norovirus rna-dependent rna-polymerase is phosphorylated ... file41 pandemic in 2007-8 (23) and...
TRANSCRIPT

Norovirus RNA-dependent RNA-polymerase is phosphorylated 1
by an important survival kinase, Akt 2
John-Sebastian Eden†, Laura J. Sharpe†, Peter A. White* and Andrew J. Brown* 3
BABS, School of Biotechnology and Biomolecular Sciences, Biosciences Building D26, 4
University of New South Wales, Sydney, NSW 2052, Australia. 5
Running Title: Akt phosphorylates Norovirus RdRp 6
Keywords: Akt; Norovirus; RdRp; phosphorylation; kinase 7
Abstract word count: 97 8
Text word count: 1624 9
† These authors contributed equally to this work. 10
* These corresponding authors contributed equally to this work. 11
Mailing address: BABS, School of Biotechnology and Biomolecular Sciences, Biosciences 12
Building D26, University of New South Wales, Sydney, NSW 2052, Australia. 13
Fax: +61-2-9385-1483; 14
AJB: Phone: +61-2-9385-2005; E-mail: [email protected] 15
PAW: Phone: +61-2-9385-3780; E-mail: [email protected] 16
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Virol. doi:10.1128/JVI.05562-11 JVI Accepts, published online ahead of print on 17 August 2011
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

1
Abstract 17
Viruses commonly use their host cells’ survival mechanisms to their own advantage. We 18
showed that Akt, an important signalling kinase involved in cell survival, phosphorylates the 19
RNA-dependent RNA-polymerase (RdRp) from Norovirus, the major cause of gastroenteritis 20
outbreaks worldwide. The Akt phosphorylation of RdRp appears to be a feature unique to the 21
more prevalent Norovirus genotypes such as GII.4 and GII.b. This phosphorylation event occurs 22
at a residue (Thr33) located at the interface where the RdRp finger and thumb domains 23
interact, and decreases de novo activity of the polymerase. This finding provides fresh insights 24
into virus-host cell interactions. 25
26
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

2
27
Norovirus (NoV) is the most common cause of acute gastroenteritis outbreaks, causing 28
vomiting and diarrhoea in affected individuals (14). Since 1996, five variants of the genogroup 29
II, genotype 4 (GII.4) lineage have been associated with global pandemics of acute 30
gastroenteritis of increased frequency (23). Overall, GII.4 variants are the cause of 62-80% of all 31
NoV outbreaks globally (11, 23). There are a number of factors contributing to this higher 32
epidemiological fitness including host-cell receptor binding patterns and duration of herd 33
immunity [reviewed in (7)]. Also, our group recently showed that the GII.4 viruses had a 34
greater capacity to evolve than other NoV genotypes through higher rates of mutation (6). 35
Therefore, the viral RNA-dependent RNA-polymerase (RdRp) is likely to play an important role 36
