nano-manipulation of actomyosin molecular motors in vitro: a new working principle
TRANSCRIPT

TIBS 1 8 - SEPTEMBER 1993
Nano-manipulation of acto'my: sin : molecular motors in vitro: a,,new
* "working principle
MUSCLE CONTRACTION and other bio- logical motions are propelled by an actomyosin motor driven by the chemi- cal energy of ATP hydrolysis. Despite numerous studies, the working prin- ciple of this molecular motor remains elusive. The underlying mechanism is not as simple as that expected from analogy with man-made machines. Since a molecular motor is only nanometers in size and has a flexible structure, it is very prone to thermal agitation. Molecular motors can thus operate under the strong influence of this thermal noise, with a high ef- ficiency of chemo-mechanical energy conversion (40% at maximum; Ref. I). This is in sharp contrast to man-made machines which operate at energies much higher than the thermal noise. To elucidate the working principle of the molecular motor, it is essential to resolve the intrinsic characteristics of the molecular machine 2.
Until recently, the mechanism of motion underlying molecular motors was studied using muscle fibers, or sus- pensions of purlfied motor proteins. These systems, however, are too com- plex to provide unambiguous infor- mation about the elementary process of energy transduction by individual molecular motors. To alleviate this problem, new techniques were needed that could directly probe the elementary process at the molecular level. Several kinds of in vitro motility models using purified motor proteins have now been designed (see Ref. 3 for a review). A model that allows the direct obser- vation of single actin filaments labeled with fluorescent phalloidin by optical microscopy 4, as well as the motion pro- duced by myosin or its subfragments bound to a substrate s,6, has received a great deal of attention. Furthermore, a very subtle technique for manipulating a single actin filament by a fine glass needle has been developed ~, which has allowed the measurement of both the motion and force of individual motors
Toshio Yanagida, Yoshie Harada and Akihiko Ishijima
T. ¥ana~da is at the Department of Biophysical Engineering, Osaka University, Toyonaka, Osaka, Japan. ¥. Harada and A. IshlJima are at the ERATO BIO-MOTRON Project, Senba-Higashi 2414, Mino, Osaka, Japan.
Techniques have been recently developed that allow the direct obser- vation of single actin filaments and their manipulation, using glass microneedles, in the nanometer range. Further development of these tech- niques has made possible the detection of subpiconewton-level forces of individual myosin heads. This in vitro motility model is sensitive in the submillisecond range and has allowed us to determine the force gener- ation of an actomyosin motor directly at the molecular level. The results have led to a new conceptual framework for chemo-mechanical energy transduction in the molecular motor.
in vitro at very high resolution (sub- nanometer and subpiconewton, respect- ively; Ref. 8). These new in vitro motility assays have greatly enhanced the progress of research on the actomyosin molecular motor. In this review, we will survey high-resolution measurements of both the motion and force of the molecular motor and introduce a new conceptual framework for chemo- mechanical energy transduction.
Direct observation of motor proteins and an In vitro motility model
The first step when studying the el- ementary process of chemo-mechanical energy transduction is to directly observe the motion of a molecular motor in solution by optical micros- copy. The resolution of conventional optical microscopes (-0.2 pin), however, is too low to observe single protein molecules, or even a small assembly of motors. Thus a great deal of effort has been made to increase the resolution of the experimental system. One method is to observe the scattered light from objects using a dark-field microscope equipped with a very strong illumi- nating source and a highly sensitive camera. This method has the advantage that objects can be observed without any type of labeling. However, the ob- jects observed need to be as large as a microtubule, about 20 nm in diameter 9. A
© 1993, Elsevier Science Publishers, (UK) 0968-0004/93/$06.00
similar resolution can be achieved if the contrast of the recording is greatly en- hanced by computer image processing I° (video-enhanced contrast method).
