nadph-diaphorase and cytosolic urea cycle enzymes in the rat spinal cord
TRANSCRIPT

NADPH-Diaphorase and Cytosolic UreaCycle Enzymes in the Rat Spinal Cord
HIROYUKI NAKAMURA*Department of Anatomy, Faculty of Medicine, Kyushu University, Fukuoka 812, Japan
Precursory Research for Embryonic Science and Technology (PRESTO),Japan Science and Technology Corporation (JST), Kawaguch, Saitoma 332, Japan
ABSTRACTNitric oxide synthase (NOS), argininosuccinate synthetase (ASS), and argininosuccinate
lyase (ASL) compose a cyclic pathway to form nitric oxide (NO). These enzymes, however, arelocalized differentially in most regions of the brain. To find out whether NOS, ASS, and ASLare colocalized in neurons of the spinal cord, we examined the distribution of these enzymes byusing a double-labeling procedure combining fluorescent immunohistochemistry with anassay for reduced nicotinamide adenine dinucleotide phosphate-diaphorase (NADPH-d).Results indicate that neurons in the dorsal horn, the intermediolateral nucleus, and thecentral canal region were NADPH-d active (1) and NOS-,ASS-, andASL-like immunoreactive(-LI). In laminae II and III of the dorsal horn, some NADPH-d (1) neurons were ASL-LI(8–30%) but only a few were ASS-LI (0.5–7%). In the nucleus intermediolateralis, a largeportion of NADPH-d (1) neurons were ASL-LI (30–60%), whereas only a small portion ofNADPH-d (1) neurons were ASS-LI (10–20%). In the central canal region, some NADPH-d(1) neurons were ASL-LI (15–40%), and a few NADPH-d (1) neurons were ASS-LI (3–16%).Thus, the results suggest that, in the nucleus intermediolateralis and the central canal region,NOS, ASS, andASL are colocalized and form a cyclic pathway to produce NO, whereas, in thedorsal horn, these enzymes are more characteristically localized in different neurons, which maytransport the substrates intercellularly. J. Comp. Neurol. 385:616–626, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: nitric oxide synthase; argininosuccinate; arginine; nucleus intermediolateralis;
dorsal horn
Nitric oxide (NO) is a neurotransmitter and modulatorin the nervous system (Bredt and Snyder, 1989; Dawsonand Snyder, 1994), which is produced by nitric oxidesynthase (NOS) from arginine, with stoichiometric produc-tion of citrulline (Bredt and Snyder, 1990). With thecatalysis of argininosuccinate synthetase (ASS), citrullinethen reacts with aspartate to form argininosuccinate.Argininosuccinate is in turn cleaved by argininosuccinatelyase (ASL) to form fumarate and arginine (Fig. 1). Thus,there exist in the brain three cytosolic members of the fiveurea cycle enzymes: ASS,ASL, and arginase (Ratner et al.,1960; Jackson et al., 1986). NOS completes the urea cyclein the brain and, together with ASS and ASL, forms acyclic enzymatic pathway for NO production.The distribution of NOS in the brain has been deter-
mined in the cat (Mizukawa et al., 1989) and rat (Bredt etal., 1990, 1991a; Arnt-Ramos et al., 1992; Vincent andKimura, 1992) by using reduced nicotinamide adeninedinucleotide phosphate-diaphorase (NADPH-d) histochem-istry or immunohistochemistry with an antibody to NOS.In the spinal cord, NOS is localized in the nucleus interme-diolateralis and the central canal region (Bredt et al.,
1991a; Anderson, 1992; Blottner and Baumgarten, 1992;Dun et al., 1992, 1993; Valtschanoff et al., 1992a,b; Ander-son et al., 1993; Vizzard et al., 1993, 1994; Saito et al.,1994; Papka et al., 1995; Tang et al., 1995), so NO isassumed to play a role in autonomic transmission (Bult etal., 1990; Anderson, 1992; Blottner and Baumgarten,1992; Briggs, 1992; Valtschanoff et al., 1992a; Anderson etal., 1993). The existence of NADPH-d and NOS in thesuperficial dorsal horn has been also reported in severalpapers, and NO is suggested to mediate hyperalgesia andsynaptic plasticity (Moore et al., 1991; Haley et al., 1992;Kitto et al., 1992; Meller et al., 1992a,b).Arginine, the substrate for NO production, may be
supplied either from the arginine pool of the cerebrospinal
Grant sponsor: Kaibara Morikazu Medical Science Promotion Founda-tion; Grant sponsor: Japan Science and Technology Corporation.*Correspondence to: Dr. Hiroyuki Nakamura, at his present address:
PRESTO, JST, Gifu University School ofMedicine, Department ofAnatomy,40 Tsukasa, Gifu 500, Japan. E-mail: [email protected] 26 June 1995; Revised 7 March 1997; Accepted 14April 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 385:616–626 (1997)
r 1997 WILEY-LISS, INC.

fluid or from aspartate and citrulline by the catalysis ofASS and ASL. However, the arginine concentration inserum and cerebrospinal fluid is not high enough to allowmaximal production of NO (Nussler et al., 1994), socolocalization of NOS, ASS, andASLmay be important foreffective NO production. Surprisingly, however, NOS andASS tend to be localized in different neurons in the ratbrain (Arnt-Ramos et al., 1992). Likewise, ASS, ASL, andarginase are localized differentially in most regions of therat brain (Nakamura et al., 1990). Therefore, the questionarises of where in the brain the enzymes are colocalizedand form a cyclic pathway within neurons.The aim of this study is to investigate whether NOS,
ASS, ASL, and arginase are colocalized in the spinal cord.Although Arnt-Ramos et al. (1992) reported the localiza-tion of ASS in relation to NADPH-d reaction in the ratbrain, the spinal cord was notmentioned. Neurons contain-ing NOS, which is a specific enzyme for the cytosolic NOsynthesis, can be stained histochemically by reacting withNADPH-d (Bredt et al., 1991a,b; Hope et al., 1991; Whiteand Marletta, 1992). In this study, we describe the neuro-nal localization of ASS-, ASL-, and arginase-like immuno-reactivity (-LI) in comparison with NADPH-d activity (1)in the spinal cord, by a fluorescent immunohistochemicalprocedure combined with NADPH-d histochemistry.
MATERIALS AND METHODS
Eight male Wistar rats weighing between 250 and 500 gwere used in this study. Three were used for immunohisto-chemistry with avidin-biotinylated-peroxidase complex(ABC) and diaminobenzidine (DAB) methods, and theother five were used for double staining with NADPH-dhistochemistry and immunohistochemistry with fluo-roisothiocyanate (FITC)-streptavidin orABC-alkaline phos-phatase (ABC-AP) reaction. Under deep anesthesia froman intraperitoneal injection of sodium pentobarbital (50mg/kg) and ether inhalation, the animals were perfusedthrough the ascending aorta with 200 ml of 0.02 Mphosphate-buffered saline (PBS), pH 7.4, followed by 500ml of a fixative solution containing 4% paraformaldehydein 0.1 M phosphate buffer (pH 7.4). The spinal cord wasthen blocked, removed, and immersed in a cold (4°C)
phosphate buffer solution containing 30% sucrose. After2–4 days, the blocks were transferred to cold isopentane(280°C) and kept in a deep freeze (280°C). Transversesections were cut at 50 µm on a freezing microtome. Everysixth section wasmounted directly on gelatinized slides forNissl staining with cresyl violet, and the remaining sec-tions were collected in PBS. For the single immunohisto-chemistry with ABC-DAB, one series of sections was usedfor NADPH-d histochemistry, and the other four serieswere used for immunohistochemistry with antisera againstNOS,ASS,ASL, and arginase. For the double staining, fiveseries of sections were collected in 0.02 M potassiumphosphate-buffered saline, pH 7.4 (KPBS), and were pro-cessed for NADPH-d histochemistry, four series of whichwere further processed for immunohistochemistry withFITC-streptavidin, by using antisera against ASS, ASL,and arginase, and with ABC-AP, by using antisera againstASS. The procedures for the animals were approved by theCommittee of Ethics in Animal Experiments, Faculty ofMedicine, Kyushu University.