in driving viral evolution and fitness (6). Post-translational modifications are frequently found 37
on viral proteins (15), including RdRps (17), and this led us to examine a potential post-38
translational modification of this polymerase from a representative pandemic NoV strain, GII.4 39
2006b (NSW696T/06/AU - GenBank EF684915). This variant was associated with a global 40
pandemic in 2007-8 (23) and caused three consecutive epidemics of gastroenteritis in Australia 41
in 2006 (26), 2007 and 2008 (12). 42
Viruses take advantage of the host cells’ existing pathways and mechanisms, and one of 43
these is the phosphatidylinositol 3-kinase (PI3K)/Akt signalling pathway (5). Akt is a 44
serine/threonine protein kinase that phosphorylates downstream targets leading to numerous 45
effects, with the net result of increasing cell growth, survival and proliferation. Consequently, 46
perturbations of the Akt signalling pathway have been implicated in the development of 47
diseases including many human cancers (2) and diabetes (28). With such a fundamental role in 48
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

3
the regulation of cell growth and proliferation, it is perhaps not surprising that a number of 49
viruses have been found to interact directly with the PI3K/Akt pathway to effect greater control 50
of their replication within the host cell. In HCV, the expression of viral proteins including 51
NS3/4A, NS4B and NS5A result in the activation of the PI3K/Akt pathway [reviewed in (3)], 52
which is required for efficient virus replication (4). Similarly, Influenza A virus activates the 53
PI3K/Akt pathway through the viral non-structural protein 1 (13). This appears to suppress the 54
induction of apoptosis, which may arrest the cell at a stage that supports viral replication. 55
Alternatively, not all viruses activate the PI3K/Akt pathway. For example, in Measles virus, 56
infection leads to the down regulation of Akt that appears to plays a role in immune 57
suppression (8). 58
Given the common exploitation by viruses of Akt signalling, we sought to determine 59
whether the NoV RdRp may be a substrate for Akt. Akt phosphorylates proteins at the minimal 60
consensus sequence of RxRxx(S/T)h (where x is any amino acid, and h is a bulky hydrophobic 61
amino acid) (1). This minimal consensus motif was supported by another peptide library 62
screening approach (20). However, there is considerable variability from this consensus in the 63
sequences surrounding Akt phosphorylation sites (22). Therefore, to help determine the 64
potential for phosphorylation to occur on the NoV GII.4 2006b RdRp, we interrogated the 65
amino acid sequence using Scansite (21). This program predicts various phosphorylation sites 66
and protein binding motifs, including likelihood (stringency), based on minimal consensus 67
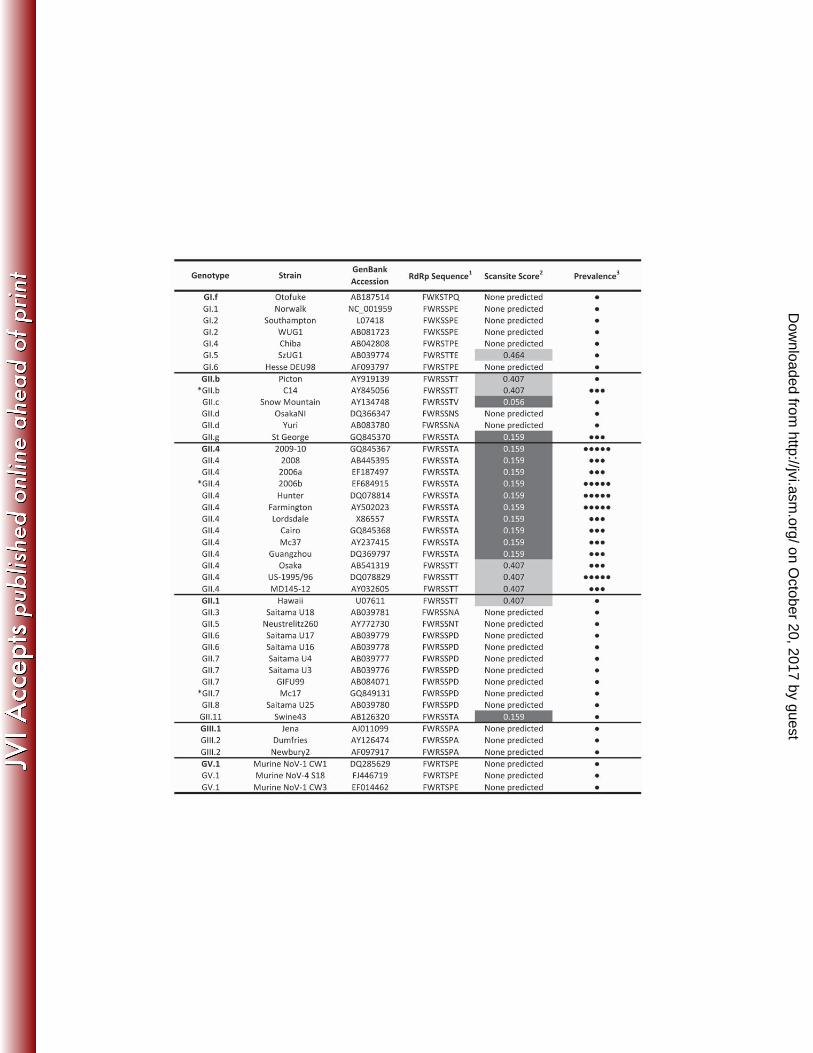
