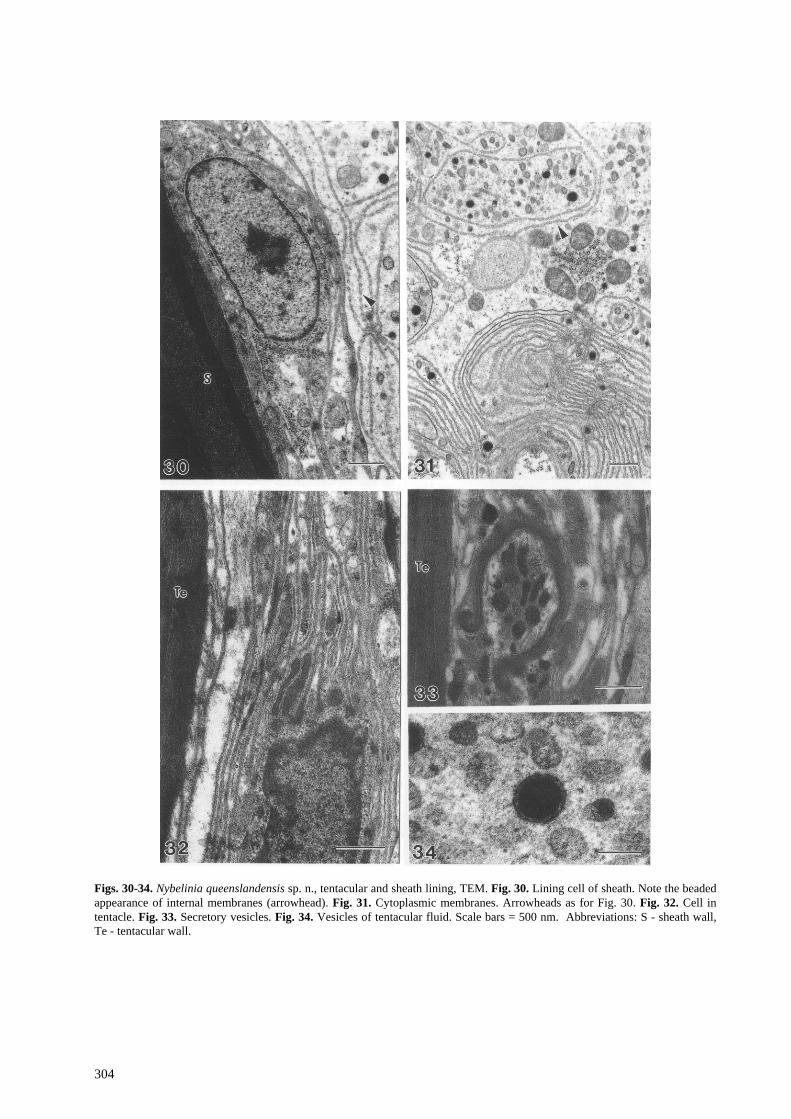
Figs. 30-34. Nybelinia queenslandensis sp. n., tentacular and sheath lining, TEM. Fig. 30. Lining cell of sheath. Note the beaded appearance of internal membranes (arrowhead). Fig. 31. Cytoplasmic membranes. Arrowheads as for Fig. 30. Fig. 32. Cell in tentacle. Fig. 33. Secretory vesicles. Fig. 34. Vesicles of tentacular fluid. Scale bars = 500 nm. Abbreviations: S - sheath wall, Te - tentacular wall.

Jones, Beveridge: Nybelinia queenslandensis sp n..
305
Figs. 35-38. Nybelinia queenslandensis sp. n., scolex, SEM. Fig. 35. Scolex. Fig. 36. Bothridium. Note the thin band of stout microtriches along the margin. Enlargement of region A is shown in Fig. 37 and region B in Fig. 38. Fig. 37. Details of hook-like microtriches. Fig. 38. Details of spine-like microtriches. Scale bars = 0.1 mm (Figs. 35, 36) and 10 µm (Figs. 37, 38).

306
Rhyncheal system. The tentacle sheath is fibrillar and composed of five to six layers of fibrils in an highly ordered array (Fig. 29). The orientation of the fibrils varies among the layers (Fig. 29). The sheath lacks a distinct outer margin, and the outermost layers of the sheath appear as a loose array of fibrils (Fig. 29) that merges with the surrounding parenchyma. The inner margin of the sheath is lined by a thin epithelium that is continuous with that of the tentacle and which surrounds the tentacular fluid (Figs 29, 30).