Fluorescence microscopy is useful when observing smaller objects n. The number of photons emitted from a single molecule of fluorescent dye can be observed by a fluorescence micro- scope equipped with conventional high- sensitivity detectors, such as a silicon intensified target (SIT) camera or an image intensifier 12, It is theoretically possible to observe single protein molecules labeled with fluorescent dyes, although it has yet to be proven in practice owing to strong photo- bleaching effects. We have demonstrated that single actin filaments labeled with phalloidin-dye complexes (which stabil- ize the filament structure of actin but do not affect the ability to produce motion with myosin 13) can be observed clearly and continuously by fluor- escence microscopy 4. This technique has allowed us to observe the motion of actin filaments during the interaction with myosin in the presence of ATP. Using this method, it has been shown that the bending motion of actin fila- ments is modulated by an interaction in solution with myosin subkagments (S-I and HMM) in the presence of ATP 4. However, the unidirectional motion of actin filaments, which is evident in
319

TIBS 18 - SEPTEMBER 1993
Box 1. Manipulation of motor proteins by optical tweezers The single-beam optical gradient field trap (optical tweezers) is a useful method to capture and manipulate small (25-100 pm) dielectric particles in solution 2s. Optical tweezers have recently attracted much attention in the biological field since with them cells and subcellular components can be easily manipulated in a non- invasive manner without significant radiation damage 28-3°. Further- more, the technique is simple. The figure shows the optical tweezers system for manipulating a single actin filament attached to a latex bead, using a microscope equipped with epifluorescence optics. A laser beam is focused on the specimen by an objective with a high numerical aperture. The scattering force, pointing in the direction of the incident light, pushes the bead down and the gradient force pulls it into the region of high light intensity, i.e. the focal spot. The bead is thus trapped just below the focal spot where both of the forces balance each other. As the laser beam used has a gaussian intensity distribution in its cross-section, the bead is securely trapped in the horizontal (X-Y) direction by the additional gradient force pulling it into the center of the beam. The trapping force depends on the intensity of the laser and the size of the particle. With a I pm diameter latex bead and 100 mW laser power, the force produced is several tens of piconewtons, which is large
To TV camera ~, Two-dimensional
Laser beam I I position sensor
High NA D M ~ p ~ = ~ ~ / j J objective rojection ~ - - ,~-,~ Bead D M / ~ of bead
~ ~ ' ~ mirrors
DM : Dichroic mirror Halogen lamp ~
enough to move the bead against the viscous drag in water at a velocity of 3700 pm s -1. The bead can be manipulated in two dimensions by moving the focal spot using the galvanometer mirrors. As the actin filament is too small to be trapped, one end of the actin filament, labeled with fluorescent phalloidin, is attached to the bead trapped in the movable beam (1 pm in diameter). The bead is coated with NEM-treated myosin (see text) and fluorescently labeled so the actin filament and the bead can be observed simultaneously under a fluorescence microscope. Thus, it is possible to manipulate actin filaments. When the other end of the actin filament is brought into contact with the myosin-coated surface, the force due to actomyosin interaction can be mea- sured. Since the light intensity distribution of the laser beam is gaussian, the trapping force increases proportionally with the deviation of the position of the center of the bead from the center of the laser beam. Therefore, the force due to actomyosin interaction can be determined by measuring this deviation. The deviation is measured by a two-dimensional position sensor with a space resolution in the nanometer range by essentially the same method as that shown in the text, which corresponds to a force resolution in the piconewton range.
intact muscle, was not observed in sol- ution. It has been shown that, when myosin or its subfragments were bound to the surface of a substrate, the actin moved In a unidirectional way along them with a velocity similar to that found in intact muscle 5,~,
Thus, it has been possible to observe the motion of molecular motors with the use of optical microscopy. This in vitro motility model has been widely used to test the functions of motor pro- teins, since it does not require any special skills or large-scale equipment.
Mlcromanipulatlon of an actin filament and force measurement of the molecular motor
Observation of the motion of actin can only provide limited information about the mechanism of force gener- ation. Recently, however, a technique for manipulating a single actin filament using a fine glass needle has been developed T. This technique enables one to exert a force on an actin filament, or to measure the force exerted on an actin filament when interacting with myosin. One end of an actin filament is caught by a glass micro-needle mounted on a mechanical manipulator; the other end is brought into contact with a glass sudace that has been coated with N-ethyl maleimide (NEM)-treated myosin in order to increase its affinity
32O
for actin. The actin filament is pulled by the force produced by the actomyosin interaction, bending the needle. The force is determined by measuring the degree of bending of the needle, as observed on a TV monitor. Using this method it is possible to measure a force of several plconewtons. Thus, the force as well as the motion of the actin fila- ment has been measured using this in vitro motility model.
