myo-inositol-1 … guard cell movement, stress response, disease resistance, and membrane and cell...
TRANSCRIPT

myo-Inositol-1-phosphate Synthase Is Required for PolarAuxin Transport and Organ Development*□S
Received for publication, March 15, 2010, and in revised form, May 7, 2010 Published, JBC Papers in Press, June 1, 2010, DOI 10.1074/jbc.M110.123661
Hao Chen‡ and Liming Xiong‡§1
From the ‡Donald Danforth Plant Science Center, St. Louis, Missouri 63132 and the §Plant Stress Genomics Research Center andDivision of Chemical and Life Science and Engineering, King Abdullah University of Science and Technology,Thuwal 23955-6900, Saudi Arabia
myo-Inositol-1-phosphate synthase is a conserved enzymethat catalyzes the first committed and rate-limiting step in ino-sitol biosynthesis. Despite its wide occurrence in all eukaryotes,the role ofmyo-inositol-1-phosphate synthase and de novo ino-sitol biosynthesis in cell signaling and organism developmenthas been unclear. In this study, we isolated loss-of-functionmutants in theArabidopsisMIPS1 gene fromdifferent ecotypes.It was found that all nullmips1mutants are defective in embry-ogenesis, cotyledon venation patterning, root growth, and rootcap development. The mutant roots are also agravitropic andhave reduced basipetal auxin transport.mips1mutants have sig-nificantly reduced levels of major phosphatidylinositols andexhibit much slower rates of endocytosis. Treatment withbrefeldin A induces slower PIN2 protein aggregation in mips1,indicating altered PIN2 trafficking. Our results demonstratethat MIPS1 is critical for maintaining phosphatidylinositol lev-els and affects pattern formation in plants likely through regu-lation of auxin distribution.
Inositol is a precursor for many inositol-containing com-pounds that play critical and diverse roles in signal transduc-tion, membrane biogenesis, vesicle trafficking, and chroma-tin remodeling (1–3). Higher plants contain abundant inositoland inositol-derived compounds such as inositol phosphates,phosphatidylinositide, and sphingolipids. These compoundsare involved in gene expression modulation, hormonal signal-ing, guard cell movement, stress response, disease resistance,and membrane and cell wall biogenesis (4–9). Plant seeds alsocontain high levels of inositol hexakisphosphate (phytate),which serves as phosphorus storage, but it also is considered tobe an anti-nutrient because its phosphorus and the associatedminerals are not available to monogastric animals (10). Toreduce the phytate content in cereal seeds has been an objectiveactively pursued by the agrobiotechnology industry (10, 11).Inositol is synthesized from glucose 6-phosphate by 1L-myo-
inositol-1-phosphate synthase (also known as 1D-myo-inositol-3-phosphate synthase) (MIPS).2 The reaction generates inositol
1-phosphate, which is then dephosphorylated by an inositolmonophosphatase to form myo-inositol (12). Because MIPScatalyzes the rate-limiting step in inositol biosynthesis, MIPSmay be important in determining the pool size of inositol andits derivatives in cells. Nonetheless, inositol can also be recycledfrom inositol-containing compounds, and it is thus not clearwhether de novo inositol synthesis plays a regulatory role inmodulating phosphoinositide levels inmulticellular organisms.Furthermore, information about the role of MIPS in plantgrowth and development is also limited.Previously, plants with reduced MIPS transcript levels ob-
tained by antisense or RNA interference suppression werefound to be defective in embryo development in maize (13),seed development in soybean (14), or apical dominance inpotato (15). Recently, the role of inositol synthesis in cell deathand pathogen resistance was also identified (16–18). Here, wereport that null mutations in the inositol-phosphate synthaseMIPS1 gene in Arabidopsis have dramatic impacts on plantdevelopment, including impaired embryogenesis, altered coty-ledon development and venation pattern, distorted root caporganization, root agravitropism, and reduced basipetal trans-port of the phytohormone auxin. The mips1 mutants havereduced contents of major phosphatidylinositol species andaltered trafficking of the auxin efflux carrier PIN2. Our studythus establishes an essential role of MIPS1 in auxin transportand organ pattern formation. It also suggests that MIPS1 maynot be a good target for controlling phytate levels in seeds.
EXPERIMENTAL PROCEDURES
Plant Materials and Growth Conditions—Arabidopsis thali-ana ecotype Columbia-0 (Col-0) was used in all experimentsexcept where indicated. Themips1-4 T-DNA in the C24 back-ground was isolated in our genetic studies of abiotic stresssignal transduction. The mips1-2 (SALK_023626), mips1-5(CS851587), and mips2-1 (SALK_031685) T-DNA lines wereisolated from the Salk T-DNA insertion library. The eir1-1 andaux1-7mutants were obtained from theArabidopsis BiologicalResource Center (Columbus, OH). The homozygous mips1-2(SALK_023626) T-DNA line was used in all experiments unlessotherwise stated. The eir1 mips1-2 and aux1 mips1-2 doublemutants were selected by genotyping progenies of geneticcrossings between the respective single mutants. The derived
* This work was supported by National Science Foundation Grant 0446359(to L. X.).
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental Figs. S1–S6.
1 To whom correspondence should be addressed. Tel.: 966-02-808-2397;E-mail: [email protected].
2 The abbreviations used are: MIPS, myo-inositol-1-phosphate synthase; BFA,brefeldin A; TGN, trans-Golgi network; ARF, ADP-ribosylation factor; GAP,
guanosine triphosphatase-activating protein; MS, Murashige and Skoog;ER, endoplasmic reticulum; NAA, naphthalene acetic acid; 2,4-D, 2,4-di-chlorophenoxyacetic acid; WT, wild type; GUS, �-glucuronidase.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 31, pp. 24238 –24247, July 30, 2010© 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
24238 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 31 • JULY 30, 2010
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

cleaved amplified polymorphic sequences (dCAPS) primers5�-TTGTTGATCATTTTACCTGGGACA and 5�-GGTTG-CAATGCCATAAATAGAC (with BseLI restriction site) wereused for genotyping of eir1-1. Because mips1-2 single mutantseedlings do not have altered 2,4-dichlorophenoxyacetic acid(2,4-D) tolerance relative to the wild type, the aux1 mips1-2doublemutant was selected from the segregating F2 populationon 100 nM 2,4-D. Seeds were surface-sterilized with bleach andplanted onto half-strength Murashige and Skoog (MS) mediasupplemented with 3% sucrose and 0.6% agar. For rescue ofmips1 mutants, myo-inositol of various concentrations wasadded into the medium. After 2 days of cold treatment, plateswere incubated at 22 °C under constant white light for seedgermination and seedling growth. For root growth assays, seed-lings grown on plates with 1.2% agar were incubated verticallyand photographed using a digital camera. Soil-grown plantswere kept in a growth room at 22 °C with a 16-h light period.Plant Transformation—The MIPS1 cDNA was amplified
with primers 5�-CACCATGTTTATTGAGAGCTTCAAAGand 5�-CCATGATCATGTTGTTCTCC. The 1.1-kb MIPS1promoter was amplified with primers 5�-CACCACTAGTGT-AACTTGGAAAAGC and 5�-GCTTCTAATCAGCGGA-GAC. The PCR fragments were ligated into the pENTR-D-TOPO vector (Invitrogen). After sequence confirmation, theywere cloned into the pMDC Gateway vectors through LR clo-nase recombination. Agrobacterium tumefaciensGV3101 weretransformed by electroporation with these constructs. Four-week-oldArabidopsis plants were then vacuum-infiltratedwiththe transformed Agrobacterium strains, and transformantswere screened using the respective antibiotics.Phosphatidylinositol Analysis—Phospholipids were ex-
tracted and analyzed using an API 4000 liquid chromatogra-phy/tandem mass spectrometry system (Applied Biosystems,Foster City, CA) as described previously (19). Five replicateswere carried out with 10-day-old seedlings.Root Curling and Root Curvature Measurements—For root
curling assay, seeds were planted on a half-strength MS plateand vertically grown for 5 days. The plate was then laid flat for 2more days before taking pictures. Formeasurement of root cur-vature, the plates with vertically grown 5-day-old seedlingswere rotated 90o, and pictures were taken every hour. The rootcurvature was measured using the NIH imaging software. Rootrealignment assay and Mann-Whitney rank order sum testwere performed as described previously (20).Auxin Polar Transport Assay—Basipetal and acropetal auxin
polar transport assays were performed as described using 3H-labeled indole-3-acetic acid (American Radiolabeled Chemical,St. Louis, MO) (21). Three replicates, each with 10 seedlings,were carried out.RNA Analysis—Total RNA was extracted from 10-day-old
seedlings grown on half-strength MS agar media by usingthe TRIzol reagent according to the manufacturer’s manual(Invitrogen). For RNA blotting analysis, 10 �g of total RNAwere loaded onto 1.5% formaldehyde-agarose gels. MIPS1gene-specific probe was generated with �300-bp 3�-untrans-lated region fragment. RNA gel blots were autoradiographed toa phosphorimager screen (Amersham Biosciences).