The laminated appearance of the sheath wall disappear at the tentacular orifice, and is replaced, for a short distance, by a single homogenous band of fibrils (Fig. 26). The tentacle wall extends distally from this short homogenous region, and consists of as many as 15-20 layers of fibrils (Figs. 26, 27). The orientation of the fibrils varies among the layers so that adjacent layers differ in electron contrast (Figs. 26, 27). The laminae of the tentacular wall disappears at the point of insertion of tentacular hooks (Fig. 28). The base of each hook appears to be anchored by a thin layer of fibrils that arise perpendicularly from the base (Fig. 28). Externally, the tentacular wall is lined by a thin tri-laminate cortex (Fig. 27). This structure resembles a unit membrane in possessing electron-opaque laminae separated by an electron-lucent region, but is thicker and lines a fibrillar matrix. The tri-laminate cortex of the tentacle wall does not extend to cover the hook (Fig. 28). Tentacular hooks are solid and have a fine granular matrix (Fig. 28). The blade contains a granular matrix that is more electron-lucent than that of the base (Fig. 28).
The sheath and tentacles are lined internally with a thin cellular lining (Figs. 26-33). The cells of this lining (Figs. 30, 32) are elongate, and have elongate nuclei with a prominent nucleolus. Membranes are prominent in the cytoplasm and occur in two main forms. The first type are thickened membranes that pass longitudinally along the cytoplasm, dividing it and organelles into strands (Figs. 30, 32). The second type are thin membranes elaborated by the cells into the tentacular fluid (Figs. 26, 30, 31). These membranes follow complex paths and form numerous whorls and laminae (Figs. 26, 30, 31). These membranes may be pinched off repeatedly, giving them a beaded appearance (Figs. 30-31). Numerous membrane-bound vesicles are evident in the tentacular fluid (Figs. 26, 31, 34) and appear to be of predominantly two kinds. The first kind of vesicle is spherical and contains a distinct electron-opaque core (Fig. 34). Numerous electron-lucent vesicles surround the electron-opaque vesicles (Fig. 34). These vesicles are irregular in shape and enclose a finely granular material (Fig. 34), and may be degradative products of the opaque vesicles.
Small aggregations of secretory vesicles are evident in pockets lined by a fibrous material beneath the
tentacular wall (Fig. 33). The vesicles are spherical to ovate and have a granular matrix (Fig. 33), resembling the vesicles synthesised by rhynchodeal glands.
The retractor muscle is a cylindrical band of musculature that extends from the bulb to the apex of the tentacle. The retractor muscle consists of numerous myofibrils surrounded and separated from each other by thin interstitial matrices (Fig. 28). Myofibrils are smooth, and contain thick and thin myofilaments, peripheral hemidesmosomes, and mitochondria.
Host-parasite interface. The scolex of Nybelinia queenslandensis is embedded in the mucosa of the pyloric stomach with the tapeworm body lying against the mucosal surface. The scolex displays numerous folds in the surface (Fig. 14). The margin of a bothridium is easily identifiable by the occurrence of stout microtriches (Figs. 15, 18). Bothridial and non-bothridial surfaces may be applied to the mucosal lining by the tapeworm (Figs. 18, 19). The bothridia did not appear to grasp the tissues of the host (Fig. 14). Tentacles did not appear to enter submucosal tissues but maintained contact with the mucosal surface (Fig. 17). The hooks penetrated apical regions of mucosal cells (Fig. 17). The scolex and tentacles caused severe desquamation of the mucosal lining (Figs. 14, 17-19) and the scolex in particular was surrounded by a channel of sloughed tissues.
DISCUSSION
Systematics A total of 46 species have been described to date in
the genus Nybelinia. Dollfus (1942) summarised information on the 14 species known at the time and subsequently (Dollfus 1960) erected 16 new species mainly from fish from the west coast of Africa. Subsequently, Yamaguti (1952) added four new species from fish off the coast of Japan, Heinz and Dailey (1974) and Carvajal et al. (1976) added species from California and Hawaii respectively while an additional eight species have been described from the Indian Ocean (Shah and Bilquees 1979, Reimer 1980, Chandra and Rao 1985, Chandra 1988, Kurshid and Bilquees 1988, Palm et al. 1998) and one species from China (Yang et al. 1995). One species from southern Australian coastal waters, N. thyrsites Korotaeva, 1971, was recently described in full (Beveridge and Campbell 1996).
Several of the descriptions of species are extremely poor and a number were regarded as species inquirendae (Dollfus 1942). Dollfus (1960) divided his new species into groups based on the diameter of the tentacles, the possession of a distinctive basal armature, whether or not hooks on the antibothridial surface of the tentacle were more slender than on the bothridial surface and the length to width ratio of the tentacular bulbs. While this is a workable sub-division for African

Jones, Beveridge: Nybelinia queenslandensis sp n..