Single immunohistochemistry
To demonstrate NOS-, ASS-, ASL-, and arginase-LI withthe ABC-DAB procedure, the sections were processedaccording to Ellis and Halliday (1992). Briefly, sectionswere rinsed with KPBS and immersed in a KPBS solutioncontaining 1% peroxide for 1–3 hours, rinsed again withKPBS, and incubated in a KPBS solution containing 0.1%bovine serum albumin overnight. They were then pro-cessed in a KPBS solution containing 1% normal goatserum, 0.3% bovine serum albumin, 0.2% sodium azide,0.3% Triton X-100, and primary antisera for 3–5 days at4°C. The sections were rinsed and incubated in 0.33%biotinylated goat antirabbit IgG (Vector Laboratories,Burlingame, CA) in KPBS containing 1% normal goatserum, 2% normal rat serum, 0.3% bovine albumin, and0.3% Triton X-100 for 15–40 hours. The sections were thenrinsed with KPBS and incubated in 0.2% Elite-ABC solu-tion (Vector Laboratories) in PBS containing 0.1% bovineserum albumin for 3 hours. Finally, the sections wereprocessed with glucose oxidase-DAB-nickel solution (Shuet al., 1988). Briefly, they were rinsed in PBS and then in0.1 M acetate buffer (pH 6.0) and incubated in 0.1 Macetate buffer containing 2% 3,38-diaminobenzidine tetra-hydrochloride (Dojin Chemical, Kumamoto, Japan), 8%¬-D-glucose, 1.6% ammonium chloride, and 0.3–0.4 unit/mlglucose oxidase (Toyobo, Tokyo, Japan) for 3–12 hours at4°C. The sections were rinsed, mounted on gelatinizedslides, dried, cleared with xylene, and coverslipped withCanada balsam.Specific antisera against purified rat NOS, ASS, ASL,
and arginase raised in rabbits were generously presentedby Drs. Mayer and Saheki (Institut fur Pharmakologie undToxikologie, Karl-Franzens-Universitat Graz, Graz, Aus-tria, and Department of Biochemistry, Kagoshima Univer-sity Faculty of Medicine, Kagoshima, Japan, respectively).The procedures for enzyme purification and characteriza-tions of the antisera have been described elsewhere (Sa-heki et al., 1977; Kawamoto et al., 1986; Kobayashi et al.,1986; Nakamura et al., 1991a; Kummer et al., 1992). Thedilutions of primary antisera were 1:5,000 for anti-ASSantiserum, 1:60,000 for anti-ASL antiserum, 1:3,000 forantiarginase antiserum, and 1:25,000 for anti-NOS antise-rum.
Fig. 1. Cyclic metabolic pathway for nitric oxide production.
NITRIC OXIDE RELATED ENZYMES IN SPINAL CORD 617

NADPH-d histochemistry
Amodified protocol of the NADPH-d histochemistry wasused. The free-floating sections were rinsed with KPBSand incubated in a KPBS solution containing 0.3% TritonX-100, 0.025% nitroblue tetrazolium (Sigma, St. Louis,MO), and 0.025% b-nicotinamide adenine dinucleotidephosphate reduced form (b-NADPH; Sigma) at room tem-perature for 1–2 hours and then were rinsed with KPBS.For the double staining of NADPH-d and fluorescenceimmunohistochemistry, theNADPH-d reactionwas stoppedbefore the cells got too dark in order not to mask orovershadow the fluorescence.For better staining, we tested the temperature at 4°C,
20°C, and 37°C for the NADPH-d incubation solution. Thereaction was stopped when the sections turned dark blue.After incubation at 4°C for 15–40 hours, the sections onlyshowed very weak staining. After incubation at 37°C for 1hour, maximal reaction was obtained for NADPH-d activeterminal-like structures, but background staining with fineblue grains also developed. At 20°C for 1–2 hours, cellular aswell as terminal staining became blue, without any blueproduct in other parts of the section. To suppress backgroundstaining, we therefore chose 20°C for theNADPH-d reaction.
Double-labeling studies
NADPH-d histochemistry and FITC immunohisto-
chemistry. For double staining with NADPH-d histo-chemistry and FITC-streptavidin immunohistochemistry,the sections were first incubated with NADPH-d solutionsas described above and then processed with the solutionsfor immunohistochemistry, using antisera against ASS,ASL, and arginase. After the NADPH-d reaction, thesections were rinsed in KPBS and incubated in a KPBSsolution containing 0.1% bovine serum albumin overnight.The sections were then transferred into KPBS solutionscontaining 1% normal goat serum, 0.3% bovine serumalbumin, 0.2% sodium azide, 0.3% Triton X-100 and pri-mary antisera for 3–5 days at 4°C. The dilutions of theprimary antisera were 1:2,000 forASS antiserum, 1:25,000for ASL antiserum, and 1:1,000 for arginase antiserum.The sections were rinsed in KPBS and incubated inbiotinylated anti-rabbit goat IgG (Vector Laboratories) inKPBS containing 1% normal goat serum, 2% normal ratserum, 0.3% bovine serum albumin, and 0.3% Triton X-100for 15–40 hours. They were then rinsed with KPBS andincubated in 1:200 FITC-streptavidin (Vector Laborato-ries) in KPBS containing 0.1% bovine serum albumin for3–12 hours. The sections were rinsed, mounted on gelati-nized slides, and coverslipped with Vectashield (VectorLaboratories).In double-stained preparations, the antisera against
ASS and arginase stained neuronal somata, as was seen inthe single FITC-stained neurons. By the dense NADPH-dstaining of the soma, the double-stained neurons appearedto have brightly fluorescent nuclei, with a black ring ofNADPH-d active soma.ASL-LI neurons sometimes showedstronger staining of the nucleus, the reason for which we donotknow;however, thedistributionpattern ofASL-LIneuronswas in agreementwith that of arginine (Aoki et al., 1991).NADPH-d histochemistry and ABC-AP immunohisto-
chemistry. As in the double-staining studies of NADPH-dhistochemistry and FITC immunohistochemistry de-scribed above, the sections were first incubated in theNADPH-d reaction medium and then rinsed in KPBS and
incubated in a KPBS solution containing 0.1% bovineserum albumin overnight. They were then transferred intoKPBS solutions containing 1% normal goat serum, 0.3%bovine serum albumin, 0.2% sodium azide, 0.3% TritonX-100 and primary antisera against ASS (1:1,500 dilution)for 3–5 days at 4°C. The sections were rinsed and incu-bated in biotinylated anti-rabbit goat IgG (Vector Labora-tories) in KPBS containing 1% normal goat serum, 2%normal rat serum, 0.3% bovine serum albumin, and 0.3%Triton X-100 for 15–40 hours. They were then rinsed withKPBS, incubated in ABC-AP solution (Vector Laborato-ries) in KPBS containing 0.1% bovine serum albumin for3–12 hours and rinsed in KPBS and then in 0.1M Tris-HCl(pH 8.2). Finally, the sections were incubated with VectorRed reaction medium (Vector Laboratories), rinsed,mounted on gelatinized slides, dried, cleared with xylene,and coverslipped with Permount.