Histochemical Staining—Seedlings or tissues from trans-genic plants were vacuumed in staining solution (25 mM so-dium phosphate buffer, pH 7.0, 10 mM EDTA, 0.5 mM ferricya-nide, 0.5 mM ferrocyanide, 0.1% Triton X-100, and 2 mM
5-bromo-4-chloro-3-indolyl�-D-glucuronide cyclohexylaminesalt) for 10min before staining at 37 °C. After staining, the solu-tion was replaced with 75% ethanol to remove chlorophyll.Confocal, Light, and Scanning Electron Microscopy—Whole
seedlings were mounted in water under glass coverslips forgreen fluorescent protein visualization using a confocal laserscanningmicroscope (NikonEclipse E-800C1) equippedwith akrypton-argon laser. Roots were stained with 10 �g per ml pro-pidium iodide for 2 min before taking root cap pictures usingthe same microscope.For FM4-64 (Invitrogen) uptake assay, 5-day-old seedlings
were incubated in 5 �M FM4-64 dye prepared in a liquid MSmedium for 5 min on ice, and images were taken immediatelyor after incubation in theMSmedium at room temperature for30 or 60min using a confocal microscope. For BFA treatments,5-day-old seedlings were first incubated in anMSmedium con-taining 50�MBFA for 2 h and then in anMSmediumwith 5�M
FM4-64 and 50�MBFA for 30min before taking pictures.Morethan 30 seedlings were examined for each treatment.The PIN2-GFP (22) or PIP2-GFP (23) transgene was intro-
duced into mips1-2 by crossing, and homozygous progenieswere observed for their green fluorescent protein fluorescenceusing a confocalmicroscope as described above. BFA treatmentof seedling was carried out at 50 �M for 30, 60, or 90 min, andimages were taken immediately or after washout in half-MS for2 h. More than 30 seedlings were examined for each treatment.Whole-mount preparation of ovules was conducted by incu-
bating the ovules in Hoyer’s solution (chloral hydrate/glycerol/water, 80:13:30 g) overnight at room temperature. To observethe cotyledon vein pattern, cotyledons from 7-day-old seed-lings were fixed in a 3:1 mixture of ethanol/acetic acid over-night. The seedlings were then cleared overnight in saturatedchloral hydrate. For starch granule staining, 3-day-old seedlingswere stained for 3 min with 1% Lugol’s solution (consists of0.05% iodine (I2) and 0.1% potassium iodide) and then rinsedwith water and cleared with saturated chloral hydrate briefly.Differential interference contrast images of cleared ovules, cot-yledons, and root caps were taken with a Nikon SMZ1500microscope and a QImaging (Burnaby, British Columbia, Can-ada) Retiga cooled camera.Scanning electron micrographs of 13-day-old cotyledons or
rosette leaves were taken with a Hitachi TM-1000 electronmicroscope. Stomatal primary and secondary complexes werecounted as described previously (24). More than 250 stomatalcomplexes from three representative seedlings were scored foreach genotype.
RESULTS
Arabidopsis MIPS1 Gene and Its Regulation—While con-ducting a genetic study of stress and abscisic acid signal trans-duction in Arabidopsis (25), we isolated a T-DNA insertionmutant in the C24 background with disruption in an inositol-phosphate synthase gene (MIPS1, At4g39800). In Arabidopsis,there are three MIPS homologs with more than 80% identity at
MIPS Required for Polar Auxin Transport, Organ Development
JULY 30, 2010 • VOLUME 285 • NUMBER 31 JOURNAL OF BIOLOGICAL CHEMISTRY 24239
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

the amino acid level:MIPS1 (AT4G39800) (also called IPS) (26),MIPS2/IPS2 (AT2G22240), and MIPS3/IPS3 (AT5G10170).The enzyme activity of MIPS1 in inositol biosynthesis has beendemonstrated by both in vitro assay and yeast complementa-tion (26). The publicArabidopsismicroarray databases indicatethat the transcript level of MIPS1 is predominant over that ofMIPS2 andMIPS3 at all development stages except seed matu-ration and early germination stages. At these stages, the tran-script level of MIPS2 is comparable with that of MIPS1,although the MIPS3 level is much lower at all developmentalstages (supplemental Fig. S1) (27, 28).To investigatewhether there is any organ-specific expression
of MIPS1, we checked the transcript levels of MIPS1 in roots,stems, leaves, flowers, and siliques. MIPS1 transcripts weredetected in all plant parts examined, with the highest level infloral tissues (Fig. 1A). Because inositol and its derivatives areknown to play roles in stress signal transduction and salt toler-ance (7, 8), we examined whetherMIPS1 is regulated by stress.However, we found that MIPS1 transcript levels were not reg-ulated by either salt stress or abscisic acid (data not shown).Transgenic plants expressing the MIPS1 promoter-driven
�-glucuronidase (GUS) reporter gene were generated to exam-ine the expression of MIPS1 at the tissue level. In young seed-lings, MIPS1 promoter activity was detected in cotyledons,
hypocotyls, and roots, with espe-cially strong signals in vascular tis-sues (Fig. 1, B–D). GUS expressionwas also observed in mature leafvascular tissues (Fig. 1E). In floralorgans, GUS expressionwas evidentin filaments (Fig. 1F), tips of siliques(Fig. 1G), and the joint region of sil-ique and pedicel (Fig. 1G).By fractionation and immunocy-
tochemistry analysis, Lackey et al.(29) showed that MIPS in Phaseolusvulgaris is widely distributed inintracellular compartments, includ-ing cytoplasm, membrane-boundorganelles, and cell walls as well. Onthe contrary, using immunogold-la-beled anti-MIPS antibody, Mitsu-hashi et al. (27) reported that MIPSproteins exclusively locate in thecytosol of developing Arabidopsisseeds. To investigate the MIPS1 lo-calization in planta, we generatedstable transgenic plants expressingthe GFP-MIPS1 fusion protein. Asshown in supplemental Fig. S2,A–C, MIPS1 was found to accumu-late mainly in the cytoplasm andpossibly at cellular membranes butnot in cell walls (supplementalFig. S2,D–F). To further investigatea possible membrane association ofMIPS1, an EYFP-MIPS1 constructwas co-infiltrated into Nicotiana
benthamiana leaves with the endoplasmic reticulum (ER)marker CFP-HDEL (30) or the Golgi marker GmMan1-CFP(31). The EYFP-MIPS1 showed a similar localization as theGFP-MIPS1 in Arabidopsis, i.e. in the cytoplasm and also likelyin membranes (supplemental Fig. S2,G andH). Although mul-tiphoton optical sections and time-lapse imaging found that theEYFP-MIPS1 signal was associated with the ERmarker in somesubcellular domains (supplemental Fig. S2K), the signal did notco-localize with the Golgi marker (supplemental Fig. S2L). InArabidopsis, the GFP-MIPS1 signal was also not internalizedupon BFA treatments (supplemental Fig. S2N), suggesting thatthe GFP-MIPS1 may be partly associated with the ER but notGolgi bodies.mips1 Mutants Exhibit Defects in Seedling Development
and Have Reduced Levels of Phosphatidylinositols—To investi-gate the role of MIPS in plant development, we identified addi-tional homozygous T-DNA insertional lines ofMIPS1 (Fig. 2A)and MIPS2 in the Columbia-0 (Col-0) background from theSALK T-DNA library (32). RNA analysis did not detect anyfull-lengthMIPS1 transcript in the mutant lines (Fig. 2B), indi-cating that these mutants are null for MIPS1. Along with themips1-4 mutant we isolated in the C24 background (Fig. 2B),these T-DNA lines were compared for their potential growthand developmental alterations. It was found that mips1
FIGURE 1. Expression pattern of MIPS1. A, MIPS1 transcript levels in different plant parts detected by RNA blotanalysis. Ten �g of total RNA from roots, stems, leaves, flowers, and siliques were loaded per lane.B–G, MIPS1::GUS expression patterns in whole seedling (B), cotyledon (C), root (D), mature leaf (E), floral organ(F), and silique (G). GUS staining was performed overnight.
