non-target-site based tolerance to herbicides in
TRANSCRIPT

University of Arkansas, FayettevilleScholarWorks@UARK
Theses and Dissertations
5-2018
Non-Target-Site Based Tolerance to Herbicides inAmaranthus palmeriReiofeli Algodon Salas-PerezUniversity of Arkansas, Fayetteville
Follow this and additional works at: http://scholarworks.uark.edu/etd
Part of the Horticulture Commons, Plant Biology Commons, and the Weed Science Commons
This Dissertation is brought to you for free and open access by ScholarWorks@UARK. It has been accepted for inclusion in Theses and Dissertations byan authorized administrator of ScholarWorks@UARK. For more information, please contact [email protected], [email protected].
Recommended CitationSalas-Perez, Reiofeli Algodon, "Non-Target-Site Based Tolerance to Herbicides in Amaranthus palmeri" (2018). Theses andDissertations. 2711.http://scholarworks.uark.edu/etd/2711

Non-Target-Site Based Tolerance to Herbicides in Amaranthus palmeri
A dissertation submitted in partial fulfillment
of the requirements for the degree of
Doctor of Philosophy in Crop, Soil, and Environmental Sciences
by
Reiofeli A. Salas-Perez
Leyte State University
Bachelor of Science in Agricultural Chemistry, 2004
University of Arkansas
Master of Science in Crop, Soil, and Environmental Sciences, 2012
May 2018
University of Arkansas
This dissertation is approved for recommendation to the Graduate Council
_______________________________
Nilda R. Burgos, Ph.D.
Dissertation Director
_______________________________ _______________________________
Robert Scott, Ph.D. Nathan Slaton, Ph.D.
Committee member Committee member
_______________________________ _______________________________
Andronikos Mauromoustakos, Ph.D. Amy Lawton-Rauh, Ph.D.
Committee member Committee member
_______________________________
Franck Dayan, Ph.D.
Ex-officio member

ABSTRACT
Palmer amaranth, one of the most aggressive and damaging broadleaf weeds in the USA,
has evolved resistance to multiple herbicide modes of action. The overall objective of this
research was to elucidate the mechanisms by which Palmer amaranth adapt to herbicide selection
stress. This research aimed (1) to evaluate the efficacy of fomesafen, glufosinate, glyphosate and
trifloxysulfuron to Amaranthus populations; (2) identify candidate genes for endowing tolerance
to glufosinate; (3) investigate the involvement of non-target-site resistance (NTSR) mechanism
in an ALS-resistant population; and (4) to examine the molecular basis of resistance to PPO
inhibitors in Palmer amaranth populations from Arkansas. For objective 1, a total of 124
populations were collected in Arkansas between 2008 and 2015. Overall, 33%, 81%, and 100%
of the populations were resistant to fomesafen, glyphosate, and trifloxysulfuron, respectively.
Thirty percent of the populations were multiple resistant to fomesafen, glyphosate, and
trifloxysulfuron. All populations were controlled >88% by glufosinate. For objective 2, the
transcriptomes of glufosinate-tolerant and –sensitive biotypes were assembled using RNA-Seq.
Thirteen candidate non-target genes were highly expressed in glufosinate-tolerant biotypes,
including glutathione S-transferase (GST), two cytochrome P450, and nine additional genes
related to stress signaling and detoxification. Validation of differential gene expression by
quantitative real-time PCR revealed increased expression CYP72A219 and GST in glufosinate-
treated tolerant biotypes, indicating their involvement in glufosinate tolerance. For objective 3, a
population with cross resistance to multiple ALS-inhibiting herbicides was investigated. Two of
the nine resistant plants harbored Ser653Asn mutation in the ALS gene. Resistant plants that
lacked ALS mutations had elevated levels of CYP81B and GSTF10 genes. This Palmer amaranth
population from Arkansas exhibit both target-site (TS) and NTSR to ALS inhibitors. For

objective 4, resistance to PPO inhibitors was first detected in a population collected in 2011 with
resistance attributed to PPO Gly210 deletion. Several PPO-resistant populations were confirmed
in 2014 and 2015; the majority (55%) of the resistant biotypes carried the same mutation. An
alternative target-site mutation Arg128Gly was also identified in at least one population. Overall,
this research showed that Palmer amaranth has multiple genetic adaptation traits to counteract
the lethal effects of herbicides.

ACKNOWLEDGEMENTS
I am incredibly grateful to my Lord and Saviour Jesus Christ for his provision and unfailing
mercies. I would not have continued the journey if not of His grace and strength that sustained
me throughout my studies.
Sincere thanks to my advisor, Dr. Nilda R. Burgos, for giving me the opportunity to pursue
graduate studies under her guidance. I thank her for the constant encouragement and support she
provided in my entire PhD journey.
I also thank my graduate advisory committee members, Dr. Robert C. Scott, Dr. Nathan Slaton,
Dr. Andy Mauromoustakos, Dr. Amy Lawton-Rauh, and Dr. Franck Dayan for their patience,
support, and constructive comments that contributed towards the improvement of this
manuscript.
Special thanks to all my Weed Physiology colleagues Gulab Rangani, Seth Bernard Abugho, Jun
Estorninos, George Botha, Vijay Singh, Shilpa Singh, Ean Alcober, Hussain Tahir, Ana Claudia
Langaro, Marlon Bastiani, Matheus Noguera, Eduardo Chagas, JP Refatti, Leo Piveta, Claudia
Oliveira, Tiago Kaspary, Fernando Ramirez, Fernando Martini, Caroline Bevilacqua, Ana
Carolina Roso, Christopher Rouse, Teal Penka, Josiane Argenta, Pâmela Carvalho de Lima, and
Sirichai for extending their help in my projects. Their camaraderie made my student life
enjoyable.
I am also grateful to Duyen, Sangeeta, Dennis, Sittie, Samantha and to my KAMPI/FSO,
NWAFAA and Grace Fellowship friends for their unwavering support, the fun-filled memories
and the prayers.

I also thank the various staffs and fellow graduate students of the CSES department for the
assistance and friendly conversations. Finally, I would like to thank Cotton Incorporated and the
Arkansas Soybean Promotion Board for funding my research, as well as BASF for the internship
opportunity.

DEDICATION
This work is dedicated to my husband (Roderic Perez) and my parents (Felix and Rosario Salas)
for their love, prayers, and support throughout the degree program. Also, I thank and pay
gratitude to my siblings and in-laws for their love, support, and trust which kept me moving.
Especially, I would like this opportunity to thank my beloved husband, the wind beneath my
wings, for being there through thick and thin, for never giving up on me, and for constantly
encouraging and motivating me to finish the race triumphantly.

TABLE OF CONTENTS
CHAPTER I .................................................................................................................................. 1
INTRODUCTION .............................................................................................................. 1
Literature Cited ................................................................................................................... 8
CHAPTER II ............................................................................................................................... 11
REVIEW OF LITERATURE ........................................................................................... 11
Palmer amaranth ................................................................................................... 12
Non-target-site based resistance mechanims ........................................................ 14
Glufosinate ............................................................................................................ 17
ALS-inhibiting herbicides ..................................................................................... 19
PPO-inhibiting herbicides ..................................................................................... 22
Literature Cited ................................................................................................................. 24
CHAPTER III ............................................................................................................................. 31
RNA-SEQ TRANSCRIPTOME ANALYSIS OF AMARANTHUS PALMERI WITH
DIFFERENTIAL TOLERANCE TO GLUFOSINATE HERBICIDE ................................ 31
Abstract ............................................................................................................................. 32
Introduction ....................................................................................................................... 34
Materials and Methods ...................................................................................................... 36
Results ............................................................................................................................... 46
Discussion ......................................................................................................................... 53
Literature Cited ................................................................................................................. 64
CHAPTER IV.............................................................................................................................. 95
MULTIPLE RESISTANCE TO ALS, PPO, AND EPSPS INHIBITORS IN
AMARANTHUS PALMERI FROM ARKANSAS, USA.......................................................... 95
ABSTRACT ...................................................................................................................... 96

INTRODUCTION ............................................................................................................ 97
MATERIALS AND METHODs .................................................................................... 100
RESULTS AND DISCUSSION ..................................................................................... 106
REFERENCES ............................................................................................................... 114
CHAPTER V ............................................................................................................................. 128
RESISTANCE TO PPO-INHIBITING HERBICDE IN PALMER AMARANTH FROM
ARKANSAS............................................................................................................................... 128
Abstract ....................................................................................................................................... 129
INTRODUCTION .......................................................................................................... 130
MATERIALS AND METHODS .................................................................................... 132
RESULTS AND DISCUSSION ..................................................................................... 137
REFERENCES ............................................................................................................... 144
CHAPTER VI............................................................................................................................ 152
FREQUENCY OF GLY210 DELETION MUTATION AMONG
PROTOPORHYRINOGEN OXIDASE INHIBITOR-RESISTANT PALMER
AMARANTH (AMARANTHUS PALMERI) POPULATIONS ............................................ 152
Abstract ........................................................................................................................... 153
Introduction ..................................................................................................................... 155
Materials and Methods .................................................................................................... 157
Results and Discussions .................................................................................................. 162
Literature Cited ............................................................................................................... 172
CONCLUSIONS ....................................................................................................................... 189

LIST OF TABLES
CHAPTER III
Table 1. Cluster analysis of A. palmeri accessions treated with glufosinate at 0.55 kg ha-1. ...... 74
Table 2. Response of A. palmeri (08-Lee-C) to foliar-applied herbicides. ................................. 75
Table 3. Glufosinate dose required to reduce growth by 50% (GR50) in A. palmeri 08-Lee-
C, C1 and SS accessions. ............................................................................................ 76
Table 4. Summary of statistics for transcriptome assembly. ....................................................... 77
Table 5. Differentially expressed genes putatively involved in differential tolerance to
glufosinate in Amaranthus palmeri. ............................................................................ 78
Table 6. Upregulated genes in glufosinate-treated and non-treated tolerant (T) plants, and in
glufosinate-treated T relative to treated susceptible (S) plants, assigned with
Gene Ontology molecular function and biological process related to metabolism
and signaling pathways. .............................................................................................. 79
Table 7. Candidate non-target genes, identified by RNA-Seq analysis, that are potentially
involved in conferring differential tolerance to glufosinate in A. palmeri. ................ 82
CHAPTER IV
Table 1. Primers used in the gene expression analysis by qRT-PCR. ........ Error! Bookmark not
defined.
Table 2. Response of 15Cri-B population to the normal dose of ALS (flumetsulam,
imazethapyr, primisulfuron, pyrithiobac, trifloxysulfuron), fomesafen, glyphosate
and glufosinate herbicides.. .......................................... Error! Bookmark not defined.
Table 3. Level of resistance to ALS, PPO, and EPSPS inhibitors in 15Cri-B Palmer
amaranth population..................................................... Error! Bookmark not defined.
Table 4. Effect of pretreatment of P450 inhibitor on the phytotoxicity of trifloxysulfuron (8
g ha-1)in 15Cri-B Palmer amaranth population. ........... Error! Bookmark not defined.
Table 5. Cytochrome P450 genes involved in NTSR to ALS inhibitors in various weed and
crop species………………………………………………………………………….125
CHAPTER V
Table 1. GR50 and LD50 values of original, C1, and C2 AR11-LAW-B populations in
Arkansas, USA. ......................................................................................................... 147
Table 2. Response of Amaranthus palmeri AR11-LAW-B population to foliar-applied
herbicides other than protoporphyrinogen oxidase inhibitors, Arkansas, USA. ....... 148

Table 3. GR50 values and resistance levels to ALS inhibitors in AR11-LAW-B Palmer
amaranth population in Arkansas, USA.................................................................... 149
CHAPTER VI
Table 1. Number of Palmer amaranth accessions in Arkansas, USA tested with foliar-
applied fomesafen at 263 g ha-1. ............................................................................... 175
Table 2. Common, trade names, and manufacturers of herbicides used in the study. .............. 176
Table 3. Response of fomesafen-resistant Palmer amaranth accessions to the recommended
rate of various foliar-applied PPO herbicides. .......................................................... 178
Table 4. Herbicide resistance profile of PPO-resistant Palmer amaranth accessions to other
foliar-applied non-PPO herbicides. ........................................................................... 180
Table 5. GR50 values of PPO-resistant Palmer amaranth accessions in Arkansas. ................... 181

LIST OF FIGURES
CHAPTER III
Figure 1. Shoot biomass reduction (%) of 08-Lee-C, C1, and SS A. palmeri accessions, 21
days after glufosinate treatment. Data were best described with nonlinear, sigmoidal, four-
parameter logistic regression function. ......................................................................................... 83
Figure 2. Ammonia content in glufosinate-tolerant (T) and –susceptible (S) A. palmeri.Error
bars represent standard error. White bars = S plants; gray bars = T plants. ................................. 84
Figure 3. Relative copy number of A. palmeri GS2 in glufosinate-susceptible (S) and -
tolerant (T) plants. Error bars represent standard deviation of the mean. Gray bars = T
plants; black bars = T plant. .......................................................................................................... 85
Figure 4. Multiple alignment of the plastidic glutamine synthetase (GS2) amino acid
sequences in Amaranthus. A. palmeri (reference A. palmeri), T1 and T2 = GS2 alleles of
glufosinate-tolerant A. palmeri biotype, S = GS2 sequence of glufosinate-susceptible A.
palmeri biotype. ............................................................................................................................ 86
Figure 5. Multidimensional scaling (MDS) plot showing the relationship between sample
types.TWO = non-treated tolerant, TWT= treated tolerant, SWO = non-treated susceptible,
SWT = treated susceptible. ........................................................................................................... 87
Figure 6. Volcano plots depicting differential gene expression between treatments. A)
Treated susceptible (S) relative to non-treated S plants (SWT vs SWO), B) Treated tolerant
(T) relative to treated S plants (TWT vs SWT), C) treated T relative to non-treated T plants
(TWT vs TWO), and D) treated T plants relative to non-treated S plants (TWO vs SWO).
The x-axis shows the log fold change or relative abundance. The P value (-log base 10) for
differential gene expression is plotted on the y axis. Black dots represent genes that did not
change in expression; colored dots on the left indicate genes with significantly
downregulated expression; colored dots on the right indicate genes with significantly
upregulated expression.................................................................................................................. 88
Figure 7. The number of differentially expressed genes common or specific to treated and
non-treated T and S plants.A 4-way Venn diagram depicting the distribution of differentially
expressed genes across all pairwise comparisons. The number within each shaded area is the
number of differentially expressed genes common between treatments. ..................................... 89
Figure 8. Biological processes (A) and molecular functions (B) of upregulated genes in
treated T relative to treated S plants. ............................................................................................ 90
Figure 9. Biological processes (A) and molecular functions (B) of downregulated genes in
treated T relative to treated S plants. ............................................................................................ 91
Figure 10. Biological processes (A) and molecular functions (B) of differentially expressed
genes that are common in treated T relative to treated S and non-treated T plants. ..................... 92

Figure 11. Heat map analysis of genes that are putatively related to abiotic stress response in
A. palmeri.TWO (non-treated tolerant), SWO (non-treated susceptible), TWT (glufosinate-
treated tolerant), SWT (glufosinate-treated susceptible). ............................................................. 93
Figure 12. Gene expression fold-change of seven candidate NTS genes in glufosinate-
tolerant Amaranthus palmeri relative to –susceptible ones. Tolerant and susceptible plants
from two populations were identified based on the level of injury incurred after glufosinate
treatment.GST and CYP72A219 was highly expressed in the tolerant biotypes in at least one
population. Untreated = before glufosinate treatment, Treated = 24 h after glufosinate
treatment. Error bars represent standard error from six biological replicates. HSP = heat
shock protein, GST = glutathione S-transferase, NMO = nitronate monooxygenase, ETF =
ethylene-responsive transcription factor, ABC = ABC transporter, NAC = NAC
transcription factor, CYP72A219 = cytochrome P450 CYP72A219. .......................................... 94
CHAPTER IV
Figure 1. Amino acid sequence alignment of acetolactate synthase (ALS) in sensitive (SS)
and resistant (15Cri-B) Palmer amaranth plant samples.Two resistant plants harbored
Ser653Asn (AGC to AAC) mutation. .............................................. Error! Bookmark not defined.
Figure 2. Gene expression analysis of ALS and NTSR genes in trifloxysulfuron-resistant
(15Cri-B) and –susceptible (SS) Palmer amaranth biotypes. Fold-change (2-ΔCt) in gene
expression was calculated relative to the susceptible samples, where ΔCt = [Ct target-gene -
Ct mean of internal control genes]. Means and standard errors from three biological
replicates are shown.. ..................................................................... Error! Bookmark not defined.
CHAPTER V
Figure 1. Shoot biomass reduction (%) of PPO-resistant and –susceptible Palmer amaranth
population, 21 d after fomesafen treatment.Treatment means (n = 10) + 1 standard error are
plotted and fitted to a regression curve. Data were best described with a nonlinear,
sigmoidal, three-parameter Gompertz regression function, y =a*exp{-exp[-b*(x-c0]}. ........... 150
Figure 2. Dose-response curves of PPO-resistant and –susceptible Palmer amaranth
population from Arkansas. Mortality (%) were obtained at 21 d after herbicide treatment.
Treatment means (n = 10) + 1 standard error are plotted and fitted to a regression curve.
Data were best described with a nonlinear, sigmoidal, three-parameter Gompertz regression
function, y =a*exp{-exp[-b*(x-c0]}. .......................................................................................... 151
CHAPTER VI
Figure 1. Variability in response to foliar-applied fomesafen (263 g ha-1) among Palmer
amaranth accessions collected between 2008 and 2015. Box plot shows median values

(horizontal line inside the box), mean values (marked X), first and third quartile values (box-
outlines), minimum and maximum values (whiskers), and outlier values (open circles). .......... 182
Figure 2. Frequency of fomesafen-resistant plants in Palmer amaranth accessions collected
in 2015 from Arkansas. Fomesafen at 263 g ha-1 was applied with 0.5% non-ionic surfactant
to 7.6-cm tall seedlings. Survivors are categorized based on visible injury. .............................. 183
Figure 3. Hierarchal clustering of 2014 and 2015 Palmer amaranth accessions treated with
foliar-applied fomesafen at 263 g ha-1.The herbicide was applied with 0.5% non-ionic
surfactant to 7.6-cm tall seedlings. Cluster 1 = most sensitive to fomesafen (50 accessions),
Cluster 2 = resistant to fomesafen with majority of survivors incurred >60% injury (19
accessions), Cluster 3 = resistant to fomesafen, majority of survivors sustained 11-30%
injury (2 accessions), Cluster 4 = resistant to fomesafen, majority of survivors incurred 0-
10% injury (3 accessions). .......................................................................................................... 184
Figure 4. (A) Cluster analysis of mortality levels in Palmer amaranth accessions collected in
2014 and 2015 treated with 870 g ha-1 glyphosate. Cluster 1 (n = 13 accessions; resistant to
glyphosate with majority of survivors incurred 31-60% injury), cluster 2 (n = 23 accessions;
resistant to glyphosate with majority of survivors incurred <11% injury), cluster 3 (n = 23
accessions; least sensitive to glyphosate), and cluster 4 (n = 14 accessions; resistant to
glyphosate with majority of its survivors incurred 61-89% injury) are depicted in box-plot.
Glyphosate was applied to 7.6-cm tall seedlings. (B) Variability in response to dicamba (280
g ae ha-1), glufosinate (549 g ha-1) and trifloxysulfuron (7.84 g ha-1) among Palmer amaranth
accessions collected in 2014 and 2015 from Arkansas, USA. Box plot shows median values
(horizontal line inside the box), mean values (marked with X), first and third quartile values
(box-outlines), minimum and maximum values (whiskers), and outlier values (open circles). . 185
Figure 5. Herbicide resistance profiles of Palmer amaranth populations from Arkansas
sampled in 2014 and 2015. ......................................................................................................... 186
Figure 6. Counties with confirmed PPO-resistant Palmer amaranth in Arkansas.Counties that
are shaded had at least one field-population with PPO-resistant Palmer amaranth biotypes
carrying the Gly210 deletion in the PPO gene.............................................................................. 187
Figure 7. Prevalence of PPO Gly210 mutation in PPO-resistant Palmer amaranth from
Arkansas. (A) Pie-chart displaying the percentage of PPO-resistant accessions that contained
and lacked the PPO Gly210 mutation. Black = all fomesafen survivors in these accessions
contained the Gly210 mutation. White = all survivors in these accessions lacked the Gly210
mutation. Gray = survivors in these accessions contained mixture of carriers and non-
carriers of Gly210 mutation. (B) Proportion of Gly210 mutation carriers in each PPO-resistant
accession. .................................................................................................................................... 188

LIST OF PUBLISHED ARTICLES
CHAPTER III
Reiofeli A. Salas, Christopher A. Saski, Rooksana E. Noorai, Subodh K. Srivastava, Amy
Lawton-Rauh, Robert L. Nichols, and Nilda R. Burgos. 2018. RNA-Seq transcriptome analysis
of Amaranthus palmeri with differential tolerance to glufosinate herbicide. PLoS ONE
(submitted)----------------------------------------------------------------------------------------------------31
CHAPTER V
Reiofeli A. Salas, Nilda R. Burgos, Patrick J. Tranel, Shilpa Singh, Les Glasgow, Robert C. Scott
and Robert L. Nichols. 2016. Resistance to PPO-inhibiting herbicide in Palmer amaranth from
Arkansas. Pest Management Science 72: 864-869. DOI: 10.1002/ps.4241------------------------128
CHAPTER V1
Reiofeli A. Salas-Perez, Nilda R. Burgos, Gulab Rangani, Shilpa Singh, Joao Paulo Refatti,
Leonard Piveta, Patrick J. Tranel, Andy Mauromoustakos, and Robert C. Scott. 2017. Frequency
of Gly-210 deletion mutation among protoporphyrinogen oxidase inhibitor-resistant Palmer
amaranth (Amaranthus palmeri) populations. Weed Science 65: 718-731. DOI:
10.1017/wsc.2017------------------------------------------------------------------------------------------152

1
CHAPTER I
INTRODUCTION

2
Introduction
Palmer amaranth (Amaranthus palmeri S Watson) is one of the most common,
troublesome, and economically damaging agronomic weeds throughout the southern US (Ward
et al., 2013). The competitive ability of Palmer amaranth is attributed to its fast growth rate (Jha
et al., 2008), high fecundity (Keeley et al., 1987), good light interception, and high water use
efficiency (Ehleringer, 1983). With estimates of over 600,000 seeds per plant, it can replenish the
seedbank tremendously (Keeley et al., 1987). Because it competes effectively with crops for
nutrients, water, light and space, it can significantly reduce crop yield. Palmer amaranth densities
of 8 and 9 plants/m2 can reduce soybean (Glycine max) yield by 78% (Bensch et al., 2003) and
corn (Zea mays) grain yield by 91% (Massinga et al., 2001), respectively. Palmer amaranth
interference for 63 d after crop emergence can cause 77% yield loss in cotton (Gossypium
hirsutum) (Fast et al., 2009). Apart from reducing yield, heavy infestation of Palmer amaranth
interfere with crop harvest and increased harvesting time by two- to four-fold (Smith et al.,
2000).
Palmer amaranth control has become a challenge because of limited herbicide resources
and its remarkable tendency to evolve herbicide resistance. As a dioecious species, Palmer
amaranth is outcrossing, allowing herbicide resistance to spread rapidly (Steckel, 2007).
Glyphosate resistance trait can be transferred up to 300 m through pollen from glyphosate-
resistant males to glyphosate-susceptible female plants (Sosnoskie et al., 2012). To date, Palmer
amaranth has been confirmed resistant to six different modes of action: acetolactate synthase
(ALS) inhibitors, dinitroanilines, triazine, glyphosate, hydroxyphenylpyruvate dioxygenase
(HPPD) inhibitors, and most recently to protoporphyrinogen oxidase (PPO) inhibitors (Heap
2018). This narrows down available control option for resistant populations. PPO herbicides like

3
Valor and Reflex are intensively used to combat herbicide-resistant Palmer amaranth. Losing
these herbicides to resistance problems would severely hamper efforts to manage weeds.
Upcoming herbicide-resistant (HR) crop technology with resistance to glyphosate and dicamba,
to glufosinate, to bleacher herbicides and other trait combinations are being developed. However,
several weed species already reported resistance to some of these herbicides (Heap 2018).
Knowing the response of Palmer amaranth populations to alternative herbicides in HR crops will
refine herbicide recommendations for resistance management.
Herbicide resistance is the inherited ability of the plant to survive and reproduce
following exposure to a dose of herbicide that would normally be lethal to the wild type (WSSA,
1998). Resistance is essentially a natural phenomenon which occurs spontaneously in weed
populations, but is only noticed when a selection pressure is applied to the weeds through the
application of a herbicide. Resistance to herbicides is the result of weed evolutionary adaptation
to herbicide application (Delye, 2013). There are two primary mechanisms of herbicide
resistance in weeds: target-site and non-target-site resistance. Target-site resistance (TSR) is
caused by alteration of the gene encoding the target protein of the herbicide. Non-target-site
resistance (NTSR) is due to mechanism(s) that minimize the amount of herbicide reaching the
target site (Powles and Yu, 2010).This includes decreased herbicide penetration and
translocation, increased herbicide sequestration, and metabolism. A study reported that rigid
ryegrass resistance to nine modes of action is attributed to NTSR (Burnet et al., 1994). Previous
research has indicated that low-dose herbicide application tend to promote non-target-site
metabolic resistance (Gardner et al., 1998). Compared with target-site resistance, non-target-site
herbicide resistance poses a greater threat because it can confer unpredictable resistance to
herbicides with various modes of action and the multi-gene involvement in the mechanism (Petit

4
et al., 2010). Despite the importance of NTSR in understanding herbicide resistance evolution,
the genetic determinants of NTSR are poorly understood because of the inherently complicated
biochemical processes and the limited genomic information available in weedy species (Delye,
2013; Yuan et al., 2007). However, the recent advances in genomics technologies will enable
identification of specific genes that mediate NTSR mechanism.
Acetolactate synthase inhibitors have been widely used for Palmer amaranth control since
their introduction in 1982 (Gaeddert et al., 1997). Inhibition of ALS enzyme affects the synthesis
of branched-chain amino acids (valine, leucine, and isoleucine), ultimately leading to plant death.
There are five chemical families of herbicide that inhibit ALS: sulfonylureas (SUs),
imidazolinones (IMIs), triazolopyrimidines (TPs), sulfonylaminocarbonyltriazolinones (SCTs),
pyrimidinylthiobenzoates (PTBs). The ALS herbicides are widely used because they provide
broad-spectrum weed control at low doses, soil residual activity, wide application windows,
excellent crop safety and low mammalian toxicity. However, these herbicides have high
propensity to select resistant weed populations due to its widespread usage, strong selection
pressure, and high levels of natural variability in the ALS gene (Tranel and Wright, 2002). To
date, 160 species including Palmer amaranth are resistant to ALS-inhibiting herbicides (Heap
2018). Populations of Palmer amaranth resistant to ALS inhibitors are prevalent and have been
identified in 14 states in the US, including Arkansas, Arizona, Delaware, Florida, Georgia,
Illinois, Kansas, Maryland, Michigan, Mississippi, Tennessee, North Carolina, South Carolina,
and Wisconsin (Nandula et al., 2012; Salas-Perez et al., 2017; Sosnoskie et al., 2011). Resistance
is frequently attributed to target-site mutation. However, various studies have reported the
presence of both target-site and non-target-site resistance in some populations of Palmer

5
amaranth, Spanish corn poppy (Papaver rhoeas) and flixweed (Descurainia Sophia L.)
populations (Nakka et al., 2017; Rey-Caballero et al., 2017; Yang et al., 2016).
The widespread distribution of glyphosate-resistant weeds compelled farmers to use
alternative herbicides including another non-selective herbicide, glufosinate, to control
herbicide-resistant weeds in glufosinate-tolerant crops. Glufosinate is a fast-acting
postemergence herbicide that controls weeds by inhibiting glutamine synthetase (GS) (E.C.
6.3.1.2), the enzyme that converts glutamate and ammonia to glutamine (Wild and
Manderscheid, 1984). Inhibition of GS by glufosinate leads to ammonia accumulation, inhibition
of amino acid synthesis, and indirect inhibition of photosynthesis, ultimately leading to plant
death (Tachibana et al., 1986). To date, resistance to glufosinate has been confirmed in
goosegrass (Eleusine indica) from Malaysia (Jalaludin et al., 2010; Seng et al., 2010) and Italian
ryegrass (Lolium perenne ssp. multiflorum) from Oregon (Avila-Garcia and Mallory-Smith,
2011). An amino acid mutation in the chloroplast-encoded GS gene, Asp171Asn, conferred
resistance to glufosinate in Italian ryegrass (Avila-Garcia et al., 2012). Resistance to glufosinate
in glufosinate-resistant crops is achieved using transgenic methods to insert the bar or pat
(phosphinothricin acetyltransferase) gene from a bacterium to the plant’s genome, allowing
detoxification of glufosinate by acetylation (Droge-Laser et al., 1994).
PPO-inhibiting herbicides became widely used during 1980-1990. However, after
introduction of glyphosate-resistant crops, usage of PPO-herbicides declined until 2006 when
economically damaging weeds evolved resistance to glyphosate (USDA-NASS). These current
situations in agriculture reinforce the high interest in PPO-inhibiting herbicides in recent years
(Dayan et al., 2010; Salas et al., 2016; Salas-Perez et al., 2017). The PPO enzyme catalyzes the
oxidation of protoporphyrinogen IX (protogen) to protoporphyrin IX (proto), the last common

6
intermediate for the tetrapyrole synthesis system (Becerril and Duke, 1989; Jacobs et al., 1991).
Several classes of PPO-inhibiting herbicides such as diphenyl ethers, thiadiazoles, oxadiazoles,
triazolinones, N-phenyl-phthalimides, and pyrimidinediones inhibit the PPO enzyme. Inhibition
of PPO results in the generation of singlet oxygen species that oxidize lipid and protein
membranes, leading to plant death (Sherman et al., 1991). However, a few species of weeds have
slowly evolved resistance from the repeated use of these herbicides in past few years. To date, 11
weed species have evolved resistance against PPO-herbicides including Palmer amaranth (Heap
2018; Salas et al., 2016; Salas-Perez et al., 2017;Giacomini et al., 2017). Many molecular and
physiological mechanisms underlie the PPO-herbicide resistance, but most often, the evolution
of resistance attributed to target-site mutations (Dayan et al., 2010; Edwards, 1996; Skipsey et
al., 1997; Thinglum et al., 2011; Wuerffel et al., 2015).
Palmer amaranth control is becoming more difficult due to its adaptability, high seed
production and resistance to many herbicides used for its management. With the evolution of
Palmer amaranth populations that are resistant to herbicides of different modes of action, new
approaches should be implemented to control and reduce the frequency of herbicide-resistant
weeds. Understanding the molecular mechanisms endowing herbicide resistance will contribute
to wiser use of herbicide resources and enable innovations that, together with integrated control
strategies, will help minimize and manage herbicide-resistance evolution (Powles and Yu, 2010).
An understanding of the fundamental molecular mechanisms behind herbicide resistance is
necessary to minimize and manage resistance evolution and increase crop yield. With the recent
advances in next-generation sequencing, tools are available for characterizing genome-wide gene
expression in Palmer amaranth using transcriptomics, which is helpful in gene discovery and
understanding the molecular mechanisms involved in plant response to herbicides and herbicide

7
resistance evolution. The goal of this study is to elucidate the mechanisms by which Palmer
amaranth (or other weeds) adapt to herbicide selection stress. The specific objectives of this
research are to (1) evaluate the efficacy of foliar-applied fomesafen, glufosinate, glyphosate and
trifloxysulfuron to Amaranthus accessions collected between 2008 and 2015 in Arkansas; (2)
identify candidate genes for endowing tolerance to glufosinate; (3) investigate the involvement
of non-target-site resistance mechanism in an ALS-resistant population; and (4) to examine the
molecular basis of resistance to PPO inhibitors in Palmer amaranth populations from Arkansas.

8
Literature Cited
Avila-Garcia, W. V., Mallory-Smith, C. 2011. Glyphosate-resistant Italian ryegrass (Lolium
perenne) populations also exhibit resistance to glufosinate. Weed Science 59: 305-309.
Avila-Garcia, W. V., Sanchez-Olguin, E., Hulting, A. G., Mallory-Smith, C. 2012. Target-site
mutation associated with glufosinate resistance in Italian ryegrass (Lolium perenne L. ssp
multiflorum). Pest Management Science 68: 1248-1254.
Becerril, J. M., Duke, S. O. 1989. Protoporphyrin IX content correlates with activity of
photobleaching herbicides. Plant Physiol 90: 1175-81.
Bensch, C. N., Horak, M. J., Peterson, D. 2003. Interference of redroot pigweed (Amaranthus
retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A. rudis) in
soybean. Weed Science 51: 37-43.
Burnet, M. W. M., Hart, Q., Holtum, J. A. M., Powles, S. B. 1994. Resistance to nine herbicide
classes in a population of rigid ryegrass (Lolium rigidum). Weed Science 42: 369-377.
Dayan, F. E. et al. 2010. Biochemical and structural consequences of a glycine deletion in the
alpha-8 helix of protoporphyrinogen oxidase. Biochimica Et Biophysica Acta-Proteins
and Proteomics 1804: 1548-1556.
Delye, C. 2013. Unravelling the genetic bases of non-target-site-based resistance (NTSR) to
herbicides: A major challenge for weed science in the forthcoming decade. Pest
Management Science 69: 176-187.
Droge-Laser, W., Siemeling, U., Puhler, A., Broer, I. 1994. The metabolites of the herbicide L-
phosphinothricin (glufosinate): Identification, stability, and mobility in transgenic,
herbicide-resistant, and untransformed plants. Plant Physiology 105: 159-166.
Edwards, R. 1996. Characterisation of glutathione transferases and glutathione peroxidases in
pea (Pisum sativum). Physiologia Plantarum 98: 594-604.
Ehleringer, J. 1983. Ecophysiology of Amaranthus palmeri, a sonoran desert summer annual.
Oecologia 57: 107-112.
Heap I. The international survey of herbicide resistant weeds. 2018.
http://www.weedscience.com. Accessed 22 January 2018.
Fast, B. J., Murdoch, S. W., Farris, R. L., Willis, J. B., Murray, D. S. 2009. Critical timing of
Palmer amaranth (Amaranthus palmeri) removal in second-generation glyphosate-
resistant cotton. The Journal of Cotton Science 13: 32-36.
Gaeddert, J. W., Peterson, D. E., Horak, M. J. 1997. Control and cross-resistance of an
acetolactate synthase inhibitor-resistant Palmer amaranth (Amaranthus palmeri) biotype.
Weed Technology 11: 132-137.
Gardner, S. N., Gressel, J., Mangel, M. 1998. A revolving dose strategy to delay the evolution of
both quantitative vs major monogene resistances to pesticides and drugs. International
Journal of Pest Management 44: 161-180.
Giacomini, D. A. et al. 2017. Two new PPX2 mutations associated with resistance to PPO-
inhibiting herbicides in Amaranthus palmeri. Pest Management Science 73: 1559-1563.

9
Jacobs, J. M., Jacobs, N. J., Sherman, T. D., Duke, S. O. 1991. Effect of diphenyl ether
herbicides on oxidation of protoporphyrinogen to protoporphyrin in organellar and
plasma membrane enriched fractions of barley. Plant Physiology 97: 197-203.
Jalaludin, A., Ngim, J., Bakar, B. H. J., Alias, Z. 2010. Preliminary findings of potentially
resistant goosegrass (Eleusine indica) to glufosinate-ammonium in Malaysia. Weed
Biology and Management 10: 256-260.
Jha, P., Norsworthy, J. K., Riley, M. B., Bielenberg, D. G., Bridges, W., Jr. 2008. Acclimation of
Palmer amaranth (Amaranthus palmeri) to shading. Weed Science 56: 729-734.
Keeley, P. E., Carter, C. H., Thullen, R. J. 1987. Influence of planting date on growth of Palmer
amaranth (Amaranthus palmeri). Weed Science 35: 199-204.
Massinga, R. A., Currie, R. S., Horak, M. J., Boyer, J. 2001. Interference of Palmer amaranth in
corn. Weed Science 49: 202-208.
Nakka, S., Thompson, C. R., Peterson, D. E., Jugulam, M. 2017. Target site-based and non-target
site based resistance to ALS inhibitors in Palmer amaranth (Amaranthus palmeri). Weed
Science 65: 681-689.
Nandula, V. K. et al. 2012. Multiple resistance to glyphosate and pyrithiobac in Palmer amaranth
(Amaranthus palmeri) from Mississippi and response to flumiclorac. Weed Science 60:
179-188.
Petit, C., Duhieu, B., Boucansaud, K., Delye, C. 2010. Complex genetic control of non-target-
site-based resistance to herbicides inhibiting acetyl-coenzyme A carboxylase and
acetolactate-synthase in Alopecurus myosuroides Huds. Plant Science 178: 501-509.
Powles, S. B., Yu, Q. 2010. Evolution in Action: Plants Resistant to Herbicides. Pages 317-347
in S. Merchant, W.R. Briggs, D. Ort, eds. Annual Review of Plant Biology, Vol 61.
Rey-Caballero, J., Menéndez, J., Osuna, M. D., Salas, M., Torra, J. 2017. Target-site and non-
target-site resistance mechanisms to ALS inhibiting herbicides in Papaver rhoeas.
Pesticide Biochemistry and Physiology 138: 57-65.
Salas, R. A. et al. 2016. Resistance to PPO-inhibiting herbicide in Palmer amaranth from
Arkansas. Pest Management Science 72: 864-869.
Salas-Perez, R. A. et al. 2017. Frequency of Gly-210 deletion mutation among
protoporphyrinogen oxidase inhibitor–resistant Palmer amaranth (Amaranthus palmeri)
populations. Weed Science 65: 718-731.
Seng, C. T., Van Lun, L., San, C. T., Bin Sahid, I. 2010. Initial report of glufosinate and paraquat
multiple resistance that evolved in a biotype of goosegrass (Eleusine indica) in Malaysia.
Weed Biology and Management 10: 229-233.
Sherman, T. D. et al. 1991. Physiological-basis for differential sensitivities of plant-species to
protoporphyrinogen oxidase-inhibiting herbicides. Plant Physiology 97: 280-287.
Skipsey, M., Andrews, C. J., Townson, J. K., Jepson, I., Edwards, R. 1997. Substrate and thiol
specificity of a stress-inducible glutathione transferase from soybean. FEBS Letters 409:
370-374.

10
Smith, D. T., Baker, R. V., Steele, G. L. 2000. Palmer amaranth (Amaranthus palmeri) impacts
on yield, harvesting, and ginning in dryland cotton (Gossypium hirsutum). Weed
Technology 14: 122-126.
Sosnoskie, L. M., Kichler, J. M., Wallace, R. D., Culpepper, A. S. 2011. Multiple resistance in
Palmer amaranth to glyphosate and pyrithiobac confirmed in Georgia. Weed Science 59:
321-325.
Sosnoskie, L. M. et al. 2012. Pollen-mediated dispersal of glyphosate-resistance in Palmer
amaranth under field conditions. Weed Science 60: 366-373.
Steckel, L. E. 2007. The dioecious Amaranthus spp.: Here to stay. Weed Technology 21: 567-
570.
Tachibana, K., Watanabe, T., Sekizawa, Y., Takematsu, T. 1986. Action mechanism of bialaphos
2: Accumulation of ammonia in plants treated with bialaphos. Journal of Pesticide
Science 11: 33-37.
Thinglum, K. A. et al. 2011. Wide Distribution of the waterhemp (Amaranthus tuberculatus)
delta G210 PPX2 mutation, which confers resistance to PPO-inhibiting herbicides. Weed
Science 59: 22-27.
Tranel, P. J., Wright, T. R. 2002. Resistance of weeds to ALS-inhibiting herbicides: What have
we learned? Weed Science 50: 700-712.
USDA-NASS. US Department of Agriculture, National Statistics Service.www.nass.usda.gov
[accessed March 2018]
Ward, S. M., Webster, T. M., Steckel, L. E. 2013. Palmer amaranth (Amaranthus palmeri): A
review. Weed Technology 27: 12-27.
Wild, A., Manderscheid, R. 1984. The effect of phosphinothricin on the assimilation of ammonia
in plants. Zeitschrift Fur Naturforschung C-a Journal of Biosciences 39: 500-504.
WSSA. 1998. Herbicide resistance and tolerance definitions. Weed Technology 12:789.
Wuerffel, R. J. et al. 2015. Distribution of the delta G210 protoporphyrinogen oxidase mutation
in Illinois waterhemp (Amaranthus tuberculatus) and an improved molecular method for
detection. Weed Science 63: 839-845.
Yang, Q. et al. 2016. Target-site and non-target-site based resistance to the herbicide tribenuron-
methyl in flixweed (Descurainia sophia L.). BMC Genomics 17: 1-13.
Yuan, J. S., Tranel, P. J., Stewart, C. N. 2007. Non-target-site herbicide resistance: a family
business. Trends in Plant Science 12: 6-13.

11
CHAPTER II
REVIEW OF LITERATURE

12
Review of Literature
Palmer amaranth
Palmer amaranth (Amaranthus palmeri S Watson, subgenus Acnida, subsection
Saueranthus) is one of the most common and problematic weed species in corn (Zea mays),
cotton (Gossypium hirsutum), grain sorghum (Sorghum bicolor), peanut (Arachis hypogea), and
soybean (Glycine max) in the US. It is described as a tall (usually about 2.5 m tall), erect,
branched summer annual plant with leaves arranged alternately on the stems. The pistillate and
staminate flowers occur in separate plants (dioecious species) in long, narrowly elongated
terminal inflorescence. The female inflorescences have sharp bracts which are distinct from male
inflorescences, which are soft to touch. Because it is a dioecious species, Palmer amaranth is an
obligate outcrosser (Ward et al., 2013). Palmer amaranth can also hybridize, to a limited extent,
with spiny amaranth, which increases the risk of spreading herbicide resistance to other
Amaranthus species (Gaines et al., 2012).
Palmer amaranth is a C4 dicot with high rates of photosynthesis (81 μmol m-2 s-1)
compared to other C4 monocots such as corn and C3 dicots like cotton and soybean (Ehleringer,
1983; Gibson, 1998). Under ideal conditions, Palmer amaranth can grow up to 5 cm daily.
(Horak and Loughin, 2000). Temperature plays an important role in the net rate of
photosynthesis in Palmer amaranth, and optimum photosynthesis occurs between 36-46°C
(Ehleringer, 1983). Furthermore, Palmer amaranth exhibits diaheliotropism, meaning the plant is
capable of tracking sunlight where the leaves orient themselves perpendicularly to the sun rays to
maximize light interception and thus photosynthesis (Ehleringer and Forseth, 1980).
Infestation of Palmer amaranth significantly affects growth and yield of most agronomic
crops, cotton being one of the most sensitive commodities. Cotton yield decreased by 5.38% for

13
every one additional Palmer amaranth plant per 9.1 m of row (Morgan et al., 2001). Palmer
amaranth interference can cause significant yield reduction in soybean (Bensch et al., 2003), corn
(Massinga et al., 2001), peanut (Burke et al., 2007), and sweet potato (Ipomoea batatas) (Meyers
et al., 2010) crop productions. The success of Palmer amaranth as troublesome weed is attributed
to its extended emergence period (Jha and Norsworthy, 2009), high fecundity (Keeley et al.,
1987), rapid growth rate (Jha et al., 2008) with a C4 photosynthetic pathway (Ehleringer, 1983),
genetic diversity, ability to tolerate adverse conditions, and its facility for evolving herbicide
resistance (Ward et al., 2013).
Palmer amaranth can be controlled by timely applications of preemergence (PRE) and
postemergence (POST) herbicides. Some of the PRE herbicides labeled on different crops for
Palmer amaranth control are diuron (e.g. corn, cotton, grain sorghum), fluometuron (e.g. cotton,
sugarcane), fomesafen (e.g. soybean, cotton), metribuzin (e.g. soybean), pendimethalin (e.g.
corn, cotton, soybean, and many other crops), pyrithiobac and trifloxysulfuron(cotton),
pyroxasulfone (e.g. corn, cotton, soybean, wheat), saflufenacil ( e.g. corn, cotton, alfalfa), S-
metolachlor (e.g. corn, cotton, soybean, and many other crops), and tembotrione (e.g. corn)
(Scott et al. 2017). Labelled post-emergence herbicides for Palmer amaranth control include
atrazine (e.g. corn, sorghum), dicamba (e.g. corn, sorghum), fomesafen (e.g. soybean),
glufosinate (e.g. LibertyLink® crops), glyphosate (e.g. Roundup Ready® crops), and mesotrione
(e.g. corn, sorghum) (Norsworthy et al., 2008).
The problem of Palmer amaranth escalated with the evolution of herbicide-resistant
populations due to intensive use of herbicides. As a dioecious species, Palmer amaranth is
outcrossing, allowing herbicide resistance to spread rapidly (Steckel, 2007). The glyphosate
resistance trait was transferred across a 300-m distance through pollen from glyphosate-resistant

14
males to glyphosate-susceptible female plants (Sosnoskie et al., 2012). Resistance to six different
modes of action (MOA) has been documented in Palmer amaranth: acetolactate synthase (ALS)
inhibitors, dinitroanilines, triazine, glyphosate, hydroxyphenylpyruvate dioxygenase (HPPD)
inhibitors, and protoporphyrinogen oxidase (PPO) inhibitors (Heap 2018). Some Palmer
amaranth populations are resistant to more than one MOA (Burgos et al., 2001). The rampant
occurrence of ALS- and glyphosate-resistant Palmer amaranth populations has raised great
challenge to farmers and researchers as this curtails the use of ALS inhibitors and glyphosate in
crop production systems. With glyphosate and ALS inhibitors out of the weed management
toolbox, weed control has returned to multiple-herbicide programs including PSII, PPO, and
shoot/root inhibitors. Genetically modified crops with tolerance to 2,4-D and dicamba are
recently commercialized and new technologies on HPPD-tolerant soybean (mesotrione and
isoxaflutole) will be available soon. These alternative modes of action should be wisely used to
preserve the utility of these herbicides and delay the evolution of resistant weeds.
Non-target-site-based resistance mechanism
There are two primary mechanisms of herbicide resistance in weeds: 1) target-site
resistance (TSR) which is caused by alterations in the gene of the herbicide target site causing a
decrease in the affinity of the herbicide for its target and 2) non-target-site resistance (NTSR)
which is endowed by mechanisms not belonging to TSR (Delye, 2013). NTSR mechanisms
include mechanisms that minimize the amount of herbicide in reaching the target site such as
decreased herbicide uptake and translocation, increases herbicide sequestration, and metabolism
(Powles and Yu, 2010). NTSR can confer unpredictable resistance to multiple herbicides with
different MOA, including herbicides not yet used (Petit et al., 2010a; Petit et al., 2010b).