307
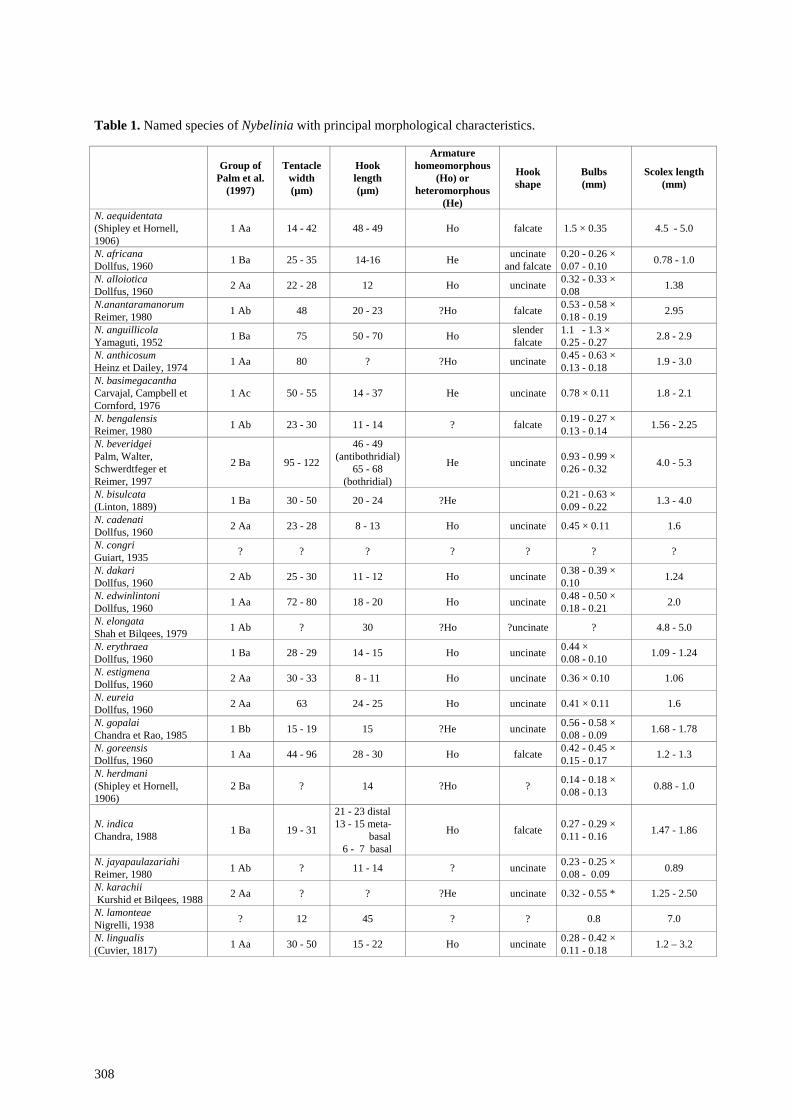
species, the requisite information is not available for many of the remaining species. The information which can be obtained from the literature is summarised in Table 1.
Palm et al. (1997) divided species into several groups based on (a) the metabasal armature being homeo-morphous or heteromorphous, (b) relative sizes of basal and metabasal hooks and (c) possession of a basal armature. The allocation of these subgroups has been indicated in Table 1. The species described above is characterised by a homeomorphous armature, a tentacle diameter of 70-90 µm and falcate hooks 20-25 µm long. Species resembling it are N. anantaramanorum Reimer, 1980, N. edwinlintoni Dollfus, 1960, N. eureia Dollfus, 1960, N. bisulcata (Linton, 1889) and N. sphyrnae Yamaguti, 1952. The material from Queensland is readily distinguishable from N. sphyrnae by the larger number of files of hooks (6 per side in N. sphyrnae, 8 in N. queenslandensis) and in the morphology of the mature segment in which the testes are distributed in two lateral groups in N. sphyrnae (see Beveridge and Campbell 1996, fig. 17) but encircle the female genitalia in the new species. N. queenslandensis differs from N. anantaramanorum in most measurements with the pars bothridialis 1.38 mm in N. anantaramanorum compared with 0.59-0.75 mm in N. queenslandensis, bulbs 0.53-0.58 by 0.18-0.19 mm for the former species compared with 0.34-0.58 by 0.09-0.13 mm in the latter and in the shape of the hooks which are much more slender and curve more sharply at the tip in N. anantaramanorum. N. queenslandensis resembles N. edwinlintoni, the description of which is based on a single post-larva from Sphyrna diplana (= S. lewini) from Senegal. N. edwinlintoni is a larger species, with a longer pars bothridialis (1.2 mm), bulbs which are much wider (0.48-0.50 by 0.18-0.21 mm) and hooks whose length (18-20 µm) lies just below the range of that of N. queenslandensis (20-25 µm). N. eureia can be distinguished from N. queenslandensis in the fact that in N. eureia, the pars bothridialis extends to the posterior ends of the bulbs and the hooks on one side of the tentacle are more sharply recurved than are hooks on the opposite surface of the tentacle whereas in N. queenslandensis, all hooks in the metabasal region are of a similar shape.
