nonsterol isoprenoids activate human constitutive...
TRANSCRIPT

1521-009X/44/4/595–604$25.00 http://dx.doi.org/10.1124/dmd.115.068551DRUG METABOLISM AND DISPOSITION Drug Metab Dispos 44:595–604, April 2016Copyright ª 2016 by The American Society for Pharmacology and Experimental Therapeutics
Nonsterol Isoprenoids Activate Human Constitutive AndrostaneReceptor in an Isoform-Selective Manner in Primary Cultured
Mouse Hepatocytes
Elizabeth A. Rondini, Zofia Duniec-Dmuchowski, and Thomas A. Kocarek
Institute of Environmental Health Sciences, Wayne State University, Detroit, Michigan
Received December 7, 2015; accepted January 20, 2016
ABSTRACT
Our laboratory previously reported that accumulation of nonsterolisoprenoids following treatment with the squalene synthase in-hibitor, squalestatin 1 (SQ1) markedly induced cytochrome P450(CYP)2B1 mRNA and reporter activity in primary cultured rathepatocytes, which was dependent on activation of the constitu-tive androstane receptor (CAR). The objective of the current studywas to evaluate whether isoprenoids likewise activate murine CAR(mCAR) or one or more isoforms of human CAR (hCAR) producedby alternative splicing (SPTV, hCAR2; APYLT, hCAR3). We foundthat SQ1 significantly induced Cyp2b10 mRNA (∼3.5-fold) in primaryhepatocytes isolated from both CAR–wild-type and humanized CARtransgenic mice, whereas the 3-hydroxy-3-methylglutaryl-CoA re-ductase inhibitor pravastatin had no effect. In the absence of CAR,basal Cyp2b10mRNA levels were reduced by 28-fold and the effect of
SQ1 on Cyp2b10 induction was attenuated. Cotransfection with anexpression plasmid for hCAR1, but not hCAR2 or hCAR3, mediatedSQ1-induced CYP2B1 and CYP2B6 reporter activation in hepatocytesisolated from CAR-knockout mice. This effect was also observedfollowing treatment with the isoprenoid trans,trans-farnesol. Thedirect agonist CITCO increased interaction of hCAR1, hCAR2, andhCAR3 with steroid receptor coactivator-1. However, no significanteffect on coactivator recruitment was observed with SQ1, suggestingan indirect activation mechanism. Further results from an in vitroligand binding assay demonstrated that neither farnesol nor otherisoprenoids are direct ligands for hCAR1. Collectively, our findingsdemonstrate that SQ1 activates CYP2B transcriptional responsesthrough farnesol metabolism in an hCAR1-dependent manner. Fur-ther, this effect probably occurs through an indirect mechanism.
Introduction
The constitutive androstane receptor (CAR, NR1I3) is a nuclearreceptor that is expressed predominately in the liver (Baes et al., 1994).Originally identified as the mediator of phenobarbital (PB)-induciblecytochrome P450 2B (CYP2B) expression (Honkakoski et al., 1998),CAR is now known to regulate a number of genes involved in thebiotransformation and clearance of xenobiotics as well as endogenouscompounds including bilirubin, bile acids, and thyroid hormone
(Maglich et al., 2002, 2004; Huang et al., 2003). Several CAR targetgenes have been identified and include enzymes of phase I and IImetabolism as well as drug transporters (Wang et al., 2012; Yang andWang, 2014). More recent evidence has linked CAR to the regulation ofgenes involved in gluconeogenesis (Yarushkin et al., 2013; Lynch et al.,2014) and lipid metabolism (Dong et al., 2009); thus CAR is emergingas a potential target for metabolic disease.Unlike other ligand-activated nuclear receptors, CAR exhibits sub-
stantial constitutive activity in the absence of ligand (Suino et al., 2004).In the uninduced state, CAR is localized in the cytoplasm as amultiprotein complex (Kobayashi et al., 2003; Yoshinari et al., 2003).Direct ligand binding or indirect activation stimulates CAR dephos-phorylation and translocation into the nucleus (Mutoh et al., 2009),where it binds as a heterodimer with retinoid X receptor (RXR) to theregulatory regions of responsive genes (Honkakoski et al., 1996, 1998;
Dr. Rondini was funded, in part, through a postdoctoral fellowship awardedthrough the Office of Vice President of Research (Wayne State University, Detroit,MI). This research was supported by the National Institutes of Health NationalHeart, Lung, and Blood Institute [Grant R01 HL050710] and the National Instituteof Environmental Health Sciences [Center Grant P30 ES020957].
dx.doi.org/10.1124/dmd.115.068551.
ABBREVIATIONS: CAR, constitutive androstane receptor (NR1I3); CAR-KO, CAR-knockout; CAR-WT, CAR–wild-type; CITCO, 6-(4-chlorophenyl)imidazo[2,1-b][1,3]thiazole-5-carbaldehyde-O-(3,4-dichlorobenzyl)oxime; CYP2B, cytochrome P450 2B; DMSO, dimethyl sulfoxide; ETOH, ethanol;FOH, farnesol; FPP, farnesyl pyrophosphate; GLuc-N, Gaussia princeps luciferase N-terminal; GLuc-C, Gaussia princeps luciferase C-terminal;hCAR, human constitutive androstane receptor (NR1I3); hCAR1, human CAR reference sequence (NCBI Reference Sequence NM_005122); hCAR2,human CAR, splice variant 2 (NCBI Reference Sequence NM_001077480, alternatively named SV3 or SV23); hCAR3, human CAR, splice variant 3(NCBI Reference Sequence XM_005245697, alternatively named SV2 or SV24); hCAR-TG, hCAR transgenic; HMGCR, 3-hydroxy-3-methylglutaryl-coenzyme A reductase; LBD, ligand binding domain; mCAR, murine CAR; PB, phenobarbital; PBRE, phenobarbital-responsive element; PBREM,phenobarbital-responsive enhancer module; PPARa, peroxisome proliferator–activated receptor alpha (NR1C1); PPARg, peroxisome proliferator–activated receptor gamma (NR1C3); PPRE, peroxisome proliferator–activated receptor response element; PCR, polymerase chain reaction;qRT-PCR, quantitative reverse transcription–polymerase chain reaction; RID, receptor interaction domain; RXR, retinoid X receptor (NR2B); SQ1,squalestatin 1 (zaragozic acid A); SRC, steroid receptor coactivator (NCOA); Srebp, sterol regulatory element-binding proteins; SV, splice variant;TCPOBOP, 1,4-bis-[2-(3,5-dichloropyridyloxy)]benzene; TR-FRET, time-resolved fluorescence resonance energy transfer; XREM, xenobiotic-responsive enhancer module.
595
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from

Sueyoshi et al., 1999). Although the general structure of CAR is highlyconserved, variations in the ligand-binding domain contribute to somespecies differences in ligand selectivity (Reschly and Krasowski, 2006;Omiecinski et al., 2011). For example, 1,4-bis-[2-(3,5-dichloropyridy-loxy)]benzene (TCPOBOP) and 6-(4-chlorophenyl)imidazo[2,1-b][1,3]thiazole-5-carbaldehyde O-(3,4-dichlorobenzyl)oxime (CITCO)are selective ligand agonists for mouse CAR (mCAR) (Tzameli et al.,2000) and human CAR (hCAR), respectively (Maglich et al., 2003).Several splice variants (SV) of hCAR have been identified that are
suggested to contribute to functional diversity in CAR responses(Auerbach et al., 2003; Savkur et al., 2003; Jinno et al., 2004; Lambaet al., 2004; DeKeyser et al., 2011). Among the major splice variantspresent in the human liver, hCAR2 (SV3 or SV23) and hCAR3 (SV2 orSV24) are estimated to account for up to ;10 and 50% of total hCARtranscripts, respectively (Jinno et al., 2004; Ross et al., 2010). Bothvariants encode proteins containing short insertions within the ligandbinding domain (LBD); i.e., either four amino acids (SPTV) for hCAR2or five (APYLT) for hCAR3 (Auerbach et al., 2003). In comparisonwith wild-type CAR (herein referred to as hCAR1), these variantsexhibit low constitutive activity and appear to function as classicligand-activated nuclear receptors, although indirect activation mech-anisms have been suggested (Faucette et al., 2007; Lau and Chang,2013). Limited information also indicates some differences in ligandselectivity among isoforms. While all variants are activated by CITCO(Auerbach et al., 2005, 2007; Lau et al., 2011), hCAR2 is also stronglyactivated by di-(2-ethylhexyl)phthalate (DeKeyser et al., 2009) and di-isobutyl phthalate (DeKeyser et al., 2011), whereas clotrimazole,bisphenol A, and pheniramine (Auerbach et al., 2005; Dring et al.,2010; DeKeyser et al., 2011) are agonists for hCAR3.Regulation of hCAR by endogenous metabolites has been less well
characterized. We previously reported that inhibition of the enzymesqualene synthasewith squalestatin 1 (SQ1) inducedCYP2B1 expressionin rat liver and primary cultured rat hepatocytes (Kocarek et al., 1998).Further studies demonstrated that this effect was CAR-dependent,independent of reduced sterol synthesis, and mediated through an endo-genous isoprenoid (Kocarek and Mercer-Haines, 2002). The substrate ofsqualene synthase, farnesyl pyrophosphate (FPP), is normally convertedto squalene, leading to cholesterol biosynthesis or used to producevarious nonsterol isoprenoids (Krag, 1998; Dallner and Sindelar, 2000).Squalene synthase inhibitors drive the accumulation of FPP (Jackson andKocarek, 2008), which is dephosphorylated to farnesol (FOH) and thenfurther metabolized to produce a family of FOH-derived dicarboxylicacids that are excreted in the urine (Bostedor et al., 1997). Treatment ofprimary rat hepatocytes (Kocarek andMercer-Haines, 2002) or rats (Hornet al., 2005) with FOH also induces CYP2B1, suggesting that FOH itselfmight be a CAR activator, although this has not been formally demon-strated. Additionally, whether SQ1 or FOH regulate mouse or humanCYP2B expression through CAR activation is not currently known.Therefore, the purpose of the current investigation was to evaluatewhether SQ1 treatment activates mouse and human CAR-mediatedresponses by using primary cultured mouse hepatocytes. Reporter-basedand coactivator recruitment assays were additionally conducted todetermine the ability of SQ1 and FOH-derived metabolites to directlytransactivate hCAR isoforms.