sequences. This indicated that the Thr33 residue of the 2006b RdRp was predicted as both a 68
high stringency Akt substrate site and 14-3-3 zeta binding site. The RdRp Thr33 residue is 69
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

4
mostly conserved between pandemic strains, and almost absent in non-pandemic strains (Table 70
1, χ2: p<0.001). 71
In light of the in silico predictions and the complementary expertise of the investigators 72
(12, 22), we decided to confirm Akt mediated phosphorylation of purified recombinant RdRp. 73
Briefly, the coding region for NoV RdRp with a C-terminal hexahistidine tag was cloned into 74
pGEX-4T-1 (GE Healthcare), then expressed and purified as previously described (6). An N-75
terminal glutathione-S-transferase (GST) tag was included to increase the size of the RdRp to 76
allow separation of the RdRp away from the Akt protein, by SDS-PAGE. An in vitro Akt kinase 77
assay (22) was employed to compare wild-type RdRp (GII.4 2006b), with a mutated version – 78
Thr33Ala, where the predicted phosphorylation site had been mutated to alanine. To detect 79
phosphorylation, we made use of an antibody designed for identification of phosphorylated Akt 80
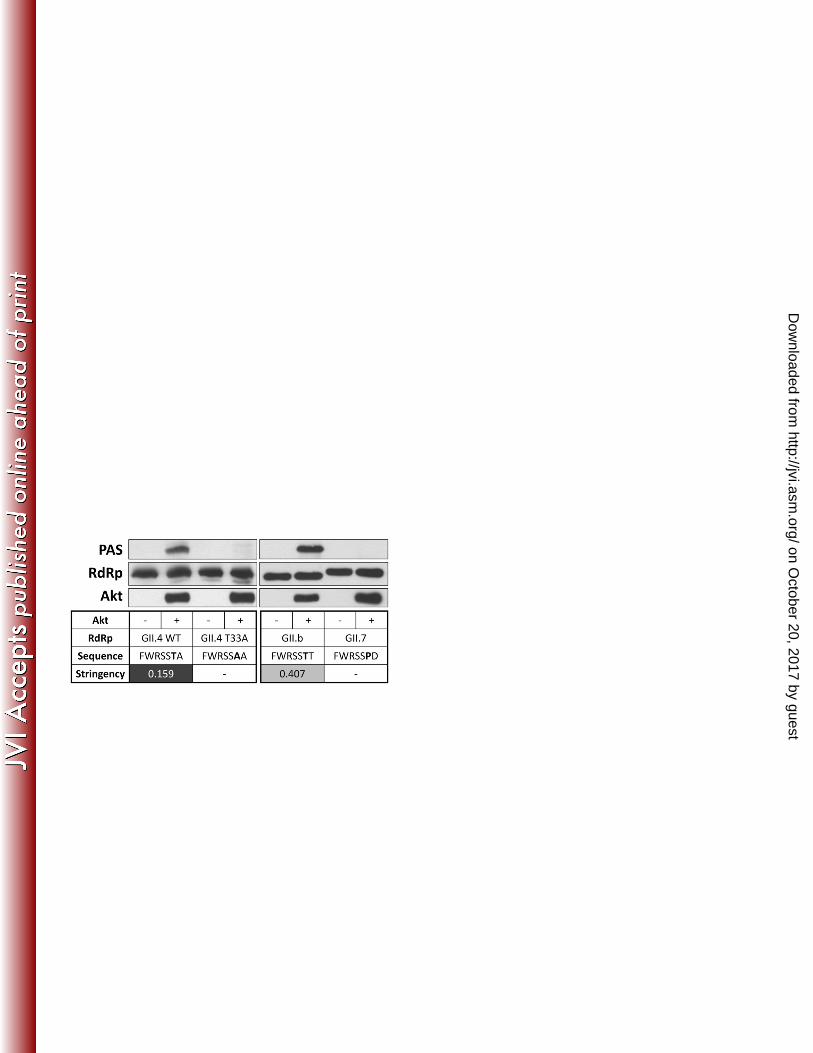
substrates [PAS, Cat #9611, Cell Signaling Technology, (29)]. Our results showed clear Akt-81
dependent phosphorylation of the wild-type GII.4 2006b RdRp (Figure 1), which was absent in 82
the mutated version. A second antibody that is more specific for phosphorylated Akt substrates 83
(Cat #9614, also from Cell Signaling Technology) also detected wild type phosphorylated RdRp 84
and not the mutated form (data not shown). In addition, we tested a representative RdRp with 85
a medium stringency site (GII.b C14) (C14/2002/AU - GenBank AY845056), and one with no 86
predicted phosphorylation site (GII.7 Mc17) (Mc17/2001/Th - GenBank AY237413). This 87
indicated that the medium stringency sequence was also phosphorylated at Thr33, but the GII.7 88
RdRp with no corresponding Thr residue was not (Figure 1). 89
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

5
Akt’s phosphorylation of NoV RdRp may provide additional insights into the 90
evolutionary strategies by which viruses exploit the host for their own benefit. The Akt 91
pathway is active when cells are proliferating rapidly, so it is possible that NoV takes advantage 92
of this situation to increase its survival. How this phosphorylation event may influence the 93
function of RdRp is unclear, since there is currently very little information linking structure and 94
function of the NoV RdRp, largely due to the lack of a cell culture system for the virus. Most of 95
the important residues have been predicted from published crystal structures (19, 27) and 96
comparison of structures to functionally defined residues in Poliovirus and Foot and Mouth 97