N. queenslandensis also resembles N. bisulcata but has more robust tentacles (30-50 µm in diameter in N. bisulcata cited by Dollfus (1942), 18-26 µm in the re-description by Sao Clemente and Gomes (1992), compared with 70-90 µm in N. queenslandensis) and the segments of N. bisulcata are acraspedote (Dollfus 1942, Sao Clemente and Gomes 1992) whereas those of N. queenslandensis are craspedote, though this character has to be assessed in fully mature segments as the velum appears to increase in length as the immature segments develop (Fig. 13). The various descriptions of
the armature of N. bisulcata are not entirely concordant (Sao Clemente and Gomes 1992). Hook sizes given by Sao Clemente and Gomes are 10 µm and uncinate at the base, 13 µm in the metabasal region and 18 µm near the apex of the tentacle. Dollfus (1942), summarising the results of Linton (1889), gave measurements of 14 µm at the base and 23 µm in the metabasal region, while Stunkard (1977) gave 18-21 µm as the lengths of the hooks. Linton (1897 - Plate LXVI, fig. 13) illustrated hooks of this species from opposite sides of the tentacle as differing in shape, in a fashion similar to those of “Section V” described by Dollfus (1960) (N. dakari, N. estigmena, N. punctatissima). However, this feature has not been commented on by subsequent authors. If this feature was correctly described by Linton (1897) then it provides an additional feature for separating N. queenslandensis from N. bisulcata.
The erection of a new species for the material described from Queensland is therefore considered justified, since it can be distinguished from those species which have been adequately described. The status of species such as N. lamonteae, N. congri, N. narinari, N. karachii, and N. indica cannot be determined at present.
Ultrastructure Frontal glands. Some features of the morphology of
N. queenslandensis warrant comment. Frontal glands are clearly evident in this species, disposed between the bothridia and appearing to be unicellular and emptying through the tegument to the surface of the cestode. Frontal glands were first described in N. syngenes by Pintner (1929) and may exist in other species of Nybelinia as well. The ultrastructure of frontal glands in Grillotia erinaceus has been described (Davydov and Biserova 1985). They considered them to be modified tegumentary cytons with 2-3 nuclei and 1-6 nucleoli which were apocrine in function and connected to the tegument by several processes. In the structure and mode of secretion of the secretory product, the frontal glands of N. queenslandensis appear to be eccrine glands (Kuperman 1982, Žďárská and Nebesářová 1997), and resemble scolex glands of the tetra-phyllidean cestode Trilocularia acanthiaevulgaris, described by McCullough and Fairweather (1989). In that species, the secretions of scolex glands are thought to be a glycoprotein and may function in adhesion of the parasite.
Rhynchodeal glands. As in other species of Nybelinia, N. queenslandensis possesses prominent rhynchodeal glands, lying immediately anterior and medial to the bulbs, with ducts leading to the tegument in the vicinity of the tentacular orifices. The histology of the rhynchodeal glands was described by Pintner (1899, 1929) for Nybelinia. It was not possible to determine where the product of rhynchodeal glands was
308
Table 1. Named species of Nybelinia with principal morphological characteristics.
Group of Palm et al.
(1997)
Tentacle width (µm)
Hook length (µm)
Armature homeomorphous
(Ho) or heteromorphous
(He)
Hook shape
Bulbs (mm)
Scolex length (mm)
N. aequidentata (Shipley et Hornell, 1906)
1 Aa 14 - 42 48 - 49 Ho falcate 1.5 × 0.35 4.5 - 5.0
N. africana Dollfus, 1960 1 Ba 25 - 35 14-16 He uncinate
and falcate0.20 - 0.26 × 0.07 - 0.10 0.78 - 1.0
N. alloiotica Dollfus, 1960 2 Aa 22 - 28 12 Ho uncinate 0.32 - 0.33 ×
0.08 1.38
N.anantaramanorum Reimer, 1980 1 Ab 48 20 - 23 ?Ho falcate 0.53 - 0.58 ×
0.18 - 0.19 2.95
N. anguillicola Yamaguti, 1952 1 Ba 75 50 - 70 Ho slender
falcate 1.1 - 1.3 × 0.25 - 0.27 2.8 - 2.9
N. anthicosum Heinz et Dailey, 1974 1 Aa 80 ? ?Ho uncinate 0.45 - 0.63 ×
0.13 - 0.18 1.9 - 3.0
N. basimegacantha Carvajal, Campbell et Cornford, 1976
1 Ac 50 - 55 14 - 37 He uncinate 0.78 × 0.11 1.8 - 2.1
N. bengalensis Reimer, 1980 1 Ab 23 - 30 11 - 14 ? falcate 0.19 - 0.27 ×
0.13 - 0.14 1.56 - 2.25
N. beveridgei Palm, Walter, Schwerdtfeger et Reimer, 1997
2 Ba 95 - 122
46 - 49 (antibothridial)
65 - 68 (bothridial)
