muscle - university of · pdf filethe term contraction, as used in muscle physiology, does not...
TRANSCRIPT

www.mhhe.com/widmaier9
MUSCLE
SECTION A Skeletal MuscleStructureMolecular Mechanisms of Contraction
Sliding-Filament MechanismRoles of Troponin, Tropomyosin, and Calcium
in ContractionExcitation-Contraction CouplingMembrane Excitation: The Neuromuscular
JunctionMechanics of Single-Fiber Contraction
Twitch ContractionsLoad-Velocity RelationFrequency-Tension RelationLength-Tension Relation
Skeletal Muscle Energy MetabolismMuscle Fatigue
Types of Skeletal Muscle FibersWhole-Muscle Contraction
Control of Muscle TensionControl of Shortening VelocityMuscle Adaptation to ExerciseLever Action of Muscles and Bones
Additional Clinical ExamplesMuscle CrampsHypocalcemic TetanyMuscular DystrophyMyasthenia Gravis
S E C T I O N A S U M M A R Y
S E C T I O N A K E Y T E R M S
S E C T I O N A C L I N I C A L T E R M S
S E C T I O N A R E V I E W Q U E S T I O N S
SECTION BSmooth MuscleStructure of Smooth MuscleSmooth Muscle Contraction and Its
ControlCross-Bridge ActivationSources of Cytosolic CalciumMembrane ActivationTypes of Smooth Muscle
S E C T I O N B S U M M A R Y
S E C T I O N B K E Y T E R M S
S E C T I O N B R E V I E W Q U E S T I O N S
C H A P T E R 9 T H O U G H T Q U E S T I O N S
9C H A P T E R N I N E
267
Color-enhanced electron microscope image of skeletal muscle sarcomeres

The ability to use chemical energy to produce force and
movement is present to a limited extent in most cells,
but in muscle cells it has become dominant. Muscles
generate force and movements used in the regulation of the
internal environment, and they also produce movements in
the external environment. In humans, the ability to
communicate, whether by speech, writing, or artistic
expression, also depends on muscle contractions. Indeed, it
is only by controlling the activity of muscles that the human
mind ultimately expresses itself.
Three types of muscle tissue can be identified on the
basis of structure, contractile properties, and control
mechanisms: (1) skeletal muscle, (2) smooth muscle, and
(3) cardiac muscle. Most skeletal muscle, as the name
implies, is attached to bone, and its contraction is responsible
for supporting and moving the skeleton. The contraction of
skeletal muscle is initiated by impulses in the neurons to the
muscle and is usually under voluntary control.
Sheets of smooth muscle surround various hollow
organs and tubes, including the stomach, intestines, urinary
bladder, uterus, blood vessels, and airways in the lungs.
Contraction of the smooth muscle surrounding hollow
organs may propel the luminal contents through the organ,
or it may regulate internal flow by changing the tube
diameter. In addition, small bundles of smooth muscle cells
are attached to the hairs of the skin and iris of the eye.
Smooth muscle contraction is controlled by the autonomic
nervous system, hormones, autocrine/paracrine agents, and
other local chemical signals. Some smooth muscles contract
autonomously, however, even in the absence of such signals.
In contrast to skeletal muscle, smooth muscle is not
normally under voluntary control.
Cardiac muscle is the muscle of the heart. Its
contraction propels blood through the circulatory system.
Like smooth muscle, it is regulated by the autonomic
nervous system, hormones, and autocrine/paracrine agents,
and it can undergo spontaneous contractions.
Although there are significant differences in these three
types of muscle, the force-generating mechanism is similar
in all of them. Skeletal muscle will be described first,
followed by a discussion of smooth muscle. Cardiac muscle,
which combines some of the properties of both skeletal and
smooth muscle, will be described in Chapter 12 in
association with its role in the circulatory system.
occurs through an increase in the size of the remainingmuscle fibers (hypertrophy).
The term muscle refers to a number of muscle fibersbound together by connective tissue (Figure 9–1). Therelationship between a single muscle fiber and a mus-cle is analogous to that between a single neuron and anerve, which is composed of the axons of many neu-rons. Muscles are usually linked to bones by bundles ofcollagen fibers known as tendons.
In some muscles the individual fibers extend the en-tire length of the muscle, but in most, the fibers areshorter, often oriented at an angle to the longitudinal axisof the muscle. The transmission of force from muscle tobone is like a number of people pulling on a rope, eachperson corresponding to a single muscle fiber and therope corresponding to the connective tissue and tendons.
Some tendons are very long, with the site of tendonattachment to bone far removed from the end of themuscle. For example, some of the muscles that movethe fingers are in the forearm (wiggle your fingers andfeel the movement of the muscles in your lower arm).These muscles are connected to the fingers by longtendons.
268
STRUCTUREA single skeletal muscle cell is known as a muscle fiber.Each muscle fiber is formed during development by thefusion of a number of undifferentiated, mononucleatedcells, known as myoblasts, into a single cylindrical,multinucleated cell. Skeletal muscle differentiation iscompleted around the time of birth, and these differen-tiated fibers continue to increase in size during growthfrom infancy to adult stature, but no new fibers areformed from myoblasts. Adult skeletal muscle fibershave diameters between 10 and 100 �m and lengths thatmay extend up to 20 cm.
If skeletal muscle fibers are destroyed after birth asa result of injury, they cannot be replaced by the divi-sion of other existing muscle fibers. New fibers can beformed, however, from undifferentiated cells known assatellite cells, which are located adjacent to the musclefibers and undergo differentiation similar to that fol-lowed by embryonic myoblasts. This capacity for form-ing new skeletal muscle fibers is considerable but willnot restore a severely damaged muscle to full strength.Much of the compensation for a loss of muscle tissue
S E C T I O N A
S k e l e t a l M u s c l e

www.mhhe.com/widmaier9 269
The most striking feature seen when observingskeletal or cardiac muscle through a light microscope(Figure 9–2) is a series of light and dark bands perpendicular to the long axis. Because of this charac-teristic striped pattern, both types are known as stri-ated muscle (Figure 9–3). Smooth muscle cells do notshow a banding pattern. The striated pattern in skele-tal and cardiac muscle results from the arrangement ofnumerous thick and thin filaments in the cytoplasminto approximately cylindrical bundles (1 to 2 �m indiameter) known as myofibrils (Figure 9–4). Most ofthe cytoplasm of a fiber is filled with myofibrils, eachof which extends from one end of the fiber to the otherand is linked to the tendons at the ends of the fiber.
The thick and thin filaments in each myofibril(Figures 9–4 and 9–5) are arranged in a repeating pat-tern along the length of the myofibril. One unit of thisrepeating pattern is known as a sarcomere (Greek, sarco,
Musclefiber
Muscle
Blood vessel
Connective tissue
Tendons
FIGURE 9–1Organization of cylindrical skeletal muscle fibers in a muscle that is attached to bones by tendons.
muscle; mer, part). The thick filaments are composedalmost entirely of the contractile protein myosin.The thin filaments (which are about half the diameterof the thick filaments) contain the contractile proteinactin, as well as two other proteins—troponin andtropomyosin—that play important roles in regulatingcontraction.
The thick filaments are located in the middle of eachsarcomere, where their orderly parallel arrangementproduces a wide, dark band known as the A band(Figure 9–4). Each sarcomere contains two sets of thinfilaments, one at each end. One end of each thin fila-ment is anchored to a network of interconnectingproteins known as the Z line, whereas the other endoverlaps a portion of the thick filaments. Two succes-sive Z lines define the limits of one sarcomere. Thus,thin filaments from two adjacent sarcomeres are an-chored to the two sides of each Z line.
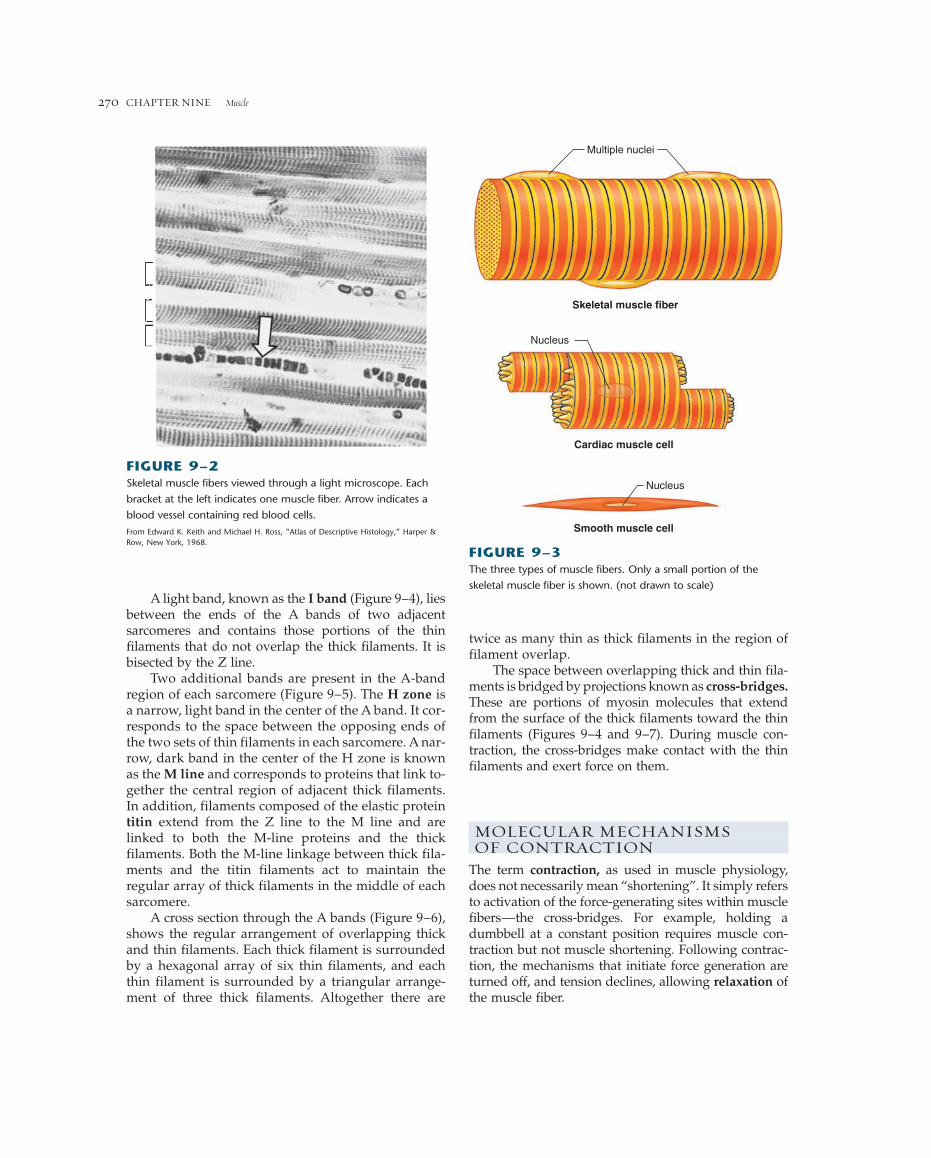
A light band, known as the I band (Figure 9–4), liesbetween the ends of the A bands of two adjacentsarcomeres and contains those portions of the thinfilaments that do not overlap the thick filaments. It isbisected by the Z line.
Two additional bands are present in the A-bandregion of each sarcomere (Figure 9–5). The H zone isa narrow, light band in the center of the A band. It cor-responds to the space between the opposing ends ofthe two sets of thin filaments in each sarcomere. A nar-row, dark band in the center of the H zone is knownas the M line and corresponds to proteins that link to-gether the central region of adjacent thick filaments.In addition, filaments composed of the elastic proteintitin extend from the Z line to the M line and arelinked to both the M-line proteins and the thick filaments. Both the M-line linkage between thick fila-ments and the titin filaments act to maintain the regular array of thick filaments in the middle of eachsarcomere.
A cross section through the A bands (Figure 9–6),shows the regular arrangement of overlapping thickand thin filaments. Each thick filament is surroundedby a hexagonal array of six thin filaments, and eachthin filament is surrounded by a triangular arrange-ment of three thick filaments. Altogether there are
CHAPTER NINE Muscle270
Multiple nuclei
Skeletal muscle fiber
Cardiac muscle cell
Nucleus
Nucleus
Smooth muscle cell
FIGURE 9–3The three types of muscle fibers. Only a small portion of the
skeletal muscle fiber is shown. (not drawn to scale)
twice as many thin as thick filaments in the region offilament overlap.
The space between overlapping thick and thin fila-ments is bridged by projections known as cross-bridges.These are portions of myosin molecules that extendfrom the surface of the thick filaments toward the thinfilaments (Figures 9–4 and 9–7). During muscle con-traction, the cross-bridges make contact with the thinfilaments and exert force on them.
MOLECULAR MECHANISMS OF CONTRACTION
The term contraction, as used in muscle physiology,does not necessarily mean “shortening”. It simply refersto activation of the force-generating sites within musclefibers—the cross-bridges. For example, holding adumbbell at a constant position requires muscle con-traction but not muscle shortening. Following contrac-tion, the mechanisms that initiate force generation areturned off, and tension declines, allowing relaxation ofthe muscle fiber.
FIGURE 9–2Skeletal muscle fibers viewed through a light microscope. Each
bracket at the left indicates one muscle fiber. Arrow indicates a
blood vessel containing red blood cells.
From Edward K. Keith and Michael H. Ross, “Atlas of Descriptive Histology,” Harper &Row, New York, 1968.

www.mhhe.com/widmaier9 271
Sliding-Filament MechanismWhen force generation produces shortening of a skele-tal muscle fiber, the overlapping thick and thin filamentsin each sarcomere move past each other, propelled bymovements of the cross-bridges. During this shorteningof the sarcomeres, there is no change in the lengths ofeither the thick or thin filaments (Figure 9–8). This isknown as the sliding-filament mechanism of musclecontraction.
During shortening, each myosin cross-bridge at-tached to a thin filament actin molecule moves in an arcmuch like an oar on a boat. This swiveling motion ofmany cross-bridges forces the thin filaments attached tosuccessive Z lines toward the center of the sarcomere,thereby shortening the sarcomere (Figure 9–9). Onestroke of a cross-bridge produces only a very small
movement of a thin filament relative to a thick filament.As long as a muscle fiber remains activated, however,each cross-bridge repeats its swiveling motion manytimes, resulting in large displacements of the filaments.Thus, the ability of a muscle fiber to generate force andmovement depends on the interaction of the contractileproteins actin and myosin.
An actin molecule is a globular protein composedof a single polypeptide that polymerizes with otheractins to form two intertwined helical chains (Figure9–10). These chains make up the core of a thin fila-ment. Each actin molecule contains a binding site formyosin. The myosin molecule, on the other hand, iscomposed of two large polypeptide heavy chains andfour smaller light chains. These polypeptides com-bine to form a molecule that consists of two globularheads (containing heavy and light chains) and a long
Muscle fiberMyofibril
I band A band
Z line Z line
Sarcomere
Myofibril
Z line Z line
Cross-bridge
Thick (myosin) filament Thin (actin) filament
FIGURE 9–4Arrangement of filaments in a skeletal muscle fiber that produces the striated banding pattern.
CHAPTER NINE Muscle272
(a)
(b)
M lineThin filament Thick filamentTitin
filament
Sarcomere
H zone
A band
Z line Z line
(c)
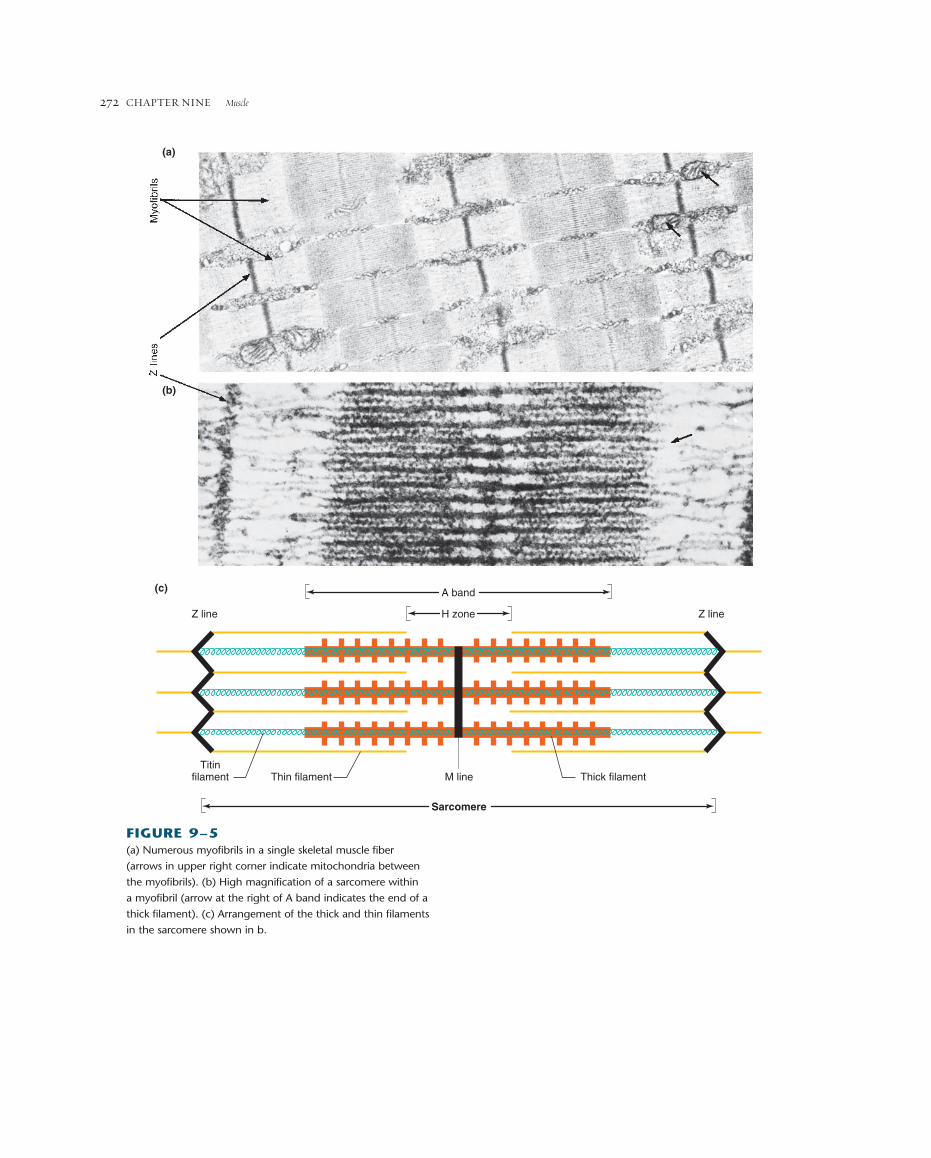
FIGURE 9–5(a) Numerous myofibrils in a single skeletal muscle fiber
(arrows in upper right corner indicate mitochondria between
the myofibrils). (b) High magnification of a sarcomere within
a myofibril (arrow at the right of A band indicates the end of a
thick filament). (c) Arrangement of the thick and thin filaments
in the sarcomere shown in b.

www.mhhe.com/widmaier9 273
Myo
fibril
Myo
fibril
Thickfilament
Thinfilament
(a) (b) Thick filament Thin filament
FIGURE 9–6(a) Electron micrograph of a cross section through several
myofibrils in a single skeletal muscle fiber.
From H. E. Huxley, J. Mol. Biol., 37:507–520 (1968).
(b) Hexagonal arrangements of the thick and thin filaments in the
overlap region in a single myofibril. Six thin filaments surround
each thick filament, and three thick filaments surround each thin
filament.
Cross-bridges
FIGURE 9–7High-magnification electron micrograph in the filament
overlap region near the middle of a sarcomere. Cross-bridges
between the thick and thin filaments can be seen at regular
intervals along the filaments.
From H. E. Huxley and J. Hanson, in G. H. Bourne (ed.), “The Structure and Function of Muscle,” Vol. 1, Academic Press, New York, 1960.