15
Sublethal dose of herbicide has been implicated to promote non-target-site metabolic resistance
(Neve and Powles, 2005; Yuan et al., 2007). NTSR within a single plant is generally endowed by
a combination of mechanisms controlled by different alleles as demonstrated in the segregation
analysis of NTSR to ACCase ad ALS herbicides in progenies from controlled pairings of black-
grass (Alopecurus myosuroides) parents (Petit et al., 2010b).
NTSR mechanisms as a part of plant stress response can interfere with herbicide
penetration and accumulation at the target site, and/or protect the plant against the phytotoxic
effects of herbicide action. Detoxification of herbicide usually follow a three-step process: 1)
transformation of the herbicide molecule into a more hydrophilic metabolite by P450
monooxygenase, 2) conjugation of metabolites to glutathione or glucose, 3) further conjugation,
cleaving and oxidizing and export of the molecule to the vacuole or cell wall for final
degradation (Delye, 2013). Increased expression and activity of the enzymes that are involved in
the detoxification process can contribute to reduced sensitivity of the plant to the herbicide.
Reduced herbicide translocation may also be connected with detoxification of herbicides.
Reduced translocation of glyphosate in horseweed (Conyza candensis) and ryegrass (Lolium spp)
is due to rapid sequestration of glyphosate into the vacuole (Ge et al., 2012; Ge et al., 2010).
Reduced herbicide absorption in an ACCase-resistant rigid ryegrass (Lolium rigidum) biotype is
due to greater epicuticular wax density in the leaf cuticle (de Prado et al., 2005).
The basis for crop-weed selectivity for many herbicides is due to differential herbicide
metabolism. The herbicide is safe for the crop because the crop metabolizes the herbicide
whereas the weed does not. Enhanced herbicide metabolism is more common in grass weeds, but
additional cases involving dicot weeds are emerging (Yu and Powles, 2014). The co-application
of malathion, a P450 inhibitor, resulted in loss or decreased level of herbicide resistance in

16
several weed species demonstrated that enhanced herbicide metabolism is attributed to the
cytochrome P450 family of proteins (Beckie et al., 2012; Breccia et al., 2017; Fu et al., 2017;
Zhao et al., 2017). An enhanced herbicide metabolism has been reported in rigid ryegrass (Yu et
al., 2013), wild mustard (Sinapis arvensis) (Veldhuis et al., 2000a), and late watergrass
(Echinochloa phyllopogon) (Yasuor et al., 2009; Yun et al., 2005). Cytochrome P450s have been
implicated in the metabolism of different herbicides in several crops and weed species
(Cotterman and Saari, 1992; Hinz and Owen, 1996; Yu and Powles, 2014). The cloning of rice
(Oryza sativa) CYP81A6 and Jerusalem artichoke (Helianthus tuberosus) CYP76B1 into
Arabidopsis conferred resistance to sulfonylurea and phenylurea herbicides, respectively (Pan et
al, 2006; Liu et al. 2012; Didierjean et al. 2002).
In addition to P450, glutathione S-transferases (GSTs) and glycosyltransferases have
been implicated in herbicide metabolism (Yuan et al., 2007). Each of these three enzyme systems
is encoded by large gene families. GSTs are ubiquitous in plants and have defined roles in
herbicide detoxification (Wagner et al., 2002). Maize (Zea mays)and giant foxtail (Setaria
faberi) are tolerant to atrazine due to its ability to rapidly detoxify the herbicide atrazine by
glutathione conjugation with GST (Hatton et al., 1999; Hatton et al., 1996). Furthermore,
induction of glutathione and GSTs by herbicide 'safeners' is employed commercially to protect
crop plants from injury by certain herbicides (Farago et al., 1994). GSTs (phi and tau class) were
identified to be involved in diclofop resistance in rigid ryegrass (Gaines et al., 2014).
Furthermore, GSTA and GST1 were also recently reported to participate in non-target resistance
or tolerance to ALS inhibitor in ryegrass and maize, respectively (Duhoux et al., 2015; Liu et al.,
2015). A transgenic tobacco plant (Nicotiana tabacum) expressing maize GST1 had enhanced
detoxification of alachlor (Karavangeli et al., 2005). In ACCase-resistant black-grass, GSTF1

17
catalyzed the conjugation of herbicide to glutathione and acted as peroxidases protecting the cell
from oxidative damage (Cummins et al., 2009; Cummins et al., 1999; Cummins et al., 2013).
Transgenic Arabidopsis expressing the black-grass GSTF1gene (AmGSTF1) has improved
tolerance to some herbicides due to increased accumulation of protective flavonoids (Cummins
et al., 2013).
Few non-target mechanisms have been elucidated at the molecular level because of the
inherently complicated biochemical processes and limited genomic information available in
weeds species (Yuan et al., 2007). Although NTSR mechanisms are widespread among weed
species, its genetic determinant is poorly understood (Delye, 2013). The de novo transcriptome
for waterhemp (Amaranthus tuberculatus) was characterized using GS-FLX 454 sequencing
which can help in establishing valuable genomic resource for weed science research (Riggins et
al., 2010). The transcriptome is useful for identifying potential candidate genes involved
herbicide resistance and may serve as a reference for gene expression and functional genomics
studies Using whole-transcriptome sequencing (RNA-Seq), several genes were identified in
ryegrass, black-grass, flixweed (Descurainia sophia L.), and shortawn foxtail (Alopecurus
aequalis) that confer NTSR to several herbicides (Gaines et al. 2014; Duhoux et al. 2017; Gardin
et al. 2015; Yang et al. 2016; Zhao et al. 2017). Identifying NTSR genes will help in
understanding NTSR evolution as well as help in understanding weed adaptation to herbicides.
Glufosinate
Glufosinate is a broad-spectrum, non-selective, post-emergence herbicide commonly
used to control weeds in vineyards, orchards and genetically modified crops with LibertyLink®
trait. Glufosinate inhibits glutamine synthetase (GS) (E. C. 6.3.1.2) enzyme which catalyzes the

18
conversion of glutamic acid and ammonia to form glutamine, the first reaction in the ammonia
assimilation in higher plants. Glutamine synthetase functions as the major assimilatory enzyme
for ammonia produced from nitrogen fixation, and nitrate or ammonia nutrition, as well as
reassimilates ammonia released as a result of photorespiration and the breakdown of proteins and
nitrogen transport compounds (Miflin and Habash, 2002). Inhibition of GS enzyme leads to rapid
ammonia accumulation and amino acid biosynthesis inhibition which cause damage in the
chloroplast structures and terminates photosynthetic activity, ultimately leading to plant death
(Tachibana et al., 1986). The availability of glufosinate-tolerant cotton, soybean, and corn
varieties allows farmers to use glufosinate in controlling broad spectrum of weeds without
damaging the crop as well as provides growers an alternative herbicide for controlling Palmer
amaranth that is resistant to other herbicides.
Despite the non-selective nature of glufosinate, variable control of goosegrass (Eleusine
indica), large crabgrass (Digitaria sanguinalis) and Palmer amaranth has been observed (Beyers
et al., 2002; Corbett et al., 2004; Everman et al., 2007). Differential tolerance to glufosinate in
weed species has been attributed to application rate, plant species, application timing, humidity,
growth stage, and variations in the level of absorption and translocation (Everman, 2007).
Resistance to glufosinate has been reported in goosegrass (Seng et al., 2010) and Italian ryegrass
(Avila-Garcia and Mallory-Smith, 2011). In glufosinate-resistant Italian ryegrass (Lolium
perenne L. ssp multiflorum) population from Oregon, resistance is attributed to mutation of the
GS gene in which aspartic acid was substituted to asparagine at the amino acid position 171
(Avila-Garcia et al., 2012). The overproduction of GS enzyme conferred resistance to glufosinate
in rice lines (Tsai et al., 2006). Glufosinate-tolerant crops are developed by expressing
phosphinothricin N-acetyltransferase (PAT), which can detoxify L-phosphinothricin

19
(glufosinate) by acetylation of the amino group (Droge et al. 1992). Although resistance to
glufosinate in Palmer amaranth has not been reported, tolerance to glufosinate has been observed
in a population of Palmer amaranth in Arkansas (Botha, 2012).
ALS-inhibiting herbicides
Acetolactate synthase, also known as acetohydroxyacid synthase or AHAS (E.C.
4.1.3.18), is the first enzyme in the biosynthesis of the branched chain amino acids Ile, Leu, and
Val. Inhibition of ALS leads to depletion of these amino acids disrupting protein synthesis,
thereby causing plant death (Whitcomb, 1999). There are five chemical families of ALS
herbicides, namely: sulfonylurea (SU), imidazolinone (IMI), triazolopyrimidine sulfonanilides
(TP), pyrimidinylthiobenzoates (PTB), and sulfonylaminocarbonyltriazolinone. The ALS
herbicides were widely used because they provide broad-spectrum weed control at low doses,
soil residual activity, wide application windows, excellent crop safety and low mammalian
toxicity. However, these herbicides have high propensity to select resistant weed populations due
to its widespread usage, strong selection pressure, and high levels of natural variability in the
ALS gene. Selection of ALS-resistant weed populations became evident in 1987, only five years
after the introduction of the first SU, with the discovery of chlorsulfuron-resistant prickly lettuce
(Lactuca serriola L.) and kochia (Kochia scoparia L. Shrad) (Mallory-Smith et al., 1990;
Primiani et al., 1990). In the United States, the first case of resistance to ALS-inhibiting
herbicides in Palmer amaranth was reported in 1993 in Kansas and cross-resistance to five ALS
herbicides was documented in Arkansas in 1994 (Heap 2018). Resistance to ALS inhibitors in
Palmer amaranth is widespread throughout the United States. As a result, ALS-inhibiting
herbicides are no longer effective on ALS-resistant populations of Palmer amaranth.

20
Most cases of ALS resistance are due to mutation in the ALS gene resulting in an enzyme
that is less sensitive to herbicide binding. To date, 29 amino acid substitutions in ALS conferring
herbicide resistance were identified at Ala122 (5), Ala205 (2), Arg377 (1), Asp376 (1), Gly654 (2),
Pro197 (11), Ser653 (3), and Trp574(4) in weed species (Heap 2018). Recent investigation on 20
Palmer amaranth populations from Arkansas revealed TSR mechanism involving Trp574Ser
mutation with a few cases of double mutations involving Ala122Thr, Pro197Ala or Ser653Asn
(Singh, 2017). Amino acid substitution at Pro197 conferred resistance to SUs with low or no
cross-resistance to IMIs (Tranel and Wright, 2002), whereas substitutions at Ala122 or Ser653
conferred resistance to IMI herbicides with low-level resistance to SUs (Bernasconi et al., 1996;
Devine et al., 1991). Substitution at Trp574 confers resistance to IMIs, PTBs, SUs, and TPs
(Tranel and Wright, 2002), whereas Asp376Gluand Ala205Phe substitution confers resistance to all
five chemical families of ALS inhibitors (Brosnan et al., 2016; Whaley et al., 2007).
An important mechanism of naturally occurring (as opposed to evolved) resistance to
ALS inhibitors is detoxification of the active herbicide in the plant. Inherent selectivity of a
particular ALS inhibitor in a given crop is based on the crops’ ability to metabolize the herbicide
to nonphytotoxic compounds rapidly enough to prevent lethal herbicide levels from reaching the
target enzyme ALS (Saari et al. 1994). Among the more common detoxification reactions
involved in crop tolerance to sulfonylureas are hydroxylation, O-dealkylation, and
deesterification (Saari et al. 1994). Maize is tolerant to nicosulfuron, a sulfonylurea herbicide,
because nicosulfuron is rapidly metabolized to 5-hydroxypyrimidinyl nicosulfuron, a
herbicidally inactive derivative which is then conjugated to glucose (Brown et al. 1991).
Similarly, flumetsulam, a triazolopyrimidine that is selective in cereals, maize, and soybeans,
also owes its selectivity to metabolic detoxification. Tolerant plants oxidize flumetsulam to one

21
or more hydroxylated metabolites, and soybean produces an open pyrimidine ring metabolite
(Swisher et al., 1991). This tolerance mechanism in crops also appears to be the same mechanism
responsible for poor control of some weeds by certain ALS herbicides (Saari et al. 1994).
Non-target-site resistance to ALS inhibitors have been reported in some weeds species.
Reduced absorption and translocation rarely underlay resistance to ALS inhibitors (Cruz-
Hipolito et al., 2013; Poston et al., 2001; Veldhuis et al., 2000b) and in only few cases have they
been reported as partial-resistance mechanism (Riar et al., 2013; White et al., 2002) . An
enhanced herbicide metabolism has been reported in rigid ryegrass (Yu et al., 2013), wild
mustard (Veldhuis et al., 2000a), and late watergrass (Yasuor et al., 2009; Yun et al., 2005). A
late watergrass biotype with multiple herbicide resistance to bispyribac-sodium, fenoxaprop-
ethyl, and thiobencarb exhibited higher P450 hydroxylation activity toward these herbicides than
the susceptible biotype, which suggests the involvement of cytochrome P450 enzymes as a
mechanism for resistance (Yun et al. 2005). Furthermore, resistance to penoxsulam in late
watergrass is conferred by an enhanced ability to detoxify the herbicide via malathion-sensitive
monooxygenases (Yasour et al. 2009). The P450 inhibitor malathion reverses chlorsulfuron
resistance in rigid ryegrass (Yu et al. 2009). Several cytochrome P450 have been identified in
ALS-resistant weeds, such as CYP72A254 (also known as CYPA72A219) in late watergrass
(bispyribac-sodium) (Iwakami et al., 2014), CYP81D in black-grass (mesosulfuron and
iodosulfuron) (Gardin et al., 2015); CYP94A1, CYP94A2, CYP71A4 and CYP734A6 in
shortawn foxtail (mesosulfuron) (Zhao et al. 2017). In addition, many CytP450s conferring
resistance to various herbicides have been documented in several major crop species, such as
CYP81A6 in rice (Pan et al., 2006); CYP76B1 in Jerusalem artichoke (Didierjean et al., 2002),
and CYP81A9 in maize (Zea mays) (Liu et al. 2015). The wheat (Triticum aestivum)

22
CYP71C6v1 expressed in yeast (Saccharomyces cerevisiae) is able to metabolize several
sulfonylurea herbicides such chlorsulfuron and triasulfuron through phenyl ring hydroxylation
(Xiang et al., 2006).
PPO-inhibiting herbicides
Herbicides that inhibit protoporphyrinogen oxidase (PPO) include lactofen and
fomesafen herbicides. Although they have been used for many years in the control of broadleaf
weeds, their use began to slowly decline in the late 1990s due to the massive adoption of
glyphosate-resistant varieties (Riggins et al., 2010). However, the advent and continued increase
of glyphosate-resistant weeds cause growers to once again rely on PPO herbicides as an
alternative approach to control weeds. Even though resistance to PPO herbicide has been slow to
evolve, it may be expected to occur in weed species that are under strong and continuous
selection pressure.
The PPO enzyme plays a major role in the biosynthesis of chlorophyll and heme by
catalyzing the oxidation of protoporphyrinogen (protogen) to porphyrin IX (Proto IX). Upon
treatment with PPO-herbicides, susceptible plants accumulate protogen IX which is then
transported to the cytoplasm where it spontaneously form Proto IX. In presence of light, Proto IX
generates singlet oxygen that causes degradation of cell membranes leading cells to break open
and ultimately to cellular death (Becerril and Duke, 1989; Jacobs et al., 1991; Sherman et al.,
1991). However, few species of weeds have slowly evolved resistance from the repeated use of
these herbicides in past few years. To, date 11 different weed species have evolved resistance to
PPO-herbicides including Palmer amaranth (Heap 2018; Giacomini et al., 2017; Salas et al.,
2016; Salas-Perez et al., 2017).

23
The resistance mechanism to PPO herbicides in tall waterhemp is not due to difference in
herbicide absorption and translocation, or metabolism (Shoup and Al-Khatib, 2005) but due to a
unique codon deletion of glycine at position 210 in the mitochondrial isoform of the PPO2 gene
(Patzoldt et al., 2006). The same mutation appears to be the only resistance mechanism to PPO
herbicides in various tall waterhemp populations in the United States (Lee et al., 2008; Wuerffel
et al., 2015). This glycine deletion alters the binding domain of the enzyme without negatively
affecting substrate affinity (Dayan et al., 2010). However, a novel PPO mutation, Arg98Leu was
detected in common ragweed (Ambrosia artemisiifolia), and recently two new substitutions
identified at the same position in Palmer amaranth as Arg128Gly and Arg128Met (numbering
changed due to the presence of 30 amino acid extension in Palmer amaranth) (Giacomini et al.,
2014; Rousonelos et al., 2012). Computational modeling revealed that replacement of this
arginine with hydrophobic glycine removed important hydrogen-bonding interactions with
acifluorfen, fomesafen, and sulfentrazone resulting in reduced binding of these inhibitors (Hao et
al., 2014).
Non-target resistance to PPO inhibitors in weed species has not yet been reported
although natural tolerance to PPO inhibitors in crops is usually due to enhanced herbicide
degradation. Diphenyl ether herbicides are detoxified in soybean by homoglutathione
conjugation (Skipsey et al., 1997). Similarly, tolerance to the diphenyl ether fluorodifen in peas
is due to rapid conjugation with glutathione (Edwards, 1996).

24
Literature Cited
Avila-Garcia, W. V., Mallory-Smith, C. 2011. Glyphosate-resistant Italian ryegrass (Lolium
perenne) populations also exhibit resistance to glufosinate. Weed Science 59: 305-309.
Avila-Garcia, W. V., Sanchez-Olguin, E., Hulting, A. G., Mallory-Smith, C. 2012. Target-site
mutation associated with glufosinate resistance in Italian ryegrass (Lolium perenne L. ssp
multiflorum). Pest Management Science 68: 1248-1254.
Becerril, J. M., Duke, S. O. 1989. Protoporphyrin IX content correlates with activity of
photobleaching herbicides. Plant Physiology 90: 1175-81.
Beckie, H. J., Warwick, S. I., Sauder, C. A. 2012. Basis for herbicide resistance in Canadian
populations of wild oat (Avena fatua). Weed Science 60: 10-18.
Bensch, C. N., Horak, M. J., Peterson, D. 2003. Interference of redroot pigweed (Amaranthus
retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A. rudis) in
soybean. Weed Science 51: 37-43.
Bernasconi, P., Woodworth, A. R., Rosen, B. A., Subramanian, M. V., Siehl, D. L. 1996. A
naturally occurring point mutation confers broad range tolerance to herbicides that target
acetolactate synthase. Journal Biological Chemistry 271: 13925.
Beyers, J. T., Smeda, R. J., Johnson, W. G. 2002. Weed management programs in glufosinate-
resistant soybean (Glycine max). Weed Technology 16: 267-273.
Botha, G. M. 2012. Alternative herbicide control options for glyphosate-resistant Palmer
amaranth (Amaranthus palmeri). Master of Science in Crop, Soil, and Environmental
Sciences: University of Arkansas. 130 p.
Breccia, G. et al. 2017. Contribution of non-target-site resistance in imidazolinone-resistant
Imisun sunflower. Bragantia 76: 536-542.
Brosnan, J. T. et al. 2016. A new amino acid substitution (Ala-205-Phe) in acetolactate synthase
(ALS) confers broad spectrum resistance to ALS-inhibiting herbicides. Planta 243: 149-
59.
Burgos, N. R., Kuk, Y.-I., Talbert, R. E. 2001. Amaranthus palmeri resistance and differential
tolerance of Amaranthus palmeri and Amaranthus hybridus to ALS-inhibitor herbicides.
Pest Management Science 57: 449-457.
Burke, I. C., Schroeder, M., Thomas, W. E., Wilcut, J. W. 2007. Palmer amaranth interference
and seed production in peanut. Weed Technology 21: 367-371.
Corbett, J. L., Askew, S. D., Thomas, W. E., Wilcut, J. W. 2004. Weed efficacy evaluations for
bromoxynil, glufosinate, glyphosate, pyrithiobac, and sulfosate. Weed Technology 18:
443-453.
Cotterman, J. C., Saari, L. L. 1992. Rapid metabolic inactivation is the basis for cross-resistance
to chlorsulfuron in diclofop-methyl-resistant rigid ryegrass (Lolium rigidum) biotype
SR484. Pesticide Biochemistry and Physiology 43: 182-192.
Cruz-Hipolito, H. et al. 2013. Resistance mechanism to tribenuron-methyl in white mustard
(Sinapis alba) from southern Spain. Weed Science 61: 341-347.

25
Cummins, I., Bryant, D. N., Edwards, R. 2009. Safener responsiveness and multiple herbicide
resistance in the weed black-grass (Alopecurus myosuroides). Plant Biotechnology
Journal 7: 807-820.
Cummins, I., Cole, D. J., Edwards, R. 1999. A role for glutathione transferases functioning as
glutathione peroxidases in resistance to multiple herbicides in black-grass. Plant Journal
18: 285-292.
Cummins, I. et al. 2013. Key role for a glutathione transferase in multiple-herbicide resistance in
grass weeds. Proceedings of the National Academy of Sciences of the United States of
America 110: 5812-5817.
Dayan, F. E. et al. 2010. Biochemical and structural consequences of a glycine deletion in the
alpha-8 helix of protoporphyrinogen oxidase. Biochimica Et Biophysica Acta-Proteins
and Proteomics 1804: 1548-1556.
de Prado, J. L., Osuna, M. D., Heredia, A., de Prado, R. 2005. Lolium rigidum, a pool of
resistance mechanisms to ACCase inhibitor herbicides. Journal of Agricultural and Food
Chemistry 53: 2185-2191.
Delye, C. 2013. Unravelling the genetic bases of non-target-site-based resistance (NTSR) to
herbicides: a major challenge for weed science in the forthcoming decade. Pest
Management Science 69: 176-187.
Devine, M. D., Maries, M. A. S., Hall, L. M. 1991. Inhibition of acetolactate synthase in
susceptible and resistant biotypes of Stellaria media. Pesticide Science 31: 273-280.
Didierjean, L. et al. 2002. Engineering herbicide metabolism in tobacco and Arabidopsis with
CYP76B1, a cytochrome P450 enzyme from Jerusalem artichoke. Plant Physiology 130.
Droge, W., Broer, I., Puhler, A. 1992. Transgenic plants containing the phosphinothricin-N-
acetyltransferase gene metabolize the herbicide L-phosphinothricin (glufosinate)
differently from untransformed plants. Planta 187: 142-51.
Duhoux, A., Carrère, S., Gouzy, J., Bonin, L., Délye, C. 2015. RNA-Seq analsis of rye-grass
transcriptomic response to an herbicide inhibiting acetolactate-synthase identifies
transcripts linked to non-target-site-based resistance. Plant Molecular Biology 87.
Edwards, R. 1996. Characterisation of glutathione transferases and glutathione peroxidases in
pea (Pisum sativum). Physiologia Plantarum 98: 594-604.
Ehleringer, J. 1983. Ecophysiology of Amaranthus palmeri, a Sonoran desert summer annual.
Oecologia 57: 107-112.
Ehleringer, J., Forseth, I. 1980. Solar tracking by plants. Science 210: 1094-1098.
Everman, W. J. 2007. Influence of environmental and physiological factors on glufosinate and
glyphosate weed management. PhD: North Carolina State University. 194 p.
Everman, W. J., Burke, I. C., Allen, J. R., Collins, J., Wilcut, J. W. 2007. Weed control and yield
with glufosinate-resistant cotton weed management systems. Weed Technology 21: 695-
701.
Farago, S., Brunold, C., Kreuz, K. 1994. Herbicide safeners and glutathione metabolism.
Physiologia Plantarum 91: 537-542.

26
Fu, D. N. et al. 2017. Bensulfuron-methyl resistant Sagittaria trifolia L.: multiple resistance,
cross-resistance and molecular basis of resistance to acetolactate synthase-inhibiting
herbicides. Archives of Biological Sciences 69: 649-658.
Gaines, T. A., Ward, S. M., Bekun, B., Preston, C., Leach, J. E., Westra, P. 2012. Interspecific
hybridization transfers a previously unknown glyphosate resistance mechanism in
Amaranthus species. Evolutionary Applications 5: 29-38.
Gaines, T. A. et al. 2014. RNA-Seq transcriptome analysis to identify genes involved in
metabolism-based diclofop resistance in Lolium rigidum. Plant Journal 78.
Gardin, J. A. C., Gouzy, J., Carrère, S., Délye, C. 2015. ALOMYbase, a resource to investigate
non-target-site-based resistance to herbicides inhibiting acetolactate-synthase (ALS) in
the major grass weed Alopecurus myosuroides (black-grass). BMC Genomics 16.
Ge, X. et al. 2012. Vacuolar glyphosate-sequestration correlates with glyphosate resistance in
ryegrass (Lolium spp.) from Australia, South America, and Europe: A P-31 NMR
Investigation. Journal of Agricultural and Food Chemistry 60: 1243-1250.
Ge, X., d'Avignon, D. A., Ackerman, J. J. H., Sammons, R. D. 2010. Rapid vacuolar
sequestration: the horseweed glyphosate resistance mechanism. Pest Management
Science 66: 345-348.
Giacomini, D., Westra, P., Ward, S. M. 2014. Impact of genetic background in fitness cost
studies: An example from glyphosate-resistant Palmer amaranth. Weed Science 62: 29-
37.
Giacomini, D. A. et al. 2017. Two new PPX2 mutations associated with resistance to PPO-
inhibiting herbicides in Amaranthus palmeri. Pest Management Science 73: 1559-1563.
Gibson, A. C. 1998. Photosynthetic organs of desert plants. Bioscience 48: 911-920.
Hao, G.-F. et al. 2014. Understanding resistance mechanism of protoporphyrinogen oxidase-
inhibiting herbicides: Insights from computational mutation scanning and site-directed
mutagenesis. Journal of Agricultural and Food Chemistry 62: 7209-7215.
Hatton, P. J., Cummins, I., Cole, D. J., Edwards, R. 1999. Glutathione transferases involved in
herbicide detoxification in the leaves of Setaria faberi (giant foxtail). Physiologia
Plantarum 105: 9-16.
Hatton, P. J., Dixon, D., Cole, D. J., Edwards, R. 1996. Glutathione transferase activities and
herbicide selectivity in maize and associated weed species. Pesticide Science 46: 267-
275.
Heap I. The international survey of herbicide resistant weeds. 2018.
http://www.weedscience.com. Accessed 22 kkJanuary 2018.
Hinz, J. R., Owen, M. D. K. 1996. Nicosulfuron and primisulfuron selectivity in corn (Zea mays)
and two annual grass weeds. Weed Science 44: 219-223.
Horak, M. J., Loughin, T. M. 2000. Growth analysis of four Amaranthus species. Weed Science
48: 347-355.

27
Iwakami, S. et al. 2014. Cytochrome P450 genes induced by bispyribac-sodium treatment in a
multiple-herbicide-resistant biotype of Echinochloa phyllopogon. Pest Management
Science 70: 549-58.
Jacobs, J. M., Jacobs, N. J., Sherman, T. D., Duke, S. O. 1991. Effect of diphenyl ether
herbicides on oxidation of protoporphyrinogen to protoporphyrin in organellar and
plasma membrane enriched fractions of barley. Plant Physiology 97: 197-203.
Jha, P., Norsworthy, J. K. 2009. Soybean canopy and tillage effects on emergence of Palmer
amaranth (Amaranthus palmeri) from a natural seed Bank. Weed Science 57: 644-651.
Jha, P., Norsworthy, J. K., Riley, M. B., Bielenberg, D. G., Bridges, W., Jr. 2008. Acclimation of
Palmer amaranth (Amaranthus palmeri) to shading. Weed Science 56: 729-734.
Karavangeli, M., Labrou, N. E., Clonis, Y. D., Tsaftaris, A. 2005. Development of transgenic
tobacco plants overexpressing maize glutathione S-transferase I for chloroacetanilide
herbicides phytoremediation. Biomolecular Engineering 22: 121-8.
Keeley, P. E., Carter, C. H., Thullen, R. J. 1987. Influence of planting date on growth of Palmer
amaranth (Amaranthus palmeri). Weed Science 35: 199-204.
Lee, R. M., Hager, A. G., Tranel, P. J. 2008. Prevalence of a novel resistance mechanism to
PPO-inhibiting herbicides in waterhemp (Amaranthus tuberculatus). Weed Science 56:
371-375.
Liu, C., Liu, S. Q., Wang, F., Wang, Y. Q., Liu, K. D. 2012. Expression of a rice CYP81A6 gene
confers tolerance to bentazon and sulfonylurea herbicides in both Arabidopsis and
tobacco. Plant Cell, Tissue and Organ Culture 109: 419-428.
Liu, X. M. et al. 2015. RNA-Seq transcriptome analysis of maize inbred carrying nicosulfuron-
tolerant and nicosulfuron-susceptible alleles. International Journal of Molecular Sciences
16: 5975-5989.
Mallory-Smith, C. A., Thill, D. C., Dial, M. J. 1990. Identification of sulfonylurea herbicide-
resistant prickly lettuce (Lactuca serriola). Weed Technology 4: 163-168.
Massinga, R. A., Currie, R. S., Horak, M. J., Boyer, J. 2001. Interference of Palmer amaranth in
corn. Weed Science 49: 202-208.
Meyers, S. L., Jennings, K. M., Schultheis, J. R., Monks, D. W. 2010. Interference of Palmer
Amaranth (Amaranthus palmeri) in sweetpotato. Weed Science 58: 199-203.
Miflin, B. J., Habash, D. Z. 2002. The role of glutamine synthetase and glutamate dehydrogenase
in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of
crops. Journal of Experimental Botany 53: 979-987.
Morgan, G. D., Baumann, P. A., Chandler, J. M. 2001. Competitive impact of Palmer amaranth
(Amaranthus palmeri) on cotton (Gossypium hirsutum) development and yield. Weed
Technology 15: 408-412.
Neve, P., Powles, S. 2005. High survival frequencies at low herbicide use rates in populations of
Lolium rigidum result in rapid evolution of herbicide resistance. Heredity 95: 485-492.

28
Norsworthy, J. K., Griffith, G. M., Scott, R. C., Smith, K. L., Oliver, L. R. 2008. Confirmation
and control of glyphosate-resistant Palmer amaranth (Amaranthus palmeri) in Arkansas.
Weed Technology 22: 108-113.
Pan, G. et al. 2006. Map based cloning of a novel rice cytochrome P450 gene CYP81A6 that
confers resistance to two different classes of herbicides. Plant Molecular Biology 61.
Patzoldt, W. L., Hager, A. G., McCormick, J. S., Tranel, P. J. 2006. A codon deletion confers
resistance to herbicides inhibiting protoporphyrinogen oxidase. Proceedings of the
National Academy of Sciences of the United States of America 103: 12329-12334.
Petit, C., Bay, G., Pernin, F., Delye, C. 2010a. Prevalence of cross- or multiple resistance to the
acetyl-coenzyme A carboxylase inhibitors fenoxaprop, clodinafop and pinoxaden in
black-grass (Alopecurus myosuroides Huds.) in France. Pest Management Science 66:
168-177.
Petit, C., Duhieu, B., Boucansaud, K., Delye, C. 2010b. Complex genetic control of non-target-
site-based resistance to herbicides inhibiting acetyl-coenzyme A carboxylase and
acetolactate-synthase in Alopecurus myosuroides Huds. Plant Science 178: 501-509.
Poston, D. H., Wu, J., Hatzios, K. K., Wilson, H. P. 2001. Enhanced sensitivity to cloransulam-
methyl in imidazolinone-resistant smooth pigweed. Weed Science 49: 711-716.
Powles, S. B., Yu, Q. 2010. Evolution in Action: Plants Resistant to Herbicides. Pages 317-347
in S. Merchant, W.R. Briggs, D. Ort, eds. Annual Review of Plant Biology, Vol 61.
Primiani, M. M., Cotterman, J. C., Saari, L. L. 1990. Resistance of kochia (Kochia scoparia) to
sulfonylurea and imidazolinone herbicides. Weed Technology 4: 169-172.
Riar, D. S. et al. 2013. Physiological and molecular basis of acetolactate synthase-inhibiting
herbicide resistance in barnyardgrass (Echinochloa crus-galli). Journal of Agricultural
and Food Chemistry 61: 278-289.
Riggins, C. W., Peng, Y., Stewart, C. N., Jr., Tranel, P. J. 2010. Characterization of de novo
transcriptome for waterhemp (Amaranthus tuberculatus) using GS-FLX 454
pyrosequencing and its application for studies of herbicide target-site genes. Pest
Management Science 66: 1042-1052.
Rousonelos, S. L., Lee, R. M., Moreira, M. S., VanGessel, M. J., Tranel, P. J. 2012.
Characterization of a common ragweed (Ambrosia artemisiifolia) population resistant to
ALS- and PPO-inhibiting herbicides. Weed Science 60: 335-344.
Salas, R. A. et al. 2016. Resistance to PPO-inhibiting herbicide in Palmer amaranth from
Arkansas. Pest Management Science 72: 864-869.
Salas-Perez, R. A. et al. 2017. Frequency of Gly-210 deletion mutation among
protoporphyrinogen oxidase inhibitor–resistant Palmer amaranth (Amaranthus palmeri)
populations. Weed Science 65: 718-731.
Seng, C. T., Van Lun, L., San, C. T., Bin Sahid, I. 2010. Initial report of glufosinate and paraquat
multiple resistance that evolved in a biotype of goosegrass (Eleusine indica) in Malaysia.
Weed Biology and Management 10: 229-233.

29
Sherman, T. D. et al. 1991. Physiological basis for differential sensitivities of plant species to
protoporphyrinogen oxidase-inhibiting herbicides. Plant Physiology 97: 280-287.
Shoup, D. E., Al-Khatib, K. 2005. Fate of acifluorfen and lactofen in common waterhemp
(Amaranthus rudis) resistant to protoporphyrinogen oxidase-inhibiting herbicides. Weed
Science 53: 284-289.
Singh, S. 2017. Characterization of glyphosate-resistant Amaranthus palmeri (Palmer amaranth)
tolerance to ALS- and HPPD-inhibiting herbicides: University of Arkansas. 125 p.
Skipsey, M., Andrews, C. J., Townson, J. K., I., J., Edwards, E. 1997. Substrate and thiol
specificity of a stress-inducible glutathione transferase from soybean. Pages 370-374
FEBS Letters.
Sosnoskie, L. M. et al. 2012. Pollen-mediated dispersal of glyphosate-resistance in Palmer
amaranth under field conditions. Weed Science 60: 366-373.
Steckel, L. E. 2007. The dioecious Amaranthus spp.: Here to stay. Weed Technology 21: 567-
570.
Swisher, B., Gerwick, B., Chang, M., Miner, V., G., d. Metabolism of the triazolopyrimidine
sulfonanilide DE-498 in plants in Proceedings of the 1991 Weed Science Society of
America. Champaign, Illinois: WSSA.
Tachibana, K., Watanabe, T., Sekizawa, Y., Takematsu, T. 1986. Action mechanism of bialaphos
.2. accumulation of ammonia in plants treated with bialaphos. Journal of Pesticide
Science 11: 33-37.
Tranel, P. J., Wright, T. R. 2002. Resistance of weeds to ALS-inhibiting herbicides: What have
we learned? Weed Science 50: 700-712.
Tsai, C. J., Wang, C. S., Wang, C. Y. 2006. Physiological characteristics of glufosinate
resistance in rice. Weed Science 54: 634-640.
Veldhuis, L. J., Hall, L. M., O'Donovan, J. T., Dyer, W., Hall, J. C. 2000a. Metabolism-based
resistance of a wild mustard (Sinapis arvensis L.) biotype to ethametsulfuron-methyl.
Journal of Agricultural and Food Chemistry 48: 2986-90.
Veldhuis, L. J., Hall, L. M., O'Donovan, J. T., Dyer, W., Hall, J. C. 2000b. Metabolism-based
resistance of a wild mustard (Sinapis arvensis L.) biotype to ethametsulfuron-methyl.
Journal of Agricultural and Food Chemistry 48: 2986-2990.
Wagner, U., Edwards, R., Dixon, D. P., Mauch, F. 2002. Probing the diversity of the Arabidopsis
glutathione S-transferase gene family. Plant Molecular Biology 49: 515-32.
Ward, S. M., Webster, T. M., Steckel, L. E. 2013. Palmer amaranth (Amaranthus palmeri): A
Review. Weed Technology 27: 12-27.
Whaley, C. M., Wilson, H. P., Westwood, J. H. 2007. A new mutation in plant ALS confers
resistance to five classes of ALS-inhibiting herbicides. Weed Science 55: 83-90.
Whitcomb, C. E. 1999. An introduction to ALS-inhibiting herbicides. Toxicology and Industrial
Health 15: 231-239.

30
White, A. D., Owen, M. D. K., Hartzler, R. G., Cardina, J. 2002. Common sunflower resistance
to acetolactate synthase–inhibiting herbicides. Weed Science 50: 432-437.
Wuerffel, R. J. et al. 2015. Distribution of the delta G210 protoporphyrinogen oxidase mutation
in iwaterhemp (Amaranthus tuberculatus) and an improved molecular method for
detection. Weed Science 63: 839-845.
Xiang, W., Wang, X., Ren, T., Ci, S. 2006. Expression of a wheat cytochrome P450
monooxygenase cDNA in yeast catalyzes the metabolism of sulfonylurea herbicides.
Pesticide Biochemistry and Physiology 85.
Yang, Q. et al. 2016. Target-site and non-target-site based resistance to the herbicide tribenuron-
methyl in flixweed (Descurainia sophia L.). BMC Genomics 17: 551. DOI
10.1186/s12864-016-2915-8.
Yasuor, H. et al. 2009. Mechanism of resistance to penoxsulam in late watergrass [ Echinochloa
phyllopogon (Stapf) Koss.]. Journal of Agricultural and Food Chemistry 57: 3653-60.
Yu, Q., Han, H., Cawthray, G. R., Wang, S. F., Powles, S. B. 2013. Enhanced rates of herbicide
metabolism in low herbicide-dose selected resistant Lolium rigidum. Plant, Cell &
Environment 36: 818-827.
Yu, Q., Powles, S. 2014. Metabolism-based herbicide resistance and cross-resistance in crop
weeds: A threat to herbicide sustainability and global crop production. Plant Physiology
166: 1106-1118.
Yuan, J. S., Tranel, P. J., Stewart, C. N. 2007. Non-target-site herbicide resistance: A family
business. Trends in Plant Science 12: 6-13.
Yun, M. S., Yogo, Y., Miura, R., Yamasue, Y., Fischer, A. J. 2005. Cytochrome P450
monooxygenase activity in herbicide-resistant and -susceptible late watergrass
(Echinochloa phyllopogon). Pesticide Biochemistry and Physiology 83: 107-114.
Zhao, B. et al. 2017. Non-target-site resistance to ALS-inhibiting herbicides in a Sagittaria
trifolia L. population. Pesticide Biochemistry and Physiology 140: 79-84.

31
CHAPTER III
RNA-SEQ TRANSCRIPTOME ANALYSIS OF AMARANTHUS PALMERI WITH
DIFFERENTIAL TOLERANCE TO GLUFOSINATE HERBICIDE

32
Abstract
Amaranthus palmeri (Amaranthaceae) is a noxious weed in several agroecosystems and in
some cases seriously threatens the sustainability of crop production in North America.
Glyphosate-resistant Amaranthus species are widespread, prompting the use of alternatives to
glyphosate such as glufosinate, in conjunction with glufosinate-resistant crop cultivars, to help
control glyphosate-resistant weeds. An experiment was conducted to analyze the transcriptome
of A. palmeri plants that survived exposure to 0.55 kg ha-1 glufosinate. Since there was no record
of glufosinate use at the collection site, survival of plants within the population are likely due to
genetic expression that pre-dates selection; in the formal parlance of weed science this is
described as natural tolerance. Leaf tissues from glufosinate-treated and non-treated seedlings
were harvested 24 h after treatment (HAT) for RNA-Seq analysis. Global gene expression was
measured using Illumina DNA sequence reads from non-treated and treated surviving
(presumably tolerant, T) and susceptible (S) plants. The same plants were used to determine the
mechanisms conferring differential tolerance to glufosinate. The S plants accumulated twice as
much ammonia as did the T plants, 24 HAT. The relative copy number of the glufosinate target
gene GS2 did not differ between T and S plants, with 1 to 3 GS2 copies in both biotypes. A
reference cDNA transcriptome consisting of 72,780 contigs was assembled, with 65,282
sequences putatively annotated. Sequences of GS2 from the transcriptome assembly did not
have polymorphisms unique to the tolerant plants. Five hundred sixty-seven genes were
differentially expressed between treated T and S plants. Of the upregulated genes in treated T
plants, 210 were more highly induced than were the upregulated genes in the treated S plants.
Glufosinate-tolerant plants had greater induction of ABC transporter, glutathione S-transferase
(GST), NAC transcription factor, nitronate monooxygenase (NMO), chitin elicitor receptor

33
kinase (CERK1), heat shock protein 83, ethylene transcription factor, heat stress transcription
factor, NADH-ubiquinone oxidoreductase, ABA 8’-hydroxylase, and cytochrome P450 genes
(CYP72A, CYP94A1). Seven candidate genes were selected for validation using quantitative real
time-PCR. While GST was upregulated in treated tolerant plants in at least one population,
CYP72A219 was consistently highly expressed in all treated tolerant biotypes. These genes are
candidates for contributing tolerance to glufosinate. Taken together, differential induction of
stress-protection genes in a population can enable some individuals to survive herbicide
application. Elevated expression of detoxification-related genes can get fixed in a population
with sustained selection pressure, leading to evolution of resistance. Alternatively, sustained
selection pressure could select for mutation(s) in the GS2 gene with the same consequence.