He uncinate 0.93 - 0.99 × 0.26 - 0.32 4.0 - 5.3
N. bisulcata (Linton, 1889) 1 Ba 30 - 50 20 - 24 ?He 0.21 - 0.63 ×
0.09 - 0.22 1.3 - 4.0
N. cadenati Dollfus, 1960 2 Aa 23 - 28 8 - 13 Ho uncinate 0.45 × 0.11 1.6
N. congri Guiart, 1935 ? ? ? ? ? ? ?
N. dakari Dollfus, 1960 2 Ab 25 - 30 11 - 12 Ho uncinate 0.38 - 0.39 ×
0.10 1.24
N. edwinlintoni Dollfus, 1960 1 Aa 72 - 80 18 - 20 Ho uncinate 0.48 - 0.50 ×
0.18 - 0.21 2.0
N. elongata Shah et Bilqees, 1979 1 Ab ? 30 ?Ho ?uncinate ? 4.8 - 5.0
N. erythraea Dollfus, 1960 1 Ba 28 - 29 14 - 15 Ho uncinate 0.44 ×
0.08 - 0.10 1.09 - 1.24
N. estigmena Dollfus, 1960 2 Aa 30 - 33 8 - 11 Ho uncinate 0.36 × 0.10 1.06
N. eureia Dollfus, 1960 2 Aa 63 24 - 25 Ho uncinate 0.41 × 0.11 1.6
N. gopalai Chandra et Rao, 1985 1 Bb 15 - 19 15 ?He uncinate 0.56 - 0.58 ×
0.08 - 0.09 1.68 - 1.78
N. goreensis Dollfus, 1960 1 Aa 44 - 96 28 - 30 Ho falcate 0.42 - 0.45 ×
0.15 - 0.17 1.2 - 1.3
N. herdmani (Shipley et Hornell, 1906)
2 Ba ? 14 ?Ho ? 0.14 - 0.18 × 0.08 - 0.13 0.88 - 1.0
N. indica Chandra, 1988 1 Ba 19 - 31
21 - 23 distal 13 - 15 meta- basal 6 - 7 basal
Ho falcate 0.27 - 0.29 × 0.11 - 0.16 1.47 - 1.86
N. jayapaulazariahi Reimer, 1980 1 Ab ? 11 - 14 ? uncinate 0.23 - 0.25 ×
0.08 - 0.09 0.89
N. karachii Kurshid et Bilqees, 1988 2 Aa ? ? ?He uncinate 0.32 - 0.55 * 1.25 - 2.50
N. lamonteae Nigrelli, 1938 ? 12 45 ? ? 0.8 7.0
N. lingualis (Cuvier, 1817) 1 Aa 30 - 50 15 - 22 Ho uncinate 0.28 - 0.42 ×
0.11 - 0.18 1.2 – 3.2

Jones, Beveridge: Nybelinia queenslandensis sp n..
309
Table 1. (continued)
Group of Palm et al.
(1997)
Tentacle width (µm)
Hook length
(µm)
Armature homeomorphous
(Ho) or heteromorphous
(He)
Hook shape
Bulbs (mm)
Scolex length (mm)
N. manazo Yamaguti, 1952 1 Ba 35 15 - 18 ? ? 0.23 - 0.25 ×
0.08 - 0.11 0.95 – 1.05
N. narinari (MacCallum, 1917) ? ? ? ? ? - 1.0
N. nipponica Yamaguti, 1952 2 Ba 30 - 60 30 - 38 Ho slender
uncinate 0.31 - 0.35 × 0.08 - 0.12 1.4 - 2.9
N. oodes Dollfus, 1960 1 Ab 24 - 27 9 - 11 Ho uncinate 0.30 × 0.10 0.92
N. palliata (Linton, 1924) 1 Aa 27 - 30 19 - 21 Ho ? 0.35 - 0.60 ×
0.14 - 0.15 1.3 - 2.5
N. perideraeus (Shipley et Hornell, 1906)
1 Ba 70 - 80 8 - 16 He uncinate and falcate 0.47 1.1 - 1.3
N. pintneri Yamaguti, 1934 1 Ab ? 15 ?Ho ? 0.38 - 0.41 ×
0.15 - 0.20 1.54
N. punctatissima Dollfus, 1960 2 Aa 25 - 35 9 - 11 Ho uncinate 0.39 × 0.11 1.24
N. rhynchobatus Yang, Lin, Liu et Peng, 1995 1 Ab ? 11 ?He ? 0.15 - 0.18 ×
0.06 - 0.09 0.81 - 0.85
N. riseri Dollfus, 1960 1 Aa 35 11 - 12 Ho uncinate 0.37 × 0.12 1.62
N. robusta (Linton, 1890) 1 Ba 15 - 27 11 - 15 Ho ? 0.27 - 0.33 ×
0.08 - 0.10 0.91 - 1.2
N. rougetcampanae Dollfus, 1960 2 Ba 40 - 46 21 - 26 He falcate 0.31 - 0.32 ×
0.08 1.22
N. senegalensis Dollfus, 1960 2 Aa 18 - 32 7 - 9 Ho uncinate 0.40 × 0.10 1.8
N. sphyrnae Yamaguti, 1952 1 Aa 34 14 - 21 Ho falcate 0.33 ×
0.09 - 0.15 0.8 - 1.0
N. strongyla Dollfus, 1960 1 Ab 55 19 - 20 Ho uncinate 0.61 × 0.19 2.3
N. surmenicola (Okada, 1929) 1 Ab 100 - 110 30 - 32 Ho uncinate 0.90 - 0.95 ×
0.35 - 0.37 5.6
N. syngenes (Pintner, 1928) 1 Ab 90 - 100 60 - 67 Ho falcate 1.0 × 0.2 2.3 - 2.5
N. tenuis (Linton, 1890) 1 Ab 20 5 ?Ho 0.09 - 0.14 ×
0.03 - 0.05 0.48 - 0.86
N. thyrsites Korotaeva, 1972 1 Aa 40 - 96 16 - 20 Ho falcate 0.31 - 0.40 ×
0.10 - 0.16 ?