The myosin molecules in the two ends of each thickfilament are oriented in opposite directions, such thattheir tail ends are directed toward the center of the fil-ament (Figure 9–11a). Because of this arrangement, thepower strokes of the cross-bridges move the attachedthin filaments at the two ends of the sarcomere towardthe center during shortening (see Figure 9–9).
The sequence of events that occurs between the timea cross-bridge binds to a thin filament, moves, and thenis set to repeat the process is known as a cross-bridgecycle. Each cycle consists of four steps: (1) attachment of
CHAPTER NINE Muscle274
tail formed by the two intertwined heavy chains(Figure 9–11b). The tail of each myosin molecule liesalong the axis of the thick filament, and the two glob-ular heads extend out to the sides, forming the cross-bridges. Each globular head contains two bindingsites, one for actin and one for ATP. The ATP bindingsite also serves as an enzyme—an ATPase that hy-drolyzes the bound ATP.
Relaxed(a)
Shortened(b)
A band H zoneI band
H zonereducedI band
reduced
Z-line
Z-line Z-line
A bandunchanged
FIGURE 9–8The sliding of thick filaments past overlapping thin filaments produces sarcomere shortening with no change in thick or thin filament
length. The I band and H zone are reduced.
Z line Z lineCross-bridgemovement
Thin filament T
FIGURE 9–9Cross-bridges in the thick filaments bind to actin in the thin
filaments and undergo a conformational change that propels the
thin filaments toward the center of a sarcomere. (Only two of the
approximately 200 cross-bridges in each thick filament are
shown.)
Thin filament
Actin molecule
FIGURE 9–10Two intertwined helical chains of actin molecules form the
primary structure of the thin filaments.

www.mhhe.com/widmaier9 275
Thickfilament
Cross-bridge
ATP binding site ATPbindingsiteLight chains
Heavy chains
Actin binding sites
Cross- bridge
(a)
(b)
Myosin
FIGURE 9–11(a) The heavy chains of myosin molecules form the core of a thick filament. The myosin molecules are oriented in opposite directions in
either half of a thick filament. (b) Structure of a myosin molecule. The two globular heads of each myosin molecule extend from the
sides of a thick filament, forming a cross-bridge.
the cross-bridge to a thin filament, (2) movement of thecross-bridge, producing tension in the thin filament, (3)detachment of the cross-bridge from the thin filament,and (4) energizing the cross-bridge so that it can againattach to a thin filament and repeat the cycle. Eachcross-bridge undergoes its own cycle of movement independently of the other cross-bridges. At any instantduring contraction only a portion of the cross-bridgesare attached to the thin filaments and producing tension,while others are in a detached portion of their cycle.
The chemical and physical events during the foursteps of the cross-bridge cycle are illustrated in Figure9–12. In a resting muscle fiber the cytoplasmic calciumconcentration is low, and the myosin cross-bridges (M)cannot bind to actin (A). The cross-bridges, however, arein an energized state produced by the splitting of ATP,and the hydrolysis products (ADP and inorganic phos-phate) are still bound to myosin. This storage of energyin myosin is analogous to the storage of potential en-ergy in a stretched spring.
Cross-bridge cycling is initiated by calcium entryinto the cytoplasm (by a mechanism that will be de-scribed shortly). The cycle begins with the binding ofan energized myosin cross-bridge to a thin filamentactin molecule (step 1):
Step 1 A � M�ADP�Pi 88n A�M�ADP�PiActin
binding
The binding of energized myosin to actin triggersthe release of the strained conformation of the energized
bridge, which produces the movement of the boundcross-bridge (sometimes called the power stroke) andthe release of Pi and ADP (step2):
Step 2 A�M�ADP�Pi 88n A�M � ADP � PiCross-bridgemovement
This sequence of energy storage and release by myosinis analogous to the operation of a mousetrap: Energy isstored in the trap by cocking the spring (ATP hydroly-sis) and released after springing the trap (binding toactin).
During the cross-bridge movement, myosin isbound very firmly to actin, and this linkage must be bro-ken in order to allow the cross-bridge to be re-energizedand repeat the cycle. The binding of a new molecule ofATP to myosin breaks the link between actin andmyosin (step 3):
Step 3 A�M � ATP 88n A � M�ATPCross-bridge
dissociation from actin
The dissociation of actin and myosin by ATP is an ex-ample of allosteric regulation of protein activity. Thebinding of ATP at one site on myosin decreases myosin’saffinity for actin bound at another site. Note that ATPis not split in this step; that is, it is not acting as an en-ergy source but only as an allosteric modulator of themyosin head that weakens the binding of myosin toactin.

movement, and (2) ATP binding (not hydrolysis) tomyosin breaks the link formed between actin and myosinduring the cycle, allowing the cycle to be repeated.
The importance of ATP in dissociating actin andmyosin during step 3 of a cross-bridge cycle is illus-trated by rigor mortis, the stiffening of skeletal musclesthat begins several hours after death and is completeafter about 12 h. The ATP concentration in cells, in-cluding muscle cells, declines after death because thenutrients and oxygen required by the metabolic path-ways to form ATP are no longer supplied by the circu-lation. In the absence of ATP, the breakage of the linkbetween actin and myosin does not occur. The thick andthin filaments remain bound to each other by immobi-lized cross-bridges, producing a rigid condition in
CHAPTER NINE Muscle276
Following the dissociation of actin and myosin, theATP bound to myosin is split (step 4), thereby re-forming the energized state of myosin.
Step 4 A � M�ATP 88n A � M�ADP�PiATP hydrolysis
If calcium is still present at this time, the cross-bridgecan reattach to a new actin molecule in the thin filamentand the cross-bridge cycle repeats. Note that the hy-drolysis of ATP (step 4) and the movement of the cross-bridge (step 2) are not simultaneous events.
To summarize, ATP performs two distinct roles in thecross-bridge cycle: (1) The energy released from ATP hy-drolysis ultimately provides the energy for cross-bridge
Cross-bridgebinds to actin
Cross-bridgemovesADP + Pi
ATP binds to myosin, causing cross-bridge to detach
Energizedcross-bridge
Thin filament (actin, A)
Thick filament (myosin, M)M line
[A + M • ADP • Pi ]Z line
[A • M • ADP • Pi ]
[A • M]
Hydrolysisof ATPenergizescross-bridge
[A + M • ATP] ATP
4 2
3
1
[Ca2+] rises
ADPPi
ADPPi
ATP
FIGURE 9–12Chemical (shown in brackets) and mechanical representations of the four stages of a cross-bridge cycle. In a resting muscle fiber,
contraction begins when calcium activates the thin filament.

www.mhhe.com/widmaier9 277
which the thick and thin filaments cannot be pulled pasteach other. The stiffness of rigor mortis disappears about48 to 60 h after death as a result of the disintegration ofmuscle tissue.
Roles of Troponin, Tropomyosin, andCalcium in ContractionHow does the presence of calcium in the cytoplasm reg-ulate the cycling of cross-bridges? The answer requiresa closer look at the thin filament proteins, troponin andtropomyosin (Figure 9–13).
Tropomyosin is a rod-shaped molecule composedof two intertwined polypeptides with a length approx-imately equal to that of seven actin molecules. Chainsof tropomyosin molecules are arranged end to endalong the actin thin filament. These tropomyosin mole-cules partially cover the myosin-binding site on eachactin molecule, thereby preventing the cross-bridgesfrom making contact with actin. Each tropomyosinmolecule is held in this blocking position by troponin,a smaller, globular protein that is bound to bothtropomyosin and actin. One molecule of troponin bindsto each molecule of tropomyosin and regulates the ac-cess to myosin-binding sites on the seven actin mole-cules in contact with tropomyosin. This is the status ofa resting muscle fiber; troponin and tropomyosin coop-eratively block the interaction of cross-bridges with thethin filament.
What enables cross-bridges to bind to actin and be-gin cycling? For this to occur, tropomyosin molecules
must be moved away from their blocking positions onactin. This happens when calcium binds to specific bind-ing sites on troponin (not tropomyosin). The binding ofcalcium produces a change in the shape of troponin,which through troponin’s linkage to tropomyosin, dragstropomyosin away from the myosin-binding site oneach actin molecule. Conversely, removal of calciumfrom troponin reverses the process, turning off contrac-tile activity.
Thus, cytosolic calcium-ion concentration deter-mines the number of troponin sites occupied by cal-cium, which in turn determines the number of actinsites available for cross-bridge binding. Changes in cytosolic calcium concentration are controlled by elec-trical events in the muscle plasma membrane, whichwe now discuss.
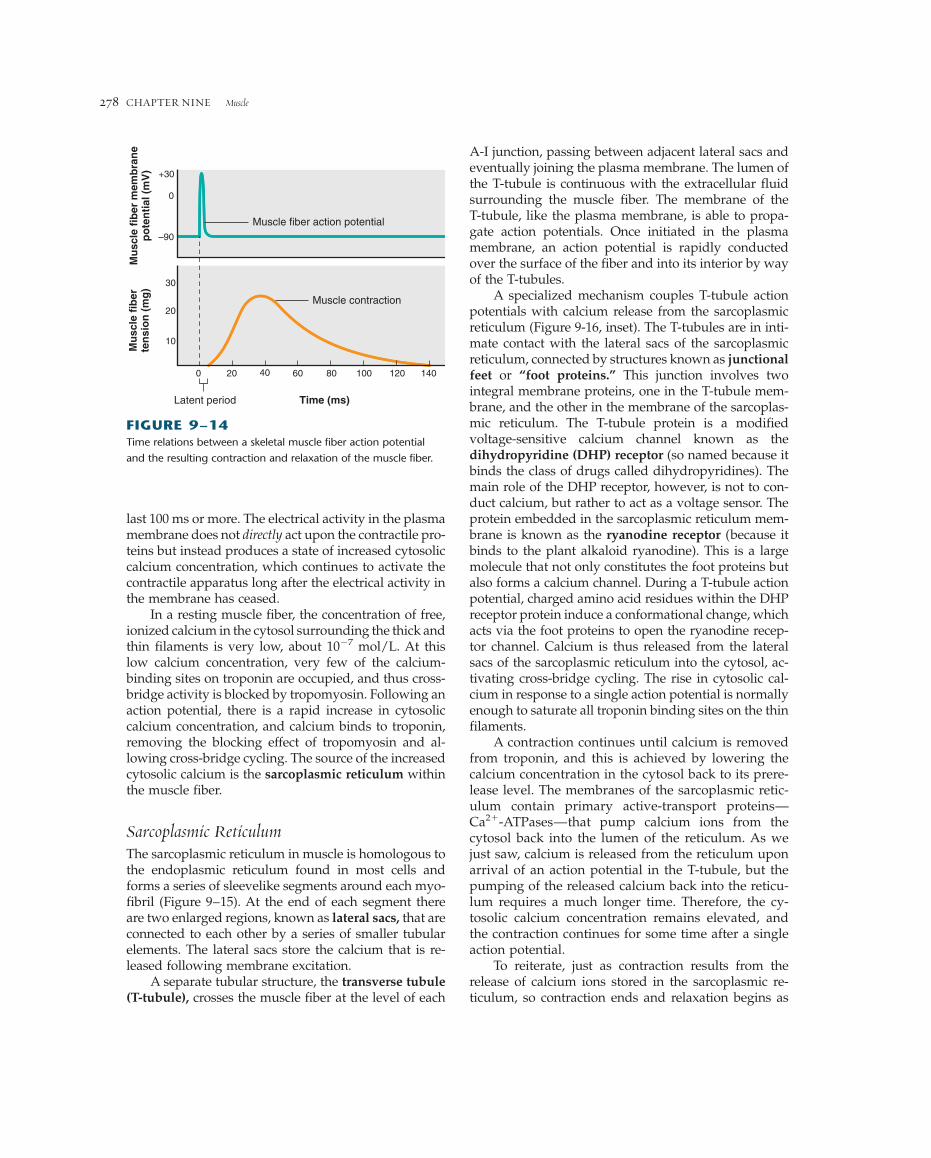
Excitation-Contraction CouplingExcitation-contraction coupling refers to the sequenceof events by which an action potential in the plasmamembrane of a muscle fiber leads to cross-bridge ac-tivity by the mechanisms just described. The skeletalmuscle plasma membrane is an excitable membrane ca-pable of generating and propagating action potentialsby mechanisms similar to those described for nerve cells(Chapter 6). An action potential in a skeletal muscle fiberlasts 1 to 2 ms and is completed before any signs of me-chanical activity begin (Figure 9–14). Once begun, themechanical activity following an action potential may
Tropomyosin
Troponin
ActinTropomyosin Troponin
(a)
(b)
Cross-bridgebinding sites
Ca2+ binding site
FIGURE 9–13(a) Molecule of troponin bound to a molecule of tropomyosin.
(b) Two chains of tropomyosin on a thin filament regulate access
of cross-bridges to binding sites on actin.
CHAPTER NINE Muscle278
last 100 ms or more. The electrical activity in the plasmamembrane does not directly act upon the contractile pro-teins but instead produces a state of increased cytosoliccalcium concentration, which continues to activate thecontractile apparatus long after the electrical activity inthe membrane has ceased.
In a resting muscle fiber, the concentration of free,ionized calcium in the cytosol surrounding the thick andthin filaments is very low, about 10�7 mol/L. At thislow calcium concentration, very few of the calcium-binding sites on troponin are occupied, and thus cross-bridge activity is blocked by tropomyosin. Following anaction potential, there is a rapid increase in cytosoliccalcium concentration, and calcium binds to troponin,removing the blocking effect of tropomyosin and al-lowing cross-bridge cycling. The source of the increasedcytosolic calcium is the sarcoplasmic reticulum withinthe muscle fiber.
Sarcoplasmic ReticulumThe sarcoplasmic reticulum in muscle is homologous tothe endoplasmic reticulum found in most cells andforms a series of sleevelike segments around each myo-fibril (Figure 9–15). At the end of each segment thereare two enlarged regions, known as lateral sacs, that areconnected to each other by a series of smaller tubularelements. The lateral sacs store the calcium that is re-leased following membrane excitation.
A separate tubular structure, the transverse tubule(T-tubule), crosses the muscle fiber at the level of each
A-I junction, passing between adjacent lateral sacs andeventually joining the plasma membrane. The lumen ofthe T-tubule is continuous with the extracellular fluidsurrounding the muscle fiber. The membrane of theT-tubule, like the plasma membrane, is able to propa-gate action potentials. Once initiated in the plasmamembrane, an action potential is rapidly conductedover the surface of the fiber and into its interior by wayof the T-tubules.
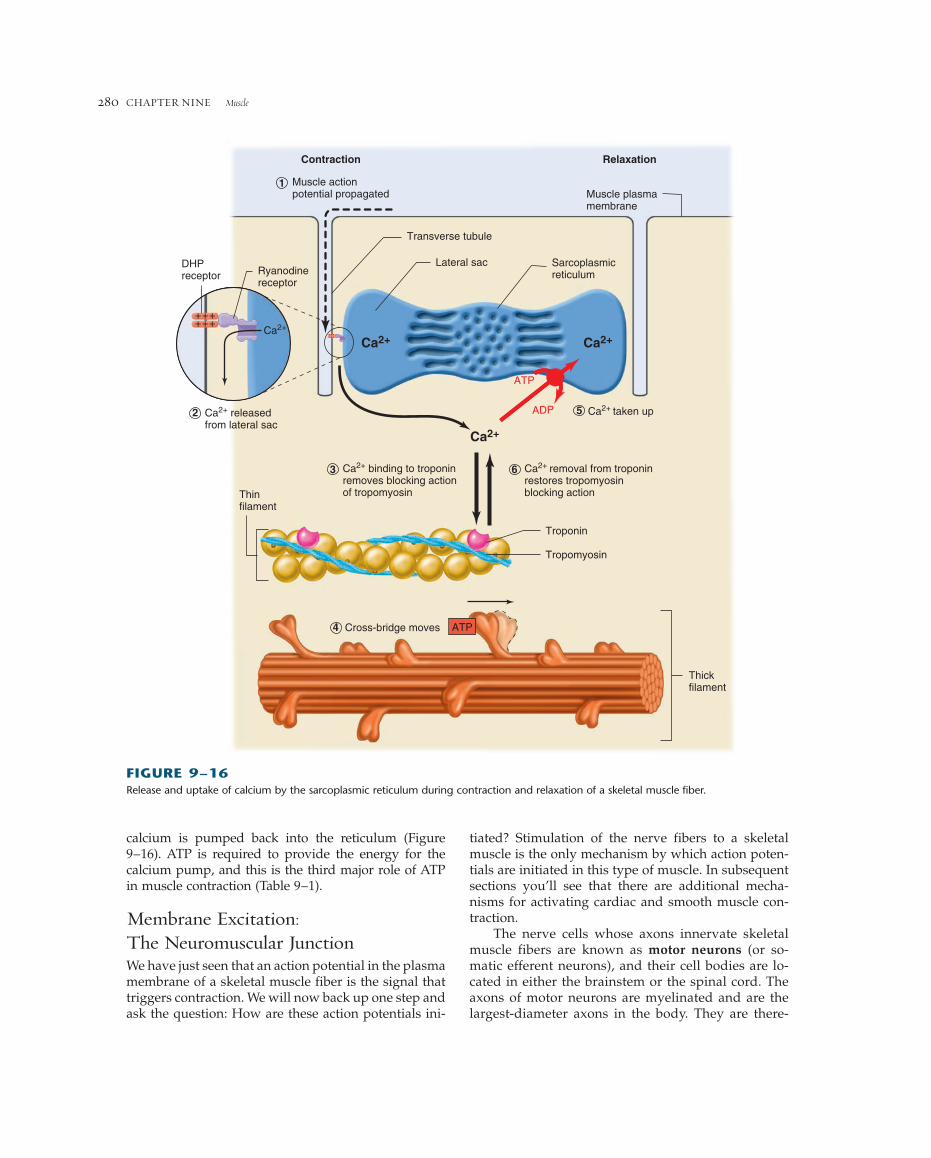
A specialized mechanism couples T-tubule actionpotentials with calcium release from the sarcoplasmicreticulum (Figure 9-16, inset). The T-tubules are in inti-mate contact with the lateral sacs of the sarcoplasmicreticulum, connected by structures known as junctionalfeet or “foot proteins.” This junction involves twointegral membrane proteins, one in the T-tubule mem-brane, and the other in the membrane of the sarcoplas-mic reticulum. The T-tubule protein is a modifiedvoltage-sensitive calcium channel known as the dihydropyridine (DHP) receptor (so named because itbinds the class of drugs called dihydropyridines). Themain role of the DHP receptor, however, is not to con-duct calcium, but rather to act as a voltage sensor. Theprotein embedded in the sarcoplasmic reticulum mem-brane is known as the ryanodine receptor (because itbinds to the plant alkaloid ryanodine). This is a largemolecule that not only constitutes the foot proteins butalso forms a calcium channel. During a T-tubule actionpotential, charged amino acid residues within the DHPreceptor protein induce a conformational change, whichacts via the foot proteins to open the ryanodine recep-tor channel. Calcium is thus released from the lateralsacs of the sarcoplasmic reticulum into the cytosol, ac-tivating cross-bridge cycling. The rise in cytosolic cal-cium in response to a single action potential is normallyenough to saturate all troponin binding sites on the thinfilaments.
A contraction continues until calcium is removedfrom troponin, and this is achieved by lowering thecalcium concentration in the cytosol back to its prere-lease level. The membranes of the sarcoplasmic retic-ulum contain primary active-transport proteins—Ca2�-ATPases—that pump calcium ions from the cytosol back into the lumen of the reticulum. As wejust saw, calcium is released from the reticulum uponarrival of an action potential in the T-tubule, but thepumping of the released calcium back into the reticu-lum requires a much longer time. Therefore, the cy-tosolic calcium concentration remains elevated, andthe contraction continues for some time after a singleaction potential.
To reiterate, just as contraction results from therelease of calcium ions stored in the sarcoplasmic re-ticulum, so contraction ends and relaxation begins as
Muscle fiber action potential
0 20 40 60 80 100 120 140
Latent period Time (ms)
+30
0
–90
30
20
10
Mu
scle
fib
erte
nsi
on
(m
g)
Mu
scle
fib
er m
emb
ran
ep
ote
nti
al (
mV
)
Muscle contraction
FIGURE 9–14Time relations between a skeletal muscle fiber action potential
and the resulting contraction and relaxation of the muscle fiber.