Cytoarchitecture
The cytoarchitecture of the spinal cord is based ondescriptions by Rexed (1952, 1954) and Molander et al.(1984, 1989). For the nomenclature of the zones in laminaII, we use lamina II externa (IIe) and lamina II interna(IIi) instead of the outer zone and inner zone of lamina II ofMolander et al. (1989).
RESULTS
Figure 2 shows the overall distribution patterns ofASS-,ASL-, and arginase-LI. ASS-LI neurons were restricted tothe superficial dorsal horn, the nucleus intermediolatera-lis, and the central canal region, whereas ASL- andarginase-LI neurons were scattered throughout the graymatter. Glial cells also were ASL-LI in the gray and whitematter. Strongly immunoreactive neuropile was seen onlyin the superficial dorsal horn.In the ventral horn, NADPH-d active fibers were seen
running dorsoventrally. We have confirmed our previousfinding that fibers and terminals in this region showedASS-LI (Nakamura et al., 1991a). We could not, however,find any fibers exhibiting both NADPH-d (1) and ASS-LI.Because we have already reported on the details of theventral horn (Nakamura et al., 1990), the following descrip-tion will be focused on the dorsal horn, intermediolateralnucleus, and region surrounding the central canal.
Dorsal horn
In single-labeled preparations, NADPH-d positive (1)neurons were observed throughout laminae II and III, anda few were also seen in laminae IV and V (Fig. 3). Smallfusiform and triangular neurons were NADPH-d (1) inboth laminae IIe and IIi. Similar neurons were NADPH-d(1) also in lamina III. A few medium-sized neurons werestained in laminae IV and V.Immunocytochemistry showed that ASS- and ASL-LI
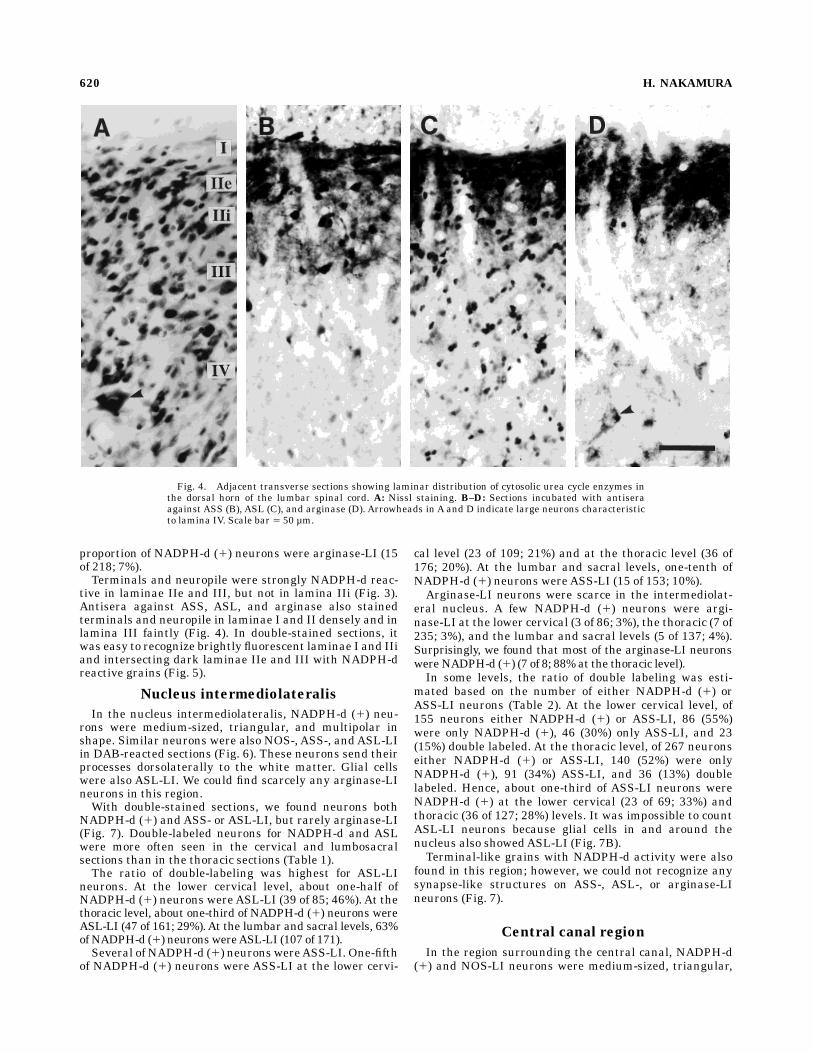
neurons were abundant through lamina I to upper laminaIII, and they were occasionally seen in lamina IV (Fig. 4).Glial cells were also ASL-LI throughout the dorsal horn.Many strongly arginase-LI neurons were found in laminaeI and II. A few large neurons in laminae III and IV wereweakly arginase-LI. Neuropile was strongly ASS-, ASL-,and arginase-LI in laminae I and IIe and was weaklyimmunoreactive in lamina IIi and upper lamina III.With double-stained sections, we found several neurons
both NADPH-d (1) and ASL-LI in laminae II and III
618 H. NAKAMURA

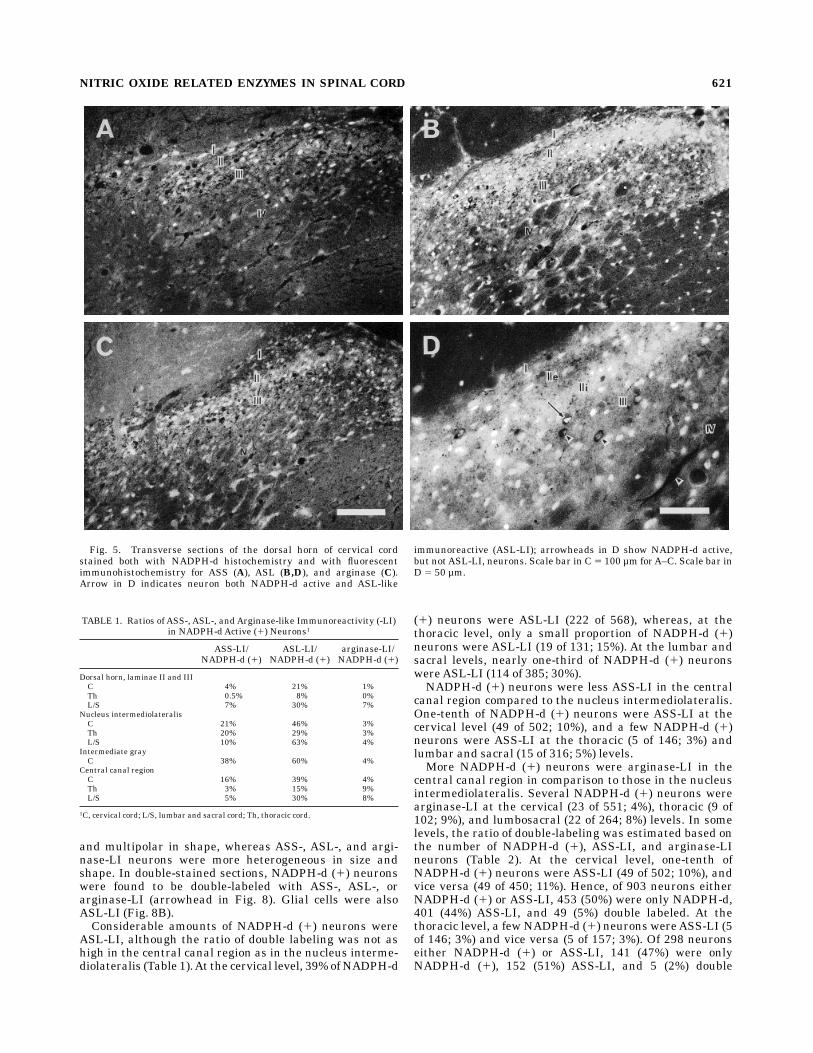
(arrow in Fig. 5D). At the cervical level, 22% of NADPH-d(1) neurons were ASL-LI (266 of 1,197), whereas, at thethoracic level, a relatively small proportion of NADPH-d(1) neurons were ASL-LI (11 of 133; 8%; Table 1). At thelumbar and sacral levels, about one-third of NADPH-d (1)neurons wereASL-LI (114 of 385; 30%).A few double-labeled neurons both NADPH-d (1) and
ASS-LI were also found in laminae II and III. At thecervical level, a small proportion of NADPH-d (1) neurons
were ASS-LI (42 of 1,170; 4%), whereas, at the thoraciclevel, few NADPH-d (1) neurons were ASS-LI (1 of 686;0.15%). At the lumbar and sacral levels, NADPH-d (1)neurons were occasionally foundASS-LI (21 of 290; 7%).A few neurons were both NADPH-d (1) and arginase-LI
in laminae II and III. At the cervical level, some NADPH-d(1) neurons were arginase-LI (19 of 137; 11%), whereas, atthe thoracic level, no NADPH-d (1) neuron was argi-nase-LI (0 of 595). At the lumbar and sacral levels, a small