N. yamagutii Dollfus, 1960 2 Ba 40 - 115 64 - 70 He falcate 0.96 - 1.01 ×
0.18 2.2 - 2.5
* Length of pars bulbosa
released, although vesicles similar to those of the rhynchodeal glands were observed in small pockets beneath the tentacular walls (Fig. 33). Pintner (1899) described the ducts of the rhynchodeal glands as running within the wall of the tentacular sheath and opening onto the external surface. He (Pintner 1899) noted that the secretion accumulated in the rhynchodeal cavity formed by the invaginated tentacle and that it might function as a lubricant. In the case of N. syngenes, Pintner (1929) suggested that the glands may also empty into the internal cavity of the tentacle. Further studies are required to clarify the nature of the secretions of rhynchodeal glands.
Rhyncheal system. The fine structure of the rhyncheal system of N. queenslandensis resembles that described for Trimacracanthus aetobatidis (Robinson, 1959) Beveridge et Campbell, 1987 by Beveridge and Smyth (1988). The hooks are solid, embedded into the tentacle wall and the tentacle lacks an apical cytoplasm with microtriches, and the musculature of the bulb is striated, in contrast with the other muscles of the scolex and strobila. Because the hooks are embedded in a fibrous matrix and not in syncytial epidermis, it seems most likely that they have not arisen from microtriches, as is the case for the hooks of cyclophyllidean cestodes (Mount 1970). The principal difference noted in N.

310
queenslandensis is the cellular lining of the sheath and tentacles with the occurrence of membranes forming whorls or laminate patterns. While morphologically striking, the functions of these cells are not known. We suggest, however, that the cells supply components for the tentacular fluid or wall.
The tentacular walls of N. queenslandensis consist of numerous laminae of fibrils. Tentacular walls in T. aetobatidis are similarly fibrillar, but lack the distinctive banding seen in N. queenslandensis, have fewer fibrils and incorporate a vesicular layer (Beveridge and Smith 1988). The sheath and tentacular walls of N. queenslandensis are highly ordered and it is of interest to determine how such organised fibrillar matrices are formed. Beveridge and Smith (1988) argued that the walls of the tentacles of T. aetobatidis were analogous to the basal laminae of tegumentary epithelia and possibly formed by an elaboration of the basal matrix of the tegument. The wall of the sheath and tentacles of N. queenslandensis differ from each other in the numbers of fibrous lamellae. The two regions are separated by a short zone lacking a laminated appearance. These observations suggest that the tentacles are not formed by outgrowth of the sheath wall during ontogeny of the worm but may develop from structurally similar but regionally distinct cells.
Microtriches. Modification of microthrix structure in the regions of the bothridia have been observed in a number of studies. Palmate microtriches occur, e.g., in the genera Callitetrarhynchus (see Carvajal et al. 1987), Floriceps (see Andersen 1987), Grillotia (see Halton and McKerr 1979, Whittaker et al. 1982), Gilquinia (see McCullough and Fairweather 1983) and Otobothrium (see Shields 1985), while enlarged, blade-like or hook-like microtriches have been observed in N. alloiotica Dollfus, 1960, Nybelinia sp. cf lingualis and
N. edwin-lintoni by Palm (Palm 1995, 1997). The microtriches of N. queenslandensis resemble those of other tentaculariid species studied so far.
Filamentous microtriches are the predominant form of microthrix on the scolex of N. queenslandensis. Modified microtriches in this species occur only in a thin band at the margins of the bothridia. When viewed by SEM, the band is distinctly different from surrounding surfaces. Yet, TEM or light microscopy reveals little specialization of the apical cytoplasm of margins relative to surrounding regions of the apical cytoplasm. The apical cytoplasm of the margins are in cytoplasmic continuity with the rest of the syncytial tegument. It would be of interest, then, to determine how such marked specialization is maintained within the syncytial tegument.