34
Introduction
Amaranthus palmeri is a dioecious, weedy Amaranthus species native to Southwestern North
America [1, 2]. It is one of the most widespread, troublesome, and economically damaging
weeds in agronomic crops throughout the southern United States [2]. Infestation of Palmer
amaranth can cause from 70% to more than 90% yield loss in cotton (Gossypium hirsutum) [3],
soybean (Glycine max) [4], and corn (Zea mays) [5]. A. palmeri is difficult to control because of
its rapid growth rate, high fecundity, tiny seeds dispersed by multiple agents, continuous
emergence pattern, high genetic diversity, high propensity for evolving herbicide resistance, and
dioecious nature with long-distance pollen dispersal [1, 6-9]. To date, resistances to six
herbicide mechanisms of action (MOAs) have been confirmed in A. palmeri: acetolactate
synthase (ALS) inhibitors, 4-hydroxyphenylpyruvate dioxygenase (HPPD) inhibitors,
enolpyruvyl shikimate-3-phosphate synthase (EPSPS) inhibitor (glyphosate), mitosis inhibitors
(dinitroanilines), photosystem II inhibitors (triazines), and protoporphyrinogen oxidase (PPO)
inhibitors [10]. The increasing resistance of A. palmeri to herbicides is a threat to corn, cotton,
peanut (Arachis hypogea), and soybean production [1, 11-14]. Alternative management
strategies are needed to combat this problem in several areas in North America [11, 15-18].
Herbicides are used as a major tool for controlling weeds and the evolution of herbicide-
resistant (HR) weeds is an increasing issue worldwide. Glyphosate-resistant (GR) crops, first
commercialized in 1996, were adopted quickly by growers because the technology allowed the
use of the nonselective herbicide, glyphosate, in-season. The technology drastically simplified
weed control with the use of a single, inexpensive, highly effective herbicide. In fact, GR crops
constituted 80% of the 175 million ha planted with transgenic crops globally [19]. However, the
over-reliance on glyphosate and its application over a vast land area has exerted unprecedented

35
selection pressure on weeds, resulting in the evolution of GR weeds including A. palmeri.
Glyphosate-resistant A. palmeri was first identified in Georgia in 2004 [20] and subsequently
became widespread across the South, Midwest and Northeast regions of the United States [10].
The widespread distribution of glyphosate-resistant weeds compelled farmers to use alternative
herbicides including another non-selective herbicide, glufosinate, to control HR weeds in
glufosinate-tolerant crops. Glufosinate is a fast-acting postemergence herbicide that controls
weeds by inhibiting glutamine synthetase (GS) (E.C. 6.3.1.2), the enzyme that converts
glutamate and ammonia to glutamine [21]. Inhibition of GS by glufosinate leads to ammonia
accumulation, inhibition of amino acid synthesis, and indirect inhibition of photosynthesis,
ultimately leading to plant death [22]. To date, resistance to glufosinate has been confirmed in
Eleusine indica from Malaysia [23, 24] and Lolium perenne ssp. multiflorum from Oregon [25].
An amino acid mutation in the chloroplast-encoded GS gene, Asp171Asn, conferred resistance to
glufosinate in L. perenne ssp. multiflorum [26]. Resistance to glufosinate in glufosinate-resistant
crops is achieved using transgenic methods to insert the bar or pat (phosphinothricin
acetyltransferase) gene from a bacterium to the plant’s genome, allowing detoxification of
glufosinate by acetylation [27].
Differential responses to glufosinate in weeds have been attributed to several factors
including light, temperature, humidity, growth stage, application rate, application timing,
species, and variation in the level of herbicide absorption, translocation, and metabolism [28,
29]. Control of A. palmeri by glufosinate is variable [30-32]. A previous study reported higher
uptake, mobility, and metabolism of glufosinate in A. palmeri compared to the more susceptible
Ipomoea lacunosa [28]. As is commonly observed, differential tolerance to herbicides are often
due to non-target-site (NTS) mechanisms, involving the detoxification of herbicide by

36
biochemical modification and/or the compartmentation of the herbicide and its metabolites [33].
Cases of weed resistance to herbicides due to NTS mechanisms have been increasing
(www.weedscience.org). The genetic bases of NTS mechanisms are not fully understood due to
the complex interaction of biochemical processes and limited genomic information on weedy
species [33-35]. In this study, we investigated A. palmeri accessions with differential tolerance to
glufosinate.
The genome of A. palmeri is not yet deciphered although the genome and transcriptome of its
cultivated relative grain amaranth (Amaranthus hypochondriacus) was completed in 2014 [36].
Also recently, the transcriptome of two weedy species Lolium rigidum [37] and Echinochloa
crus-galli [38] were sequenced to identify genes involved in herbicide resistance. Understanding
the molecular mechanisms underlying herbicide resistance could be used to mitigate and manage
resistance evolution and reduce weed impact on crops. This study assembled the transcriptome
sequence of A. palmeri exposed to glufosinate compared to controls to elucidate candidate genes
involved in differential tolerance to glufosinate.
Materials and Methods
Plant Materials
Amaranthus palmeri seed samples from 120 fields were collected in Arkansas, USA between
2008 and 2014. The collection of plant samples from the field was done with permission of farm
owners, managers, consultants, or Extension Agents. In the process of collecting samples, no
endangered species were affected. Inflorescences of at least 10 female plants per field were
harvested, dried, threshed, and cleaned for herbicide bioassays in the greenhouse. One accession

37
of interest (08-Lee-C) was collected from a field that was planted with glyphosate-tolerant
(Roundup Ready®) soybean in 2008 and glyphosate-tolerant cotton in 2006 and 2007. Although
this field had no record of being sprayed with glufosinate, some plants survived exposure to
glufosinate (0.55 kg ha-1) in the greenhouse. The survivors were grown and allowed to cross-
pollinate to produce the first cycle of purified (intercrossed) progeny (C1).
To study the potential survival mechanisms, seeds of 08-Lee-C and the C1 progeny were
planted in 4-cm-diameter pots using commercial potting soil mix (Sunshine Mix, Bellevue, WA,
USA). Seedlings (100) were grown at one plant per pot in a growth chamber maintained at 32/26
°C day/night temperature with a 16-h photoperiod. Plants were watered daily and fertilized with
a water-soluble all-purpose plant food containing 15-30-15% NPK (Miracle-Gro®, Marysville,
OH, USA), every 2 wk. Fifty plants per accession (9-cm tall) were sprayed with glufosinate
(0.55 kg ai ha-1) (Liberty®, Bayer) mixed with 20 g L-1 ammonium sulfate to identify S and T
plants. Susceptible and T plants from the non-treated control were identified by ammonia
accumulation assay. Six confirmed S plants from 08-Lee-C and T plants from C1 were used for
ammonia accumulation assay, determination of chloroplast-encoded glutamine synthetase (GS2)
copy number, and RNA-Seq experiment.
Phenotypic evaluation of A. palmeri response to glufosinate
The response of A. palmeri, collected between 2008 and 2014, was evaluated in the
greenhouse. A known herbicide-susceptible accession (SS) was included in each experiment as
control [39]. Five-hundred mg of seeds from each field-collected plant were mixed to make a
composite seed sample representing each accession. The experiment was conducted twice in a
randomized complete block design with two replications. Each replication consisted of one
cellular tray (28 X 54 cm) with 50 cells (Redway Feed Garden and Pet Supply, Reedway, CA,

38
USA) filled with a commercial medium (Sunshine Mix, Bellevue, WA). Composite seeds from
each accession were planted in each cell and seedlings were thinned to one per cell. Glufosinate
was applied at 0.55 kg ha-1 when seedlings were 7.5- 9 cm tall using a laboratory sprayer fitted
with a flat fan nozzle tip (800067 TeeJet, Spraying Systems Co., Wheaton, IL, USA) delivering
187 L ha-1 at 269 kPa. The herbicide was applied with 20 g L-1 ammonium sulfate. The plants
were assessed visually relative to the non-treated control 21 d after treatment (21 DAT) using a
scale of 0 to 100, where 0 = no visible injury and 100% = complete desiccation. The number of
survivors was recorded. Survivors from glufosinate treatment were grown to produce seed. Data
were analyzed using hierarchal clustering in JMP Pro v. 12.
Herbicide resistance profiling of a selected A. palmeri accession
Data from the differential tolerance evaluation were used to select an accession for further
study. Accession 08-Lee-C had the most number of survivors with minimum injury. Seeds from
08-Lee-C and SS accessions were planted as described in the previous section. Seedlings (7.5-9
cm tall) were treated with the recommended dose of fomesafen (264 g ha-1) (Flexstar®,
Syngenta), glyphosate (870 g ha-1) (Roundup PowerMAX®, Monsanto), dicamba (280 g ha-1)
(Clarity®, BASF), and ALS inhibitors pyrithiobac (73 g ha-1) (Staple LX®, DuPont) and
trifloxysulfuron (8 g ha-1) (Envoke®, Syngenta). The ALS inhibitors were applied with 0.25%
non-ionic surfactant (Induce®, Helena Chemical Co., Collierville, TN, USA), respectively.
Herbicide treatments were applied as described in the previous section. Mortality was evaluated
21 d after treatment. The experiment was conducted in a randomized complete block design as in
the previous section. Data were analyzed using ANOVA in JMP Pro v. 12.

39
Evaluation of tolerance level to Glufosinate
A dose-response bioassay was conducted in the greenhouse to determine the tolerance level
of 08-Lee-C and C1 relative to the SS accession to glufosinate. Seeds were sown in 15-cm
diameter pots filled with commercial potting soil. Seedlings, 7.5-cm tall, were sprayed with 11
doses of glufosinate using a laboratory sprayer as described in the previous section. The 08-Lee-
C and C1 accessions were sprayed with glufosinate from 0.0012 to 0.5940 kg ai ha-1, whereas the
SS accession was sprayed at 0.0006 to 0.5950 kg ha-1, with a non-treated check. The herbicide
was applied with 20 g L-1 ammonium sulfate. Shoot biomass was harvested 21 DAT, dried at
60°C for 72 h, and weighed. The experiment was conducted in a randomized complete block
design with four replications. Five plants were used per replication (20 plants total) per herbicide
concentration.
Data were analyzed using SAS JMP Pro v. 13 in conjunction with SigmaPlot v.13 (Systat
Software, Inc., San Jose, CA, USA) for nonlinear regression analysis. The percentage biomass
reduction was fitted to a nonlinear, sigmoid, four-parameter logistic regression model defined by
𝑦 = 𝑐 + [(𝑑 − 𝑐)/(1 + 𝑒{−𝑎(𝑥 − 𝑏)})]
where y represents the biomass reduction expressed as percentage relative to the non-treated
control, a is the growth rate, b is the inflection point, c is the lower asymptote, d is the upper
asymptote, and x is the glufosinate dose. The herbicide doses that would cause 50% growth
reduction (GR50) were estimated using the fitted regression equation.
Ammonia accumulation assay
To identify S and T plants without glufosinate treatment, a leaf disc assay was conducted to
measure ammonia accumulation caused by the inhibition of photorespiration by glufosinate [40].

40
The assay was conducted using 50 non-treated plants each from 08-Lee-C and the C1 progeny.
In addition, leaf tissues from three 08-Lee-C plants that were controlled (S) and three C1 plants
that survived (T) glufosinate application at the whole plant level were also tested. From each
plant, two leaf discs (5-mm diameter) were excised from the youngest, fully expanded leaf of
6.4-cm tall seedlings. One leaf disc was placed per well in a microtiter plate containing 200 µM
glufosinate. The plate was sealed with micropore tape and placed on a bench under light for 24 h.
The plate was moved to a -80 °C freezer to stop the reaction. After two freeze-thaw cycles,
ammonia content was measured in a spectrophotometer (Pharma Spec UV-1700, Shimadzu,
Columbia, MD) at 630 nm using a modified method by Molin and Khan [41]. Leaf discs from S
plants were expected to have higher ammonia content than those from T plants.
Glutamine synthetase (GS2) relative copy number
Leaf tissues were harvested from confirmed S and T plants (three each) without glufosinate
treatment and stored at -80°C until processing. Leaf tissues were harvested also from plants
treated with glufosinate, 24 HAT. Upon evaluation of plant response 21 DAT, leaf tissues from
three S and T plants were used also to determine the relative copy number of GS2 gene. Genomic
DNA was extracted using the modified hexadecyltrimethylammonium bromide (CTAB) method
[42]. Quantitative real-time PCR (qPCR) was used to determine the genomic copy number of
GS2 relative to a housekeeping RNA dead box helicase gene GS2 in A. palmeri. The primer pair
GS2-F (5’- ATACGGAGAAGGAAGGCAAAG -3’) and GS2-R (5’-
TGTGGGTTCCCAAAGTAGTG-3’) were designed to amplify a region of the chloroplast GS.
RNA dead box helicase gene primers A36-F (5’- TTGGAACTGTCAGAGCAACC-3’) and A36-
R (5’-GAACCCACTTCCACCAAAAC-3’) were used as internal primers to normalize the
samples for any differences in DNA quantities. Reactions were conducted in three technical

41
replicates, and a negative control consisting of primer pairs with no template was included. An 8-
fold serial dilution of genomic DNA samples, ranging from 0.00064 to 50 ng, was used to
construct a standard curve. The slope of the standard curve was used to determine amplification
efficiency (E). The qPCR reaction efficiency was 97% with an R2 of 0.9907 and a slope of 3.271
indicating good assay validation. Genomic DNA templates (2 ng) were amplified in a 25-µL
reaction containing 12.5-µL Bio-Rad iQ SYBR Green Supermix, 2-µL of primers (1:1 mix of
forward and reverse primers), and nuclease-free water. Reaction conditions included 10 min
incubation at 94°C, then 40 cycles of 94°C for 15 s and 60°C for 1 min, followed by a melt-curve
analysis to confirm single PCR product amplification. Data were analyzed using CFX Manager
software (v.1.5). Relative GS2 copy number was calculated as ∆𝐶𝑡 = (𝐶𝑡, 𝐴36 − 𝐶𝑡, 𝐺𝑆2)
according to the method described by Gaines et al [43]. Increase in GS2 copy number was
expressed as 2ΔCt. Results were expressed as the fold increase in GS2 copy number relative to
RNA dead box helicase.
RNA-Seq Analysis
Sample preparation for RNA-Seq
This experiment used leaf tissues from non-treated and treated, confirmed S and T plants.
These were the same plants used for ammonia assay and GS2 copy number determination.
Tissues were collected 24 h after glufosinate application for RNA extraction. This collection
time was selected to capture herbicide stress adaptation genes and because maximum absorption
of glufosinate occurs 24 HAT [28]. Treatments were designated as non-treated S (susceptible
without treatment, SWO), non-treated tolerant (tolerant without treatment, TWO), treated
susceptible (SWT), and treated tolerant (TWT) plants with three biological replicates. Total RNA

42
was extracted from young leaf tissues of S and T plants using PureLink RNA Mini Kit (Life
Technologies, Carlsbad, CA, USA) following the manufacturer’s protocol. The extracted RNA
was treated with DNase (Invitrogen, Carlsbad, CA, USA) to remove potential genomic DNA
contamination, according to the manufacturer’s instructions. The samples were then sent to the
Clemson University Genomics and Computational Biology Laboratory, South Carolina for
sequencing the transcriptome.
Transcriptome sequencing and assembly
Total RNA was normalized and converted to cDNA using the TruSeq RNA library kit v2.0
(Illumina). Final sequencing products were validated for size on an Agilent Bioanalyzer 2100
(Agilent Technologies, Waldbronn, Germany) and sequenced using a 2x125bp paired-end
sequencing module on an Illumina HiSeq 2500 (Illumina). Raw sequence reads were assessed for
quality using the FastQC software package
(http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) and preprocessed to remove
sequence adapters and low quality bases with the Trimmomatic software [44]. A reference
unigene assembly comprehensive of developmental stage, tissue source, and experimental
conditions was prepared by concatenating all preprocessed reads and assembling with the Trinity
software package [45]. The resulting unigene assembly was filtered for genuine coding
sequences (e.g., sequences without internal stop codons or chimeras) with the TransDecoder
software, and clustered by identity with the CD-HIT software [46] in an attempt to collapse
homologs, but not paralogs, at high identity thresholds.
Differential gene expression (DGE)
Paired-end reads from each individual were aligned to the de novo transcriptome using the
Subread package [47, 48]. Samtools was used to convert alignments from sam to bam format,

43
sort, and index [49, 50]. Subread’s featureCounts counted the number of reads that aligned to
each gene in the transcriptome [47, 48]. The final gene counts were loaded into Bioconductor’s
edgeR package for statistical analysis [51-55]. Variance between samples was visualized by a
multidimensional scaling (MDS) plot. Volcano plots were generated for each comparison of
samples. The criteria for differential gene expression included a fold-change >2 between
compared groups and statistical significance at P<0.05 [56]. Expression differences were
compared between non-treated T and non-treated S (TWO vs SWO), treated T and treated S
(TWT vs SWT), treated T and non-treated T (TWT vs TWO), and between treated S and non-
treated S (SWT vs SWO).
Transcriptome annotation
The final reference assembly was annotated by blastx and blastp alignment to the non-
redundant protein database at NCBI, the UniProt-swissprot database, and the uni-ref database to
determine homology to known genes. Protein domains were determined by alignment to the
HMMER database (http://hmmer.janelia.org). Signal peptides were determined with the SignalP
software [57] and transmembrane regions predicted with tmHMM
(http://www.cbs.dtu.dk/index.shtml). Gene ontology terms were derived from the best BLAST
match [58] and clustered to determine enrichment using the Agbase tool [59].
The entire dataset was submitted as NCBI BioProject (PRJNA390774), which is a part of the
U.S. National Library of Medicine of the National Institutes of Health. The 12 samples that were
used to construct the transcriptome, and to run the differential gene expression comparisons were
submitted as 12 separate BioSamples, SAMN07260017-SAMN07260028. The trimmed, paired-
end fastq files for each of the 12 samples were submitted to the Sequence Read Archive,
SRR5759376 – SRR5759387. Finally, the transcriptome was submitted to the Transcriptome

44
Shotgun Assembly Sequence Database. The transcriptome, consisting of 72,780 transcripts, is
under TSA submission: SUB2788796.
Sequence analysis of the glutamine synthetase 2 (GS2) gene
Glutamine synthetase 2 (GS2) gene sequences of the T and S biotypes were extracted and
assembled from the transcriptome data. A 1296-bp GS2 gene (431 amino acids) from S and T
plants was sequenced. The full length GS2 sequences of S and T plants were aligned using
Sequencher 5.4.6 and BioEdit software packages to identify amino acid substitutions. Sequence
alignment also included GS2 sequences of other Amaranthus species (A. viridis, A. albus, A.
spinosus, A. hybridus, A. lividus and A. thunbergii) available in the database at
http://www.weedscience.org.
Heat Map Analysis
Differentially expressed genes associated with abiotic stress response were subsampled and
subjected to heat map analysis. Normalized read count averages were calculated to produce
biological expression profiles followed by hierarchical clustering to recursively merge
expression based on pair-wise distances between non-treated T (TWO), non-treated S (SWO),
glufosinate-treated T (TWT), and glufosinate-treated S (SWT) samples. Digital expressions were
visualized between rows for normalized read count numbers (minimum & maximum) expression
patterns. The expression pattern was generated using GENE-E tool
(http://www.broadinstitute.org/cancer/software/GENE-E/).
Selection of candidate non-target genes
Genes that were commonly expressed between treated S and T, and between treated T and
non-treated T plants were selected for further evaluation based on their gene ontologies (GO).

45
Genes that were assigned with GO molecular function and biological process related to
metabolism and signaling pathways (oxidoreductase activity, nuclear acid binding transcription
factor activity, hydrolase activity, transferase activity, transmembrane transporter activity,
transferase activity, protein transporter activity, biosynthetic process, small molecule metabolic
process, signal transduction, homeostatic process, immune system process, cell wall
organization, secondary metabolic process, nitrogen cycle metabolic process) were evaluated
based on UniProt and their fold change. Contig assemblies that were consistently upregulated in
the treated T (relative to treated S and non-treated T) with a significant P-value in the DESeq
analysis were selected, for a total of 49 contigs. A subset of this list was generated based on
known gene function. Contigs with predicted annotations related to stress response, signaling,
transcription factors, and herbicide metabolism were selected as potential candidate NTS genes
involved in glufosinate tolerance.
Candidate gene validation by qRT-PCR
Two A. palmeri populations were treated with glufosinate at 0.37 kg ha-1 using the previously
described procedure. Leaf tissues were collected three days before and 24 h after herbicide
treatment. Tolerant and susceptible plants from each population were identified three weeks after
herbicide treatment based on level of injury. Three biological replicates from the non-treated and
treated samples from each biotype within each population were used for the validation
experiment. Total RNA was extracted from leaf tissues using PureLink RNA Mini kit (Life
Technologies, Carlsbad, CA). Genomic DNA was removed using DNAse I (Thermo Scientific,
Waltham, MA). cDNA was generated from 5 µg total RNA using Reverse Transcription System
first-strand cDNA synthesis kit ( Promega). A 4-fold serial dilution of cDNA samples (1:1, 1:5,
1:25, 1:125) was used to construct a standard curve. Seven of the candidate NTS genes were

46
subjected to real-time quantitative PCR with primers designed using Primer3 tool. Two genes (β-
tubulin, RNA helicase) were used as internal controls for normalization of gene expression.
Primers had an amplification efficiency of 96 to 110%.
The expression level of 7 candidate NTS genes was measured in 24 plants. Quantitative real-
time PCR reactions were conducted in a 12-µL volume containing 6.25 µL of SyberGreen Master
Mix, 1 µL of 1:25 diluted cDNA, and 0.5 µL of 10 µM primers (1:1 mix of forward and reverse
primers). Amplification was performed in a Bio-Rad MiniOpticon System PCR machine (CFX96,
Bio-Rad Laboratories, Inc., Hercules, CA, USA) under the following conditions: 10 min at 94 oC,
40 cycles of 94 oC for 15 s and 60 oC for 1 min, followed by a melt-curve analysis to confirm
single PCR product amplification. Data were analyzed using CFX Manager software (v.1.5).
Slopes for target and internal control genes were equivalent as observed in amplification plots.
Comparative CT method was used to calculate relative expression levels as 2−ΔΔCt where ΔCT =
[CT target gene–geometric mean (CT internal control genes)] and ΔΔCT = [ΔCt tolerant - ΔCt
susceptible]. Wilcoxon non-parametric test (α=0.05) was used to determine statistical difference
in gene expression between tolerant and susceptible biotypes.
Results
Differential response of A. palmeri accessions to glufosinate
The majority of accessions were 100% sensitive to 0.55 kg ha-1 glufosinate, except for some,
which had few survivors. The 120 accessions differentiated into three groups based on mortality
and levels of injury of survivors (Table 1). The first group is composed of the 88 most sensitive
accessions. The second group, composed of 28 accessions, were controlled 94 to 99% with
survivors incurring 60-99% injury. The third group consisted of four apparently segregating

47
accessions with 88 to 97% mortality. Survivors from these accessions incurred 30-80% injury.
Of the possibly segregating accessions, only survivors from 08-Lee-C were able to produce
sufficient seeds to continue further experiments and evaluation. Seven percent of 08-Lee-C
survived glufosinate treatment, of which 4% of treated plants had <61% injury and produced
seeds. Survivors from other recalcitrant accessions were not able to produce enough seeds due to
either having high injury (>75%) or low number of survivors, which were all males. Considering
that plants growing in the field tend to be more robust than those in the greenhouse, the
likelihood of expected escapes in the field may be higher than that in the greenhouse. Plants in
the field also do not receive uniform amounts of herbicide for various reasons such as partial
coverage by other plants or differential plant size. In addition, plants maintained in the
greenhouse that are well-watered and cultured under warm temperatures grow faster and reach
the recommended spraying heights earlier than those growing in the field due to less variation in
ambient conditions in the greenhouse [60].
Response of 08-Lee-C to other foliar herbicides
The 08-Lee-C accession was susceptible to dicamba and fomesafen, but resistant to
glyphosate and ALS inhibitors, which are commonly used herbicides. The mortality of 08-Lee-C
was 98% and 99% with fomesafen and dicamba, respectively (Table 2). On the other hand, 08-
Lee-C was controlled poorly with glyphosate (EPSPS inhibitor) (61%) as well as with ALS
inhibitors trifloxysulfuron (25%), and pyrithiobac (21%). This accession is resistant to two
commonly used modes of action, thus limiting the herbicide options for post-emergence weed
mitigation.

48
Resistance level to glufosinate
The response of A. palmeri to 11 doses of glufosinate fitted a sigmoidal, logistic function
(Fig. 1). The glufosinate doses required to reduce growth by 50% (GR50) were 0.076, 0.110, and
0.214 kg ha-1 for SS, 08-Lee-C, and C1 accessions, respectively (Table 3). Based on these GR50
values, the level of tolerance to glufosinate in 08-Lee-C and and C1 accessions was 1.4- and 2.8-
fold, respectively, relative to the susceptible standard (SS). The GR50 increased 2-fold, from 110
g ha-1 in 08-Lee-C to 214 g glufosinate ha-1 in C1.
Ammonia accumulation in response to glufosinate
Glutamine synthetase, the target site of glufosinate, is a nuclear-coded enzyme that catalyzes
the conversion of L-glutamate to L-glutamine by assimilating ammonia in the cytoplasm and
plastids, but predominantly in the chloroplast of green tissues [21]. Ammonia accumulation is a
direct response to the inhibition of this pathway by glufosinate. The ammonia concentration in S
plants was 830 µg g-1 fresh leaf tissue (+60) and was 394 µg g-1 fresh weight (+40) in T plants
(Fig. 2). The S plants accumulated 2X more ammonia than the T plants, indicating rapid
depletion of functional glutamine synthetase as a consequence of glufosinate treatment.
Glutamine synthetase 2 (GS2) relative copy number
The relative GS2 copy number of S and T plants ranged from 1 to 3 (Fig. 3). Similar GS2
copies were detected in both S and T plants, indicating that differential tolerance to glufosinate is
not due to amplification of the GS2 gene. Transcriptome analysis also revealed that GS2 was not

49
differentially expressed between T and S plants, which indicated that differential tolerance to
glufosinate in A. palmeri was not due to changes in expression of the target enzyme.
Glutamine synthetase gene sequence analysis
Glutamine synthetase plays a primary role in plant nitrogen metabolism by catalyzing the
conversion of glutamate to glutamine [21]. Glutamine synthetase in higher plants exists in two
major isoforms: GS1 in the cytosol and GS2 in the chloroplast/plastids [61]. The cytosolic form
(GS1) is the predominant isoform in roots and non-green tissues [62]. The chloroplast form of
glutamine synthetase (GS2) is the major isoform in leaves, which is primarily responsible for
recycling ammonia during photorespiration and synthesis of glutamine [63]. In our study, two
different alleles of GS2 were observed in the T plants and one in the S plants. The nucleotide
sequences obtained from the two biotypes had 97-99% identity with GS2 sequences from other
Amaranthus species (A. albus, A. hybridus, A. spinosus, A. lividus, A. thunbergii, and A. viridis).
The T biotype differed in six amino acids in the upstream region of GS2 when compared to the S
biotype. Seven nonsynonymous point mutations (Tyr8Asn, Ser25Leu, Asn26Ser, Lys37Gln,
Gly39Lys, Gln54Lys, Asp56Glu) in the upstream region were detected in one of the GS2 alleles of
the T biotype (Fig. 4). The second allele of the T biotype harbored only the Tyr8Asn substitution.
These nonsynonymous substitutions identified in the A. palmeri T biotype also occur in
herbicide-susceptible A. viridis, indicating that these substitutions do not contribute to tolerance
to glufosinate. Other nucleotide polymorphisms between T and S plants were synonymous
mutations.

50
Global transcriptional changes in A. palmeri 24 h after glufosinate application
A reference cDNA transcriptome consisting of 72,794 sequences was assembled (Table 4).
Treatment samples were similar as indicated in the multidimensional scaling (MDS) plot (Fig.
5). Biological replicates from the same treatment clustered together indicating low bias and
variation among treatment samples. One or more GO terms were assigned to 33,516 sequences
with 76,455 GO assignments in total for biological process (31.9%), cellular component (10.8%)
and molecular function (57.3%) categories.
Background differences between susceptible and tolerant plants. Pairwise comparison
between non-treated S and T plants showed 438 differentially expressed genes, 158 of which
were downregulated and 280 were upregulated in the T plants relative to S plants (Table 5).
Genes that were notably more expressed in the T plants relative to the S plants without herbicide
treatment included cytochrome P450s (Cyp72A219, Cyp86b, Cyt77A, Cyt71A, Cyt76A, Cyt86A),
transporters (ABC transporter), transferases (glycosyltransferase, acyltransferases), antioxidants
(glutathione-S-transferase, superoxide dismutase), and genes related to lipid metabolism
(esterase lipase).
Genes induced by glufosinate application. Relative to the respective non-treated checks,
8154 genes were affected by glufosinate application in the T plants and 6034 genes in the S
plants (Table 5, Figs. 6 and 7). Comparison between treated T and S plants revealed 567 genes
that were more repressed or more induced by glufosinate in the treated T plants. Overall, there
were 210 upregulated and 357 downregulated genes in the treated T relative to the treated S
plants (Figs. 8 and 9). One hundred-five glufosinate-responsive genes were differentially
expressed in both treated T (32 genes) and S plants (73 genes) (Fig. 7). In addition, 239 genes
that were differentially expressed between treated (52 genes) and non-treated (187 genes) T

51
plants were more highly repressed or more induced in treated T plants than in treated S plants. Of
these 239 genes, the majority were related to biosynthetic process, cellular nitrogen compound
and small molecule metabolic processes, response to stress, and oxidoreductase activity (Fig. 9).
Among the upregulated genes of this 239-gene subset, 91 were induced by glufosinate in T
plants, including genes putatively annotated as NAC transcription factor, CYP94a1, and ABC
transporter b. The majority of upregulated genes that were differentially expressed between
treated T and S plants, and that were also differentially expressed relative to their respective non-
treated counterparts, were related to nitrogen compound metabolic processes, oxidoreductase
activity, nucleotide binding, and transferase activity. Two genes that were repressed exclusively
in the treated T plants relative to the treated S plants were folypolyglutamate synthase and
caffeic acid 3-o-methyltransferase (Fig. 7).
The functional classification of selected differentially expressed genes (DEGs) associated
with stress response and herbicide metabolism was examined to investigate the pattern of
transcriptome regulation that occurred during glufosinate treatment (Fig. 11). Glufosinate
treatment triggered the expression of genes related to stress response and xenobiotic
detoxification as expected. Some genes associated with photosynthesis, structural stabilization,
cell membrane binding, stress response, and detoxification were repressed. Increased expression
of non-target site (NTS) genes related to stress response, stress signaling, detoxification, abiotic
response, cell structure stabilization, and growth and senescence was observed in treated T
plants. Genes that were exclusively induced in treated T plants were annotated to code for
transmembrane protein 45b, heat stress transcription factor B, hypersensitive-induced response
protein, cytochrome P450 (Cyp72A219, 94A2, Cyt86b1-like), transcription factor, ethylene-
responsive transcription factor, glutathione S-transferase (GST), zinc finger protein constans-like

52
10, and NAC transcription factor (Fig 8, Tables 5 and 6). These candidate genes likely play a role
in the adaptation of A. palmeri to glufosinate and, possibly, also to other herbicides.
Of the 239 genes that were more highly induced or repressed in the treated T plants, 49
were consistently upregulated. These consistently upregulated genes are involved in biosynthetic
process, cellular nitrogen compound metabolic process, nucleic acid binding transcription factor
activity, oxidoreductase activity, stress response, signal transduction, transferase activity,
transmembrane transporter activity, and transport (Table 6 and Fig. 10). A subset of 13
glufosinate-inducible genes which are related to detoxification, stress signaling, and transport are
candidate genes involved in conferring some tolerance to glufosinate. These include ABA 8’-
hydroxylase, ABC transporter, chitin elicitor receptor kinase, cytochrome P450 72A, cytochrome
P450 94A, GST, heat stress transcription factor, heat shock protein 83, ethylene response
transcription factor, NAC transcription factor, NAC transcription factor 25, NADH ubiquinone
oxidoreductase, and nitronate monooxygenase (NMO) (Table 7). These genes were induced >2-
fold in treated T plants relative to treated S plants and non-treated T plants.
Validation of selected genes using qRT-PCR
The expression of seven candidate NTSR genes was measured in two A. palmeri accessions
using quantitative real-time PCR. Gene expression was similar in the non-treated tolerant and
susceptible plants. The 7 genes were induced in T and S plants upon glufosinate treatment. Five
genes (HSP, NMO, ETF, ABC, NAC) were not differentially expressed between treated tolerant
and treated susceptible plants. GST was differentially expressed in only one of the treated tolerant
plants. On the other hand, CYP72A219 was expressed eight times higher in all the treated tolerant
plants relative to the susceptible ones (Fig. 12).

53
Discussion
Differential tolerance to glufosinate in A. palmeri accessions
The differential response to glufosinate in the 120 Amaranthus palmeri accessions
demonstrates variation in herbicide efficacy on a weed population (Table 1). Several factors
affect glufosinate activity even on a single species; these include temperature, light, relative
humidity, time of day, plant size/age, and dose [30, 64-69]. Variation in environmental
conditions was minimal in the greenhouse. Variation in plant factors was minimized by
maintaining seedlings of the same size. The impact of time of day on glufosinate activity was
eliminated by applying the herbicide at about the same time in each repetition of the experiment.
Accessions with a few survivors reflect some heterogeneity within the population, as expected of
this highly diverse species. Projecting such diversity to field conditions, where all the factors
mentioned above can vary, we expect to see a higher frequency of individuals that would escape
weed control activities in the field. Many such escapes could have been subjected to sub-lethal
doses as a consequence of plant, environmental, and application variables. Controlling these
escaped individuals, or adopting a weed management strategy that controls escaping genotypes,
is a critical step in mitigating the accumulation of non-target-site genes that could eventually
endow resistance to herbicides [70].
Resistance profiling of the 08-Lee-C recalcitrant accession
Herbicides impose strong abiotic stresses to weeds in crop fields, managed turfgrass,
gardens, and roadsides. The evolution of resistance in populations of weedy species is an
increasing problem worldwide. The recalcitrant A. palmeri accession, 08-Lee-C, was resistant to

54
glyphosate and ALS-inhibiting herbicides, trifloxysulfuron and pyrithiobac (Table 2). The
occurrence of resistance to multiple herbicides in 08-Lee-C is not surprising because this field
was sprayed with glyphosate and ALS inhibitors for several years. This field was planted with
glyphosate-tolerant crops for more than three years and had been exposed to ALS inhibitors in
the years prior when the grower was planting conventional soybean. Resistance to glyphosate
and ALS inhibitors among A. palmeri in Arkansas is widespread [71]. Fomesafen, a PPO
herbicide with soil and foliar activity, was first commercialized in the 1960s and had been used
by farmers mainly for soybean. The usage of fomesafen, and almost all other herbicides in
soybean, dropped when glyphosate-tolerant soybean was introduced in the mid-1990s. Upon the
explosion of glyphosate-resistant Amaranthus species, soybean farmers reverted to using
fomesafen and its use was expanded to cotton to control glyphosate-resistant Amaranthus
species. The farmer of this field, like many others, had been using only glyphosate to control
weeds. Although glufosinate had not been used in this field, some A. palmeri individuals
displayed differential tolerance to glufosinate. Being an obligate outcrossing species, A. palmeri
exhibits high genetic diversity, which facilitates its tendency to evolve herbicide resistance.
Intensive use of glufosinate in this field, in a manner that allows escapes to produce seed, will
accelerate the evolution of resistance through accumulation of multiple low-impact tolerance
genes as demonstrated already in some species, including Lolium rigidum [72] and A. palmeri
[73]. Tolerance traits can accumulate and get fixed in the population as selection pressure
continues.

55
Ammonia accumulation in response to glufosinate
Ammonia accumulation is directly related to glufosinate toxicity. Inhibition of glutamine
synthetase and ammonia accumulation triggers a cascade of reactions, including inhibition of
ribulose-1,5-bisphosphate carboxylase/oxygenase (RUBISCO) enzyme [74] and photosystem
electron flow [75], affecting photosynthesis [30, 76] leading to plant death. Ammonia reduces
pH gradient across the membrane, which uncouples photophosphorylation [75]. Elevated levels
of ammonia accumulated in glufosinate-treated rice and soybean cell cultures [77, 78]. In our
study, glufosinate-sensitive plants accumulated 2X more ammonia than the T plants (Fig. 2).
Similarly, glufosinate-sensitive L. perenne ssp. multiflorum from Oregon accumulated 1.6X
more ammonia than the resistant population [25]. Increased ammonia level in S plants after
glusofinate treatment is the consequence of rapid depletion of functional glutamine synthetase.
Reduced ammonia accumulation in T plants indicates the presence of mechanism(s) that reduce
the impact of glufosinate on plant function. Such mechanisms could either be reduced binding
affinity of glufosinate by target site modification or, mechanisms external to the herbicide-
binding site (NTSM) including detoxification and others. The latter applies to the glufosinate-
tolerant A. palmeri plants.
Glutamine synthetase (GS2) copy number
Gene amplification conferring herbicide resistance has been identified in GR weeds such as
A. palmeri, A. tuberculatus, K. scoparia, and L. multiflorum [43, 79, 80]. The GR plants contain
multiple copies of EPSPS, the target site of glyphosate, which results in increased production of
EPSPS enzyme allowing the plant to function normally despite the presence of glyphosate. This
mechanism has not been observed with other herbicide target genes either because it is exclusive

56
to the EPSPS regulatory process, or simply because it has not been investigated in other
herbicide target genes. Amplification of GS2, in glufosinate-resistant weeds, is not yet reported.
However, a 4- to 11-fold amplification of GS2 in alfalfa cell culture lines resulted in increased
GS enzyme production, endowing resistance to glufosinate [81]. In our study, S and T A. palmeri
had similar copies (1-3) of GS2 indicating that tolerance to glufosinate was not due to GS2
amplification (Fig. 3). This was supported by the fact that GS2 transcripts were not different
between S and T plants.
Glutamine synthetase (GS2) gene sequence
A rare individual in a population may harbor a mutation in the herbicide-binding site that can
alter the folding structure of the protein, resulting in reduced binding affinity of the herbicide.
Resistance to ALS-inhibiting herbicides in weeds, in most cases, is due to mutation(s) in one or
more of the binding domains in the ALS enzyme [82]. The higher frequency of SNPs in GS2 of
T plants could predispose such individuals in the population to accumulate nonsynonymous
nucleotide substitutions. However, genetic polymorphisms may not always get translated to
protein polymorphisms, and only certain amino acid mutations will result in herbicide resistance
[82]. In the current study, most of the polymorphisms observed in the nucleotide sequence of
GS2 in T plants were synonymous. Although seven amino acid substitutions in the upstream
region were detected in one of the alleles of the T biotype, these substitutions also occur in
glufosinate-sensitive A. viridis and is in a region outside of the substrate-binding domain (Fig. 4).
A Tyr8Asn substitution was detected in the two GS2 alleles of the T biotype. Asparagine and
tyrosine are both polar, uncharged amino acids, hence Tyr8Asn substitution may not alter the
physiological and physicochemical stability of the plastid GS enzyme. The presence of this

57
trivial amino acid substitution in glufosinate-tolerant plants as well as similar GS2 copies as that
of the S plants suggests that non-target-site tolerance factors are involved. Transcriptome
analysis could inform us on differential tolerance mechanisms. A glufosinate-resistant L. perenne
ssp. multiflorum from Oregon, USA, which showed similar level of ammonia accumulation to
the tolerant A. palmeri in our study, harbors a Asn171Asp mutation (GAC TO AAC) in the GS2
gene which confers resistance to glufosinate [25].
Tolerance level to glufosinate in 08-Lee-C and C1
After one cycle of selection, the GR50 for C1 increased from 1.44-fold to 2.80-fold relative to
SS, reflecting increased frequency of tolerant plants in C1 (Table 3). Although a low frequency
(<10%) of the plants survived exposure to glufosinate, the increase in GR50 after one cycle of
glufosinate selection is indicative that the population has become less sensitive to glufosinate
after one cycle of purifying selection. Because glufosinate had not been used in the field, the
frequency of survivors was low, and the tolerance level was only 1.44 to 2.80-fold, it is likely
that these plant variants have low-level, non-target site resistance. We are possibly capturing an
early phase of herbicide resistance evolution. Should the population continue to be under
selection pressure from glufosinate, the probability of the population acquiring additional
adaptive alleles and expressing resistance to field use rate of glufosinate would increase.
Transcriptome of A. palmeri and candidate NTS genes involved in glufosinate tolerance
A reference cDNA transcriptome consisting of 72,794 sequences was assembled for A.
palmeri (BioProject PRJNA390774, TaxId 107608). The transcriptome of A. hypochondriacus

58
had 57,658 assembled sequences [36]. Our data demonstrated broad effects of glufosinate on
several metabolic pathways, as expected of a herbicidal compound, including nitrogen
assimilation and metabolism similar to what is reported in Arabidopsis [83]. One of the apparent
functional categories to which glufosinate-responsive genes belong is protein families known to
participate in metabolism, stress response, and defense, with the majority of these genes
potentially associated with abiotic stress response signaling and chemical detoxification
pathways (Figs. 10 and 11). Stress response genes are inducible by many other herbicides or
stress factors. Generally, abiotic stress such as herbicide, salinity, and drought modulates the
expression of genes that are involved in signaling cascades and in transcriptional control [84],
genes that code for proteins involved in membrane protection [85], and genes that are involved in
water and ion uptake and transport [86, 87]. These stress-regulated genes are activated to
counteract the stress effects, maintain homeostasis, and adapt. Cell membrane receptor-kinases,
stress signaling genes, detoxification-related genes, and antioxidants were activated upon
glufosinate treatment in both S and T plants (Fig. 11). Peroxidase and superoxide dismutase, for
example, were upregulated to help counteract the oxidative stress caused by lipid peroxidation
resulting from glufosinate treatment. This indicates that plants undergo extensive transcriptional
adjustment in response to herbicide-induced stress. Activation of herbicide-stress-response genes
is hypothesized to be initiated by a herbicide sensor, which triggers the activation of regulator
genes, which causes a cascade of reactions to either detoxify the herbicide or protect the plant
from herbicide-mediated stress [35].
Two genes, folypolyglutamate synthase and caffeic acid 3-o-methyltransferase, were
repressed in the treated T relative to S plants, but were not differentially expressed in other
pairwise comparisons. Both are involved in one-carbon transfer and phenylpropanoid

59
biosynthesis [88, 89]. The phenylpropanoid pathway serves as a rich source of metabolites in
plants, especially for lignin biosynthesis and the production of flavonoids, coumarins,
hydrocinnamic acid conjugate, cutins and lignins [90, 91]. Phenylpropanoids are involved in
plant defense, structural support and survival [91, 92]. Repression of genes involved in
phenylpropanoid biosynthesis indicate less allocation of carbon resources to these plant products
in the T plants after glufosinate treatment compared to the S plants, indicating carbon allocation
to other intermediates is more critical for survival under herbicide stress.
Thirteen candidate genes were identified which included ABA 8’-hydroxylase, ABC
transporter (ABC), chitin elicitor receptor kinase (CERK1), cytochrome P450 72A
(CYP72A219), cytochrome P450 94A, glutathione S-transferase (GST), heat stress transcription
factor, NAC transcription factor, NAC transcriptor 25, ethylene-response transcription factor
(ETF), heat shock protein 83 (HSP), NADH ubiquinone oxidoreductase and nitronate
monooxygenase (NMO) (Table 6). The expression of these candidate genes was induced by
glufosinate treatment. Delye [35] proposed a model of NTS resistance mechanism in which
herbicide stress triggers the expression of ‘protectors’ and ‘regulators’, as well as epigenetic
modifiers which enable the plant to survive herbicide stress. Protector genes include cytochrome
P450, oxidase, peroxidases, esterases, hydrolases, glutathione S-transferases and transporters,
which play roles in reducing the efficacy of herbicide by detoxification. ‘Regulator’ genes are
involved in transcriptional, post-transcriptional, and post-transductional control such as
transcription factors, micro-RNAs, and kinases [35]. It is noteworthy that the candidate NTS
genes could act as either ‘protector’ or ‘regulator’ based on their functions. Cytochrome P450,
GST, NADH ubiquinone oxidoreductase, and ABC transporter proteins have roles in pesticide
detoxification [93-98]. Chitin elicitor receptor kinase, ABA 8’-hydroxylase, ethylene-response

60
transcription factor, heat stress transcription factor, and NAC transcription factor are involved in
stress response signaling and regulation [35, 89, 99-102].
Of the seven candidate genes subjected to qRT-PCR validation experiment, only two genes,
(cytochrome P450 CYP72A219 and GST) were associated with tolerance to glufosinate (Fig. 12).
The induction of GST and CYP72A219 suggests that T plants are able to deactivate glufosinate to
some extent. Induction of cytochrome P450 and GST facilitate the conversion of the herbicide
into a less toxic metabolite. The biochemical role of cytochrome P450-mediated herbicide
metabolism has been well established in herbicide-resistant weed species. Plant cytochrome
P450s facilitate the detoxification of toxic xenobiotics by catalyzing oxygen- and NADPH-
dependent mono-oxygenation reactions which convert herbicide into a more hydrophilic
metabolite [103]. Hundreds of P450 genes exist in higher plants. For example, Arabidopsis
thaliana and Oryza sativa possess 272 and 458 putative P450 genes, respectively [104]. RNA-
Seq transcriptome analysis of L. rigidum identified CYP72A genes to be involved in metabolic
resistance to diclofop [37]. Non-target-site ACCase and ALS resistance in Alopecurus
myosuroides [95, 96], Stellaria media [105], Lolium [96, 106-108], Sinapis arvensis [109],
Echinochloa phyllopogon [110], and Digitaria sanguinalis [111] were reported previously to be
facilitated by cytochrome P450 enzymes. Upregulation of CYP72A and CYP94A was reported in
a multiple-herbicide-resistant E. phyllopogon population [112]. Similarly, CYP94A1, a plant
cytochrome P450-catalyzing fatty acid omega hydroxylase, was induced by chemical stress in
Vicia sativa and by bentazon treatment in soybean [113, 114]. Some cytochrome P450 genes in
the CYP71A family were also demonstrated to be involved directly in herbicide metabolism in
crops, such as O. sativa CYP71A31 and Zea mays CYP71A28 [115, 116].