Disease Virus [e.g. (25)]. The phosphorylated Thr33 residue is located at the tip of the finger 98
domain which interacts with the top of the thumb and encloses the active site (Figure 2A & B). 99
A study by Thompson et al. suggests that this structure, which is unique to RdRps, plays a role 100
in stabilising Poliovirus RdRp (25). These same residues may play a similar stabilising role in 101
NoV RdRp, allowing a conformation that facilitates RNA synthesis. Homology modelling 102
predicts extensive inter-domain interactions within this region of the RdRp including two polar 103
bridging interactions that link the finger and thumb domains at Thr33 in the GII.4 2006b RdRp 104
(Figure 2C). When the Thr33 residue is phosphorylated, the negative charge of the phosphate 105
should repel the negatively charged side chain of Glu468, altering the dynamics of these inter-106
domain finger/thumb interactions. 107
The HCV RdRp (NS5B) has been shown to be phosphorylated by Protein Kinase C-related 108
Kinase 2, and that phosphorylation event played a role in regulating HCV RNA replication (16). 109
To explore the possibility that the phosphorylation of Thr33 is functionally significant, we 110
compared the enzyme kinetics of the wild-type 2006b RdRp with a mutated form that should 111
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

6
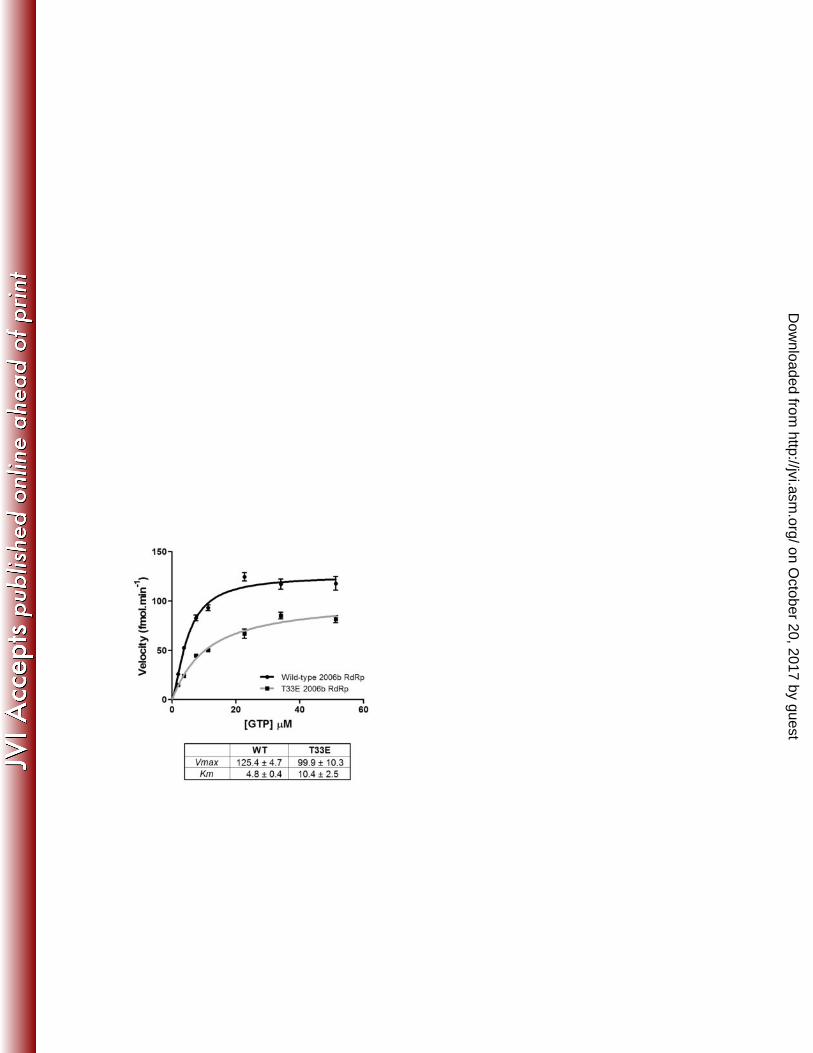
mimic phosphorylation (Thr33Glu), using a de novo GTP incorporation assay as previously 112
described (6). The phosphomimetic mutant had a lower maximum enzyme velocity (Vmax = 113
100 fmol.min-1) compared to the wild type RdRp (Vmax = 125 fmol.min-1), indicating a slower 114
rate of RNA synthesis (Figure 3). The phosphomimetic mutant also had a lower affinity for the 115
GTP substrate with a Km value of 10.4 µM, compared to 4.8 µM for the wild-type RdRp. The 116
differences in enzyme kinetics between the wild-type and phosphomimetic RdRp suggest that 117
the phosphorylation of Thr33 may modulate the function of the polymerase. In HCV, it has 118
been shown that disruptions to the interactions between the finger and thumb domains will 119
lead to a reduction in de novo RdRp activity, while efficient primer extension activity is 120
maintained (9, 10, 24). Since NoV utilises both de novo and protein primed RNA synthesis, it is 121
possible that phosphorylation of NoV RdRp could alter the balance between these modes of 122
RNA synthesis. 123
In contrast to examples of viral exploitation of the Akt pathway (3, 8, 13), a decrease in 124
polymerase activity could indicate that Akt phosphorylation of the NoV RdRp is an anti-viral 125
strategy by the infected cell. However, this may be hard to reconcile with our observation that 126
the Akt phosphorylation event occurs in the most epidemiologically fit genotype, GII.4 and is 127
generally absent from others. Further work will be needed to distinguish between these 128