The bothridia of N. queenslandensis are plainly evident in living worms and whole mounts, but are difficult to discern in transverse section. During attachment, the bothridia do not grasp onto the mucosal surface as appears to be the case for those of other trypanorhynchs and tetraphyllideans (Borucinska and Caira 1993). It is possible then that the scolex of N. queenslandensis maintains attachment primarily by means of the tentacles and hooks, aided by surface area interactions between the scolex surface and the mucosa. The stout microtriches may interdigitate with the mucosal surface to hold the scolex in place.
Acknowledgments. Collection of material described herein was supported financially by the Australian Biological Resources Study. Bruce Robertson is thanked for excellent technical assistance. The authors are grateful to Dr. B. Diggles for providing a specimen of Carcharinus melanopterus and to Dr. Deborah Stenzel, whose late-night provision of EM fixatives made the ultrastructural part of this study possible.
REFERENCES
ANDERSEN K. 1987: S.E.M. observations on plerocercus larvae of Floriceps saccatus Cuvier, 1817 and Molicola horridus (Goodsir, 1841) (Cestoda; Trypanorhyncha) from sunfish (Mola mola). Fauna Norveg. 8: 25-28.
BEVERIDGE I., CAMPBELL R.A. 1996: New records and descriptions of trypanorhynch cestodes from Australian fishes. Rec. S. Aust. Mus. 29: 1-22.
BEVERIDGE I., SMITH K. 1988: Ultrastructure of the rhyncheal system of Trimacracanthus aetobatidis (Cestoda: Trypanorhyncha). Int. J. Parasitol. 18: 623-632.
BORUCINSKA J., CAIRA J.N. 1993: A comparison of mode of attachment and histopathogenicity of four tapeworm species representing two orders infecting the spiral intestine of the nurse shark, Ginglymostoma cirratum. J. Parasitol. 79: 238-246.
CARVAJAL J., BARROS C., WHITTAKER F.H. 1987: Scanning electron microscopy of the scolex of the plerocercous Callitetrarhynchus gracilis (Rudolphi, 1819) (Cestoda: Trypanorhyncha). J. Parasitol. 73: 1265-1267.
CARVAJAL J., CAMPBELL R.A., CORNFORD E.M. 1976: Some trypanorhynch cestodes from Hawaiian fishes, with description of four new species. J. Parasitol. 62: 70-77.
CHANDRA K.J. 1988: Nybelinia indica n. sp. (Cestoda: Trypanorhyncha) from teleost fishes of Waltair Coast, Bay of Bengal. Riv. Parassitol. 3: 190-202.
CHANDRA K.J., RAO K.H. 1985: Two new species of Tentaculariidae Poche, 1926 (Cestoda: Trypanorhyncha) from marine fishes of Waltair. Riv. Parassitol. 2: 439-443.
DAVYDOV V.G., BISEROVA N.M. 1985: Morphology of frontal glands of Grillotia erinaceus (Cestoda: Trypano-rhyncha). Parazitologiya 19: 32-38. (In Russian).
DOLLFUS R.-P. 1942: Etudes critiques sur les Tétrarhynques du Muséum de Paris. Archives du Muséum National d’Histoire Naturelle, Paris, 6 ème série 19: 1-466.
DOLLFUS R.-P. 1960: Sur une collection de Tétrarhynques homéacanthes de la famille des Tentaculariidae, récoltés principalement dans la région de Dakar. Bull. Inst. Fr. d’ Afrique Noire, série A., 22: 788-852.

Jones, Beveridge: Nybelinia queenslandensis sp n..
311
HALTON D.W., MCKERR G. 1979: SEM observations on the spiral valve of rays and their cestode parasites. Parasitology 79: xlvii.
HEINZ M.L., DAILEY M.D. 1974: The Trypanorhyncha (Cestoda) of elasmobranch fishes from southern California and Northern Mexico. Proc. Helminthol. Soc. Wash. 41: 161-169.
HOLY J.M., OAKS J.A. 1986: Ultrastructure of the tegumental microvilli (microtriches) of Hymenolepis diminuta. Cell Tiss. Res. 244: 457-466.
KUPERMAN B.I., DAVYDOV V.G. 1982: The fine structure of frontal glands in adult cestodes. Int. J. Parasitol. 24: 285-293.
KURSHID N., BILQUEES F.M. 1988: Nybelinia karachii new species from the fish Cybium guttatum of Karachi Coast. Pak. J. Zool. 20: 239-242.
LAST P.R., STEVENS J.D. 1994: Sharks and Rays of Australia. C.S.I.R.O., Canberra, Australia., 513 pp.
LINTON E. 1889: Notes on Entozoa of marine fishes of New England, with descriptions of several new species. Ann. Rep. Comm. Fisheries, 1886: 453-511.