Fig. 2. Photomicrographs showing the overall distribution of immunoreactivities for argininosucci-nate synthetase (ASS; A), argininosuccinate lyase (ASL; B), and arginase (C) in adjacent transversesections of the upper sacral cord. CC, central canal; IML, nucleus intermediolateralis; VH, ventral horn.Scale bar 5 200 µm.
Fig. 3. Nissl-stained (A) and NADPH-d-reacted (B) adjacent transverse sections of the dorsal horn ofthe thoracic cord showing laminar correspondence. Scale bar 5 100 µm.
NITRIC OXIDE RELATED ENZYMES IN SPINAL CORD 619
proportion of NADPH-d (1) neurons were arginase-LI (15of 218; 7%).Terminals and neuropile were strongly NADPH-d reac-
tive in laminae IIe and III, but not in lamina IIi (Fig. 3).Antisera against ASS, ASL, and arginase also stainedterminals and neuropile in laminae I and II densely and inlamina III faintly (Fig. 4). In double-stained sections, itwas easy to recognize brightly fluorescent laminae I and IIiand intersecting dark laminae IIe and III with NADPH-dreactive grains (Fig. 5).
Nucleus intermediolateralis
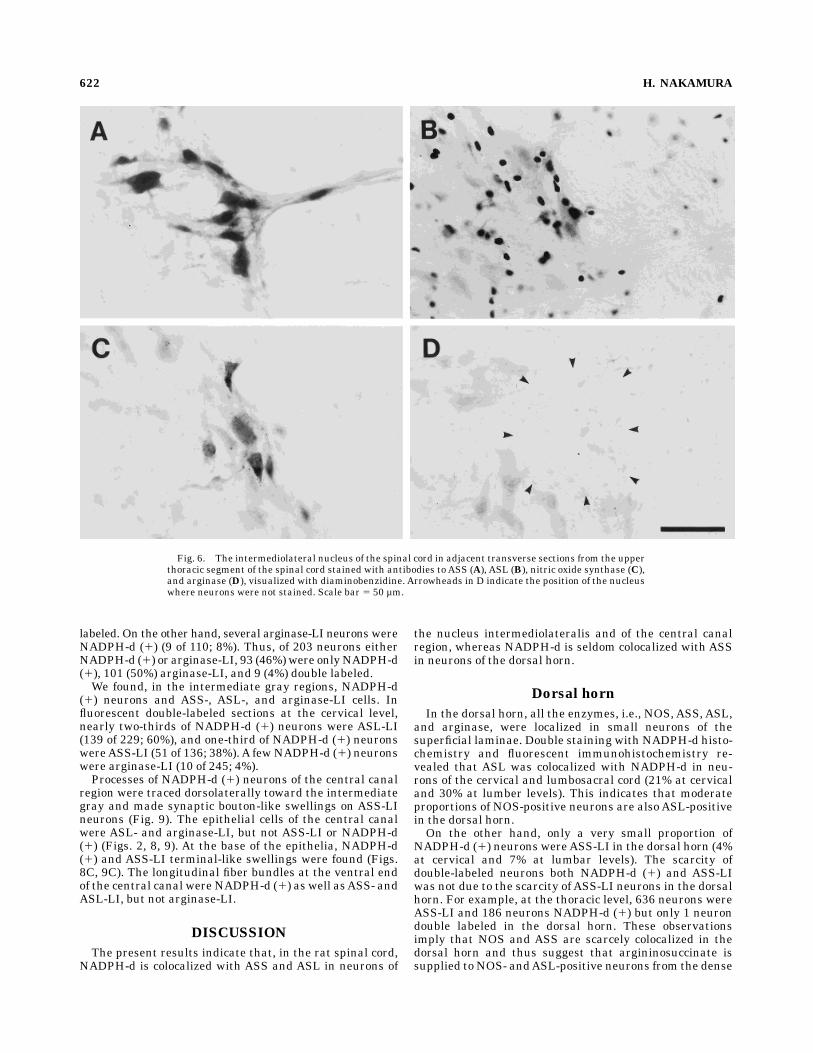
In the nucleus intermediolateralis, NADPH-d (1) neu-rons were medium-sized, triangular, and multipolar inshape. Similar neurons were also NOS-, ASS-, andASL-LIin DAB-reacted sections (Fig. 6). These neurons send theirprocesses dorsolaterally to the white matter. Glial cellswere also ASL-LI. We could find scarcely any arginase-LIneurons in this region.With double-stained sections, we found neurons both
NADPH-d (1) and ASS- or ASL-LI, but rarely arginase-LI(Fig. 7). Double-labeled neurons for NADPH-d and ASLwere more often seen in the cervical and lumbosacralsections than in the thoracic sections (Table 1).The ratio of double-labeling was highest for ASL-LI
neurons. At the lower cervical level, about one-half ofNADPH-d (1) neurons wereASL-LI (39 of 85; 46%). At thethoracic level, about one-third of NADPH-d (1) neurons wereASL-LI (47 of 161; 29%).At the lumbar and sacral levels, 63%ofNADPH-d (1) neuronswereASL-LI (107 of 171).Several of NADPH-d (1) neurons wereASS-LI. One-fifth
of NADPH-d (1) neurons were ASS-LI at the lower cervi-
cal level (23 of 109; 21%) and at the thoracic level (36 of176; 20%). At the lumbar and sacral levels, one-tenth ofNADPH-d (1) neurons wereASS-LI (15 of 153; 10%).Arginase-LI neurons were scarce in the intermediolat-
eral nucleus. A few NADPH-d (1) neurons were argi-nase-LI at the lower cervical (3 of 86; 3%), the thoracic (7 of235; 3%), and the lumbar and sacral levels (5 of 137; 4%).Surprisingly, we found that most of the arginase-LI neuronswereNADPH-d (1) (7 of 8; 88%at the thoracic level).In some levels, the ratio of double labeling was esti-
mated based on the number of either NADPH-d (1) orASS-LI neurons (Table 2). At the lower cervical level, of155 neurons either NADPH-d (1) or ASS-LI, 86 (55%)were only NADPH-d (1), 46 (30%) only ASS-LI, and 23(15%) double labeled. At the thoracic level, of 267 neuronseither NADPH-d (1) or ASS-LI, 140 (52%) were onlyNADPH-d (1), 91 (34%) ASS-LI, and 36 (13%) doublelabeled. Hence, about one-third of ASS-LI neurons wereNADPH-d (1) at the lower cervical (23 of 69; 33%) andthoracic (36 of 127; 28%) levels. It was impossible to countASL-LI neurons because glial cells in and around thenucleus also showedASL-LI (Fig. 7B).Terminal-like grains with NADPH-d activity were also
found in this region; however, we could not recognize anysynapse-like structures on ASS-, ASL-, or arginase-LIneurons (Fig. 7).