possibilities and to determine the viral outcomes of this phosphorylation event. 129
Phosphorylation may not just alter function directly, but could act as a molecular signal 130
for a binding partner. In line with this, a high stringency 14-3-3 zeta binding site was predicted 131
by Scansite on the phosphorylated Thr33 residue. 14-3-3s are ubiquitous proteins that bind to 132
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

7
phosphorylated residues; therefore further alterations of protein function could occur (18). In 133
this case, due to the high intrinsic disorder within this region of the polymerase, it is possible 134
that 14-3-3 binding may act as a clamp that fixes the polymerase into a functionally significant 135
conformation. The interaction of 14-3-3 zeta and NoV RdRp may also affect the localisation 136
within the cell to aid evasion of the host cell recognition system. The possibility that 137
phosphorylation of NoV RdRp may affect intermolecular interactions represents another 138
interesting avenue for further research. 139
Here, we have discovered a novel Akt substrate, a viral polymerase, and identified its 140
phosphorylation site. This residue appears to be critical for the activity of the NoV RdRp, based 141
on our in vitro polymerase assay. Importantly, it is routinely present in pandemic strains, but 142
mostly lacking in non-pandemic genotypes. Future work should include confirming the NoV 143
RdRp phosphorylation event in vivo, determining its impact on viral replication and whether 144
this favours the host or the virus. In this respect, it would be worthwhile to engineer the 145
phosphorylation site into the existing murine NoV infectious cell culture system. The current 146
work provides a good basis for future efforts to better understand how phosphorylation of viral 147
proteins may affect replication. 148
149
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

8
References 150
1. Alessi, D. R., F. B. Caudwell, M. Andjelkovic, B. A. Hemmings, and P. Cohen. 1996. 151
Molecular basis for the substrate specificity of protein kinase B; Comparison with 152
MAPKAP kinase-1 and p70 S6 kinase. Febs Letters 399:333-338. 153
2. Altomare, D. A., and J. R. Testa. 2005. Perturbations of the AKT signaling pathway in 154
human cancer. Oncogene 24:7455-7464. 155
3. Bode, J. G., E. D. Brenndorfer, J. Karthe, and D. Haussinger. 2009. Interplay between 156
host cell and hepatitis C virus in regulating viral replication. Biol Chem 390:1013-32. 157
4. Brenndorfer, E. D., J. Karthe, L. Frelin, P. Cebula, A. Erhardt, J. Schulte am Esch, H. 158
Hengel, R. Bartenschlager, M. Sallberg, D. Haussinger, and J. G. Bode. 2009. 159
Nonstructural 3/4A protease of hepatitis C virus activates epithelial growth factor-160
induced signal transduction by cleavage of the T-cell protein tyrosine phosphatase. 161
Hepatology 49:1810-20. 162