LINTON E. 1897: Notes on larval cestode parasites of fishes. Proc. U. S. Nat. Mus. 19: 787-824.
MCCULLOUGH J.S., FAIRWEATHER I. 1983: A SEM study of the cestodes Trilocularia acanthiaevulgaris, Phyllobothrium squali and Gilquinia squali from the spiny dogfish. Z. Parasitenkd. 69: 655-665.
MCCULLOUGH J.S., FAIRWEATHER I. 1989: The fine structure and possible function of scolex gland cells in Trilocularia acanthiaevulgaris (Cestoda, Tetraphyllidea). Parasitol. Res. 75: 575-582.
MOUNT P.M. 1970: Histogenesis of the rostellar hooks of Taenia crassiceps (Zeder, 1800) (Cestoda). J. Parasitol. 56: 947-961.
PALM H. 1995: Untersuchungen zur Systematik von Rüsselbandwürmen (Cestoda: Trypanorhyncha) aus atlantischen Fischen. Berichte aus dem Institut für Meereskunde an der Christian-Albrechts-Universität Kiel. 238 pp.
PALM H., POYNTON S., RUTLEDGE P. 1998: Surface ultrastructure of plerocercoids of Bombycirhynchus sphyraenaicum (Pintner, 1930) (Cestoda: Trypano-rhyncha). Parasitol. Res. 84: 195-204.
PALM H.W., WALTER T., SCHWERDTFEGER G., REIMER L.W. 1997: Nybelinia Poche, 1926 (Cestoda: Trypanorhyncha from the Mocambique coast, with description of N. beveridgei sp. nov. and systematic consideration on the genus. S. Afr. J. Mar. Sci. 18: 273-285.
PINTNER T. 1899: Die Rhynchodäaldrüsen der Tetra-rhynchiden. Arb. Zool. Inst. Univ. Wien 12: 1-24.
PINTNER T. 1929: Tetrarhynchen von den Forschungsreisen des Dr. Sixten Bock. Göteborgs Kungl. Vetenskaps och Vitterhets Samhäller Handlingar, Femte Foldjen Serie B 1: 3-48.
PRITCHARD M.H., KRUSE G.O.W. 1982: The Collection and Preservation of Animal Parasites. University of Nebraska Press, Lincoln, 141pp.
REIMER L.W. 1980: Larven der Ordnung Trypanorhyncha (Cestoda) aus Teleostiern des Indischen Ozeans. Angew. Parasitol. 21: 221-231.
RICHMOND C., CAIRA J.N. 1991: Morphological investiga-tions into Floriceps minacanthus (Trypanorhyncha: Lacistorhynchidae) with analysis of the systematic utility of scolex microtriches. Syst. Parasitol. 19: 25-32.
SAO CLEMENTE S.C., GOMES D.C. 1992: Description of the adult form of Nybelinia (Syngenes) rougetcampanae Dollfus, 1960 and some new data on N. (N.) bisulcata (Linton, 1889) (Trypanorhyncha: Tentaculariidae). Mem. Inst. Oswaldo Cruz 87: 251-255.
SHAH M., BILQUEES F.M. 1979: Nybelinia elongata new species from the fish Pellona elongata of Karachi coast. Pak. J. Parasitol. 11: 231-233.
SHIELDS J.D. 1985: Surface morphology and description of Otobothrium kurisi new species (Cestoda: Trypano-rhyncha) from a hammerhead shark, Sphyrna lewini. Int. J. Parasitol. 15: 635-643.
STUNKARD H. 1977: Studies on tetraphyllidean and tetrarhynchidean metacestodes from squids taken on the New England coast. Biol. Bull. 153: 387-412.
WARD S.M., MCKERR G., ALLEN J.M. 1986: Structure and ultrastructure of muscle systems within Grillotia erinaceus metacestodes (Cestoda: Trypanorhyncha). Parasitology 93: 587-597.
WHITTAKER F.H., CARVAJAL J., APKARIAN R.A. 1982: Scanning electron microscopy of the scolex of Grillotia dollfusi Carvajal 1971 (Cestoda; Trypanorhyncha). J. Parasitol. 68: 1173-1175.
YAMAGUTI S. 1952: Studies on the helminth fauna of Japan. Part 49. Cestodes of fishes. Acta Med. Okayama 8: 1-76.
YANG W., LIN Y., LIU G., PENG W. 1995: Five species of Trypanorhyncha from marine fishes in Xiamen, Fujian, China. J. Xiamen Univ. Nat. Sci. 34: 811-817.
ŽĎÁRSKÁ Z., NEBESÁŘOVÁ J. 1997: Ultrastructure of three types of scolex gland cells in adult Bothriocephalus claviceps (Cestoda: Pseudophyllidea). Folia Parasitol. 44: 139-146.
Received 26 October 1997 Accepted 16 April 1998