Measurements of subnanometer motion and subplconewton force of the molecular motor
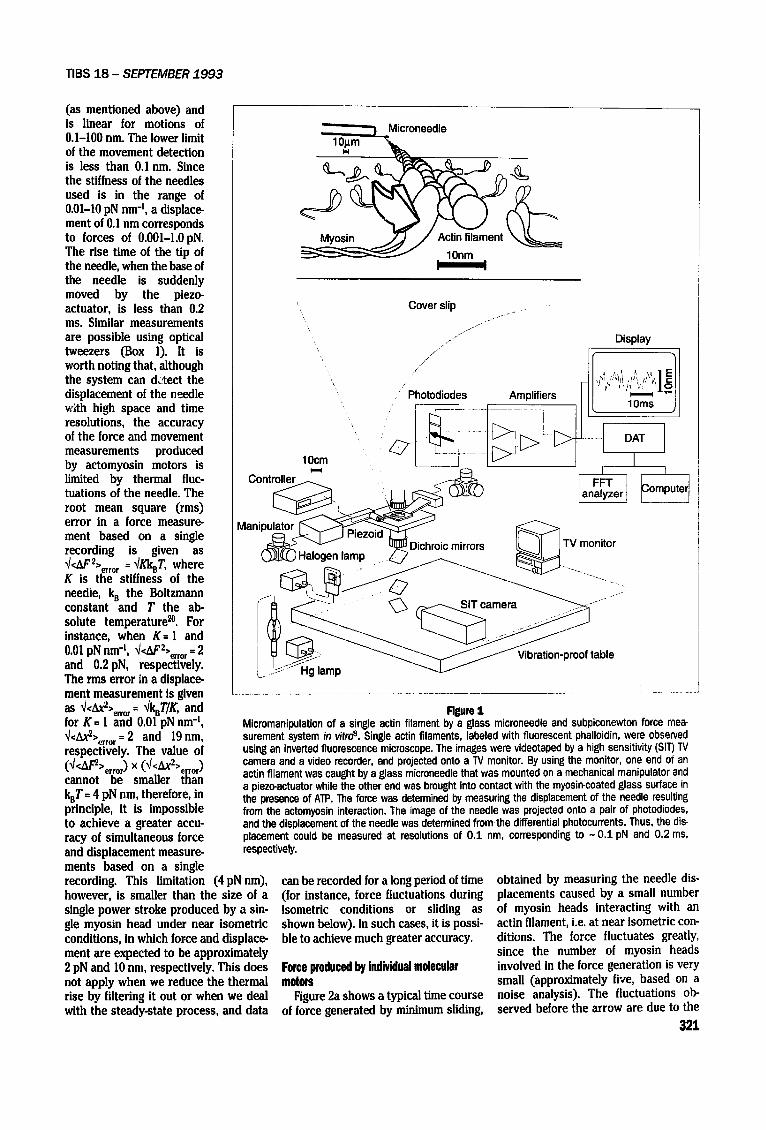
Another new technique that directly probes the process of the force gener- ation of individual molecular motors in vitro has been developed s, In order to minimize the deterioration of the time resolution resulting from the needle's mass and viscous drag in the solution, a finer glass needle, 50-70 pm in length and 0.2 pm in diameter, has been used. This fine glass needle was attached to a rigid holding rod mounted on a piezo- actuator and a mechanical manipulator that could move the needle over a range from less than 0.I nm to several tens of micrometers. The force was determined by measuring the displacement of the needle, as described above. In turn, the displacement was detected at very high time and space resolutions as follows.
The static position of the needle can- not be determined at a space resolution of less than about half of the wave- length of light owing to the diffraction limit. The displacement, however, is different :~-:8. For instance, if the image of the needle is projected onto a pair of photo-dlodes and the displacement Is determined by measuring the difference in the photo-currents of the photo- diodes, a displacement of less than I/I000 of the wavelength can be detected. This method is similar to that used in an atomic force microscope (AFM) to determine the position of the probe. The function of the pair of photo- diodes can be replaced by computer processing images of the needles, or small particles attached to objects, which have been recorded on video tape. The time resolution, however, is not as high 19 (several tens of milli- seconds).