Central canal region
In the region surrounding the central canal, NADPH-d(1) and NOS-LI neurons were medium-sized, triangular,
Fig. 4. Adjacent transverse sections showing laminar distribution of cytosolic urea cycle enzymes inthe dorsal horn of the lumbar spinal cord. A: Nissl staining. B–D: Sections incubated with antiseraagainst ASS (B), ASL (C), and arginase (D). Arrowheads in A and D indicate large neurons characteristicto lamina IV. Scale bar 5 50 µm.
620 H. NAKAMURA
and multipolar in shape, whereas ASS-, ASL-, and argi-nase-LI neurons were more heterogeneous in size andshape. In double-stained sections, NADPH-d (1) neuronswere found to be double-labeled with ASS-, ASL-, orarginase-LI (arrowhead in Fig. 8). Glial cells were alsoASL-LI (Fig. 8B).Considerable amounts of NADPH-d (1) neurons were
ASL-LI, although the ratio of double labeling was not ashigh in the central canal region as in the nucleus interme-diolateralis (Table 1).At the cervical level, 39% ofNADPH-d
(1) neurons were ASL-LI (222 of 568), whereas, at thethoracic level, only a small proportion of NADPH-d (1)neurons were ASL-LI (19 of 131; 15%). At the lumbar andsacral levels, nearly one-third of NADPH-d (1) neuronswereASL-LI (114 of 385; 30%).NADPH-d (1) neurons were less ASS-LI in the central
canal region compared to the nucleus intermediolateralis.One-tenth of NADPH-d (1) neurons were ASS-LI at thecervical level (49 of 502; 10%), and a few NADPH-d (1)neurons were ASS-LI at the thoracic (5 of 146; 3%) andlumbar and sacral (15 of 316; 5%) levels.More NADPH-d (1) neurons were arginase-LI in the
central canal region in comparison to those in the nucleusintermediolateralis. Several NADPH-d (1) neurons werearginase-LI at the cervical (23 of 551; 4%), thoracic (9 of102; 9%), and lumbosacral (22 of 264; 8%) levels. In somelevels, the ratio of double-labeling was estimated based onthe number of NADPH-d (1), ASS-LI, and arginase-LIneurons (Table 2). At the cervical level, one-tenth ofNADPH-d (1) neurons were ASS-LI (49 of 502; 10%), andvice versa (49 of 450; 11%). Hence, of 903 neurons eitherNADPH-d (1) or ASS-LI, 453 (50%) were only NADPH-d,401 (44%) ASS-LI, and 49 (5%) double labeled. At thethoracic level, a few NADPH-d (1) neurons wereASS-LI (5of 146; 3%) and vice versa (5 of 157; 3%). Of 298 neuronseither NADPH-d (1) or ASS-LI, 141 (47%) were onlyNADPH-d (1), 152 (51%) ASS-LI, and 5 (2%) double
Fig. 5. Transverse sections of the dorsal horn of cervical cordstained both with NADPH-d histochemistry and with fluorescentimmunohistochemistry for ASS (A), ASL (B,D), and arginase (C).Arrow in D indicates neuron both NADPH-d active and ASL-like
immunoreactive (ASL-LI); arrowheads in D show NADPH-d active,but notASL-LI, neurons. Scale bar in C5 100 µm forA–C. Scale bar inD 5 50 µm.
TABLE 1. Ratios of ASS-, ASL-, and Arginase-like Immunoreactivity (-LI)in NADPH-d Active (1) Neurons1
ASS-LI/NADPH-d (1)
ASL-LI/NADPH-d (1)
arginase-LI/NADPH-d (1)
Dorsal horn, laminae II and IIIC 4% 21% 1%Th 0.5% 8% 0%L/S 7% 30% 7%
Nucleus intermediolateralisC 21% 46% 3%Th 20% 29% 3%L/S 10% 63% 4%
Intermediate grayC 38% 60% 4%
Central canal regionC 16% 39% 4%Th 3% 15% 9%L/S 5% 30% 8%
1C, cervical cord; L/S, lumbar and sacral cord; Th, thoracic cord.
NITRIC OXIDE RELATED ENZYMES IN SPINAL CORD 621
labeled. On the other hand, several arginase-LI neurons wereNADPH-d (1) (9 of 110; 8%). Thus, of 203 neurons eitherNADPH-d (1) or arginase-LI, 93 (46%)were onlyNADPH-d(1), 101 (50%) arginase-LI, and 9 (4%) double labeled.We found, in the intermediate gray regions, NADPH-d
(1) neurons and ASS-, ASL-, and arginase-LI cells. Influorescent double-labeled sections at the cervical level,nearly two-thirds of NADPH-d (1) neurons were ASL-LI(139 of 229; 60%), and one-third of NADPH-d (1) neuronswereASS-LI (51 of 136; 38%).A fewNADPH-d (1) neuronswere arginase-LI (10 of 245; 4%).Processes of NADPH-d (1) neurons of the central canal
region were traced dorsolaterally toward the intermediategray and made synaptic bouton-like swellings on ASS-LIneurons (Fig. 9). The epithelial cells of the central canalwere ASL- and arginase-LI, but not ASS-LI or NADPH-d(1) (Figs. 2, 8, 9). At the base of the epithelia, NADPH-d(1) and ASS-LI terminal-like swellings were found (Figs.8C, 9C). The longitudinal fiber bundles at the ventral endof the central canal were NADPH-d (1) as well asASS- andASL-LI, but not arginase-LI.
DISCUSSION
The present results indicate that, in the rat spinal cord,NADPH-d is colocalized with ASS and ASL in neurons of
the nucleus intermediolateralis and of the central canalregion, whereas NADPH-d is seldom colocalized with ASSin neurons of the dorsal horn.