MIPS Required for Polar Auxin Transport, Organ Development
24240 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 31 • JULY 30, 2010
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from
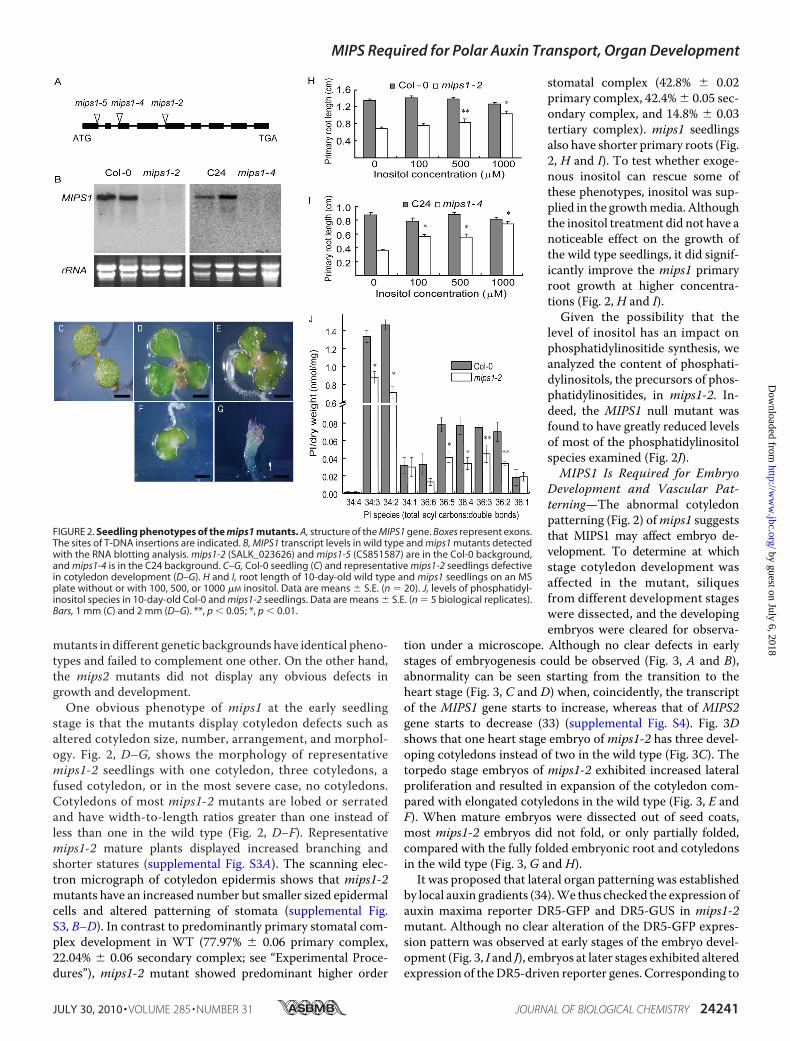
mutants in different genetic backgrounds have identical pheno-types and failed to complement one other. On the other hand,the mips2 mutants did not display any obvious defects ingrowth and development.One obvious phenotype of mips1 at the early seedling
stage is that the mutants display cotyledon defects such asaltered cotyledon size, number, arrangement, and morphol-ogy. Fig. 2, D–G, shows the morphology of representativemips1-2 seedlings with one cotyledon, three cotyledons, afused cotyledon, or in the most severe case, no cotyledons.Cotyledons of most mips1-2 mutants are lobed or serratedand have width-to-length ratios greater than one instead ofless than one in the wild type (Fig. 2, D–F). Representativemips1-2 mature plants displayed increased branching andshorter statures (supplemental Fig. S3A). The scanning elec-tron micrograph of cotyledon epidermis shows that mips1-2mutants have an increased number but smaller sized epidermalcells and altered patterning of stomata (supplemental Fig.S3, B–D). In contrast to predominantly primary stomatal com-plex development in WT (77.97% � 0.06 primary complex,22.04% � 0.06 secondary complex; see “Experimental Proce-dures”), mips1-2 mutant showed predominant higher order
stomatal complex (42.8% � 0.02primary complex, 42.4% � 0.05 sec-ondary complex, and 14.8% � 0.03tertiary complex). mips1 seedlingsalso have shorter primary roots (Fig.2, H and I). To test whether exoge-nous inositol can rescue some ofthese phenotypes, inositol was sup-plied in the growthmedia. Althoughthe inositol treatment did not have anoticeable effect on the growth ofthe wild type seedlings, it did signif-icantly improve the mips1 primaryroot growth at higher concentra-tions (Fig. 2, H and I).