The bending of the needle is meas- ured by the system shown in Fig. I. To increase the contrast, a small nickel particle of about 1 Bm in diameter is attached to the tip of the needle. An image of the particle is projected onto a pair of photo-diodes, and the displace- ment of the needle is obtained from the differential output of the photodetec- tor. The differential output is very sen- sitive to any movement of the particle
TIBS 18 - SEPTEMBER 1993
(as mentioned above) and is linear for motions of 0.1-100 nm. The lower limit of the movement detection is less than 0.I nm. Since the stiffness of the needles used is in the range of 0.01-I0 pN nm -t, a displace- ment of 0.I nm corresponds to forces of 0.001-I.0 pN. The rise time of the tip of the needle, when the base of the needle is suddenly moved by the piezo- actuator, is less than 0.2 ms. Similar measurements are possible using optical tweezers (Box I). It is worth noting that, although the system can d~tect the displacement of the needle with high space and time resolutions, the accuracy of the force and movement measurements produced by actomyosin motors is limited by thermal fluc- tuations of the needle. The root mean square (rms) error in a force measure- ment based on a single recording is given as ~/<Z~C'2>erro r = ~/KkBT, where K is the stiffness of the needle, k a the Boltzmann constant and T the ab- solute temperature 2°. For instance, when K= 1 and 0.01 pN nm -I, ~J<AF')%rro, = 2 and 0.2pN, respectively. The rms error in a displace- ment measurement is given as ~/<Ax2%,or = ~kBT/K, and for K = 1 and 0.01 pN nm -I, 'J<AX2%rro r = 2 and 19 nm, respectively. The value of (~<~J~Z>error) X (~<~X2>error) cannot be smaller than kBT = 4 pN nm, therefore, in principle, it is impossible to achieve a greater accu- racy of simultaneous force and displacement measure- ments based on a single recording. This limitation (4pN nm), however, is smaller than the size of a single power stroke produced by a sin- gle myosin head under near isometric conditions, in which force and displace- ment are expected to be approximately 2 pN and I0 nm, respectively. This does not apply when we reduce the thermal rise by filtering it out or when we deal with the steady-state process, and data
Microneedle
L
lOcm
Cover slip ....
/ / / /.
Photodiodes Amplifiers
@
Controller ~ . ~
anip,, or ~ ~ Dichroic mirrors LQ II I TV monitor
/ ?_ ~ : ' - ' ~ - ~ ~ ~ Vibration-prooftable L. =S : IHg lamp
Display
,,,q A ,, TEl .f:~.l"t( . vl ;.'~ , \ /El l
j ,Ores Jl
I
Rgure 1 Micromanipulation of a single actin filament by a glass microneedle and subpiconewton force mea- surement system in vitro s. Single actin filaments, labeled with fluorescent phalloidin, were observed using an inverted fluorescence microscope. The images were videotaped by a high sensitivity (SIT) TV camera and a video recorder, and projected onto a TV monitor. By using the monitor, one end of an actin filament was caught by a glass microneedle that was mounted on a mechanical manipulator and a piezo-actuator while the other end was brought into contact with the myosin-coated glass surface in the presence of ATP. The force was determined by measuring the displacement of the needle resulting from the actomyosin interaction. The image of the needle was projected onto a pair of photodiodes, and the displacement of the needle was determined from the differential photocurrants. Thus, the dis- placement could be measured at resolutions of 0.1 nm, corresponding to ~ 0.1 pN and 0.2 ms, respectively.