Dorsal horn
In the dorsal horn, all the enzymes, i.e., NOS,ASS, ASL,and arginase, were localized in small neurons of thesuperficial laminae. Double staining with NADPH-d histo-chemistry and fluorescent immunohistochemistry re-vealed that ASL was colocalized with NADPH-d in neu-rons of the cervical and lumbosacral cord (21% at cervicaland 30% at lumber levels). This indicates that moderateproportions of NOS-positive neurons are also ASL-positivein the dorsal horn.On the other hand, only a very small proportion of
NADPH-d (1) neurons wereASS-LI in the dorsal horn (4%at cervical and 7% at lumbar levels). The scarcity ofdouble-labeled neurons both NADPH-d (1) and ASS-LIwas not due to the scarcity of ASS-LI neurons in the dorsalhorn. For example, at the thoracic level, 636 neurons wereASS-LI and 186 neurons NADPH-d (1) but only 1 neurondouble labeled in the dorsal horn. These observationsimply that NOS and ASS are scarcely colocalized in thedorsal horn and thus suggest that argininosuccinate issupplied to NOS- andASL-positive neurons from the dense
Fig. 6. The intermediolateral nucleus of the spinal cord in adjacent transverse sections from the upperthoracic segment of the spinal cord stained with antibodies to ASS (A), ASL (B), nitric oxide synthase (C),and arginase (D), visualized with diaminobenzidine. Arrowheads in D indicate the position of the nucleuswhere neurons were not stained. Scale bar 5 50 µm.
622 H. NAKAMURA

plexus of ASS-positive terminals or nearby ASS-positiveneurons in the superficial laminae of the dorsal horn.We have previously reported that urea cycle enzymes
are localized differentially in neurons in several regions ofthe brain (Nakamura et al., 1990), but ASS-, ASL-, andarginase-LI cells are closely related, sometimes makingsynaptic contact with each other. These closely relatedenzymes mediate consecutive catalytic activity, so thatarginine and argininosuccinate can be transneuronallytransported. The idea of transneuronal transport wasstrengthened in our previous study showing that, inisolated cerebellar neurons, argininosuccinate modulates
the intracellular calcium increase by glutamate receptoractivation (Nakamura et al., 1991b), suggesting the exis-tence of intercellular communication with argininosucci-nate.
Nucleus intermediolateralisand central canal region
We confirmed the existence of NADPH-d (1) cells andNOS-positive neurons in the intermediolateral nucleus ofthe spinal cord (Anderson, 1992; Blottner and Baumgar-ten, 1992; Dun et al., 1992; Valtschanoff et al., 1992a;Anderson et al., 1993; Spike et al., 1993; Vizzard et al.,1993, 1994; Saito et al., 1994; Okamura et al., 1995; Papkaet al., 1995; Tang et al., 1995). These NADPH-d (1)neurons were reported to be sympathetic and parasympa-thetic preganglionic neurons (Vizzard et al., 1993; Oka-mura et al., 1995).Our results indicate that, in the intermediolateral
nucleus, NOS,ASS, andASL are colocalized in the pregan-glionic neurons, suggesting the existence of the cyclicpathway to form NO here. Thus, our results furthersupport the idea of NO as a neurotransmitter of theautonomic preganglionic neurons.Colocalization ratio of ASS and NADPH-d is not as high
as that of ASL and NADPH-d in the intermediolateralisand central canal region. Recent studies indicate that NOSandASS are induced simultaneously by bacterial lipopoly-saccharide (LPS) in vascular smooth muscle cells andmacrophages (Hattori et al., 1994; Nussler et al., 1994).Thus, ASS and NOS may be induced in the nervous tissueas well.The induction of ASS and NOS by LPS also suggests the
importance of recycling of citrulline to arginine via thecitrulline-NO cycle in NO production. Because arginine isthe only physiological substrate for NOS, its concentrationmay determine the rate of NO production.Arginine concen-tration in the brain is 167–229 nmol/g, which is almost thesame as that in the serum (177–284 nmol/ml; in mouse;personal communication from Dr. Takeyori Saheki, Ka-goshima University Faculty of Medicine, Kagoshima, Ja-pan). This is about one-half of the concentration formaximal production of NO (Nussler et al., 1994). Tosupplement the shortage of arginine, colocalization of ASSand ASL with NOS, and thus the existence of an NO cyclewithin the neuron to recycle citrulline to arginine, may beof great importance.
Rostrocaudal difference incolocalization ratio
As is shown in Table 1, a rostrocaudal difference exists incolocalization ratio in the dorsal horn, the nucleus in-
Fig. 7. Fluorescent photomicrographs of the nucleus intermediolat-eralis of the lower cervical cord stained with NADPH-d histochemistryas well as with antisera against ASS (A), ASL (B), and arginase (C).Arrowheads indicate double-labeled neurons. Scale bar 5 50 µm.
TABLE 2. Proportion of NADPH-dActive (1), ASS-like Immunoreactive(-LI), and Double-labeled Neurons per Neurons Either NADPH-d Active or
Immunoreactive for ASS1
NADPH-d(1) only
ASS-LIonly
NADPH-d (1)1 ASS-LI
Nucleus intermediolateralisC 30% 55% 15%Th 52% 34% 13%
Central canal regionC 50% 44% 5%Th 47% 51% 2%
1C, cervical cord; Th, thoracic cord.
NITRIC OXIDE RELATED ENZYMES IN SPINAL CORD 623
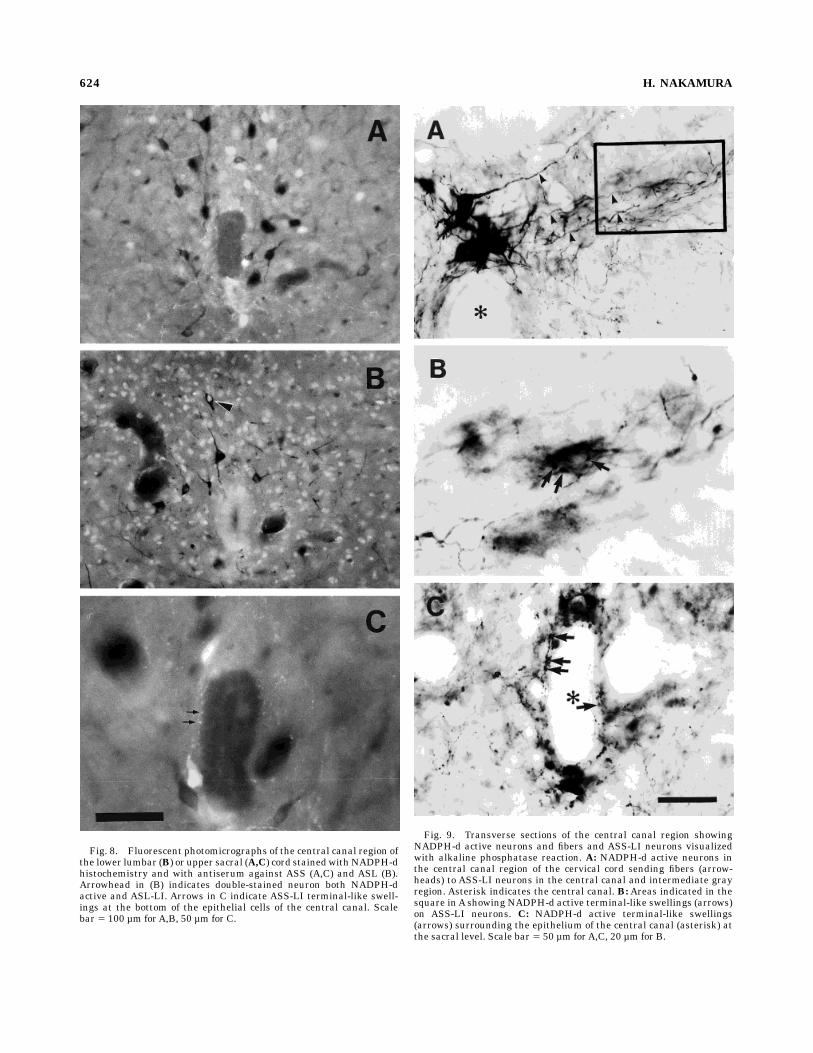
Fig. 8. Fluorescent photomicrographs of the central canal region ofthe lower lumbar (B) or upper sacral (A,C) cord stainedwithNADPH-dhistochemistry and with antiserum against ASS (A,C) and ASL (B).Arrowhead in (B) indicates double-stained neuron both NADPH-dactive and ASL-LI. Arrows in C indicate ASS-LI terminal-like swell-ings at the bottom of the epithelial cells of the central canal. Scalebar 5 100 µm forA,B, 50 µm for C.