Materials and Methods
Materials. SQ1 was a gift from GlaxoSmithKline (Research Triangle Park,NC) and pravastatin from Bristol-Myers Squibb Co. (Stamford, CT). Dimethylsulfoxide (DMSO), CITCO, TCPOBOP, and trans,trans-FOH were purchasedfrom MilliporeSigma (St. Louis, MO), other isoprenoids were purchased fromEchelon Biosciences (Salt Lake City, UT), Matrigel from Corning Life Sciences(Tewksbury, MA), PureCol from Advanced BioMatrix (San Diego, CA),
recombinant human insulin (Novolin R) from Novo Nordisk Pharmaceuticals,Inc. (Princeton, NJ), and culture medium, supplements, and Lipofectamine 2000reagent from Invitrogen (Carlsbad, CA). Additional sources of reagents areindicated with the methods descriptions that are provided below.
Primary Cultures of Mouse Hepatocytes. Male C57BL/6 mice CAR–wild-type (CAR-WT), C57BL/6-Nr1i3tm1.1Arte CAR-knockout mice (CAR-KO), andC57BL/6-Nr1i3tm1(NR1I3)Arte humanized CAR transgenic mice (hCAR-TG), all5–6 weeks of age, were purchased from Taconic Farms (Germantown, NewYork) and maintained in an Association for Assessment and Accreditation ofLaboratory Animal Care–approved animal facility with free access to chow andhouse-distilled water for at least 1 week prior to use. Generation of the hCAR-TGmouse model has been described previously (Scheer et al., 2008). Briefly, thehCAR (NR1I3) gene sequence from the first coding exon (exon 2) through exon9 was knocked into the coding region of the murine Nr1i3 gene; therefore,expression of hCAR is controlled by the mouse Nr1i3 promoter. Because theentire hCAR genomic coding region is present, all potential major mRNA splicevariants of human CAR (hCAR1, hCAR2, hCAR3) (Auerbach et al., 2003; Jinnoet al., 2004) are expressed (Scheer et al., 2008).
Hepatocytes were isolated from mouse livers using a collagenase perfusionmethod, which has been described in detail elsewhere (Wu et al., 2001).Following isolation, 1.2 million hepatocytes per well were plated onto collagen I-coated six-well plates (for RNA experiments), or 0.5 million hepatocytes per wellonto 12-well plates (for transfection experiments), and hepatocytes were culturedwith Williams’ E medium supplemented with 0.1 mM triamcinolone acetonide,0.25 IU/ml insulin, 100 IU/ml penicillin, and 100 mg/ml streptomycin. Culturemedium was renewed every 24 hours and, when appropriate, drugs or syntheticCAR ligands were added to the culture medium as concentrated stock solutions(1000X). Pravastatin and SQ1 were diluted in water, CITCO and TCPOBOP inDMSO, and FOH in ethanol (ETOH) at the concentrations specified in theindividual figure legends. FOH (60 mM) and ETOH (0.1%) were complexed tofatty acid-free bovine serum albumin (0.05% w/v) in Williams’ E medium for atleast 10 minutes prior to use.
Total RNA Isolation and Quantitative Reverse Transcription–Polymerase Chain Reaction. Freshly isolated hepatocytes from CAR-WT,CAR-KO, or hCAR-TG mice were plated onto six-well plates, as describedearlier. Beginning 24 hours after plating, culture medium was replaced withWilliams’ E medium containing Matrigel (1:50 dilution) and then furtherincubated overnight at 37�C. The following day, cells were treated withcholesterol synthesis inhibitors or prototypical CAR activators (CITCO,TCPOBOP) at the concentrations described in the individual figure legends.Culture medium containing treatments was renewed once after 24 hours. Forty-eight hours following the initial treatment, total RNA was extracted and columnpurified using the PureLink RNA isolation kit (Ambion, Carlsbad, CA), andcDNAwas synthesized from total RNA using the High-Capacity cDNAReverseTranscription Kit (Life Technologies/ThermoFisher Scientific, Grand Island,NY), each according to the manufacturer’s instructions.
Gene-specific primers to detect murine Cyp2b10, 3-hydroxy-3-methylglutaryl-coenzyme A reductase (Hmgcr), and TATA box binding protein (product numberMm.PT.39a.22214839) mRNAs as well as the hCAR splice variants hCAR1,hCAR2, and hCAR3were purchased from IntegratedDNATechnologies (Coralville,IA). Primer sequences to detect the individual splice variants of hCAR have beenpublished previously (Jinno et al., 2004; Zhang et al., 2013). The sequences (59 to 39)of primer pairs used to detect Cyp2b10 and Hmgcr are as follows: Cyp2b10: forward59-GCTTTGAGTACACAGACCGT-39 and reverse 59-CTCAAACATCTGGCTG-GAGA-39; Hmgcr: forward 59- ATCCTGACGATAACGCGGTG-39 and reverse59-AAGAGGCCAGCAATACCCAG-39. Determination of mRNA levels was con-ducted using a StepOnePlus Real-Time PCR System (Applied Biosystems, FosterCity, CA) and SYBR Green Master Mix (Life Technologies/ThermoFisherScientific). Each reaction (20 ml) contained 10 ml of SYBR green master mix,50 ng of RNA equivalents, and 150 nM primers. The polymerase chain reaction(PCR) cycling conditions included an initial activation step (95�C, 15 minutes)followed by 40 cycles of melting (95�C, 15 seconds), and annealing/extension (60�C,1minute). TATAbox binding protein was used as the endogenous control. All assayswere performed in duplicate, and the comparative cycle threshold (DDCT) methodwas used to calculate relative fold changes.
Generation of Plasmid Constructs. Generation of a CAR-responsiveluciferase (firefly) reporter plasmid containing the phenobarbital-responsiveelement (PBRE) within ;2.4 kb of the rat CYP2B1 59-flanking region has
596 Rondini et al.
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from

been described previously (Kocarek et al., 1998; Kocarek and Mercer-Haines,2002). A CYP2B6 luciferase reporter plasmid containing both the proximalphenobarbital-responsive enhancer module (PBREM) and the distal xenobiotic-responsive enhancer module (XREM) of the human CYP2B6 gene (Wang et al.,2003) was generously provided by Dr. Thomas Chang (University of BritishColumbia; British Columbia, CA). Expression plasmids containing the full-lengthcoding sequence for the major human CAR splice variants (hCAR1, hCAR2, andhCAR3) and empty vector control were gifts from Dr. Curtis Omiecinski(Pennsylvania State University, College Park, PA) (Auerbach et al., 2003).
Isoform-specific hCAR interaction with the coactivator, steroid receptorcoactivator (SRC)1, was evaluated using a protein-fragment complementationassay (PCA), as described elsewhere (Remy and Michnick, 2006, 2007).Expression plasmids containing either the N-terminal region of the Gaussiaprinceps luciferase (Gluc-N) cDNA (corresponding to amino acids 1–93) ligatedinto the pEYFP-N1 expression plasmid or the C-terminal sequence (correspond-ing to amino acids 94–169; Gluc-C) ligated into pECFP-C1 (Clontech, MountainView, CA) were prepared and kindly provided by Dr. James Granneman (WayneState University, Detroit, MI). Chimeric plasmids were then constructed for theexpression of fusion proteins containing either the full-length coding sequenceof hCAR (isoform 1, 2, or 3) ligated downstream of Gluc-N cDNA (hCAR–Gluc-N) or the receptor interaction domain (RID) of hSRC1 ligated upstream ofthe Gluc-C sequence [hSRC1(RID)-Gluc-C]. The CAR and SRC1 inserts wereligated in-frame with and separated from the Gluc sequences by a polypeptidelinker. Constructs were prepared by PCR using expression plasmids astemplates, PfuUltraII Fusion DNA polymerase (Agilent, Santa Clara, CA), andgene-specific primers. The sequences of the primer pairs for generating all of thehCAR splice variants were: forward 59-GCGAAGCTTTGGGCGGAGGCG-GAAGCGGCGGAGGCGGAAGTAGGGAAGATGAGCTGAGGAACT-39(underscored sequence = HindIII site; bold = 2 nt to maintain frame; doubleunderscored = GGGGSGGGG amino acid spacer) and reverse 59-GCGGGATCCTCAGCTGCAGATCTCCTGGAGCA-39 (underscored =BamHI site), corresponding to nt 210–1250 of the reference sequence forhCAR1 (NM_005122). For SRC1-RID, the forward primer sequence was 59-GCGAAGCTTGCCACCATGGATTACAAGGATGACGACGATAAG-GATGGAGACAGTAAATACTCTCAAAC-39 (underscored sequence = HindIIIsite; bold = Kozak consensus sequence; italicized = start codon; doubleunderscored= flag tag sequence), and the reverse primer sequence was 59-GCGGGATCCAGGCTTCCGCCTCCGCCGCTTCCGCCTCCGCCGTTTG-GAGTTGATCTTAAATCTTTCT-39 (underscored sequence = BamHI; bold =2 nt to maintain frame; double underscored = GGGGSGGGGS spacer),corresponding to nt 2119–2553 of reference sequence NM_003743. Theunderscored nucleotides indicate restriction sites for ligating the PCR productsinto the respective expression plasmid. Sequences of cloned fragments wereverified by the Applied Genomics Technology Center at Wayne State University.