Given the possibility that thelevel of inositol has an impact onphosphatidylinositide synthesis, weanalyzed the content of phosphati-dylinositols, the precursors of phos-phatidylinositides, in mips1-2. In-deed, the MIPS1 null mutant wasfound to have greatly reduced levelsof most of the phosphatidylinositolspecies examined (Fig. 2J).MIPS1 Is Required for Embryo
Development and Vascular Pat-terning—The abnormal cotyledonpatterning (Fig. 2) ofmips1 suggeststhat MIPS1 may affect embryo de-velopment. To determine at whichstage cotyledon development wasaffected in the mutant, siliquesfrom different development stageswere dissected, and the developingembryos were cleared for observa-
tion under a microscope. Although no clear defects in earlystages of embryogenesis could be observed (Fig. 3, A and B),abnormality can be seen starting from the transition to theheart stage (Fig. 3, C and D) when, coincidently, the transcriptof the MIPS1 gene starts to increase, whereas that of MIPS2gene starts to decrease (33) (supplemental Fig. S4). Fig. 3Dshows that one heart stage embryo ofmips1-2 has three devel-oping cotyledons instead of two in the wild type (Fig. 3C). Thetorpedo stage embryos of mips1-2 exhibited increased lateralproliferation and resulted in expansion of the cotyledon com-pared with elongated cotyledons in the wild type (Fig. 3, E andF). When mature embryos were dissected out of seed coats,most mips1-2 embryos did not fold, or only partially folded,compared with the fully folded embryonic root and cotyledonsin the wild type (Fig. 3, G and H).It was proposed that lateral organ patterning was established
by local auxin gradients (34).We thus checked the expression ofauxin maxima reporter DR5-GFP and DR5-GUS in mips1-2mutant. Although no clear alteration of the DR5-GFP expres-sion pattern was observed at early stages of the embryo devel-opment (Fig. 3, I and J), embryos at later stages exhibited alteredexpression of theDR5-driven reporter genes. Corresponding to
FIGURE 2. Seedling phenotypes of the mips1 mutants. A, structure of the MIPS1 gene. Boxes represent exons.The sites of T-DNA insertions are indicated. B, MIPS1 transcript levels in wild type and mips1 mutants detectedwith the RNA blotting analysis. mips1-2 (SALK_023626) and mips1-5 (CS851587) are in the Col-0 background,and mips1-4 is in the C24 background. C–G, Col-0 seedling (C) and representative mips1-2 seedlings defectivein cotyledon development (D–G). H and I, root length of 10-day-old wild type and mips1 seedlings on an MSplate without or with 100, 500, or 1000 �M inositol. Data are means � S.E. (n � 20). J, levels of phosphatidyl-inositol species in 10-day-old Col-0 and mips1-2 seedlings. Data are means � S.E. (n � 5 biological replicates).Bars, 1 mm (C) and 2 mm (D–G). **, p � 0.05; *, p � 0.01.
MIPS Required for Polar Auxin Transport, Organ Development
JULY 30, 2010 • VOLUME 285 • NUMBER 31 JOURNAL OF BIOLOGICAL CHEMISTRY 24241
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from
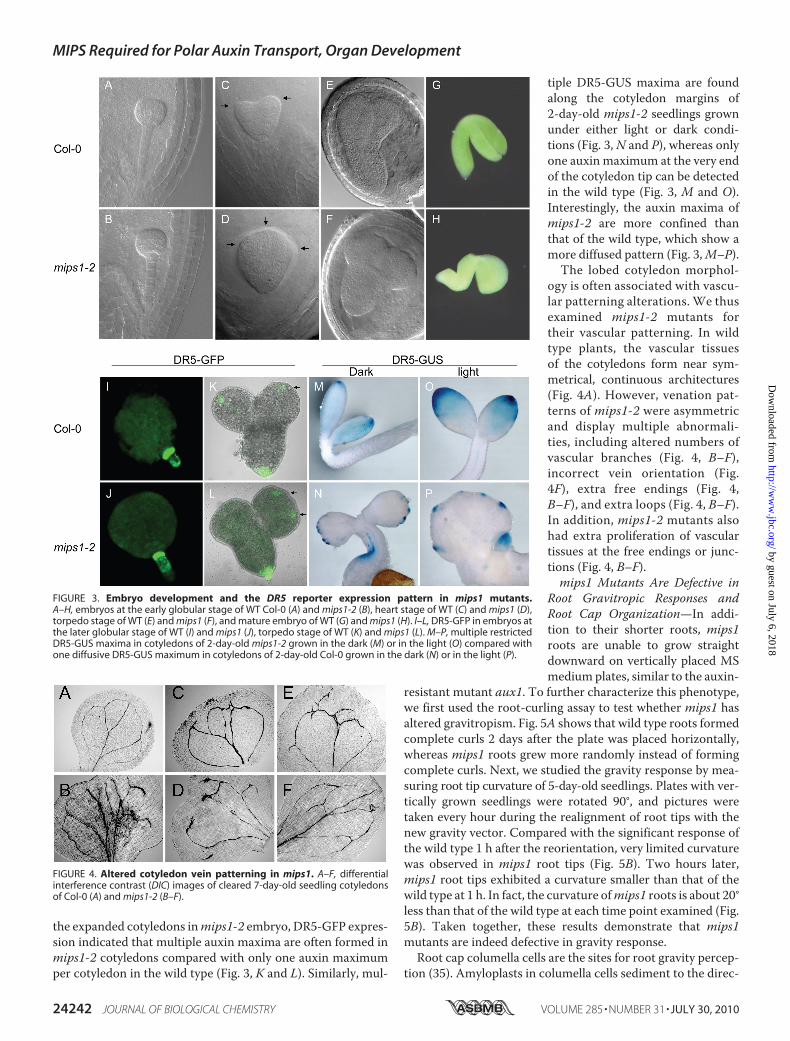
the expanded cotyledons inmips1-2 embryo, DR5-GFP expres-sion indicated that multiple auxin maxima are often formed inmips1-2 cotyledons compared with only one auxin maximumper cotyledon in the wild type (Fig. 3, K and L). Similarly, mul-
tiple DR5-GUS maxima are foundalong the cotyledon margins of2-day-old mips1-2 seedlings grownunder either light or dark condi-tions (Fig. 3, N and P), whereas onlyone auxin maximum at the very endof the cotyledon tip can be detectedin the wild type (Fig. 3, M and O).Interestingly, the auxin maxima ofmips1-2 are more confined thanthat of the wild type, which show amore diffused pattern (Fig. 3,M–P).
The lobed cotyledon morphol-ogy is often associated with vascu-lar patterning alterations. We thusexamined mips1-2 mutants fortheir vascular patterning. In wildtype plants, the vascular tissuesof the cotyledons form near sym-metrical, continuous architectures(Fig. 4A). However, venation pat-terns of mips1-2 were asymmetricand display multiple abnormali-ties, including altered numbers ofvascular branches (Fig. 4, B–F),incorrect vein orientation (Fig.4F), extra free endings (Fig. 4,B–F), and extra loops (Fig. 4, B–F).In addition, mips1-2 mutants alsohad extra proliferation of vasculartissues at the free endings or junc-tions (Fig. 4, B–F).mips1 Mutants Are Defective in
Root Gravitropic Responses andRoot Cap Organization—In addi-tion to their shorter roots, mips1roots are unable to grow straightdownward on vertically placed MSmediumplates, similar to the auxin-
resistant mutant aux1. To further characterize this phenotype,we first used the root-curling assay to test whether mips1 hasaltered gravitropism. Fig. 5A shows that wild type roots formedcomplete curls 2 days after the plate was placed horizontally,whereas mips1 roots grew more randomly instead of formingcomplete curls. Next, we studied the gravity response by mea-suring root tip curvature of 5-day-old seedlings. Plates with ver-tically grown seedlings were rotated 90°, and pictures weretaken every hour during the realignment of root tips with thenew gravity vector. Compared with the significant response ofthe wild type 1 h after the reorientation, very limited curvaturewas observed in mips1 root tips (Fig. 5B). Two hours later,mips1 root tips exhibited a curvature smaller than that of thewild type at 1 h. In fact, the curvature ofmips1 roots is about 20°less than that of the wild type at each time point examined (Fig.5B). Taken together, these results demonstrate that mips1mutants are indeed defective in gravity response.Root cap columella cells are the sites for root gravity percep-
tion (35). Amyloplasts in columella cells sediment to the direc-
FIGURE 3. Embryo development and the DR5 reporter expression pattern in mips1 mutants.A–H, embryos at the early globular stage of WT Col-0 (A) and mips1-2 (B), heart stage of WT (C) and mips1 (D),torpedo stage of WT (E) and mips1 (F), and mature embryo of WT (G) and mips1 (H). I–L, DR5-GFP in embryos atthe later globular stage of WT (I) and mips1 (J), torpedo stage of WT (K) and mips1 (L). M–P, multiple restrictedDR5-GUS maxima in cotyledons of 2-day-old mips1-2 grown in the dark (M) or in the light (O) compared withone diffusive DR5-GUS maximum in cotyledons of 2-day-old Col-0 grown in the dark (N) or in the light (P).