can be recorded for a long period of time (for instance, force fluctuations during isometric conditions or sliding as shown below). In such cases, it is possv ble to achieve much greater accuracy.
Force produced by individual molecular motors
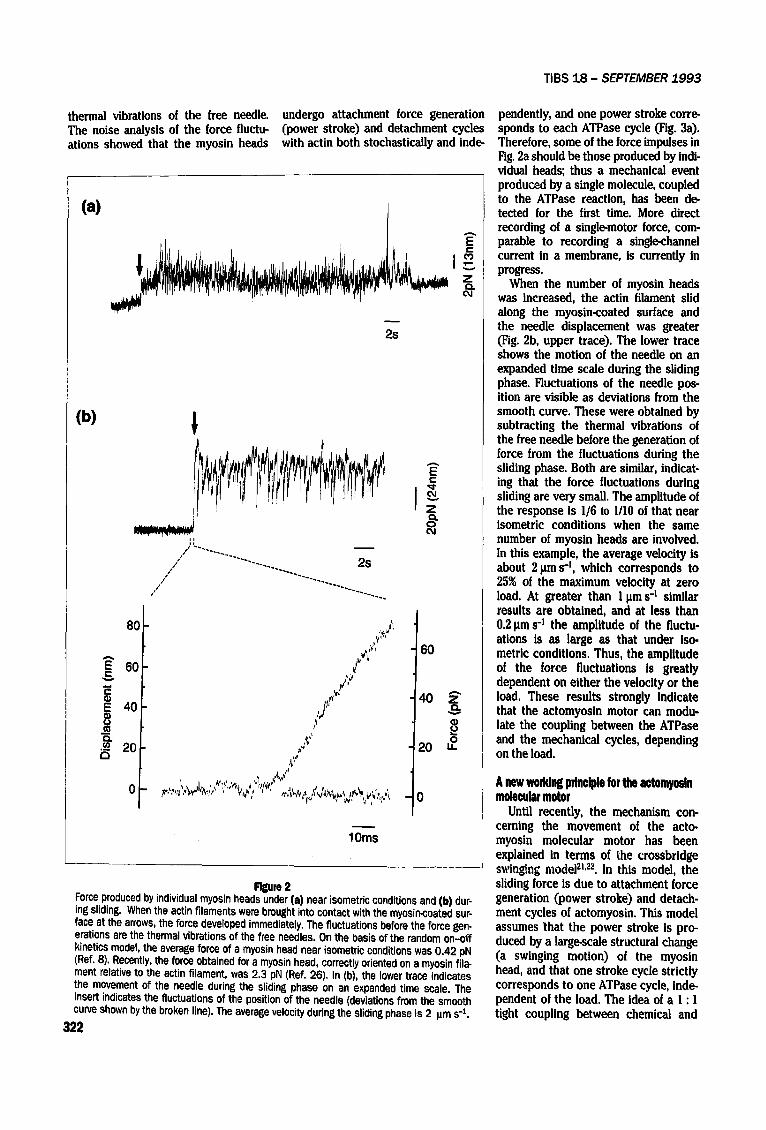
Figure 2a shows a typical time course of force generated by minimum sliding,
obtained by measuring the needle dis- placements caused by a small number of myosin heads interacting with an actin filament, i.e. at near isometric con- ditions. The force fluctuates greatly, since the number of myosin heads involved in the force generation is very small (approximately five, based on a noise analysis). The fluctuations ob- served before the arrow are due to the
321
TIBS 18 - SEPTEMBER 1993
thermal vibrations of the free needle. The noise analysis of the force fluctu- ations showed that the myosin heads
undergo attachment force generation (power stroke.) and detachment cycles with actin both stochastically and inde-
(a)
2s
(b) |
80
E" 60
E 40
}
_
p ~ '~'7~'
//;
lOms
E c
z o
60
40 Q,
20 ,,o
R~m 2 Force produced by individual myosin heads under (a) near isometric conditions and (b) dur- ing sliding. When the actin filaments were brought into contact with the myosin-coated sur- face at the arrows, the force developed immediately. The fluctuations before the force gen- erations are the thermal vibrations of the free needles. On the basis of the random on-off kinetics model, the average force of a myosin head near isometric conditions was 0.42 pN (Ref. 8), Recently, the force obtained for a myosin head, correctly oriented on a myosin fila- merit relative to the actin filament, was 2.3 pN (Ref. 26). In (b), the lower trace indicates the movement of the needle during the sliding phase on an expanded time scale. The insert indicates the fluctuations of the position of the needle (deviations from the smooth curve shown by the broken line). The average velocity during the sliding phase is 2 pms -1.
322
pendently, and one power stroke corre- sponds to each ATPase cycle (Fig. 3a). Therefore, some of the force impulses in Fig. 2a should be those produced by indi- vidual heads; thus a mechanical event produced by a single molecule, coupled to the ATPase reaction, has been de- tected for the first time. More direct recording of a single-motor force, com- parable to recording a single-channel current in a membrane, is currently in progress.
When the number of myosin heads was increased, the actin filament slid along the myosin-coated surface and the needle displacement was greater (Fig. 2b, upper trace). The lower trace shows the motion of the needle on an expanded time scale during the sliding phase. Fluctuations of the needle pos- ition are visible as deviations from the smooth curve. These were obtained by subtracting the thermal vibrations of the free needle before the generation of force from the fluctuations during the sliding phase. Both are similar, indicat- ing that the force fluctuations during sliding are very small. The amplitude of the response is 1/6 to 1/10 of that near isometric conditions when the same number of myosin heads are involved. In this example, the average velocity is about 2 Ixms 4, which corresponds to 25% of the maximum velocity at zero load. At greater than I l~m s-' similar results are obtained, and at less than 0.2 pms -z the amplitude of the fluctu- ations is as large as that under iso- metric conditions. Thus, the amplitude of the force fluctuations Is greatly dependent on either the velocity or the load. These results strongly Indicate that the actomyosin motor can modu- late the coupling between the ATPase and the mechanical cycles, depending on the load.