Transient Transfection of Primary Cultured Hepatocytes. Primaryhepatocytes isolated from CAR-KO mice were transiently transfected withCYP2B1-PBRE or CYP2B6-PBREM/XREM reporter plasmids as previouslydescribed (Kocarek and Mercer-Haines, 2002) with minor modifications. Briefly,hepatocytes were plated onto collagen I-coated 12-well plates (0.5 millionhepatocytes per well), and 24 hours after plating, culture medium was replacedwith fresh Williams’ E medium. For reporter assays, cells were transientlytransfected with premixed complexes of Lipofectamine 2000 (4 ml), a CYP2Bfirefly reporter plasmid (1.5mg), an hCAR expression plasmid or empty expressionvector (50 ng), and 1 ng of the Renilla luciferase-CMV reporter plasmid (PromegaCorporation, Madison, WI) diluted in 0.2 ml Opti-MEM medium. For the PCAassay, the transfection mixture contained Lipofectamine 2000 (3 ml), hCAR-Gluc-N (50 ng), and hSRC1(RID)-Gluc-C (200 ng) expression plasmids, pBluescriptKS+ (1 mg), and 5 ng of the pGL3-Control firefly luciferase reporter plasmid(Promega Corporation) to allow for normalization. Culture medium was removedafter 5 hours and replaced with Williams’ E medium containing Matrigel (1:50dilution). The next day cells were treated with medium either alone or containingdrug treatments as described in the individual figure legends. Culture mediumcontaining the drug treatments was renewed after 24 hours. Hepatocytes were thenharvested for measurement of luciferase activity (firefly and Renilla or firefly andGaussia) 48 hours following initial treatment using the Dual Luciferase ReporterAssay System (Promega Corporation) and a GloMax luminometer (PromegaCorporation). Treatments were performed in triplicate and repeated in two to fiveindependent hepatocyte culture experiments (n = 3wells/treatment per experiment).
Time-Resolved Fluorescence Resonance Energy Transfer CAR Coac-tivator Recruitment Assay. FOH was diluted in either ETOH or DMSO tomake a 10 mM stock concentration. Other isoprenoid stock solutions wereprepared in water (FPP, isopentenyl pyrophosphate, dimethylallyl pyrophos-phate, geranyl pyrophosphate, and geranylgeranyl pyrophosphate), DMSO(farnesoic acid), or ETOH (methyl farnesoate, geranylgeraniol). CITCO,prepared in DMSO, was used as a positive control hCAR agonist ligand, andclotrimazole (in DMSO) as an inverse agonist. The LanthaScreen TR-FRETCAR Coactivator Assay was used according to the manufacturer’s instructions(Invitrogen) and has been recently described (Carazo and Pavek, 2015). Adilution series of each test compound was prepared at 100� and then diluted to2� with Complete TR-FRET Coregulator Buffer G. A 10-ml aliquot of eachdilution was transferred to the wells of a 384-well plate in triplicate, and theglutathione S-transferase–hCAR ligand-binding-domain fusion protein (20 nM)was added to each well followed by a mixture containing 0.5 mM fluorescein-labeled peroxisome proliferator-activated receptor gamma, coactivator 1 alpha(PGC1a) peptide, and 20 nM terbium-conjugated anti-glutathione S-transferaseantibody. The plate was sealed, protected from light, and incubated with gentleshaking for 1 hour at room temperature. Time-resolved fluorescence resonanceenergy transfer (TR-FRET) fluorescence was measured using the Victor2VMultilabel Reader (PerkinElmer, Waltham, MA) with an excitation wavelengthof 340 nm and fluorescent emission signals detected at 520 (fluorescein) and 490(terbium) nm. Emission ratios were calculated as the quotients of the emissionvalues at 520 nm divided by the emission values at 490 nm. Binding curves weregenerated using the sigmoidal dose-response (variable slope) algorithm ofGraphPad Prism 6 (GraphPad Software; La Jolla, CA).
Statistical Analysis. Statistical analyses were performed using the SigmaStatstatistical program (Version 3.5; Point Richmond, CA). Data for quantitativereverse-transcription polymerase chain reaction (qRT-PCR) and transienttransfections were analyzed using one-way or two-way analysis of variance asappropriate. When statistical differences were detected, the Student-Newman-Keuls test was used for individual comparisons. Results were consideredsignificant at P , 0.05.
Results
Effect of SQ1 Treatment on Cyp2b10 and Hmgcr mRNA Levelsin Primary Cultured Hepatocytes Isolated from CAR-WT andCAR-KO Mice. To determine whether the effect of squalene synthaseinhibition on isoprenoid-mediated CYP2B expression is conserved andfurther investigate whether this effect is attributable to activation of oneor more of the hCAR isoforms that are generated by alternative splicing,we first evaluated treatment effects on Cyp2b10 mRNA levels in primaryhepatocytes isolated from CAR-WT and CAR-KO mice. As shown inFig. 1, both SQ1 and the prototypical mouse CAR activator TCPOBOPsignificantly upregulated Cyp2b10 expression in a CAR-dependentmanner. In hepatocytes isolated from CAR-WT mice, Cyp2b10 mRNAlevels were increased ;3.5-fold by SQ1 and 6.8-fold by TCPOBOP 48hours after treatment (P, 0.05; Fig. 1A), whereas the HMGCR inhibitorpravastatin nonsignificantly decreased Cyp2b10 levels. In the absence ofCAR, the basal level of Cyp2b10 mRNA was 28-fold lower than thatmeasured in CAR-WT hepatocytes; however, no further significanttreatment effects on Cyp2b10 levels were observed (P. 0.05; Fig. 1A).Depletion of intracellular cholesterol initiates transcriptional upre-
gulation of cholesterol biosynthetic enzymes through proteolyticcleavage and activation of the sterol-sensitive transcription factors(sterol regulatory element–binding proteins, Srebps) (Sakakura et al.,2001). To demonstrate that SQ1 and pravastatin were both effective inblocking cholesterol synthesis, we evaluated Hmgcr mRNA levels inCAR-WT and CAR-KO hepatocytes (Fig. 1B). As shown in Fig. 1B,SQ1 and pravastatin, but not TCPOBOP, induced Hmgcr mRNA levelsto a similar extent in both CAR-WT and CAR-KO hepatocytes (P ,0.05), consistent with sterol depletion. Because both drugs inducedHmgcr and only SQ1 induced Cyp2b10 mRNA levels, our resultsindicate that the effect of SQ1 on Cyp2b10 induction is independent of
Nonsterol Isoprenoids Activate hCAR1 597
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from

Srebp-mediated transcription, as previously established in rats(Kocarek and Mercer-Haines, 2002).SQ1 Induces Cyp2b10 mRNA Levels in Primary Cultured
Hepatocytes Isolated from hCAR-TG Mice. Having demonstratedthat responsiveness of Cyp2b10 to SQ1 was attenuated in CAR-KOmouse hepatocytes, we next evaluated whether SQ1 mediates Cyp2b10induction in primary hepatocytes isolated from humanized CARtransgenic mice (hCAR-TG). In this model, expression of hCAR iscontrolled by the mCAR promoter; however, activation of CAR-mediated responses is hCAR-dependent (Scheer et al., 2008). Treat-ment effects were measured after 48 hours, and relative expressionlevels of the major splice variants of hCAR were also evaluated. Asshown in Fig. 2, both SQ1 and the prototypical hCAR activator CITCOincreased Cyp2b10 mRNA levels, although this only reached signif-icance in SQ1-treated hepatocytes (P, 0.05), whereas pravastatin hadno effect on Cyp2b10 mRNA levels. Consistent with the effects