FIGURE 4. Altered cotyledon vein patterning in mips1. A–F, differentialinterference contrast (DIC) images of cleared 7-day-old seedling cotyledonsof Col-0 (A) and mips1-2 (B–F).
MIPS Required for Polar Auxin Transport, Organ Development
24242 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 31 • JULY 30, 2010
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

tion of gravity, which initiates an undefined signaling cascadethat eventually directs auxin flux toward the lower side of theroot presumably through PIN3 and AUX1 (36–38). Auxin inthe lower side of the root can be redistributed to the elongationzone by PIN2, where it inhibits cell elongation and leads to rootbending toward the gravity vector (36, 39, 40).We first checkedwhether starch granules are distorted in mips1. Staining withthe Lugol’s solution could detect starch granules in both wildtype and mips1-2 root columella cells, with a smaller regionstained in mips1-2 (Fig. 5, C–F), suggesting that the root capcolumella cell identity might not have been changed, and grav-ity sensing may still be normal inmips1-2. Nonetheless,mips1mutant roots had distorted root cap organization. Comparedwith the clear three-layered columella cells in thewild type root
cap (Fig. 5G), both the shape and thealignment of these cells in mips1-2are irregular, and no single recog-nizable columella cell layer couldbe identified (Fig. 5H). The sever-ity of these cell-patterning defectsin the root cap can be significantlymitigated by exogenous inositol orinositol 1-phosphate (supplemen-tal Fig. S5), although no significantrestoration of root gravitropismby inositol feeding was foundunder the current experimentalconditions (supplemental Fig. S6).
Because the auxin transportersAUX1 and PIN2 both are involvedin gravity response, to investigatethe extent to which the gravitro-pism alteration in mips1-2 is con-tributed by PIN2 or AUX1, we com-pared pin2 (eir1) or aux1 single andpin2 (eir1)mips1-2 or aux1mips1-2doublemutants for their root curva-ture response (Fig. 5I). Althougheir1 seedlings had a more severedefect in the assay thanmips1-2, nosignificant difference was observedbetween eir1 and eir1 mips1-2 dou-ble mutant (p value obtained fromMann-Whitney, two-tailed non-parametric test was 0.128, n � 105),indicating that MIPS1 and PIN2may act in the same pathway, andthe gravitropic defect in root bend-ing inmips1-2was largely caused byPIN2 dysfunction. On the contrary,aux1 and aux1mips1-2 differed sig-nificantly from each other (p �0.00013, n � 105). Although aux1 ismore agravitropic than mips1-2,surprisingly, aux1 mips1-2 doublemutant exhibited a gravitropicresponse similar to themips1-2 sin-gle mutant (p � 0.052, n � 105),
suggesting that the mips1 mutation partially suppressed aux1agravitropic response. Therefore, AUX1 andMIPS1 may act indistinct pathways in gravitropism, although the phenotypes ofthe respective mutants are partly overlapping. In addition, theaux1 mips1-2 double mutant has greatly reduced fertility (sil-ique length), although no difference was observed among wildtype, aux1,mips1-2, eir1, and eir1 mips1-2 (Fig. 5J and data notshown), indicating that AUX1 and MIPS1 may act in distinctpathways and in the mips1-2 mutant background, AUX1-me-diated auxin influx becomes limiting for fertility.Altered Auxin Transport and Response in mips1 Mutants—
The various auxin-related phenotypes of mips1 mutants de-scribed above suggest that the mutants may have changedauxin responsiveness or transport. We first measured auxin
FIGURE 5. Root gravitropism and columella cell patterning in mips1. A, root curling assay of the wild typeand mips1 seedlings. The pictures were taken 2 days after the plate was placed horizontally. B, root curvature of5-day-old wild type and mips1 roots. Shown are means � S.E. (n � 33). C and D, Lugol’s solution stained Col-0(C) and mips1-2 (D) root tips. E and F, high magnification of Col-0 (E) and mips1-2 (F) root cap stained withLugol’s solution. G and H, propidium iodide-stained Col-0 (G) and mips1-2 (H) root tips. QC, quiescent center; ci,columella initials; t1, t2, t3, the first three columella cell layers of the root cap. I, quantitative analyses of rootre-orientation of mips1, eir1, aux1 single and double mutants. Root angles were determined as the deviationfrom 0o representing complete re-orientation to the vertical and grouped in 12 sectors of 30o. Bars representrelative numbers of roots as percentage of the total (n). J, inflorescence of 6-week-old Col-0, aux1–7, mips1-2,and mips1 aux1 double mutant.
MIPS Required for Polar Auxin Transport, Organ Development
JULY 30, 2010 • VOLUME 285 • NUMBER 31 JOURNAL OF BIOLOGICAL CHEMISTRY 24243
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

transport rates in mips1 roots. Although no significant differ-ence in acropetal auxin transport was detected between thewild type andmips1-2 (Fig. 6B), basipetal auxin polar transportrate in mips1-2 roots was less than half that in wild type roots(Fig. 6A). To determine whether the mips1-2 mutants exhibitaltered responses to auxin, mutant and wild type seedlings withDR5-GUS transgenewere treatedwith different concentrationsof 2,4-D or naphthalene acetic acid (NAA). No clear differ-
ences in DR5-GUS induction by2,4-D were observed between wildtype andmips1-2, but we found thatmips1-2 mutant exhibited reducedinduction of reporter gene by NAA(Fig. 6,C andD). 2,4-D is a relativelypoor substrate for auxin efflux car-rier, although NAA can freely dif-fuse into the cell independent ofauxin influx carrier; therefore, re-duced response to NAA but intactresponse to 2,4-D induction ofDR5-GUS in mips1-2 may suggestthat rather than changed auxinresponse, mips1-2 may be morelikely impaired in its auxin effluxcapacity.mips1 Mutants Exhibit Defects in
Vesicle and PlasmaMembrane Pro-tein Trafficking—Previous studiesin animal cells have documentedthe pivotal role of phosphatidyli-nositides in many membrane traf-ficking events, including vesicletargeting, interactions between themembrane and the cytoskeleton,and membrane budding and fus-ing (2, 41). Given that auxin influxand efflux carriers undergo vesicle-dependent trafficking (42, 43) andthe fact that mips1 has reducedphosphatidylinositol contents (Fig.2J), it is likely that impaired vesicletrafficking might underlie the polarauxin transport-related phenotypesin mips1. We thus investigated thepossible impacts of mips1 muta-tions on vesicle trafficking.The fluorescent dye FM4-64 can
stain plasma membrane and hasbeen used as a marker to monitorthe endocytosis of plasma mem-brane. Thirty minutes after treat-ment, wild type root tips showedclear internalization of the dye (Fig.7, B and J). However, no apparentaccumulation of the dye was ob-served in mips1 roots (Fig. 7, F andN). One hour after the treatment,when large chunks of fluorescence
were evident in the wild type (Fig. 7, C and K),mips1 started toshow limited cytosolic endosome fluorescence (Fig. 7,G andO).Therefore, mips1 mutants have significantly slower endocyto-sis rates.BFA inhibits vesicle trafficking to and from the Golgi/TGN
and induces the formation of BFA compartments (44). Consis-tentwith its slowed endocytosis rate,mips1mutants also exhib-ited reduced BFA compartmentalization (Fig. 7,D,H, L, and P).
FIGURE 6. Impaired polar auxin transport and altered auxin response in mips1. A and B, relative rate ofbasipetal (A) and acropetal (B) auxin transport in mips1 roots. Data are means � S.E. (n � 3). C and D, DR5-GUSactivity (A.U., arbitrary units) in Col-0 and mips1-2 seedlings treated with different concentrations of 2,4-D (C) orNAA (D). Five-day-old seedlings were treated with the indicated concentrations of auxins for 24 h beforeextracting the protein for GUS assay. Data are means � S.E. (n � 3).