5. Buchkovich, N. J., Y. Yu, C. A. Zampieri, and J. C. Alwine. 2008. The TORrid affairs of 163
viruses: effects of mammalian DNA viruses on the PI3K-Akt-mTOR signalling pathway. 164
Nature Reviews: Microbiology 6:266-275. 165
6. Bull, R. A., J. S. Eden, W. D. Rawlinson, and P. A. White. 2010. Rapid evolution of 166
pandemic noroviruses of the GII.4 lineage. PLoS Pathogens 6:e1000831. 167
7. Bull, R. A., and P. A. White. 2011. Mechanisms of GII.4 norovirus evolution. Trends 168
Microbiol 19:233-40. 169
8. Carsillo, M., D. Kim, and S. Niewiesk. 2010. Role of AKT kinase in measles virus 170
replication. J Virol 84:2180-3. 171
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

9
9. Chinnaswamy, S., A. Murali, P. Li, K. Fujisaki, and C. C. Kao. 2010. Regulation of de 172
novo-initiated RNA synthesis in hepatitis C virus RNA-dependent RNA polymerase by 173
intermolecular interactions. J Virol 84:5923-35. 174
10. Chinnaswamy, S., I. Yarbrough, S. Palaninathan, C. T. Kumar, V. Vijayaraghavan, B. 175
Demeler, S. M. Lemon, J. C. Sacchettini, and C. C. Kao. 2008. A locking mechanism 176
regulates RNA synthesis and host protein interaction by the hepatitis C virus 177
polymerase. J Biol Chem 283:20535-46. 178
11. Donaldson, E. F., L. C. Lindesmith, A. D. Lobue, and R. S. Baric. 2010. Viral shape-179
shifting: norovirus evasion of the human immune system. Nat Rev Microbiol 8:231-41. 180
12. Eden, J. S., R. A. Bull, E. Tu, C. J. McIver, M. J. Lyon, J. A. Marshall, D. W. Smith, J. 181
Musto, W. D. Rawlinson, and P. A. White. 2010. Norovirus GII.4 variant 2006b caused 182
epidemics of acute gastroenteritis in Australia during 2007 and 2008. Journal of Clinical 183
Virology 49:265-71. 184
13. Ehrhardt, C., T. Wolff, S. Pleschka, O. Planz, W. Beermann, J. G. Bode, M. Schmolke, 185
and S. Ludwig. 2007. Influenza A virus NS1 protein activates the PI3K/Akt pathway to 186
mediate antiapoptotic signaling responses. J Virol 81:3058-67. 187
14. Estes, M. K., B. V. Prasad, and R. L. Atmar. 2006. Noroviruses everywhere: has 188
something changed? Current Opinion in Infectious Diseases 19:467-74. 189
15. Kadaveru, K., J. Vyas, and M. R. Schiller. 2008. Viral infection and human disease--190
insights from minimotifs. Front Biosci 13:6455-71. 191
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

10
16. Kim, S. J., J. H. Kim, Y. G. Kim, H. S. Lim, and J. W. Oh. 2004. Protein kinase C-related 192
kinase 2 regulates hepatitis C virus RNA polymerase function by phosphorylation. J Biol 193
Chem 279:50031-41. 194
17. Mackenzie, J. M., M. T. Kenney, and E. G. Westaway. 2007. West Nile virus strain 195
Kunjin NS5 polymerase is a phosphoprotein localized at the cytoplasmic site of viral RNA 196
synthesis. J Gen Virol 88:1163-1168. 197
18. Muslin, A. J., J. W. Tanner, P. M. Allen, and A. S. Shaw. 1996. Interaction of 14-3-3 with 198