observed in wild-type mouse hepatocytes, SQ1 and pravastatin alsoincreased Hmgcr mRNA levels to a similar extent (P, 0.05; Fig. 2B),demonstrating that both treatments were equally effective in blockingsterol synthesis and the cumulative effect of SQ1 was attributable toisoprenoid accumulation. Relative mRNA levels for the major splicevariants of hCAR were also assessed (Fig. 2C). We found that amongthe major splice variants, hCAR1 (SV0, 59%) was most abundantlyexpressed, followed by hCAR3 (;33%) and hCAR2 (;8%), whenexpressed as a percentage of the total of the three isoforms. Theseresults are consistent with levels observed in human livers and otherhCAR transgenic mouse models (Jinno et al., 2004; Ross et al., 2010;Zhang et al., 2013).SQ1 Activates CYP2B1-PBRE and CYP2B6-PBREM/XREM
Reporter Activity in an hCAR1-Dependent Manner. The previousresults suggested that SQ1 treatment could activate hCAR dependingon the cellular context. Therefore, we next evaluated whether the effectsof SQ1 were mediated through one or more of the individual splicevariants of hCAR using hepatocytes from a CAR-KO cellularbackground. To consider whether species differences in CAR targetgene sequences could contribute to hCAR-mediated effects, luciferasereporter assays were conducted with constructs containing either thePBREM/XREM of the human CYP2B6 gene or the PBRE of the ratCYP2B1 gene. Primary hepatocytes isolated from CAR-KO mice werecotransfected with a reporter construct and each individual hCARexpression plasmid or an empty vector control, and treatment effectswere evaluated after 48 hours. Results are presented in Fig. 3. For bothCYP2B reporter plasmids, a significant interaction was detectedbetween treatment and expression plasmid (P , 0.0001). In general,transfection of cells with hCAR1 significantly increased basal activityof both reporters, compared with the empty expression vector (Fig. 3, Aand B; P , 0.05). Treatment of hCAR1-transfected hepatocytes withSQ1, the hCAR ligand agonist CITCO, or the indirect CAR activatorPB significantly increased the activity of both reporters, compared withvehicle controls (P , 0.05; Fig. 3, A and B).Some treatment-induced increases in luciferase activity for both
reporters were evident in the empty expression vector–transfectedCAR-KO hepatocytes, which were stronger for CITCO than the othertreatments and somewhat variable among different hepatocyte prepa-rations. In cells transfected with the CYP2B1 reporter, CITCOtreatment significantly increased luciferase activity in the hCAR3-transfected cells (Fig. 3A). However, this CITCO-mediated increasewas not significantly different from the luciferase activity measured inCITCO-treated empty vector controls (P . 0.05), indicating a weakeffect of CITCO/hCAR3 on CYP2B1 activation in this system. Afteradjusting for background levels in the empty expression vector–transfected controls and the basal activity in vector-matched untreatedcontrols, the adjusted fold-changes for hCAR1 were 1.5, 1.5, and 2.4;for hCAR2, 0.95, 0.87, and 1.1; and for hCAR3, 1.0, 1.0, and 1.0 inhepatocytes treated with SQ1, CITCO, and PB, respectively. In cellstransfected with the CYP2B6-PBREM/XREM reporter, significantbasal luciferase activity was detected with all hCAR isoforms, althoughthis effect was weaker for the hCAR2 and hCAR3 isoforms (,2-fold)than for hCAR1 (;6-fold) (Fig. 3B). Similar to the findings for theCYP2B1 reporter, CITCO or SQ1 treatment increased CYP2B6-PBREM/XREM reporter activity ,2-fold in hCAR2 or hCAR3-transfected hepatocytes, and more strongly in hCAR1-transfected cells.After adjusting for basal activity detected among the different hCARexpression plasmids and for treatment effects within the empty vectorcontrols, the adjusted fold-changes for hCAR1 were 1.4, 2.7, and 2.3; forhCAR2, 1.0, 1.1, and 1.1; and for hCAR3, 1.1, 1.1, and 1.1 in hepatocytestreated with SQ1, CITCO, and PB, respectively. Thus, these resultsindicate the main treatment effects occurred in hCAR1-transfected cells
Fig. 1. Effect of the cholesterol synthesis inhibitors SQ1 and pravastatin on mRNAlevels of (A) Cyp2b10 and (B) Hmgcr in primary cultured hepatocytes isolated fromCAR-WT and CAR-KO mice. Hepatocytes were isolated from C57BL/6 (CAR-WT)and CAR-KO mice and plated onto collagen-coated six-well plates. Forty-eighthours after plating, cultures were incubated in Williams’ E medium either alone(CON) or containing 0.1% DMSO, 0.1 mM SQ1, 30 mM pravastatin, or theprototypical mouse CAR ligand agonist TCPOBOP (0.25 mM). Ninety-six hoursafter the initial plating, cultures were processed for the measurement of Cyp2b10and Hmgcr mRNA levels by qRT-PCR. Values are normalized to those of theuntreated (CON), CAR-WT hepatocytes. Each bar represents the mean 6 S.E.M. ofnormalized values from 3–4 independent hepatocyte preparations. *Significantlydifferent compared with untreated (CON) controls (P , 0.05). †Significantlydifferent compared with DMSO controls (P , 0.05).
598 Rondini et al.
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from

for both the CYP2B1 and CYP2B6-PBREM/XREM reporters (Fig. 3,A and B).FOH Treatment Enhances CYP2B6-PBREM/XREM Reporter
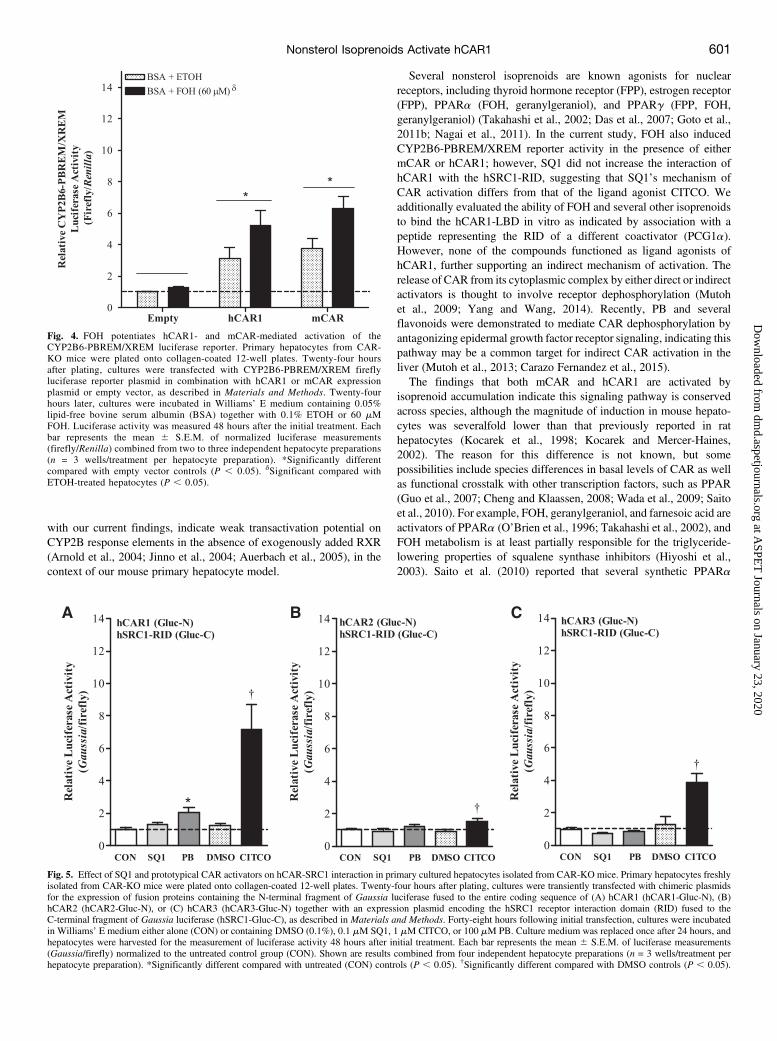
Activity in Primary Cultured CAR-KO Hepatocytes Transfectedwith Either hCAR1 or mCAR Expression Plasmid. Inhibition ofsqualene synthase causes accumulation of the branch point intermediateFPP, which can be dephosphorylated to produce the isoprenoid alcoholFOH (Bansal and Vaidya, 1994). FOH is then oxidized to farnesoic acidand a family of dicarboxylic acids that are then excreted in the urine(Gonzalez-Pacanowska et al., 1988; Bostedor et al., 1997; Vaidya et al.,1998). Treatment of animals with squalene synthesis inhibitors or FOHproduces a similar profile of metabolites in the urine, indicating that thispathway is the primary route of FPP catabolism following squalenesynthase inhibition (Bostedor et al., 1997; Vaidya et al., 1998).Treatment of primary cultured rat hepatocytes or rat livers with FOHwas previously shown to upregulate CYP2B1 mRNA levels (Kocarekand Mercer-Haines, 2002; Horn et al., 2005). Therefore, we addition-ally determined whether FOH treatment could directly influencehCAR1- or mCAR-mediated activation of the CYP2B6-PBREM/XREM reporter plasmid. As shown in Fig. 4, transfection of cells witheither hCAR1 or mCAR significantly increased CYP2B6-PBREM/XREM reporter activity compared with empty vector controls (P ,0.05). Treatment of cells with FOH (60 mM) further enhanced reporteractivity in the hCAR1- and mCAR-transfected cells by ;1.5-foldcompared with ETOH-treated controls (Fig. 4).Treatment Effects of SQ1 on hCAR-SRC1 Coactivator In-
teraction in Primary Cultured Hepatocytes. Unlike most ligand-activated nuclear receptors, hCAR1 can weakly interact with coactivatorsin the absence of a ligand, which is stabilized upon heterodimerizationwith RXR (Dussault et al., 2002) and even further in the presence of aligand agonist (Wright et al., 2007). A number of coactivators have beenidentified that bind to CAR-WT (Muangmoonchai et al., 2001; Arnoldet al., 2004; Kachaylo et al., 2011). Because all major splice variants ofhCAR (hCAR1, hCAR2, and hCAR3) interact with the nuclear receptorcoactivator SRC1 (Auerbach et al., 2003; Lau and Chang, 2013), weevaluated whether SQ1 treatment influenced isoform-specific hCAR-SRC1(RID) interactions using a protein-fragment complementationassay. As shown in Fig. 5, SQ1 had no significant effect on the
interaction of hSRC1(RID) with any hCAR isoform, whereas theindirect CAR activator PB modestly increased hSRC1(RID) interactionwith hCAR1 (P , 0.05). By comparison, the ligand agonist CITCO(1 mM) strongly increased the interaction between hSRC1(RID)and hCAR1 (;7-fold), weakly increased the interaction with hCAR2(1.7-fold), and moderately increased the interaction with hCAR3(;4-fold) (P , 0.05) (Fig. 5).FOH and Nonsterol Isoprenoids Are Not Direct Ligand