FIGURE 7. Impaired vesicle trafficking in mips1 mutants. Shown are fluorescence images of FM4-64 uptakein the root tips of Col-0 (A–D), mips1-2 (E–H), C24 (I–L), and mips1-4 (M–P). Roots of 5-day-old seedlings wereincubated in a half-strength MS medium containing 5 �M FM4-64 for 5 min on ice. Pictures were taken imme-diately or after 30 or 60 min of incubation in a half-strength MS solution at 22 °C. For BFA-induced compart-ments, 5-day-old seedlings were incubated in 50 �M BFA for 2 h before incubation in 50 �M BFA plus 5 �M
FM4-64 for 30 min. Pictures were taken immediately after the FM4-64 treatment. More than 30 seedlings wereexamined for each treatment. Bars, 25 �m.
MIPS Required for Polar Auxin Transport, Organ Development
24244 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 31 • JULY 30, 2010
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

The slower rate of endocytosis inmips1mutants may affect thetrafficking of membrane proteins, including auxin carriers. Totest this, we introduced the PIN2-GFP protein fusion constructinto themips1-2mutant plants by crossing with the PIN2-GFPline (22). Although 30 min of BFA treatment induces the for-mation of clear BFA compartments in the wild type seedlings(Fig. 8B), no clear BFA compartments were formed in themips1-2 mutant (Fig. 8G). With longer BFA treatment, one ortwo large BFA compartments were observed in the wild type(Fig. 8, C andD). Under the same conditions, smaller andmorenumerous BFA compartments were detected in mips1-2mutants than in the wild type (Fig. 8,H and I). Two hours afterwashout of BFA, PIN2 resumed its asymmetric localization inthe wild type (Fig. 8E), whereas in mips1-2 mutants, a certainamount of PIN2 remained in the cytosol (Fig. 8J). To testwhether the mips1 mutation affects the trafficking of othermembrane proteins, we checked another plasma membraneprotein PIP2-GFP (23) inmips1-2. Similar to PIN2 protein, theBFA-induced compartmentalization of PIP2 protein was alsoimpaired in mips1-2 (Fig. 8, K–N). Similar BFA compartmentresults have been reported by Sieburth et al. (45) in ARF-GAPmutant sfc for PIN1, suggesting thatmips1-2mutant may havesuppressed scarface activity and consequently impaired endo-cytosis of PINproteins.However, it is also possible that a step(s)before GNOMARF-GEFmay be affected inmips1 that leads tothe altered BFA compartment.
DISCUSSION
Inositol and its derivatives are essential for numerous cellularprocesses such as chromatin remodeling, gene regulation,mRNA export, hormone signaling, vesicle trafficking, stress
adaptation, and membrane and cellwall biogenesis (1–4). As a result,defects in MIPS or lack of inositolmay have dramatic impacts on cel-lular organisms. For instance, theTrypanosoma brucei mipsmutant isnonviable (46), and the Saccharo-myces cerevisiae ino1mutant is ino-sitol auxotrophic (47); yet, it isunclear which process is primarilyresponsible for the lethality in theseunicellular organisms. In this study,we isolated Arabidopsis MIPS1 nullmutants that allow us to reveal thein vivo role of this group of highlyconserved enzymes in regulatingvesicle trafficking and patterndevelopment in a higher eukaryote.MIPS1 Is Required for Multiple
Developmental Processes—Null mips1mutants exhibited multiple defectsin embryogenesis (Fig. 3,C–H), cot-yledon vein patterning (Fig. 4), epi-dermal cell division (supplementalFig. S3, B–D), root growth and grav-itropism (Fig. 2, H and I, and Fig. 5,A, B, and I), root cap cell patterning
(Fig. 5, C–H), apical dominance (supplemental Fig. S3A), andauxin response (Fig. 6, C andD). Similar to some of the pheno-types we observed, smaller seedling size, wrinkled seeds, irreg-ular cotyledons, altered vascular patterning, and altered epider-mal cell division in mips1 have also been reported recentlytogether with its compromised cell death suppression and basalimmunity (16, 18). The maize low phytic acid mutant lpa241with a reduced MIPS transcript level caused by an unknownepigenetic mutation(s) also exhibits defects related to embryodevelopment such as displacement of the scutellum and alteredsymmetry (13). Transgenic potato plants expressing antisenseMIPS have altered leaf morphology and reduced apical domi-nance (15). In addition, RNA interference silencing of soybeanGmMIPS1 leads to aborted seed development (14). Consistentwith this, mutants in inositol polyphosphate or phosphatidyl-inositolmetabolism also exhibit defects in vein formation, grav-itropic response, or auxin response (48, 49). Our currentgenetic study of the MIPS1 gene, together with previous geneexpression studies, demonstrate that de novo biosynthesis ofinositol is essential for normal embryo development and seed-ling growth. For this reason,MIPSmay not be a good target forengineering low phytate crops.There are some intriguing observations frommips1mutants.
First, some of these defects appear to occur at specific develop-mental stages. For instance, the defects in embryos weredetected relatively late in embryogenesis when theMIPS1 geneexpression starts to increase (supplemental Fig. S4). Perhaps atthese stages, the demand for de novo inositol biosynthesis fromMIPS1 is the highest. The absence of clear defects during earlyembryogenesis and the relatively normal epidermal cell prolif-eration and stomatal patterning in rosette leaves suggest that
FIGURE 8. Altered sorting of membrane proteins in mips1 mutants. A–J, PIN2 localization in root tips ofCol-0 (A–E) and mips1-2 (F–J). PIN2-GFP in root epidermal and cortical cells of Col-0 (A) and mips1 (F) withoutBFA treatment. PIN2 accumulation after 30, 60, and 90 min of BFA treatment in Col-0 (B–D) and mips1 (G–I).Note that as compared with one or two large PIN2 compartments in the Col-0 after 90 min of BFA treatment,multiple smaller PIN2 compartments are evident in mips1. Two hours after BFA washout, PIN2 polar localizationis reestablished in Col-0 (E), although considerable amount of PIN2 is still in the cytosol in mips1-2 (J). K–N, PIP2localization in root tips of Col-0 (K and L) and mips1-2 (M and N). PIP2-GFP in epidermal cells of Col-0 (K) andmips1-2 (M) without BFA treatment. PIP2 accumulation after 90 min of BFA treatment in Col-0 (L) and mips1-2(N). More than 30 seedlings were examined for each treatment. Bars, 20 �m.