A new wmkl~ principle for the aotomyosln molecular motor
Until recently, the mechanism con- cerning the movement of the acto- myosin molecular motor has been explained in terms of the crossbridge swingtng model 2z,22. ]n this model, the sliding force is due to attachment force generation (power stroke) and detach- ment cycles of actomyosin. This model assumes 'that the power stroke is pro- duced by a large-scale structural change (a swinging motion) of the myosin head, and that one stroke cycle strictly corresponds to one ATPase cycle, inde- pendent of the load. The idea of a l : 1 tight coupling between chemical and
TIBS 18 - SEPTEMBER 1 9 9 3
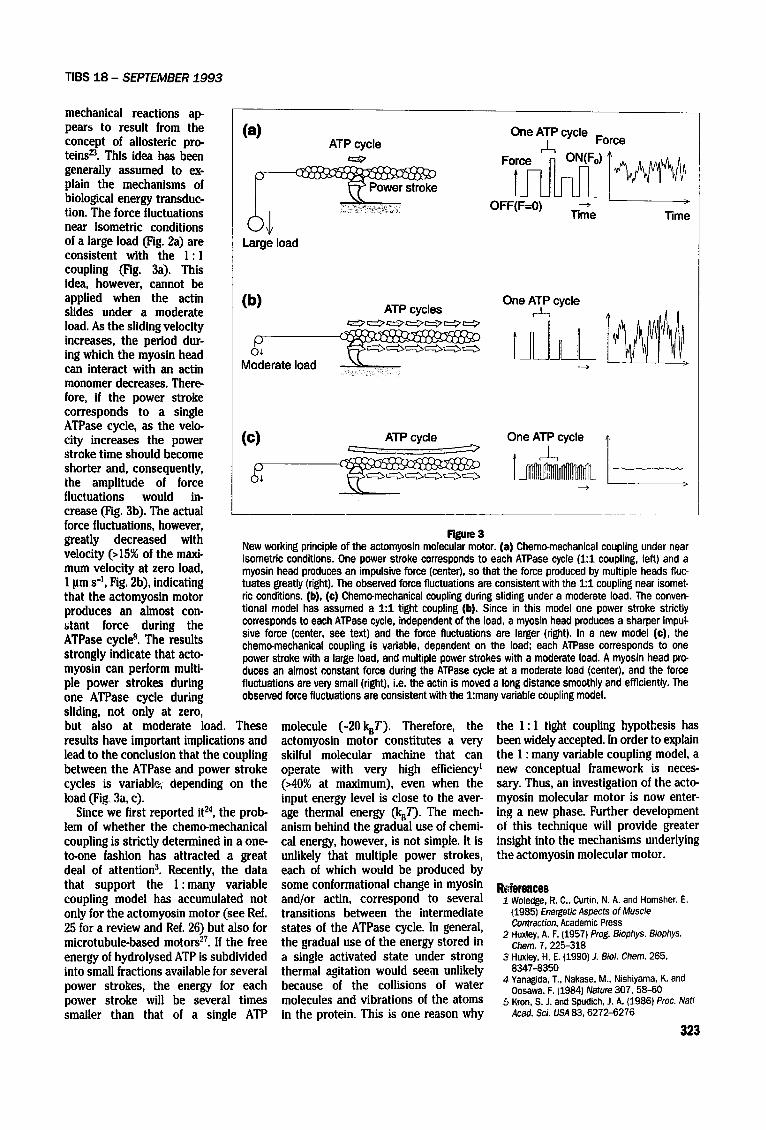
mechanical reactions ap- pears to result from the concept of allosteric pro- teins 23. This idea has been generally assumed to ex- plain the mechanisms of biological energy transduc- tion. The force fluctuations near isometric conditions of a large load (Fig. 2a) are consistent with the 1 : 1 coupling (Fig. 3a). This idea, however, cannot be applied when the actin (b) slides under a moderate load. As the sliding velocity increases, the period dur- ing which the myosin head can interact with an actin monomer decreases. There- fore, if the power stroke corresponds to a single ATPase cycle, as the velo- city increases the power (C) stroke time should become shorter and, consequently, p the amplitude of force o+
fluctuations would in- crease (Fig. 3b). The actual force fluctuations, however, greatly decreased with velocity (>15% of the maxi- mum velocity at zero load, 1 pms -1, Fig. 2b), indicating that the actomyosin motor produces an almost con- atant force during the ATPase cycle 8. The results strongly indicate that acto- myosin can perform multi- ple power strokes during one ATPase cycle during sliding, not only at zero, but also at moderate load. These results have important implications and lead to the conclusion that the coupling between the ATPase and power stroke cycles is variablr+; depending on the load (Fig 3a, c).
Since we first reported it 24, the prob- lem of whether the chemo-mechanical coupling is strictly determined in a one- to-one fashion has attracted a great deal of attention 3. Recently, the data that support the 1 :many variable coupling model has accumulated not only for the actomyosin motor (see Ref. 25 for a review and Ref. 26) but also for microtubule-based motors 27. If the free energy of hydrolysed ATP is subdivided into small fractions available for several power strokes, the energy for each power stroke will be several times smaller than that of a single ATP
(a) ATP cycle
Large load
One ATP cycle Force
F i r ~ ) l ~ / ~ ~
OFF(F=O) --> "rime "time
ATP cycles
Moderate load
One ATP cycle
~ _ _ A T P c y c l e One ATP cycle !