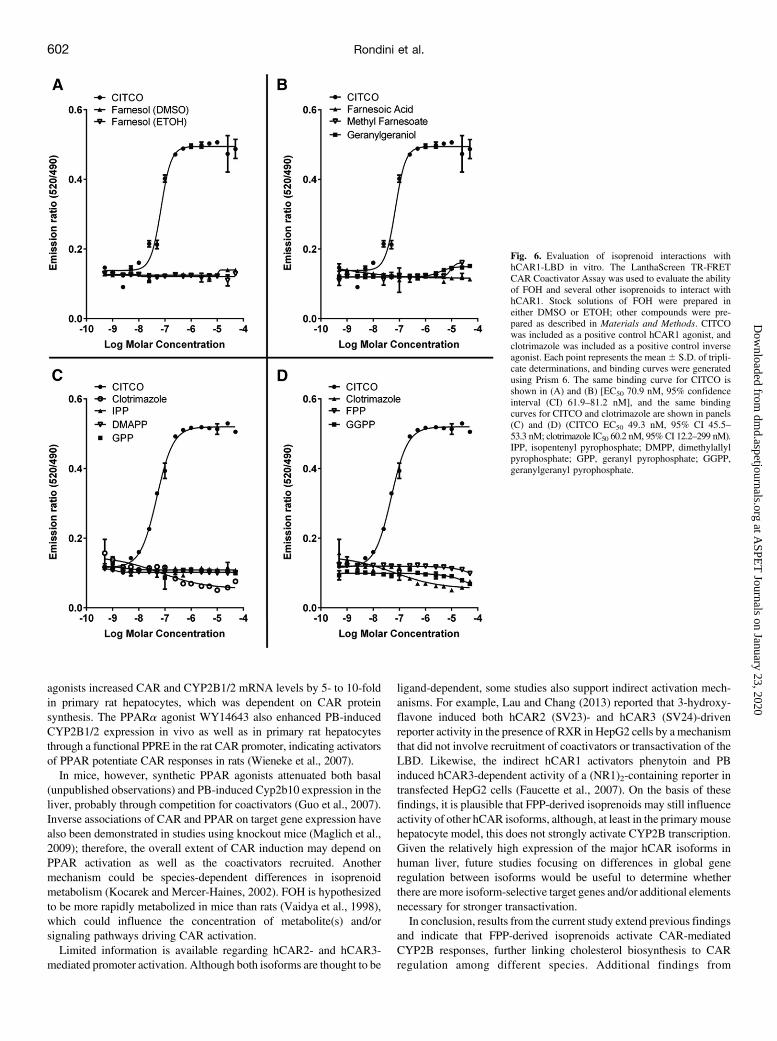
Agonists of hCAR1. The failure of SQ1 treatment to induce asignificant interaction between hCAR1 and SRC1, as was seen forthe CAR ligand agonist CITCO, suggested that SQ1 treatment may notactivate hCAR1 by causing accumulation of an hCAR ligand. To testwhether FOH or several other isoprenoids could interact directly withthe hCAR1 LBD, we used the LanthaScreen TR-FRET hCARCoactivator Assay (Fig. 6). Whereas the positive control hCAR ligandagonist CITCO bound to hCAR1 with an average EC50 of 60.1 nM, andthe hCAR1 inverse agonist clotrimazole bound with an IC50 of 60.2nM, FOH showed no ability to bind to hCAR1 at concentrations up to50 mM, whether added in DMSO or ETOH as the vehicle. Several otherfarnesoids (i.e., methyl farnesoate and farnesoic acid), isoprenoidpyrophosphates (i.e., dimethylallyl pyrophosphate, isopentenyl pyro-phosphate, geranyl pyrophosphate, FPP, and geranylgeranyl pyrophos-phate), and isoprenoid alcohols (geranylgeraniol) were also tested fortheir abilities to bind to hCAR1, and all showed little or no ability tobind (Fig. 6), although at the highest concentrations tested, geranylger-aniol and methyl farnesoate showed slight trends toward agonist-typebinding, whereas FPP and geranylgeranyl pyrophosphate showed slighttrends toward inverse agonist-type binding.
Discussion
Nonsterol isoprenoids are intermediates produced in the cholesterolbiosynthetic pathway that serve as substrates for prenylation reactionsand are precursors for a number of important cellular products (Edwardsand Ericsson, 1999). FOH and geranylgerianol, in particular, arebiologically active molecules that can influence the expression of drug-and lipid-metabolizing enzymes through modulation of nuclear re-ceptor signaling pathways (Kocarek and Mercer-Haines, 2002;
Fig. 2. Relative mRNA levels of the major human CAR splice variants (hCAR1, hCAR2, and hCAR3) and induction of Cyp2b10 mRNA expression by SQ1 treatment inprimary hepatocytes isolated from hCAR-TG mice. Hepatocytes were prepared from hCAR-TG mice and plated onto collagen-coated six-well plates. Forty-eight hours afterplating, cultures were incubated in Williams’ E medium either alone (CON) or containing 0.1% DMSO, 0.1 mM SQ1, 30 mM pravastatin, or the prototypical human CARligand agonist, CITCO (1 mM). Cultures were processed for the measurement of hCAR splice variants and Cyp2b10 and Hmgcr mRNA levels by qRT-PCR 48 hours afterthe initial treatment. (A) SQ1 and CITCO, but not pravastatin, induce Cyp2b10 mRNA levels in primary hepatocytes isolated from hCAR-TG mice. (B) SQ1 and pravastatin,but not CITCO, induce Hmgcr mRNA levels. (C) Levels of the major splice variants of human CAR (hCAR1, hCAR2, hCAR3) expressed relative to hCAR1 mRNA levels.Each bar represents the mean 6 S.E.M. of normalized values from two mouse hepatocyte preparations (n = 2 wells/treatment per preparation). *Significant compared withuntreated (CON) controls (P , 0.05). dSignificantly different compared with hCAR1 mRNA levels (P , 0.05).
Nonsterol Isoprenoids Activate hCAR1 599
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from

Takahashi et al., 2002; Goto et al., 2011a). We previously reportedthat accumulation of isoprenoids in response to squalene synthaseinhibition increased CYP2B1 expression in primary rat hepatocytes in aCAR-dependent manner (Kocarek et al., 1998; Kocarek and Mercer-Haines, 2002). In a continuation of these studies, we investigatedwhether mCAR or any of the major hCAR isoforms are activated bysimilar mechanisms. Using primary hepatocytes from wild-type, CAR-KO, and hCAR-TG mice, we found that squalene synthase inhibitionsignificantly induced Cyp2b10 mRNA levels in mCAR- or hCAR-expressing hepatocytes, whereas this effect was attenuated in theabsence of CAR. Consistent with previous findings (Kocarek andReddy, 1996), pravastatin had no effect on Cyp2b10 expression,although both pravastatin and SQ1 induced Hmgcr mRNA to a similarextent. These findings indicate that the main effects of SQ1 on Cyp2b10induction are mediated through CAR, and that both human and murineCAR are activated by endogenous isoprenoids.The splice variants hCAR2 and hCAR3 are estimated to account for
up to ;10 and 50% of total hCAR transcripts in human liver (Jinno
et al., 2004; Zhang et al., 2013), respectively, which was corroboratedin the current study. The amino acid insertion into hCAR2 is predictedto alter CAR’s ligand binding pocket, thereby modifying the activationprofile (DeKeyser et al., 2009). In comparison, ligand selectivity ofhCAR3 is similar to that of the reference receptor (Omiecinski et al.,2011). In the current study, we evaluated the hCAR isoforms for theability to activate CYP2B reporter activity and found that the effects ofSQ1 were mediated primarily in an hCAR1-dependent manner. Thenormalized activation of hCAR1 by SQ1 was significant and onlyslightly less than that observed with either the ligand agonist CITCO orthe indirect CAR activator PB. In comparison, CITCO did not stronglyinduce CYP2B reporter activity above background in the presence ofeither hCAR2 or hCAR3. Consistent with previous findings (Auerbachet al., 2003; Lau et al., 2011), however, CITCO was capable ofactivating all three receptors in the order hCAR1. hCAR3. hCAR2as indicated by increased interaction with the hSRC1-RID. BothhCAR2 and hCAR3 are reported to have compromised ability toheterodimerize with RXR (Auerbach et al., 2005, 2007). This, together
Fig. 3. SQ1 modulates (A) CYP2B1-PBRE and(B) CYP2B6-PBREM/XREM reporter activityin an hCAR1-dependent manner. Primary hepa-tocytes freshly isolated from CAR-KO micewere plated onto collagen-coated 12-well plates.Twenty-four hours after plating, cultures weretransfected with either a CYP2B1-PBRE or aCYP2B6-PBREM/XREM firefly luciferase re-porter plasmid in combination with either anhCAR expression plasmid or empty vector, asdescribed in Materials and Methods. Forty-eighthours after plating, cultures were incubated inWilliams’ E medium either alone (CON) or con-taining 0.1% DMSO or SQ1 (0.1 or 0.25 mM).The prototypical human CAR activators CITCO(1 mM) and PB (100 mM) were used as positivecontrols. Ninety-six hours after initial plating,cultures were processed for the measurement ofluciferase activity. Each bar represents the mean6S.E.M. of normalized luciferase measurements(firefly/Renilla) combined from four to five hepa-tocyte preparations (n = 3 wells/treatment perhepatocyte preparation). For both reporters, a sig-nificant interaction was detected (P , 0.05). Thefigures show significant treatment effects withinindividual expression plasmids and significantdifferences among the different expression plas-mids. *Significantly different compared with un-treated controls within each expression plasmid(P, 0.05). †Significantly different compared withDMSO controls within each expression plasmid(P , 0.05). Letters denote significant effects ofexpression plasmid on CYP2B reporter activity;groups with different letters are significantlydifferent (P , 0.05).
600 Rondini et al.
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from
with our current findings, indicate weak transactivation potential onCYP2B response elements in the absence of exogenously added RXR(Arnold et al., 2004; Jinno et al., 2004; Auerbach et al., 2005), in thecontext of our mouse primary hepatocyte model.