MIPS Required for Polar Auxin Transport, Organ Development
JULY 30, 2010 • VOLUME 285 • NUMBER 31 JOURNAL OF BIOLOGICAL CHEMISTRY 24245
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

MIPS2, and perhaps also MIPS3, may play a redundant roleduring early embryogenesis. Second, although the MIPS1promoter GUS was expressed mainly in the root vasculature(Fig. 1, B–D), reduced basipetal rather than acropetal auxintransport was detected in mips1 roots (Fig. 6, A and B). Thismay be explained by the transport of MIPS1 product from vas-culature to out layers in root.Mitsuhashi et al. (27) also showedthat MIPS1 protein primarily located in endosperm but not inembryo, although mips1 mutant exhibits severe defects inembryo development (Fig. 3, B, D, F, and H). Finally, whereasexogenous inositol or inositol 1-phosphate partially rescuessome of the phenotypes of mips1 (Fig. 2, H and I, andsupplemental Fig. S5) it could not rescue the defects in auxinresponse in the gravitropism in roots (supplemental Fig. S6).The failure of rescue of this defect by inositol feeding mayreflect the limitation of the particular experimental conditionswhere these assays were conducted or, alternatively, MIPS1might have unknown functions independent of inositol biosyn-thesis. Interestingly, a recent study also reported that exoge-nous inositol could not substitute the loss ofMIPS1 gene in T.brucei (46).Altered Trafficking of PIN Proteins in mips1 Mutants—Al-
though a role of MIPS1 in vesicular trafficking has not previ-ously been reported in any organism, phosphatidylinositidesare widely known for their involvement in vesicular trafficking.Specific phosphatidylinositides are proposed to define theidentity of discrete vesicle trafficking organelles. The phosphor-ylation status of the polar heads of phosphatidylinositides inspecific intracellular locations may signal either the recruit-ment or the activation of proteins essential for vesicular trans-port (50). For example, plasma membrane is specificallyenriched in phosphatidylinositol 4,5-bisphosphate, and endo-somes are enriched in phosphatidylinositol 3-phosphate,whereas phosphatidylinositol 4-phosphate is the predominantGolgi membrane marker (51). Thus, the significantly reducedlevels of major phosphatidylinositols inmips1mutants (Fig. 2J)(16) may have important implications for vesicle trafficking. Itis also tempting to speculate that certain vesicle trafficking pro-cesses may be specifically affected due to the characteristicchanges in phosphatidylinositol species (Fig. 2J). Indeed, themips1 mutants have a slowed rate of FM4-64 internalizationand are defective in BFA compartmentalization (Fig. 7, A–P).
Consistent with impaired vesicle trafficking in mips1, theBFA compartment of PIN2 proteins (Fig. 8, A–J) is similar tothat of PIN1 protein in the sfc/van3 mutant (45). SFC/VAN3encodes an ADP-ribosylation factor-guanosine triphosphatase(GTPase)-activating protein (ARF-GAP) that bears a phospha-tidylinositol 4-phosphate-binding pleckstrin homology domainand is located in the TGN (52). AtSNX1, a component of retro-mer complex that retrieves protein from prevacuole compart-ment to the TGN, contains a phosphatidylinositol-binding PXdomain (53) and co-localizes with a marker for phosphatidyl-inositol 3-phosphate-enriched membrane subdomains (54).Therefore, the changed sorting of PIN2protein inmips1 ismostlikely linked to the significantly reduced content of phosphati-dylinositols, which may disrupt the homeostasis among thephosphatidylinositide species and consequently either in-creased sorting to vacuole or decreased retrieval to TGN or
both. Indeed, the regulation of SFC/VAN3 by phosphatidyli-nositide has recently been reported in Arabidopsis by geneticanalysis of sfc/van3 and inositol polyphosphate 5�-phospha-tases mutants cvp2 and cvl1 (55, 56).It worth noting that although mips1 mutant has changed
sorting of both PIN2 and PIP2 protein, it does not necessarilymean that this mutant has changed sorting of all plasma mem-brane proteins. Unlike PIN2 and PIP2, AUX1 sorting to vacuoleis not regulated by light (57), and our double mutant analysesalso indicate that MIPS1 and AUX1 act in distinct pathways(Fig. 5, I and J), suggesting AUX1 protein sorting may not bealtered in themips1mutant. We also cannot exclude the possi-bility that mips1 may be defective in auxin response indepen-dent of auxin transport, because inositol hexakisphosphate(phytate) might act as a cofactor of auxin receptor TIR1 (5),although supplementing the growth media or spraying withphytate could not rescue the mips1 mutant phenotypes (datanot shown). Notably, our discovery of MIPS regulation of vesi-cle trafficking and protein sorting in plantsmay be applicable toother eukaryotic systems as well. In fact, an inositol monophos-phatase that could function in both inositol biosynthesis andcycling was recently found to regulate the localization of syn-aptic components in the Caenorhabditis elegans nervous sys-tem (58), demonstrating that inositol is also required for pro-tein trafficking in animals.
Acknowledgments—We thank Drs. Jian Xu and Ben Scheres for thePIN2-GFP line and Dr. Thomas Guilfoyle for the DR5-GUS line. Wethank Dr. Ming Chen for providing ER and Golgi cyan fluorescentprotein marker plasmids, Dr. Howard Berg for helping with themicroscopy work, and Dr. Mark Running for comments on the exper-iments. The Kansas Lipidomics Research Center at the Kansas StateUniversity assisted with the phospholipid analysis.
REFERENCES1. Michell, R. H. (2008) Nat. Rev. Mol. Cell Biol. 9, 151–1612. Martin, T. F. (1998) Annu. Rev. Cell Dev. Biol. 14, 231–2643. Roth, M. G. (2004) Physiol. Rev. 84, 699–7304. Stevenson, J. M., Perera, I. Y., Heilmann, I., Persson, S., and Boss, W. F.
(2000) Trends Plant Sci. 5, 252–2585. Tan, X., Calderon-Villalobos, L. I., Sharon,M., Zheng, C., Robinson, C. V.,
Estelle, M., and Zheng, N. (2007) Nature 446, 640–6456. Nagy, R., Grob, H., Weder, B., Green, P., Klein, M., Frelet-Barrand, A.,
Schjoerring, J. K., Brearley, C., andMartinoia, E. (2009) J. Biol. Chem. 284,33614–33622
7. Xiong, L., Lee, Bh., Ishitani, M., Lee, H., Zhang, C., and Zhu, J. K. (2001)Genes Dev. 15, 1971–1984
8. Nelson, D. E., Rammesmayer, G., and Bohnert, H. J. (1998) Plant Cell 10,753–764
9. Lemtiri-Chlieh, F.,MacRobbie, E. A.,Webb, A. A.,Manison, N. F., Brown-lee, C., Skepper, J. N., Chen, J., Prestwich, G. D., and Brearley, C. A. (2003)Proc. Natl. Acad. Sci. U.S.A. 100, 10091–10095
10. Raboy, V. (2003) Phytochemistry 64, 1033–104311. Shi, J., Wang, H., Schellin, K., Li, B., Faller, M., Stoop, J. M., Meeley, R. B.,
Ertl, D. S., Ranch, J. P., and Glassman, K. (2007) Nat. Biotechnol. 25,930–937
12. Loewus, F. A., and Murthy, P. P. (2000) Plant Sci. 150, 1–1913. Pilu, R., Landoni,M., Cassani, E., Doria, E., andNielsen, E. (2005)Crop Sci.