61
The involvement of glutathione S-transferases (GSTs) in herbicide resistance is reported in
several weed species. Glutathione S-transferases are ubiquitous enzymes that catalyze the
conjugation of harmful xenobiotics to reduced glutathione, facilitating their metabolism,
sequestration or removal [117]. The primary factor for atrazine selectivity in corn is the activity
of a soluble GST, which detoxifies atrazine by forming an atrazine-glutathione conjugate [118].
In a recent transcriptome study, increased expression of GST is associated with diclofop
resistance in L. rigidum and nicosulfuron tolerance in Z. mays [37, 116]. GST also functions as
an antioxidant, protecting plants from herbicide-mediated oxidative stress by scavenging reactive
oxygen species [119]. Increased expression of glutathione transferase gene (AmGSTF1) in
multiple-resistant A. myosuroides led to accumulation of flavonoids which protects the plant
from herbicide injury [120]. It has been reported that glutathione transferase orchestrate
tolerance to abiotic stress through their ability to regulate redox signaling pathways that activate
defense genes [121]. In glufosinate-tolerant A. palmeri, GST is possibly involved in converting
glufosinate into a less toxic metabolite following possible minimal phase I detoxification by
CYP72A219 as well as in protecting the plants against oxidative stress and lipid peroxidation
from glufosinate phytoxicity.
Other candidate genes were identified by RNASeq, but were not differentially expressed in
the validation experiment, including as the ABC transporter, NAC transcription factor, NMO,
HSP, and ethylene transcription factor. Although these might not be involved in conferring some
level of tolerance to glufosinate, their involvement in detoxification of toxic xenobiotic
compounds and in stress response have been reported. For example, NMO in A. thaliana is
associated with detoxification of the allelochemical benzoxazolin [122]. Increased expression of
NADH ubiquinone oxidoreductase and induction of P450 genes were involved in resistance to

62
pyriproxyfen insecticide in Bemisia tabaci [123]. Induction of heat shock proteins is associated
with drought and oxidative stress in Brassica juncea [124] and A. thaliana [125], respectively.
Plant ABC transporters have been associated with the movement of herbicide conjugates [126,
127]. Modifications of ABC transporters have been suspected in some cases of weed resistance
to glyphosate or paraquat [128, 129]. Tolerance of Arabidopsis thaliana to paraquat is endowed
by a mutation in the plasma membrane-localized ABC transporter, which resulted in reduced
herbicide uptake in plant cells [130]. NAC transcription factors and ethylene-response
transcription factors play an important role in the regulation of the transcriptional reprogramming
associated with plant stress response such as cold, drought, and salinity [131-134].
Diversity in gene expression and regulation is an important factor driving herbicide
resistance evolution [135]. As gene expression regulation also involves post-transcriptional and
post-translational controls, protein expression of the identified genes may need to be further
investigated. Because of genetic diversity, plants have the potential to overcome herbicide stress
through a concerted effort of multiple genes. Plants with low-level tolerance showed greater
induction of abiotic stress-protection- and detoxification-related genes than S plants. Thus,
survival from glufosinate treatment is facilitated by stress-protection/stress-adaptation genes.
Differential expression of stress-protection genes in a population can enable some individuals to
survive herbicide application. Tolerance-related genes can get fixed in a population upon the
exertion of sustained selection pressure. Selection pressure coupled with genetic diversity drives
evolutionary processes leading to herbicide resistance.
A low frequency of a low-level presumed tolerance in the phenotype was observed across a
two-stage screening of survivors from a recalcitrant, segregating population. The surviving
individuals were described as tolerant because there was no record of their exposure to

63
glufosinate in this field. However, a three-year record, which is what growers can provide
generally, is insufficient to assert that the gene regulation demonstrated here is truly ancestral
and that it would have been expressed in the total absence of glufosinate selection. Amaranthus
palmeri is a prolific dioecious plant. Seed could have been transported to the field from
elsewhere where glufosinate has been used. Similarly, pollen from other fields with a history of
glufosinate use, could have blown through the field and fertilized female plants. Differential
gene expression of CYP72A219 and GST and the presence of surviving progeny in intentionally
intercrossed populations show that genetic mechanisms exist for the evolution of low-level, or
potentially incipient, resistance to glufosinate in some A. palmeri populations.
Natural variation in sensitivity to herbicides among individuals exists in A. palmeri
population. The inherent ability of A. palmeri to tolerate glufosinate, or any other herbicides, due
to non-target mechanism(s) will impact the dynamics and evolution of herbicide-resistant
populations. The frequency of glufosinate-tolerant biotypes in a population will increase with
continued herbicide exposure. Non-target-genes could accumulate and get fixed in the population
leading to evolution of resistance. Vigilance will be required to detect elevated glufosinate
tolerance, especially because it is multi-genic, impossible to eliminate from populations, and
eventually might confer resistance to more than one class of herbicide chemistries and abiotic
stresses. If proactive measures are ignored, it is just a matter of time that glufosinate-resistant A.
palmeri populations will evolve and cause formidable task in weed management.

64
References
1. Ward SM, Webster TM, Steckel LE. Palmer Amaranth (Amaranthus palmeri): A Review.
Weed Technol. 2013;27: 12-27.
2. Webster TM, Nichols RL. Changes in the prevalence of weed species in the major
agronomic crops of the southern United States: 1994/1995 to 2008/2009. Weed Sci.
2012;60: 145-157.
3. Fast BJ, Murdoch SW, Farris RL, Willis JB, Murray DS. Critical timing of Palmer
amaranth (Amaranthus palmeri) removal in second-generation glyphosate-resistant
cotton. J. Cotton Sci. 2009; 32-36.
4. Bensch CN, Horak MJ, Peterson D. Interference of redroot pigweed (Amaranthus
retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A. rudis) in
soybean. Weed Sci. 2003;51: 37-43.
5. Massinga RA, Currie RS, Horak MJ, Boyer J. Interference of Palmer amaranth in corn.
Weed Sci. 2001;49: 202-208.
6. Sosnoskie LM, Webster TM, Kichler JM, MacRae AW, Grey TL, Culpepper AS. Pollen-
mediated dispersal of glyphosate-resistance in Palmer Amaranth under field conditions.
Weed Sci. 2012;60: 366-73.
7. Steckel LE. The dioecious Amaranthus spp.: Here to stay. Weed Technol. 2007;21 :567-
570.
8. Ehleringer JR, Cerling TE, Helliker BR. C-4 photosynthesis, atmospheric CO2 and
climate. Oecologia. 1997;112: 285-299.
9. Ehleringer J, Forseth I. Solar tracking by plants. Science. 1980;210: 1094-1098.
10. Heap I.The international survey of herbicide resistant weeds.2016. Available from:
www.weedscience.org.
11. Wiggins MS, McClure MA, Hayes RM, Steckel LE. Integrating cover crops and post
herbicides for glyphosate-resistant Palmer amaranth (Amaranthus palmeri) control in
corn. Weed Technol. 2015;29: 412-418.
12. Wiggins MS, Hayes RM, Steckel LE. Evaluating cover crops and herbicides for
glyphosate-resistant Palmer amaranth (Amaranthus palmeri) control in cotton. Weed
Technol. 2016;30: 415-422.
13. Whitaker JR, York AC, Jordan DL, Culpepper S. Palmer amaranth (Amaranthus palmeri)
control in soybean with glyphosate and conventional herbicide systems. Weed Technol.
2010;24: 403-410.
14. Burke IC, Schroeder M, Thomas WE, Wilcut JW. Palmer amaranth interference and seed
production in peanut. Weed Technol. 2007;21: 367-371.
15. Inman MD, Jordan DL, York AC, Jennings KM, Monks DW, Everman WJ, et al. Long-
term management of Palmer amaranth (Amaranthus palmeri) in dicamba-tolerant cotton.
Weed Sci. 2016;64: 161-169.

65
16. Price AJ, Monks CD, Culpepper AS, Duzy LM, Kelton JA, Marshall MW, et al. High-
residue cover crops alone or with strategic tillage to manage glyphosate-resistant Palmer
amaranth (Amaranthus palmeri) in southeastern cotton (Gossypium hirsutum). J. Soil
Water Conserv. 2016;71: 1-11.
17. Meyer CJ, Norsworthy JK, Young BG, Steckel LE, Bradley KW, Johnson WG, et al.
Herbicide program approaches for managing glyphosate-resistant palmer amaranth
(Amaranthus palmeri) and waterhemp (Amaranthus tuberculatus and Amaranthus rudis)
in future soybean-trait technologies. Weed Technol. 2015;29: 716-729.
18. Merchant RM, Culpepper AS, Eure PM, Richburg JS, Braxton LB. Controlling
glyphosate-resistant Palmer amaranth (Amaranthus palmeri) in cotton with resistance to
glyphosate, 2,4-D, and glufosinate. Weed Technol. 2014;28: 291-297.
19. James C. Global status of commercialized biotech/GM crops. Ithaca, New York, USA:
ISAAA, 2013.
20. Culpepper AS, Grey TL, Vencill WK, Kichler JM, Webster TM, Brown SM, et al.
Glyphosate-resistant Palmer amaranth (Amaranthus palmeri) confirmed in Georgia.
Weed Sci. 2006;54:620-626.
21. Wild A, Manderscheid R. The effect of phosphinothricin on the assimilation of ammonia
in plants. Z. Naturforsch.. 1984;39: 500-504.
22. Tachibana K, Watanabe T, Sekizawa Y, Takematsu T. Action mechanism of bialaphos 2:
Accumulation of ammonia in plants treated with bialaphos. J. Pestic. Sci. 1986;11: 33-37.
23. Jalaludin A, Ngim J, Bakar BHJ, Alias Z. Preliminary findings of potentially resistant
goosegrass (Eleusine indica) to glufosinate-ammonium in Malaysia. Weed Biol. Manag.
2010;10: 256-260.
24. Seng CT, Van Lun L, San CT, Bin Sahid I. Initial report of glufosinate and paraquat
multiple resistance that evolved in a biotype of goosegrass (Eleusine indica) in Malaysia.
Weed Biol. Manag. 2010;10: 229-233.
25. Avila-Garcia WV, Mallory-Smith C. Glyphosate-resistant Italian ryegrass (Lolium
perenne) populations also exhibit resistance to glufosinate. Weed Sci. 2011;59: 305-309.
26. Avila-Garcia WV, Sanchez-Olguin E, Hulting AG, Mallory-Smith C. Target-site
mutation associated with glufosinate resistance in Italian ryegrass (Lolium perenne L. ssp
multiflorum). Pest Manag. Sci. 2012;68: 1248-1254.
27. Droge-Laser W, Siemeling U, Puhler A, Broer I. The metabolites of the herbicide L-
phosphinothricin (glufosinate): Identification, stability, and mobility in transgenic,
herbicide-resistant, and untransformed plants. Plant Physiol. 1994;105: 159-166.
28. Everman WJ, Thomas WE, Burton JD, York AC, Wilcut JW. Absorption, translocation,
and metabolism of glufosinate in transgenic and nontransgenic cotton, Palmer amaranth
(Amaranthus palmeri), and pitted morningglory (Ipomoea lacunosa). Weed Sci. 2009;57:
357-361.
29. Everman WJ. Influence of environmental and physiological factors on glufosinate and
glyphosate weed management. PhD Dissertation, North Carolina State University. 2007.

66
Available from: http://0-
search.proquest.com.library.uark.edu/docview/304553564?accountid=8361.
30. Coetzer E, Al-Khatib K. Photosynthetic inhibition and ammonium accumulation in
Palmer amaranth after glufosinate application. Weed Sci. 2001;49: 454-459.
31. Corbett JL, Askew SD, Thomas WE, Wilcut JW. Weed efficacy evaluations for
bromoxynil, glufosinate, glyphosate, pyrithiobac, and sulfosate. Weed Technol. 2004;18:
443-453.
32. Everman WJ, Burke IC, Allen JR, Collins J, Wilcut JW. Weed control and yield with
glufosinate-resistant cotton weed management systems. Weed Technol. 2007;21: 695-
701.
33. Yuan JS, Tranel PJ, Stewart CN. Non-target-site herbicide resistance: a family business.
Trends Plant Sci. 2007;12: 6-13.
34. Delye C, Gardin JAC, Boucansaud K, Chauvel B, Petit C. Non-target-site-based
resistance should be the centre of attention for herbicide resistance research: Alopecurus
myosuroides as an illustration. Weed Res. 2011;51: 433-437.
35. Delye C. Unravelling the genetic bases of non-target-site-based resistance (NTSR) to
herbicides: a major challenge for weed science in the forthcoming decade. Pest Manag.
Sci. 2013;69: 176-187.
36. Sunil M, Hariharan AK, Nayak S, Gupta S, Nambisan SR, Gupta RP, et al. The draft
genome and transcriptome of Amaranthus hypochondriacus: A C4 dicot producing high-
lysine edible pseudo-cereal. DNA Res. 2014;21: 585-602.
37. Gaines TA, Lorentz L, Figge A, Herrmann J, Maiwald F, Ott MC, et al. RNA-Seq
transcriptome analysis to identify genes involved in metabolism-based diclofop resistance
in Lolium rigidum. Plant J. 2014;78: 865-876.
38. Yang X, Yu XY, Li YF. De novo assembly and characterization of the barnyardgrass
(Echinochloa crus-galli) transcriptome using next-generation pyrosequencing. Plos One.
2013;8(7). doi: 10.1371/journal.pone.0069168
39. Salas RA, Burgos NR, Tranel PJ, Singh S, Glasgow L, Scott RC, et al. Resistance to
PPO-inhibiting herbicide in Palmer amaranth from Arkansas. Pest Manag. Sci. 2016;72:
864-869. doi: 10.1002/ps.4241
40. Dayan FE, Owens DK, Corniani N, Silva FML, Watson SB, Howell J, et al. Biochemical
markers and enzyme assays for herbicide mode of action and resistance studies. Weed
Sci. 2015;63: 23-63.
41. Molin WT, Khan RA. Microbioassays to determine the activity of membrane disrupter
herbicides. Pestic. Biochem. Physiol. 1995;53: 17217-17219. doi:
10.1006/pest.1995.1065
42. Sales MA, Shivrain VK, Burgos NR, Kuk YI. Amino acid substitutions in the
acetolactate synthase gene red rice (Oryza sativa) confer resistance to imazethapyr. Weed
Sci. 2008;56: 485-489.

67
43. Gaines TA, Zhang W, Wang D, Bukun B, Chisholm ST, Shaner DL, et al. Gene
amplification confers glyphosate resistance in Amaranthus palmeri. Proc. Natl. Acad.
Sci. U.S.A. 2010;107: 1029-1034. doi: 10.1073/pnas.0906649107
44. Bolger AM, Lohse M, Usadel B. Trimmomatic: A flexible trimmer for Illumina sequence
data. Bioinformatics. 2014;30: 2114-2120. doi: 10.1093/Bioinformatics/Btu170
45. Grabherr MG, Haas BJ, Yassour M, Levin JZ, Thompson DA, Amit I, et al. Full-length
transcriptome assembly from RNA-Seq data without a reference genome. Nat.
Biotechnol. 2011;29: 644-U130. doi: 10.1038/Nbt.1883
46. Li W, Godzik A. Cd-hit: A fast program for clustering and comparing large sets of
protein or nucleotide sequences. Bioinformatics. 2006;22: 1658-1659. doi:
10.1093/bioinformatics/btl158
47. Liao Y, Smyth GK, Shi W. FeatureCounts: An efficient general purpose program for
assigning sequence reads to genomic features. Bioinformatics. 2014;30: 923-930. doi:
10.1093/bioinformatics/btt656
48. Liao Y, Smyth GK, Shi W. The Subread aligner: fFast, accurate and scalable read
mapping by seed-and-vote. Nucleic Acids Res. 2013;41:e108. doi: 10.1093/nar/gkt214
49. Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, et al. The Sequence
Alignment/Map format and SAMtools. Bioinformatics. 2009;25: 2078-2079. doi:
10.1093/bioinformatics/btp352
50. Li H. A statistical framework for SNP calling, mutation discovery, association mapping
and population genetical parameter estimation from sequencing data. Bioinformatics.
2011;27: 2987-2993. doi: 10.1093/bioinformatics/btr509
51. Robinson MD, McCarthy DJ, Smyth GK. EdgeR: A Bioconductor package for
differential expression analysis of digital gene expression data. Bioinformatics. 2010;26:
139-140. doi: 10.1093/bioinformatics/btp616
52. McCarthy DJ, Chen YS, Smyth GK. Differential expression analysis of multifactor RNA-
Seq experiments with respect to biological variation. Nucleic Acids Res. 2012;40: 4288-
4297. doi: 10.1093/nar/gks042
53. Robinson MD, Smyth GK. Moderated statistical tests for assessing differences in tag
abundance. Bioinformatics. 2007;23: 2881-2887. doi: 10.1093/bioinformatics/btm453
54. Robinson MD, Smyth GK. Small-sample estimation of negative binomial dispersion,
with applications to SAGE data. Biostatistics. 2008;9: 321-332. doi:
10.1093/biostatistics/kxm030
55. Zhou XB, Lindsay H, Robinson MD. Robustly detecting differential expression in RNA
sequencing data using observation weights. Nucleic Acids Res. 2014;42:e91. doi:
10.1093/nar/gku310
56. Gaines TA, Lorentz L, Figge A, Herrmann J, Maiwald F, Ott MC. RNA-Seq
transcriptome analysis to identify genes involved in metabolism-based diclofop resistance
in Lolium rigidum. Plant J. 2014;78: 865-876. doi: 10.1111/tpj.12514

68
57. Petersen TN, Brunak S, von Heijne G, Nielsen H. SignalP 4.0: discriminating signal
peptides from transmembrane regions. Nat. Methods. 2011;8: 785-786. doi:
10.1038/nmeth.1701
58. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al. Gene ontology:
tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000;25:
25-29. doi: 10.1038/75556
59. Davis MJ, Sehgal MS, Ragan MA. Automatic, context-specific generation of Gene
Ontology slims. BMC Bioinformatics. 2010; 11:498. doi: 10.1186/1471-2105-11-498
60. Botha GM. Alternative herbicide control options for glyphosate-resistant Palmer
amaranth (Amaranthus palmeri). M.Sc. Thesis, University of Arkansas. 2012. Available
from: http://scholarworks.uark.edu/etd/609
61. Unno H, Uchida T, Sugawara H, Kurisu G, Sugiyama T, Yamaya T, et al. Atomic
structure of plant glutamine synthetase - a key enzyme for plant productivity. J. Biol.
Chem. 2006;281: 29287-29296. doi: 10.1074/jbc.M601497200
62. Miflin BJ, Habash DZ. The role of glutamine synthetase and glutamate dehydrogenase in
nitrogen assimilation and possibilities for improvement in the nitrogen utilization of
crops. J. Exp.Bot. 2002;53: 979-987. doi: 10.1093/jexbot/53.370.979
63. Ireland RJ, Lea, P. J. In: Singh BK, editor. Plants Amino Acids, Biochemistry and
Biotechnology. New York: Dekker, M. Inc.; 1999.
64. Anderson DM, Swanton CJ, Hall JC, Mersey BG. The influence of temperature and
relative humidity on the efficacy of glufosinare-ammonium. Weed Res. 1993;33: 139-
147.
65. Bond JA, Oliver LR, Stephenson DO. Response of Palmer amaranth (Amaranthus
palmeri) accessions to glyphosate, fomesafen, and pyrithiobac. Weed Technol. 2006;20:
885-892. doi: 10.1614/wt-05-189.1
66. Kumaratilake AR, Preston C. Low temperature reduces glufosinate activity and
translocation in wild radish (Raphanus raphanistrum). Weed Sci. 2005;53: 10-16. doi:
10.1614/ws-03-140r
67. Martinson KB, Durgan BR, Gunsolus JL, Sothern RB. Time of day of application effect
on glyphosate and glufosinate efficacy. Crop Manag. 2005:4(1). doi: 10.1094/CM-2005-
0718-02-RS
68. Sellers BA, Smeda RJ, Li JM. Glutamine synthetase activity and ammonium
accumulation is influenced by time of glufosinate application. Pestic. Biochem. Physiol.
2004;78: 9-20. doi: 10.1016/j.pestbp.2003.09.006
69. Steckel GJ, Wax LM, Simmons FW, Phillips WH. Glufosinate efficacy on annual weeds
is influenced by rate and growth stage. Weed Technol. 1997;11: 484-488.
70. Mithila J, Godar A. Understanding genetics of herbicide resistance in weeds:
Implications for weed management. Adv. Crop Sci. and Tech. 2013;1:115. doi:
10.4172/2329-8863.1000115.

69
71. Burgos NR, Alcober, E. A. L., Lawton-Rauh, A., Estorninos, L., Tseng, T. M., Smith,
K. L. The spread and population genetics of glyphosate-resistant Palmer amaranth in
Arkansas.In: Oosterhuis DM, editor. Research Series 573: Summaries of Arkansas Cottn
Research 2008; University of Arkansas, 2009. pp 103-109.
72. Neve P, Powles S. Recurrent selection with reduced herbicide rates results in the rapid
evolution of herbicide resistance in Lolium rigidum. Theor. Appl. Genet. 2005;110: 1154-
1166. doi: 10.1007/s00122-005-1947-2
73. Tehranchian P, Norsworthy JK, Powles S, Bararpour MT, Bagavathiannan MV, Barber
T, et al. Recurrent sublethal-dose selection for reduced susceptibility of Palmer amaranth
(Amaranthus palmeri) to dicamba. Weed Sci. 2017;65: 206-212.
74. Wild A, Wendler C. Inhibitory action of glufosinate on photosynthesis. Z. Naturforsch.
C. 1993;48: 367-373.
75. Logusch EW, Walker DM, McDonald JF, Franz JE. Inhibition of plant glutamine
synthetase by substituted phosphinothricins. Plant Physiol. 1991;95: 1057-1062. doi:
10.1104/pp.95.4.1057
76. Wendler C, Barniske M, Wild A. Effect of phosphinotricin (glufosinate) on
photosynthesis and photorespiration of C3 and C4 plants. Photosynth. Res. 1990;24: 55-
61. doi: 10.1007/bf00032644
77. Tsai CJ, Wang CS, Wang CY. Physiological characteristics of glufosinate resistance in
rice. Weed Sci. 2006;54: 634-640. doi: 10.1614/ws-06-017r.1
78. Pornprom T, Surawattananon S, Srinives P. Ammonia accumulation as an index of
glufosinate-tolerant soybean cell lines. Pestic. Biochem. Physiol. 2000;68: 102-106. doi:
10.1006/pest.2000.2502
79. Jugulam M, Niehues K, Godar AS, Koo DH, Danilova T, Friebe B, et al. Tandem
amplification of a chromosomal segment harboring 5-enolpyruvylshikimate-3-phosphate
synthase locus confers glyphosate resistance in Kochia scoparia. Plant Physiol.
2014;166: 1200-1207. doi: 10.1104/pp.114.242826
80. Salas RA, Dayan FE, Pan ZQ, Watson SB, Dickson JW, Scott RC, et al. EPSPS gene
amplification in glyphosate-resistant Italian ryegrass (Lolium perenne ssp multiflorum)
from Arkansas. Pest Manag. Sci. 2012;68: 1223-1230. doi: 10.1002/ps.3342
81. Donn G, Tischer E, Smith JA, Goodman HM. Herbicide-resistant alfalfa cells: an
example of gene amplification in plants. J. Mol. Appl. Genet. 1984;2: 621-635
82. Tranel PJ, Wright TR. Resistance of weeds to ALS-inhibiting herbicides: What have we
learned? Weed Sci. 2002;50: 700-712. d
83. Abdeen A, Miki B. The pleiotropic effects of the bar gene and glufosinate on the
Arabidopsis transcriptome. Plant Biotechnol. J. 2009;7: 266-282. doi: 10.1111/j.1467-
7652.2009.00398.x
84. Zhu J-K. Cell signaling under salt, water and cold stresses. Curr. Opin. Plant Biol.
2001;4: 401-406. doi: http://dx.doi.org/10.1016/S1369-5266(00)00192-8

70
85. Bray EA. Plant responses to water deficit. Trends Plant Sci. 1997;2: 48-54. doi:
http://dx.doi.org/10.1016/S1360-1385(97)82562-9
86. Blumwald E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol.
2000;12:431-434.
87. Macedo AF. Abiotic stress responses in plants: Metabolism and productivity. In: Ahmad
P, Prasad MNV, editors. Abiotic stress responses in plants: Metabolism, productivity and
sustainability. 1 ed. Spring Stree, New York, USA: Springer-Verlag New York; 2012. p.
473.
88. Srivastava AC, Chen F, Ray T, Pattathil S, Pena MJ, Avci U, et al. Loss of function of
folylpolyglutamate synthetase 1 reduces lignin content and improves cell wall
digestibility in Arabidopsis. Biotechnol. Biofuels. 2015; 8: 224 doi: 10.1186/s13068-015-
0403-z
89. Guo DJ, Chen F, Inoue K, Blount JW, Dixon RA. Downregulation of caffeic acid 3-O-
methyltransferase and caffeoyl CoA 3-O-methyltransferase in transgenic alfalfa: Impacts
on lignin structure and implications for the biosynthesis of G and S lignin. Plant Cell.
2001;13: 73-88. doi: 10.1105/tpc.13.1.73
90. D'Auria JC, Gershenzon J. The secondary metabolism of Arabidopsis thaliana: growing
like a weed. Curr. Opin. Plant Biol. 2005;8:308-16. doi: 10.1016/j.pbi.2005.03.012
91. Vogt T. Phenylpropanoid Biosynthesis. Mol. Plant. 2010;3: 2-20. doi:
10.1093/mp/ssp106
92. La Camera S, Gouzerh G, Dhondt S, Hoffmann L, Fritig B, Legrand M, et al. Metabolic
reprogramming in plant innate immunity: The contributions of phenylpropanoid and
oxylipin pathways. Immunol. Rev. 2004;198: 267-284. doi: 10.1111/j.0105-
2896.2004.0129.x
93. Inui H, Ohkawa H. Herbicide resistance in transgenic plants with mammalian P450
monooxygenase genes. Pest Manag. Sci. 2005;61: 286-291. doi: 10.1002/ps.1012
94. Letouze A, Gasquez J. Enhanced activity of several herbicide-degrading enzymes: A
suggested mechanism responsible for multiple resistance in blackgrass (Alopecurus
myosuroides Huds.). Agronomie. 2003;23:601-608. doi: 10.1051/agro:2003036
95. Reade JPH, Milner LJ, Cobb AH. A role for glutathione S-transferases in resistance to
herbicides in grasses. Weed Sci. 2004;52: 468-474. doi: 10.1614/p2002-168d
96. Cocker KM, Northcroft DS, Coleman JOD, Moss SR. Resistance to ACCase-inhibiting
herbicides and isoproturon in UK populations of Lolium multiflorum: mechanisms of
resistance and implications for control. Pest Manag. Sci. 2001;57: 587-597. doi:
10.1002/ps.330.abs
97. Robineau T, Batard Y, Nedelkina S, Cabello-Hurtado F, LeRet M, Sorokine O, et al. The
chemically inducible plant cytochrome P450 CYP76B1 actively metabolizes phenylureas
and other xenobiotics. Plant Physiol. 1998;118: 1049-1056. doi: 10.1104/pp.118.3.1049
98. Bartholomew DM, Van Dyk DE, Lau SM, O'Keefe DP, Rea PA, Viitanen PV. Alternate
energy-dependent pathways for the vacuolar uptake of glucose and glutathione
conjugates. Plant Physiol. 2002;130: 1562-1572. doi: 10.1104/pp.008334

71
99. Antolin-Llovera M, Petutsching EK, Ried MK, Lipka V, Nurnberger T, Robatzek S, et al.
Knowing your friends and foes--plant receptor-like kinases as initiators of symbiosis or
defence. New Phytol. 2014;204: 791-802. doi: 10.1111/nph.13117
100. Kondo S, Sugaya S, Sugawa S, Ninomiya M, Kittikorn M, Okawa K, et al. Dehydration
tolerance in apple seedlings is affected by an inhibitor of ABA 8 '-hydroxylase
CYP707A. J. Plant Physiol. 2012;169: 234-241. doi: 10.1016/j.jplph.2011.09.007
101. Müller M, Munné-Bosch S. Ethylene Response Factors: A key regulatory hub in
hormone and stress signaling. Plant Physiol. 2015;169: 32-41. doi: 10.1104/pp.15.00677
102. Nuruzzaman M, Sharoni AM, Kikuchi S. Roles of NAC transcription factors in the
regulation of biotic and abiotic stress responses in plants. Front. Microbiol. 2013;4: 248.
doi: 10.3389/fmicb.2013.00248
103. Werck-Reichhart D, Hehn A, Didierjean L. Cytochromes P450 for engineering herbicide
tolerance. Trends Plant Sci. 2000;5: 116-123. doi: 10.1016/s1360-1385(00)01567-3
104. Nelson DR, Schuler MA, Paquette SM, Werck-Reichhart D, Bak S. Comparative
genomics of rice and Arabidopsis. Analysis of 727 cytochrome P450 genes and
pseudogenes from a monocot and a dicot. Plant Physiol. 2004;135: 756-772. doi:
10.1104/pp.104.039826
105. Coupland D, Lutman PJW, Heath C. Uptake, translocation, and metabolism of mecoprop
in a sensitive and a resistant biotypes of Stellaria media. Pestic. Biochem. Physiol.
1990;36: 61-67. doi: 10.1016/0048-3575(90)90021-s
106. Christopher JT, Powles SB, Liljegren DR, Holtum JAM. Cross-resistance to herbicides in
annual ryegrass (Lolium rigidum) 2: Chlorsulfuron resistance involves a wheat-like
detoxification system. Plant Physiol. 1991;95: 1036-1043. doi: 10.1104/pp.95.4.1036
107. Christopher JT, Preston C, Powles SB. Malathion antagonizes metabolism-based
chlorsulfuron resistance in Lolium rigidum. Pestic. Biochem. Physiol. 1994;49: 172-182.
doi: http://dx.doi.org/10.1006/pest.1994.1045
108. Bravin F, Zanin G, Preston C. Resistance to diclofop-methyl in two Lolium spp.
populations from Italy: studies on the mechanism of resistance. Weed Res. 2001;41: 461-
473. doi: 10.1046/j.1365-3180.2001.00250.x
109. Veldhuis LJ, Hall LM, O'Donovan JT, Dyer W, Hall JC. Metabolism-based resistance of
a wild mustard (Sinapis arvensis L.) biotype to ethametsulfuron-methyl. J. Agric. Food
Chem. 2000;48: 2986-2990. doi: 10.1021/jf990752g
110. Yun MS, Yogo Y, Miura R, Yamasue Y, Fischer AJ. Cytochrome P-450 monooxygenase
activity in herbicide-resistant and -susceptible late watergrass (Echinochloa
phyllopogon). Pestic. Biochem. Physiol. 2005;83: 107-114. doi:
10.1016/j.pestbp.2005.04.002
111. Hidayat I, Preston C. Cross-resistance to imazethapyr in a fluazifop-P-butyl-resistant
population of Digitaria sanguinalis. Pestic. Biochem. Physiol. 2001;71: 190-195. doi:
10.1006/pest.2001.2576
112. Iwakami S, Uchino A, Kataoka Y, Shibaike H, Watanabe H, Inamura T. Cytochrome
P450 genes induced by bispyribac-sodium treatment in a multiple-herbicide-resistant

72
biotype of Echinochloa phyllopogon. Pest Manag Sci. 2014;70: 549-58. doi:
10.1002/ps.3572
113. Benveniste I, Bronner R, Wang Y, Compagnon V, Michler P, Schreiber L, et al.
CYP94A1, a plant cytochrome P450-catalyzing fatty acid ω-hydroxylase, is selectively
induced by chemical stress in Vicia sativa seedlings. Planta. 2005;221: 881-890. doi:
10.1007/s00425-005-1503-y
114. Zhu J. An investigation of plant hypersensitive response and photosynthesis inhibition on
a genomic and physiological scale. PhD Dissertation, University of Illinois at Urbana-
Champaign. 2009. Available from: http://0-
search.proquest.com.library.uark.edu/docview/304894215?accountid=8361
115. Saika H, Horita J, Taguchi-Shiobara F, Nonaka S, Nishizawa-Yokoi A, Iwakami S, et al.
A novel rice cytochrome P450 gene, CYP72A31, confers tolerance to acetolactate
synthase-inhibiting herbicides in rice and Arabidopsis. Plant Physiol. 2014;166: 1232-
1240. doi: 10.1104/pp.113.231266
116. Liu XM, Xu X, Li BH, Wang XQ, Wang GQ, Li MR. RNA-Seq transcriptome analysis
of maize inbred carrying nicosulfuron-tolerant and nicosulfuron-susceptible alleles. Int. J.
Mol. Sci. 2015;16:5975-5989. doi: 10.3390/ijms16035975
117. Dalton DA, Boniface C, Turner Z, Lindahl A, Kim HJ, Jelinek L, et al. Physiological
roles of glutathione s-transferases in soybean root nodules. Plant Physiol. 2009;150: 521-
530. doi: 10.1104/pp.109.136630
118. Shimabukuro RH, Frear DS, Swanson HR, Walsh WC. Glutathione conjugation. An
enzymatic basis for atrazine resistance in corn. Plant Physiol. 1971;47: 10-14.
119. Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress
tolerance in crop plants. Plant Physiol. Biochem. 2010;48: 909-930. doi:
10.1016/j.plaphy.2010.08.016
120. Cummins I, Cole DJ, Edwards R. A role for glutathione transferases functioning as
glutathione peroxidases in resistance to multiple herbicides in black-grass. Plant J.
1999;18: 285-292. doi: 10.1046/j.1365-313X.1999.00452.x
121. Roxas VP, Smith RK, Allen ER, Allen RD. Overexpression of glutathione S-transferase
glutathione peroxidase enhances the growth of transgenic tobacco seedlings during stress.
Nature Biotechnol. 1997;15: 988-991. doi: 10.1038/nbt1097-988
122. Baerson SR, Sanchez-Moreiras A, Pedrol-Bonjoch N, Schulz M, Kagan IA, Agarwal AK,
et al. Detoxification and transcriptome response in Arabidopsis seedlings exposed to the
allelochemical benzoxazolin-2(3H)-one. J. Biol. Chem. 2005;280: 21867-81. doi:
10.1074/jbc.M500694200
123. Ghanim M, Kontsedalov S. Gene expression in pyriproxyfen-resistant Bemisia tabaci Q
biotype. Pest Manag Sci. 2007;63: 776-783. doi: 10.1002/ps.1410
124. Aneja B, Yadav NR, Kumar N, Yadav RC. Hsp transcript induction is correlated with
physiological changes under drought stress in Indian mustard. Physiol. Mol. Biol. Plants.
2015;21: 305-316. doi: 10.1007/s12298-015-0305-3

73
125. Scarpeci TE, Zanor MI, Valle EM. Investigating the role of plant heat shock proteins
during oxidative stress. Plant Signal. Behav. 2008;3: 856-857.
126. Gaillard C, Dufaud A, Tommasini R, Kreuz K, Amrhein N, Martinoia E. A herbicide
antidote (safener) induces the activity of both the herbicide detoxifying enzyme and of a
vacuolar transporter for the detoxified herbicide. FEBS Lett. 1994;352: 219-221.
127. Klein M, Weissenbock G, Dufaud A, Gaillard C, Kreuz K, Martinoia E. Different
energization mechanisms drive the vacuolar uptake of a flavonoid glucoside and a
herbicide glucoside. J. Biol. Chem. 1996;271:29666-29671.
128. Powles SB, Yu Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant
Biol. 2010; 61: 317-347. doi: 10.1146/annurev-arplant-042809-112119
129. Ge X, d'Avignon DA, Ackerman JJH, Sammons RD. Rapid vacuolar sequestration: the
horseweed glyphosate resistance mechanism. Pest Manag. Sci. 2010;66: 345-348. doi:
10.1002/ps.1911
130. Xi J, Xu P, Xiang CB. Loss of AtPDR11, a plasma membrane-localized ABC transporter,
confers paraquat tolerance in Arabidopsis thaliana. Plant J. 2012;69: 782-791. doi:
10.1111/j.1365-313X.2011.04830.x
131. Nuruzzaman M, Sharoni AM, Satoh K, Karim MR, Harikrishna JA, Shimizu T, et al.
NAC transcription factor family genes are differentially expressed in rice during
infections with Rice dwarf virus, Rice black-streaked dwarf virus, Rice grassy stunt virus,
Rice ragged stunt virus, and Rice transitory yellowing virus. Front. Plant Sci. 2015;6:
676. doi: 10.3389/fpls.2015.00676
132. Tian Y, Zhang H, Pan X, Chen X, Zhang Z, Lu X, et al. Overexpression of ethylene
response factor TERF2 confers cold tolerance in rice seedlings. Transgenic Res. 2011;20:
857-66. doi: 10.1007/s11248-010-9463-9
133. Zhang Z, Li F, Li D, Zhang H, Huang R. Expression of ethylene response factor JERF1
in rice improves tolerance to drought. Planta. 2010;232: 765-774. doi: 10.1007/s00425-
010-1208-8
134. Zhai Y, Wang Y, Li Y, Lei T, Yan F, Su L, et al. Isolation and molecular characterization
of GmERF7, a soybean ethylene-response factor that increases salt stress tolerance in
tobacco. Gene. 2013;513: 174-183. doi: 10.1016/j.gene.2012.10.018
135. Neve P, Vila-Aiub M, Roux F. Evolutionary-thinking in agricultural weed management.
New Phytol. 2009;184:783-93. doi: 10.1111/j.1469-8137.2009.03034.x

74
Table 1. Cluster analysis of A. palmeri accessions treated with glufosinate at 0.55 kg ha-1.
Cluster Number of
accessions
Mortality (%) Mean frequency of plants at different levels of injury (%)
Mean Min Max 0-10%
injury
11-30%
injury
31-60%
injury
61-80%
injury
81-99%
injury
100%
injury
1 88 100 100 100 0 0 0 0 0 100
2 28 98 94 99 0 0 1 1 0 98
3 4 92 88 97 0 1 3 4 0 92

75
Table 2. Response of A. palmeri (08-Lee-C) to foliar-applied herbicides.
Herbicide Mortality (%)a Mode of actionb
Dicamba 99 Synthetic auxin
Fomesafen 98 PPO inhibitor
Glufosinate 93 Glutamine synthetase inhibitor
Glyphosate 61 EPSP synthase inhibitor
Pyrithiobac 21 ALS inhibitor
Trifloxysulfuron 25 ALS inhibitor
aUniform-sized plants (7.5-9 cm tall) were sprayed with dicamba (280 g ha-1), fomesafen (264 g
ha-1) glufosinate (0.55 kg ha-1), glyphosate (870 g ha-1), pyrithiobac (73 g ha-1), and
trifloxysulfuron (8 g ha-1). Mortality was recorded 21 d after herbicide application. bPPO- protoporphyrinogen oxidase, EPSP- enolpyruvyl shikimate-3-phosphate, ALS- acetolactate synthase

76
Table 3. Glufosinate dose required to reduce growth by 50% (GR50) in A. palmeri 08-Lee-C, C1
and SS accessions.
Accession GR50
Confidence
Intervalsa
T/Sb
kg ai ha-1
08-Lee-C 0.110
0.097 - 0.123 1.44
C1 0.214
0.184 - 0.244 2.80
SSc 0.076
0.064 - 0.088
a 95% confidence intervals.
b Tolerance levels (T/S) calculated using the GR50
of the tolerant accession relative to the susceptible standard. cHerbicide-susceptible standard accession.