Several nonsterol isoprenoids are known agonists for nuclearreceptors, including thyroid hormone receptor (FPP), estrogen receptor(FPP), PPARa (FOH, geranylgeraniol), and PPARg (FPP, FOH,geranylgeraniol) (Takahashi et al., 2002; Das et al., 2007; Goto et al.,2011b; Nagai et al., 2011). In the current study, FOH also inducedCYP2B6-PBREM/XREM reporter activity in the presence of eithermCAR or hCAR1; however, SQ1 did not increase the interaction ofhCAR1 with the hSRC1-RID, suggesting that SQ1’s mechanism ofCAR activation differs from that of the ligand agonist CITCO. Weadditionally evaluated the ability of FOH and several other isoprenoidsto bind the hCAR1-LBD in vitro as indicated by association with apeptide representing the RID of a different coactivator (PCG1a).However, none of the compounds functioned as ligand agonists ofhCAR1, further supporting an indirect mechanism of activation. Therelease of CAR from its cytoplasmic complex by either direct or indirectactivators is thought to involve receptor dephosphorylation (Mutohet al., 2009; Yang and Wang, 2014). Recently, PB and severalflavonoids were demonstrated to mediate CAR dephosphorylation byantagonizing epidermal growth factor receptor signaling, indicating thispathway may be a common target for indirect CAR activation in theliver (Mutoh et al., 2013; Carazo Fernandez et al., 2015).The findings that both mCAR and hCAR1 are activated by
isoprenoid accumulation indicate this signaling pathway is conservedacross species, although the magnitude of induction in mouse hepato-cytes was severalfold lower than that previously reported in rathepatocytes (Kocarek et al., 1998; Kocarek and Mercer-Haines,2002). The reason for this difference is not known, but somepossibilities include species differences in basal levels of CAR as wellas functional crosstalk with other transcription factors, such as PPAR(Guo et al., 2007; Cheng and Klaassen, 2008; Wada et al., 2009; Saitoet al., 2010). For example, FOH, geranylgeraniol, and farnesoic acid areactivators of PPARa (O’Brien et al., 1996; Takahashi et al., 2002), andFOH metabolism is at least partially responsible for the triglyceride-lowering properties of squalene synthase inhibitors (Hiyoshi et al.,2003). Saito et al. (2010) reported that several synthetic PPARa
Fig. 4. FOH potentiates hCAR1- and mCAR-mediated activation of theCYP2B6-PBREM/XREM luciferase reporter. Primary hepatocytes from CAR-KO mice were plated onto collagen-coated 12-well plates. Twenty-four hoursafter plating, cultures were transfected with CYP2B6-PBREM/XREM fireflyluciferase reporter plasmid in combination with hCAR1 or mCAR expressionplasmid or empty vector, as described in Materials and Methods. Twenty-fourhours later, cultures were incubated in Williams’ E medium containing 0.05%lipid-free bovine serum albumin (BSA) together with 0.1% ETOH or 60 mMFOH. Luciferase activity was measured 48 hours after the initial treatment. Eachbar represents the mean 6 S.E.M. of normalized luciferase measurements(firefly/Renilla) combined from two to three independent hepatocyte preparations(n = 3 wells/treatment per hepatocyte preparation). *Significantly differentcompared with empty vector controls (P , 0.05). dSignificant compared withETOH-treated hepatocytes (P , 0.05).
Fig. 5. Effect of SQ1 and prototypical CAR activators on hCAR-SRC1 interaction in primary cultured hepatocytes isolated from CAR-KO mice. Primary hepatocytes freshlyisolated from CAR-KO mice were plated onto collagen-coated 12-well plates. Twenty-four hours after plating, cultures were transiently transfected with chimeric plasmidsfor the expression of fusion proteins containing the N-terminal fragment of Gaussia luciferase fused to the entire coding sequence of (A) hCAR1 (hCAR1-Gluc-N), (B)hCAR2 (hCAR2-Gluc-N), or (C) hCAR3 (hCAR3-Gluc-N) together with an expression plasmid encoding the hSRC1 receptor interaction domain (RID) fused to theC-terminal fragment of Gaussia luciferase (hSRC1-Gluc-C), as described in Materials and Methods. Forty-eight hours following initial transfection, cultures were incubatedin Williams’ E medium either alone (CON) or containing DMSO (0.1%), 0.1 mM SQ1, 1 mM CITCO, or 100 mM PB. Culture medium was replaced once after 24 hours, andhepatocytes were harvested for the measurement of luciferase activity 48 hours after initial treatment. Each bar represents the mean 6 S.E.M. of luciferase measurements(Gaussia/firefly) normalized to the untreated control group (CON). Shown are results combined from four independent hepatocyte preparations (n = 3 wells/treatment perhepatocyte preparation). *Significantly different compared with untreated (CON) controls (P , 0.05). †Significantly different compared with DMSO controls (P , 0.05).
Nonsterol Isoprenoids Activate hCAR1 601
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from
agonists increased CAR and CYP2B1/2 mRNA levels by 5- to 10-foldin primary rat hepatocytes, which was dependent on CAR proteinsynthesis. The PPARa agonist WY14643 also enhanced PB-inducedCYP2B1/2 expression in vivo as well as in primary rat hepatocytesthrough a functional PPRE in the rat CAR promoter, indicating activatorsof PPAR potentiate CAR responses in rats (Wieneke et al., 2007).In mice, however, synthetic PPAR agonists attenuated both basal
(unpublished observations) and PB-induced Cyp2b10 expression in theliver, probably through competition for coactivators (Guo et al., 2007).Inverse associations of CAR and PPAR on target gene expression havealso been demonstrated in studies using knockout mice (Maglich et al.,2009); therefore, the overall extent of CAR induction may depend onPPAR activation as well as the coactivators recruited. Anothermechanism could be species-dependent differences in isoprenoidmetabolism (Kocarek and Mercer-Haines, 2002). FOH is hypothesizedto be more rapidly metabolized in mice than rats (Vaidya et al., 1998),which could influence the concentration of metabolite(s) and/orsignaling pathways driving CAR activation.Limited information is available regarding hCAR2- and hCAR3-
mediated promoter activation. Although both isoforms are thought to be
ligand-dependent, some studies also support indirect activation mech-anisms. For example, Lau and Chang (2013) reported that 3-hydroxy-flavone induced both hCAR2 (SV23)- and hCAR3 (SV24)-drivenreporter activity in the presence of RXR in HepG2 cells by amechanismthat did not involve recruitment of coactivators or transactivation of theLBD. Likewise, the indirect hCAR1 activators phenytoin and PBinduced hCAR3-dependent activity of a (NR1)2-containing reporter intransfected HepG2 cells (Faucette et al., 2007). On the basis of thesefindings, it is plausible that FPP-derived isoprenoids may still influenceactivity of other hCAR isoforms, although, at least in the primary mousehepatocyte model, this does not strongly activate CYP2B transcription.Given the relatively high expression of the major hCAR isoforms inhuman liver, future studies focusing on differences in global generegulation between isoforms would be useful to determine whetherthere are more isoform-selective target genes and/or additional elementsnecessary for stronger transactivation.In conclusion, results from the current study extend previous findings
and indicate that FPP-derived isoprenoids activate CAR-mediatedCYP2B responses, further linking cholesterol biosynthesis to CARregulation among different species. Additional findings from
Fig. 6. Evaluation of isoprenoid interactions withhCAR1-LBD in vitro. The LanthaScreen TR-FRETCAR Coactivator Assay was used to evaluate the abilityof FOH and several other isoprenoids to interact withhCAR1. Stock solutions of FOH were prepared ineither DMSO or ETOH; other compounds were pre-pared as described in Materials and Methods. CITCOwas included as a positive control hCAR1 agonist, andclotrimazole was included as a positive control inverseagonist. Each point represents the mean6 S.D. of tripli-cate determinations, and binding curves were generatedusing Prism 6. The same binding curve for CITCO isshown in (A) and (B) [EC50 70.9 nM, 95% confidenceinterval (CI) 61.9–81.2 nM], and the same bindingcurves for CITCO and clotrimazole are shown in panels(C) and (D) (CITCO EC50 49.3 nM, 95% CI 45.5–53.3 nM; clotrimazole IC50 60.2 nM, 95%CI 12.2–299 nM).IPP, isopentenyl pyrophosphate; DMPP, dimethylallylpyrophosphate; GPP, geranyl pyrophosphate; GGPP,geranylgeranyl pyrophosphate.
602 Rondini et al.
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from

coactivator recruitment assays further suggest that the effect ofisoprenoids on mediating CAR responses occurs in an indirect manner.Endogenous isoprenoid levels are known to be influenced by a numberof pharmaceutical drugs, including squalene synthase inhibitors,N-bisphosphonates, as well as statins (Park et al., 2014). Therefore, ourfindings have implications for future studies aimed at evaluating thebiologic effects of these compounds on CAR-mediated signaling pathwaysas well as understanding species differences on hepatocellular physiology.
Acknowledgments
The authors thank Mary Gargano for technical skills in isolating the mouseprimary hepatocytes; and Drs. Curtis Omiecinski (Pennsylvania State Univer-sity), Thomas Chang (University of British Columbia), and James Granneman(Wayne State University) for providing plasmids that were essential for thisstudy.
Authorship ContributionsParticipated in research design: Rondini, Kocarek.Conducted experiments: Rondini, Duniec-Dmuchowski.Performed data analysis: Rondini, Duniec-Dmuchowski, Kocarek.Wrote or contributed to the writing of the manuscript: Rondini, Duniec-
Dmuchowski, Kocarek.
References
Arnold KA, Eichelbaum M, and Burk O (2004) Alternative splicing affects the function andtissue-specific expression of the human constitutive androstane receptor. Nucl Recept 2:1.