Fig. 9. Transverse sections of the central canal region showingNADPH-d active neurons and fibers and ASS-LI neurons visualizedwith alkaline phosphatase reaction. A: NADPH-d active neurons inthe central canal region of the cervical cord sending fibers (arrow-heads) to ASS-LI neurons in the central canal and intermediate grayregion. Asterisk indicates the central canal. B: Areas indicated in thesquare inA showing NADPH-d active terminal-like swellings (arrows)on ASS-LI neurons. C: NADPH-d active terminal-like swellings(arrows) surrounding the epithelium of the central canal (asterisk) atthe sacral level. Scale bar 5 50 µm forA,C, 20 µm for B.
624 H. NAKAMURA

termediolateralis, and the central canal regions. Thedouble-labeling ratio in the thoracic cord was about one-half of that in the cervical and lumbosacral cord. Forexample, in the nucleus intermediolateralis, the double-staining ratio of ASL with NADPH-d reached 45–60% inthe cervical and lumbar cords but was 30% in the thoraciccord.There are two possible reasons for the different ratio of
colocalization at different levels of the spinal cord. Onepossibility is the technical problem inherent in spinal cordfixation and anatomical differences in vascularization: Theblood supply of the spinal cord is provided mainly by theanterior spinal artery branching from the vertebral arteryand by the lower thoracic and upper lumbar branches ofthe intercostal and lumbar arteries. Thus, the cervical,lower thoracic, and upper lumbar cord may be betterperfused and fixed than the upper and middle thoraciccord. The difference of fixation may reflect the colocaliza-tion rate between different segments, because theNADPH-d reaction is influenced by the fixation of thetissue. The other possibility is that the difference incolocalization ratio actually depends on the segmentalrelation to the autonomic nervous system. A preciselycontrolled and systematic examination will be necessary toanswer this question.
ACKNOWLEDGMENTS
The author thanks Dr. M. Kawabuchi for his supportand encouragement; Dr. T. Saheki for kindly providingantisera against ASS, ASL, and arginase as well as forinvaluable discussion; Dr. Bernd Mayer for the gift ofantiserum against NOS; Drs. Teruyoshi Kondoh andAkikoYamashita for discussion; and Y. Hirakawa and A.T.M.S.Islam for technical assistance.
LITERATURE CITED
Anderson, C.R. (1992) NADPH diaphorase-positive neurons in the ratspinal cord including a subpopulation of autonomic preganglionicneurons. Neurosci. Lett. 139:280–284.
Anderson, C.R., S.L. Edwards, J.B. Furness, D.S. Bredt, and S.H. Snyder(1993) The distribution of nitric oxide synthase-containing autonomicpreganglionic terminals in the rat. Brain Res. 614:78–85.
Aoki, E., R. Semba, K. Mikoshiba, and S. Kashiwamata (1991) Predomi-nant localization of glial cells of free L-arginine. Immunocytochemicalevidence. Brain Res. 547:190–192.
Arnt-Ramos, L.R., W.E. O’Brien, and S.R. Vincent (1992) Immunohisto-chemical localization of argininosuccinate synthetase in the rat brain inrelation to nitric oxide synthase-containing neurons. Neuroscience51:773–789.
Blottner, D., and H. Baumgarten (1992) Nitric oxide synthetase (NOS)containing sympathoadrenal cholinergic neurons of the rat IML-cellcolumn: Evidence from histochemistry, immunohistochemistry, andretrograde labeling. J. Comp. Neurol. 316:45–55.
Bredt, D.A., and S.H. Snyder (1989) Nitric oxide mediates glutamate-linked enhancement of cGMP levels in the cerebellum. Proc. Natl. Acad.Sci. U.S.A. 86:9030–9033.
Bredt, D.S., and S.H. Snyder (1990) Isolation of nitric oxide synthetase, acalmodulin-requiring enzyme. Proc. Natl. Acad. Sci. U.S.A. 87:682–685.
Bredt, D.S., P.M. Hwang, and S.H. Snyder (1990) Localization of nitricoxide synthase indicating a neural role for nitric oxide. Nature 347:768–770.
Bredt, D.S., C.E. Glatt, P.M. Hwang, M. Fotuhi, T.M. Dawson, and S.H.Snyder (1991a) Nitric oxide synthase protein and mRNA are discretelylocalized in neuronal populations of the mammalian CNS together withNADPH diaphorase. Neuron 7:615–624.
Bredt, D.S., P.M. Hwang, C.E. Glatt, C. Lowenstein, R.R. Reed, and S.H.Snyder (1991b) Cloned and expressed nitric oxide synthase structurallyresembles cytochrome P-450 reductase. Nature 351:714–718.
Briggs, C.A. (1992) Potentiation of nicotinic transmission in the ratsuperior cervical sympathetic ganglion: Effects of cyclic GMP and nitricoxide generators. Brain Res. 573:139–146.
Bult, H., G.E. Boeckxstaens, P.A. Pelckmans, F.H. Jordaens, Y.M. VanMaercke, and A.G. Herman (1990) Nitric oxide as an inhibitorynonadrenergic noncholinergic neurotransmitter. Nature 345:346–347.
Dawson, T.M., and S.H. Snyder (1994) Gases as biological messengers:Nitric oxide and carbon monoxide in the brain. J. Neurosci. 14:5147–5159.
Dun, N.J., S.L. Dun, U. Forstermann, and L.F. Tseng (1992) Nitric oxidesynthase immunoreactivity in the rat spinal cord. Neurosci. Lett.147:217–220.
Dun, N.J., S.L. Dun, S.Y. Wu, U. Forstermann, H.H.H.W. Schmidt, and L.F.Tseng (1993) Nitric oxide synthase immunoreactivity in the rat, mouse,cat and squirrel monkey spinal cord. Neuroscience 54:845–857.
Ellis, J., and G. Halliday (1992) A comparative study of avidin-biotin-peroxidase complexes for the immunohistochemical detection of anti-gens in neural tissue. Biotech. Histochem. 67:367–371.
Haley, J.E., A.H. Dickenson, and M. Schachter (1992) Electrophysiologicalevidence for a role of nitric oxide in prolonged chemical nociception inthe rat. Neuropharmacology 31:93–96.
Hattori, Y., E.B. Campbell, and S.S. Gross (1994)Argininosuccinate synthe-tase mRNA and activity are induced by immunostimulants in vascularsmooth muscle. Role in the regeneration of arginine for nitric oxidesynthesis. J. Biol. Chem. 269:9405–9408.
Hope, B.T., G.J. Michael, K.M. Knigge, and S.R. Vincent (1991) NeuronalNADPH diaphorase is a nitric oxide synthetase. Proc. Natl. Acad. Sci.U.S.A. 88:2811–2814.