77
Table 4. Summary of statistics for transcriptome assembly.
Reads (n) Bases (Mb) Average
length (bp)
Illumina raw reads 1,667,277, 670 8409.7 125
Assembled contigs 72,780 49.15 675
Annotated sequences (blastX) 65,282 - -
Sequences assigned with GO terms 33,294 - -

78
Table 5. Differentially expressed genes putatively involved in differential tolerance to glufosinate in Amaranthus palmeri.
Level of
gene
expression
Number of differentially expressed genes
TWO vs SWO SWT vs SWO TWT vs TWO TWT vs SWT
Repressed Induced Repressed Induced Repressed Induced Repressed Induced
>1-2 2 0 589 353 1277 1048 0 0
>2-3 43 26 1228 994 1846 1229 33 11
>3-4 58 72 662 677 756 679 94 65
>4-5 27 64 328 389 299 351 74 41
>5-6 13 36 109 218 125 182 63 31
>6-7 4 22 72 146 55 135 23 22
>7-10 8 51 45 175 37 113 47 33
>10 3 9 10 39 1 21 23 7
Total 158 280 3043 2991 4396 3758 357 210
438 6034 8254 567
aTWO vs SWO: non-treated tolerant (T) relative to non-treated susceptible (S) plants; SWT vs SWO: treated S relative to non-treated S plants;
TWT vs TWO: treated T relative to non-treated T plants; TWT vs SWT: treated T relative to treated S plants

79
Table 6. Upregulated genes in glufosinate-treated and non-treated tolerant (T) plants, and in
glufosinate-treated T relative to treated susceptible (S) plants, assigned with Gene Ontology
molecular function and biological process related to metabolism and signaling pathways.
GO function Contig Gene annotation Fold change
TWTn/TWOn TWTn/SWTn
biosynthetic process Pa27529 50S ribosomal protein
chloroplastic 8.47 9.02
biosynthetic process Pa29824 60s ribosomal protein l13a-
2 7.60 6.25
biosynthetic process Pa38623 phenazine biosynthesis-
like domain-containing
protein 1 isoform x2
6.93 3.92
biosynthetic process, cellular
nitrogen compound metabolic
process, nucleic acid binding
transcription factor activity
Pa65724 cyclic dof factor 1-like
3.67 4.46
biosynthetic process, small molecule
metabolic process
Pa35601 phosphoribosylaminoimida
zole chloroplastic-like 9.24 11.22
cellular amino acid metabolic
process, biosynthetic process, small
molecule metabolic process
Pa17844 shikimate chloroplastic
2.49 4.41
cellular amino acid metabolic
process, secondary metabolic process
Pa39917 3-isopropylmalate
dehydratase -like protein 4.86 5.79
cellular amino acid metabolic
process, small molecule metabolic
process
Pa60555 probable low-specificity l-
threonine aldolase 1 7.04 5.40
cellular nitrogen compound
metabolic process
Pa52820 putative polyprotein 2.55 4.88
cellular nitrogen compound
metabolic process
Pa63676 CTP synthase 1.64 3.18
cellular nitrogen compound
metabolic process
Pa71553 gag-pol polyprotein 5.93 7.09
cellular nitrogen compound
metabolic process
Pa8879 zinc finger bed domain-
containing protein
ricesleeper 1-like
7.63 9.05
cellular nitrogen compound
metabolic process, biosynthetic
process, signal transduction
Pa63868 two-component response
regulator arr9 isoform x1 2.20 3.43
cellular nitrogen compound
metabolic process, biosynthetic
process
Pa37812 NAC transcription factor
8.25 7.80
cellular nitrogen compound
metabolic process, biosynthetic
process, signal transduction
Pa51700 two-component response
regulator arr5-like isoform
x2
1.62 2.58
cellular nitrogen compound
metabolic process, response to stress,
biosynthetic process
Pa37809 heat stress transcription
factor b-2b-like 3.01 2.88
cellular nitrogen compound
metabolic process, response to stress,
biosynthetic process, signal
transduction
Pa13900 RNA polymerase ii c-
terminal domain
phosphatase-like 1 3.52 3.83
hydrolase activity, acting on glycosyl
bonds
Pa26833 PREDICTED: alpha-
glucosidase-like [Beta
vulgaris subsp. vulgaris]
6.20 4.47

80
Table 6 (Cont.)
GO function Contig Gene annotation Fold change
TWTn/TWOn TWTn/SWTn
hydrolase activity, acting on glycosyl
bonds
Pa42036 alkaline neutral invertase
cinv2-like 3.40 3.15
hydrolase activity, acting on glycosyl
bonds, response to stress
Pa69030 beta-amylase chloroplastic 3.94 5.41
nucleic acid binding transcription
factor activity, biosynthetic process,
signal transduction
Pa49594 auxin-responsive protein
iaa29 4.11 4.51
nucleic acid binding transcription
factor activity, cellular nitrogen
compound metabolic process,
biosynthetic process
Pa47424 NAC transcription factor 25-
like 1.98 3.34
nucleic acid binding transcription
factor activity, cellular nitrogen
compound metabolic process,
response to stress, biosynthetic
process, signal transduction
Pa38292 ethylene-responsive
transcription factor abr1
5.24 4.92
oxidoreductase activity Pa10467 cytochrome P450
cyp72A219-like 8.56 4.42
oxidoreductase activity Pa40402 internal alternative NAD H-
ubiquinone oxidoreductase
mitochondrial
5.22 3.49
oxidoreductase activity Pa45867 nitronate monooxygenase 6.26 4.28
oxidoreductase activity Pa51578 -dopa dioxygenase extradiol-
like protein 6.44 5.06
oxidoreductase activity Pa56011 short-chain type
dehydrogenase reductase-like 7.89 3.99
oxidoreductase activity Pa60473 cytochrome P450 94a1-like 2.90 3.34
oxidoreductase activity, cellular
amino acid & metabolic process,
cellular amino acid metabolic
process, homeostatic process,
oxidoreductase activity
Pa10326 5 -adenylylsulfate reductase
chloroplastic- partial
3.47 4.04
oxidoreductase activity, small
molecule metabolic process
Pa44392 abscisic acid 8 -hydroxylase
2
4.153201 4.072155
response to stress, immune system
response
Pa52955 macpf domain-containing
protein at1g14780
1.801976 3.089814
response to stress, immune system
response
Pa62900 heat shock protein 83 4.420622 4.392136
response to stress, signal
transduction, immune system process
Pa42133 chitin elicitor receptor kinase
1-like
3.354966 3.353194
response to stress, signal
transduction, immune system process
Pa69811 receptor-like protein kinase
at3g47110
3.401606 6.459389
response to stress, transport,
transmembrane transport
Pa53135 mitochondrial phosphate
carrier protein mitochondrial-
like
11.42314 4.210532
signal transduction Pa60381 receptor-like serine
threonine-protein kinase sd1-
8 isoform x1
5.532361 3.68854
signal transduction Pa63442 PREDICTED:
uncharacterized protein
LOC104887975
3.319583 5.558233

81
Table 6. (Cont.)
GO function Contig Gene annotation Fold change
TWTn/TWOn TWTn/SWTn
transferase activity, transferring acyl
groups
Pa67068 uncharacterized
acetyltransferase at3g50280-
like
10.27052 4.376485
transferase activity, transferring alkyl
or aryl (other than methyl) groups
Pa19271 glutathione s-transferase-like
protein
10.47473 6.70153
transferase activity, transferring
glycosyl groups, biosynthetic process
Pa49933 7-deoxyloganetin
glucosyltransferase-like
6.037006 4.239084
transferase activity, transferring
glycosyl groups, response to stress,
bioysnthetic process, small molecule
metabolic process, cell wall
organization or biogenesis
Pa57353 gdp-l-galactose
phosphorylase 2-like
5.457352 5.232313
transmembrane transporter activity Pa14919 peroxisomal nicotinamide
adenine dinucleotide carrier-
like
1.675916 4.465376
transmembrane transporter activity Pa21499 calcium-transporting atpase
plasma membrane-type
4.459466 3.358566
transmembrane transporter activity Pa35784 mate efflux family protein 9-
like
3.45508 3.260395
transmembrane transporter activity Pa63432 anoctamin-like protein
at1g73020
3.788903 3.221265
transmembrane transporter activity,
cellular nitrogen compound
metabolic process, transport, small
molecule metabolic process
Pa63215 ABC transporter b family
member 2-like
4.662064 6.567808
transport Pa60553 outer envelope protein
mitochondrial
3.146434 7.2662
aTWT = treated T plants
TWO = non-treated T plants
SWT = treated S plants

82
Table 7. Candidate non-target genes, identified by RNA-Seq analysis, that are potentially
involved in conferring differential tolerance to glufosinate in A. palmeri.
Contig Gene annotation Fold changea Function
TWTn/TWOn TWTn/SWTn
19271 Glutathione S-transferase (GST) 10.47 6.7 Detoxification
10467 Cytochrome P450 CYP72A219 8.55 4.42 Heme-thiolate
monoxygenase;
detoxification
37812 NAC transcription factor 8.24 7.8 Transcription
regulator in plant
stress response
38292 Ethylene-response transcription
factor abr1
5.24 4.91 ABA signaling
pathway in
response to stress
response
40402 NAD H-ubiquinone
oxidoreductase
5.21 3.49 Detoxification
63215 ABC transporter b family
member 2
4.66 6.56 Transmembrane
transport
62900 Heat shock protein 83 4.31 8.2 Molecular
chaperone; stress
signaling
44392 ABA 8’-hydroxylase 4.15 4.07 ABA catabolism
45867 Nitronate monooxygenase (NMO) 6.26 4.27 Detoxification
42133 Chitin elicitor receptor kinase 1
(CERK1)
3.35 3.35 Cell surface
receptor toward
biotic and abiotic
stresses
37809 Heat stress transcription factor b 3.01 2.87 Transcription
regulator for heat
shock proteins;
stress signaling
60473 Cytochrome P450 94a1 2.9 3.33 Detoxification
47424 NAC transcription factor 25-like 1.97 3.34 Abiotic stress
response aTWT = treated tolerant plants
TWO = non-treated tolerant plants
SWT = treated sensitive plants

83
Figure 1. Shoot biomass reduction (%) of 08-Lee-C, C1, and SS A. palmeri accessions, 21 days
after glufosinate treatment. Data were best described with nonlinear, sigmoidal, four-parameter
logistic regression function.

84
Figure 2. Ammonia content in glufosinate-tolerant (T) and –susceptible (S) A. palmeri.Error bars
represent standard error. White bars = S plants; gray bars = T plants.

85
Figure 3. Relative copy number of A. palmeri GS2 in glufosinate-susceptible (S) and -tolerant
(T) plants. Error bars represent standard deviation of the mean. Gray bars = T plants; black bars
= T plant.

86
Figure 4. Multiple alignment of the plastidic glutamine synthetase (GS2) amino acid sequences
in Amaranthus. A. palmeri (reference A. palmeri), T1 and T2 = GS2 alleles of glufosinate-
tolerant A. palmeri biotype, S = GS2 sequence of glufosinate-susceptible A. palmeri biotype.

87
Figure 5. Multidimensional scaling (MDS) plot showing the relationship between sample
types.TWO = non-treated tolerant, TWT= treated tolerant, SWO = non-treated susceptible, SWT
= treated susceptible.

88
Figure 6. Volcano plots depicting differential gene expression between treatments. A) Treated susceptible (S) relative to non-treated S
plants (SWT vs SWO), B) Treated tolerant (T) relative to treated S plants (TWT vs SWT), C) treated T relative to non-treated T plants
(TWT vs TWO), and D) treated T plants relative to non-treated S plants (TWO vs SWO). The x-axis shows the log fold change or
relative abundance. The P value (-log base 10) for differential gene expression is plotted on the y axis. Black dots represent genes that
did not change in expression; colored dots on the left indicate genes with significantly downregulated expression; colored dots on the
right indicates genes with significantly upregulated expression.

89
Figure 7. The number of differentially expressed genes common or specific to treated and non-
treated T and S plants.A 4-way Venn diagram depicting the distribution of differentially
expressed genes across all pairwise comparisons. The number within each shaded area is the
number of differentially expressed genes common between treatments.

90
Figure 8. Biological processes (A) and molecular functions (B) of upregulated genes in treated T
relative to treated S plants.

91
Figure 9. Biological processes (A) and molecular functions (B) of downregulated genes in
treated T relative to treated S plants.

92
Figure 10. Biological processes (A) and molecular functions (B) of differentially expressed
genes that are common in treated T relative to treated S and non-treated T plants.

93
Figure 11. Heat map analysis of genes that are putatively related to abiotic stress response in A.
palmeri.TWO (non-treated tolerant), SWO (non-treated susceptible), TWT (glufosinate-treated
tolerant), SWT (glufosinate-treated susceptible).

94
Figure 12. Gene expression fold-change of seven candidate NTS genes in glufosinate-tolerant
Amaranthus palmeri relative to –susceptible ones. Tolerant and susceptible plants from two
populations were identified based on the level of injury incurred after glufosinate treatment. GST
and CYP72A219 were highly expressed in the tolerant biotypes in at least one population. Non-
treated = before glufosinate treatment, Treated = 24 h after glufosinate treatment. Error bars
represent standard error from six biological replicates. HSP = heat shock protein, GST =
glutathione S-transferase, NMO = nitronate monooxygenase, ETF = ethylene-responsive
transcription factor, ABC = ABC transporter, NAC = NAC transcription factor, CYP72A219 =
cytochrome P450 CYP72A219.

95
CHAPTER IV
MULTIPLE RESISTANCE TO ALS-, PPO, AND EPSPS INHIBITORS IN
AMARANTHUS PALMERI FROM ARKANSAS, USA

96
ABSTRACT
Palmer amaranth (Amaranthus palmeri) is a major weed problem in agronomic crops in the
United States. The majority of the Palmer amaranth populations from Arkansas are resistant to
ALS inhibitors. Resistance to ALS inhibitors in Palmer amaranth populations is frequently
attributed to target-site mutation, however non-target-site based mechanisms can also be
involved. This study aimed to characterize the herbicide resistance profile and investigate target-
site and non-target-site resistance mechanisms to ALS inhibitors in a Palmer amaranth
population (15Cri-B) from Arkansas. Resistance level (R/S) for flumetsulam, imazethapyr,
trifloxysulfuron, fomesafen, and glyphosate were 4.8, 60.9, 4.4, 8.3, and 32 times, respectively,
for the 15Cri-B relative to the susceptible population. The addition of cytochrome P450 inhibitor
increased trifloxysulfuron phytotoxicity from 25% to 49% with malathion and to 66% with
piperonyl butoxide, suggesting the involvement of P450-mediated herbicide metabolism. A
Ser653Asn substitution in the ALS gene was detected in two out of seven resistant plants tested.
Resistant plants without any known resistance-conferring mutations had increased expression of
CYP81B and GSTF10 genes after trifloxysulfuron treatment, indicating their involvement in
metabolic resistance to ALS inhibitors. This Palmer amaranth population from Arkansas exhibit
both target-site and non-target-site based resistance to ALS inhibitors.

97
INTRODUCTION
Palmer amaranth (Amaranthus palmeri S Watson) is a dioecious, aggressive weedy
species native to the area of the southwest United States and northern Mexico. It has become one
the most damaging and difficult-to-control weeds in major agronomic crops in the United States.
1Palmer amaranth is considered as an opportunistic, invasive and competitive weed due to its
rapid growth rate, continuous emergence period, high fecundity, genetic diversity, and its
capacity to evolve traits that increase its potential to grow and reproduce in various cropping
systems and environmental conditions.2,3 With its invasiveness and competitiveness, Palmer
amaranth can cause significant yield reduction in cotton (Gossypium hirsutum) (77%),4 soybean
(Glycine max) (79%)5, corn (Zea mays) (91%),6 grain sorghum (Sorghum bicolor) (63%),7 and
sweet potato (Ipomoea batatas) (94%).8 In addition to reducing yields, its large size (up to 3 m)
also decreases harvest efficiency as it hampers the operation of combines harvesters. Palmer
amaranth is an obligate outcrossing species which enables it to adapt and spread herbicide
resistance genes quickly. To date, various populations had evolved resistance to six different
mechanisms of action (MOA), namely: acetolactate synthase (ALS), 5-enol-pyruvylshikimate-3-
phosphate synthase (EPSPS), photosystem II, 4-hydroxyphenylpyruvate dioxygenase (HPPD),
microtubule, and protoporphyrinogen oxidase (PPO) inhibitors.9 Palmer amaranth populations in
Arkansas had evolved resistance to ALS inhibitors and glyphosate, and recently to PPO-
inhibiting herbicides which limits herbicide options for growers.10
Herbicides are a major tool in weed management; however, its usefulness is threatened
by the evolution of herbicide-resistant weeds. Herbicide resistance is an evolutionary adaptation
in weed species as a consequence of intensive herbicide selection pressure. Herbicide resistance
mechanism can be classified as either target-site (TSR) or non-target-site (NTSR). Target-site

98
mechanisms include structural changes in the herbicide binding-site due to amino acid
substitution or increased expression of the target protein.11 Compared to TSR, NTSR is less often
reported and its molecular mechanisms are not well understood. With NTSR mechanism, the
number of herbicide molecules reaching the target site is minimized either by reduced herbicide
absorption or herbicide translocation or metabolism of the herbicide into non-toxic metabolites.
Reduction in herbicide absorption is due difference in cuticle properties or other structural
barriers that prevents that affects herbicide retention on the leaves and/or herbicide absorption
through the cuticle.11 Reduced herbicide translocation is characterized by restricted movement of
the herbicide within the plant, often due to sequestration of the herbicide into the vacuole or
inhibition of cellular transport.11-14
Herbicide detoxification usually involves oxidation of the herbicide molecules, usually
carried out by cytochrome P450 monooxygenases (P450s), conjugation of the activated
xenobiotic using thiols or sugars, transport of the conjugated molecules into the vacuole or
extracellular space by active transport, and degradation of the conjugated molecule into the
vacuole or extracellular space.15 NTSR mechanisms involve multiple genes and usually are part
of plant stress response.11 NTSR mechanisms pose a serious concern in weed management
because it can confer unpredictable resistance to a broad range of herbicides, including
herbicides that are yet to be commercialized.
Acetolactate synthase (ALS) inhibitors have been widely used for Palmer amaranth
control since their introduction in 1982.16 Inhibition of ALS enzyme affects the synthesis of
branched-chain amino acids (valine, leucine, and isoleucine), ultimately leading to plant death.
There are five chemical families of herbicide that inhibit ALS: sulfonylureas (SUs),
imidazolinones (IMIs), triazolopyrimidines (TPs), sulfonylaminocarbonyltriazolinones (SACTs),

99
pyrimidinylthiobenzoates (PTBs). The ALS herbicides are widely used because they provide
broad-spectrum weed control at low doses, soil residual activity, wide application windows,
excellent crop safety and low mammalian toxicity.17 However, these herbicides have high
propensity to select resistant weed populations due to strong selection pressure and high levels of
natural variability in the ALS gene.18 The widespread occurrence of Palmer amaranth populations
that had evolved resistance to ALS inhibitors has threatened the continued use of ALS-inhibiting
herbicides.
Most cases of ALS resistance are due to mutation in the ALS gene resulting in an enzyme
that is less sensitive to herbicide binding. To date, 29 amino acid substitutions in ALS that confer
herbicide resistance were identified at Ala122 (5), Ala205 (2), Arg377 (1), Asp376 (1), Gly654 (2),
Pro197 (11), Ser653 (3), and Trp574(4) in weed species.9 Substitution at Trp574 confers resistance to
IMIs, PTBs, SUs, and TPs,18 whereas Asp376Glu and Ala205Phe substitutions confer resistance to
all five chemical families of ALS inhibitors.19, 20 Amino acid substitution at Pro197 conferred
resistance to SUs with low or no cross-resistance to IMIs,18 whereas substitutions at Ala122 or
Ser653 conferred resistance to IMI herbicides with low-level resistance to SUs.21, 22 Recent
investigation on 20 Palmer amaranth populations from Arkansas revealed TSR mechanism
involving Trp574Ser mutation with a few cases of double mutations involving Ala122Thr,
Pro197Ala or Ser653Asn.23
Another way whereby plants can become resistant to ALS inhibitors is reducing the
amount of herbicide reaching ALS to below what is lethal (NTSR). Resistance to ALS inhibitors
is rarely attributed to reduced absorption and translocation,24-26 and in only few cases have they
been reported as partial-resistance mechanism.7, 28 Many studies have attributed enhanced
herbicide metabolism to the cytochrome P450 family of proteins by demonstrating that co-

100
application of the pesticide malathion, a P450 inhibitor, results in loss of resistance or reduced
level of resistance.29-32 An enhanced herbicide metabolism has been reported in ALS-resistant
rigid ryegrass (Lolium rigidum),33 wild mustard (Sinapis arvensis),34 and late watergrass
(Echinochloa phyllopogon).35, 36 Cytochrome P450 genes, CYP81A6 and CYP76B1, confer
resistance to sulfonylurea and phenylurea herbicides, respectively.37, 38 Multiple resistance
mechanisms may also exist in a weed population. Resistance to ALS inhibitor in Spanish corn
poppy (Papaver rhoeas) and flixweed (Descurainia Sophia L.) populations was controlled by
both target-site mutation (Pro197 substitution) and enhanced herbicide metabolism.39, 40
Several studies have identified the involvement of several NTSR genes in conferring
herbicide resistance by whole-transcriptome sequencing.39-42 With the availability of the Palmer
amaranth transcriptome,43 candidate NTSR genes conferring resistance to ALS inhibitors can be
validated. The objectives of this study were to determine the resistance profile of a Palmer
amaranth population from Arkansas to ALS herbicides, fomesafen, and glyphosate as well as to
unravel the resistance mechanism to ALS-inhibiting herbicides.
MATERIALS AND METHODS
Greenhouse experiments were conducted at Fayetteville, Arkansas. One known
susceptible (SS) and one ALS-resistant Palmer amaranth population (hereafter referred to as
15Cri-B) were used in the experiments. The SS seeds were collected from a vegetable field in
Crawford County, Arkansas and have been used routinely for resistance screening due to its
limited exposure to herbicides.44 The 15Cri-B population was collected from a soybean field in
the late summer of 2015. Little is known about the crop management history in this field, except
that the field has been planted with continuous soybean for several years. Inflorescence from at

101
least 10 female plants were collected, dried, threshed, and cleaned for bioassay in the
greenhouse.
Cross-resistance to multiple herbicides
Seeds were planted in 28- X 54- cm cellular trays filled with commercial potting soil
(Sunshine premix No. 1, Sun Gro Horticulture, Bellevue, WA). The experiment was set up in a
randomized complete block design with four replications. Each replication consisted of a tray
with 50 seedlings. Treatments included glufosinate at 549 g ha-1 (Liberty®, Bayer CropScience
LP, Research Triangle Park, NC), fomesafen at 263 g ha-1 (Flexstar®, Syngenta Crop Protection
LLC, Greensboro, NC) and glyphosate at 840 g ae ha-1 (Roundup POWERMAX®, Monsanto
Co., St. Louis, MO) and five ALS inhibitors namely; trifloxysulfuron at 8 g ha-1 (Envoke®,
Syngenta Crop Protection LLC, Greensboro, NC), pyrithiobac at 73 g ha-1 (Staple LX®, DuPont
Crop Protection, Wilington, DE), flumetsulam at 7 g ha-1 (Python®, Dow AgroSciences LLC,
Indianapolis, IN), imazethapyr at 70 g ae ha-1 (Pursuit®, BASF Corp., Research Triangle Park,
NC), and primisulfuron at 40 g ha-1 (Beacon®, Syngenta Crop Protection LLC, Greensboro, NC).
Pyrithiobac and trifloxysulfuron treatments included 0.25% non-ionic surfactant (NIS) (Induce®
non-ionic surfactant, Helena Chemical Co. Collierville, TN) by volume. Imazethapyr was
applied with 0.25% (v/v) NIS and 1.4% (w/v) ammonium sulfate (AMS), whereas primisulfuron
was applied with 0.25% (v/v) NIS and 2244 g AMS ha-1. Flumetsulam, fomesafen, and
glufosinate treatments included 0.5% (v/v) crop oil concentrate (COC) (Agri-Dex® crop oil
concentrate, Helena Chemical Co. Collierville, TN), 0.5% NIS and 3366 g AMS ha-1,
respectively. Herbicides were applied to 7.5-cm tall seedlings using a compressed air, motorized
boom, cabinet sprayer equipped with Teejet XR800067 flat fan nozzle (Teejet spray nozzles;

102
Spraying Systems Co., Wheaton, IL) calibrated to deliver 187 L ha-1 at 269 kPa. Mortality was
assessed 21 d after treatment (DAT) and each surviving plant was visually evaluated for injury
relative to the non-treated control. Injury was recorded on a scale of 0-100% where 0 had no
injury and 100% was dead. Data were analyzed using ANOVA in JMP Pro v.13.
Herbicide dose-response assay
Dose-response experiments were conducted as described by Salas et. al (2016).44 Seeds
were planted in 15-cm-diameter pots filled with commercial potting soil and thinned to 5 plants
per pot. The experiment was conducted in a completely randomized design with four
replications. Seedlings (7.5-cm tall) were treated with flumetsulam (1X = 7 g ha-1), imazethapyr
(1X = 70 g ae ha-1), trifloxysulfuron (1X = 8 g ha-1), glyphosate (1X = 840 g ae ha-1) and
fomesafen (1X = 263 g ha-1). The resistant plants were sprayed with eight herbicide doses
ranging from a 0X to a 8X dose; susceptible plants were ware sprayed with eight doses ranging
from 0X to 2X of the recommended herbicide dose. Visible injury and mortality were evaluated
21 DAT. Plants were cut at the soil surface, dried for 48 h, and weighed. Data were analyzed
using JMP Pro v. 13 in conjunction with SigmaPlot v. 13. Nonlinear regression analysis was
conducted and the data were fitted with a three-parameter log-logistic model (equation 1) to
determine the herbicide dose that would cause 50% control
𝑦 = 𝑐/[1 + 𝑒)−𝑎 (𝑥−𝑏) [1]
where y is the percent biomass reduction, a is the asymptote, b is the slope, c is the inflection
point, and x is the herbicide dose.

103
Whole-plant response to trifloxysulfuron with or without P450 inhibitors
Palmer amaranth plants (15Cri-B and SS) were grown in 15-cm diameter pots filled with
commercial soil mixture potting medium. Seedlings were thinned to 5 plants per pot after
emergence. Uniform sized-plants (7.5-cm tall) were treated with cytochrome P450 inhibitors
malathion (1000 g ha-1) (Hi-Yield Chemical Co., Bonham, TX), carbaryl (1100 g ha-1)
(Tessenderlo Kerley Inc., Phoenix, AZ) and piperonyl butoxide (1200 g ai ha-1) (MGK Co.,
Minneapolis, MN) 45 min before applying trifloxysulfuron at 8 g ha-1. Herbicide and P450
inhibitor applications were made using a laboratory sprayer equipped with a flat fan nozzle
(TeeJet spray nozzles, Spraying Systems Co., Wheaton, IL 60189) delivering 187 L ha-1. A
non-treated control was also provided for each population. At 21 DAT, plant injury was
evaluated visually. Aboveground biomass was harvested and dried at 60oC for 3 d. Results were
expressed as the percent biomass reduction relative to the non-treated control. The experiment
was conducted in a completely randomized design in a factorial treatment arrangement with
P450 inhibitor and herbicide as the main factors. Each treatment had four replications and the
experiment was conducted twice. Data were subjected to analysis of variance in JMP Pro v.13
software. Significant means were separated using Fisher’s protected LSD0.05.
ALS gene sequencing
Leaf tissues from four sensitive (SS) and seven trifloxysulfuron-resistant (15Cri-B) plants
were collected and stored immediately at -80oC freezer. Genomic DNA was extracted using the
modified hexadecytrimethylammonium chloride, previously described by Salas et al.44
Polymerase chain reaction (PCR) was conducted in a 40 µL mixture of 20 µL 2X PCR master

104
mix (Takara Bio USA Inc., San Francisco, CA), 2 µL of both forward and reverse primers (10
µM), 2 µL of genomic DNA (50 ng µL-1), and 16 µL water using MJ Research thermal cycler
(PTC-200, MJ Research, Inc., Waltham, MA). The following primers were used to sequence the
full ALS gene (2 kb): forward primer PAALS_F (5’-ATGGCGTCCACTTCAACAAAC-3’),
reverse primer PAALS_R (5’-GGTGATGGAAGAAGGGCTTATTAG-3); and internal primers
PAALS_F2 (5’-AGGATATTCCTAGAATTGTTAAGG-3’), PAALS_F3 (5’-
ATGCGGTTGTAAGTACCGGTGT-3’), PAALS_R1 (5’-CCTGGACCTGTTTTGATTGATA-
3’). PCR was performed under the following conditions; initial denaturation at 94°C for 2 min,
35 cycles of denaturation at 94°C for 1 min, annealing at 60°C for 1 min, extension at 72°C for 2
min, then final extension at 72°C for 10 min. The PCR amplification products were resolved on a
0.8% agarose gel to confirm the expected fragment size (2 kb). The PCR product was purified
using EZNA gel extraction kit (Omega Bio-Tek, Inc., Norcross, GA) following the
manufacturer’s instructions. The purified PCR product was sequenced at Eurofins Genomics,
Louisville, KY. The resulting DNA sequences were cleaned, aligned and analyzed using
Sequencher, BioEdit and ClustalW softwares.
Gene expression using qRT-PCR
The Palmer amaranth populations (15Cri-B and SS) were treated with trifloxysulfuron at
8 g ha-1 using the previously described procedure. Leaf tissues were collected 3 days before and
24 h after herbicide treatment. Genomic DNA from leaf tissues were subjected to PCR to amplify
the ALS gene following the previously described procedure. The purified PCR products were sent
for ALS gene sequencing. Plant samples which lacked any previously reported ALS point mutation
were used for the gene expression. The experiment was conducted in a completely randomized

105
design with herbicide treatment, plant biotype, and gene expression as the variables. Three
biological replicates from non-treated and treated samples of 15Cri-B and SS populations were
used. Total RNA was extracted from leaf samples using PureLink RNA Mini kit (Life
Technologies, Carlsbad, CA). cDNA synthesis was conducted using 5 µg total DNAse-treated
RNA using Reverse Transcription System first-strand cDNA synthesis kit (Promega). A 4-fold
serial dilution of cDNA samples (1:1, 1:5, 1:25, 1:125) was used to construct a standard curve.
Primers were designed to amplify ALS and six NTSR genes that were reported to be associated
with resistance to ALS-inhibiting herbicides (Table 1). DEAD-box RNA helicase and β-tubulin
were used as internal control for normalization of gene expression 45, 46
Quantitative real-time PCR (qRT-PCR) was conducted to determine changes in expression of
ALS and NTSR candidate genes relative to the reference genes. The qRT-PCR experiments were
conducted in a 12-µL volume containing 6.25 µL of SyberGreen Master Mix, 1 µL of 1:10 diluted
cDNA, and 0.5 µL of 10 µM primers (1:1 mix of forward and reverse primers). Amplification was
performed with three technical replicates using a Mastercycler EP Realplex machine (Eppendorf)
under the following conditions: 10 min at 94 oC, 40 cycles of 94 oC for 15 s and 60 oC for 1 min,
followed by a melt-curve analysis. Data were analyzed using Realplex 2.2 software. Fold-change
in gene expression as (2-ΔCt) was calculated by the comparative method,47 relative to the susceptible
samples, where ΔCT = [CT target gene –Ct mean of the two internal control genes]. Wilcoxon non-
parametric test (α=0.05) was used to determine statistical difference in gene expression between
resistant and susceptible biotypes.

106
RESULTS AND DISCUSSION
Cross-resistance to ALS-, EPSPS- and PPO-inhibitors
The 15Cri-B population was resistant to ALS inhibitors flumetsulam, imazethapyr,
primisulfuron, pyrithiobac, trifloxysulfuron with mortality ranging from 0 to 35% (Table 2).
Acetolactate synthase inhibitors such as pyrithiobac and trifloxysulfuron were widely used in
cotton fields in the past to control broadleaf weed species including Palmer amaranth species.48,49
Similarly, imazethapyr used to be highly effective in managing Amaranthus species in soybean
fields.50,51 These once-effective herbicides now fail to provide the desired Palmer amaranth
control as observed in numerous fields. Cross-resistance to multiple ALS-inhibiting herbicides is
common in Palmer amaranth.2 Previous studies indicated imidazolinone-resistant populations
from Arkansas and Georgia that were also resistant to chlorimuron, diclosulam, and
pyrithiobac.52,53 Herbicide dose-response experiments conducted on whole plants confirmed
resistance to ALS inhibitors (flumetsulam, imazethapyr, trifloxysulfuron), fomesafen, and
glyphosate. Based on GR50, its resistance level was 4.8-, 60.9-, and 4.4-fold with flumetsulam
(triazolopyrimidine), imazethapyr (imidazolinone), and trifloxysulfuron (sulfonylurea),
respectively, relative to the SS population. The level of resistance in 15Cri-B is lower than what
has been previously reported for other populations of ALS-resistant Palmer amaranth
populations.54,55 A population of Palmer amaranth from Kansas and Argentina had >2,800- and
288-fold resistance to imazethapyr, respectively.54, 57 Resistant population of Palmer amaranth
from Mississippi and Georgia had 8- and 303-fold resistance to pyrithiobac, respectively.55, 56
The difference in resistance levels is partly due to the sensitivity of the standard susceptible
population used in greenhouse bioassay or due to differences in the mechanism of resistance or

107
both.58 Even though the15Cri-B population had a lower resistance ratio than other populations,
some plants survived at 8X the normal dose of flumetsulam, imazethapyr, and trifloxysulfuron.
The 15Cri-B population exhibited resistance not only to ALS inhibitors but also to
fomesafen (PPO inhibitor) and glyphosate (EPSPS inhibitor) herbicides. Fomesafen and
glyphosate had 77 and 61% control, respectively, of this population whereas glufosinate killed
91% of the population (Table 2). The resistance level was 8.3-fold with fomesafen and 32-fold
with glyphosate, relative to the SS population (Table 3). The mechanism of resistance to
fomesafen and glyphosate in 15Cri-B population is target-site based, via amino acid substitution
(Arg128Gly mutation in the PPO gene) and amplified EPSPS copy number, respectively.10, 59
Populations of Palmer amaranth resistant to both glyphosate and ALS inhibitors are prevalent
and have been identified in 10 states, including Arkansas, Arizona, Delaware, Florida, Georgia,
Illinois, Maryland, Michigan, Mississippi, Tennessee, and South Carolina.9,10,55,56 Other case of
multiple resistance was also documented such as the Palmer amaranth population from Michigan
that was resistant to ALS, EPSPS, and PSII inhibitors.58 Several Palmer amaranth populations in
Arkansas are resistant to glyphosate, ALS-, and PPO-inhibiting herbicides posing serious
challenge to growers.10, 60 With Palmer amaranth resistant to multiple modes of action, limited
herbicide options remain for its control. The use of photosystem II and long chain fatty acid
inhibitors for pre-emergence and foliar application of glufosinate in glufosinate-resistant crops
can be adopted. Herbicide-resistant trait technologies (LibertyLink®, Xtend®, or Enlist®) are
available and can be used along with integrated weed management practices such as applying
herbicides with different modes of action, crop rotation and other cultural and mechanical
practices (e.g. cover crops, tillage, hand weeding).61

108
P450 inhibitor interaction with trifloxysulfuron
The involvement of P450 monooxygenase in metabolic resistance has often been detected
when specific P450 inhibitors enhance injury to resistant biotypes when applied in conjunction
with the herbicide.36, 62 The P450 inhibitors carbaryl, malathion, and piperonyl butoxide applied
alone caused little to no effect on the growth of Palmer amaranth. Therefore, these compounds,
by themselves, are not harmful to this weed. However, plants sprayed with malathion and
piperonyl butoxide prior to trifloxysulfuron treatment incurred higher phytoxicity compared to
plants treated with trifloxysulfuron alone (Table 4). Biomass reduction was 25% with
trifloxysulfuron alone but pretreatment with malathion or piperonyl butoxide increased Palmer
amaranth control to 49% and 66%, respectively. On the other hand, pretreatment with carbaryl
did not change the plant response compared to trifloxysulfuron alone. The increased efficacy of
trifloxysulfuron applied 45 min after malathion or piperonyl butoxide indicates P450-mediated
metabolism of the herbicide in resistant plants. Malathion is an excellent synergist for ALS-
inhibiting herbicides in ALS-resistant late watergrass, rigid ryegrass, and tall waterhemp
(Amaranthus tuberculatus).36, 62, 63 The synergistic interaction of malathion with sulfonylurea
herbicide is likely caused by competitive inhibition of cytochrome P450 degradation enzymes.64
The inhibition of herbicide activity by malathion occurs when atomic sulfur released from the
oxygenated organophosphate inhibits the P450 apoprotein.65 On the other hand, piperonyl
butoxide inhibits certain cytochrome P450 enzymes by causing steric hindrance created by its
long side-chain in the substrate access channel of the enzyme.38 Cytochrome P450 enzymes have
long been implicated in herbicide metabolism. These enzymes catalyze nonsynthetic
modifications, such as oxidation and hydroxylation.66 In fact, herbicide selectivity in weeds and
crops is attributed to the plant’s ability to metabolize the herbicide. Herbicide safeners protect

109
monocot crops from herbicide damage by inducing the production of P450 enzymes, glutathione
S-transferase, and glutathione peroxidase which can enhance the herbicide detoxification
processes.65 Hundreds of P450 genes exist in plants genomes, and each P450 participates in
various biochemical pathways to produce primary and secondary metabolites.67 The complexity
of cytochrome P450s increases the difficulty of elucidating the genetics of resistance, however,
the use of advanced molecular biology and genomics approaches are beginning to provide new
information on the identity and role of P450 genes in endowing metabolic resistance in plants.
Sequence analysis of acetolactate synthase gene
Previous researches have shown that amino acid substitution in the ALS gene is the most
common resistance mechanism in ALS-resistant Amaranthus species18, 23, 68 The ALS gene of
seven resistant plants from 15Cri-B contained nucleotide changes at multiple loci; however most
of the polymorphisms were silent mutations. A guanine to adenine change at position 653 was
detected in two resistant plants which resulted in Ser653Asn amino acid mutation (number
relative to Arabidopsis) (Fig. 1). The other five resistant plants did not have any of the known
resistance-conferring mutations to ALS herbicides. Variability in the ALS gene within a
population was also reported in ALS-resistant Palmer amaranth from Argentina in which five
allelic versions of the ALS gene were detected.57 Given the high genetic diversity of Palmer
amaranth, it is not surprising to have several genotypes of Palmer amaranth in a population. Our
results agree with the findings of other researchers, who also reported that Ser653Asn mutation
conferred high-level resistance to imidazolinone herbicides in tall waterhemp and downy brome
(Bromus tectorum).69,70 In greenhouse bioassays 15Cri-B had high level of resistance to
imazethapyr (60.9-fold), and low level of resistance to trifloxysulfuron (4.4-fold) and

110
flumetsulam (4.8-fold). The Ser653Asn mutation has been almost exclusively linked to
imidazolinone resistance, conferring low resistance to sulfonylurea in some cases,18 however,
15Cri-B was also cross-resistant to TPs (flumetsulam) and PTBs (pyrithiobac). NTSR
mechanisms possibly exist in these plants that would enable broad resistance to ALS herbicides
belonging to different chemical families. Furthermore, the presence of plants that carry the
Ser653Asn mutation and plants that do not harbor ALS mutation suggest the existence of both TS
and NTSR mechanisms in 15Cri-B population.
ALS and NTSR gene expression analysis
Target gene overexpression has been reported to confer resistance to herbicides.
Glyphosate-resistant Palmer amaranth populations contain multiple copies of the EPSPS gene
resulting in overproduction of the target enzyme EPSPS.71-73 In our experiment, ALS-resistant
and –susceptible Palmer amaranth biotypes had similar ALS transcript abundance (1-fold)
suggesting that the ALS gene is not overexpressed in resistant plants. The ALS gene expression
did not differ before and after trifloxysulfuron treatment in both biotypes. Stable expression of
the ALS gene has been documented; in fact, it has been used as reference in several gene
expression.72, 74 However, in shortawn foxtail (Alopecuros aequalis), the ALS copy number is
variable.75 The ALS copy number variation in shortawn foxtail is not likely an adaptation for
herbicide resistance since the additional ALS copies were deemed to be non-functional
pseudogenes. In the case of Palmer amaranth, the ALS gene was not differentially expressed
between ALS-resistant and –susceptible plants, which indicates that other mechanism(s), apart
from target gene overexpression, endow resistance to other ALS inhibitors.

111
The expression patterns of six genes [four cytochrome P450s (CYP72A, CYPA219A,
CYP81D, CYP81B), one glycosyltransferase (GT), and one glutathione-S-transferase (GSTF10)]
were examined before and after trifloxysulfuron treatment in resistant and susceptible plants.
CYP81B and GSTF10 were induced upon trifloxysulfuron application and were not
constitutively expressed in non-treated plants (Fig. 2). The expression of GSTF10 and CYP81B
in resistant plants ranged from 3- to 5-fold relative to the susceptible plant suggesting their
involvement in NTSR to trifloxysulfuron. Cytochrome P450 and glutathione-S-transferase have
been implicated in herbicide degradation; the former catalyzes herbicide oxidation and the latter
facilitates conjugation with the herbicide metabolite.11
The biochemical role of cytochrome P450-mediated herbicide metabolism have been
reported in herbicide-resistant rigid ryegrass and late watergrass populations.42, 62,76,77,79 For
example, high expression of CYP81B1, CYP81A21, and CYPA12 were associated with
resistance to SUs and TPs in Lolium and late watergrass populations.77,79,80 Several other
cytochrome P450 genes have been linked to ALS resistance in various weed populations such as
CYP72A254 (also known as CYPA72A219) in late watergrass (bispyribac),78 CYP81D in black-
grass (Alopecurus myosuroides) (mesosulfuron and iodosulfuron),81 CYP94A1, CYP94A2,
CYP71A4 and CYP734A6 in shortawn foxtail (mesosulfuron) (Table 5).41 In addition, many
CytP450s conferring resistance to ALS herbicides have been documented in several major crop
species, such as CYP81A6 and CYP72A31 in rice (Oryza sativa) and CYP81A9 in maize (Zea
mays) (Table 5).37,82,83 It was demonstrated that the wheat (Triticum aestivum) CYP71C6v1
expressed in yeast (Saccharomyces cerevisiae) is able to metabolize several sulfonylurea
herbicides such chlorsulfuron and triasulfuron through phenyl ring hydroxylation.84 The

112
CYP81B gene identified in Palmer amaranth (15Cri-B) may also confer resistance to multiple
herbicides, but this will require further investigation.
Glutathione S-transferases (GSTs) are ubiquitous in plants and have defined roles in
herbicide detoxification.85 Differences in glutathione availability and in the portfolio of GST
isoenzymes are associated with herbicide selectivity and herbicide resistance.86 For example,
tolerance of maize and giant foxtail (Setaria faberi) to atrazine is due to the high levels of GST
that facilitate the detoxification of the herbicide via glutathione conjugation.86, 87 Furthermore,
induction of glutathione and GSTs by herbicide 'safeners' is employed commercially to protect
crop plants from injury by certain herbicides.88 GSTs (phi and tau class) were involved Lolium
resistance to diclofop.89 Furthermore, GSTA and GST1 were also recently reported to participate
in non-target resistance or tolerance to ALS inhibitors in Lolium and maize, respectively.82,90 It
was once demonstrated that transgenic tobacco plants (Nicotiana tabacum) expressing maize
GST1 had enhanced detoxification of the herbicide alachlor.91 In ACCase-resistant black-grass,
GSTF1 catalyzed the conjugation of herbicide to glutathione as well as acted as peroxidase
protecting the cell from oxidative damage.92-94 Transgenic Arabidopsis (Arabidopsis thaliana)
expressing the black-grass GSTF1 gene (AmGSTF1) has improved tolerance to alachlor and
atrazine due to increased accumulation of protective flavonoids.94 Glutathione S-transferases
contribute herbicide resistance by detoxifying and transporting herbicide metabolites and/or
counteracting oxidative stress. It remains to be investigated whether the GSTF10 of Palmer
amaranth is involved in herbicide conjugation or protection from oxidative damage. An ortholog
of GSTF10 in Arabidopsis (AtFSTF10) possesses only transferase activity and not peroxidase
activity.95 Thus, Palmer amaranth GST10 is likely to be involved in detoxification of ALS-
inhibiting herbicides by herbicide conjugation with glutathione.

113
Our study showed ALS target-site mutation (Ser653Asn) conferring resistance to
trifloxysulfuron in ~29% (2/7) of 15Cri-B Palmer amaranth population. Previous reports
associated Ser653Asn mutation with high-level resistance to IMIs and low-level resistance to
SUs,18 however, the 15Cri-B population exhibited resistance not only to IMIs and SUs but to
PTBs and TPs. It is likely that resistant plants that carried the ALS Ser653Asn mutation may
harbor NTSR mechanisms, but this needs to be verified. The majority of the resistant plants did
not carry the Ser653Asn mutation but showed induction of NTSR (GSTF10 and CYP81B) genes,
which would endow resistance to multiple ALS herbicides. The occurrence of both TSR and
NTSR to ALS inhibitors in the same population was also reported in a Palmer amaranth from
Kansas.96 In that study, 30% of the chlorsulfuron-resistant plants harbored ALS Pro197Ser
mutation and the remaining 70% was deduced to have cytochrome P450-mediated metabolism
based on a malathion pretreatment assay.96 Other weed species displaying both TSR and NTSR
to ALS inhibitors include corn poppy,40 flixweed,39 and rigid ryegrass populations.39,40,97 The
evolution of TSR and NTSR in weed species predisposes resistance to multiple herbicide
families and modes of action. With Palmer amaranth being a prolific and obligate crossing
species, plants in a population can accumulate multiple resistance mechanisms which would
likely drive resistance evolution faster and is therefore detrimental to chemical weed
management.