www.mhhe.com/widmaier9 279
FIGURE 9–15(a) Diagrammatic representation of the sarcoplasmic reticulum,
the transverse tubules, and the myofibrils. (b) Anatomical
structure of transverse tubules and sarcoplasmic reticulum in
a single skeletal muscle fiber.
Myofibrils
Cytosol
Sarcoplasmic reticulum
Plasmamembrane
Transverse tubules
Lateral sacs
Mitochondrion
(b)
(a)Opening of transversetubule to extracellular fluid Muscle-fiber plasma membrane
Transversetubules
Segments ofsarcoplasmic reticulum
Myofibrils
tiated? Stimulation of the nerve fibers to a skeletalmuscle is the only mechanism by which action poten-tials are initiated in this type of muscle. In subsequentsections you’ll see that there are additional mecha-nisms for activating cardiac and smooth muscle con-traction.
The nerve cells whose axons innervate skeletalmuscle fibers are known as motor neurons (or so-matic efferent neurons), and their cell bodies are lo-cated in either the brainstem or the spinal cord. Theaxons of motor neurons are myelinated and are thelargest-diameter axons in the body. They are there-
CHAPTER NINE Muscle280
calcium is pumped back into the reticulum (Figure 9–16). ATP is required to provide the energy for the calcium pump, and this is the third major role of ATPin muscle contraction (Table 9–1).
Membrane Excitation:The Neuromuscular JunctionWe have just seen that an action potential in the plasmamembrane of a skeletal muscle fiber is the signal thattriggers contraction. We will now back up one step andask the question: How are these action potentials ini-
Muscle actionpotential propagated
Contraction Relaxation
Muscle plasmamembrane
Transverse tubule
Lateral sac Sarcoplasmicreticulum
Ca2+ binding to troponinremoves blocking action of tropomyosin
Ca2+ removal from troponinrestores tropomyosinblocking action
Ca2+
Ca2+ taken up
Troponin
Tropomyosin
Cross-bridge moves
Thickfilament
Thinfilament
Ca2+ Ca2+
ATP
ADP
ATP
1
Ca2+ releasedfrom lateral sac
2 5
63
4
++++++
++++++
Ca2+
DHPreceptor Ryanodine
receptor
FIGURE 9–16Release and uptake of calcium by the sarcoplasmic reticulum during contraction and relaxation of a skeletal muscle fiber.
www.mhhe.com/widmaier9 281
fore able to propagate action potentials at high ve-locities, allowing signals from the central nervoussystem to be transmitted to skeletal muscle fiberswith minimal delay.
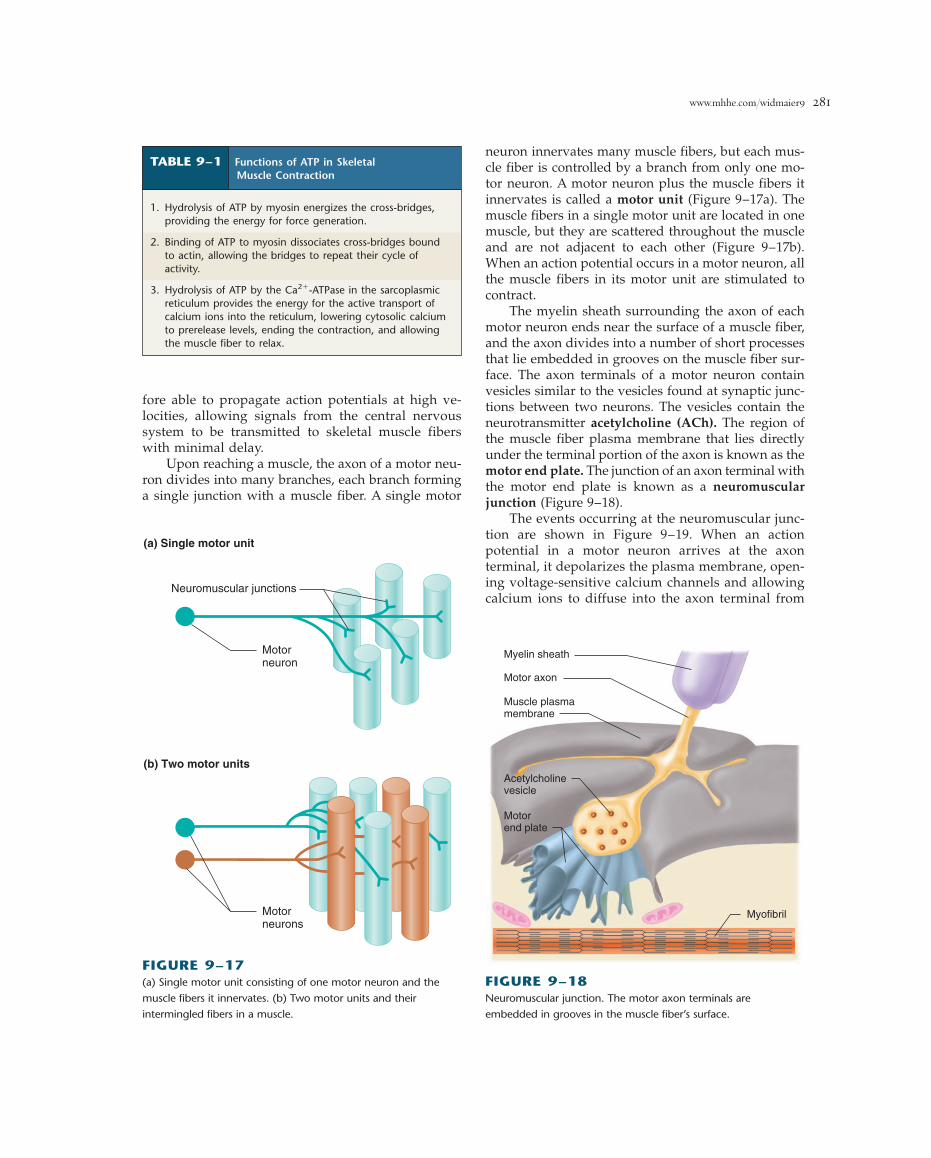
Upon reaching a muscle, the axon of a motor neu-ron divides into many branches, each branch forminga single junction with a muscle fiber. A single motor
neuron innervates many muscle fibers, but each mus-cle fiber is controlled by a branch from only one mo-tor neuron. A motor neuron plus the muscle fibers itinnervates is called a motor unit (Figure 9–17a). Themuscle fibers in a single motor unit are located in onemuscle, but they are scattered throughout the muscleand are not adjacent to each other (Figure 9–17b).When an action potential occurs in a motor neuron, allthe muscle fibers in its motor unit are stimulated tocontract.
The myelin sheath surrounding the axon of eachmotor neuron ends near the surface of a muscle fiber,and the axon divides into a number of short processesthat lie embedded in grooves on the muscle fiber sur-face. The axon terminals of a motor neuron containvesicles similar to the vesicles found at synaptic junc-tions between two neurons. The vesicles contain theneurotransmitter acetylcholine (ACh). The region ofthe muscle fiber plasma membrane that lies directlyunder the terminal portion of the axon is known as themotor end plate. The junction of an axon terminal withthe motor end plate is known as a neuromuscularjunction (Figure 9–18).
The events occurring at the neuromuscular junc-tion are shown in Figure 9–19. When an action potential in a motor neuron arrives at the axon terminal, it depolarizes the plasma membrane, open-ing voltage-sensitive calcium channels and allowingcalcium ions to diffuse into the axon terminal from
TABLE 9–1 Functions of ATP in SkeletalMuscle Contraction
1. Hydrolysis of ATP by myosin energizes the cross-bridges,providing the energy for force generation.
2. Binding of ATP to myosin dissociates cross-bridges boundto actin, allowing the bridges to repeat their cycle ofactivity.
3. Hydrolysis of ATP by the Ca2�-ATPase in the sarcoplasmicreticulum provides the energy for the active transport ofcalcium ions into the reticulum, lowering cytosolic calciumto prerelease levels, ending the contraction, and allowingthe muscle fiber to relax.
(a) Single motor unit
Neuromuscular junctions
Motorneuron
Motorneurons
(b) Two motor units
FIGURE 9–17(a) Single motor unit consisting of one motor neuron and the
muscle fibers it innervates. (b) Two motor units and their
intermingled fibers in a muscle.
Myelin sheath
Motor axon
Muscle plasmamembrane
Acetylcholinevesicle
Motorend plate
Myofibril
FIGURE 9–18Neuromuscular junction. The motor axon terminals are
embedded in grooves in the muscle fiber’s surface.
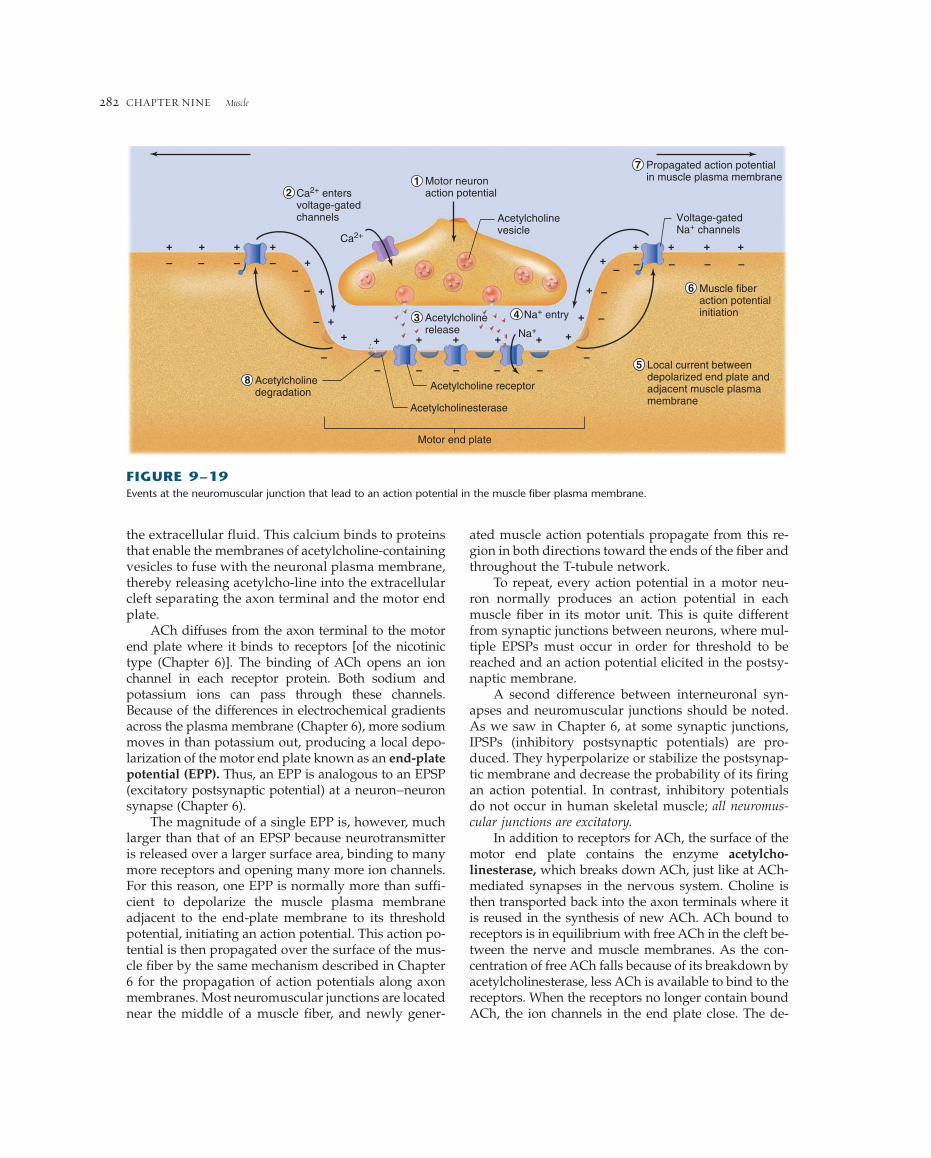
ated muscle action potentials propagate from this re-gion in both directions toward the ends of the fiber andthroughout the T-tubule network.
To repeat, every action potential in a motor neu-ron normally produces an action potential in eachmuscle fiber in its motor unit. This is quite differentfrom synaptic junctions between neurons, where mul-tiple EPSPs must occur in order for threshold to bereached and an action potential elicited in the postsy-naptic membrane.
A second difference between interneuronal syn-apses and neuromuscular junctions should be noted.As we saw in Chapter 6, at some synaptic junctions,IPSPs (inhibitory postsynaptic potentials) are pro-duced. They hyperpolarize or stabilize the postsynap-tic membrane and decrease the probability of its firingan action potential. In contrast, inhibitory potentialsdo not occur in human skeletal muscle; all neuromus-cular junctions are excitatory.
In addition to receptors for ACh, the surface of themotor end plate contains the enzyme acetylcho-linesterase, which breaks down ACh, just like at ACh-mediated synapses in the nervous system. Choline isthen transported back into the axon terminals where itis reused in the synthesis of new ACh. ACh bound toreceptors is in equilibrium with free ACh in the cleft be-tween the nerve and muscle membranes. As the con-centration of free ACh falls because of its breakdown byacetylcholinesterase, less ACh is available to bind to thereceptors. When the receptors no longer contain boundACh, the ion channels in the end plate close. The de-
CHAPTER NINE Muscle282
the extracellular fluid. This calcium binds to proteinsthat enable the membranes of acetylcholine-containingvesicles to fuse with the neuronal plasma membrane,thereby releasing acetylcho-line into the extracellularcleft separating the axon terminal and the motor endplate.
ACh diffuses from the axon terminal to the motorend plate where it binds to receptors [of the nicotinictype (Chapter 6)]. The binding of ACh opens an ionchannel in each receptor protein. Both sodium andpotassium ions can pass through these channels.Because of the differences in electrochemical gradientsacross the plasma membrane (Chapter 6), more sodiummoves in than potassium out, producing a local depo-larization of the motor end plate known as an end-platepotential (EPP). Thus, an EPP is analogous to an EPSP(excitatory postsynaptic potential) at a neuron–neuronsynapse (Chapter 6).
The magnitude of a single EPP is, however, muchlarger than that of an EPSP because neurotransmitteris released over a larger surface area, binding to manymore receptors and opening many more ion channels.For this reason, one EPP is normally more than suffi-cient to depolarize the muscle plasma membraneadjacent to the end-plate membrane to its thresholdpotential, initiating an action potential. This action po-tential is then propagated over the surface of the mus-cle fiber by the same mechanism described in Chapter6 for the propagation of action potentials along axonmembranes. Most neuromuscular junctions are locatednear the middle of a muscle fiber, and newly gener-
+ ++ +
+
+
+
+ + + +
+
+
++ + + +
+++
Acetylcholinerelease
Motor neuronaction potential
Muscle fiberaction potentialinitiation
Local current betweendepolarized end plate andadjacent muscle plasmamembrane
Acetylcholine receptorAcetylcholinedegradation
Acetylcholinesterase
Motor end plate
Acetylcholinevesicle
Propagated action potentialin muscle plasma membrane
Voltage-gatedNa+ channels
– – – ––
–
–– – –
–
–
–
– – – – –
– –
–
12
8
3
7
6
4
5
Na+ entry
Na+
Ca2+ entersvoltage-gatedchannels
Ca2+
FIGURE 9–19Events at the neuromuscular junction that lead to an action potential in the muscle fiber plasma membrane.

www.mhhe.com/widmaier9 283
polarized end plate returns to its resting potential andcan respond to the subsequent arrival of ACh releasedby another neuron action potential.
Table 9–2 summarizes the sequence of events thatlead from an action potential in a motor neuron to thecontraction and relaxation of a skeletal muscle fiber.
Disruption of Neuromuscular Signaling
There are many ways by which events at the neuro-muscular junction can be modified by disease or drugs.For example, the deadly South American arrowheadpoison curare binds strongly to nicotinic ACh recep-tors, but it does not open their ion channels and is notdestroyed by acetylcholinesterase. When a receptor isoccupied by curare, ACh cannot bind to the receptor.Therefore, although the motor nerves still conduct nor-mal action potentials and release ACh, there is no re-sulting EPP in the motor end plate and no contraction.
Since the skeletal muscles responsible for breathing,like all skeletal muscles, depend upon neuromusculartransmission to initiate their contraction, curare poi-soning can lead to death by asphyxiation. Drugs sim-ilar to curare are used in small amounts to preventmuscular contractions during certain types of surgicalprocedures when it is necessary to immobilize the sur-gical field (gallamine is one example). The use of suchparalytic agents also reduces the required dose of gen-eral anesthetic, allowing patients to recover faster withfewer complications. Patients are artificially ventilatedin order to maintain respiration until the drug has beenremoved from the system.
Neuromuscular transmission can also be blockedby inhibiting acetylcholinesterase. Some organophos-phates, which are the main ingredients in certain pes-ticides and “nerve gases” (the latter developed forbiological warfare), inhibit this enzyme. In the pres-ence of such agents, ACh is released normally upon
TABLE 9–2 Sequence of Events Between a Motor Neuron Action Potential and Skeletal Muscle Fiber Contraction
1. Action potential is initiated and propagates to motor neuron axon terminals.
2. Calcium enters axon terminals through voltage-gated calcium channels.
3. Calcium entry triggers release of ACh from axon terminals.
4. ACh diffuses from axon terminals to motor end plate in muscle fiber.
5. ACh binds to nicotinic receptors on motor end plate, increasing their permeability to Na� and K�.
6. More Na� moves into the fiber at the motor end plate than K� moves out, depolarizing the membrane, producing the end platepotential (EPP).
7. Local currents depolarize the adjacent muscle cell plasma membrane to its threshold potential, generating an action potential thatpropagates over the muscle fiber surface and into the fiber along the T-tubules.
8. Action potential in T-tubules triggers release of Ca2� from lateral sacs of sarcoplasmic reticulum.
9. Ca2� binds to troponin on the thin filaments, causing tropomyosin to move away from its blocking position, thereby uncoveringcross-bridge binding sites on actin.
10. Energized myosin cross-bridges on the thick filaments bind to actin:
A � M � ADP � Pi 88n A � M � ADP � Pi
11. Cross-bridge binding triggers release of ATP hydrolysis products from myosin, producing an angular movement of each cross-bridge:
A � M � ADP � Pi 88n A � M � ADP � Pi
12. ATP binds to myosin, breaking linkage between actin and myosin and thereby allowing cross-bridges to dissociate from actin:
A � M � ATP 88n A � M � ATP
13. ATP bound to myosin is split, energizing the myosin cross-bridge:
M � ATP 88n M � ADP � Pi
14. Cross-bridges repeat steps 10 to 13, producing movement (sliding) of thin filaments past thick filaments. Cycles of cross-bridgemovement continue as long as Ca2� remains bound to troponin.
15. Cytosolic Ca2� concentration decreases as Ca2� is actively transported into sarcoplasmic reticulum by Ca2�-ATPase.
16. Removal of Ca2� from troponin restores blocking action of tropomyosin, the cross-bridge cycle ceases, and the muscle fiberrelaxes.