Jackson, M.J., A.L. Beaudet, and W.E. O’Brien (1986) Mammalian ureacycle enzymes. Annu. Rev. Genet. 20:431–464.
Kawamoto, S., Y. Amaya, T. Oda, T. Kuzumi, T. Saheki, S. Kimura, and M.Mori (1986) Cloning and expression in Escherichia coli of cDNA forarginase of rat liver. Biochem. Biophys. Res. Commun. 136:955–961.
Kitto, K.F., J.E. Haley, and G.L. Wilcox (1992) Involvement of nitric oxide inspinally mediated hyperalgesia in the mouse. Neurosci. Lett. 148:1–5.
Kobayashi, K., Y. Itakura, T. Saheki, K. Nakano, M. Sase, K. Oyanagi, R.Okamoto, and M. Mino (1986) Absence of argininosuccinate lyaseprotein in the liver of two patients with argininosuccinic aciduria. Clin.Chim.Acta 159:59–67.
Kummer, W., A. Fischer, P. Mundel, B. Mayer, B. Hoba, B. Philippin, and U.Preissler (1992) Nitric oxide synthase in VIP-containing cholinergicvasodilator nerves. Neuroreport 3:653–655.
Meller, S.T., C. Dykstra, and G.F. Gebhart (1992a) Production of endog-enous nitric oxide and activation of soluble guanylate cyclase arerequired for N-methyl-D-aspartate-produced facilitation of the nocicep-tive tail-flick reflex. Eur. J. Pharmacol. 214:93–96.
Meller, S.T., P.S. Pechman, G.F. Gebhart, and Y.J. Maves (1992b) Nitricoxide mediates the thermal hyperalgesia produced in a model ofneuropathic pain in the rat. Neuroscience 50:7–10.
Mizukawa, K., S.R. Vincent, P.L. McGeer, and E.G.McGeer (1989) Distribu-tion of reduced-nicotininamide-adenine-dinucleotide-phosphate-diapho-rase-positive cells and fibers in the cat central nervous system. J. Comp.Neurol. 279:281–311.
Molander, C., Q. Xu, and G. Grant (1984) The cytoarchitectonic organiza-tion of the spinal cord in the rat. I. The lower thoracic and lumbosacralcord. J. Comp. Neurol. 230:133–141.
Molander, C., Q. Xu, C. Rivero-Melian, and G. Grant (1989) Cytoarchitec-tonic organization of the spinal cord in the rat: II. The cervical andupper thoracic cord. J. Comp. Neurol. 289:375–385.
Moore, P.K., A.O. Oluyomi, R.C. Babbedge, P. Wallace, and S.L. Hart (1991)L-NG-nitro argininemethyl ester exhibits antinociceptive activity in themouse. Br. J. Pharmacol. 102:198–202.
Nakamura, H., T. Saheki, and S. Nakagawa (1990) Differential cellularlocalization of enzymes of L-arginine metabolism in the rat brain. BrainRes. 530:108–112.
Nakamura, H., T. Saheki, H. Ichiki, K. Nakata, and S. Nakagawa (1991a)Immunocytochemical localization of argininosuccinate synthetase inthe rat brain. J. Comp. Neurol. 312:652–679.
Nakamura, H., T. Yada, T. Saheki, T. Noda, and S. Nakagawa (1991b)L-argininosuccinate modulates L-glutamate response in acutely iso-lated cerebellar neurons of immature rat brain. Brain Res. 539:312–315.
NITRIC OXIDE RELATED ENZYMES IN SPINAL CORD 625

Nussler,A.K., T.R. Billiar, Z.-Z. Liu, and S.M.Morris, Jr. (1994) Coinductionof nitric oxide synthase and argininosuccinate synthetase in a murinemacrophage cell line. J. Biol. Chem. 269:1257–1261.
Okamura, H., K. Umehara, N. Tadaki, Y. Hisa, H. Esumi, and Y. Ibata(1995) Sympathetic preganglionic neurons contain nitric oxide syn-thase and project to the superior cervical ganglion: Combined applica-tion of retrograde neuronal tracer andNADPH-diaphorase histochemis-try. Brain Res. Bull. 36:491–494.
Papka, R.E., J.R. McCurdy, S.J. Williams, B. Mayer, L. Marson, and K.B.Platt (1995) Parasympathetic preganglionic neurons in the spinal cordinvolved in uterine innervation are cholinergic and nitric oxide-containing. Anat. Rec. 241:554–562.
Ratner, S., H. Morell, and E. Carvalho (1960) Enzymes of argininemetabolism in brain. Arch. Biochem. Biophys. 91:280–289.
Rexed, B. (1952) The cytoarchitectonic organization of the spinal cord in thecat. J. Comp. Neurol. 96:415–495.
Rexed, B. (1954) A cytoarchitectonic atlas of the spinal cord in the cat. J.Comp. Neurol. 100:297–379.
Saheki, T., T. Kusumi, S. Takada, and T. Katsunuma (1977) Studies of ratliver argininosuccinate synthetase. I. Physicochemical, catalytic, andimmunochemical properties. J. Biochem. 81:687–696.
Saito, A., G.J. Kidd, B.D. Trapp, T.M. Dawson, D.S. Bredt, D. Wilson, R.J.Traystman, S.H. Snyder, and D.F. Hanley (1994) Rat spinal cordneurons contain nitric oxide synthase. Neuroscience 59:447–456.
Shu, S., G. Ju, and L. Fan (1988) The glucose oxidase-DAB-nickel method inperoxidase histochemistry of the nervous system. Neurosci. Lett.85:169–171.
Spike, R.C., A.J. Todd, and H.M. Johnston (1993) Coexistence of NADPHdiaphorase with GABA, glycine, and acetylcholine in rat spinal cord. J.Comp. Neurol. 335:320–333.
Tang, F.R., C.K. Tan, and E.A. Ling (1995) Light and electron microscopicstudies of the distribution of NADPH-diaphorase in the rat upperthoracic spinal cord with special reference to the spinal autonomicregion. Arch. Histol. Cytol. 58:493–505.
Valtschanoff, J.G., R.J. Weinberg, and A. Rustioni (1992a) NADPH diapho-rase in the spinal cord in rats. J. Comp. Neurol. 321:209–222.
Valtschanoff, J.G., R.J. Weinberg, A. Rustioni, and H.H.H.W. Schmidt(1992b) Nitric oxide synthase and GABA colocalize in lamina II of ratspinal cord. Neurosci. Lett. 148:6–109.
Vincent, S.R., and H. Kimura (1992) Histochemical mapping of nitric oxidesynthase in the rat brain. Neuroscience 46:755–784.
Vizzard, M.A., S.L. Erdman, and W.C. de Groat (1993) Localization ofNADPH-diaphorase in pelvic afferent and efferent pathways of the rat.Neurosci. Lett. 152:72–76.
Vizzard, M.A., S.L. Erdman, V.L. Erickson, R.J. Stewart, J.R. Roppolo, andW.C. De Groat (1994) Localization of NADPH diaphorase in thelumbosacral spinal cord and dorsal root ganglia of the cat. J. Comp.Neurol. 339:62–75.
White, K.A., and M.A. Marletta (1992) Nitric oxide synthase is a cyto-chrome P-450 type hemoprotein. Biochemistry 31:6627–6631.
Wu, G., and J.T. Brosnan (1992) Macrophages can convert citrulline intoarginine. Biochem. J. 281:45–48.
626 H. NAKAMURA