114
REFERENCES
1. Webster, T. M.; Nichols, R. L., Changes in the prevalence of weed species in the major
agronomic crops of the southern United States: 1994/1995 to 2008/2009. Weed Sci. 2012,
60, 145-157.
2. Ward, S. M.; Webster, T. M.; Steckel, L. E., Palmer amaranth (Amaranthus palmeri): A
review. Weed Technol. 2013, 27, 12-27.
3. Bravo, W.; Leon, R. G.; Ferrell, J. A.; Mulvaney, M. J.; Wood, C. W., Differentiation of
life-history traits among Palmer amaranth populations (Amaranthus palmeri) and its
relation to cropping systems and glyphosate sensitivity. Weed Sci. 2017, 65, 339-349.
4. Fast, B. J.; Murdoch, S. W.; Farris, R. L.; Willis, J. B.; Murray, D. S., Critical timing of
Palmer amaranth (Amaranthus palmeri) removal in second-generation glyphosate-resistant
cotton. J. Cot. Sci.: 2009, 13, 32-36.
5. Bensch, C. N.; Horak, M. J.; Peterson, D., Interference of redroot pigweed (Amaranthus
retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A. rudis) in
soybean. Weed Sci. 2003, 51, 37-43.
6. Massinga, R. A.; Currie, R. S.; Horak, M. J.; Boyer, J., Interference of Palmer amaranth in
corn. Weed Sci. 2001, 49, 202-208.
7. Moore, J. W.; Murray, D. S.; Westerman, R. B., Palmer amaranth (Amaranthus palmeri)
effects on the harvest and yield of grain sorghum (Sorghum bicolor). Weed Technol. 2004,
18, 23-29.
8. Meyers, S. L.; Jennings, K. M.; Schultheis, J. R.; Monks, D. W., Interference of Palmer
amaranth (Amaranthus palmeri) in sweetpotato. Weed Sci. 2010, 58, 199-203.
9. Heap I. The international survey of herbicide resistant weeds. 2018.
http://www.weedscience.com. Accessed 22 January 2018.
10. Salas-Perez, R. A.; Burgos, N. R.; Rangani, G.; Singh, S.; Paulo Refatti, J.; Piveta, L.;
Tranel, P. J.; Mauromoustakos, A.; Scott, R. C., Frequency of Gly-210 deletion mutation
among protoporphyrinogen oxidase inhibitor–resistant Palmer amaranth (Amaranthus
palmeri) populations. Weed Sci. 2017, 65, 718-731.
11. Delye, C., Unravelling the genetic bases of non-target-site-based resistance (NTSR) to
herbicides: A major challenge for weed science in the forthcoming decade. Pest Manage.
Sci. 2013, 69, 176-187.
12. Ge, X.; d'Avignon, D. A.; Ackerman, J. J. H.; Collavo, A.; Sattin, M.; Ostrander, E. L.;
Hall, E. L.; Sammons, R. D.; Preston, C., Vacuolar glyphosate-sequestration correlates
with glyphosate resistance in ryegrass (Lolium spp.) from Australia, South America, And
Europe: a P-31 NMR investigation. J. Agri. Food Chem. 2012, 60, 1243-1250.
13. Brunharo, C. A. C. G.; Hanson, B. D., Vacuolar sequestration of paraquat is involved in
the resistance mechanism in Lolium perenne. spp. multiflorum. Fron. Plant Sci. 2017, 8,
1485.
14. Goggin, D. E.; Cawthray, G. R.; Powles, S. B., 2,4-D resistance in wild radish: reduced
herbicide translocation via inhibition of cellular transport. J. Exp. Bot. 2016, 67, 3223-35.

115
15. Yuan, J. S.; Tranel, P. J.; Stewart, C. N., Non-target-site herbicide resistance: A family
business. Trends Plant Sci. 2007, 12.
16. Gaeddert, J. W.; Peterson, D. E.; Michael, J. H., Control and cross-resistance of an
acetolactate synthase inhibitor-resistant Palmer amaranth (Amaranthus palmeri) biotype.
Weed Technol. 1997, 11, 132-137.
17. Mazur, B. J.; Falco, S. C., The development of herbicide resistant crops. Annu. Rev. Plant
Physiol. Plant Mol. Biol. 1989, 40, 441-470.
18. Tranel, P. J.; Wright, T. R., Resistance of weeds to ALS-inhibiting herbicides: What have
we learned? Weed Sci. 2002, 50, 700-712.
19. Cory, M. W.; Henry, P. W.; Westwood, J. H., A new mutation in plant ALS confers
resistance to five classes of ALS-inhibiting herbicides. Weed Sci. 2007, 55, 83-90.
20. Brosnan, J. T.; Vargas, J. J.; Breeden, G. K.; Grier, L.; Aponte, R. A.; Tresch, S.; Laforest,
M., A new amino acid substitution (Ala-205-Phe) in acetolactate synthase (ALS) confers
broad spectrum resistance to ALS-inhibiting herbicides. Planta 2016, 243, 149-59.
21. Bernasconi, P.; Woodworth, A. R.; Rosen, B. A.; Subramanian, M. V.; Siehl, D. L., A
naturally occurring point mutation confers broad range tolerance to herbicides that target
acetolactate synthase. J. Biol. Chem. 1996, 271, 13925.
22. Devine, M. D.; Maries, M. A. S.; Hall, L. M., Inhibition of acetolactate synthase in
susceptible and resistant biotypes of Stellaria media. J. Pestic. Sci. 1991, 31, 273-280.
23. Singh, S. Characterization of glyphosate-resistant Amaranthus palmeri (Palmer amaranth)
tolerance to ALS- and HPPD-Inhibiting Herbicides. University of Arkansas, 2017.
24. Veldhuis, L. J.; Hall, L. M.; O'Donovan, J. T.; Dyer, W.; Hall, J. C., Metabolism-based
resistance of a wild mustard (Sinapis arvensis L.) biotype to ethametsulfuron-methyl. J.
Agric. Food Chem. 2000, 48, 2986-2990.
25. Cruz-Hipolito, H.; Rosario, J.; Ioli, G.; Osuna, M. D.; Smeda, R. J.; Gonzalez-Torralva,
F.; De Prado, R., resistance mechanism to tribenuron-methyl in white mustard (Sinapis
alba) from southern Spain. Weed Sci. 2013, 61, 341-347.
26. Poston, D. H.; Wu, J.; Hatzios, K. K.; Wilson, H. P., Enhanced sensitivity to cloransulam-
methyl in imidazolinone-resistant smooth pigweed. Weed Sci. 2001, 49, 711-716.
27. White, A. D.; Owen, M. D. K.; Hartzler, R. G.; Cardina, J., Common sunflower resistance
to acetolactate synthase–inhibiting herbicides. Weed Sci. 2002, 50, 432-437.
28. Riar, D. S.; Norsworthy, J. K.; Srivastava, V.; Nandula, V.; Bond, J. A.; Scott, R. C.,
Physiological and molecular basis of acetolactate synthase-inhibiting herbicide resistance
in barnyardgrass (Echinochloa crus-galli). J. Agric. Food Chem. 2013, 61, 278-289.
29. Délye, C.; Jasieniuk, M.; Corre, V. L., Deciphering the evolution of herbicide resistance in
weeds. Trends Genet. 2013, 29.
30. Yuan, J. S.; Tranel, P. J.; Stewart, C. N., Non-target-site herbicide resistance: A family
business. Trends Plant Sci. 2007, 12 (1), 6-13.

116
31. Zhao, B.; Fu, D.; Yu, Y.; Huang, C.; Yan, K.; Li, P.; Shafi, J.; Zhu, H.; Wei, S.; Ji, M.,
Non-target-site resistance to ALS-inhibiting herbicides in a Sagittaria trifolia L.
population. Pestic. Biochem. Physiol. 2017, 140, 79-84.
32. Beckie, H. J.; Warwick, S. I.; Sauder, C. A., Basis for herbicide resistance in canadian
populations of wild oat (Avena fatua). Weed Sci. 2012, 60, 10-18.
33. Yu, Q.; Han, H.; Cawthray, G. R.; Wang, S. F.; Powles, S. B., Enhanced rates of herbicide
metabolism in low herbicide-dose selected resistant Lolium rigidum. Plant Cel. Environ.
2013, 36, 818-827.
34. Veldhuis, L. J.; Hall, L. M.; O'Donovan, J. T.; Dyer, W.; Hall, J. C., Metabolism-based
resistance of a wild mustard (Sinapis arvensis L.) biotype to ethametsulfuron-methyl. J.
Agric. Food Chem. 2000, 48, 2986-90.
35. Yun, M. S.; Yogo, Y.; Miura, R.; Yamasue, Y.; Fischer, A. J., Cytochrome P-450
monooxygenase activity in herbicide-resistant and -susceptible late watergrass
(Echinochloa phyllopogon). Pestic. Biochem. Physiol. 2005, 83, 107-114.
36. Yasuor, H.; Osuna, M. D.; Ortiz, A.; Saldain, N. E.; Eckert, J. W.; Fischer, A. J.,
Mechanism of resistance to penoxsulam in late watergrass [Echinochloa phyllopogon
(Stapf) Koss.]. J. Agric. Food Chem. 2009, 57, 3653-60.
37. Pan, G.; Zhang, X.; Liu, K.; Zhang, J.; Wu, X.; Zhu, J.; Tu, J., Map based cloning of a
novel rice cytochrome P450 gene CYP81A6 that confers resistance to two different
classes of herbicides. Plant Mol. Biol. 2006, 61, 933-943,
38. Didierjean, L.; Gondet, L.; Perkins, R.; Lau, S. M. C.; Schaller, H.; O’Keefe, D. P.;
Werck-Reichhart, D., Engineering herbicide metabolism in tobacco and Arabidopsis with
CYP76B1, a cytochrome P450 enzyme from Jerusalem artichoke. Plant Physiol. 2002,
130, 179-189.
39. Yang, Q.; Deng, W.; Li, X.; Yu, Q.; Bai, L.; Zheng, M., Target-site and non-target-site
based resistance to the herbicide tribenuron-methyl in flixweed (Descurainia sophia L.).
BMC Genomics 2016, 17, 1-13.
40. Rey-Caballero, J.; Menéndez, J.; Osuna, M. D.; Salas, M.; Torra, J., Target-site and non-
target-site resistance mechanisms to ALS inhibiting herbicides in Papaver rhoeas. Pestic.
Biochem. Physiol. 2017, 138, 57-65.
41. Zhao, N.; Li, W.; Bai, S.; Guo, W.; Yuan, G.; Wang, F.; Liu, W.; Wang, J., Transcriptome
profiling to identify genes involved in mesosulfuron-methyl resistance in Alopecurus
aequalis. Fron. Plant Sci. 2017, 8, 1391.
42. Gaines, T. A.; Lorentz, L.; Figge, A.; Herrmann, J.; Maiwald, F.; Ott, M. C.; Han, H. P.;
Busi, R.; Yu, Q.; Powles, S. B.; Beffa, R., RNA-Seq transcriptome analysis to identify
genes involved in metabolism-based diclofop resistance in Lolium rigidum. Plant J. 2014,
78, 865-876.
43. Salas, R. A.; Saski, C.;Noorai, R. E.; Lawton-Rauh, A.; Srivastava, S.; Nichols, R. L.;
Burgos, N. R., RNA-Seq analysis of Amaranthus palmeri transcriptomic response to
glufosinate. Proceedings of the 57th Weed Science Society of America Annual Meeting,
Tucson, Arizona, USA, 2017.

117
44. Salas, R. A.; Burgos, N. R.; Tranel, P. J.; Singh, S.; Glasgow, L.; Scott, R. C.; Nichols, R.
L., Resistance to PPO-inhibiting herbicide in Palmer amaranth from Arkansas. Pest
Manage. Sci. 2016, 72, 864-869.
45. Feng, L.; Yu, Q.; Li, X.; Ning, X.; Wang, J.; Zou, J.; Zhang, L.; Wang, S.; Hu, J.; Hu, X.;
Bao, Z., Identification of reference genes for qRT-PCR Analysis in yesso scallop
Patinopecten yessoensis. PLoS One 2013, 8, e75609.
46. Nakka, S.; Godar, A. S.; Wani, P. S.; Thompson, C. R.; Peterson, D. E.; Roelofs, J.;
Jugulam, M., Physiological and molecular characterization of hydroxyphenylpyruvate
dioxygenase (HPPD)-inhibitor resistance in Palmer amaranth (Amaranthus palmeri
S.Wats.). Fron. Plant Sci. 2017, 8, 555.
47. Schmittgen, T. D.; Livak, K. J., Analyzing real-time PCR data by the comparative CT
method. Nat. Protoc. 2008, 3.
48. Porterfield, D.; Wilcut, J. W.; Askew, S. D., Weed management with CGA-362622,
fluometuron, and prometryn in cotton. Weed Sci. 2002, 50, 642-647.
49. Dotray, P. A.; Keeling, J. W.; Henniger, C. G.; Abernathy, J. R., Palmer Amaranth
(Amaranthus palmeri) and devil's-claw (Proboscidea louisianica) control in cotton
(Gossypium hirsutum) with pyrithiobac. Weed Technol. 1996, 10, 7-12.
50. Mayo, C. M.; Horak, M. J.; Peterson, D. E.; Boyer, J. E., Differential control of 4
Amaranthus species by 6 postemergence herbicides in soybean (Glycine max). Weed
Technol. 1995, 9, 141-147.
51. Cantwell, J. R.; Liebl, R. A.; Slife, F. W., Imazethapyr for weed control in soybean
(Glycine max). Weed Technol. 1989, 3, 596-601.
52. Burgos, N. R.; Kuk, Y.-I.; Talbert, R. E., Amaranthus palmeri resistance and differential
tolerance of Amaranthus palmeri and Amaranthus hybridus to ALS-inhibitor herbicides.
Pest Manage. Sci. 2001, 57, 449-457.
53. Wise, A. M.; Grey, T. L.; Prostko, E. P.; Vencill, W. K.; Webster, T. M., Establishing the
geographical distribution and level of acetolactate synthase resistance of Palmer amaranth
(Amaranthus palmeri) accessions in Georgia. Weed Technol. 2009, 23, 214-220.
54. Sprague, C. L.; Stoller, E. W.; Wax, L. M.; Horak, M. J., Palmer amaranth (Amaranthus
palmeri) and common waterhemp (Amaranthus rudis) resistance to selected ALS-
inhibiting herbicides. Weed Sci. 1997, 45, 192-197.
55. Nandula, V. K.; Reddy, K. N.; Koger, C. H.; Poston, D. H.; Rimando, A. M.; Duke, S. O.;
Bond, J. A.; Ribeiro, D. N., Multiple resistance to glyphosate and pyrithiobac in Palmer
amaranth (Amaranthus palmeri) from Mississippi and response to flumiclorac. Weed Sci.
2012, 60, 179-188.
56. Sosnoskie, L. M.; Kichler, J. M.; Wallace, R. D.; Culpepper, A. S., Multiple resistance in
Palmer amaranth to glyphosate and pyrithiobac confirmed in Georgia. Weed Sci. 2011, 59,
321-325.
57. Larran, A. S.; Palmieri, V. E.; Perotti, V. E.; Lieber, L.; Tuesca, D.; Permingeat, H. R.,
Target-site resistance to acetolactate synthase (ALS)-inhibiting herbicides in Amaranthus
palmeri from Argentina. Pest Manage. Sci. 2017, 73, 2578-2584.

118
58. Kohrt, J. R.; Sprague, C. L.; Nadakuduti, S. S.; Douches, D., Confirmation of a three-way
(glyphosate, ALS, and atrazine) herbicide-resistant population of Palmer amaranth
(Amaranthus palmeri) in Michigan. Weed Sci. 2017, 65, 327-338.
59. Salas, R. A.; Oliveira, C.; Refatti, J. P.; Piveta, L.; Scott, R.; Burgos, N. R. In
Investigating multiple resistance mechanisms of Amaranthus palmeri populations from
Arkansas, 57th Annual Meeting of the Weed Science Society of America, Tucson,
Arizona, Weed Science Society of America: Tucson, Arizona, 2017.
60. Schwartz-Lazaro, L. M.; Norsworthy, J. K.; Scott, R. C.; Barber, L. T., Resistance of two
Arkansas Palmer amaranth populations to multiple herbicide sites of action. Crop Prot.
2017, 96, 158-163.
61. Norsworthy, J. K.; Ward, S. M.; Shaw, D. R.; Llewellyn, R. S.; Nichols, R. L.; Webster,
T. M.; Bradley, K. W.; Frisvold, G.; Powles, S. B.; Burgos, N. R.; Witt, W. W.; Barrett,
M., Reducing the risks of herbicide resistance: best management practices and
recommendations. Weed Sci. 2012, 60, 31-62.
62. Christopher, J. T.; Preston, C.; Powles, S. B., Malathion antagonizes metabolism-based
chlorsulfuron resistance in Lolium rigidum. Pestic. Biochem. Physiol. 1994, 49, 172-182.
63. Guo, J. Q.; Riggins, C. W.; Hausman, N. E.; Hager, A. G.; Riechers, D. E.; Davis, A. S.;
Tranel, P. J., Nontarget-site resistance to ALS inhibitors in waterhemp (Amaranthus
tuberculatus). Weed Sci. 2015, 63, 399-407.
64. Tardif, F. J.; Powles, S. B., Effect of malathion on resistance to soil-applied herbicides in
a population of rigid ryegrass (Lolium rigidum). Weed Sci. 1999, 47, 258-261.
65. Werck-Reichhart, D.; Hehn, A.; Didierjean, L., Cytochromes P450 for engineering
herbicide tolerance. Trends Plant Sci. 2000, 5, 116-123.
66. Ohkawa, H.; Tsujii, H.; Ohkawa, Y., The use of cytochrome P450 genes to introduce
herbicide tolerance in crops: A review. Pestic. Sci. 1999, 55, 867-874.
67. Mizutani, M.; Ohta, D., Diversification of P450 Genes during land plant evolution. Annual
Rev. Plant Biol. 2010, 61, 291-315.
68. Whaley, C. M.; Wilson, H. P.; Westwood, J. H., A aew mutation in plant als confers
resistance to five classes of ALS-inhibiting herbicides. Weed Sci. 2007, 55, 83-90.
69. Patzoldt, W. L.; Tranel, P. J., Multiple ALS mutations confer herbicide resistance in
waterhemp (Amaranthus tuberculatus). Weed Sci. 2007, 55, 421-428.
70. Kumar, V.; Jha, P., First report of Ser653Asn mutation endowing high-level resistance to
imazamox in downy brome (Bromus tectorum L.). Pest Manag. Sci. 2017, 73, 2585-2591.
71. Chandi, A.; York, A. C.; Jordan, D. L.; Beam, J. B., Resistance to acetolactate synthase
and acetyl co-a carboxylase inhibitors in North Carolina Italian ryegrass (Lolium perenne).
Weed Technol. 2011, 25, 659-666.
72. Gaines, T. A.; Zhang, W.; Wang, D.; Bukun, B.; Chisholm, S. T.; Shaner, D. L.; Nissen,
S. J.; Patzoldt, W. L.; Tranel, P. J.; Culpepper, A. S.; Grey, T. L.; Webster, T. M.; Vencill,
W. K.; Sammons, R. D.; Jiang, J.; Preston, C.; Leach, J. E.; Westra, P., Gene amplification
confers glyphosate resistance in Amaranthus palmeri. PNAS 2010, 107, 1029-1034.

119
73. Mohseni-Moghadam, M.; Schroeder, J.; Ashigh, J., Mechanism of resistance and
inheritance in glyphosate resistant Palmer amaranth (Amaranthus palmeri) populations
from New Mexico, USA. Weed Sci. 2013, 61, 517-525.
74. Chen, J.; Huang, Z.; Huang, H.; Wei, S.; Liu, Y.; Jiang, C.; Zhang, J.; Zhang, C., Selection
of relatively exact reference genes for gene expression studies in goosegrass (Eleusine
indica) under herbicide stress. Sci. Rep. 2017, 7, 46494.
75. Iwakami, S.; Shimono, Y.; Manabe, Y.; Endo, M.; Shibaike, H.; Uchino, A.; Tominaga,
T., Copy number variation in acetolactate synthase genes of thifensulfuron-methyl
resistant Alopecurus aequalis (shortawn foxtail) accessions in Japan. Fron. Plant Sci.
2017, 8, 254.
76. Duhoux, A.; Delye, C., Reference genes to study herbicide stress response in Lolium sp.:
up-regulation of P450 genes in plants resistant to acetolactate-synthase inhibitors. PLoS
One 2013, 8, e63576.
77. Duhoux, A.; Carrere, S.; Gouzy, J.; Bonin, L.; Delye, C., RNA-Seq analysis of rye-grass
transcriptomic response to an herbicide inhibiting acetolactate-synthase identifies
transcripts linked to non-target-site-based resistance. Plant Mol Biol 2015, 87, 473-87.
78. Iwakami, S.; Uchino, A.; Kataoka, Y.; Shibaike, H.; Watanabe, H.; Inamura, T.,
Cytochrome P450 genes induced by bispyribac-sodium treatment in a multiple-herbicide-
resistant biotype of Echinochloa phyllopogon. Pest Manag. Sci. 2014, 70, 549-58.
79. Duhoux, A.; Carrere, S.; Delye, C., Transcriptional markers enable identification of rye-
grass (Lolium sp.) plants with non-target -site-based resistance to herbicides inhibiting
acetolactate-synthase. Plant Sci. 2017, 257, 22-36.
80. Iwakami, S.; Endo, M.; Saika, H.; Okuno, J.; Nakamura, N.; Yokoyama, M., Cytochrome
P450 CYP81A12 and CYP81A21 are associated with resistance to two acetolactate
synthase inhibitors in Echinochloa phyllopogon. Plant Physiol. 2014, 165, 618-629.
81. Gardin, J. A. C.; Gouzy, J.; Carrère, S.; Délye, C., ALOMYbase, a resource to investigate
non-target-site-based resistance to herbicides inhibiting acetolactate-synthase (ALS) in the
major grass weed Alopecurus myosuroides (black-grass). BMC Genomics 2015, 16, 590.
82. Liu, X. M.; Xu, X.; Li, B. H.; Wang, X. Q.; Wang, G. Q.; Li, M. R., RNA-Seq
transcriptome analysis of maize inbred carrying nicosulfuron-tolerant and nicosulfuron-
susceptible alleles. Int. J. Mol. Sci. 2015, 16, 5975-5989.
83. Saika, H.; Horita, J.; Taguchi-Shiobara, F.; Nonaka, S.; Nishizawa-Yokoi, A.; Iwakami,
S.; Hori, K.; Matsumoto, T.; Tanaka, T.; Itoh, T.; Yano, M.; Kaku, K.; Shimizu, T.; Toki,
S., A novel rice cytochrome P450 gene, CYP72A31, confers tolerance to acetolactate
synthase-inhibiting herbicides in rice and arabidopsis. Plant Physiol. 2014, 166, 1232-
1240.
84. Xiang, W.; Wang, X.; Ren, T.; Ci, S., Expression of a wheat cytochrome P450
monooxygenase cDNA in yeast catalyzes the metabolism of sulfonylurea herbicides.
Pestic. Biochem. Physiol. 2006, 85, 402-406.
85. Wagner, U.; Edwards, R.; Dixon, D. P.; Mauch, F., Probing the diversity of the
Arabidopsis glutathione S-transferase gene family. Plant Mol. Biol. 2002, 49, 515-32.

120
86. Hatton, P. J.; Dixon, D.; Cole, D. J.; Edwards, R., Glutathione transferase activities and
herbicide selectivity in maize and associated weed species. Pestic. Sci. 1996, 46, 267-275.
87. Hatton, P. J.; Cummins, I.; Cole, D. J.; Edwards, R., Glutathione transferases involved in
herbicide detoxification in the leaves of Setaria faberi (giant foxtail). Physiol. Plant. 1999,
105, 9-16.
88. Farago, S.; Brunold, C.; Kreuz, K., Herbicide safeners and glutathione metabolism.
Physiol. Plant. 1994, 91, 537-542.
89. Gaines, T. A.; Lorentz, L.; Figge, A.; Herrmann, J.; Maiwald, F.; Ott, M. C., RNA-Seq
transcriptome analysis to identify genes involved in metabolism-based diclofop resistance
in Lolium rigidum. Plant J. 2014, 78, 865-876.
90. Duhoux, A.; Carrère, S.; Gouzy, J.; Bonin, L.; Délye, C., RNA-Seq analsis of rye-grass
transcriptomic response to an herbicide inhibiting acetolactate-synthase identifies
transcripts linked to non-target-site-based resistance. Plant Mol. Biol. 2015, 87, 473-487.
91. Karavangeli, M.; Labrou, N. E.; Clonis, Y. D.; Tsaftaris, A., Development of transgenic
tobacco plants overexpressing maize glutathione S-transferase I for chloroacetanilide
herbicides phytoremediation. Biomol. Eng. 2005, 22, 121-8.
92. Cummins, I.; Cole, D. J.; Edwards, R., A role for glutathione transferases functioning as
glutathione peroxidases in resistance to multiple herbicides in black-grass. Plant J. 1999,
18, 285-292.
93. Cummins, I.; Bryant, D. N.; Edwards, R., Safener responsiveness and multiple herbicide
resistance in the weed black-grass (Alopecurus myosuroides). Plant Biotechnol. J. 2009, 7,
807-820.
94. Cummins, I.; Wortley, D. J.; Sabbadin, F.; He, Z. S.; Coxon, C. R.; Straker, H. E.; Sellars,
J. D.; Knight, K.; Edwards, L.; Hughes, D.; Kaundun, S. S.; Hutchings, S. J.; Steel, P. G.;
Edwards, R., Key role for a glutathione transferase in multiple-herbicide resistance in
grass weeds. PNAS 2013, 110, 5812-5817.
95. Sappl, P. G.; Carroll, A. J.; Clifton, R.; Lister, R.; Whelan, J.; Harvey Millar, A.; Singh,
K. B., The Arabidopsis glutathione transferase gene family displays complex stress
regulation and co-silencing multiple genes results in altered metabolic sensitivity to
oxidative stress. Plant J. 2009, 58, 53-68.
96. Nakka, S.; Thompson, C. R.; Peterson, D. E.; Jugulam, M., Target site-based and non-
target site based resistance to ALS inhibitors in Palmer amaranth (Amaranthus palmeri).
Weed Sci. 2017, 65, 681-689.
97. Christopher, J. T., Powles, S. B., Holtum, J. A., Resistance to acetolactate synthase-
inhibiing herbicides in annual ryegrass (Lolium rigidum) involves at least two
mechanisms. Plant Physiol. 1992, 100, 1909-1913.

121
Table 1. Primers used in the gene expression analysis by qRT-PCR.a
Gene Primer Sequence Name Reference
Cytochrome P450
CYP72A219
GGACAAGAAACTACATCGACA Cyp72A219_F3 78
TGTTTGCAGGACTTCTTCC Cyp72A219_R3
Cytochrome P450
CYP81D
GTTTATACCATTCGGGTCAGG Cyp81D_F4 80
TTCTCCCATTCTTTCCCATTC Cyp81D_R4
Cytochrome P450
CYP81B
AACGACTTCTAGCACAATGG Cyp81B_F2 76
GTCCTACGTGCTCTAAGATTTC Cyp81B_R2
UDP-
glycosyltransferase
(GT)
GAATTGATGGAGGGAAGGAATG GT_F3 39,76
CCTTCAACTTTAGTAGCCTCTTG GT_R3
Glutathione-S-
transferase
(GSTF10)
GCTATTTAGCTGGAGACGAC GST_F1 76
CCCACCATCTACTTACATTCTC GST_R1
Cytochrome P450
CYP72A215
TGCACAATCTCTCGAATGG Cyp72_F2 76,83
GGATCGTGGTGGGTATAAAG Cyp72_R2
ALS GCTGCTGAAGGCTACGCT ALS_F2 72
GCGGGACTGAGTCAAGAAGTG ALS_R2
DEAD-box RNA
helicase
TTGGAACTGTCAGAGCAACC A36_F 45
GAACCCACTTCCACCAAAAC A36_R
β-tubulin ATTCCCTCGGCTTCATTTC BT_F1 46
TCCCACATTTGCTGAGTTAG BT_R2
a Herbicide-treated and non-treated Palmer amaranth plants (15Cri-B and SS populations) with
three replications per treatment were used for the qPCR assay. The herbicide-treated plants were
harvested 24 h after application of trifloxysulfuron at 8 g ha-1.

122
Table 2. Response of 15Cri-B Palmer amaranth population to the normal dose of ALS
(flumetsulam, imazethapyr, primisulfuron, pyrithiobac, trifloxysulfuron), fomesafen, glyphosate
and glufosinate herbicidesa.
Mode of Action Herbicide Mortality
(%)
Acetolactate synthase (ALS) inhibitor
Flumetsulam 35
Imazethapyr 21
Primisulfuron 4
Pyrithiobac 0
Trifloxysulfuron 3
Protoporphyrinogen oxidase (PPO) inhibitor Fomesafen 77
Enolpyruvyl shikimate-3-phosphate synthase
(EPSPS) inhibitor
Glyphosate 61
Glutamine synthetase (GS) inhibitor Glufosinate 91
a Flumetsulam (7 g ai ha-1) , imazethapyr (70 g ae ha-1), primisulfuron (40 g ai ha-1), pyrithiobac
(73 g ai ha-1), trifloxysulfuron (8 g ai ha-1), fomesafen (263 g ai ha-1), glyphosate (840 g ae ha-1),
and glufosinate (549 g ai ha-1) were applied to 7.5-cm tall seedlings. Each herbicide was applied
to 200 seedlings.

123
Table 3. Level of resistance to ALS, PPO, and EPSPS inhibitors in 15Cri-B Palmer amaranth
population.
Mode of Action Herbicide GR50a
(g ai ha-1)
R/Sb
SSc 15Cri-B
Acetolactate synthase
(ALS) inhibitor
Flumetsulam 1.68
(1.04-2.32)d
8.16
(5.24-11.08)
4.8
Imazethapyr 6.42
(4.45-8.39)
67.35
(48.27-86.43)
60.9
Trifloxysulfuron 0.85
(0.63-1.07)
3.8
(2.75-4.87)
4.4
Protoporphyrinogen
oxidase (PPO)
inhibitor
Fomesafen 15.10
(10-21)
125.26
(85-164)
8.3
Enolpyruvyl
shikimate-3-
phosphate synthase
(EPSPS) inhibitor
Glyphosate 11.9
(9.0-14.7)
385
(287-410)
32
a Herbicide dose required to cause 50% biomass reduction. b Resistance level (R/S) calculated using the GR50 of the resistant population relative to the
susceptible population. c SS = susceptible standard population d Values inside the parenthesis denote 95% confidence intervals.

124
Table 4. Effect of pretreatment with P450 inhibitorsa on the phytotoxicity of trifloxysulfuron (8 g
ha-1) on 15Cri-B Palmer amaranth population.
P450 inhibitor Biomass reduction relative to the non-
treated control (%)
No herbicide With
trifloxysulfuron
Carbaryl 0 22
Malathion 5 49
Piperonyl butoxide 0 66
No P450 inhibitor 0 25
LSD0.05
Main effect of P450 inhibitor 10
Main effect of herbicide 14
P450 inhibitor x herbicide interaction 16
aP450 inhibitors were applied 45 min before trifloxysulfuron treatment.

125
Table 5. Cytochrome P450 genes involved in NTSR to ALS inhibitors in various weed and crop
species.
Cytochrome P450 gene
Species Herbicide Reference
CYP71A Alopecurus myosuroides iodosulfuron-methyl,
mesosulfuron-methyl
81
CYP71A4 Alopecuros aequalis mesosulfuron-methyl 41
CYP734A6 Alopecuros aequalis mesosulfuron-methyl 41
CYP72A254 Echinochloa phyllopogon bispyribac-sodium 78
CYP71B Alopecurus myosuroides iodosulfuron-methyl,
mesosulfuron-methyl
81
CYP71A31 Oryza sativa bispyribac-sodium 83
CYP71C6v1 Triticum aestivum chlorsulfuron, triasulfuron 84
CYP81A6 Oryza sativa bensulfuron-methyl 37
CYP81A9 Zea mays nicosulfuron 82
CYP81A12 Echinochloa phyllopogon bensulfuron-methyl, penoxsulam 80
CYP81A21 Echinochloa phyllopogon bensulfuron-methyl, penoxsulam 80
CYP81B1 Lolium pyroxsulam, iodosulfuron-methyl,
mesosulfuron-methyl
79
CYP81D Alopecurus myosuroides iodosulfuron-methyl,
mesosulfuron-methyl
81
CYP94A1 Alopecuros aequalis mesosulfuron-methyl 41
CYP94A2 Alopecuros aequalis mesosulfuron-methyl 41

126
Figure 1. Amino acid sequence alignment of acetolactate synthase (ALS) in sensitive (SS) and
resistant (15Cri-B) Palmer amaranth plant samples. Two resistant plants harbored Ser653Asn
(AGC to AAC) mutation.

127
Figure 2. Gene expression analysis of ALS and NTSR genes in trifloxysulfuron-resistant (15Cri-
B) and –susceptible (SS) Palmer amaranth biotypes. Fold-change (2-ΔCt) in gene expression was
calculated relative to the susceptible samples, where ΔCt = [Ct target-gene - Ct mean of internal
control genes]. Means and standard errors from three biological replicates are shown. Wilcoxon
non-parametric test was used to compare differential gene expression between ALS-resistant and
susceptible plants. An asterisk denotes fold change significance at P<0.05.

128
CHAPTER V
RESISTANCE TO PPO-INHIBITING HERBICIDE IN PALMER AMARANTH FROM
ARKANSAS
© 2016 Pest Management Science published by JohnWiley & Sons Ltd on behalf of Society of
Chemical Industry.

129
Abstract
BACKGROUND: The widespread occurrence of ALS inhibitor- and glyphosate-resistant
Amaranthus palmeri has led to increasing use of protoporphyrinogen oxidase (PPO)-inhibiting
herbicides in cotton and soybean. Studies were conducted to confirm resistance to fomesafen (a
PPO inhibitor), determine the resistance frequency, examine the resistance profile to other foliar-
applied herbicides, and investigate the resistance mechanism of resistant plants in a population
collected in 2011 (AR11-LAW B), and its progenies from two cycles of fomesafen selection (C1
and C2).
RESULTS: The frequency of fomesafen-resistant plants increased from 5% in the original
AR11-LAW-B to 17% in the C2 population. The amounts of fomesafen that caused 50% growth
reduction were 6-, 13-, and 21-fold greater in AR11-LAW-B, C1, and C2 populations,
respectively, than the sensitive ecotype. The AR11-LAW-B population was sensitive to atrazine,
dicamba, glufosinate, glyphosate, and mesotrione but resistant to ALS-inhibiting herbicides
pyrithiobac and trifloxysulfuron. Fomesafen survivors from C1 and C2 populations tested
positive for the PPO glycine 210 deletion previously reported in waterhemp (Amaranthus
tuberculatus).
CONCLUSION: These studies confirmed that Palmer amaranth in Arkansas has evolved
resistance to foliar-applied PPO-inhibiting herbicide.

130
INTRODUCTION
Palmer amaranth (Amaranthus palmeri S. Watson) is one of the most common,
troublesome, and economically damaging agronomic weeds throughout the southern United
States1. This weed continues to emerge throughout the summer making control critical from crop
emergence to harvest. The competitive ability of Palmer amaranth is attributed to its fast growth
rate2, high fecundity3, good light interception, and high water use efficiency4. With estimates of
over 600,000 seeds per plant, it can replenish the seedbank3 in one generation. Because it is
highly competitive with crops, it can reduce crop yield. Palmer amaranth densities of 8 and 9
plants m-2 can reduce soybean yield by 78%5 and corn grain yield by 91% 6, respectively. Fast et
al. (2009)7 reported that Palmer amaranth interference for 63 d after crop emergence caused 77%
cotton yield loss. Apart from reducing yield, more severely in stripper cotton, high infestation of
Palmer amaranth interferes with cotton harvest and can increase harvest time by two- to four-
fold8.
Palmer amaranth control has become a challenge because of its high propensity to evolve
herbicide resistance, resulting in reduced herbicide options in infested crops such as cotton and
soybean. To date, Palmer amaranth has been confirmed resistant to herbicides spanning five
modes of action: acetolactate synthase (ALS) inhibitors, carotenoid biosynthesis (4-
hydroxyphenylpyruvate dioxygenase) inhibitors, enolpyruvyl shikimate-3-phosphate synthase
inhibitor (glyphosate), mitosis inhibitors (dinitroanilines), and photosystem II inhibitors
(triazines)9. Being dioecious, Palmer amaranth is an obligate cross-pollinated species, allowing
herbicide resistance to spread rapidly10. Sosnoskie et al. (2012)11 reported that glyphosate
resistance trait was transferred across a distance of at least 300 m through pollen flow.

131
The widespread occurrence of resistance to ALS inhibitors and glyphosate in Palmer
amaranth has led to increasing use of protoporphyrinogen oxidase (PPO)-inhibitor herbicides
such as fomesafen, flumioxazin, saflufenacil, and sulfentrazone. Advantageous characteristics of
PPO inhibitors include broad herbicidal spectrum as these are active against many
monocotyledon and dicotyledon weeds, have low mammalian toxicity, low effective rates, rapid
onset of action, and long residual activity on some herbicides in this group12. The PPO enzyme
catalyzes the conversion of protoporphyrinogen IX (protogen IX) to protoporphyrin IX (proto
IX) which is the last common step in the biosynthesis of heme and chlorophyll13. In plants, two
PPO isoforms are encoded by two different PPO nuclear genes, PPX1 and PPX2. These isoforms
share little sequence identity (25%) and differ in their subcellular targeting. PPO1 and PPO2 are
localized in plastids and mitochondria, respectively; however, at least some PPO2 isoforms are
dual-targeted to both organelles14-16. Inhibition of PPO by herbicides results in the generation of
singlet oxygen species that attack lipid and protein membranes leading to plant death17.
The PPO enzyme is inhibited by several herbicide chemical classes (e.g. diphenyl ethers,
heterocyclic phenyl ethers, oxadiazoles, phenyl imides, triazolinones and pyrazoles). Herbicides
that inhibit PPO have been in the market since the 1960s and were primarily used in soybean.
One of these is fomesafen, a diphenyl ether herbicide, which can be applied preplant,
preemergence or postemergence for control or suppression of broadleaf weeds, grasses and
sedges in soybean. Resistance to PPO herbicides has been slow to evolve (about four decades
from first commercialization), and to date, has been confirmed only in seven weed species9. The
first weed to evolve resistance to PPO herbicides was waterhemp (Amaranthus tuberculatus) in
200118. Resistance to PPO herbicides in weedy species has been attributed to target-site mutation
in the PPX2 gene19-20. A unique target-site amino acid deletion (Gly210) and Arg98Leu

132
substitution confers PPO resistance in waterhemp19 and common ragweed (Ambrosia
artemisiifolia)20, respectively.
PPO herbicides are widely utilized for controlling glyphosate-resistant Palmer amaranth
in conventional and RoundupReady® soybeans and in cotton. Intensive use of herbicides exerts
high selection pressure leading to the evolution of herbicide-resistant weed populations. This
paper describes fomesafen-resistance in Palmer amaranth populations from Arkansas.
2 MATERIALS AND METHODS
2.1 Plant materials
In the late summer of 2011, Palmer amaranth samples escaping from either glufosinate or
PPO-inhibiting herbicide applications were collected from several fields in Arkansas.
Inflorescences from 10 female Palmer amaranth plants per field were collected, dried, threshed,
and cleaned for bioassay in the greenhouse. One such field in Lawrence County has been planted
with LibertlyLink® soybean and RoundupReady® corn in alternate years since 2011 and treated
with either fomesafen and glufosinate or glyphosate. Fomesafen was applied during the soybean
production year. Five-hundred mg of seeds from each of the 10 plants per field were mixed to
make a composite, which was used for subsequent experiments. Susceptible Palmer amaranth
seeds (SS) were collected from a vegetable field in Crawford County, Arkansas. This vegetable
field was not exposed to glyphosate or PPO-inhibiting herbicides.
Plants were grown in a greenhouse maintained at 32/25 °C ± 3 °C day/night temperature
with a 16-h photoperiod. Plants were watered daily and fertilized with MiracleGro®, a water-
soluble all-purpose plant food containing 15-30-15% NPK, every two weeks.

133
2.2 Fomesafen resistance bioassay in the greenhouse
Palmer amaranth seeds were planted in 28 x 54 cm cellular trays (Redwayfeed Garden
and Pet supply, 290 Briceland Rd, Reedway, CA 95560) using Sunshine® premix soil (Sunshine
premix #1®, Sun Gro Horticulture, 15831 NE 8th Street, Suite 100, Bellevue, WA 98008). The
experiment was conducted in a randomized complete block design with five replications. Each
replication consisted of one cellular tray with 50 seedlings, grown at one seedling per cell. The
test was repeated. Thus, from each composite seed sample, a total of 500 plants were sprayed
with the recommended dose of fomesafen at 264 g ha-1 (Flexstar 1.88 EC, Syngenta) when
seedlings were 7.5-9 cm tall. The herbicide was applied with 0.5% by volume nonionic
surfactant (NIS), using a laboratory sprayer equipped with a flat fan spray nozzle (TeeJet spray
nozzles, Spraying Systems Co., Wheaton, IL) delivering 187 L ha-1 at 269 kPa. After 21 d, the
overall effects of fomesafen (stunting, chlorosis, necrosis, and desiccation) were assessed
visually relative to the nontreated control using a scale of 0 to 100 where 0 = no visible injury
and 100 = complete death.
Survivors from fomesafen treatment were grown, and allowed to interbreed, to produce
C1 and C2 populations of progenies. A subset of nine survivors (which eventually consisted of 3
females and 6 males) from AR11-LAW-B produced the C1 population. C1 plants were sprayed
with fomesafen following the same procedure described previously. A subset of six survivors
(which eventually consisted of 1 female and 5 males) from C1 plants were grown for seed
production to produce C2 population.

134
2.3 Progression of resistance from the original AR11-LAW-B to C2 Palmer amaranth
population
Seeds from the C1 and C2 Palmer amaranth populations were planted in cellular trays to
determine the frequency of fomesafen-resistant plants and the progression of resistance from the
original population to the C2 population. The experiment was conducted twice in a randomized
complete block design with 5 replications; the C1 population was bioassayed four times. In each
cellular tray or replication, 50 plants were grown separately and sprayed with 264 g ha-1 of
fomesafen using the herbicide-application method described previously. Mortality and plant
injury were recorded at 21 DAT.
2.3 Fomesafen dose-response bioassay
Palmer amaranth seeds of the susceptible, original AR15-LAW-B, C1, and C2
populations were planted in 11 x 11 cm square pots filled with Sunshine Mix LC1 potting soil
(Sun Gro Horticulture Canada Ltd., Vancouver, British Columbia, Canada). Seedlings were
thinned to five per pot. Seedlings at 7.5-9 cm tall were sprayed with eight doses of fomesafen
from 0 to 2109 g ha-1, which corresponds to 0 to 8 times the recommended field dose. The SS
population was sprayed with 7 doses from 4 to 264 g ha-1 corresponding to 1/64 to 1X
recommended dose, with a nontreated check. The herbicide was applied with a 0.5% NIS as
described in Section 2.2. The experiment was conducted twice in randomized complete block
design with five replications. At 21 DAT, visible injury and the number of survivors were
recorded. The above-ground plant tissue was harvested, placed in brown paper bag, dried at 60°C
for 48 h, and dry weights recorded. Data were expressed as percentage of biomass reduction
relative to the nontreated control. The biomass data generated from two runs were pooled as the
test for homogeneity of variance showed that the variance across runs was similar. Regression

135
analysis was conducted using SigmaPlot v.13. The percent biomass reduction and mortality were
fitted to nonlinear, sigmoid, three-parameter Gompertz regression model defined by (Equation
1),
y = a*exp [-exp {-b*(x-c)}] [1]
where Y is the biomass reduction expressed as a percentage of the nontreated control or
mortality percentage; a is the asymptote; b is the growth rate; c is the inflection point; and x is
the fomesafen dose. The dose needed to kill 50% (LD50) of the population or cause 50% biomass
reduction (GR50) was calculated from the above equation.
2.4 Response of Palmer amaranth population to other foliar-applied herbicides
Palmer amaranth seeds from the susceptible and original AR11-LAW-B populations were
planted in cellular trays in the greenhouse. Uniform-sized plants (7.5-9 cm tall) were treated with
atrazine at 2244 g ha-1, dicamba at 280 g ha-1, glufosinate at 547 g ha-1, glyphosate at 870 g ha-1,
mesotrione at 105 g ha-1, and ALS inhibitors. Glufosinate and mesotrione treatments included
3366 g ammonium sulfate (AMS) ha-1 and 1% crop oil concentrate (COC), respectively. The
ALS inhibitors and their respective rates included pyrithiobac at 73 g ha-1 and trifloxysulfuron at
8 g ha-1, applied with 0.25% NIS by volume. Herbicide treatments were applied as described in
Section 2.2. Following application, plants were placed on greenhouse benches in a randomized
complete block design. Each treatment was replicated twice, with each replication consisting of
50 plants. Mortality was assessed at 21 DAT.
Because AR11-LAW-B was found resistant to pyrithiobac and trifloxysulfuron, dose-
response assays were conducted to determine the level of resistance to these herbicides. Seeds
were planted in 13-cm round pots filled with commercial potting soil and seedlings were thinned

136
to five plants per pot. The SS population was sprayed with 8 herbicide doses from 1/16X to 4X
the recommended dose of pyrithiobac (1X = 73 g ha-1) and trifloxysulfuron (1X = 8 g ha-1),
including a nontreated check. AR11-LAW-B population was treated with eight doses of
pyrithiobac (0 to 1166 g ha-1) and eight doses of trifloxysulfuron (0 to 31 g ha-1), which
corresponds to 0 to 16X the recommended herbicide dose. Herbicides were applied following the
procedure described previously. The experiment was conducted in a completely randomized
design with five replications. At 28 DAT, plants were cut at the soil surface, stored in the dryer
for 2 d, and the dry weights recorded. Percent biomass reduction relative to the nontreated
control were fitted to nonlinear, sigmoid, three-parameter Gompertz regression model. The dose
needed to reduce the aboveground biomass by 50% was obtained from the above equation using
SigmaPlot v. 13.
2.5 Mechanism of resistance in PPO-resistant Palmer amaranth
Fomesafen survivors from C1 and C2 populations were tested for the presence of the
PPO glycine 210 deletion (ΔG210). This deletion confers resistance to PPO herbicides in
waterhemp, a relative weedy species of Palmer amaranth19. Young leaf tissues from 81 C1 and
13 C2 plants that survived the application of 264 g fomesafen ha-1 were collected and stored at -
80°C. Tissues from three sensitive plants (SS) were also collected for DNA extraction. Genomic
DNA from 100 mg leaf tissue was extracted using hexadecytrimethylammonium bromide
(CTAB) method21 following the modification of Sales et al22. The extracted genomic DNA was
quantified using a NanoDrop spectrophotometer model ND-1000 (Thermo Scientific,
Wilmington DE).