45, 2096–210514. Nunes, A. C., Vianna, G. R., Cuneo, F., Amaya-Farfan, J., de Capdeville, G.,
Rech, E. L., and Aragao, F. J. (2006) Planta 224, 125–132
MIPS Required for Polar Auxin Transport, Organ Development
24246 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 31 • JULY 30, 2010
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

15. Keller, R., Brearley, C. A., Trethewey, R. N., and Muller-Rober, B. (1998)Plant J. 16, 403–410
16. Donahue, J. L., Alford, S. R., Torabinejad, J., Kerwin, R. E., Nourbakhsh, A.,Ray,W. K., Hernick, M., Huang, X., Lyons, B. M., Hein, P. P., and Gillaspy,G. E. (2010) Plant Cell 22, 888–903
17. Murphy, A.M., Otto, B., Brearley, C. A., Carr, J. P., andHanke, D. E. (2008)Plant J. 56, 638–652
18. Meng, P. H., Raynaud, C., Tcherkez, G., Blanchet, S., Massoud, K., Do-menichini, S., Henry, Y., Soubigou-Taconnat, L., Lelarge-Trouverie, C.,Saindrenan, P., Renou, J. P., and Bergounioux, C. (2009) PLoS ONE 4,e7364
19. Welti, R., Li, W., Li, M., Sang, Y., Biesiada, H., Zhou, H. E., Rajashekar,C. B., Williams, T. D., and Wang, X. (2002) J. Biol. Chem. 277,31994–32002
20. Men, S., Boutte, Y., Ikeda, Y., Li, X., Palme, K., Stierhof, Y. D., Hartmann,M. A., Moritz, T., and Grebe, M. (2008) Nat. Cell Biol. 10, 237–244
21. Rashotte, A. M., Brady, S. R., Reed, R. C., Ante, S. J., and Muday, G. K.(2000) Plant Physiol. 122, 481–490
22. Xu, J., and Scheres, B. (2005) Plant Cell 17, 525–53623. Cutler, S. R., Ehrhardt, D. W., Griffitts, J. S., and Somerville, C. R. (2000)
Proc. Natl. Acad. Sci. U.S.A. 97, 3718–372324. Kutter, C., Schob, H., Stadler,M.,Meins, F., Jr., and Si-Ammour, A. (2007)
Plant Cell 19, 2417–242925. Ishitani, M., Xiong, L., Stevenson, B., and Zhu, J. K. (1997) Plant Cell 9,
1935–194926. Johnson, M. D., and Sussex, I. M. (1995) Plant Physiol. 107, 613–61927. Mitsuhashi, N., Kondo, M., Nakaune, S., Ohnishi, M., Hayashi, M., Hara-
Nishimura, I., Richardson, A., Fukaki, H., Nishimura, M., andMimura, T.(2008) J. Exp. Bot. 59, 3069–3076
28. Zimmermann, P., Hirsch-Hoffmann, M., Hennig, L., and Gruissem, W.(2004) Plant Physiol. 136, 2621–2632
29. Lackey, K. H., Pope, P. M., and Johnson, M. D. (2003) Plant Physiol. 132,2240–2247
30. Chen, M., Han, G., Dietrich, C. R., Dunn, T. M., and Cahoon, E. B. (2006)Plant Cell 18, 3576–3593
31. Zeng, Q., Wang, X., and Running, M. P. (2007) Plant Physiol. 143,1119–1131
32. Alonso, J. M., Stepanova, A. N., Leisse, T. J., Kim, C. J., Chen, H., Shinn, P.,Stevenson, D. K., Zimmerman, J., Barajas, P., Cheuk, R., Gadrinab, C.,Heller, C., Jeske, A., Koesema, E., Meyers, C. C., Parker, H., Prednis, L.,Ansari, Y., Choy, N., Deen, H., Geralt, M., Hazari, N., Hom, E., Karnes,M.,Mulholland, C., Ndubaku, R., Schmidt, I., Guzman, P., Aguilar-Henonin,L., Schmid, M., Weigel, D., Carter, D. E., Marchand, T., Risseeuw, E.,Brogden, D., Zeko, A., Crosby, W. L., Berry, C. C., and Ecker, J. R. (2003)Science 301, 653–657
33. Winter, D., Vinegar, B., Nahal, H., Ammar, R., Wilson, G. V., and Provart,
N. J. (2007) PLoS ONE 2, e71834. Benkova, E., Michniewicz, M., Sauer, M., Teichmann, T., Seifertova, D.,
Jurgens, G., and Friml, J. (2003) Cell 115, 591–60235. Blancaflor, E. B., Fasano, J. M., and Gilroy, S. (1999) Adv. Space Res. 24,
731–73836. Chen, R., Guan, C., Boonsirichai, K., and Masson, P. H. (2002) Plant Mol.
Biol. 49, 305–31737. Ottenschlager, I., Wolff, P., Wolverton, C., Bhalerao, R. P., Sandberg, G.,
Ishikawa, H., Evans, M., and Palme, K. (2003) Proc. Natl. Acad. Sci. U.S.A.100, 2987–2991
38. Friml, J., Vieten, A., Sauer, M., Weijers, D., Schwarz, H., Hamann, T.,Offringa, R., and Jurgens, G. (2003) Nature 426, 147–153
39. Swarup, R., Friml, J., Marchant, A., Ljung, K., Sandberg, G., Palme, K., andBennett, M. (2001) Genes Dev. 15, 2648–2653
40. Muday, G. K., and DeLong, A. (2001) Trends Plant Sci. 6, 535–54241. Yin, H. L., and Janmey, P. A. (2003) Annu. Rev. Physiol. 65, 761–78942. Geldner, N., Anders, N., Wolters, H., Keicher, J., Kornberger, W., Muller,
P., Delbarre, A., Ueda, T., Nakano, A., and Jurgens, G. (2003) Cell 112,219–230
43. Steinmann, T., Geldner, N., Grebe, M., Mangold, S., Jackson, C. L., Paris,S., Galweiler, L., Palme, K., and Jurgens, G. (1999) Science 286, 316–318
44. Nebenfuhr, A., Ritzenthaler, C., and Robinson, D. G. (2002) Plant Physiol.130, 1102–1108
45. Sieburth, L. E., Muday, G. K., King, E. J., Benton, G., Kim, S., Metcalf, K. E.,Meyers, L., Seamen, E., and Van Norman, J. M. (2006) Plant Cell 18,1396–1411
46. Martin, K. L., and Smith, T. K. (2006)Mol. Microbiol. 61, 89–10547. Culbertson,M. R., Donahue, T. F., andHenry, S. A. (1976) J. Bacteriol. 126,
232–24248. Li, G., and Xue, H. W. (2007) Plant Cell 19, 281–29549. Carland, F. M., and Nelson, T. (2004) Plant Cell 16, 1263–127550. De Camilli, P., Emr, S. D., McPherson, P. S., and Novick, P. (1996) Science
271, 1533–153951. Di Paolo, G., and De Camilli, P. (2006) Nature 443, 651–65752. Koizumi, K., Naramoto, S., Sawa, S., Yahara, N., Ueda, T., Nakano, A.,
Sugiyama, M., and Fukuda, H. (2005) Development 132, 1699–171153. Vanoosthuyse, V., Tichtinsky, G., Dumas, C., Gaude, T., and Cock, J. M.
(2003) Plant Physiol. 133, 919–92954. Kleine-Vehn, J., Leitner, J., Zwiewka,M., Sauer, M., Abas, L., Luschnig, C.,
and Friml, J. (2008) Proc. Natl. Acad. Sci. U.S.A. 105, 17812–1781755. Carland, F., and Nelson, T. (2009) Plant J. 59, 895–90756. Naramoto, S., Sawa, S., Koizumi, K., Uemura, T., Ueda, T., Friml, J., Na-
kano, A., and Fukuda, H. (2009) Development 136, 1529–153857. Laxmi, A., Pan, J., Morsy, M., and Chen, R. (2008) PLoS ONE 3, e151058. Tanizawa, Y., Kuhara, A., Inada, H., Kodama, E., Mizuno, T., and Mori, I.
(2006) Genes Dev. 20, 3296–3310
MIPS Required for Polar Auxin Transport, Organ Development
JULY 30, 2010 • VOLUME 285 • NUMBER 31 JOURNAL OF BIOLOGICAL CHEMISTRY 24247
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from

Hao Chen and Liming XiongOrgan Development
-Inositol-1-phosphate Synthase Is Required for Polar Auxin Transport andmyo
doi: 10.1074/jbc.M110.123661 originally published online June 1, 20102010, 285:24238-24247.J. Biol. Chem.
10.1074/jbc.M110.123661Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2010/06/01/M110.123661.DC1
http://www.jbc.org/content/285/31/24238.full.html#ref-list-1
This article cites 58 references, 31 of which can be accessed free at
by guest on July 6, 2018http://w
ww
.jbc.org/D
ownloaded from