Signaling Proteins Is Mediated by the Recognition of Phosphoserine. Cell 84:889-897. 199
19. Ng, K. K., N. Pendas-Franco, J. Rojo, J. A. Boga, A. Machin, J. M. Alonso, and F. Parra. 200
2004. Crystal structure of norwalk virus polymerase reveals the carboxyl terminus in the 201
active site cleft. J Biol Chem 279:16638-45. 202
20. Obata, T., M. B. Yaffe, G. G. Leparc, E. T. Piro, H. Maegawa, A. Kashiwagi, R. Kikkawa, 203
and L. C. Cantley. 2000. Peptide and protein library screening defines optimal substrate 204
motifs for AKT/PKB. Journal of Biological Chemistry 275:36108-36115. 205
21. Obenauer, J. C., L. C. Cantley, and M. B. Yaffe. 2003. Scansite 2.0: proteome-wide 206
prediction of cell signaling interactions using short sequence motifs. Nucleic Acids 207
Research 31:3635-3641. 208
22. Sharpe, L. J., W. Luu, and A. J. Brown. 2011. Akt Phosphorylates Sec24: New Clues into 209
the Regulation of ER-to-Golgi Trafficking. Traffic 12:19-27. 210
23. Siebenga, J. J., H. Vennema, D. P. Zheng, J. Vinje, B. E. Lee, X. L. Pang, E. C. Ho, W. Lim, 211
A. Choudekar, S. Broor, T. Halperin, N. B. Rasool, J. Hewitt, G. E. Greening, M. Jin, Z. J. 212
Duan, Y. Lucero, M. O'Ryan, M. Hoehne, E. Schreier, R. M. Ratcliff, P. A. White, N. 213
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

11
Iritani, G. Reuter, and M. Koopmans. 2009. Norovirus illness is a global problem: 214
emergence and spread of norovirus GII.4 variants, 2001-2007. J Infect Dis 200:802-12. 215
24. Simister, P., M. Schmitt, M. Geitmann, O. Wicht, U. H. Danielson, R. Klein, S. 216
Bressanelli, and V. Lohmann. 2009. Structural and functional analysis of hepatitis C virus 217
strain JFH1 polymerase. J Virol 83:11926-39. 218
25. Thompson, A. A., R. A. Albertini, and O. B. Peersen. 2007. Stabilization of poliovirus 219
polymerase by NTP binding and fingers-thumb interactions. J Mol Biol 366:1459-74. 220
26. Tu, E. T. V., R. A. Bull, G. E. Greening, J. Hewitt, M. J. Lyon, J. A. Marshall, C. J. McIver, 221
W. D. Rawlinson, and P. A. White. 2008. Epidemics of Gastroenteritis during 2006 Were 222
Associated with the Spread of Norovirus GII.4 Variants 2006a and 2006b. Clinical 223
Infectious Diseases 46:413-420. 224
27. Zamyatkin, D. F., F. Parra, A. Machin, P. Grochulski, and K. K. Ng. 2009. Binding of 2'-225
amino-2'-deoxycytidine-5'-triphosphate to norovirus polymerase induces 226
rearrangement of the active site. J Mol Biol 390:10-6. 227
28. Zdychova, J., and R. Komers. 2005. Emerging role of Akt kinase/protein kinase B 228
signaling in pathophysiology of diabetes and its complications. Physiological Research 229
54:1-16. 230
29. Zhang, H., X. M. Zha, Y. Tan, P. V. Hornbeck, A. J. Mastrangelo, D. R. Alessi, R. D. 231
Polakiewicz, and M. J. Comb. 2002. Phosphoprotein analysis using antibodies broadly 232
reactive against phosphorylated motifs. Journal of Biological Chemistry 277:39379-233
39387. 234
235