Auerbach SS, Dekeyser JG, Stoner MA, and Omiecinski CJ (2007) CAR2 displays unique ligandbinding and RXRalpha heterodimerization characteristics. Drug Metab Dispos 35:428–439.
Auerbach SS, Ramsden R, Stoner MA, Verlinde C, Hassett C, and Omiecinski CJ (2003) Al-ternatively spliced isoforms of the human constitutive androstane receptor. Nucleic Acids Res31:3194–3207.
Auerbach SS, Stoner MA, Su S, and Omiecinski CJ (2005) Retinoid X receptor-alpha-dependenttransactivation by a naturally occurring structural variant of human constitutive androstanereceptor (NR1I3). Mol Pharmacol 68:1239–1253.
Baes M, Gulick T, Choi HS, Martinoli MG, Simha D, and Moore DD (1994) A new orphanmember of the nuclear hormone receptor superfamily that interacts with a subset of retinoicacid response elements. Mol Cell Biol 14:1544–1552.
Bansal VS and Vaidya S (1994) Characterization of two distinct allyl pyrophosphatase activitiesfrom rat liver microsomes. Arch Biochem Biophys 315:393–399.
Bostedor RG, Karkas JD, Arison BH, Bansal VS, Vaidya S, Germershausen JI, Kurtz MM,and Bergstrom JD (1997) Farnesol-derived dicarboxylic acids in the urine of animals treatedwith zaragozic acid A or with farnesol. J Biol Chem 272:9197–9203.
Carazo A and Pávek P (2015) The Use of the LanthaScreen TR-FRET CAR Coactivator Assay inthe Characterization of Constitutive Androstane Receptor (CAR) Inverse Agonists. Sensors(Basel) 15:9265–9276.
Carazo Fernández A, Smutny T, Hyrsová L, Berka K, and Pavek P (2015) Chrysin, baicalein andgalangin are indirect activators of the human constitutive androstane receptor (CAR). ToxicolLett 233:68–77.
Cheng X and Klaassen CD (2008) Perfluorocarboxylic acids induce cytochrome P450 enzymes inmouse liver through activation of PPAR-alpha and CAR transcription factors. Toxicol Sci 106:29–36.
Dallner G and Sindelar PJ (2000) Regulation of ubiquinone metabolism. Free Radic Biol Med 29:285–294.
Das S, Schapira M, Tomic-Canic M, Goyanka R, Cardozo T, and Samuels HH (2007) Farnesylpyrophosphate is a novel transcriptional activator for a subset of nuclear hormone receptors.Mol Endocrinol 21:2672–2686.
DeKeyser JG, Laurenzana EM, Peterson EC, Chen T, and Omiecinski CJ (2011) Selectivephthalate activation of naturally occurring human constitutive androstane receptor splice var-iants and the pregnane X receptor. Toxicol Sci 120:381–391.
DeKeyser JG, Stagliano MC, Auerbach SS, Prabhu KS, Jones AD, and Omiecinski CJ (2009) Di(2-ethylhexyl) phthalate is a highly potent agonist for the human constitutive androstane re-ceptor splice variant CAR2. Mol Pharmacol 75:1005–1013.
Dong B, Saha PK, Huang W, Chen W, Abu-Elheiga LA, Wakil SJ, Stevens RD, Ilkayeva O,Newgard CB, and Chan L, et al. (2009) Activation of nuclear receptor CAR amelioratesdiabetes and fatty liver disease. Proc Natl Acad Sci USA 106:18831–18836.
Dring AM, Anderson LE, Qamar S, and Stoner MA (2010) Rational quantitative structure-activityrelationship (RQSAR) screen for PXR and CAR isoform-specific nuclear receptor ligands.Chem Biol Interact 188:512–525.
Dussault I, Lin M, Hollister K, Fan M, Termini J, Sherman MA, and Forman BM (2002) Astructural model of the constitutive androstane receptor defines novel interactions that mediateligand-independent activity. Mol Cell Biol 22:5270–5280.
Edwards PA and Ericsson J (1999) Sterols and isoprenoids: signaling molecules derived from thecholesterol biosynthetic pathway. Annu Rev Biochem 68:157–185.
Faucette SR, Zhang TC, Moore R, Sueyoshi T, Omiecinski CJ, LeCluyse EL, Negishi M,and Wang H (2007) Relative activation of human pregnane X receptor versus constitutiveandrostane receptor defines distinct classes of CYP2B6 and CYP3A4 inducers. J PharmacolExp Ther 320:72–80.
Gonzalez-Pacanowska D, Arison B, Havel CM, and Watson JA (1988) Isopentenoid synthesis inisolated embryonic Drosophila cells. Farnesol catabolism and omega-oxidation. J Biol Chem263:1301–1306.
Goto T, Kim YI, Funakoshi K, Teraminami A, Uemura T, Hirai S, Lee JY, Makishima M, NakataR, and Inoue H, et al. (2011a) Farnesol, an isoprenoid, improves metabolic abnormalities inmice via both PPARa-dependent and -independent pathways. Am J Physiol Endocrinol Metab301:E1022–E1032.
Goto T, Nagai H, Egawa K, Kim YI, Kato S, Taimatsu A, Sakamoto T, Ebisu S, Hohsaka T,and Miyagawa H, et al. (2011b) Farnesyl pyrophosphate regulates adipocyte functions as anendogenous PPARg agonist. Biochem J 438:111–119.
Guo D, Sarkar J, Suino-Powell K, Xu Y, Matsumoto K, Jia Y, Yu S, Khare S, Haldar K, and RaoMS, et al. (2007) Induction of nuclear translocation of constitutive androstane receptor byperoxisome proliferator-activated receptor alpha synthetic ligands in mouse liver. J Biol Chem282:36766–36776.
Hiyoshi H, Yanagimachi M, Ito M, Yasuda N, Okada T, Ikuta H, Shinmyo D, Tanaka K, KurusuN, and Yoshida I, et al. (2003) Squalene synthase inhibitors suppress triglyceride biosynthesisthrough the farnesol pathway in rat hepatocytes. J Lipid Res 44:128–135.
Honkakoski P, Moore R, Gynther J, and Negishi M (1996) Characterization of phenobarbital-induciblemouse Cyp2b10 gene transcription in primary hepatocytes. J Biol Chem 271:9746–9753.
Honkakoski P, Zelko I, Sueyoshi T, and Negishi M (1998) The nuclear orphan receptor CAR-retinoid X receptor heterodimer activates the phenobarbital-responsive enhancer module of theCYP2B gene. Mol Cell Biol 18:5652–5658.
Horn TL, Long L, Cwik MJ, Morrissey RL, Kapetanovic IM, and McCormick DL (2005)Modulation of hepatic and renal drug metabolizing enzyme activities in rats by subchronicadministration of farnesol. Chem Biol Interact 152:79–99.
Huang W, Zhang J, Chua SS, Qatanani M, Han Y, Granata R, and Moore DD (2003) Induction ofbilirubin clearance by the constitutive androstane receptor (CAR). Proc Natl Acad Sci USA100:4156–4161.
Jackson NM and Kocarek TA (2008) Suppression of CYP2B induction by alendronate-mediatedfarnesyl diphosphate synthase inhibition in primary cultured rat hepatocytes. Drug MetabDispos 36:2030–2036.
Jinno H, Tanaka-Kagawa T, Hanioka N, Ishida S, Saeki M, Soyama A, Itoda M, Nishimura T,Saito Y, and Ozawa S, et al. (2004) Identification of novel alternative splice variants of humanconstitutive androstane receptor and characterization of their expression in the liver. MolPharmacol 65:496–502.
Kachaylo EM, Pustylnyak VO, Lyakhovich VV, and Gulyaeva LF (2011) Constitutive androstanereceptor (CAR) is a xenosensor and target for therapy. Biochemistry (Mosc) 76:1087–1097.
Kobayashi K, Sueyoshi T, Inoue K, Moore R, and Negishi M (2003) Cytoplasmic accumulationof the nuclear receptor CAR by a tetratricopeptide repeat protein in HepG2 cells. Mol Phar-macol 64:1069–1075.
Kocarek TA, Kraniak JM, and Reddy AB (1998) Regulation of rat hepatic cytochrome P450expression by sterol biosynthesis inhibition: inhibitors of squalene synthase are potent inducers ofCYP2B expression in primary cultured rat hepatocytes and rat liver. Mol Pharmacol 54:474–484.
Kocarek TA and Mercer-Haines NA (2002) Squalestatin 1-inducible expression of rat CYP2B:evidence that an endogenous isoprenoid is an activator of the constitutive androstane receptor.Mol Pharmacol 62:1177–1186.
Kocarek TA and Reddy AB (1996) Regulation of cytochrome P450 expression by inhibitors ofhydroxymethylglutaryl-coenzyme A reductase in primary cultured rat hepatocytes and in ratliver. Drug Metab Dispos 24:1197–1204.
Krag SS (1998) The importance of being dolichol. Biochem Biophys Res Commun 243:1–5.Lamba JK, Lamba V, Yasuda K, Lin YS, Assem M, Thompson E, Strom S, and Schuetz E (2004)Expression of constitutive androstane receptor splice variants in human tissues and theirfunctional consequences. J Pharmacol Exp Ther 311:811–821.
Lau AJ and Chang TK (2013) Indirect activation of the SV23 and SV24 splice variants of humanconstitutive androstane receptor: analysis with 3-hydroxyflavone and its analogues. Br JPharmacol 170:403–414.
Lau AJ, Yang G, and Chang TK (2011) Isoform-selective activation of human constitutiveandrostane receptor by Ginkgo biloba extract: functional analysis of the SV23, SV24, andSV25 splice variants. J Pharmacol Exp Ther 339:704–715.