the muscle remains constant, is said to be isotonic (con-stant tension). Shortening contractions are also referredto as concentric contractions.
A third type of contraction is a lengthening con-traction (eccentric contraction). This occurs when anunsupported load on a muscle is greater than the ten-sion being generated by the cross-bridges. In this sit-uation, the load pulls the muscle to a longer length inspite of the opposing force being produced by thecross-bridges. Such lengthening contractions occurwhen an object being supported by muscle contractionis lowered, like when the knee extensors in your thighsare used to lower you to a seat from a standing posi-tion. It must be emphasized that in these situations thelengthening of muscle fibers is not an active processproduced by the contractile proteins, but a conse-quence of the external forces being applied to the mus-cle. In the absence of external lengthening forces, afiber will only shorten when stimulated; it will neverlengthen. All three types of contractions—isometric,isotonic, and lengthening—occur in the natural courseof everyday activities.
During each type of contraction the cross-bridgesrepeatedly go through the four steps of the cross-bridge cycle illustrated in Figure 9–12. During step 2of an isotonic contraction, the cross-bridges bound toactin move to their angled positions, causing short-ening of the sarcomeres. In contrast, during an iso-metric contraction, the bound cross-bridges are unableto move the thin filaments because of the load on themuscle fiber, but they do exert a force on the thin fil-aments—isometric tension. If isometric contraction isprolonged, cycling cross-bridges repeatedly rebind tothe same actin molecule. During a lengthening con-traction, the cross-bridges in step 2 are pulled back-ward toward the Z lines by the load while still boundto actin and exerting force. The events of steps 1, 3,and 4 are the same in all three types of contractions.Thus, the chemical changes in the contractile proteinsduring each type of contraction are the same. The endresult (shortening, no length change, or lengthening)is determined by the magnitude of the load on themuscle.
Contraction terminology applies to both singlefibers and whole muscles. In this section, we describethe mechanics of single fiber contractions. Later we willdiscuss the factors controlling the mechanics of whole-muscle contraction.
Twitch ContractionsThe mechanical response of a single muscle fiber to asingle action potential is known as a twitch. Figure9–20a shows the main features of an isometric twitch.Following the action potential, there is an interval of a
CHAPTER NINE Muscle284
the arrival of an action potential at the axon terminaland binds to the end-plate receptors. The ACh is notdestroyed, however, because the acetylcholinesteraseis inhibited. The ion channels in the end plate thereforeremain open, producing a maintained depolarizationof the end plate and the muscle plasma membrane adjacent to the end plate. A skeletal muscle membranemaintained in a depolarized state cannot generate action potentials because the voltage-gated sodiumchannels in the membrane have entered an inactivestate, which requires repolarization to remove. Thus,the muscle does not contract in response to subsequentnerve stimulation, and the result is skeletal muscleparalysis and death from asphyxiation. Note that nervegases also cause ACh to build up at muscarinicsynapses, for example where parasympathetic neuronsinhibit cardiac pacemaker cells (Chapter 12). Thus, theantidote for nerve gas exposure must include the mus-carinic receptor antagonist atropine.
A third group of substances, including the toxinproduced by the bacterium Clostridium botulinum,blocks the release of acetylcholine from nerve termi-nals. Botulinum toxin is an enzyme that breaks downa protein required for the binding and fusion of AChvesicles with the plasma membrane of the axon ter-minal. This toxin, which produces the food poisoningcalled botulism, is one of the most potent poisonsknown because of the very small amount necessary toproduce an effect. Local application of botulinumtoxin is increasingly being used for clinical and cos-metic procedures, including the inhibition of overac-tive extraocular muscles, prevention of excessivesweat gland activity, and reduction of aging-relatedskin wrinkles.
MECHANICS OF SINGLE-FIBERCONTRACTION
The force exerted on an object by a contracting mus-cle is known as muscle tension, and the force exertedon the muscle by an object (usually its weight) is theload. Muscle tension and load are opposing forces.Whether or not a fiber shortens depends on the rela-tive magnitudes of the tension and the load. In orderfor muscle fibers to shorten, and thereby move a load,muscle tension must be greater than the opposingload.
When a muscle develops tension but does notshorten (or lengthen), the contraction is said to be iso-metric (constant length). Such contractions occur whenthe muscle supports a load in a constant position or at-tempts to move an otherwise supported load that isgreater than the tension developed by the muscle. Acon-traction in which the muscle shortens, while the load on

www.mhhe.com/widmaier9 285
few milliseconds, known as the latent period, beforethe tension in the muscle fiber begins to increase.During this latent period, the processes associated withexcitation-contraction coupling are occurring. The timeinterval from the beginning of tension development atthe end of the latent period to the peak tension is thecontraction time. Not all skeletal muscle fibers havethe same twitch contraction time. Some fast fibers havecontraction times as short as 10 ms, whereas slowerfibers may take 100 ms or longer. The duration of thecontraction time depends in part on the time that cy-tosolic calcium remains elevated so that cross-bridgescan continue to cycle. It is closely related to the Ca2�-ATPase activity in the sarcoplasmic reticulum; activity
0 20 40 60 80 100 120 140
Time (ms)Single actionpotential
Ten
sio
n (
mg
)
30
20
10
Latent period
Stimulator
Contraction time
(a) Isometric contraction
Motor neuron4
3
2
1
0mm
Muscle fiber
Force transducer
Dis
tan
ce s
ho
rten
ed (
mm
)
3
2
1
0 20 40 60 80 100 120 140
Time (ms)Single actionpotential
(b) Isotonic contraction
4
3
2
1
0mm Load
Shorteningdistance
FIGURE 9–20(a) Measurement of tension during a single isometric twitch of a skeletal muscle fiber. (b) Measurement of shortening during a single
isotonic twitch of a skeletal muscle fiber.
is greater in fast-twitch fibers and less in slow-twitchfibers.
Comparing isotonic and isometric twitches in thesame muscle fiber, one can see from Figure 9–20b thatthe latent period in an isotonic twitch is longer than thatin an isometric contraction, while the duration of themechanical event—shortening—is briefer in an isotonictwitch than the duration of force generation in an iso-metric twitch.
Moreover, the characteristics of an isotonic twitchdepend upon the magnitude of the load being lifted(Figure 9–21): At heavier loads: (1) the latent periodis longer, (2) the velocity of shortening (distanceshortened per unit of time) is slower, (3) the duration

of the twitch is shorter, and (4) the distance shortenedis less.
A closer look at the sequence of events in an iso-metric twitch explains this load-dependent behavior.Following excitation, the cross-bridges begin to developforce, but shortening does not begin until the muscletension just exceeds the load on the fiber. Thus, beforeshortening, there is a period of isometric contraction dur-ing which the tension increases. The heavier the load,the longer it takes for the tension to increase to the valueof the load, when shortening will begin. If the load ona fiber is increased, eventually a load is reached that themuscle is unable to lift, the velocity and distance ofshortening will be zero, and the contraction will becomecompletely isometric.
Load-Velocity RelationIt is a common experience that light objects can bemoved faster than heavy objects. That is, the velocityat which a muscle fiber shortens decreases with in-creasing loads (Figure 9–22). The shortening velocityis maximal when there is no load and is zero whenthe load is equal to the maximal isometric tension. Atloads greater than the maximal isometric tension, thefiber will lengthen at a velocity that increases withload.
The shortening velocity is determined by the rate atwhich individual cross-bridges undergo their cyclicalactivity. Because one ATP is split during each cross-bridge cycle, the rate of ATP splitting determines theshortening velocity. Increasing the load on a cross-bridge slows its forward movement during the powerstroke. This reduces the overall rate of ATP hydrolysis,and thus the velocity of shortening.
CHAPTER NINE Muscle286
Maximum shortening velocity(zero load)
Maximum isometric tension(zero velocity)
Load
Isotonicshortening
Lengtheningcontraction
Sh
ort
enin
gve
loci
tyL
eng
then
ing
velo
city
0
FIGURE 9–22Velocity of skeletal muscle fiber shortening and lengthening as a
function of load. Note that the force on the cross-bridges during
a lengthening contraction is greater than the maximum isometric
tension.
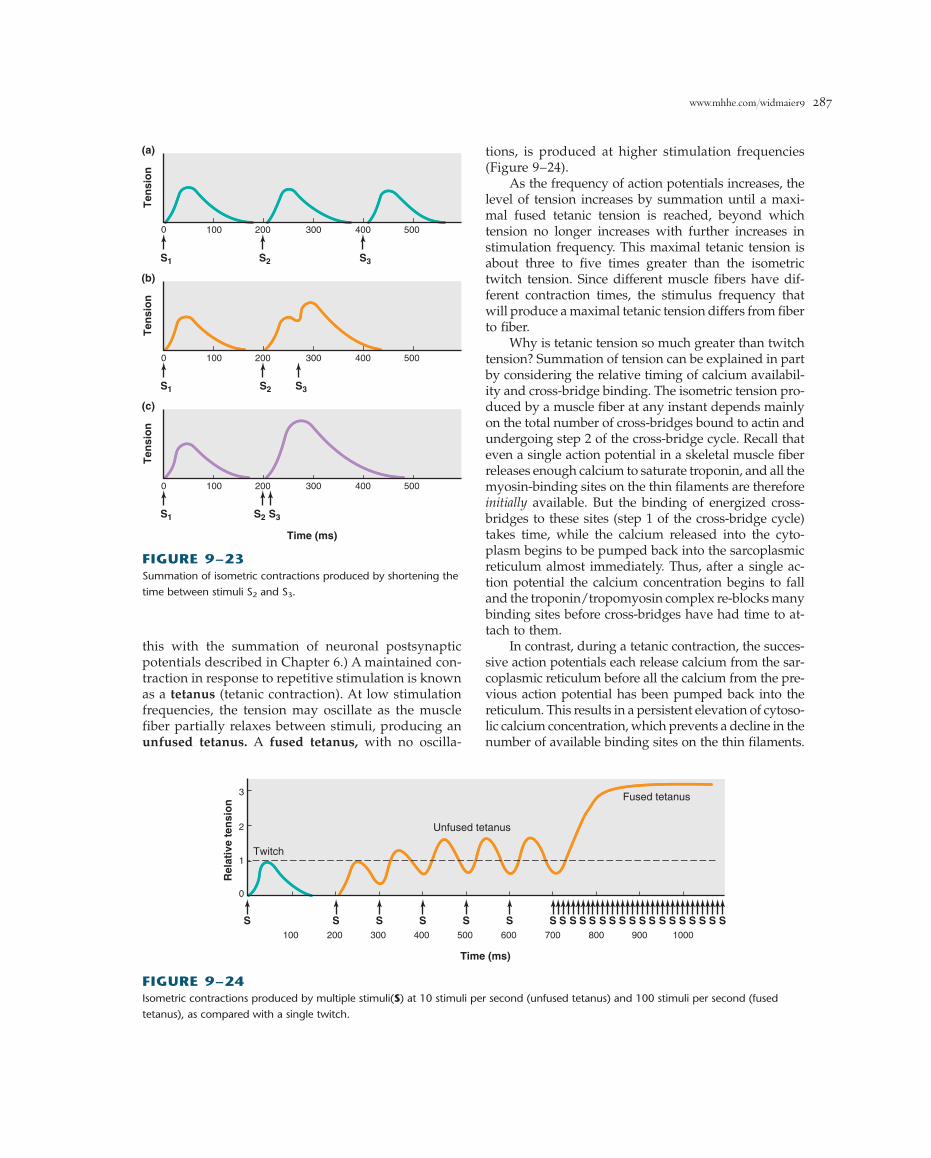
Frequency-Tension RelationSince a single action potential in a skeletal musclefiber lasts 1 to 2 ms but the twitch may last for 100ms, it is possible for a second action potential to beinitiated during the period of mechanical activity.Figure 9–23 illustrates the tension generated duringisometric contractions of a muscle fiber in responseto three successive stimuli. In Figure 9–23a, the iso-metric twitch following the first stimulus S1 lasts 150ms. The second stimulus S2, applied to the musclefiber 200 ms after S1 when the fiber has completelyrelaxed, causes a second identical twitch, and a thirdstimulus S3, equally timed, produces a third identicaltwitch. In Figure 9–23b, the interval between S1 andS2 remains 200 ms, but a third stimulus is applied 60ms after S2, when the mechanical response resultingfrom S2 is beginning to decrease but has not yetended. Stimulus S3 induces a contractile responsewhose peak tension is greater than that produced byS2. In Figure 9–23c, the interval between S2 and S3 isfurther reduced to 10 ms, and the resulting peak ten-sion is even greater. Indeed, the mechanical responseto S3 is a smooth continuation of the mechanical re-sponse already induced by S2.
The increase in muscle tension from successive ac-tion potentials occurring during the phase of mechan-ical activity is known as summation. (Do not confuse
Light load
Intermediate load
Heavy load
Time (ms)Single actionpotential
Dis
tan
ce s
ho
rten
ed (
mm
) 4
3
2
1
0 20 40 60 80 100 120 140
FIGURE 9–21Isotonic twitches with different loads. The distance shortened,
velocity of shortening, and duration of shortening all decrease
with increased load, whereas the time from stimulation to the
beginning of shortening increases with increasing load.
www.mhhe.com/widmaier9 287
this with the summation of neuronal postsynapticpotentials described in Chapter 6.) A maintained con-traction in response to repetitive stimulation is knownas a tetanus (tetanic contraction). At low stimulationfrequencies, the tension may oscillate as the musclefiber partially relaxes between stimuli, producing anunfused tetanus. A fused tetanus, with no oscilla-
tions, is produced at higher stimulation frequencies(Figure 9–24).
As the frequency of action potentials increases, thelevel of tension increases by summation until a maxi-mal fused tetanic tension is reached, beyond whichtension no longer increases with further increases instimulation frequency. This maximal tetanic tension isabout three to five times greater than the isometrictwitch tension. Since different muscle fibers have dif-ferent contraction times, the stimulus frequency thatwill produce a maximal tetanic tension differs from fiberto fiber.
Why is tetanic tension so much greater than twitchtension? Summation of tension can be explained in partby considering the relative timing of calcium availabil-ity and cross-bridge binding. The isometric tension pro-duced by a muscle fiber at any instant depends mainlyon the total number of cross-bridges bound to actin andundergoing step 2 of the cross-bridge cycle. Recall thateven a single action potential in a skeletal muscle fiberreleases enough calcium to saturate troponin, and all themyosin-binding sites on the thin filaments are thereforeinitially available. But the binding of energized cross-bridges to these sites (step 1 of the cross-bridge cycle)takes time, while the calcium released into the cyto-plasm begins to be pumped back into the sarcoplasmicreticulum almost immediately. Thus, after a single ac-tion potential the calcium concentration begins to falland the troponin/tropomyosin complex re-blocks manybinding sites before cross-bridges have had time to at-tach to them.
In contrast, during a tetanic contraction, the succes-sive action potentials each release calcium from the sar-coplasmic reticulum before all the calcium from the pre-vious action potential has been pumped back into thereticulum. This results in a persistent elevation of cytoso-lic calcium concentration, which prevents a decline in thenumber of available binding sites on the thin filaments.
Ten
sio
n
(b)
100 200 300 400 5000
S1 S2 S3
Ten
sio
n
(c)
100 200 300 400 5000
S1 S2 S3
Time (ms)
Ten
sio
n(a)
100 200 300 400 5000
S1 S3S2
FIGURE 9–23Summation of isometric contractions produced by shortening the
time between stimuli S2 and S3.
FIGURE 9–24Isometric contractions produced by multiple stimuli(S) at 10 stimuli per second (unfused tetanus) and 100 stimuli per second (fused
tetanus), as compared with a single twitch.
0
1
2
3
Rel
ativ
e te
nsi
on
S S S S S S100 200 300 400 500 600
S S S S S S S S S S S S S S S S700 800 900 1000
S S
Time (ms)
Twitch
Fused tetanus
Unfused tetanus

The length at which the fiber develops the greatest iso-metric active tension is termed the optimal length, lo.
When a muscle fiber length is 60 percent of lo, thefiber develops no tension when stimulated. As length isincreased from this point, the isometric tension at eachlength is increased up to a maximum at lo. Furtherlengthening leads to a drop in tension. At lengths of 175percent lo or beyond, the fiber develops no tension whenstimulated.
When all the skeletal muscles in the body are re-laxed, the lengths of most fibers are near lo and thusnear the optimal lengths for force generation. The lengthof a relaxed fiber can be altered by the load on the mus-cle or the contraction of other muscles that stretch therelaxed fibers, but the extent to which the relaxed lengthcan be changed is limited by the muscle’s attachmentsto bones. It rarely exceeds a 30 percent change from loand is often much less. Over this range of lengths, theability to develop tension never falls below about halfof the tension that can be developed at lo (Figure 9–25).
The relationship between fiber length and the fiber’s capacity to develop active tension during contractioncan be partially explained in terms of the sliding-filament mechanism. Stretching a relaxed muscle fiberpulls the thin filaments past the thick filaments, chang-ing the amount of overlap between them. Stretching afiber to 1.75 lo pulls the filaments apart to the pointwhere there is no overlap. At this point there can be no cross-bridge binding to actin and no development of tension. Between 1.75 lo and lo, more and more filaments overlap, and the tension developed uponstimulation increases in proportion to the increasednumber of cross-bridges in the overlap region. Filamentoverlap is greatest at lo, allowing the maximal numberof cross-bridges to bind to the thin filaments, therebyproducing maximal tension.
The tension decline at lengths less than lo is the re-sult of several factors. For example, (1) the overlappingsets of thin filaments from opposite ends of the sar-comere may interfere with the cross-bridges’ ability tobind and exert force, and (2) at very short lengths the Zlines collide with the ends of the relatively rigid thickfilaments, creating an internal resistance to sarcomereshortening.
SKELETAL MUSCLE ENERGYMETABOLISM
As we have seen, ATP performs three functions directlyrelated to muscle fiber contraction and relaxation (seeTable 9–1). In no other cell type does the rate of ATPbreakdown increase so much from one moment to thenext as in a skeletal muscle fiber (20 to several hun-
CHAPTER NINE Muscle288
number of binding sites remains available, and manymore cross-bridges are bound to the thin filaments at anyinstant.
Other causes of the lower tension seen in a singletwitch are elastic structures, such as muscle tendons andthe protein titin, which delay the transmission of cross-bridge force to the ends of a fiber. Because a single twitchis so brief, cross-bridge activity is already declining be-fore force has been fully transmitted through thesestructures. This is less of a factor during tetanic stimu-lation because of the much longer duration of cross-bridge activity and force generation.
Length-Tension RelationThe springlike characteristics of the protein titin, whichis attached to the Z line at one end and the thick fila-ments at the other, is responsible for most of the passiveelastic properties of relaxed muscles. With increasedstretch, the passive tension in a relaxed fiber increases,not from active cross-bridge movements but from elon-gation of the titin filaments. If the stretched fiber is re-leased, its length will return to an equilibrium length,much like releasing a stretched rubber band. The criti-cal point for this section is that the amount of active ten-sion developed by a muscle fiber during contraction canalso be altered by changing the length of the fiber. If youstretch a muscle fiber to various lengths and tetanicallystimulate it at each length, the magnitude of the activetension will vary with length as shown in Figure 9–25.
40 60 80 100 120 140 160Percent of muscle length
100
80
60
40
20
0
Per
cen
t o
f m
axim
um
iso
met
ric
teta
nic
ten
sio
n
lo
FIGURE 9–25Variation in active isometric tetanic tension with muscle fiber
length. The blue band represents the range of length changes
that can normally occur in the body.

www.mhhe.com/widmaier9 289
dredfold depending on the type of muscle fiber) whenit goes from rest to a state of contractile activity. The smallsupply of preformed ATP that exists at the start of con-tractile activity would only support a few twitches. If afiber is to sustain contractile activity, molecules of ATPmust be produced by metabolism as rapidly as they arebroken down during the contractile process.
There are three ways a muscle fiber can form ATP(Figure 9–26): (1) phosphorylation of ADP by creatinephosphate, (2) oxidative phosphorylation of ADP in themitochondria, and (3) phosphorylation of ADP by theglycolytic pathway in the cytosol.
Phosphorylation of ADP by creatine phosphate (CP)provides a very rapid means of forming ATP at the on-set of contractile activity. When the chemical bond be-tween creatine (C) and phosphate is broken, the amountof energy released is about the same as that releasedwhen the terminal phosphate bond in ATP is broken.This energy, along with the phosphate group, can betransferred to ADP to form ATP in a reversible reactioncatalyzed by creatine kinase:
Creatinekinase
CP � ADP 34 C � ATP
Although creatine phosphate is a high-energy molecule,its energy cannot be released by myosin to drive cross-bridge activity. During periods of rest, muscle fibersbuild up a concentration of creatine phosphate approx-
imately five times that of ATP. At the beginning of con-traction, when the concentration of ATP begins to falland that of ADP to rise owing to the increased rate ofATP breakdown by myosin, mass action favors the for-mation of ATP from creatine phosphate. This transfer ofenergy is so rapid that the concentration of ATP in amuscle fiber changes very little at the start of contrac-tion, whereas the concentration of creatine phosphatefalls rapidly.
Although the formation of ATP from creatine phos-phate is very rapid, requiring only a single enzymaticreaction, the amount of ATP that can be formed by thisprocess is limited by the initial concentration of creatinephosphate in the cell. If contractile activity is to be con-tinued for more than a few seconds, the muscle mustbe able to form ATP from the other two sources listedpreviously. The use of creatine phosphate at the start ofcontractile activity provides the few seconds necessaryfor the slower, multienzyme pathways of oxidativephosphorylation and glycolysis to increase their rates ofATP formation to levels that match the rates of ATPbreakdown.
At moderate levels of muscular activity, most of theATP used for muscle contraction is formed by oxidativephosphorylation, and during the first 5 to 10 min of suchexercise, breakdown of muscle glycogen to glucoseprovides the major fuel contributing to oxidative phos-phorylation. For the next 30 min or so, blood-bornefuels become dominant, blood glucose and fatty acids
ATP
OxidativephosphorylationGlycolysis
Lactic acid
Glycogen
Creatine phosphate
Creatine
ADP + Pi
Fatty acids
Amino acids Proteins
Ca2+-ATPase
Myosin ATPase contraction
relaxation
Muscle fiber
Glucose
Oxygen
Fatty acids
Blood
(1)
(3) (2)
FIGURE 9–26The three sources of ATP production during muscle contraction: (1) creatine phosphate, (2) oxidative phosphorylation, and
(3) glycolysis.