Rgure 3 New working principle of the actomyosin molecular motor. (a) Chemo-mechanical coupling under near isometric conditions. One power stroke corresponds to each ATPase cycle (1:1 coupling, left) and a myosin head produces an impulsive force (center), so that the force produced by multiple heads fluc- tuates greatly (right). The observed force fluctuations are consistent with the 1:1 coupling near isomet- ric conditions. (b), (e) Chemo-mechanical coupling during sliding under a moderate load. The conven- tional model has assumed a 1:1 tight coupling (b). Since in this model one power stroke strictly corresponds to each ATPase cycle, independent of the load, a myosin head produces a sharper impul- sire force (center, see text) and the force fluctuations are larger (right). In a new model (©), the chemo-mechanical coupling is variable, dependent on the load; each ATPase corresponds to one power stroke with a large load, and multiple power strokes with a moderate load. A myosin head pro- duces an almost constant force during the ATPase cycle at a moderate load (center), and the force fluctuations are veff small (right), i.e. the actin is moved a long distance smoothly and efficiently. The observed force fluctuations are consistent with the 1:many variable coupling model.
molecule (-20kBT). Therefore, the actomyosin motor constitutes a very skilful molecular machine that can operate with very high efficiency = (>40% at maximum), even when the input energy level is close to the aver- age thermal energy (kBT). The mech- anism behind the gradual use of chemi- cal energy, however, is not simple, it is unlikely that multiple power strokes, each of which would be produced by some conformational change in myosin and/or actin, correspond to several transitions between the intermediate states of the ATPase cycle. In general, the gradual use of the energy stored in a single activated state under strong thermal agitation would seem unlikely because of the collisions of water molecules and vibrations of the atoms in the protein. This is one reason why
the I :1 tight coupling hypothesis has been widely accepted. In order to explain the 1 : many variable coupling model, a new conceptual framework is neces- sary. Thus, an investigation of the acto- myosin molecular motor is now enter- ing a new phase. Further development of this technique will provide greater insight into the mechanisms underlying the actomyosin molecular motor.
R~.ferences i Woledge, R. C.. Curtin, N. A. and Homsher, E.
~1985) Energetic Aspects of Muscle Contraction, Academic Press
2 Huxley, A. F. (1957) Prog. Biophys. Biophys. Chem. 7,225-318
3 Huxley, H. E. (1990) J. Biol. Chem. 265, 8347-8350
4 Yanagida, T., Nakase, M.. Nishiyama. K. and Oosawa. F. (1984) Nature 307, 58-60
5 Kron, S. J. and Spudich, J. A. (1986) Proc. Nat/ Acad. Sci. USA 83, 6272-6276
323

TIBS 18 - SEPTEMBER 1993
6 Toyoshima, ¥. Y. et aL (1987) Nature 328, 536-539
7 Kishino, A. and Yanagida, T. (1988) Nature 334, 74-76
8 Ishijima, A., Doi, T., Sakurada, K. and Yanagida, T. (1991) Nature 352, 301-306
9 Hotani, H. (1976) J. MoL BioL 106, 151-166 10 Inoue, S. (1981) J. Cell Biol. 89, 346-356 11 Morikawa, K. and Yanagida, M. (1981)
J. Biochem. 89, 693-696 12 Harada, Y. and Yanagida, T. (1988) Cell Mot#.
Cytoskel. 10, 71-76 13 Wieland, T. and Faulstich, H. (1978) CRC Crit.
Biochem. 5, 185-260 14 Borejdo, J. and Morales, M. F. (1977) Biophys.
J. 20, 315-334 15 Iwazurni, T. (1987) Am. J. Physiol. 252, 253-262 16 Crawford, A. C. and Fettiplace, R. (1985)
J. PhysioL 364, 359-379 17 Kamimura, S. and Kamiya, R. (1989) Nature
340, 476-478 18 Denk, W., Webb, W. W. and Hudspeth, A. J.
(1989) Prec. Natl Acad. Sci. USA 86, 5371-5375 19 Sheetz, M. P., Turney, S., Qian, H. and Elson,
E. L. (1989) Nature 340, 284-288 20 Kittel, C. (1964) Elementary Statistical Physics,
Chap. 30, John Wiley & Sons 21 Huxley, H. E. (1969) Science 164, 1356-1366 22 Huxley, A. F. and Simmons, R. M. (1971) Nature
233, 533-538
23 Ebashi, S. (1991) Annu, Rev. Physiol. 53,1-16 24 Yanagida, T,, Arata, T. and Oosawa, F. (1985)
Nature 316, 366-369 25 Burton, K. (1992) J. Muscle Res. Cell Mot#. 13,
590-607 26 Yanagida, T. (1993) Abs. 32nd Int. Physiol.