137
The ΔG210 codon deletion was detected using an allele-specific PCR assay as previously
described for waterhemp23. The same assay was predicted to work for Palmer amaranth based on
PPX2 sequence data previously generated from this species24.
3 RESULTS AND DISCUSSION
3.1 Progression of PPO-resistance in PA-AR11-LAW-B population
The frequency of fomesafen-resistant plants increased from 5% in the original AR11-
LAW-B to 17% in the C2 population, in response to the 264 g ha-1 dose of fomesafen. Of the 500
plants treated, 25 survived in the original population. The number of survivors increased in the
C1 (n=56) and C2 (n=86) population. By the practical description of a resistant population25, the
field population in 2011 (5% resistant individuals) would still be considered a susceptible
population. The few remnant plants in the field did not cause any economic loss nor were noticed
by the farmer. Until the number of resistant plants reach a level that presents a problem for
management and yield loss, the population would not be considered resistant25. This current
work showed that once a few individuals carrying the resistance trait is selected, sustained
herbicide selection pressure on a prolific species like Palmer amaranth could produce a resistant
population in two years. This would be when the farmer would call for assistance from crop
advisors or Extension Service (personal experience). By the time a population-level resistance is
noticed, it is already too late for that field; by then the resistant plants would have had already
deposited a large amount of seeds in the soil and the resistant allele(s) would remain in the
population.
Not all progenies of fomesafen-resistant survivors carried the PPO-mutation as shown by
the resistance frequencies. This suggests that the resistance trait is segregating and progenies are
heterogeneous. Palmer amaranth has wide genetic variability owing to its cross-pollinating

138
behavior. However, for every round of selection, the population becomes more homogeneous.
The C2 population had more frequency of resistant plants than C1 or its parent population AR11-
LAW-B. After two cycles of selection, the frequency of resistant plants increased about three
times. There are examples of rapid evolution of herbicide resistance in response to intense and
sustained selection pressure26. The Weed Science Society of America (WSSA) defines herbicide
resistance as the inherited ability of a plant to survive and reproduce following exposure to a
dose of herbicide that is normally lethal to the wild type27. In herbicide resistance, there is a
change in the weed population response (i.e. reduced efficacy of the herbicide) with time as
observed with AR11-LAW-B. Thus, the field-collected sample in 2011 was in the early state of
resistance evolution to PPO herbicides. The large-scale testing of our Palmer amaranth collection
(2008 – 2011) for differential tolerance to fomesafen was conducted in 2013. The tests for
heritability and confirmation of G210 deletion among fomesafen survivors were completed in
2015. Thus, the detection and confirmation of resistance to PPO herbicides in Palmer amaranth
happened years later from collection of the resistance-bearing population.
The farmer of AR11-LAW-B field adopted a corn-soybean crop rotation system. In 2015,
the field was clean, except for a sparse remnant Palmer amaranth (about 1 ha-1). Three plants
were sampled and tested for resistance to fomesafen; one of these tested positive for the G210
deletion mutation. This demonstrates that once the resistant allele has been selected for, it could
remain in the population unless the seedbank is depleted.
3.2 Resistance level to fomesafen
Increasing the dose of fomesafen reduced the dry weight of SS, AR11-LAW-B, C1, and
C2 populations. The fomesafen dose that caused 50% growth reduction or GR50 (+ 1 standard
error) was 13 (+ 0.86), 82 (+ 6.1), 168 (+ 11.9), and 265 (+ 20.4) g fomesafen ha-1 for SS, AR11-

139
LAW-B, C1, and C2 population, respectively (Figure 1, Table 1). Based on these GR50 values,
the level of resistance to fomesafen in AR11-LAW-B, C1, and C2 population was 6-, 13-, and
21-fold relative to the sensitive population (SS). The GR50 increased from 82 g ha-1 in the
original AR11-LAW-B to 265 g fomesafen ha-1 in the C2 population. The commercial field dose
of fomesafen (264 g ha-1) is required to reduce aboveground biomass by 50% in the C2
population indicating that the normal field dose would no longer provide effective control. After
two more cycles of selection from the point where 5% resistant individuals were detected, the
GR50 increased three times, reflecting the observed increase in the frequency of resistant plants.
The herbicide dose that caused 50% mortality (LD50) was 16 (+ 1.3), 45 (+ 8.8), 181 (+
16.8), and 262 (+ 30.1) g fomesafen ha-1 for SS, AR11-LAW-B, C1, and C2 population,
respectively (Figure 2, Table 1). On the basis of LD50 values, AR11-LAW-B, C1, and C2
populations had 3-, 11-, and 16-fold resistance relative to the SS population. The LD50 values of
C1 and C2 populations were relatively similar to the GR50 values. The 0.7X and 1X of the
commercial field dose of fomesafen would kill 50% of the C1 and C2 population, respectively.
Thus, the normal field dose would not control all plants in the C1 and C2 populations, allowing
the resistant plants to proliferate progressively in the next growing season. The PPO-resistant
common waterhemp in Kansas had about the same GR50 value (270 g fomesafen ha-1) as that of
the C2 population18. Although AR11-LAW-B had higher GR50 and LD50 than SS, the
commercial dose of fomesafen can still kill >90% of the AR11-LAW-B population. However,
continued selection had shifted the population response in just two cycles. If each cycle of
selection represents one cropping season, we can predict that resistance to PPO herbicides in the
source field (in 2011) would be apparent in 2013. Unaware of the impending resistance problem
in the field, the farmer happened to have adopted a RoundupReady corn-LibertyLink soybean

140
cropping system, and kept the field clean, except for a few remnant Palmer amaranth in 2015
(1/3 of which, still carried the resistance trait). Thus, prevention of seed production of survivors
cannot be overstated. With good management practices, the evolution of resistant populations
can be slowed down.
3.3 Response of Palmer amaranth population to other foliar-applied herbicides
The AR11-LAW-B population was found to be susceptible to atrazine, dicamba,
glufosinate, glyphosate, and mesotrione but resistant to ALS inhibitors pyrithiobac and
trifloxysulfuron (Table 2). The commercial field dose of pyrithiobac and trifloxysulfuron
controlled AR11-LAW-B 17% and 44%, respectively. The GR50 for pyrithiobac was 3.2 (+ 1.2)
and 44.5 (+ 6.2) g ha-1, respectively, for SS and AR11-LAW-B (Table 3). Similarly, the GR50 for
trifloxysulfuron for SS and AR11-LAW-B was 0.9 (+0.7) and 5.1 (+ 1.1) g ha-1, respectively.
About 0.6X the recommended dose of pyrithiobac and trifloxysulfuron were needed to reduce
the aboveground biomass of AR11-LAW-B by 50%. The AR11-LAW-B population had 14-fold
resistance to pyrithiobac and 5-fold resistance to trifloxysulfuron. The occurrence of multiple
resistances in AR11-LAW-B is not surprising, considering a widespread resistance to ALS
inhibitors among Palmer amaranth in Arkansas28. With the prevalence of glyphosate-resistant
Palmer amaranth population, it is interesting to note that this population is controlled 84% by
glyphosate. The grower of this field had been alternating RoundupReady soybean with rice in the
past years (before 2011), then shifted to LibertyLink soybean in 2011, and used other
management practices for weed control; thus, keeping the population responsive to glyphosate.
This population may or may not be resistant to other PPO herbicides, thus further experiments
need to be conducted to verify its response to other foliar and soil-applied PPO herbicides. A
recent study indicated that soil-applied PPO-inhibitor herbicides remain efficacious in PPO-

141
resistant waterhemp populations in Illinois, although the length of residual activity is shortened
compared with PPO-susceptible waterhemp29. With this population evolving resistance to ALS
and PPO inhibitors, AR11-LAW-B can still be controlled with atrazine, dicamba, glufosinate,
glyphosate, and mesotrione. Reliance on PPO herbicides should be mitigated to hinder or slow
the evolution of PPO resistance in Palmer amaranth. Adequate control of Palmer amaranth can
be achieved with tank-mixes of residual herbicides such as S-metolachlor + metribuzin applied
preemergence followed by pyroxasulfone or S-metolachlor applied early postemergence.
Glufosinate is still an effective option in LibertyLink soybeans; however, it should be coupled
with a preemergence program followed by glufosinate tankmixed with S-metolachlor or
pyroxasulfone early postemergence30. In addition, integration of cultural and mechanical control
practices would be helpful in managing PPO-resistant Palmer amaranth populations31.
3.4 Resistance mechanism in PPO-resistant Palmer amaranth
Results revealed 74 out of 81 C1 plants and all 13 C2 samples tested positive for the
ΔG210 mutation, whereas all the sensitive plants tested negative for the said mutation (data not
shown). This indicated that these survivors were resistant to fomesafen due to target-site
mutation in the PPO gene. Thinglum et al.32 reported that ΔG210 mutation confers resistance to
PPO herbicides in waterhemp populations in Illinois, Kansas, and Missouri suggesting that the
ΔG210 mutation is likely the only known mechanism of resistance to PPO inhibitors in
waterhemp. Palmer amaranth and waterhemp belong to the Amaranthus genus and may,
therefore, have some common morphological, biological, and physiological characteristics and
genomic tendencies. In fact, based on previous DNA sequence comparisons, it was predicted that
the ΔG210 mutation would evolve in Palmer amaranth24. The loss of a glycine residue in the PPO

142
gene alters the architecture of the substrate-binding domain of the PPO enzyme33. As a result, the
mutated PPO enzyme has reduced affinity for several PPO-inhibiting herbicides.
Some survivors that did not show the PPO mutation may harbor other resistance
mechanisms such as non-target site-based resistance mechanisms. Diphenyl ether herbicides are
detoxified in soybeans by homoglutathione conjugation34. Similarly, tolerance to the diphenyl
ether fluorodifen in peas is due to rapid conjugation with glutathione35. Alternatively, the lack of
detection of the ΔG210 mutation in some resistant plants could be due to sequence
polymorphisms at the primer binding sites, resulting in false negatives.
3.5 Implications and future research
PPO inhibitors have been used heavily in the past years to combat herbicide-resistant
Palmer amaranth. Before this, PPO herbicides were among the most widely used herbicides for
soybean. As history has shown, intensive use of any herbicides often results in the selection for
genes conferring herbicide resistance in weed populations. This research confirmed the
occurrence of the first Palmer amaranth population to have evolved resistance to a PPO-
inhibiting herbicide. Recurrent selection with the same herbicide significantly increased the
frequency of resistant plants. Because resistance to glyphosate is also rampant, it is expected that
populations with multiple resistance to PPO and ALS inhibitors as well as PPO inhibitors and
glyphosate will evolve. With Palmer amaranth resistant to these modes of action, limited
herbicide options remain for Palmer amaranth control. The evolution of resistance to PPO
inhibitors in Palmer amaranth is a recent phenomenon in the southern US; thus, best
management practices (including diversification of herbicide modes of action) are vital to
manage the spread of resistance, especially in soybean and cotton acres. Since PPO resistance

143
may still be localized, best management practices should be employed on a broader scale
immediately.
Future research will investigate the genetics and mechanism of inheritance to PPO
inhibitors in AR11-LAW-B C1 and C2 population, the efficacy of other PPO herbicides (soil or
foliar) on PPO-resistant populations, the distribution and population genetics of PPO-resistant
Palmer amaranth populations and fitness of multiple-resistant plants.

144
REFERENCES
1. Ward SM, Webster TM and Steckel LE, Palmer Amaranth (Amaranthus palmeri): A
Review. Weed Technol 27:12-27 (2013).
2. Jha P, Norsworthy JK, Riley MB, Bielenberg DG and Bridges W, Jr., Acclimation of
Palmer amaranth (Amaranthus palmeri) to shading. Weed Sci 56:729-734 (2008).
3. Keeley PE, Carter CH and Thullen RJ, Influence of planting date on growth of Palmer
amaranth (Amaranthus palmeri). Weed Sci 35:199-204 (1987).
4. Ehleringer J, Ecophysiology of Amaranthus palmeri, a sonoran desert summer annual.
Oecologia 57:107-112 (1983).
5. Bensch CN, Horak MJ and Peterson D, Interference of redroot pigweed (Amaranthus
retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A. rudis) in
soybean. Weed Sci 51:37-43 (2003).
6. Massinga RA, Currie RS, Horak MJ and Boyer J, Interference of Palmer amaranth in
corn. Weed Sci 49:202-208 (2001).
7. Fast BJ, Murdoch SW, Farris RL, Willis JB and Murray DS, Critical timing of Palmer
amaranth (Amaranthus palmeri) removal in second-generation glyphosate-resistant
cotton. J. Cotton Sci 13:32-36 (2009).
8. Smith DT, Baker RV and Steele GL, Palmer amaranth (Amaranthus palmeri) impacts on
yield, harvesting, and ginning in dryland cotton (Gossypium hirsutum). Weed Technol
14:122-126 (2000).
9. Heap I. The International Survey of Herbicide Resistant Weeds.[Online]. Available:
http.www.weedscience.org/[20 October 2015].
10. Steckel LE, The dioecious Amaranthus spp.: Here to stay. Weed Technol 21:567-570
(2007).
11. Sosnoskie LM, Webster TM, Kichler JM, MacRae AW, Grey TL and Culpepper AS,
Pollen-mediated dispersal of glyphosate-resistance in Palmer amaranth under field
conditions. Weed Sci 60:366-373 (2012).
12 Hao GF, Zuo Y, Yang SH and Yang GF, Protoporphynogen oxidase inhibitor: an ideal
target for herbicide discovery. Chimia 65:961-969 (2011).
13. Deybach JC, Dasilva V, Grandchamp B and Nordmann Y, The mitochondrial location of
protoporphyrinogen oxidase. Eur J Biochem 149:431-435 (1985).
14. Lermontova I, Kruse E, Mock HP and Grimm B, Cloning and characterization of a
plastidal and a mitochondrial isoform of tobacco protoporphyrinogen IX oxidase. Proc.
Natl Acad Sci USA 94:8895-8900 (1997).
15. Lermontova I and Grimm B, Overexpression of plastidic protoporphyrinogen IX oxidase
leads to resistance to the diphenylether herbicide acifluorfen Plant Physiol 122:987-987
(2000).

145
16. Watanabe N, Che FS, Iwano M, Takayama S, Yoshida S and Isogai A, Dual targeting of
spinach protoporphyrinogen oxidase II to mitochondria and chloroplasts by alternative
use of two in-frame initiation codons. J of Bio Chem 276:20474-20481 (2001).
17. Sherman TD, Becerril JM, Matsumoto H, Duke MV, Jacobs JM, Jacobs NJ and Duke
SO, Physiological basis for differential sensitivities of plant species to
protoporphyrinogen oxidase-inhibiting herbicides. Plant Physiol 97:280-287 (1991).
18. Shoup DE, Al-Khatib K and Peterson DE, Common waterhemp (Amaranthus rudis)
resistance to protoporphyrinogen oxidase-inhibiting herbicides. Weed Sci 51:145-150
(2003).
19. Patzoldt WL, Hager AG, McCormick JS and Tranel PJ, A codon deletion confers
resistance to herbicides inhibiting protoporphyrinogen oxidase. Proc. Natl Acad Sci USA
103:12329-12334 (2006).
20. Rousonelos SL, Lee RM, Moreira MS, VanGessel MJ and Tranel PJ, Characterization of
a common ragweed (Ambrosia artemisiifolia) population resistant to ALS- and PPO-
inhibiting herbicides. Weed Sci 60:335-344 (2012).
21. Doyle JJ and Doyle JL, A rapid DNA isolation procedure for small quantities of fresh
leaf tissue. In Phytochem Bull, pp. 11-15 (1987).
22. Sales MA, Shivrain VK, Burgos NB and Kuk YI, Amino acid substitutions in the
acetolactate synthase gene of red rice (Oryza sativa) confer resistance to imazethapyr.
Weed Sci 56:485-489 (2008).
23. Lee RM, Hager AG and Tranel PJ, Prevalence of a novel resistance mechanism to PPO-
inhibiting herbicides in waterhemp. Weed Sci 56:371-375 (2008).
24. Riggins CW and Tranel PJ, Will the Amaranthus tuberculatus resistance mechanism to
PPO-inhibiting herbicides evolve in other Amaranthus species? Int J Agron Vol. 212,
doi:10.1155/2012/305764 (2012).
25. Heap I, Criteria for confirmation of herbicide-resistant weeds. [Online]. Available:
http.www.weedscience.org/[25 November 2015].
26. Weed Science Society of America, Herbicide resistance and herbicide tolerance
definition. Weed Technol 12:789 (1998).
27. Powles SB and Yu Q, Evolution in action: plants resistant to herbicides. Ann Rev Plant
Bio 61: 317-347 (2010).
28. Burgos NR, Alcober EAL, Lawton-Rauh A, Rauh B, Estorninos L, Tseng TM and Smith
KL, The spread and population genetics of glyphosate-resistant Palmer amaranth in
Arkansas. Summ Ark Cot Res 582:94-104 (2009).
29. Wuerffel RJ, Young JM, Matthews JL and Young BG, Characterization of PPO-
inhibitor-resistant waterhemp (Amaranthus tuberculatus) response to soil-applied ppo-
inhibiting herbicides. Weed Sci 63:511-521 (2015).

146
30. Steckel L, PPO-resistant Palmer amaranth likely in Tennessee. In Southeast Farm Press
[Online]. Available: http://southeastfarmpress.com/[22 July 2015].
31. Schultz JL, Chatham LA, Riggins CW, Tranel PJ and Bradley KW, Distribution of
herbicide resistances and molecular mechanisms conferring resistance in Missouri
waterhemp (Amaranthus rudis Sauer) populations. Weed Sci 63:336-345 (2015).
32. Thinglum KA, Riggins CW, Davis AS, Bradley KW, Al-Khatib K and Tranel PJ, Wide
distribution of the waterhemp (Amaranthus tuberculatus) G210 PPX2 mutation, which
confers resistance to PPO-inhibiting herbicides. Weed Sci 59:22-27 (2011).
33. Dayan FE, Daga PR, Duke SO, Lee RM, Tranel PJ and Doerksen RJ, Biochemical and
structural consequences of a glycine deletion in the alpha-8 helix of protoporphyrinogen
oxidase. Biochim Biophys Acta 1804:1548-1556 (2010).
34. Skipsey M, Andrews CJ, Townson JK, Jepson I and Edwards E, Substrate and thiol
specificity of a stress-inducible glutathione transferase from soybean. FEBS Letters 409:
370-374 (1997).
35. Edwards S. Characterizationof glutathione transferase and glutathione peroxidases in pea
(Pisum sativum). Physiol Planta 98:594-604 (1996).
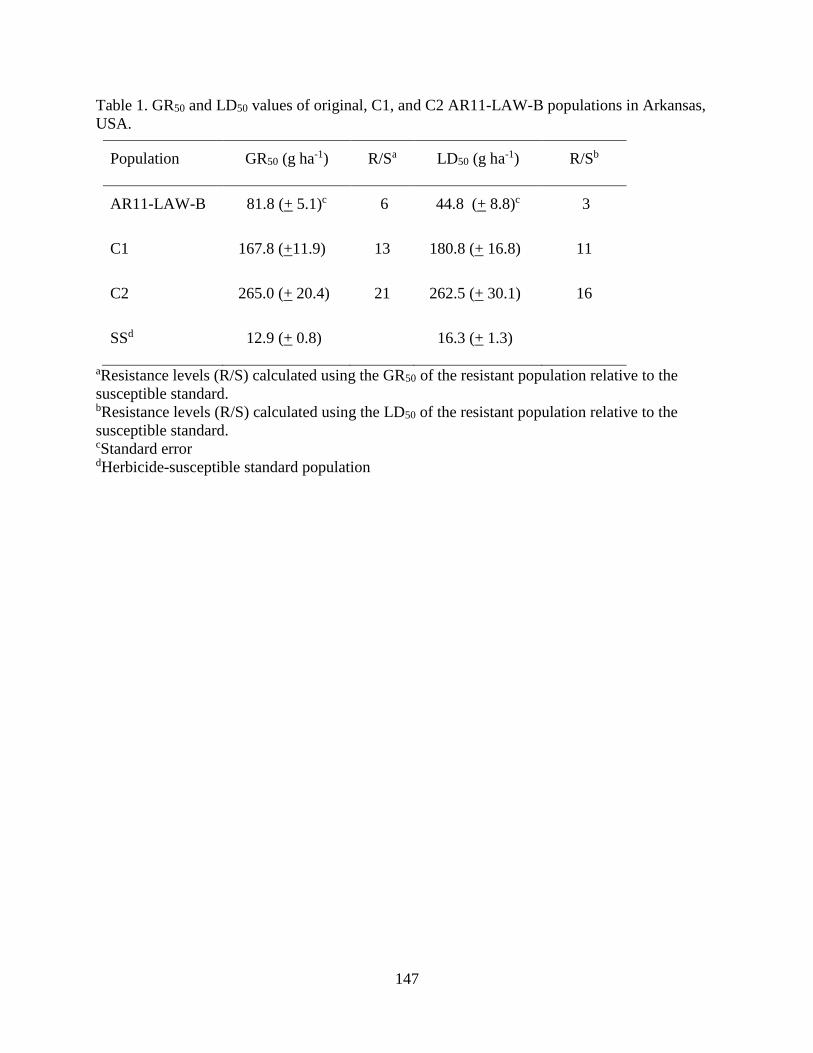
147
Table 1. GR50 and LD50 values of original, C1, and C2 AR11-LAW-B populations in Arkansas,
USA.
Population GR50 (g ha-1) R/Sa LD50 (g ha-1) R/Sb
AR11-LAW-B 81.8 (+ 5.1)c 6 44.8 (+ 8.8)c 3
C1 167.8 (+11.9) 13 180.8 (+ 16.8) 11
C2 265.0 (+ 20.4) 21 262.5 (+ 30.1) 16
SSd 12.9 (+ 0.8) 16.3 (+ 1.3)
aResistance levels (R/S) calculated using the GR50 of the resistant population relative to the
susceptible standard. bResistance levels (R/S) calculated using the LD50 of the resistant population relative to the
susceptible standard. cStandard error dHerbicide-susceptible standard population

148
Table 2. Response of Amaranthus palmeri AR11-LAW-B population to foliar-applied herbicides
other than protoporphyrinogen oxidase inhibitors, Arkansas, USA.
Herbicide Mortality (%)a Resistance classification
atrazine 100 susceptible
dicamba 100 susceptible
glufosinate 100 susceptible
glyphosate 84 susceptibleb
mesotrione 94 susceptible
pyrithiobac 17 resistant
trifloxysulfuron 44 resistant
aUniform-sized plants (7.5-9 cm tall) were sprayed with atrazine at 2244 g ha-1, dicamba at 280 g
ha-1, glufosinate at 547 g ha-1, glyphosate at 870 g ha-1, mesotrione at 105 g ha-1, pyrithiobac at
73 g ha-1 and trifloxysulfuron at 8 g ha-1. Glufosinate and mesotrione treatments included 3366 g
ammonium sulfate (AMS) ha-1 and 1% crop oil concentrate (COC), respectively. Pyrithiobac and
trifloxysulfuron were applied with 0.25% NIS by volume. Mortality was recorded 21 d after
herbicide application. bPlants were not dead at evaluation time, but survivors incurred high injury and did not grow to
maturity.

149
Table 3. GR50 values and resistance levels to ALS inhibitors in AR11-LAW-B Palmer amaranth
population in Arkansas, USA.
Population Pyrithiobac Trifloxysulfuron
GR50 (g ha-1) R/Sa GR50 (g ha-1) R/S
AR11-LAW-B 44.5 (+6.2)b 14 5.1 (+1.1) 5
SSc 3.2 (+1.2) - 0.9 (+0.7) -
aResistance levels (R/S) calculated using the GR50 of the resistant population relative to the
susceptible standard. bStandard error of estimate cHerbicide-susceptible standard population

150
Figure 1. Shoot biomass reduction (%) of PPO-resistant and –susceptible Palmer amaranth
population, 21 d after fomesafen treatment.Treatment means (n = 10) + 1 standard error are
plotted and fitted to a regression curve. Data were best described with a nonlinear, sigmoidal,
three-parameter Gompertz regression function, y =a*exp{-exp[-b*(x-c0]}.

151
Figure 2. Dose-response curves of PPO-resistant and –susceptible Palmer amaranth population
from Arkansas.Mortality (%) were obtained at 21 d after herbicide treatment. Treatment means
(n = 10) + 1 standard error are plotted and fitted to a regression curve. Data were best described
with a nonlinear, sigmoidal, three-parameter Gompertz regression function, y =a*exp{-exp[-
b*(x-c0]}.

152
CHAPTER VI
FREQUENCY OF GLY-210 DELETION MUTATION AMONG
PROTOPORPHYRINOGEN OXIDASE INHIBITOR-RESISTANT PALMER
AMARANTH (AMARANTHUS PALMERI) POPULATIONS
© Weed Science Society of America, 2017

153
Abstract
The widespread occurrence of Palmer amaranth resistant to acetolactate synthase
inhibitors and/or glyphosate led to the increased use of protoporphyrinogen oxidase (PPO)-
inhibiting herbicides. This research aimed to: (1) evaluate the efficacy of foliar-applied
fomesafen to Palmer amaranth, (2) evaluate cross-resistance to foliar PPO inhibitors and efficacy
of foliar herbicides with different mechanisms of action, (3) survey the occurrence of the PPO
Gly-210 deletion mutation among PPO inhibitor– resistant Palmer amaranth, (4) identify other
PPO target-site mutations in resistant individuals, and (5) determine the resistance level in
resistant accessions with or without the PPO Gly-210 deletion. Seedlings were sprayed with
fomesafen (263 g ai ha−1), dicamba (280 g ai ha−1), glyphosate (870 g ai ha−1), glufosinate (549 g
ai ha−1), and trifloxysulfuron (7.84 g ai ha−1). Selected fomesafen-resistant accessions were
sprayed with other foliar-applied PPO herbicides. Mortality and injury were evaluated 21d after
treatment (DAT). The PPX2L gene of resistant and susceptible plants from a selected accession
was sequenced. The majority (70%) of samples from putative PPO-resistant populations in 2015
were confirmed resistant to foliar-applied fomesafen. The efficacy of other foliar PPO herbicides
on fomesafen-resistant accessions was saflufenacil>acifluorfen = flumioxazin>carfentrazone =
lactofen >pyraflufen-ethyl>fomesafen>fluthiacet-methyl. With small seedlings, cross-resistance
occurred with all foliar-applied PPO herbicides except saflufenacil (i.e., 25% with acifluorfen,
42% with flumioxazin). Thirty-two PPO-resistant accessions were multiple resistant to
glyphosate and trifloxysulfuron. Resistance to PPO herbicides in Palmer amaranth occurred in at
least 13 counties in Arkansas. Of 316 fomesafen survivors tested, 55% carried the PPO Gly-210
deletion reported previously in common waterhemp. The PPO gene (PPX2L) in one accession
(15CRI-B), which did not encode the Gly-210 deletion, encoded an Arg-128-Gly substitution.

154
The 50% growth reduction values for fomesafen in accessions with Gly-210 deletion were 8- to
15-fold higher than that of a susceptible population, and 3- to 10-fold higher in accessions
without the Gly-210 deletion.

155
Introduction
Palmer amaranth is one of the most troublesome and economically damaging agronomic
weeds in the southern United States. It is able to adapt to diverse climatic and agricultural
conditions. The photosynthetic rate of Palmer amaranth (81µmol m−2s−1) is three to four times
that of corn (Zea mays L.), cotton (Gossypium hirsutum L.), and soybean [Glycine max (L.)
Merr.] (Ehleringer 1983). This high photosynthetic rate translates into its rapid growth rate of
5cm day−1 under optimum growing conditions (Horak and Loughin 2000). Consequently, this
rapid growth rate results in a narrow window of opportunity for effective POST herbicide
application. Palmer amaranth control becomes difficult when it is >10-cm tall (Riar et al. 2013).
A female Palmer amaranth can produce up to 1 million seeds; however, its seeds are relatively
short-lived in the soil, with about 80% mortality in 3yr (Sosnoskie et al. 2014). With its fast
growth rate, high fecundity, season-long emergence, high photosynthetic rate, and high
propensity to evolve herbicide resistance, Palmer amaranth has become a serious weed in row
crops and vegetables (Ehleringer et al.1997; Guo and Al-Khatib 2003; Jha and Norsworthy 2012;
Steckel 2007). Palmer amaranth infestation can reduce corn, cotton, and soybean yield 91%,
77%, and 78%, respectively (Bensch et al. 2003; Fast et al. 2009; Massinga et al. 2001). In the
past decade, reports abound of Palmer amaranth evolving resistance to glyphosate and other
herbicides (Culpepper et al. 2006; Jha et al. 2008; Norsworthy et al. 2008). To date, Palmer
amaranth has been confirmed resistant to acetolactate synthase (ALS) inhibitors (Burgos et al.
2001; Horak and Peterson 1995), dinitroanilines (Gossett et al. 1992), 4-hydroxyphenylpyruvate
dioxygenase (HPPD) inhibitors (Jhala et al. 2014), glyphosate (Culpepper et al. 2006;
Norsworthy et al. 2008), photosystem II herbicides (Vencill et al. 2011), and most recently,
protoporphyrinogen oxidase (PPO) inhibitors (Salas et al. 2016).

156
Several chemical families inhibit PPO activity, including diphenylethers (e.g.,
acifluorfen, fomesafen, lactofen, oxyfluorfen), N-phenylphthalimide (e.g., flumioxazin,
flumiclorac), phenylpyrazoles (e.g., fluazolate, pyraflufen-ethyl), oxadiazole (e.g., oxadiazon),
oxazolidinones (e.g., pentoxazone), pyrimidinediones (e.g., saflufenacil), thiadiazole (e.g.,
fluthiacet-methyl), and triazolinone (e.g., carfentrazone, sulfentrazone) (Heap2017).
Diphenylether herbicides are used PRE or POST. Some triazolinones and N-phenylphthalimide
have high soil activity and are phytotoxic to the crop when applied foliar, hence most are
commonly used PRE (Dayan and Duke 2010). There are two known nuclear PPO genes in
plants, PPX1 and PPX2, which encode plastid- and mitochondrial-targeted PPO isoforms,
respectively; however, some PPX2 isoforms are dual targeted to both organelles (Lermontova
and Grimm 2000; Lermontova et al. 1997; Watanabe et al. 2001). The inhibition of PPO causes
accumulation of protophorphyrinogen IX, which leaks from the plastid to the cytoplasm, where it
is oxidized rapidly into photosensitive protoporphyrin IX (Becerril and Duke 1989; Jacobs et al.
1991; Lee et al. 1993). Upon exposure to light, protoporphyrin IX generates singlet oxygen
molecules that cause lipid peroxidation, membrane destruction, and ultimately cell death
(Becerril and Duke 1989; Jacobs et al. 1991). The first weed species to evolve resistance to PPO-
inhibiting herbicides was common waterhemp in 2001 (Shoup et al. 2003). Resistance to PPO
inhibiting herbicides in Palmer amaranth was first reported in Arkansas and is confirmed also in
Tennessee and Illinois (Heap 2017; Salas et al. 2016; L Steckel and K Gage, personal
communication). When this research was conducted, resistance to PPO herbicides in Amaranthus
spp. was attributed to a target-site mutation in the PPX2 gene only (Patzoldt et al. 2006; Salas et
al. 2016). The Gly-210 deletion mutation was present in all PPO-resistant waterhemp
populations in Illinois, Kansas, and Missouri (Thinglum et al. 2011). However, a novel PPO

157
mutation, Arg-98-Leu, was detected in PPO-resistant common ragweed (Ambrosia artemisiifolia
L.) (Rousonelos et al. 2012).
This study was conducted to investigate the response of Palmer amaranth from Arkansas
to various foliar-applied PPO-inhibiting herbicides (acifluorfen, carfentrazone, flumioxazin,
fluthiacetmethyl, fomesafen, pyraflufen-ethyl, saflufenacil) and to foliar-applied herbicides with
different modes of action (dicamba, glyphosate, glufosinate, trifloxysulfuron). The study also
aimed to determine the frequency of Gly-210 deletion among the PPO-resistant biotypes,
identify other PPO target-site mutations that endow resistance to PPO-inhibiting herbicides, and
compare the resistance level to fomesafen in accessions that contained or lacked the Gly-210
deletion.
Materials and Methods
Plant Materials. A total of 124 Palmer amaranth accessions were collected in the summer
between 2008 and 2015 from 23 counties in Arkansas, primarily from the eastern half of the
state. Sampling sites were identified by county extension agents and crop consultants. Samples in
2008 and 2009 were collected from 18 counties (17 from eastern Arkansas and 1 from central
Arkansas) to survey resistance to glyphosate. Seven counties were sampled in 2011 from fields
that were planted with LibertyLink® crops to verify differential response to glufosinate. Fifty-
two fields from 16 counties were sampled in 2014. Samples in 2015 (23 accessions) were
collected specifically to survey fields with remnant Palmer amaranth after having been sprayed
with PPO herbicides. Inflorescences from at least 10 mature female plants−1 field were
collected, dried, and threshed. Equal amounts of seeds from each plant were mixed to make a
composite seed sample to represent the field. A sensitive standard population (SS) was included
in each experiment for comparison. Plants were grown in a greenhouse maintained at 32/25±3°C

158
day/night temperature with a 16:8-h day:night regime and photon flux density of 0.0005 mol m−2
s−1. Plants were planted using commercial potting soil (Sunshine® Premix No. 1; Sun Gro
Horticulture, Bellevue, WA), watered daily, and fertilized with Miracle-Gro® (Scotts Miracle-
Gro, Marysville, OH) every 2 wk.
Palmer Amaranth Response to Foliar-applied Fomesafen. One hundred-twenty four Palmer
amaranth accessions (Table 1) were tested in the greenhouse for resistance to fomesafen.
Composite seed samples were planted in 24 by 54 cm cellular trays. Seedlings were thinned to 1
plant cell−1 and sprayed with 263g ai ha−1 fomesafen (Flexstar®, Syngenta Crop Protection,
Greensboro, NC 27419) when seedlings were 7- to 8-cm tall. The herbicide was applied with
0.5% by volume nonionic surfactant (NIS) (Induce®, Helena Chemical, Collierville, TN 38017)
using a laboratory sprayer equipped with a flat-fan spray nozzle delivering 187 L ha−1 at 221
kPa. The experiment was conducted in a randomized complete block design with two
replications and repeated. Each replication consisted of 50 seedlings grown in a cellular tray at 1
seedling cell−1. The plants were assessed visually relative to nontreated plants at 21 d after
treatment (DAT) using an injury scale of 0 to 100, where 0% = no visible injury and 100% =
complete desiccation. Survivors with 0% to 10%, 11% to 30%, 31% to 60%, and 61% to 89%
injury were classified as highly resistant, resistant, moderately resistant, and slightly resistant,
respectively. Individuals with 90% injury or higher were considered sensitive. In this
experiment, an accession with >10% survivors that were at least slightly resistant was considered
to be PPO resistant. Data were analyzed using hierarchal clustering in JMP Pro v. 13 (SAS
Institute, Cary, NC.).
Response to Other Foliar-applied PPO Herbicides. Twelve accessions that had low mortality
(<83%) from foliar-applied fomesafen were bioassayed in the greenhouse for their response to

159
other foliar-applied PPO herbicides. A sensitive standard accession was also included. Seedlings
(100 per accession, 10-cm tall) were treated with the recommended doses of acifluorfen,
carfentrazone, flumioxazin, fluthiacet-methyl, lactofen, pyraflufenethyl, or saflufenacil (Table 2).
Carfentrazone, pyraflufen-ethyl, flumioxazin, and fluthiacet-methyl treatments included 0.25%
NIS (v/v), whereas acifluorfen included 0.125% NIS (v/v). Lactofen was sprayed with 0.5% crop
oil concentrate (v/v). Saflufenacil was sprayed with 1% methylated seed oil and 1% ammonium
sulfate (w/v). Following herbicide applications, the plants were placed in the greenhouse,
grouped by herbicide, and the accessions were randomized within each herbicide group.
Mortality and injury of survivors were evaluated at 21 DAT. In the second run of the experiment,
herbicides were sprayed when seedlings were 5- to 8-cm tall. Data were analyzed, by herbicide,
using JMP Pro v. 13 (SAS Institute, Cary, NC).
Response to Foliar-applied Non-PPO Herbicides. Seventy-three accessions collected in 2014
and 2015 were tested in the greenhouse for response to dicamba, glufosinate, glyphosate, and
trifloxysulfuron (Table 2). The SS accession was also included for comparison. The experiment
was conducted in a completely randomized design with two replications and two runs. Each
replication consisted of 50 plants grown in a cellular tray. Composite seed samples were planted
as described earlier. The foliar herbicides were applied to uniform-sized plants (7.6-cm tall).
Glufosinate and trifloxysulfuron were sprayed with 3,366g AMS ha−1 and 0.25% NIS (v/v),
respectively. Herbicide applications were made as described earlier. Plants were evaluated for
injury and mortality at 21 DAT. Data were analyzed by herbicide using JMP Pro v. 13 (SAS
Institute, Cary, NC). Response to glyphosate was analyzed using hierarchal clustering,
collectively considering mortality and injury of survivors.

160
Mechanism of Resistance to PPO Herbicides. A total of 316 survivors from 47 accessions
treated with foliar-applied fomesafen were analyzed for the Gly-210 deletion mutation (Patzoldt
et al. 2006; Salas et al. 2016). Young leaf tissues were collected from up to 10 survivors each of
35 accessions sprayed with the recommended dose of foliar-applied fomesafen in the greenhouse
bioassays. In addition, leaf tissues were collected from 82 plants in 14 fields with high
population density of Palmer amaranth after having been sprayed with PPO inhibitors, among
other herbicides. DNA was extracted using a modified Cetyltrimethylammonium bromide
(CTAB) protocol (Sales et al. 2008) and quantified using a NanoDrop spectrophotometer (ND-
1000, Thermo Scientific, Wilmington, DE). An allelespecific PCR assay was used to detect the
codon Gly210 following the protocol described for common waterhemp, which also worked for
Palmer amaranth (Lee et al. 2008; Salas et al. 2016). Some PPO-resistant plants that did not have
the Gly-210 deletion mutation were selected for sequencing of the PPX2L gene. Total RNA was
extracted using the RNeasy extraction kit (Qiagen 74903, Valencia, CA) and converted to cDNA
using the Reverse Transcription kit (Promega A3500, Madison, WI). Initially a partial sequence
of the PPX2L gene was amplified using the primer pair ppx2-1F (5′-
GTAATTCAATCCATTACCCACCTT-3′) and ppx2-3R (5′-TTACGCGGTCTTCTCATCCAT-
3′) and sequenced using the internal primers: ppx2-1R (5′-TTCCATACGTCGGGAAATGT-3′),
ppx2-2F (5′-TGTTGGAACCATTTCTCTGG-3′), ppx2-2R (5′-
GGGGATAAGAACTCCGAAGC-3′) and ppx2-3F (5′-GATGCTGTGGTTGTCACTGC-3′).
Eventually, the full-length sequences of PPX2L in PPO-sensitive and PPO-resistant plants were
obtained (GenBank accession nos. MF583744 and MF583746) by designing primers based on
the upstream and downstream gene sequence of Prince-of-Wales feather (Amaranthus
hypochondriacus L.) (GenBank accession no. EU024569.1). These additional primers were:

161
ppx2-5′UTR (5′-TGGCAGATTGAGACAAAATT GG-3′) and ppx2-3 ′UTR (5′-
GGCAGAAAAGTCAC TGCACA-3′).
Fomesafen Dose–Response Bioassay. Seven PPO-resistant accessions, including the SS, were
used in whole-plant bioassays to determine the resistance level to fomesafen. Three accessions
that contained the Gly-210 deletion (14MIS-H, 15MIS-C, 15MIS-E) and four accessions that
lacked the Gly-210 deletion mutation (14CRI-C, 15CRI-B, 15PHI-A) were selected. Seedlings
were grown in 15-cm-diameter pots and thinned to 5 plants pot−1. Seedlings, 5- to 7-cm tall, were
sprayed with 10 doses of fomesafen from 0 to 2,107g ai ha−1. The SS was sprayed with 9 doses
from 2 to 263 g ha −1, corresponding to 1/128 to 1X the recommended dose. A nontreated check
was included. The herbicide was applied with 0.5% NIS. The experiment was conducted in a
randomized complete block design with four replications. At 21 DAT, plants were cut at the soil
surface, shoots were dried for 48 h at 60°C, and the dry weights were recorded. Data were
analyzed using SAS JMP Pro v.13 (SAS Institute, Cary, NC) in conjunction with SigmaPlot v.13
(Systat Software, San Jose, CA) for regression analysis. The percentage biomass reduction was
fit to a nonlinear, sigmoid, four-parameter logistic regression model defined by
𝑦 = 𝑐 + [(𝑑 − 𝑐)/(1 + 𝑒{−𝑎(𝑥 − 𝑏)})]
where y is the biomass reduction expressed as percentage relative to the untreated control, a is
the growth rate, b is the inflection point, c is the lower asymptote, d is the upper asymptote, and x
is the fomesafen dose. Estimates of herbicide dose that cause 50% growth reduction (GR50) were
determined using the fitted regression equation.