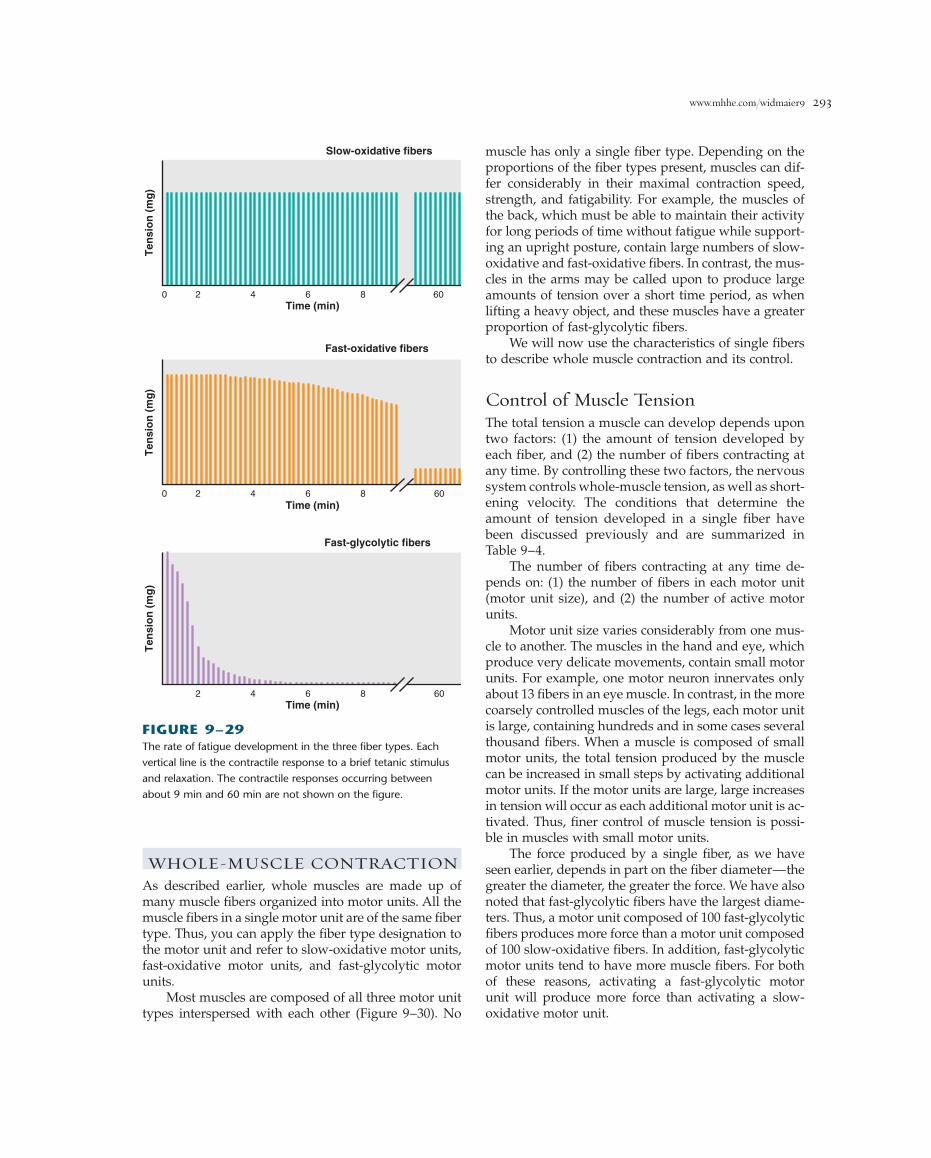
Muscle FatigueWhen a skeletal muscle fiber is repeatedly stimulated,the tension developed by the fiber eventually decreaseseven though the stimulation continues (Figure 9–27).This decline in muscle tension as a result of previouscontractile activity is known as muscle fatigue.Additional characteristics of fatigued muscle are a de-creased shortening velocity and a slower rate of relax-ation. The onset of fatigue and its rate of developmentdepend on the type of skeletal muscle fiber that is ac-tive, the intensity and duration of contractile activity,and the degree of an individual’s fitness.
If a muscle is allowed to rest after the onset of fa-tigue, it can recover its ability to contract upon restim-ulation (Figure 9–27). The rate of recovery dependsupon the duration and intensity of the previous activ-ity. Some muscle fibers fatigue rapidly if continuouslystimulated but also recover rapidly after a brief rest. This is the type of fatigue (high-frequency fatigue) that accompanies high-intensity, short-duration exercise,such as weight lifting. In contrast, low-frequency fatiguedevelops more slowly with low-intensity, long-durationexercise, such as long-distance running, during whichthere are cyclical periods of contraction and relaxation.This type of fatigue requires much longer periods of rest,often up to 24 h, before the muscle achieves completerecovery.
It might seem logical that depletion of energy inthe form of ATP would account for fatigue, but the ATPconcentration in fatigued muscle is found to be onlyslightly lower than in a resting muscle, and not lowenough to impair cross-bridge cycling. If contractile activity were to continue without fatigue, the ATP con-centration could decrease to the point that the cross-bridges would become linked in a rigor configuration,
CHAPTER NINE Muscle290
contributing approximately equally; beyond this period,fatty acids become progressively more important, andglucose utilization by muscle decreases.
If the intensity of exercise exceeds about 70 percentof the maximal rate of ATP breakdown, however, gly-colysis contributes an increasingly significant fraction ofthe total ATP generated by the muscle. The glycolyticpathway, although producing only small quantities ofATP from each molecule of glucose metabolized, canproduce large quantities of ATP when enough enzymesand substrate are available, and it can do so in the ab-sence of oxygen (anaerobic). The glucose for glycolysiscan be obtained from two sources: the blood or the storesof glycogen within the contracting muscle fibers. As theintensity of muscle activity increases, a greater fractionof the total ATP production is formed by anaerobic gly-colysis. This is associated with a corresponding increasein the production of lactic acid.
At the end of muscle activity, creatine phosphateand glycogen levels in the muscle have decreased. Toreturn a muscle fiber to its original state, therefore, theseenergy-storing compounds must be replaced. Bothprocesses require energy, and so a muscle continues toconsume increased amounts of oxygen for some timeafter it has ceased to contract. In addition, extra oxygenis required to metabolize accumulated lactic acid andreturn the blood and interstitial fluid oxygen concen-trations to pre-exercise values. These processes are evi-denced by the fact that you continue to breathe deeplyand rapidly for a period of time immediately followingintense exercise. This elevated consumption of oxygenfollowing exercise repays what has been called the oxy-gen debt—that is, the increased production of ATP byoxidative phosphorylation following exercise that isused to restore the energy reserves in the form ofcreatine phosphate and glycogen.
Stimuli
Iso
met
ric
ten
sio
n
Tetanus
Fatigue Fatigue
Time
Restperiod
FIGURE 9–27Muscle fatigue during a maintained isometric tetanus and recovery following a period of rest.

www.mhhe.com/widmaier9 291
which is very damaging to muscle fibers. Thus, mus-cle fatigue may have evolved as a mechanism for pre-venting the onset of rigor.
Many factors can contribute to the fatigue of skele-tal muscle. Fatigue from high-intensity, short durationexercise is thought to involve at least three differentmechanisms:
1. Conduction Failure. The muscle actionpotential can fail to be conducted into the fiberalong the T-tubules, which halts the release ofcalcium from the sarcoplasmic reticulum. Theconduction failure results from the buildup ofpotassium ions in the small volume of the T-tubule during the repolarization of repetitiveaction potentials. Elevated external potassiumconcentration leads to a persistentdepolarization of the membrane potential, andeventually a failure to produce action potentialsin the T-tubular membrane (due to inactivationof sodium channels). Recovery is rapid with restas the accumulated potassium diffuses out ofthe tubule, restoring excitability.
2. Lactic Acid Buildup. Elevated hydrogen ionconcentration alters protein conformation andactivity. Thus, the acidification of muscle by lacticacid alters a number of muscle proteins, includingactin and myosin, as well as the proteins involvedin calcium release. The function of theCa2�-ATPase pumps of the sarcoplasmic reticulumis also affected, which may in part explain theimpaired relaxation of fatigued muscle.
3. Inhibition of Cross-Bridge Cycling. Thebuildup of ADP and Pi within muscle fibersduring intense activity may directly inhibitcross-bridge cycling (in particular step 2) bymass action. Slowing the rate of this step delayscross-bridge detachment from actin, and thusslows the overall rate of cross-bridge cycling.These changes contribute to the reducedshortening velocity and impaired relaxationobserved in muscle fatigue resulting from high-intensity exercise.
With low-intensity, long-duration exercise a num-ber of processes have been implicated in fatigue, butno single process can completely account for it. Thethree factors just listed may play minor roles in thistype of exercise as well, but it appears that depletionof fuel substrates may be more important. Althoughdepletion of ATP is not a cause of fatigue, the decreasein muscle glycogen, which is supplying much of thefuel for contraction, correlates closely with fatigue on-set. In addition, low blood glucose (hypoglycemia) anddehydration have been demonstrated to increase fa-
tigue. Thus a certain level of carbohydrate metabolismappears necessary to prevent fatigue during low-intensity exercise, but the mechanism of this require-ment is unknown.
Another type of fatigue quite different from mus-cle fatigue is due to failure of the appropriate regionsof the cerebral cortex to send excitatory signals to themotor neurons. This is called central command fa-tigue, and it may cause a person to stop exercisingeven though the muscles are not fatigued. An athlete’sperformance depends not only on the physical stateof the appropriate muscles but also upon the “will towin”—that is, the ability to initiate central commandsto muscles during a period of increasingly distressfulsensations.
TYPES OF SKELETAL MUSCLE FIBERS
Skeletal muscle fibers do not all have the same me-chanical and metabolic characteristics. Different typesof fibers can be identified on the basis of (1) their max-imal velocities of shortening—fast or slow— and (2)the major pathway used to form ATP—oxidative orglycolytic.
Fast and slow fibers contain forms of myosin thatdiffer in the maximal rates at which they split ATP.This, in turn, determines the maximal rate of cross-bridge cycling and thus the maximal shortening velocity. Fibers containing myosin with high ATPaseactivity are classified as fast fibers, and those con-taining myosin with lower ATPase activity are slowfibers. Although the rate of cross-bridge cycling isabout four times faster in fast fibers than in slowfibers, the force produced by both types of cross-bridges is about the same.
The second means of classifying skeletal musclefibers is according to the type of enzymatic machineryavailable for synthesizing ATP. Some fibers contain numerous mitochondria and thus have a high capacityfor oxidative phosphorylation. These fibers are classi-fied as oxidative fibers. Most of the ATP produced bysuch fibers is dependent upon blood flow to deliveroxygen and fuel molecules to the muscle. Not sur-prisingly, therefore, these fibers are surrounded bymany small blood vessels. They also contain largeamounts of an oxygen-binding protein known as myo-globin, which increases the rate of oxygen diffusionwithin the fiber and provides a small store of oxygen.The large amounts of myoglobin present in oxidativefibers give the fibers a dark-red color, and thus ox-idative fibers are often referred to as red musclefibers.

In addition to these biochemical differences, thereare also size differences. Glycolytic fibers generallyhave much larger diameters than oxidative fibers(Figure 9–28). This fact has significance for tension de-velopment. The number of thick and thin filaments perunit of cross-sectional area is about the same in alltypes of skeletal muscle fibers. Therefore, the larger thediameter of a muscle fiber, the greater the total numberof thick and thin filaments acting in parallel to pro-duce force, and the greater the maximum tension it candevelop (greater strength). Accordingly, the averageglycolytic fiber, with its larger diameter, develops moretension when it contracts than does an average oxida-tive fiber.
These three types of fibers also differ in their ca-pacity to resist fatigue. Fast-glycolytic fibers fatigue rap-idly, whereas slow-oxidative fibers are very resistant tofatigue, which allows them to maintain contractile ac-tivity for long periods with little loss of tension. Fast-oxidative fibers have an intermediate capacity to resistfatigue (Figure 9–29).
The characteristics of the three types of skeletalmuscle fibers are summarized in Table 9–3.
CHAPTER NINE Muscle292
In contrast, glycolytic fibers have few mitochon-dria but possess a high concentration of glycolytic en-zymes and a large store of glycogen. Corresponding totheir limited use of oxygen, these fibers are surroundedby relatively few blood vessels and contain little myo-globin. The lack of myoglobin is responsible for the palecolor of glycolytic fibers and their designation as whitemuscle fibers.
On the basis of these two characteristics, three typesof skeletal muscle fibers can be distinguished:
1. Slow-oxidative fibers (type I) combine lowmyosin-ATPase activity with high oxidativecapacity.
2. Fast-oxidative fibers (type IIa) combine highmyosin-ATPase activity with high oxidativecapacity.
3. Fast-glycolytic fibers (type IIb) combine highmyosin-ATPase activity with high glycolyticcapacity.
Note that the fourth theoretical possibility—slow-glycolytic fibers—is not found.
FIGURE 9–28Cross sections of skeletal muscle. (a) The capillaries surrounding the muscle fibers have been stained. Note the large number of capillaries
surrounding the small-diameter oxidative fibers. (b) The mitochondria have been stained, indicating the large numbers of mitochondria
in the small-diameter oxidative fibers.
Courtesy of John A. Faulkner.
www.mhhe.com/widmaier9 293
muscle has only a single fiber type. Depending on theproportions of the fiber types present, muscles can dif-fer considerably in their maximal contraction speed,strength, and fatigability. For example, the muscles ofthe back, which must be able to maintain their activityfor long periods of time without fatigue while support-ing an upright posture, contain large numbers of slow-oxidative and fast-oxidative fibers. In contrast, the mus-cles in the arms may be called upon to produce largeamounts of tension over a short time period, as whenlifting a heavy object, and these muscles have a greaterproportion of fast-glycolytic fibers.
We will now use the characteristics of single fibersto describe whole muscle contraction and its control.
Control of Muscle TensionThe total tension a muscle can develop depends upontwo factors: (1) the amount of tension developed byeach fiber, and (2) the number of fibers contracting atany time. By controlling these two factors, the nervoussystem controls whole-muscle tension, as well as short-ening velocity. The conditions that determine theamount of tension developed in a single fiber havebeen discussed previously and are summarized inTable 9–4.
The number of fibers contracting at any time de-pends on: (1) the number of fibers in each motor unit(motor unit size), and (2) the number of active motorunits.
Motor unit size varies considerably from one mus-cle to another. The muscles in the hand and eye, whichproduce very delicate movements, contain small motorunits. For example, one motor neuron innervates onlyabout 13 fibers in an eye muscle. In contrast, in the morecoarsely controlled muscles of the legs, each motor unitis large, containing hundreds and in some cases severalthousand fibers. When a muscle is composed of smallmotor units, the total tension produced by the musclecan be increased in small steps by activating additionalmotor units. If the motor units are large, large increasesin tension will occur as each additional motor unit is ac-tivated. Thus, finer control of muscle tension is possi-ble in muscles with small motor units.
The force produced by a single fiber, as we haveseen earlier, depends in part on the fiber diameter—thegreater the diameter, the greater the force. We have alsonoted that fast-glycolytic fibers have the largest diame-ters. Thus, a motor unit composed of 100 fast-glycolyticfibers produces more force than a motor unit composedof 100 slow-oxidative fibers. In addition, fast-glycolyticmotor units tend to have more muscle fibers. For bothof these reasons, activating a fast-glycolytic motorunit will produce more force than activating a slow-oxidative motor unit.
Ten
sio
n (
mg
)T
ensi
on
(m
g)
Ten
sio
n (
mg
)
2 4 6 8
Fast-oxidative fibers
60Time (min)
0
2 4 6 8 60Time (min)
Fast-glycolytic fibers
0
2 4 6 8 60Time (min)
Slow-oxidative fibers
FIGURE 9–29The rate of fatigue development in the three fiber types. Each
vertical line is the contractile response to a brief tetanic stimulus
and relaxation. The contractile responses occurring between
about 9 min and 60 min are not shown on the figure.
WHOLE-MUSCLE CONTRACTIONAs described earlier, whole muscles are made up ofmany muscle fibers organized into motor units. All themuscle fibers in a single motor unit are of the same fibertype. Thus, you can apply the fiber type designation tothe motor unit and refer to slow-oxidative motor units,fast-oxidative motor units, and fast-glycolytic motorunits.
Most muscles are composed of all three motor unittypes interspersed with each other (Figure 9–30). No

294TABLE 9–3 Characteristics of the Three Types of Skeletal Muscle Fibers
SLOW-OXIDATIVE FIBERS FAST-OXIDATIVE FIBERS FAST-GLYCOLYTIC FIBERS
Primary source of ATP Oxidative phosphorylation Oxidative phosphorylation Glycolysisproduction
Mitochondria Many Many Few
Capillaries Many Many Few
Myoglobin content High (red muscle) High (red muscle) Low (white muscle)
Glycolytic enzyme Low Intermediate Highactivity
Glycogen content Low Intermediate High
Rate of fatigue Slow Intermediate Fast
Myosin-ATPase activity Low High High
Contraction velocity Slow Fast Fast
Fiber diameter Small Intermediate Large
Motor unit size Small Intermediate Large
Size of motor neuron Small Intermediate Largeinnervating fiber
I. Tension developed by each fibera. Action potential frequency (frequency-tension relation)b. Fiber length (length-tension relation)c. Fiber diameterd. Fatigue
II. Number of active fibersa. Number of fibers per motor unitb. Number of active motor units
TABLE 9–4 Factors Determining Muscle Tension
Motor unit 2: fast-oxidative fibers
Motor unit 3: fast-glycolytic fibers
Motor unit 1: slow-oxidative fibers
Motor unit1 recruited
Motor unit2 recruited
Motor unit3 recruited
Time0
Wh
ole
-mu
scle
ten
sio
n
(b)
(a)
FIGURE 9–30(a) Diagram of a cross section through a muscle composed of
three types of motor units. (b) Tetanic muscle tension resulting
from the successive recruitment of the three types of motor units.
Note that motor unit 3, composed of fast-glycolytic fibers,
produces the greatest rise in tension because it is composed of
the largest-diameter fibers and contains the largest number of
fibers per motor unit.
The process of increasing the number of motor unitsthat are active in a muscle at any given time is calledrecruitment. It is achieved by activating excitatorysynaptic inputs to more motor neurons. The greater thenumber of active motor neurons, the more motor unitsrecruited, and the greater the muscle tension.
Motor neuron size plays an important role in the re-cruitment of motor units. The size of a motor neuronrefers to the diameter of the nerve cell body, which isusually correlated with the diameter of its axon. Giventhe same number of sodium ions entering a cell at a sin-gle excitatory synapse in a large and in a small motorneuron, the small neuron will undergo a greater depo-larization because these ions will be distributed over asmaller membrane surface area. Accordingly, given thesame level of synaptic input, the smallest neurons willbe recruited first—that is, will begin to generate actionpotentials first. The larger neurons will be recruited onlyas the level of synaptic input increases. Since the small-est motor neurons innervate the slow-oxidative motorunits (see Table 9–3), these motor units are recruitedfirst, followed by fast-oxidative motor units, and finally,during very strong contractions, by fast-glycolytic mo-tor units (Figure 9–30).