Lynch C, Pan Y, Li L, Heyward S, Moeller T, Swaan PW, and Wang H (2014) Activation of theconstitutive androstane receptor inhibits gluconeogenesis without affecting lipogenesis or fattyacid synthesis in human hepatocytes. Toxicol Appl Pharmacol 279:33–42.
Maglich JM, Lobe DC, and Moore JT (2009) The nuclear receptor CAR (NR1I3) regulates serumtriglyceride levels under conditions of metabolic stress. J Lipid Res 50:439–445.
Maglich JM, Parks DJ, Moore LB, Collins JL, Goodwin B, Billin AN, Stoltz CA, Kliewer SA,Lambert MH, and Willson TM, et al. (2003) Identification of a novel human constitutiveandrostane receptor (CAR) agonist and its use in the identification of CAR target genes. J BiolChem 278:17277–17283.
Maglich JM, Stoltz CM, Goodwin B, Hawkins-Brown D, Moore JT, and Kliewer SA (2002)Nuclear pregnane x receptor and constitutive androstane receptor regulate overlapping butdistinct sets of genes involved in xenobiotic detoxification. Mol Pharmacol 62:638–646.
Maglich JM, Watson J, McMillen PJ, Goodwin B, Willson TM, and Moore JT (2004) The nuclearreceptor CAR is a regulator of thyroid hormone metabolism during caloric restriction. J BiolChem 279:19832–19838.
Muangmoonchai R, Smirlis D, Wong SC, Edwards M, Phillips IR, and Shephard EA (2001)Xenobiotic induction of cytochrome P450 2B1 (CYP2B1) is mediated by the orphan nuclearreceptor constitutive androstane receptor (CAR) and requires steroid co-activator 1 (SRC-1)and the transcription factor Sp1. Biochem J 355:71–78.
Mutoh S, Osabe M, Inoue K, Moore R, Pedersen L, Perera L, Rebolloso Y, Sueyoshi T,and Negishi M (2009) Dephosphorylation of threonine 38 is required for nucleartranslocation and activation of human xenobiotic receptor CAR (NR1I3). J Biol Chem 284:34785–34792.
Mutoh S, Sobhany M, Moore R, Perera L, Pedersen L, Sueyoshi T, and Negishi M (2013)Phenobarbital indirectly activates the constitutive active androstane receptor (CAR) by in-hibition of epidermal growth factor receptor signaling. Sci Signal 6:ra31.
Nagai H, Ebisu S, Abe R, Goto T, Takahashi N, Hosaka T, and Kawada T (2011) Development ofa novel PPARg ligand screening system using pinpoint fluorescence-probed protein. BiosciBiotechnol Biochem 75:337–341.
O’Brien ML, Rangwala SM, Henry KW, Weinberger C, Crick DC, Waechter CJ, Feller DR,and Noonan DJ (1996) Convergence of three steroid receptor pathways in the mediation ofnongenotoxic hepatocarcinogenesis. Carcinogenesis 17:185–190.
Omiecinski CJ, Coslo DM, Chen T, Laurenzana EM, and Peffer RC (2011) Multi-speciesanalyses of direct activators of the constitutive androstane receptor. Toxicol Sci 123:550–562.
Nonsterol Isoprenoids Activate hCAR1 603
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from

Park J, Matralis AN, Berghuis AM, and Tsantrizos YS (2014) Human isoprenoid synthaseenzymes as therapeutic targets. Front Chem 2:50.
Remy I and Michnick SW (2006) A highly sensitive protein-protein interaction assay based onGaussia luciferase. Nat Methods 3:977–979.
Remy I and Michnick SW (2007) Application of protein-fragment complementation assays in cellbiology. Biotechniques 42:137–, 139, 141 passim.
Reschly EJ and Krasowski MD (2006) Evolution and function of the NR1I nuclear hormonereceptor subfamily (VDR, PXR, and CAR) with respect to metabolism of xenobiotics andendogenous compounds. Curr Drug Metab 7:349–365.
Ross J, Plummer SM, Rode A, Scheer N, Bower CC, Vogel O, Henderson CJ, Wolf CR,and Elcombe CR (2010) Human constitutive androstane receptor (CAR) and pregnane Xreceptor (PXR) support the hypertrophic but not the hyperplastic response to the murinenongenotoxic hepatocarcinogens phenobarbital and chlordane in vivo. Toxicol Sci 116:452–466.
Saito K, Kobayashi K, Mizuno Y, Fukuchi Y, Furihata T, and Chiba K (2010) Peroxisome proliferator-activated receptor alpha (PPARalpha) agonists induce constitutive androstane receptor (CAR) andcytochrome P450 2B in rat primary hepatocytes. Drug Metab Pharmacokinet 25:108–111.
Sakakura Y, Shimano H, Sone H, Takahashi A, Inoue N, Toyoshima H, Suzuki S, and Yamada N(2001) Sterol regulatory element-binding proteins induce an entire pathway of cholesterolsynthesis. Biochem Biophys Res Commun 286:176–183.
Savkur RS, Wu Y, Bramlett KS, Wang M, Yao S, Perkins D, Totten M, Searfoss G, 3rd, RyanTP, and Su EW, et al. (2003) Alternative splicing within the ligand binding domain of thehuman constitutive androstane receptor. Mol Genet Metab 80:216–226.
Scheer N, Ross J, Rode A, Zevnik B, Niehaves S, Faust N, and Wolf CR (2008) A novel panel ofmouse models to evaluate the role of human pregnane X receptor and constitutive androstanereceptor in drug response. J Clin Invest 118:3228–3239.
Sueyoshi T, Kawamoto T, Zelko I, Honkakoski P, and Negishi M (1999) The repressed nuclearreceptor CAR responds to phenobarbital in activating the human CYP2B6 gene. J Biol Chem274:6043–6046.
Suino K, Peng L, Reynolds R, Li Y, Cha JY, Repa JJ, Kliewer SA, and Xu HE (2004) Thenuclear xenobiotic receptor CAR: structural determinants of constitutive activation and het-erodimerization. Mol Cell 16:893–905.
Takahashi N, Kawada T, Goto T, Yamamoto T, Taimatsu A, Matsui N, Kimura K, Saito M, HosokawaM, and Miyashita K, et al. (2002) Dual action of isoprenols from herbal medicines on both PPAR-gamma and PPARalpha in 3T3-L1 adipocytes and HepG2 hepatocytes. FEBS Lett 514:315–322.
Tzameli I, Pissios P, Schuetz EG, and Moore DD (2000) The xenobiotic compound 1,4-bis[2-(3,5-dichloropyridyloxy)]benzene is an agonist ligand for the nuclear receptor CAR. Mol CellBiol 20:2951–2958.
Vaidya S, Bostedor R, Kurtz MM, Bergstrom JD, and Bansal VS (1998) Massive production offarnesol-derived dicarboxylic acids in mice treated with the squalene synthase inhibitorzaragozic acid A. Arch Biochem Biophys 355:84–92.
Wada T, Gao J, and Xie W (2009) PXR and CAR in energy metabolism. Trends EndocrinolMetab 20:273–279.
Wang H, Faucette S, Sueyoshi T, Moore R, Ferguson S, Negishi M, and LeCluyse EL (2003) Anovel distal enhancer module regulated by pregnane X receptor/constitutive androstanereceptor is essential for the maximal induction of CYP2B6 gene expression. J Biol Chem 278:14146–14152.
Wang YM, Ong SS, Chai SC, and Chen T (2012) Role of CAR and PXR in xenobiotic sensingand metabolism. Expert Opin Drug Metab Toxicol 8:803–817.
Wieneke N, Hirsch-Ernst KI, Kuna M, Kersten S, and Püschel GP (2007) PPARalpha-dependentinduction of the energy homeostasis-regulating nuclear receptor NR1i3 (CAR) in rat hepato-cytes: potential role in starvation adaptation. FEBS Lett 581:5617–5626.
Wright E, Vincent J, and Fernandez EJ (2007) Thermodynamic characterization of the interactionbetween CAR-RXR and SRC-1 peptide by isothermal titration calorimetry. Biochemistry 46:862–870.
Wu W, Kocarek TA, and Runge-Morris M (2001) Sex-dependent regulation by dexamethasone ofmurine hydroxysteroid sulfotransferase gene expression. Toxicol Lett 119:235–246.
Yang H and Wang H (2014) Signaling control of the constitutive androstane receptor (CAR).Protein Cell 5:113–123.
Yarushkin AA, Kachaylo EM, and Pustylnyak VO (2013) The constitutive androstane receptoractivator 4-[(4R,6R)-4,6-diphenyl-1,3-dioxan-2-yl]-N,N-dimethylaniline inhibits the gluco-neogenic genes PEPCK and G6Pase through the suppression of HNF4a and FOXO1 tran-scriptional activity. Br J Pharmacol 168:1923–1932.
Yoshinari K, Kobayashi K, Moore R, Kawamoto T, and Negishi M (2003) Identification of thenuclear receptor CAR:HSP90 complex in mouse liver and recruitment of protein phosphatase2A in response to phenobarbital. FEBS Lett 548:17–20.
Zhang YK, Lu H, and Klaassen CD (2013) Expression of human CAR splicing variants inBAC-transgenic mice. Toxicol Sci 132:142–150.
Address correspondence to: Dr. Thomas A. Kocarek, Institute of EnvironmentalHealth Sciences, 6135 Woodward Avenue, IBio Building, Room 2126, WayneState University, Detroit, MI 48202. E-mail: [email protected]
604 Rondini et al.
at ASPE
T Journals on January 23, 2020
dmd.aspetjournals.org
Dow
nloaded from