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

12
Figure legends 236
Figure 1: Akt phosphorylates RdRp at Thr33 237
Wild-type (WT) or mutant (T33A) recombinant NoV GII.4 2006b, GII.b C14, or GII.7 Mc17 RdRp 238
(1 µg) were incubated with or without recombinant Akt (400 ng) as indicated at 30°C for 30 239
min. Reactions were stopped by addition of loading buffer, and samples were subjected to 240
SDS-PAGE and Western blotting with Phospho-Akt Substrates (PAS) antibody, followed by 241
stripping and probing for RdRp and Akt. Representative of three independent experiments. 242
The bold residue in the sequence indicates the residue corresponding to Thr33 in WT GII.4 243
2006b RdRp. Stringency values refer to Scansite stringency: <0.2 = high stringency and 0.2-1 = 244
medium stringency. 245
246
Figure 2: 3D-modelling of NoV RdRp showing finger-thumb domain interactions at Thr33 247
(A) Front view of the NoV GII.4 2006b RdRp highlighting the finger (red), thumb (blue) and palm 248
(yellow) domains. (B) Top view highlighting the fingertips (red) and C-terminal region (cyan). 249
(C) Close-up view of the boxed region of (B) showing the finger-thumb interactions at Thr33 for 250
2006b RdRp. Images produced using PyMOL, Version 1.3, Schrödinger, LLC. 251
252
253
254
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from

13
Figure 3: Comparison of enzyme kinetics of the wild-type 2006b RdRp with a Thr33Glu 255
phosphomimetic mutant 256
The enzyme kinetics of the wild-type 2006b RdRp (Wild) were compared with a Thr33Glu 257
phosphomimetic mutant (T33E). Briefly, 50 ng RdRp was added to initiate a reaction where 3H-258
GTP was incorporated onto homopolymeric C RNA. After a ten minute incubation, the reaction 259
was stopped, harvested onto a filterplate, and then the incorporated nucleotides detected via 260
scintillation counting. Samples were tested in triplicate and background values were subtracted 261
to obtain the measure of incorporated nucleotides as fmole.min-1. This was measured across a 262
range of GTP substrate concentrations, increasing to 51 µM. The data were analysed using the 263
Hill equation in GraphPad Prism, Version 5.04. The kinetic parameters are presented with 264
standard deviations. 265
266
Table 1 Footnotes 267
1RdRp residues 28 - 34 relative to GII4 2006b variant - NSW696T/06/AUS (GenBank Accession - 268
EF684915). The bold residue is the predicted phosphorylation site. 269
2Values refer to Scansite stringency: <0.2 = high stringency and 0.2-1 = medium stringency. 270
3Prevalence: ●●●●● indicates strain was associated with a global pandemic, ●●● indicates 271
strain was associated with a regional epidemic and ● indicates strain was associated with a local 272
outbreak or sporadic disease in the natural host. 273
*Strains used in Figure 1. 274
on October 20, 2017 by guest
http://jvi.asm.org/
Dow
nloaded from