www.mhhe.com/widmaier9 295
Thus, during moderate-strength contractions, suchas are used in most endurance types of exercise, rela-tively few fast-glycolytic motor units are recruited, andmost of the activity occurs in oxidative fibers, which aremore resistant to fatigue. The large fast-glycolytic mo-tor units, which fatigue rapidly, begin to be recruitedwhen the intensity of contraction exceeds about 40 per-cent of the maximal tension that can be produced by themuscle.
In conclusion, the neural control of whole-muscletension involves (1) the frequency of action potentialsin individual motor units (to vary the tension gener-ated by the fibers in that unit) and (2) the recruitmentof motor units (to vary the number of active fibers).Most motor neuron activity occurs in bursts of actionpotentials, which produce tetanic contractions of indi-vidual motor units rather than single twitches. Recallthat the tension of a single fiber increases only three-to fivefold when going from a twitch to a maximaltetanic contraction. Therefore, varying the frequency ofaction potentials in the neurons supplying them pro-vides a way to make only three- to fivefold adjustmentsin the tension of the recruited motor units. The force awhole muscle exerts can be varied over a much widerrange than this, from very delicate movements to extremely powerful contractions, by the recruitment of motor units. Thus, recruitment provides the primarymeans of varying tension in a whole muscle. Re-cruitment is controlled by the central commands fromthe motor centers in the brain to the various motor neu-rons (Chapter 10).
Control of Shortening VelocityAs we saw earlier, the velocity at which a single musclefiber shortens is determined by (1) the load on the fiberand (2) whether the fiber is a fast fiber or a slow fiber.Translated to a whole muscle, these characteristics be-come (1) the load on the whole muscle and (2) the typesof motor units in the muscle. For the whole muscle,however, recruitment becomes a third very importantfactor, one that explains how the shortening velocity canbe varied from very fast to very slow even though theload on the muscle remains constant. Consider, for thesake of illustration, a muscle composed of only two mo-tor units of the same size and fiber type. One motor unitby itself will lift a 4-g load more slowly than a 2-g loadbecause the shortening velocity decreases with increas-ing load. When both units are active and a 4-g load islifted, each motor unit bears only half the load, and itsfibers will shorten as if it were lifting only a 2-g load.In other words, the muscle will lift the 4-g load at ahigher velocity when both motor units are active.Recruitment of motor units thus leads to an increase inboth force and velocity.
Muscle Adaptation to ExerciseThe regularity with which a muscle is used, as well asthe duration and intensity of its activity, affects the prop-erties of the muscle. If the neurons to a skeletal muscleare destroyed or the neuromuscular junctions becomenonfunctional, the denervated muscle fibers will be-come progressively smaller in diameter, and the amountof contractile proteins they contain will decrease. Thiscondition is known as denervation atrophy. A musclecan also atrophy with its nerve supply intact if the mus-cle is not used for a long period of time, as when a bro-ken arm or leg is immobilized in a cast. This conditionis known as disuse atrophy.
In contrast to the decrease in muscle mass that re-sults from a lack of neural stimulation, increased amountsof contractile activity—in other words, exercise—canproduce an increase in the size (hypertrophy) of mus-cle fibers as well as changes in their capacity for ATPproduction.
Since the number of fibers in a muscle remains es-sentially constant throughout adult life, the changes inmuscle size with atrophy and hypertrophy do not re-sult from changes in the number of muscle fibers but inthe metabolic capacity and size of each fiber.
Exercise that is of relatively low intensity but of longduration (popularly called “aerobic exercise”), such asrunning and swimming, produces increases in the num-ber of mitochondria in the fibers that are recruited inthis type of activity. In addition, there is an increase inthe number of capillaries around these fibers. All thesechanges lead to an increase in the capacity for enduranceactivity with a minimum of fatigue. (Surprisingly, fiberdiameter decreases slightly, and thus there is a small de-crease in the maximal strength of muscles as a result ofendurance exercise.) As we shall see in later chapters,endurance exercise produces changes not only in theskeletal muscles but also in the respiratory and circula-tory systems, changes that improve the delivery of oxy-gen and fuel molecules to the muscle.
In contrast, short-duration, high-intensity exercise(popularly called “strength training”), such as weightlifting, affects primarily the fast-glycolytic fibers, whichare recruited during strong contractions. These fibersundergo an increase in fiber diameter (hypertrophy)due to the increased synthesis of actin and myosin fila-ments, which form more myofibrils. In addition, the gly-colytic activity is increased by increasing the synthesisof glycolytic enzymes. The result of such high-intensityexercise is an increase in the strength of the muscle andthe bulging muscles of a conditioned weight lifter. Suchmuscles, although very powerful, have little capacity forendurance, and they fatigue rapidly.
Exercise produces limited change in the types ofmyosin enzymes formed by the fibers and thus little

of exercise leads to muscle soreness the next day. Thissoreness is the result of a mild inflammation in the mus-cle, which occurs whenever tissues are damaged(Chapter 18). The most severe inflammation occursfollowing a period of lengthening contractions, indicat-ing that the lengthening of a muscle fiber by an externalforce produces greater muscle damage than do eitherisotonic or isometric contractions. Thus, exercising by gradually lowering weights will produce greatermuscle soreness than an equivalent amount of weightlifting.
The effects of anabolic steroids on skeletal musclegrowth and strength are described in Chapter 17.
Lever Action of Muscles and BonesA contracting muscle exerts a force on bones throughits connecting tendons. When the force is great enough,the bone moves as the muscle shortens. A contractingmuscle exerts only a pulling force, so that as the mus-cle shortens, the bones to which it is attached are pulledtoward each other. Flexion refers to the bending of a limbat a joint, whereas extension is the straightening of a limb(Figure 9–31). These opposing motions require at leasttwo muscles, one to cause flexion and the other exten-sion. Groups of muscles that produce oppositely di-rected movements at a joint are known as antagonists.For example, from Figure 9–31 it can be seen that con-traction of the biceps causes flexion of the arm at the el-bow, whereas contraction of the antagonistic muscle, thetriceps, causes the arm to extend. Both muscles exertonly a pulling force upon the forearm when theycontract.
Sets of antagonistic muscles are required not onlyfor flexion-extension, but also for side-to-side move-ments or rotation of a limb. The contraction of some mus-cles leads to two types of limb movement, depending onthe contractile state of other muscles acting on the samelimb. For example, contraction of the gastrocnemiusmuscle in the leg causes a flexion of the leg at the knee,as in walking (Figure 9–32). However, contraction of thegastrocnemius muscle with the simultaneous contrac-tion of the quadriceps femoris (which causes extensionof the lower leg) prevents the knee joint from bending,leaving only the ankle joint capable of moving. The footis extended, and the body rises on tiptoe.
The muscles, bones, and joints in the body arearranged in lever systems. The basic principle of a leveris illustrated by the flexion of the arm by the biceps mus-cle (Figure 9–33), which exerts an upward pulling forceon the forearm about 5 cm away from the elbow joint.In this example, a 10-kg weight held in the hand exertsa downward force of 10 kg about 35 cm from the elbow.A law of physics tells us that the forearm is in mechan-
CHAPTER NINE Muscle296
change in the proportions of fast and slow fibers in amuscle. As described previously, however, exercise doeschange the rates at which metabolic enzymes are syn-thesized, leading to changes in the proportion of ox-idative and glycolytic fibers within a muscle. With endurance training, there is a decrease in the number offast-glycolytic fibers and an increase in the number offast-oxidative fibers as the oxidative capacity of thefibers is increased. The reverse occurs with strengthtraining as fast-oxidative fibers are converted to fast-glycolytic fibers.
The signals responsible for all these changes in mus-cle with different types of activity are unknown. Theyare related to the frequency and intensity of the con-tractile activity in the muscle fibers and thus to the pat-tern of action potentials produced in the muscle over anextended period of time.
Because different types of exercise produce quitedifferent changes in the strength and endurance capac-ity of a muscle, an individual performing regularexercises to improve muscle performance must choosea type of exercise that is compatible with the type ofactivity he or she ultimately wishes to perform. Thus,lifting weights will not improve the endurance of a long-distance runner, and jogging will not produce theincreased strength desired by a weight lifter. Most ex-ercises, however, produce some effects on both strengthand endurance.
These changes in muscle in response to repeated pe-riods of exercise occur slowly over a period of weeks.If regular exercise is stopped, the changes in the mus-cle that occurred as a result of the exercise will slowlyrevert to their unexercised state.
The maximum force generated by a muscle de-creases by 30 to 40 percent between the ages of 30 and80. This decrease in tension-generating capacity is dueprimarily to a decrease in average fiber diameter. Someof the change is simply the result of diminishing phys-ical activity with age and can be prevented by exerciseprograms. The ability of a muscle to adapt to exercise,however, decreases with age: The same intensity andduration of exercise in an older individual will not pro-duce the same amount of change as in a younger person.
This effect of aging, however, is only partial, andthere is no question that even in the elderly, exercise canproduce significant adaptation. Aerobic training has re-ceived major attention because of its effect on the car-diovascular system (Chapter 12). Strength training of amodest degree, however, is also strongly recommendedbecause it can partially prevent the loss of muscle tis-sue that occurs with aging. Moreover, it helps maintainstronger bones (Chapter 14).
Extensive exercise by an individual whose muscleshave not been used in performing that particular type
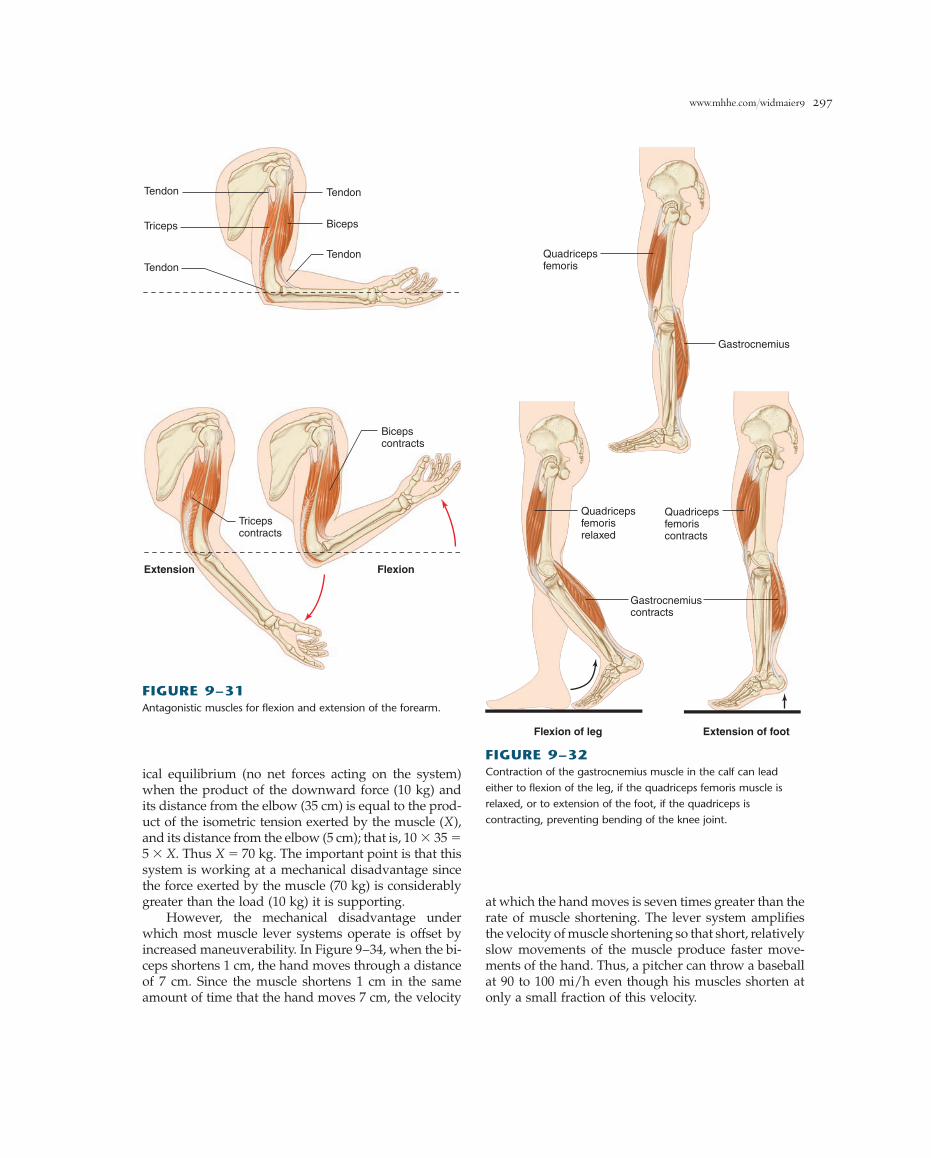
www.mhhe.com/widmaier9 297
ical equilibrium (no net forces acting on the system)when the product of the downward force (10 kg) andits distance from the elbow (35 cm) is equal to the prod-uct of the isometric tension exerted by the muscle (X),and its distance from the elbow (5 cm); that is, 10 � 35 �5 � X. Thus X � 70 kg. The important point is that thissystem is working at a mechanical disadvantage sincethe force exerted by the muscle (70 kg) is considerablygreater than the load (10 kg) it is supporting.
However, the mechanical disadvantage underwhich most muscle lever systems operate is offset byincreased maneuverability. In Figure 9–34, when the bi-ceps shortens 1 cm, the hand moves through a distanceof 7 cm. Since the muscle shortens 1 cm in the sameamount of time that the hand moves 7 cm, the velocity
at which the hand moves is seven times greater than therate of muscle shortening. The lever system amplifiesthe velocity of muscle shortening so that short, relativelyslow movements of the muscle produce faster move-ments of the hand. Thus, a pitcher can throw a baseballat 90 to 100 mi/h even though his muscles shorten atonly a small fraction of this velocity.
Tendon
Tendon
Tendon
Tendon
Triceps
Tricepscontracts
Biceps
Bicepscontracts
Extension Flexion
FIGURE 9–31Antagonistic muscles for flexion and extension of the forearm.
Quadricepsfemoris
Gastrocnemius
Gastrocnemiuscontracts
Quadricepsfemorisrelaxed
Quadricepsfemoriscontracts
Flexion of leg Extension of foot
FIGURE 9–32Contraction of the gastrocnemius muscle in the calf can lead
either to flexion of the leg, if the quadriceps femoris muscle is
relaxed, or to extension of the foot, if the quadriceps is
contracting, preventing bending of the knee joint.

CHAPTER NINE Muscle298
A D D I T I O N A L C L I N I C A L E X A M P L E S
A number of diseases can affect the contraction of skeletalmuscle. Many of them are due to defects in the parts of thenervous system that control contraction of the muscle fibersrather than to defects in the muscle fibers themselves. For ex-ample, poliomyelitis is a viral disease that destroys motorneurons, leading to the paralysis of skeletal muscle, and mayresult in death due to respiratory failure.
MUSCLE CRAMPSInvoluntary tetanic contraction of skeletal muscles producesmuscle cramps. During cramping, nerve action potentials fireat abnormally high rates, a much greater rate than occurs dur-ing maximal voluntary contraction. The specific cause of thishigh activity is uncertain but is probably related to electrolyteimbalances in the extracellular fluid surrounding both themuscle and nerve fibers. These imbalances may arise fromoverexercise or persistent dehydration, and they can directlyinduce action potentials in motor neurons and muscle fibers.Another theory is that sensory receptors in the muscle arestimulated by chemical imbalances within the muscle, and themotor neurons to the area are reflexly activated when thosesignals reach the spinal cord.
HYPOCALCEMIC TETANYSimilar in symptoms to muscular cramping is hypocalcemictetany, the involuntary tetanic contraction of skeletal musclesthat occurs when the extracellular calcium concentration fallsto about 40 percent of its normal value. This may seem sur-prising, since we have seen that calcium is required forexcitation-contraction coupling. However, recall that this cal-cium is sarcoplasmic reticulum calcium, not extracellularcalcium. The effect of changes in extracellular calcium is ex-erted not on the sarcoplasmic reticulum calcium, but directlyon the plasma membrane. Low extracellular calcium(hypocalcemia) increases the opening of sodium channels inexcitable membranes, leading to membrane depolarizationand the spontaneous firing of action potentials. It is this thatcauses the increased muscle contractions. The mechanismscontrolling the extracellular concentration of calcium ions arediscussed in Chapter 14.
MUSCULAR DYSTROPHYThis disease is one of the most frequently encountered geneticdiseases, affecting one in every 3500 males (but much lesscommonly in females) born in America. Muscular dystrophy
X = 70 kg
10 kg x 35 cm = X x 5 cmX = 70 kg
5 cm 30 cm 10 kg
10 kg
FIGURE 9–33Mechanical equilibrium of forces acting on the forearm while
supporting a 10-kg load.
Force
1 cm
7 cm
Vm = musclecontractionvelocity
Vh = handvelocity
= 7 x Vm
FIGURE 9–34Velocity of the biceps muscle is amplified by the lever system of
the arm, producing a greater velocity of the hand. The range of
movement is also amplified (1 cm of shortening by the muscle
produces 7 cm of movement by the hand).

www.mhhe.com/widmaier9 299
1 2 3 4 5
FIGURE 9–35Boy with Duchenne muscular dystrophy. Muscles of the hip girdle and trunk are the first to weaken, requiring patients to use their
arms to climb up the legs in attempting to go from lying to standing.
is associated with the progressive degeneration of skeletal andcardiac muscle fibers, weakening the muscles and leading ul-timately to death from respiratory or cardiac failure (Figure9 – 35). The symptoms become evident at about 2 to 6 yearsof age, and most affected individuals do not survive much be-yond the age of 20.
The recessive gene responsible for a major form of mus-cular dystrophy (Duchenne muscular dystrophy) has beenidentified on the X chromosome, and muscular dystrophy isa sex-linked recessive disease. (As described in Chapter 17,girls have two X chromosomes and boys only one.Accordingly, a girl with one abnormal X chromosome andone normal one will not generally develop the disease. Thisis why the disease is so much more common in boys.) Thisgene codes for a protein known as dystrophin, which is ei-ther absent or present in a nonfunctional form in patientswith the disease. Dystrophin is a large protein that links cy-toskeletal proteins to membrane glycoproteins. It resemblesother known cytoskeletal proteins and may be involved inmaintaining the structural integrity of the plasma membraneor of elements within the membrane, such as ion channels,in fibers subjected to repeated structural deformation dur-ing contraction. Preliminary attempts are being made to treatthe disease by inserting the normal gene into dystrophicmuscle cells.
Myasthenia GravisMyasthenia gravis is a collection of neuromuscular disor-ders characterized by muscle fatigue and weakness that pro-
gressively worsens as the muscle is used. It affects about12,000 Americans. One type of the disease results from a de-crease in the number of ACh receptors on the motor endplate. The release of ACh from the nerve terminals is nor-mal, but the magnitude of the end-plate potential ismarkedly reduced because of the decreased number of re-ceptors. Even in normal muscle, the amount of ACh releasedwith each action potential decreases with repetitive activity,and thus the magnitude of the resulting EPP falls. In normalmuscle, however, the EPP remains well above the thresholdnecessary to initiate a muscle action potential. In contrast, af-ter a few motor nerve impulses in a myasthenia gravis pa-tient, the magnitude of the EPP falls below the threshold forinitiating a muscle action potential. As described in Chapter18, the destruction of the ACh receptors is brought about bythe body’s own defense mechanisms gone awry, specificallybecause of the formation of antibodies to the nicotinic ACh-receptor proteins.
A number of approaches are currently used to treat thedisease. One is to administer acetylcholinesterase inhibitors(e.g., neostygmine). This can partially compensate for the re-duction in ACh receptor numbers by prolonging the timethat acetylcholine is available at the synapse. Other thera-pies aim at blunting the immune response. Removal of thethymus gland (thymectomy) reduces the production of anti-bodies and reverses symptoms in about 50 percent of pa-tients. Plasmapheresis is a treatment that involves removingthe liquid fraction of blood (plasma), which contains the of-fending antibodies. A combination of these treatments hasgreatly reduced the mortality rate for myasthenia gravis.