Congress 7.4/0, 10 27 Taylor, E. W. (1993) Nature 361, 115-116. 28 Askin, A. and Dziedzic, J. M. (1987) Science
235, 1517-1520 29 Block, S. M. (1990) in Noninvasive Techniques
in Cell Biology(Fosbett and Grinstein, eds), pp. 375-402, Wiley-Liss
30 Kuo, S. C. and Sheetz, M. P. (1992) Trends Cell 8ioL 2, 116-118
TALKING POINT
• Tetanus and botulism
neurotoxins: a new group of zinc
proteases
TETANUS AND BOTULINUM neurotox- ins are the most potent toxins known (mouse lethal dose <0.I ng kg-X). They are released by bacteria of the genus Clostridium as a single polypeptide chain of 150 kDa, later cleaved to gener- ate two disulfide-linked fragments. The heavy chain (H, I00 kDa) is involved in cell binding and penetration, while the light chain (L, 50 kDa) is responsible for the intracellular activity ~,~.
The tremendous potency of these toxins is primarily due to their absolute neurospeclficity. They bind to the neuromuscular junction and are intern- alized inside vesicles 3. Botullnum neurotoxln (BoNT, seven different serotypes: A-G) penetrates into the cytosol and blocks the release of acetyl- choline, thus causing a flaccid paral- ysis. By contrast, tetanus neurotoxin (TeTx) migrates retroaxonally (in the reverse direction to nerve impu]ses) and, by transcytosis, reaches the spinal inhibitory interneurons, where it blocks neurotransmitter release with a conse- quent spastic paralysis R. Despite the opposing clinical symptoms of botulism and tetanus, these toxins act in a simi- lar way at the cellular level. Since TeTx and BoNT are entirely responsible for the diseases tetanus and botulism, respectively, understanding their intra- cellular activity would also lead to the description of the molecular patho- genesis of these diseases. At the same time, we would expect to understand better the process of neuroexocytosis.
c. Monteeu©©o and G. S©hla¥o are at the Department of Biomedical Sciences of the University of Padova, Via Trieste 75 1-35121, Padova, Italy.
324
The active forms of tetanus :,nd botulinum neurotoxins, released from the precursor molecule by specific proteolysis and reduction, block the release of neurotransmitters via a Zn2*-dependent protease activity. VAMP/synaptobrevin, an integral membrane protein of the synaptic ves- icles, is cleaved at a single site by tetanus and botuliaum B, D and F neurotoxins. The unique sequence, mechanism of activation and site of activity of clostridial neurotoxins mark them out as an independent group of Zn2*-endopeptidases.
Tetanus and botulinum neurotoxins are Zn a÷ proteins
The enormous potency of these neurotoxins, and analogy with other bacterial toxins with intracellular ac- tivity, suggests that TeTx and BoNT possess or induce a catalytic activity inside cells.
Their amino acid sequences show a limited degree of similarity, concen- trated mainly in the L chain. The seg- ment with the closest homology is located in the central part and contains the Zn2*-binding motif, HEXXH, that is a characteristic feature of Zn 2~ endopepti- dases 4"~2, Atomic absorption measure- ments showed that indeed TeTx and BoNT/A, B, C, E and F contain approxi- mately one Zn 2~ ion bound to the L chainS-7. As with other Zn ~ proteases, the Zn 2÷ ion can be removed with EDTA or ortho-phenanthroline, thus forming
an apotoxin, and it can be replaced by incubation in ZnZ'-containing buffers s'7.
The three-dimensional structures of four Zn 2÷ endopeptidases have been solved by X-ray diffraction. In thermo- lysin 13, neutral protease t4 and Pseudo- monas aeruginosa elastase L~, the Zn 2~ ion is at the center of a tetrahedron, co- ordinated by the two histidine residues of the motif, by one water molecule that is bound to the glutamate residue of the motif, and by the carboxylate group of another glutamic acid residue. By con- trast, the recently reported structure of astacin ~6, a Zn 2~ protease of the crayfish digestive tract, shows the Zn z~ ion penta-coordinated by the two histidine residues, the water molecule that is bound to the glutamate residue of the motif, an additional histidine residue, and a tyrosine residue. On the basis of these findings, Zn 2÷ endopeptidases
© 1993, Elsevier Science Publishers, (UK) 0968-0004/93/$06.00