162
Results and Discussion
Palmer Amaranth Response to Foliar-applied Fomesafen. The frequency distribution of
mortality from fomesafen treatment was highly skewed, with a greater proportion of high
mortality; therefore, the median values were used to describe the data, except for the 2015
accessions (Figure 1). The 2008 accessions generally incurred 97% mortality with the field dose
of fomesafen, except for three accessions with <90% mortality. One rare accession from Phillips
County had only 74% mortality; however, the survivors incurred >75% injury and did not
produce seeds. Thus, the early samples had individuals that were relatively more tolerant to
fomesafen than others. The accessions in 2009 and 2011 were all susceptible, but one of two
accessions with 95% mortality, 11LAW-B, contained rare individuals carrying the Gly-210
deletion mutation (Salas et al. 2016). Eleven accessions (22%) collected in 2014 and 16
accessions (70%) in 2015 were classified resistant. The 2015 samples were from fields with
suspected resistance to PPO herbicides, and the results confirmed most growers’ suspicions. The
mortality data with fomesafen treatment was normally distributed from 23% to 100%, with an
average of 71%. Accessions classified as PPO resistant in 2015 consisted of individuals with
variable response to fomesafen (Figure 2). For example, 15CLA-A had 50% survivors and
almost all showed ≤10% injury. On the other hand, 15GRE-A had 75% survivors, and only about
one-half showed ≤10% injury. Overall in 2015, 16 accessions from 10 counties showed mostly
<90% mortality with fomesafen and were classified resistant based on further evaluations
discussed in succeeding sections. This shows that resistance to PPO herbicides among Palmer
amaranth has evolved quasi-simultaneously across counties in Arkansas.
Prior to 2015, there were fields with a few Palmer amaranth remaining after PPO
herbicide application, but such low numbers did not catch the growers’ attention because of the

163
expected variability in plant response under various environmental conditions. In some cases,
any escapes would have been blamed on various factors. Continued selection with different
herbicides, but with the same PPO-inhibitor mode of action, has increased the frequency of
surviving individuals to a field-scale, observable level. PPO-inhibiting herbicides are used
extensively to control glyphosate- and ALS-resistant Palmer amaranth in soybean and cotton
(Salas et al. 2016). Prior to the widespread glyphosate-resistance problem in Amaranthus,
fomesafen had been used primarily as PRE or POST herbicide for broadleaf weed control in
soybean. Since fomesafen commercialization in the 1960s, several other herbicides targeting the
PPO enzyme have been commercialized. With the expansion of PPO-herbicide use pattern to
cotton and PPO-herbicide use in preplant applications, the selection pressure from this mode-of-
action group has intensified greatly. Considering that rare PPO-resistant Palmer amaranth
individuals were detected retroactively in a population collected in 2011, it took 4 yr before
several reports arose of field-level escapes from PPO herbicide application in 2015.
Fields sampled in 2015 were those with remnant infestations of Palmer amaranth after a
weed management program that included multiple applications of a PPO herbicide. Most of
those fields had plants resistant to 263 g ha−1 fomesafen. The frequency of resistance reported
here pertained mostly to fields with putative PPO herbicide– resistance problem. A random
survey of fields infested with Palmer amaranth across the state, irrespective of cropping history,
is expected to produce a lower frequency of resistance to PPO herbicides. Of the 23 fields
verified for resistance in 2015, only two had 100% mortality with a field dose of fomesafen. The
rest had resistant individuals showing different levels of injury of survivors (Figure 2). The
majority of survivors from three accessions (15CLA-A and 15CRI-A) incurred minimal injury

164
(0% to 10%). High frequency of PPO-resistant individuals among the 2015 accessions threatens
the continued use of PPO herbicides.
Multivariate cluster analysis was conducted to group the 2014 and 2015 accessions based
on their response to the field use rate of fomesafen, taking into account both mortality data and
injury of survivors. The 2014 and 2015 accessions differentiated into four clusters (Figure 3).
The 50 accessions belonging to the first cluster were the most sensitive, the majority of which
were from the quasi-random collection in 2014. The second cluster is composed of 19 resistant
accessions with the majority of survivors showing >60% injury, from the 2014 and 2015
collection. The third cluster comprises two resistant accessions in which most survivors incurred
low (11% to 30%) injury. The fourth cluster contains the three resistant accessions in which a
large number of survivors incurred the lowest (<11%) injury. This indicates that a large number
of PPO-resistant plants incurred low to moderate levels of injury from field use rate of
fomesafen, allowing them to mature and reproduce. The evolution of observable, population-
level resistance to PPO herbicides in multiple fields in 2015 demonstrated the quasi-
simultaneous evolution of resistance to PPO herbicides in Palmer amaranth. This occurred after
several decades of use primarily in soybean, then more recently in cotton. The proportion of
resistant populations that evolved via gene flow (pollen or seed) or via independent selection is
yet to be determined.
Response to Other Foliar-applied PPO-inhibiting Herbicides. The response of Palmer
amaranth to other foliar-applied PPO herbicides and to other non-PPO foliar-applied herbicides
was evaluated, because PPO herbicides are used heavily to control ALS- and glyphosate-
resistant Palmer amaranth. Twelve of the most fomesafen-resistant accessions were tested with
other PPO-inhibiting herbicides (Table 3). Mortality with pyraflufen-ethyl and fluthiacet-methyl

165
was similar across runs; data were therefore combined. These herbicides controlled the SS 100%.
The mortality of all accessions, excluding the SS, was <72% with fluthiacet-methyl. Palmer
amaranth generally has variable sensitivity to this PPO herbicide and is therefore not listed on
the label; whereas this herbicide is expected to provide only partial control of common
waterhemp (Anonymous 2011). The mortality rating of all PPOresistant accessions was at least
17% lower than the SS when treated with pyraflufen-ethyl, with the exception of 14CRI-C
(>90% mortality). Pyraflufen-ethyl also is naturally weak on Palmer amaranth, although it killed
the SS in these experiments, as did the other herbicides.
Mortality was different between runs for acifluorfen, carfentrazone, flumioxazin,
lactofen, and saflufencil; data were therefore analyzed separately. In general, mortality was
higher in the second run than in the first run due to smaller plant size at the time of herbicide
treatment. With acifluorfen, only 2 of 12 fomesafen-resistant accessions tested (14CRI-C and
15CRI-C) were effectively controlled (>90% mortality) in the first run; however, 75% of
fomesafen-R accessions were susceptible to acifluorfen when smaller plants were sprayed in the
second run. With carfentrazone, all 12 fomesafen-resistant accessions showed only 48% to 83%
mortality in the first run. Similar results were observed in the second run. With flumioxazin,
eight accessions that had ≥74% mortality in the first run were verified to be sensitive in the
second run. About 42% of fomesafen-resistant accessions tested were cross-resistant to foliar-
applied flumioxazin. The mortality of all accessions was <70% with lactofen in the first run, with
the exception of 15CRI-C (93% mortality). In the second run, all accessions were still poorly to
moderately controlled, except for 14CRI-C and 15CRI-C (>90% mortality). Saflufenacil was
effective only on five accessions (>90% mortality) with larger seedlings. With smaller seedlings,
all 12 fomesafen-resistant accessions showed >97% mortality with saflufenacil. Across all

166
chemistries, regardless of herbicidal strength, application on small seedlings is necessary for
maximum possible efficacy. Application on bigger seedlings is risky. Previous reports indicated
that resistance of common waterhemp to foliar-applied PPO herbicides becomes prevalent at the
4- to 6-leaf stage (Falk et al. 2006). Plant size is a critical factor in the efficacy and consistency
of performance of most foliar-applied herbicides. When growers miss the application window,
selection for resistance is expected to be stronger. If survival in greenhouse bioassays is seen to
be consistent across replications and repetitions, then the risk of having survivors in the field
populations is high.
Consistent in both runs, fluthiacet-methyl was the least effective and saflufenacil was the
most effective of all foliar-applied PPO herbicides. The response to PPO herbicides, in order of
decreasing efficacy, was as follows: saflufenacil>acifluorfen = flumioxazin> carfentrazone =
lactofen>pyraflufen-ethyl>fluthiacetmethyl. In this series, fomesafen would fall between the last
two herbicides. The fomesafen-resistant accessions were generally cross-resistant to other foliar-
applied PPO herbicides, with the exception of saflufenacil. However, saflufenacil is not an in-
crop option for soybean or cotton. Although saflufenacil showed the greatest efficacy, some
accessions already showed reduced sensitivity to the field use rate of saflufenacil when applied
to 10-cm-tall seedlings. Previous studies on diphenylether-resistant tall waterhemp [Amaranthus
tuberculatus (Moq.) Sauer] also showed cross-resistance to other foliar-applied PPO herbicide
families (Patzoldt et al. 2005; Shoup et al. 2003; Wuerffel et al. 2015a).
Response to Foliar-applied Non–PPO Inhibiting Herbicides. Palmer amaranth accessions
collected between 2014 and 2015 were tested with dicamba, glufosinate, glyphosate, and
trifloxysulfuron. The 73 accessions, including the SS accession, differentiated into four clusters
in response to glyphosate (Figure 4A). The first cluster was composed of 13 resistant accessions

167
that had 0% to 36% mortality with glyphosate, with the majority of survivors incurring 31% to
60% injury. Twenty-three accessions constituted the second cluster, with resistance to glyphosate
in which a large number of the survivors incurred <11%injury. The third cluster was composed
of 24 accessions with ≥65% mortality, 17 of which were classified as sensitive (at least 90%
mortality). The fourth cluster included 14 resistant accessions (36% to 68% mortality), in which
most of the survivors sustained >60% injury from glyphosate treatment. Overall, about 50% of
accessions (Clusters 1 and 2) were resistant to glyphosate, with mortality ranging from 0%
to42%. In addition, all accessions were also resistant to trifloxysulfuron, showing <86%
mortality (Figure 4B). The widespread occurrence of ALS- and glyphosate-resistant Palmer
amaranth in Arkansas was reported previously (Burgos et al. 2009). The ALS-inhibiting
herbicides and glyphosate had been used extensively in the past. Overall, the 27 PPO-resistant
accessions were also resistant to either ALS inhibitor or glyphosate, or both (Figure 5; Table 4).
The majority of PP-Oresistant accessions (85%) exhibited resistance to both glyphosate and
trifloxysulfuron.
The 2014 and 2015 accessions were all sensitive to the 549g ai ha−1 glufosinate (>90%
mortality) (Figure 4B). All accessions incurred >95% mortality, except for 15CRI-B (91%). In
the same manner, dicamba controlled all accessions (Figure 4B). Dicamba caused >90%
mortality in 61% of the accessions. Mortality of the remaining accessions was <89%; however,
the survivors incurred >75% injury. Anecdotal reports, however, indicated poor performance of
dicamba in fields infested with PPO-resistant Palmer amaranth.
Prevalence of the PPO-Gly-210 Deletion Mutation in Resistant Palmer Amaranth. The Gly-
210 deletion in the PPX2L gene confers resistance to PPO-inhibiting herbicides in PPO-resistant
common waterhemp and Palmer amaranth (Patzoldt et al. 2006; Salas et al. 2016; Wuerffel et al.

168
2015b). A point mutation in PPX2 of common ragweed also confers resistance to PPO
herbicides; however, whereas a plastid-targeting signal was not found to be encoded by PPX2 in
common ragweed (Rousonelos et al. 2012), PPX2L in common waterhemp encodes plastidic and
mitochondrial isoforms of PPO (Patzoldt et al. 2006). This dual-targeting phenomenon has been
reported previously for spinach (Spinacia oleracea L.) and corn (Watanabe et al. 2001). The
Gly-210 deletion in imparts herbicide resistance to the dualtargeted protein by altering the
architecture of the substrate binding domain (Dayan et al. 2010).
The molecular survey was carried out on 316 fomesafen-resistant plants from 38
accessions representing 13 counties in Arkansas including Clay, Conway, Crittenden, Greene,
Independence, Jackson, Lawrence, Lee, Independence, Mississippi, Phillips, White, and
Woodruff (Figure 6). The Gly-210 codon deletion mutation was found in 46% and 60% of the
survivors from 2014 and 2015 accessions, respectively (unpublished data). The PPO Gly-210
was prevalent among the PPO-resistant accessions; however, a substantial number of the
resistant plants did not carry this mutation (Figure 7, A and B). In most of the PPO-resistant
accessions (68%), resistant individuals were mixtures of carriers and noncarriers of the Gly-210
deletion. Only 13% of the PPO-resistant accessions (n = 6) contained individuals that were all
carriers of the Gly-210 deletion. In nine accessions, none of the resistant individuals carried the
Gly-210 deletion. This indicates that either an alternative target-site mutation is present or
another resistance mechanism is occurring within and among resistant populations in the field.
To verify the occurrence of alternative target-site mutations, we generated a 1,542-bp sequence
of PPX2L in selected resistant individuals without the Gly-210 deletion (GenBank accession no.
MF583745) from the noncarrier accession 15CRI-B. Resistant plants in this accession contained
a different amino acid mutation, Arg-128-Gly. This mutation was reported recently in parallel

169
investigations of PPO-resistant Palmer amaranth, along with an Arg-128-Met mutation
(Giacomini et al. 2017). A mutation at the homologous site was identified previously in PPO-
resistant common ragweed in the form of an Arg-98-Leu substitution (Rousonelos et al. 2012).
Our survey of more than 300 PPO inhibitor– resistant individuals representing 38 field
populations showed that a resistant population may contain a mixture of plants carrying either
one of these mutations. Henceforth, testing for these mutations should be done simultaneously on
suspected PPO inhibitor–resistant plants using available tools (Giacomini et al. 2017).
Sequencing of Palmer amaranth with different resistance levels to PPO inhibitors, with or
without the Gly-210 mutation, is ongoing.
The occurrence of other resistance-conferring mutations in other loci of the PPO gene is
rare, as indicated by previous research. Many single or double point mutations were reported in
mutant PPO genes in an attempt to obtain PPO herbicide– resistant Arabidopsis (Li and Nicholl
2005). Those authors’ data showed that single point mutations either provided low resistance or
resulted in substantial fitness penalty.
Resistance Level to Fomesafen. The fomesafen dose that caused 50% growth reduction (GR50)
ranged from 116 to 232g ha−1 in accessions that contained the Gly-210 deletion, whereas GR50
ranged from 51 to 153g ha−1 in accessions that lacked the Gly-210 deletion (Table 5). Based on
these GR50 values, the level of resistance to fomesafen relative to the SS ranged from 8- to 15-
fold in accessions that contained the Gly-210 and from 3- to 10-fold in accessions that lacked the
Gly-210 deletion. The dose–response bioassay indicated high resistance level in accessions
carrying the Gly-210 deletion mutation. However, some accessions (14CRI-G and 15CRI-B) that
lacked the Gly-210 deletion had GR50 values comparable to those of accessions that contained
the Gly-210 deletion. Accession 15CRI-B, which contained the Arg-128-Gly mutation, had

170
similar GR50 values to those of the resistant accessions carrying the Gly-210 deletion. This
indicates that Gly-210 and Arg-128-Gly mutations result in comparable levels of resistance to
fomesafen. Accessions 14CRI-C and 15PHI-A, which lacked the Gly-210 deletion, had lower
GR50 values than the other PPO-resistant accessions but higher GR50 values than the SS. These
accessions have not yet been studied for the occurrence of other PPO mutations or other
resistance mechanisms. Although our results indicated that the Arg-128-Gly mutation may
confer a resistance level similar to that of the Gly-210 deletion, additional data are needed to
quantify the impact of this mutation on resistance to PPO herbicides. In addition, we have not
ruled out the possibility that other resistance mechanism (s) exist in some PPO inhibitor–resistant
accessions, particularly those exhibiting low-level resistance to PPO inhibitors. It is also possible
also the populations showing high-level resistance harbor multiple resistance mechanisms, as in
the case of ALS-resistant turnipweed [Rapistrum rugosum (L.) All.] (Hatami et al. 2016).
The increasing number of Palmer amaranth populations with resistance to PPO herbicides
is a great concern, because this limits herbicide options for cotton and soybean. Most PPO-
resistant populations were also resistant to glyphosate and trifloxysulfuron, which leaves almost
no herbicides for POST Palmer amaranth control. The remaining POST herbicide options for
PPO-resistant Palmer amaranth include glufosinate in LibertyLink® crops, dicamba in Roundup
Xtend® or Engenia®, or 2,4-D in Enlist Duo® crops. However, overdependence on glufosinate
and phenoxy herbicides must be avoided, as some Palmer amaranth populations show high
tolerance to dicamba (unpublished data). If we lose glufosinate and dicamba, there will be zero
POST options for weed control in soybean and cotton, unless HPPD and 2,4-D traits are
commercialized soon.

171
Overall, the majority of the PPO-resistant individuals carried the PPO Gly-210 deletion.
An alternative target-site mutation, Arg-128-Gly, was identified in resistant plants of one
accession analyzed that did not carry the Gly-210 mutation. The occurrence of other resistance-
conferring mutations or other resistance mechanisms is being investigated. The combination of
these mutations in one plant may be lethal, but the presence of these mutations in different plants
in one field, or different mutations in proximal fields, may accelerate the evolution and spread of
resistance to PPO inhibitors in Palmer amaranth.

172
Literature Cited
Anonymous (2011) Cadet herbicide product label. Philadelphia, PA: FMC. 4 p
Becerril JM, Duke SO (1989) Protoporphyrin IX content correlates with activity of
photobleaching herbicides. Plant Physiol 90: 1175-81
Bensch CN, Horak MJ, Peterson D (2003) Interference of redroot pigweed (Amaranthus
retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A. rudis) in
soybean. Weed Sci 51: 37-43
Burgos NR, Kuk YI, Talbert RE (2001) Amaranthus palmeri resistance and differential tolerance
of Amaranthus palmeri and Amaranthus hybridus to ALS-inhibitor herbicides. Pest
Manag Sci 57: 449-457
Burgos NR, Alcober EAL, Lawton-Rauh A, Estorninos L, Tseng TM, Smith KL (2009) The
spread and population genetics of glyphosate-resistant Palmer amaranth in Arkansas:
University of Arkansas. 94-104 p
Culpepper AS, Grey TL, Vencill WK, Kichler JM, Webster TM, Brown SM, York AC, Davis
JW, Hanna WW (2006) Glyphosate-resistant Palmer amaranth (Amaranthus palmeri)
confirmed in Georgia. Weed Sci 54: 620-626
Dayan FE, Daga PR, Duke SO, Lee RM, Tranel PJ, Doerksen RJ (2010) Biochemical and
structural consequences of a glycine deletion in the α-8 helix of protoporphyrinogen
oxidase. Biochim Biophy Acta 1804: 1548-1556
Dayan FE, Duke SO (2010) Protoporphyrinogen oxidase-inhibiting herbicides. Pages 1722-1751
in Krieger R, Doull J, Hodgson E, Maibach H, Reiter L, Ritter L, Ross J, Slikker WJ,
Van Hemmen J, eds. Haye's Handbook of Pesticide Toxicology. San Diego: Academic
Ehleringer J (1983) Ecophysiology of Amaranthus palmeri, a sonoran desert summer annual.
Oecologia 57: 107-112
Ehleringer JR, Cerling TE, Helliker BR (1997) C-4 photosynthesis, atmospheric CO2 and
climate. Oecologia 112: 285-299
Falk JS, Shoup DE, Al-Khatib K, Peterson DE (2006) Protox-resistant common waterhemp
(Amaranthus rudis) response to herbicides applied at different growth stages. Weed Sci
54: 793-799
Fast BJ, Murdoch SW, Farris RL, Willis JB, Murray DS (2009) Critical timing of Palmer
amaranth (Amaranthus palmeri) removal in second-generation glyphosate-resistant
cotton. J Cotton Sci 13: 32-36
Giacomini DA, Umphres AM, Nie H, Mueller TC, Steckel LE, Young BG, Scott RC, Tranel PJ
(2017) Two new PPX2 mutation associated with resistance to PPO-inhibiting herbicides
in Amaranthus palmeri. Pest Manag Sci doi:10.1002/ps.4581
Gossett BJ, Murdock EC, Toler JE (1992) Resistance of Palmer amaranth (Amaranthus palmeri)
to the dinitroaniline herbicides. Weed Technol 6: 587-591
Guo PG, Al-Khatib K (2003) Temperature effects on germination and growth of redroot pigweed
(Amaranthus retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A.
rudis). Weed Sci 51: 869-875

173
Hatami ZM, Gherekhloo J, Rojano-Delgado AM, Osuna MD, Alcantara R, Fernandez P,
Sadeghipor HR, de Prado R (2016) Multiple mechanisms increase levels of resistance in
Rapistrum rugosum to ALS herbicides. Front Plant Sci 7: 169
Heap I. (2016) The International Survey of Herbicide Resistant Weeds.
http://www.weedscience.org. Accessed: December 15, 2016
Horak MJ, Loughin TM (2000) Growth analysis of four Amaranthus species. Weed Sci 48: 347-
355
Horak MJ, Peterson DE (1995) Biotypes of Palmer amaranth (Amaranthus palmeri) and common
waterhemp (Amaranthus rudis) are resistant to imazathapyr and thifensulfuron. Weed
Technol 9: 192-195
Jacobs JM, Jacobs NJ, Sherman TD, Duke SO (1991) Effect of diphenyl ether herbicides on
oxidation of protoporphyrinogen to protoporphyrin in organellar and plasma membrane
enriched fractions of barley. Plant Physiol 97: 197-203
Jha P, Norsworthy JK (2012) Influence of late-season herbicide applications on control,
fecundity, and progeny fitness of glyphosate-resistant Palmer amaranth (Amaranthus
palmeri) biotypes from Arkansas. Weed Technol 26: 807-812
Jha P, Norsworthy JK, Riley MB, Bielenberg DG, Bridges W (2008) Acclimation of Palmer
amaranth (Amaranthus palmeri) to shading. Weed Sci 56: 729-734
Jhala AJ, Sandell LD, Rana N, Kruger GR, Knezevic SZ (2014) Confirmation and control of
traizine and 4-hydroxyphenylpyruvate dioxygenase-inhibiting herbicide-resistant Palmer
amaranth (Amaranthus palmeri) in Nebraska. Weed Technol 28: 28-38
Lee HJ, Duke MV, Duke SO (1993) Cellular localization of protoporphyrinogen oxidizing
activities of etiolated barley (Hordeum vulgare L) leaves - relationship to mechanism of
action of protoporphyrinogen oxidase-inhibiting herbicides. Plant Physiol 102: 881-889
Lee RM, Hager AG, Tranel PJ (2008) Prevalence of a novel resistance mechanism to PPO-
inhibiting herbicides in waterhemp (Amaranthus tuberculatus). Weed Sci 56: 371-375
Lermontova I, Grimm B (2000) Overexpression of plastidic protoporphyrinogen IX oxidase
leads to resistance to the diphenyl-ether herbicide acifluorfen. Plant Physiol 122: 75-83
Lermontova I, Kruse E, Mock HP, Grimm B (1997) Cloning and characterization of a plastidal
and a mitochondrial isoform of tobacco protoporphyrinogen IX oxidase. Proc Natl Acad
Sci USA 94: 8895-8900
Li X, Nicholl D (2005) Development of PPO inhibitor-resistant cultures and crops. Pest Manag
Sci 61: 277-285
Massinga RA, Currie RS, Horak MJ, Boyer J (2001) Interference of Palmer amaranth in corn.
Weed Sci 49: 202-208
Norsworthy JK, Scott RC, Smith KL, Oliver LR (2008) Response of northeastern Arkansas
Palmer Amaranth (Amaranthus Palmeri) accessions to glyphosate. Weed Technol 22:
408-413

174
Patzoldt WL, Hager AG, McCormick JS, Tranel PJ (2006) A codon deletion confers resistance
to herbicides inhibiting protoporphyrinogen oxidase. Proc Natl Acad Sci USA 103:
12329-12334
Riar DS, Norsworthy JK, Steckel LE, Stephenson DO, Eubank TW, Scott RC (2013) Assessment
of weed management practices and problem weeds in the midsouth Unites States-
soybean: A consultant's perspective. Weed Technol 27: 612-622
Rousonelos SL, Lee RM, Moreira MS, VanGessel MJ, Tranel PJ (2012) Characterization of a
common ragweed (Ambrosia artemisiifolia) population resistant to ALS- and PPO-
inhibiting herbicides. Weed Sci 60: 335-344
Salas RA, Burgos, NR, Tranel PJ, Singh S, Glasgow L, Scott RC, Nichols RL (2016) Resistance
to PPO-inhibiting herbicide in Palmer amaranth from Arkansas. Pest Manag Sci 72: 864-
869
Sales MA, Shivrain VK, Burgos NR, Kuk YI (2008) Amino acid substitutions in the acetolactate
synthase gene red rice (Oryza sativa) confer resistance to imazethapyr. Weed Sci 56: 485-
489
Shoup DE, Al-Khatib K, Peterson DE (2003) Common waterhemp (Amaranthus rudis)
resistance to protoporphyrinogen oxidase-inhibiting herbicides. Weed Sci 51: 145-150
Sosnoskie LM, Webster TM, Culpepper SA, Kichler J (2014) The biology and ecology of
Palmer amaranth: Implications for control.
http://extension.uga.edu/publications/files/pdf/C%201000_2.PDF. Accessed: December
15, 2016
Steckel LE (2007) The dioecious Amaranthus spp.: Here to stay. Weed Technol 21: 567-570
Thinglum KA, Riggins CW, Davis AS, Bradley KW, Al-Khatib K, Tranel PJ (2011) Wide
distribution of the waterhemp (Amaranthus tuberculatus) ΔG210 PPX2 mutation, which
confers resistance to PPO-inhibiting herbicides. Weed Sci 59: 22-27
Vencill W, Grey W, Culpepper S (2011) Resistance of weeds to herbicides. Pages 585-594 in
Kortekamp A, ed. Herbicides and Environment. Croatia: InTech
Watanabe N, Che FS, Iwano M, Takayama S, Yoshida S, Isogai A (2001) Dual targeting of
spinach protoporphyrinogen oxidase II to mitochondria and chloroplasts by alternative
use of two in-frame initiation codons. J Biol Chem 276: 20474-20481
Wuerffel RJ, Young JM, Lee RM, Tranel PJ, Lightfoot DA, Young BG (2015). Distribution of
the ΔG210 protoporphyrinogen oxidase mutation in Illinois waterhemp (Amaranthus
tuberculatus) and an improved molecular method for detection. Weed Sci 63: 839-845
Wuerffel RJ, Young JM, Matthews JL, Young BG (2015a) Characterization of PPO-inhibitor-
resistant waterhemp (Amaranthus tuberculatus) response to soil-applied PPO-inhibiting
herbicides. Weed Sci 63:511–521

175
Table 1. Number of Palmer amaranth accessions in
Arkansas, USA tested with foliar-applied fomesafen
at 263 g ha-1.
a Palmer amaranth samples were collected from
fields with history of glyphosate, glufosinate, or
PPO-inhibiting herbicide use. Sampling in 2015
was done specifically to survey fields with remnant
Palmer amaranth population after having been
sprayed with PPO herbicides.
Year collecteda Number of accessions
2008 25
2009 10
2011 16
2014 50
2015 23
Total 124

176
17
6
Table 2. Common, trade names, and manufacturers of herbicides used in the study.
Site of
action
Application Common name Trade Rate Company Company
website
g ai ha-1
PPO foliar acifluorfen Ultra Blazer 2SL 560 United Phosphorus, Inc.,
King of Prussia, PA 19406
www.upi-
usa.com
PPO foliar carfentrazone Aim 2EC 280 FMC Corporation,
Philadelphia, PA 19103
www.fmc.com
PPO foliar flumioxazin Valor SX 51 WDG 70.6 Valent USA Corporation,
Walnut Creek, CA
www.valent.com
PPO foliar fluthiacet-methyl Cadet 0.91EC 0.672 FMC Corporation,
Philadelphia, PA 19103
www.fmc.com
PPO foliar lactofen Cobra 2EC 224 Valent USA Corporation,
Walnut Creek, CA
www.valent.com
PPO foliar pyraflufen-ethyl ET 0.2EC 3.64 Nichino America,
Wilmington, DE 19808
www.nichino.net
PPO foliar saflufenacil Sharpen 4F 24.6 FMC Corporation,
Philadelphia, PA 19103
www.fmc.com
PPO soil fomesafen Reflex 2SL 280 Syngenta Crop Protection,
LLC, Greensboro, NC
27419
www.syngenta-
us.com
PPO soil flumioxazin Valor SX 51 WDG 70.6 Valent USA Corporation,
Walnut Creek, CA
www.valent.com
PPO soil saflufenacil Sharpen 4F 49.3 FMC Corporation,
Philadelphia, PA 19103
www.fmc.com
PPO soil sulfentrazone Spartan 4F 280 FMC Corporation,
Philadelphia, PA 19103
www.fmc.com

177
17
6
Table 2 (Cont.)
Site of
action
Application Common name Trade Rate Company Company
website
g ai ha-
1
PPO soil oxyfluorfen GoalTender 4E 280 Dow AgroSciences LLC,
Indianapolis, IN 46268
www.dowagro.c
om
Auxin foliar dicamba Clarity 4SL 280a BASF Corporation,
Research Triangle Park, NC
27709
https://www.basf
.com
Glutamine
synthetase
foliar glufosinate Liberty 280SL 549 Bayer CropScience LP,
Research Triangle Park, NC
27709
www.bayer.com
EPSPS foliar glyphosate Roundup PowerMAX
4.5SL
870a Monsanto Company, St.
Louis, MO 63167
www.monsanto.
com
ALS foliar trifloxysulfuron Envoke 75 7.84 Syngenta Crop Protection,
LLC, Greensboro, NC
27419
www.syngenta-
us.com
a g ae ha-1
177

178
178
Table 3. Response of fomesafen-resistant Palmer amaranth accessions to the recommended rate of various foliar-applied PPO
herbicides.
Mortalityb
Accessiona Acifluorfen Carfentrazone Flumioxazin Fluthiacet Lactofen Pyraflufen Saflufenacil
---------------------------------------------------------------%----------------------------------------------------
14CRI-C 100c (100)d 84 (99) 100 (100) 71f 93 (95) 94f 98 (100)
14MIS-H 55 (97) 67 (81) 66 (75) 18 42 (73) 37 79 (100)
15CLA-A 38 (64) 48 (45) 55 (44) 13 23 (66) 29 50 (98)
15CRI-A 68 (94) 54 (55) 74 (90) 39 49 (72) 53 73 (100)
15CRI-C 91 (100) 74 (87) 100 (100) 48 67 (92) 73 92 (100)
15CRI-D 72 (98) 60 (66) 91 (98) 19 44 (75) 60 93 (100)
15GRE-A 31 (78) 66 (72) 48 (81) 21 21 (29) 35 38 (99)
15IND-A 70 (94) 63 (74) 87 (90) 44 70 (79) 64 78 (100)
15LAW-C 73 (93) 58 (84) 81 (98) 33 49 (80) 57 91 (99)
15MIS-D 83 (99) 63 (80) 80 (60) 31 45 (77) 34 89 (100)
15MIS-E 73 (86) 69 (66) 79 (91) 30 54 (78) 54 95 (100)
15MIS-F 60 (93) 67 (67) 68 (89) 21 39 (69) 43 84 (100)

179
179
Table 3 (Cont.)
Mortalityb
Accessiona Acifluorfen Carfentrazone Flumioxazin Fluthiacet Lactofen Pyraflufen Saflufenacil
---------------------------------------------------------------%----------------------------------------------------
SSf 100 (100) 100 (100) 100 (100) 100 100 (100) 100 100 (100)
LSD0.05g 14 (15) 19 (19) 15 (19) 23 16 (23) 22 12
a Accessions were collected between 2014 and 2015 in Arkansas, USA. Accessions were confirmed resistant to foliar-
applied fomesafen (263 g ha-1) except for the susceptible standard (SS).
b Mortality ratings from acifluorfen, carfentrazone, flumioxazin, lactofen, and saflufenacil treatments were different across
runs, thus data were analyzed separately. Carfentrazone (280 g ha-1), pyraflufen-ethyl (3.64 g ha-1), and flumioxazin (70.6
g ha-1) treatments included 0.25% v/v non-ionic surfactant (NIS). Acifluorfen at 560 g ha-1 included 0.125% v/v NIS.
Lactofen (224 g ha-1) was sprayed with 0.5% v/v crop oil concentrate. Saflufenacil (24.6 g ha-1) was applied with 1%
methylated seed oil and 1% ammonium sulfate.
c Values obtained from the first run of experiment. Herbicides were applied to 10-cm tall seedlings.
d Values obtained from the second run of experiment. Herbicides were applied to 5- to 8-cm tall seedlings.
eAveraged mortality across two runs. Mortality ratings across runs were similar in pyraflufen-ethyl and fluthiacet-methyl treatments,
thus data were combined.
f SS = sensitive standard population
g Fisher’s protected LSD to compare accessions within each herbicide
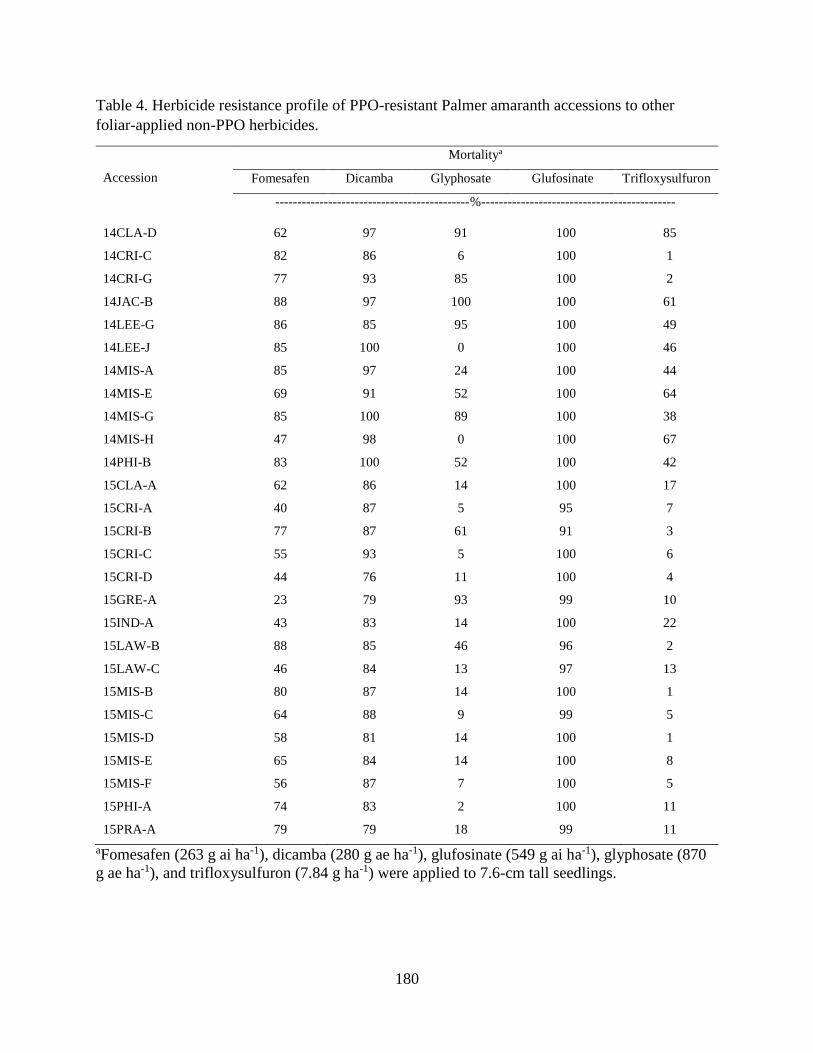
180
Table 4. Herbicide resistance profile of PPO-resistant Palmer amaranth accessions to other
foliar-applied non-PPO herbicides.
Accession
Mortalitya
Fomesafen Dicamba Glyphosate Glufosinate Trifloxysulfuron
--------------------------------------------%--------------------------------------------
14CLA-D 62 97 91 100 85
14CRI-C 82 86 6 100 1
14CRI-G 77 93 85 100 2
14JAC-B 88 97 100 100 61
14LEE-G 86 85 95 100 49
14LEE-J 85 100 0 100 46
14MIS-A 85 97 24 100 44
14MIS-E 69 91 52 100 64
14MIS-G 85 100 89 100 38
14MIS-H 47 98 0 100 67
14PHI-B 83 100 52 100 42
15CLA-A 62 86 14 100 17
15CRI-A 40 87 5 95 7
15CRI-B 77 87 61 91 3
15CRI-C 55 93 5 100 6
15CRI-D 44 76 11 100 4
15GRE-A 23 79 93 99 10
15IND-A 43 83 14 100 22
15LAW-B 88 85 46 96 2
15LAW-C 46 84 13 97 13
15MIS-B 80 87 14 100 1
15MIS-C 64 88 9 99 5
15MIS-D 58 81 14 100 1
15MIS-E 65 84 14 100 8
15MIS-F 56 87 7 100 5
15PHI-A 74 83 2 100 11
15PRA-A 79 79 18 99 11
aFomesafen (263 g ai ha-1), dicamba (280 g ae ha-1), glufosinate (549 g ai ha-1), glyphosate (870
g ae ha-1), and trifloxysulfuron (7.84 g ha-1) were applied to 7.6-cm tall seedlings.

181
Table 5. GR50 values of PPO-resistant Palmer amaranth accessions in Arkansas.
Gly210 mutation carrier Accession GR50a R/S
g ai ha-1
Yes 14MIS-H 232
(148-315)b
15
Yes 15MIS-C 116
(80-153)
8
Yes 15MIS-E 141
(94-188)
9
No 14CRI-C 70
(43-97)
4
No 14CRI-G 153
(112-195)
10
No 15CRI-B 125
(85-164)
8
No 15PHI-A 51
(32-70)
3
No SS 15
(10-21)
1
aGR50: dose of herbicide required to cause 50% biomass reduction bValues in parenthesis indicate 95% confidence intervals.

182
Figure 1. Variability in response to foliar-applied fomesafen (263 g ha-1) among Palmer
amaranth accessions collected between 2008 and 2015.Box plot shows median values (horizontal
line inside the box), mean values (marked X), first and third quartile values (box-outlines),
minimum and maximum values (whiskers), and outlier values (open circles).

183
Figure 2. Frequency of fomesafen-resistant plants in Palmer amaranth accessions collected in
2015 from Arkansas.Fomesafen at 263 g ha-1 was applied with 0.5% non-ionic surfactant to 7.6-
cm tall seedlings. Survivors are categorized based on visible injury.

184
Figure 3. Hierarchal clustering of 2014 and 2015 Palmer amaranth accessions treated with foliar-
applied fomesafen at 263 g ha-1.The herbicide was applied with 0.5% non-ionic surfactant to 7.6-
cm tall seedlings. Cluster 1 = most sensitive to fomesafen (50 accessions), Cluster 2 = resistant
to fomesafen with majority of survivors incurred >60% injury (19 accessions), Cluster 3 =
resistant to fomesafen, majority of survivors sustained 11-30% injury (2 accessions), Cluster 4 =
resistant to fomesafen, majority of survivors incurred 0-10% injury (3 accessions).
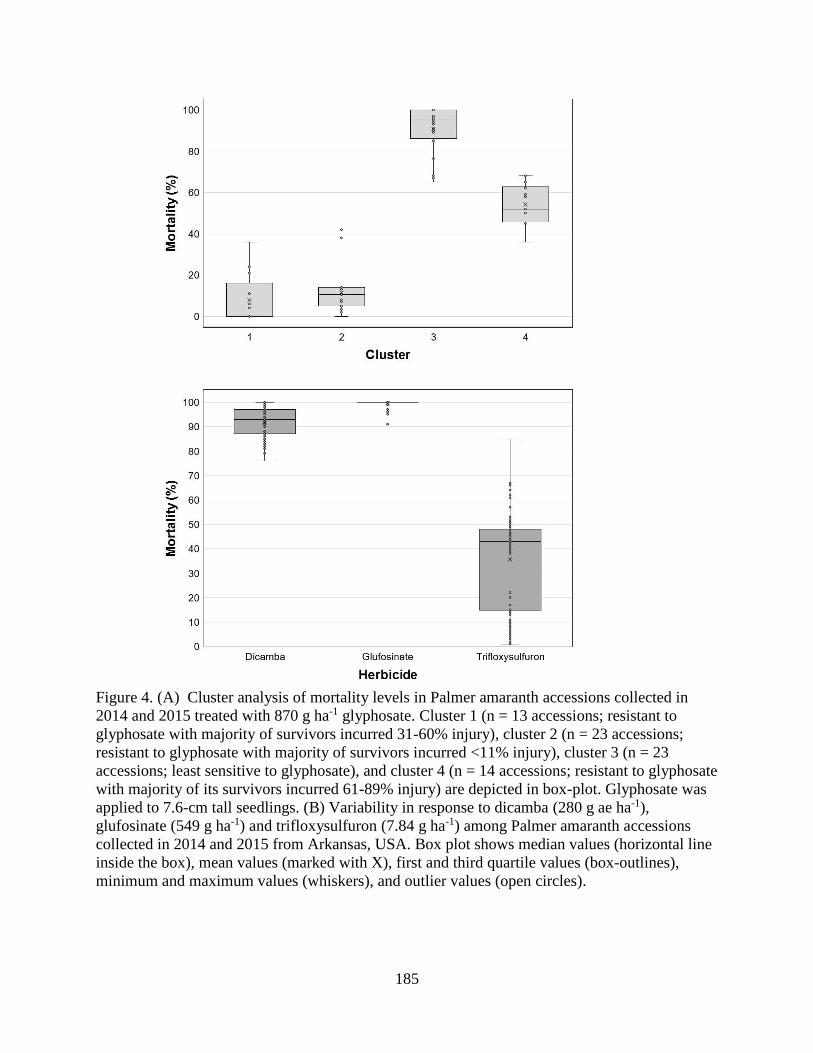
185
Figure 4. (A) Cluster analysis of mortality levels in Palmer amaranth accessions collected in
2014 and 2015 treated with 870 g ha-1 glyphosate. Cluster 1 (n = 13 accessions; resistant to
glyphosate with majority of survivors incurred 31-60% injury), cluster 2 (n = 23 accessions;
resistant to glyphosate with majority of survivors incurred <11% injury), cluster 3 (n = 23
accessions; least sensitive to glyphosate), and cluster 4 (n = 14 accessions; resistant to glyphosate
with majority of its survivors incurred 61-89% injury) are depicted in box-plot. Glyphosate was
applied to 7.6-cm tall seedlings. (B) Variability in response to dicamba (280 g ae ha-1),
glufosinate (549 g ha-1) and trifloxysulfuron (7.84 g ha-1) among Palmer amaranth accessions
collected in 2014 and 2015 from Arkansas, USA. Box plot shows median values (horizontal line
inside the box), mean values (marked with X), first and third quartile values (box-outlines),
minimum and maximum values (whiskers), and outlier values (open circles).

186
Figure 5. Herbicide resistance profiles of Palmer amaranth populations from Arkansas sampled
in 2014 and 2015.

187
Figure 13. Counties with confirmed PPO-resistant Palmer amaranth in Arkansas.Counties that
are shaded had at least one field-population with PPO-resistant Palmer amaranth biotypes
carrying the Gly210 deletion in the PPO gene.

188
Figure 7. Prevalence of PPO Gly210 mutation in PPO-resistant Palmer amaranth from Arkansas.
(A) Pie-chart displaying the percentage of PPO-resistant accessions that contained and lacked the
PPO Gly210 mutation. Black = all fomesafen survivors in these accessions contained the Gly210
mutation. White = all survivors in these accessions lacked the Gly210 mutation. Gray = survivors
in these accessions contained mixture of carriers and non-carriers of Gly210 mutation. (B)
Proportion of Gly210 mutation carriers in each PPO-resistant accession.

189
CONCLUSIONS
Majority of the Palmer amaranth accessions from Arkansas were sensitive to glufosinate
but resistant to trifloxysulfuron and glyphosate. Increased expression of GST and CYPA219
genes were associated with differential tolerance to glufosinate. The elevated expression of these
detoxification-related genes can get fixed in population with sustained selection pressure, leading
to evolution of resistance. Resistance to ALS inhibitors in a Palmer amaranth population from
Arkansas is attributed to ALS Ser653Asn mutation and elevated levels of CYP81B and GSTF10
genes. Induced expression of these non-target genes endow resistance to various ALS herbicide
classes.
Palmer amaranth in Arkansas had evolved resistance to PPO-inhibiting herbicides. PPO-
resistant Palmer amaranth biotype was first detected in a population collected in 2011. (4) Since
then, the number of PPO-resistant populations increased in at least 13 counties in Arkansas in
2015. Fomesafen-resistant populations were generally resistant to trifloxysulfuron and
glyphosate and were also cross-resistant to other PPO-inhibiting herbicides such as
carfentrazone, flumioxazin, and lactofen. The PPO Gly210 deletion is the prevalent (55%)
resistance mechanism among PPO-resistant Palmer amaranth populations from Arkansas. Plants
carrying PPO Gly210 deletion or Arg128Gly mutations had similar level of resistance to
fomesafen. Palmer amaranth has multiple genetic weapons, within or outside of the target site,
which it can use to counteract the lethal effects of herbicides. Resistance to multiple modes of
action, coupled with target and non-target-site mechanisms, severely limits herbicide option for
Palmer amaranth control. With Palmer amaranth being an obligate crossing species, plants in a
population can accumulate multiple resistance mechanisms which would likely drive resistance
evolution faster and is therefore detrimental to chemical weed management.