CHAPTER NINE Muscle300
IV. Branches of a motor neuron axon formneuromuscular junctions with the muscle fibers in itsmotor unit. Each muscle fiber is innervated by abranch from only one motor neuron.a. Acetylcholine released by an action potential in
a motor neuron binds to receptors on the motor endplate of the muscle membrane, opening ion channelsthat allow the passage of sodium and potassiumions, which depolarize the end-plate membrane.
b. A single action potential in a motor neuron issufficient to produce an action potential in askeletal muscle fiber.
c. Figure 9–19 summarizes events at theneuromuscular junction.
V. Table 9–2 summarizes the events leading to thecontraction of a skeletal muscle fiber.
Mechanics of Single-Fiber ContractionI. Contraction refers to the turning on of the cross-
bridge cycle. Whether there is an accompanyingchange in muscle length depends upon the externalforces acting on the muscle.
II. Three types of contractions can occur followingactivation of a muscle fiber: (1) an isometriccontraction in which the muscle generates tension butdoes not change length; (2) an isotonic contraction inwhich the muscle shortens, moving a load; and (3) alengthening contraction in which the external load onthe muscle causes the muscle to lengthen during theperiod of contractile activity.
III. Increasing the frequency of action potentials in amuscle fiber increases the mechanical response(tension or shortening), up to the level of maximaltetanic tension.
IV. Maximum isometric tetanic tension is produced at theoptimal sarcomere length lo. Stretching a fiber beyondits optimal length or decreasing the fiber lengthbelow lo decreases the tension generated.
V. The velocity of muscle fiber shortening decreaseswith increases in load. Maximum velocity occurs atzero load.
Skeletal Muscle Energy MetabolismI. Muscle fibers form ATP by the transfer of phosphate
from creatine phosphate to ADP, by oxidativephosphorylation of ADP in mitochondria, and bysubstrate-level phosphorylation of ADP in theglycolytic pathway.
II. At the beginning of exercise, muscle glycogen isthe major fuel consumed. As the exercise proceeds,glucose and fatty acids from the blood provide mostof the fuel, fatty acids becoming progressively moreimportant during prolonged exercise. When theintensity of exercise exceeds about 70 percent ofmaximum, glycolysis begins to contribute anincreasing fraction of the total ATP generated.
III. Muscle fatigue is caused by a variety of factors,including internal changes in acidity, cross-bridgeinhibition, glycogen depletion, and excitation-contraction coupling failure, but not by a lack of ATP.
S E C T I O N A S U M M A R Y
I. There are three types of muscle—skeletal, smooth,and cardiac. Skeletal muscle is attached to bones andmoves and supports the skeleton. Smooth musclesurrounds hollow cavities and tubes. Cardiac muscleis the muscle of the heart.
StructureI. Skeletal muscles, composed of cylindrical muscle
fibers (cells), are linked to bones by tendons at eachend of the muscle.
II. Skeletal muscle fibers have a repeating, striatedpattern of light and dark bands due to thearrangement of the thick and thin filaments withinthe myofibrils.
III. Actin-containing thin filaments are anchored to theZ lines at each end of a sarcomere, while their freeends partially overlap the myosin-containing thickfilaments in the A band at the center of thesarcomere.
Molecular Mechanisms of ContractionI. When a skeletal muscle fiber actively shortens, the
thin filaments are propelled toward the center of theirsarcomere by movements of the myosin cross-bridgesthat bind to actin.a. The two globular heads of each cross-bridge
contain a binding site for actin and an enzymaticsite that splits ATP.
b. The four steps occurring during each cross-bridgecycle are summarized in Figure 9–12. The cross-bridges undergo repeated cycles during acontraction, each cycle producing only a smallincrement of movement.
c. The three functions of ATP in muscle contractionare summarized in Table 9–1.
II. In a resting muscle, attachment of cross-bridges toactin is blocked by tropomyosin molecules that arein contact with the actin subunits of the thinfilaments.
III. Contraction is initiated by an increase in cytosoliccalcium concentration. The calcium ions bind totroponin, producing a change in its shape that istransmitted via tropomyosin to uncover the bindingsites on actin, allowing the cross-bridges to bind tothe thin filaments.a. The rise in cytosolic calcium concentration is
triggered by an action potential in the plasmamembrane. The action potential is propagated intothe interior of the fiber along the transversetubules to the region of the sarcoplasmicreticulum, where it produces a release of calciumions from the reticulum.
b. Relaxation of a contracting muscle fiber occurs asa result of the active transport of cytosoliccalcium ions back into the sarcoplasmicreticulum.

www.mhhe.com/widmaier9 301
Types of Skeletal Muscle FibersI. Three types of skeletal muscle fibers can be
distinguished by their maximal shortening velocitiesand the predominate pathway used to form ATP:slow-oxidative, fast-oxidative, and fast-glycolyticfibers.a. Differences in maximal shortening velocities are
due to different myosin enzymes with high or lowATPase activities, giving rise to fast and slowfibers.
b. Fast-glycolytic fibers have a larger averagediameter than oxidative fibers and thereforeproduce greater tension, but they also fatiguemore rapidly.
II. All the muscle fibers in a single motor unit belong tothe same fiber type, and most muscles contain allthree types.
III. Table 9–3 summarizes the characteristics of the threetypes of skeletal muscle fibers.
Whole-Muscle ContractionI. The tension produced by whole-muscle contraction
depends on the amount of tension developed by eachfiber and the number of active fibers in the muscle(Table 9–4).
II. Muscles that produce delicate movements have asmall number of fibers per motor unit, whereas largepostural muscles have much larger motor units.
III. Fast-glycolytic motor units not only have large-diameter fibers but also tend to have large numbersof fibers per motor unit.
IV. Increases in muscle tension are controlled primarilyby increasing the number of active motor units in amuscle, a process known as recruitment. Slow-oxidative motor units are recruited first during weakcontractions, then fast-oxidative motor units, andfinally fast-glycolytic motor units during very strongcontractions.
V. Increasing motor-unit recruitment increases thevelocity at which a muscle will move a given load.
VI. The strength and susceptibility to fatigue of a musclecan be altered by exercise.a. Long-duration, low-intensity exercise increases a
fiber’s capacity for oxidative ATP production byincreasing the number of mitochondria and bloodvessels in the muscle, resulting in increasedendurance.
b. Short-duration, high-intensity exercise increasesfiber diameter as a result of increased synthesis ofactin and myosin, resulting in increased strength.
VII. Movement around a joint requires two antagonisticgroups of muscles: one flexes the limb at the joint,and the other extends the limb.
VIII. The lever system of muscles and bones requiresmuscle tensions far greater than the load in order tosustain a load in an isometric contraction, but thelever system produces a shortening velocity at theend of the lever arm that is greater than the muscle-shortening velocity.
A band 270acetylcholine (ACh) 280acetylcholinesterase 282actin 269antagonist 295cardiac muscle 268central command
fatigue 290concentric contraction 283contraction 270contraction time 284creatine phosphate 288cross-bridge 270cross-bridge cycle 272dihydropyridine (DHP)
receptor 279eccentric contraction 283end-plate potential
(EPP) 281excitation-contraction
coupling 277extension 295fast fiber 290fast-glycolytic fiber 291fast-oxidative fiber 291flexion 295foot proteins 278fused tetanus 285glycolytic fiber 291H zone 270heavy chains 271hypertrophy 268hypocalcemia 297I band 270isometric contraction 283isotonic contraction 283junctional feet 278latent period 284lateral sac 277lengthening contraction 283light chains 271load 283M line 270motor end plate 281
motor neuron 280motor unit 280muscle 268muscle fatigue 289muscle fiber 268myoblast 268myofibril 269myoglobin 291myosin 269neuromuscular junction 281optimal length (lo) 287oxidative fiber 291oxygen debt 289power stroke 274recruitment 293red muscle fiber 291relaxation 270rigor mortis 276ryanodine receptor 279sarcomere 269sarcoplasmic reticulum 277satellite cell 268skeletal muscle 268sliding-filament
mechanism 270slow fiber 290slow-oxidative fiber 291smooth muscle 268striated muscle 269summation 285tendon 268tension 283tetanus 285thick filament 269thin filament 269titin 270transverse tubule
(T-tubule) 277tropomyosin 276troponin 276twitch 284unfused tetanus 285white muscle fiber 291Z line 270
S E C T I O N A C L I N I C A L T E R M S
atropine 283botulism 283curare 283denervation atrophy 294disuse atrophy 294duchenne muscular
dystrophy 298gallamine 283
hypocalcemic tetany 297muscle cramps 297muscular dystrophy 298myasthenia gravis 298neostigmine 298plasmapheresis 298poliomyelitis 297thymectomy 298
S E C T I O N A K EY T E R M S

CHAPTER NINE Muscle302
S E C T I O N A R E V I E W Q U E S T I O N S
1. List the three types of muscle cells and their locations.2. Diagram the arrangement of thick and thin filaments
in a striated muscle sarcomere, and label the majorbands that give rise to the striated pattern.
3. Describe the organization of myosin and actinmolecules in the thick and thin filaments.
4. Describe the four steps of one cross-bridge cycle.5. Describe the physical state of a muscle fiber in rigor
mortis and the conditions that produce this state.6. What three events in skeletal muscle contraction and
relaxation are dependent on ATP?7. What prevents cross-bridges from attaching to sites
on the thin filaments in a resting skeletal muscle?8. Describe the role and source of calcium ions in
initiating contraction in skeletal muscle.9. Describe the location, structure, and function of the
sarcoplasmic reticulum in skeletal muscle fibers.10. Describe the structure and function of the transverse
tubules.11. Describe the events that result in the relaxation of
skeletal muscle fibers.12. Define a motor unit and describe its structure.13. Describe the sequence of events by which an action
potential in a motor neuron produces an actionpotential in the plasma membrane of a skeletalmuscle fiber.
14. What is an end-plate potential, and what ionsproduce it?
15. Compare and contrast the transmission of electricalactivity at a neuromuscular junction with that at asynapse.
16. Describe isometric, isotonic, and lengtheningcontractions.
17. What factors determine the duration of an isotonictwitch in skeletal muscle? An isometric twitch?
18. What effect does increasing the frequency of actionpotentials in a skeletal muscle fiber have upon theforce of contraction? Explain the mechanismresponsible for this effect.
19. Describe the length-tension relationship in striatedmuscle fibers.
20. Describe the effect of increasing the load on a skeletalmuscle fiber on the velocity of shortening.
21. What is the function of creatine phosphate in skeletalmuscle contraction?
22. What fuel molecules are metabolized to produce ATPduring skeletal muscle activity?
23. List the factors responsible for skeletal musclefatigue.
24. What component of skeletal muscle fibers accountsfor the differences in the fibers’ maximal shorteningvelocities?
25. Summarize the characteristics of the three types ofskeletal muscle fibers.
26. Upon what two factors does the amount of tension developed by a whole skeletal muscledepend?
27. Describe the process of motor-unit recruitment incontrolling (a) whole-muscle tension and (b) velocityof whole-muscle shortening.
28. During increases in the force of skeletal musclecontraction, what is the order of recruitment of thedifferent types of motor units?
29. What happens to skeletal muscle fibers when themotor neuron to the muscle is destroyed?
30. Describe the changes that occur in skeletal musclesfollowing a period of (a) long-duration, low-intensityexercise training; and (b) short-duration, high-intensity exercise training.
31. How are skeletal muscles arranged around joints sothat a limb can push or pull?
32. What are the advantages and disadvantages of themuscle-bone-joint lever system?
We now examine the second of the three types of mus-cle found in the body—smooth muscle. Two character-istics are common to all smooth muscles: they lack thecross-striated banding pattern found in skeletal and car-diac fibers (which makes them “smooth”), and thenerves to them are derived from the autonomic divisionof the nervous system rather than the somatic division.Thus, smooth muscle is not normally under direct vol-untary control.
Smooth muscle, like skeletal muscle, uses cross-bridge movements between actin and myosin filamentsto generate force, and calcium ions to control cross-bridge activity. However, the organization of the
contractile filaments and the process of excitation-contraction coupling are quite different in these twotypes of muscle. Furthermore, there is considerable di-versity among smooth muscles with respect to themechanism of excitation-contraction coupling.
STRUCTURE OF SMOOTH MUSCLE
Each smooth muscle fiber is a spindle-shaped cell witha diameter ranging from 2 to 10 �m, as compared to arange of 10 to 100 �m for skeletal muscle fibers (see
S E C T I O N B
S m o o t h M u s c l e

www.mhhe.com/widmaier9 303
a banding pattern (Figure 9–37). Nevertheless, smoothmuscle contraction occurs by a sliding-filament mech-anism.
The concentration of myosin in smooth muscle isonly about one-third of that in striated muscle, whereasthe actin content can be twice as great. In spite of thesedifferences, the maximal tension per unit of cross-sectional area developed by smooth muscles is similarto that developed by skeletal muscle.
The isometric tension produced by smooth musclefibers varies with fiber length in a manner qualitativelysimilar to that observed in skeletal muscle. There is anoptimal length at which tension development is maxi-mal, and less tension is generated at lengths shorter orlonger than this optimal length. The range of musclelengths over which smooth muscle is able to developtension is greater, however, than it is in skeletal muscle.This property is highly adaptive since most smoothmuscles surround hollow organs that undergo changesin volume with accompanying changes in the lengthsof the smooth muscle fibers in their walls. Even withrelatively large increases in volume, as during the ac-cumulation of large amounts of urine in the bladder, thesmooth muscle fibers in the wall retain some ability todevelop tension, whereas such distortion might stretchskeletal muscle fibers beyond the point of thick- andthin-filament overlap.
SMOOTH MUSCLECONTRACTION AND ITS CONTROL
Changes in cytosolic calcium concentration control thecontractile activity in smooth muscle fibers, as in stri-ated muscle. However, there are significant differencesbetween the two types of muscle in the way in which
Figure 9–3). While skeletal muscle fibers are multinu-cleate cells that are unable to divide once they have dif-ferentiated, smooth muscle fibers have a single nucleusand have the capacity to divide throughout the life ofan individual. Smooth muscle cells can be stimulated todivide by a variety of paracrine agents, often in responseto tissue injury.
Two types of filaments are present in the cyto-plasm of smooth muscle fibers (Figure 9–36): thickmyosin-containing filaments and thin actin-containingfilaments. The thin filaments are anchored either tothe plasma membrane or to cytoplasmic structuresknown as dense bodies, which are functionally similarto the Z lines in skeletal muscle fibers. Note in Figure9–36 that the filaments are oriented slightly diago-nally to the long axis of the cell. When the fiber short-ens, the regions of the plasma membrane betweenthe points where actin is attached to the membraneballoon out. The thick and thin filaments are not organized into myofibrils, as in striated muscles, and there is no regular alignment of these filamentsinto sarcomeres, which accounts for the absence of
Relaxed Contracted
Dense bodies
Nucleus
ThickfilamentsThin
filaments
FIGURE 9–36Thick and thin filaments in smooth muscle are arranged in
slightly diagonal chains that are anchored to the plasma
membrane or to dense bodies within the cytoplasm. When
activated, the thick and thin filaments slide past each other,
causing the smooth muscle fiber to shorten and thicken.
FIGURE 9–37Photomicrograph of a sheet of smooth muscle cells. Note the
spindle shape, single nucleus, and lack of striations.

calcium activates cross-bridge cycling and in the mech-anisms by which stimulation leads to alterations incalcium concentration.
Cross-Bridge ActivationThe thin filaments in smooth muscle do not have thecalcium-binding protein troponin that mediates cal-cium-triggered cross-bridge activity in both skeletal andcardiac muscle. Instead, cross-bridge cycling in smoothmuscle is controlled by a calcium-regulated enzyme that phos-phorylates myosin. Only the phosphorylated form ofsmooth muscle myosin can bind to actin and undergocross-bridge cycling.
The following sequence of events occurs after a risein cytosolic calcium in a smooth muscle fiber (Figure9–38): (1) Calcium binds to calmodulin, a calcium-bind-
ing protein that is present in the cytoplasm of most cells(Chapter 5) and whose structure is related to that of tro-ponin. (2) The calcium-calmodulin complex binds to an-other cytosolic protein, myosin light-chain kinase,thereby activating the enzyme. (3) Active myosin light-chain kinase then uses ATP to phosphorylate myosinlight chains in the globular head of myosin. (4) Thephosphorylated cross-bridge binds to actin. A key dif-ference here is that cross-bridge activity in smooth mus-cle is turned on by calcium-mediated changes in thethick filaments, whereas in striated muscle, calcium me-diates changes in the thin filaments.
The smooth muscle form of myosin has a very lowmaximal rate of ATPase activity, on the order of 10to 100 times less than that of skeletal muscle myosin.Since the rate of ATP splitting determines the rate ofcross-bridge cycling and thus shortening velocity,
CHAPTER NINE Muscle304
Cytosolic Ca2+
Ca2+ binds to calmodulinin cytosol
Ca2+–calmodulin complexbinds to myosin
light-chain kinase
Myosin light-chain kinaseuses ATP to phosphorylate
myosin cross-bridges
Phosphorylatedcross-bridges
bind to actin filaments
Cross-bridge cycleproduces tension and
shortening
Ca2+ binds to troponinon thin filaments
Conformational changein troponin movestropomyosin out ofblocking position
Myosin cross-bridgesbind to actin
Cross-bridge cycleproduces tension and
shortening
Smooth muscle Skeletal muscle
Cytosolic Ca2+
FIGURE 9–38Pathways leading from increased cytosolic calcium to cross-bridge cycling in smooth and skeletal muscle fibers.

www.mhhe.com/widmaier9 305
tubules connected to the plasma membrane in smoothmuscle. The small fiber diameter and the slow rate ofcontraction do not require such a rapid mechanism forgetting an excitatory signal into the muscle fiber.Portions of the sarcoplasmic reticulum are located nearthe plasma membrane, however, forming associationssimilar to the relationship between T tubules and thelateral sacs in skeletal muscle. Action potentials in theplasma membrane can be coupled to the release of sar-coplasmic reticulum calcium at these sites. In addition,second messengers released from the plasma mem-brane or generated in the cytosol in response to thebinding of extracellular chemical messengers toplasma-membrane receptors, can trigger the release ofcalcium from the more centrally located sarcoplasmicreticulum.
What about extracellular calcium in excitation-contraction coupling? There are voltage-sensitive cal-cium channels in the plasma membranes of smoothmuscle cells, as well as calcium channels controlledby extracellular chemical messengers. Since the con-centration of calcium in the extracellular fluid is10,000 times greater than in the cytosol, the openingof calcium channels in the plasma membrane resultsin an increased flow of calcium into the cell. Becauseof the small cell size, the entering calcium does nothave far to diffuse to reach binding sites within thecell.
Removal of calcium from the cytosol to bring aboutrelaxation is achieved by the active transport of calciumback into the sarcoplasmic reticulum as well as out ofthe cell across the plasma membrane. The rate of cal-cium removal in smooth muscle is much slower than inskeletal muscle, with the result that a single twitch lastsseveral seconds in smooth muscle but lasts only a frac-tion of a second in skeletal muscle.
Moreover, whereas in skeletal muscle a single ac-tion potential releases sufficient calcium to saturate alltroponin sites on the thin filaments, only a portion ofthe cross-bridges are activated in a smooth muscle fiberin response to most stimuli. Therefore, the tension gen-erated by a smooth muscle fiber can be graded byvarying cytosolic calcium concentration. The greaterthe increase in calcium concentration, the greater thenumber of cross-bridges activated, and the greater thetension.
In some smooth muscles, the cytosolic calcium con-centration is sufficient to maintain a low level of basalcross-bridge activity in the absence of external stimuli.This activity is known as smooth muscle tone. Its in-tensity can be varied by factors that alter the cytosoliccalcium concentration.
As in our description of skeletal muscle, we haveapproached the question of excitation-contraction
smooth muscle shortening is much slower than that ofskeletal muscle. Due to this slow rate of energy usage,smooth muscle does not undergo fatigue during pro-longed periods of activity.
To relax a contracted smooth muscle, myosinmust be dephosphorylated because dephosphorylatedmyosin is unable to bind to actin. This dephosphoryla-tion is mediated by the enzyme myosin light-chainphosphatase, which is continuously active in smoothmuscle during periods of rest and contraction. Whencytosolic calcium rises, the rate of myosin phosphory-lation by the activated kinase exceeds the rate of de-phosphorylation by the phosphatase, and the amountof phosphorylated myosin in the cell increases, pro-ducing a rise in tension. When the cytosolic calciumconcentration decreases, the rate of dephosphoryla-tion exceeds the rate of phosphorylation, and thea-mount of phosphorylated myosin decreases, producingrelaxation.
In some smooth muscles, when stimulation ispersistent and cytosolic calcium concentration re-mains elevated, the rate of ATP splitting by the cross-bridges declines even though isometric tension ismaintained. This condition is known as the latch stateand occurs when a phosphorylated cross-bridge be-comes dephosphorylated while still attached to actin.In this circumstance it can maintain tension in a rig-orlike state without movement. Dissociation of thesedephosphorylated cross-bridges from actin by thebinding of ATP occurs at a much slower rate than dis-sociation of phosphorylated bridges. The net result isthe ability to maintain tension for long periods oftime with a very low rate of ATP consumption. Agood example of the usefulness of this mechanism isseen in blood vessel walls, where smooth musclemust maintain diameter for prolonged periodsagainst persistent pressure.
Sources of Cytosolic CalciumTwo sources of calcium contribute to the rise in cytoso-lic calcium that initiates smooth muscle contraction: (1)the sarcoplasmic reticulum and (2) extracellular calciumentering the cell through plasma-membrane calciumchannels. The amount of calcium contributed by thesetwo sources differs among various smooth muscles,some being more dependent on extracellular calciumthan the stores in the sarcoplasmic reticulum, and viceversa.
First we’ll examine the role of the sarcoplasmicreticulum. The total quantity of this organelle insmooth muscle is smaller than in skeletal muscle, andit is not arranged in any specific pattern in relation tothe thick and thin filaments. Moreover, there are no T-

coupling in smooth muscle by first describing thecoupling (the changes in cytosolic calcium). Now wemust back up a step and ask what constitutes the ex-citation that elicits these changes in calcium concen-tration.
Membrane ActivationIn contrast to skeletal muscle, in which membrane acti-vation is dependent on a single input—the somatic neu-rons to the muscle—many inputs to a smooth muscleplasma membrane can alter the contractile activity ofthe muscle (Table 9–5). Some of these increase contrac-tion while others inhibit it. Moreover, at any one time,multiple inputs may be occurring, with the contractilestate of the muscle dependent on the relative intensityof the various inhibitory and excitatory stimuli. All theseinputs influence contractile activity by altering cytoso-lic calcium concentration as described in the previoussection.
Some smooth muscles contract in response to mem-brane depolarization, whereas others can contract inthe absence of any membrane potential change. In-terestingly, in smooth muscles in which action poten-tials occur, calcium ions, rather than sodium ions, carrypositive charge into the cell during the rising phase ofthe action potential—that is, depolarization of the mem-brane opens voltage-gated calcium channels, producingcalcium-mediated action potentials rather than sodium-mediated ones.
Another very important point needs to be madeabout electrical activity and cytosolic calcium concen-tration in smooth muscle. Unlike the situation in skeletalmuscle, in smooth muscle cytosolic calcium concentra-tion can be increased (or decreased) by graded depolar-izations (or hyperpolarizations) in membrane potential,which increase or decrease the number of open calciumchannels.
Spontaneous Electrical ActivitySome types of smooth muscle fibers generate actionpotentials spontaneously in the absence of any neu-ral or hormonal input. The plasma membranes ofsuch fibers do not maintain a constant resting poten-tial. Instead, they gradually depolarize until theyreach the threshold potential and produce an actionpotential. Following repolarization, the membraneagain begins to depolarize (Figure 9–39), so that a se-quence of action potentials occurs, producing a tonicstate of contractile activity. The membrane potentialchange occurring during the spontaneous depolar-ization to threshold is known as a pacemaker potential.Pacemaker cells are found throughout the gastroin-testinal tract, and thus gut smooth muscle tends to con-tract rhythmically even in the absence of neural input.As described in other chapters, some cardiac musclefibers and a few neurons in the central nervous systemalso have pacemaker potentials and can spontaneouslygenerate action potentials in the absence of externalstimuli.
Nerves and HormonesThe contractile activity of smooth muscles is influ-enced by neurotransmitters released by autonomicnerve endings. Unlike skeletal muscle fibers, smoothmuscle fibers do not have a specialized motor end-plate region. As the axon of a postganglionic auto-nomic neuron enters the region of smooth musclefibers, it divides into many branches, each branch con-taining a series of swollen regions known as varicosi-ties. Each varicosity contains many vesicles filled withneurotransmitter, some of which are released when
CHAPTER NINE Muscle306
1. Spontaneous electrical activity in the plasma membrane ofthe muscle fiber
2. Neurotransmitters released by autonomic neurons
3. Hormones
4. Locally induced changes in the chemical composition(paracrine agents, acidity, oxygen, osmolarity, and ionconcentrations) of the extracellular fluid surrounding thefiber
5. Stretch
TABLE 9–5 Inputs Influencing Smooth MuscleContractile Activity
Actionpotential
Pacemakerpotential
Thresholdpotential
Time
0
+30
-60
Mem
bra
ne
po
ten
tial
(m
V)
FIGURE 9–39Generation of action potentials in a smooth muscle fiber
resulting from spontaneous depolarizations of the membrane
(pacemaker potentials).

www.mhhe.com/widmaier9 307
inositol trisphosphate, can cause the release of calciumfrom the sarcoplasmic reticulum, producing a contrac-tion without a change in membrane potential.
Local FactorsLocal factors, including paracrine agents, acidity, oxy-gen concentration, osmolarity, and the ion compositionof the extracellular fluid, can also alter smooth muscletension. Responses to local factors provide a means foraltering smooth muscle contraction in response tochanges in the muscle’s immediate internal environ-ment, which can lead to regulation that is independentof long-distance signals from nerves and hormones.
Many of these local factors induce smooth musclerelaxation. Nitric oxide (NO) is one of the most com-monly encountered paracrine agents that producessmooth muscle relaxation. NO is released from somenerve terminals as well as a variety of epithelial and en-dothelial cells. Because of the short life span of this re-active molecule, it acts as a paracrine agent, influencingonly those cells that are very near its release site.
Some smooth muscles can also respond by con-tracting when they are stretched. Stretching opensmechanosensitive ion channels, leading to membranedepolarization. The resulting contraction opposes theforces acting to stretch the muscle.
It is well to remember that seldom is a single agentacting on a smooth muscle, but rather the state of con-tractile activity at any moment depends on the simul-taneous magnitude of the signals promoting contractionversus those promoting relaxation.
Types of Smooth MuscleThe great diversity of the factors that can influence thecontractile activity of smooth muscles from variousorgans has made it difficult to classify smooth musclefibers. Many smooth muscles can be placed, however,
an action potential passes the varicosity. Varicositiesfrom a single axon may be located along several mus-cle fibers, and a single muscle fiber may be located near varicosities belonging to postganglionic fibers ofboth sympathetic and parasympathetic neurons (Figure9–40). Therefore, a number of smooth muscle fibers areinfluenced by the neurotransmitters released by a sin-gle nerve fiber, and a single smooth muscle fiber maybe influenced by neurotransmitters from more thanone neuron.
Whereas some neurotransmitters enhance contrac-tile activity, others produce a lessening of contractileactivity. Thus, in contrast to skeletal muscle, which re-ceives only excitatory input from its motor neurons,smooth muscle tension can be either increased or de-creased by neural activity.
Moreover, a given neurotransmitter may produceopposite effects in different smooth muscle tissues. Forexample, norepinephrine, the neurotransmitter releasedfrom most postganglionic sympathetic neurons, en-hances contraction of most vascular smooth muscle byacting on alpha-adrenergic receptors. By contrast, thesame neurotransmitter produces relaxation of airway(bronchiolar) smooth muscle by acting on beta-2 adren-ergic receptors. Thus, the type of response (excitatoryor inhibitory) depends not on the chemical messengerper se, but on the receptors to which the chemical mes-senger binds in the membrane and the intracellularsignaling mechanisms activated by those receptors.
In addition to receptors for neurotransmitters,smooth muscle plasma membranes contain receptorsfor a variety of hormones. Binding of a hormone to itsreceptor may lead to either increased or decreased con-tractile activity.
Although most changes in smooth muscle contrac-tile activity induced by chemical messengers are ac-companied by a change in membrane potential, this isnot always the case. Second messengers, for example,
FIGURE 9–40Innervation of smooth muscle by postganglionic autonomic neurons. Neurotransmitter is released from the varicosities along the
branched axons and diffuses to receptors on muscle fiber plasma membranes.
Smooth musclefiber
Axonvaricosities
Postganglionicsympatheticneuron
Postganglionicparasympatheticneuron

into one of two groups, based on the electrical charac-teristics of their plasma membrane: single-unit smoothmuscles and multiunit smooth muscles.
Single-Unit Smooth MuscleThe muscle fibers in a single-unit smooth muscleundergo synchronous activity, both electrical and mechanical; that is, the whole muscle responds tostimulation as a single unit. This occurs because eachmuscle fiber is linked to adjacent fibers by gap junc-tions, through which action potentials occurring inone cell are propagated to other cells by local cur-rents. Therefore, electrical activity occurring any-where within a group of single-unit smooth musclefibers can be conducted to all the other connected cells(Figure 9–41).
Some of the fibers in a single-unit muscle are pace-maker cells that spontaneously generate action poten-tials. These action potentials are conducted by way ofgap junctions to the rest of the fibers, most of which arenot capable of pacemaker activity.
The contractile activity of single-unit smooth mus-cles can be altered by nerves, hormones, and localfactors, using the variety of mechanisms described pre-viously for smooth muscles in general. The extent towhich these muscles are innervated varies considerablyin different organs. The nerve terminals are oftenrestricted to the regions of the muscle that contain pace-maker cells. The activity of the entire muscle can be con-trolled by regulating the frequency of the pacemakercells’ action potentials.
One additional characteristic of single-unit smoothmuscles is that a contractile response can often beinduced by stretching the muscle. In several holloworgans—the stomach, for example—stretching the
smooth muscles in the walls of the organ as a result ofincreases in the volume of material in the lumen initi-ates a contractile response.
The smooth muscles of the intestinal tract, uterus,and small-diameter blood vessels are examples ofsingle-unit smooth muscles.
Multiunit Smooth MuscleMultiunit smooth muscles have no or few gap junctions.Each fiber responds independently of its neighbors, andthe muscle behaves as multiple units. Multiunit smoothmuscles are richly innervated by branches of the auto-nomic nervous system. The contractile response of thewhole muscle depends on the number of muscle fibersthat are activated and on the frequency of nerve stimu-lation. Although stimulation of the nerve fibers to themuscle leads to some degree of depolarization and acontractile response, action potentials do not occur inmost multiunit smooth muscles. Circulating hormonescan increase or decrease contractile activity in multiunitsmooth muscle, but stretching does not induce contrac-tion in this type of muscle. The smooth muscle in thelarge airways to the lungs, in large arteries, and attachedto the hairs in the skin are examples of multiunit smoothmuscles.
It must be emphasized that most smooth musclesdo not show all the characteristics of either single-unitor multiunit smooth muscles. These two prototypes rep-resent the two extremes in smooth muscle characteris-tics, with many smooth muscles having characteristicsthat overlap the two groups.
Table 9–6 compares the properties of the differenttypes of muscle. Cardiac muscle has been included forcompleteness although its properties are discussed inChapter 12.
CHAPTER NINE Muscle308
Postganglionicsympatheticneuron Gap junctions
Postganglionicparasympatheticneuron
FIGURE 9–41Innervation of a single-unit smooth muscle is often restricted to only a few fibers in the muscle. Electrical activity is conducted from
fiber to fiber throughout the muscle by way of the gap junctions between the fibers.

www.mhhe.com/widmaier9 309
TABLE 9–6 Characteristics of Muscle Fibers
Smooth Muscle
CHARACTERISTIC SKELETAL MUSCLE SINGLE UNIT MULTIUNIT CARDIAC MUSCLE
Thick and thin filaments Yes Yes Yes Yes
Sarcomeres—banding pattern Yes No No Yes
Transverse tubules Yes No No Yes
Sarcoplasmic reticulum (SR)* ���� � � ��
Gap junctions between fibers No Yes Few Yes
Source of activating calcium SR SR and SR and SR andextracellular extracellular extracellular
Site of calcium regulation Troponin Myosin Myosin Troponin
Speed of contraction Fast-slow Very slow Very slow Slow
Spontaneous production of No Yes No Yes in certainaction potentials by fibers, but mostpacemakers not spontaneously
active
Tone (low levels of maintained No Yes No Notension in the absence ofexternal stimuli)
Effect of nerve stimulation Excitation Excitation or Excitation or Excitation orinhibition inhibition inhibition
Physiological effects of No Yes Yes Yeshormones on excitabilityand contraction
Stretch of fiber produces No Yes No Nocontraction
*Number of plus signs (�) indicates the relative amount of sarcoplasmic reticulum present in a given muscle type.
S E C T I O N B S U M M A R Y
StructureI. Smooth muscle fibers are spindle-shaped cells that
lack striations, have a single nucleus, and are capableof cell division. They contain actin and myosinfilaments and contract by a sliding-filamentmechanism.
Contraction and Its ControlI. An increase in cytosolic calcium leads to the binding
of calcium by calmodulin. The calcium-calmodulincomplex then binds to myosin light-chain kinase,activating the enzyme, which uses ATP tophosphorylate smooth muscle myosin. Onlyphosphorylated myosin can bind to actin andundergo cross-bridge cycling.
II. Smooth muscle myosin has a low rate of ATPsplitting, resulting in a much slower shorteningvelocity than is found in striated muscle. However,
the tension produced per unit cross-sectional area isequivalent to that of skeletal muscle.
III. Two sources of the cytosolic calcium ions that initiatesmooth muscle contraction are the sarcoplasmicreticulum and extracellular calcium. The opening ofcalcium channels in the smooth muscle plasmamembrane and sarcoplasmic reticulum, mediated bya variety of factors, allows calcium ions to enter thecytosol.
IV. The increase in cytosolic calcium resulting from moststimuli does not activate all the cross-bridges.Therefore smooth muscle tension can be increased byagents that increase the concentration of cytosoliccalcium ions.
V. Table 9–5 summarizes the types of stimuli that caninitiate smooth muscle contraction by opening orclosing calcium channels in the plasma membrane orsarcoplasmic reticulum.
VI. Most, but not all, smooth muscle cells can generateaction potentials in their plasma membrane uponmembrane depolarization. The rising phase of the

smooth muscle action potential is due to the influx ofcalcium ions into the cell through voltage-gatedcalcium channels.
VII. Some smooth muscles generate action potentialsspontaneously, in the absence of any external input,because of pacemaker potentials in the plasmamembrane that repeatedly depolarize the membraneto threshold.
VIII. Smooth muscle cells do not have a specialized end-plate region. A number of smooth muscle fibers may be influenced by neurotransmittersreleased from the varicosities on a single nerveending, and a single smooth muscle fiber may beinfluenced by neurotransmitters from more thanone neuron. Neurotransmitters may have eitherexcitatory or inhibitory effects on smooth musclecontraction.
IX. Smooth muscles can be classified broadly as single-unitor multiunit smooth muscle (Table 9–6).
CHAPTER NINE Muscle310
S E C T I O N B K EY T E R M S
dense body 00latch state 00multiunit smooth
muscle 00myosin light-chain
kinase 00
myosin light-chainphosphatase 00
pacemaker potential 00single-unit smooth
muscle 00smooth muscle tone 00varicosity 00
S E C T I O N B R E V I E W Q U E S T I O N S
1. How does the organization of thick and thinfilaments in smooth muscle fibers differ from that instriated muscle fibers?
2. Compare the mechanisms by which a rise in cytosoliccalcium concentration initiates contractile activity inskeletal and smooth muscle fibers.
3. What are the two sources of calcium that lead to theincrease in cytosolic calcium that triggers contractionin smooth muscle?
4. What types of stimuli can trigger a rise in cytosoliccalcium in smooth muscle fibers?
5. What effect does a pacemaker potential have on asmooth muscle cell?
6. In what ways does the neural control of smoothmuscle activity differ from that of skeletal muscle?
7. Describe how a stimulus may lead to the contractionof a smooth muscle cell without a change in theplasma membrane potential.
8. Describe the differences between single-unit andmultiunit smooth muscles.
C H A P T E R 9 T H O U G H T Q U E S T I O N S
(Answers are given in appendix A.)1. Which of the following corresponds to the state of
myosin (M) under resting conditions and in rigormortis? (a) M � ATP, (b) M � ADP � Pi, (c) A � M �ADP � Pi, (d) A � M.
2. If the transverse tubules of a skeletal muscle aredisconnected from the plasma membrane, will actionpotentials trigger a contraction? Give reasons.
3. When a small load is attached to a skeletal musclethat is then tetanically stimulated, the muscle lifts theload in an isotonic contraction over a certain distance,but then stops shortening and enters a state ofisometric contraction. With a heavier load, thedistance shortened before entering an isometriccontraction is shorter. Explain these shortening limitsin terms of the length-tension relation of muscle.
4. What conditions will produce the maximum tensionin a skeletal muscle fiber?
5. A skeletal muscle can often maintain a moderate levelof active tension for long periods of time, eventhough many of its fibers become fatigued. Explain.
6. If the blood flow to a skeletal muscle were markedlydecreased, which types of motor units would mostrapidly have their ability to produce ATP for musclecontraction severely reduced? Why?
7. As a result of an automobile accident, 50 percent ofthe muscle fibers in the biceps muscle of a patientwere destroyed. Ten months later, the biceps musclewas able to generate 80 percent of its original force.Describe the changes that took place in the damagedmuscle that enabled it to recover.
8. In the laboratory, if an isolated skeletal muscle isplaced in a solution that contains no calcium ions,will the muscle contract when it is stimulated(1) directly by depolarizing its membrane, or (2) bystimulating the nerve to the muscle? What wouldhappen if it were a smooth muscle?
9. The following experiments were performed on asingle-unit smooth muscle in the gastrointestinal tract.a. Stimulating the parasympathetic nerves to the
muscle produced a contraction.b. Applying a drug that blocks the voltage-sensitive
sodium channels in most plasma membranes ledto a failure to contract upon stimulating theparasympathetic nerves.
c. Applying a drug that binds to muscarinic receptors(Chapter 6), and hence blocks the action of ACh atthese receptors, did not prevent the muscle fromcontracting when the parasympathetic nerve wasstimulated.
From these observations, what might one concludeabout the mechanism by which parasympatheticnerve stimulation produces a contraction of thesmooth muscle?