modifications in low-density lipoprotein receptor expression affects cyclosporin a cellular uptake...
TRANSCRIPT

Modifications in Low-Density Lipoprotein ReceptorExpression Affects Cyclosporin A Cellular Uptakeand Cytotoxicity
CARLOS LEON,1 JESSICA JIA,1 GUOSONG QIU,2 JOHN S. HILL,2 KISHOR M. WASAN1
1Division of Pharmaceutics and Biopharmaceutics, Faculty of Pharmaceutical Sciences,The University of British Columbia, Vancouver British Columbia, Canada V6T 1Z3
2Atherosclerosis Specialty Laboratory, Healthy Heart Program, James Hogg iCAPTURE Centre for Cardiovascularand Pulmonary Research, Department of Pathology and Laboratory Medicine, St. Paul’s Hospital,University of British Columbia, Vancouver British Columbia, Canada
Received 16 April 2007; revised 20 June 2007; accepted 26 June 2007
Published online in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/jps.21141
Carlos Leonwork.
Abbreviationnium persulfateCE, cholesterylporine A; Dil-LDtein; DMEM, Ddouble-strandedradish peroxidadensity lipoproted with LDLr-slipoprotein recedimethylthiazolphenyl)-2H-tetr
2350 JOURN
ABSTRACT: The purpose of this study was to test the effect of modulating the expres-sion of the human low-density lipoprotein receptor (LDLr) in human embryonic kidney(293T) cells on Cyclosporin A (CsA) cellular uptake and CsA-mediated cytotoxicity. LDLrexpression was modulated using RNA interference (RNAi) and an LDLr overexpressionplasmid. One of the small-interfering RNA (siRNA) constructs, LDLr-792, showed a 60%decrease in LDLr protein expression. The downregulation effect was specific as trans-fection with an annexin V (AxV) siRNA construct did not decrease LDLr expressionlevels. AxV and ABCA1 expression levels were not affected in the cells transfected withLDLr-792 (LDLrLOW cells) compared to the controls. At a functional level, fluorescentlow-density lipoprotein (LDL) (DiI-LDL) internalization in the LDLrLOW cells wasdecreased (30%) compared to control cells. We tested the dose-dependent cytotoxicityinduced by CsA using a respiration assay. We found a decrease in CsA-mediatedcytotoxicity in the range of CsA doses studied (1–10 mg/mL) in the LDLrLOW cellscompared to the pSHAG-transfected cells, reaching a statistical significance at 10mg/mLCsA. At higher CsA doses we found a significant decrease in LDLr expression. When thecontrol and LDLrLOW cells were treated with another cytotoxic drug, gentamycin, therewas no difference in the cell viability, suggesting that this effect is specific for CsA. Weconfirmed the association of LDLr expression levels with CsA uptake by overexpressingthe LDLr. The LDLr overexpressing cells showed an enhanced uptake of radiolabelledCsA. Taken together these results suggest that CsA internalization and cytotoxicity areaffected by the LDL receptor expression levels. � 2007 Wiley-Liss, Inc. and the American
Pharmacists Association J Pharm Sci 97:2350–2361, 2008
proximal tubule; RISC, RNA-induced silencing complex; RNAi,RNA interference; RT-PCR, reverse transcriptase PCR; SDS,sodium dodecyl sulfate; shRNA, short hairpin RNA; TCR,T-cell receptor; TEMED, N,N, N,N-tetra-methyl-ethelenedia-mine; TG, triglycerides; VLDL, very low-density lipoprotein.
Correspondence to: Kishor M. Wasan (Telephone: þ1-604-822-4889; fax: þ1-604-822-3035;E-mail: [email protected])
Journal of Pharmaceutical Sciences, Vol. 97, 2350–2361 (2008)
� 2007 Wiley-Liss, Inc. and the American Pharmacists Association
and Jessica Jia contributed equally to this
s: ANOVA, analysis of variance; APS, ammo-; AxV, Annexin V; BSA, bovine serum albumin;esters; CO, cholesterol oxidases; CsA, Cyclos-L, fluorescently labeled low density lipopro-ulbecco Modified Eagle’s medium; dsRNA,RNA; FBS, fetal bovine serum; HRPO, horse-
se; LDL, low-density lipoprotein; LDLr, low-ein receptor; LDLrLOW cells, cells transfect-iRNA which express low levels of low-densityptor; mAb, monoclonal antibody; MTS, 3-(4,5--2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfo-azolium; PCR, polymerase chain reaction; PT,
AL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008

LDL RECEPTOR LEVELS AFFECT CYCLOSPORIN A-MEDIATED CYTOTOXICITY 2351
Keywords: renal proximal tubule cell
DOI 10.1002/jps
line; LDL receptors; RT-PCR analysis; DiI-LDL; Western blot analysis; RNA interference
INTRODUCTION
The low-density lipoprotein receptor (LDLr) isan endocytic receptor that transports relevantmacromolecules, mainly the cholesterol-rich low-density lipoprotein (LDL), into cells through aprocess called receptor-mediated endocytosis.1,2
This process involves the cell surface receptorrecognizing an LDL particle from the extracel-lular membrane (ECM), internalizing it throughclathrin-coated pits and transporting it intracel-lularly via a vesicle.2–4 Subsequently, the vesiclebecomes degraded upon fusion with the lysosome,releasing lipids into the cytoplasm for cell use;meanwhile the receptor recycles back to the cellsurface to bind to another LDL particle.5,6 Much ofour current knowledge of receptor-mediatedendocytosis originated from the pioneering stud-ies on the LDL receptor pathway conducted byGoldstein and Brown.2,5 Their work involvedcharacterizing the biochemistry and genetics ofthe LDL receptor and determining its importantrole in maintaining intracellular cholesterolhomeostasis. Inherited mutations within thisreceptor have been linked to familial hypercho-lesterolemia, a genetic disease that results inpathologically elevated blood cholesterol andpremature coronary disease.5,7,8
Lipoproteins are biological carriers in whichlipids and proteins can be transported systemi-cally. It is well established that specific drugs suchas halofantrine, amphotericin B, and Cyclospor-ine A associate with lipoproteins.9–12 There havebeen a number of studies suggesting that the LDLreceptor and members of its super-family maybe playing a role in cellular drug uptake,specifically, aminoglycosides, type-I ribosome-inactivating proteins (RIP), anionic liposomes,and Cyclosporine A.11–13 Therefore, it would be ofinterest to examine the role of the LDL receptorfamily as a mechanism of cellular drug uptake.One of the most widely used methods to decreasegene expression is RNA interference (RNAi).Because that degradation of mRNA therebyshuts-down (quells) production of the correspond-ing protein, RNAi constitutes a pathway that cellsutilize to regulate/silence gene expression. Fol-lowing the introduction of double-stranded RNA(dsRNA) into the target cells RNAi mediatessequence-specific posttranscriptional gene silen-
J
cing. RNAi as an experimental strategy is basedupon an ancient, natural regulatory mechanismwidely distributed in nature-including plants,Caenorhabditis elegans, Drosophila, and mam-malian cells-designed to protect cells from viralinfection. RNAi is initiated when dsRNA isprocessed into short (<30 bp) dsRNA duplexescalled small-interfering RNA (siRNA) by dicer, ahost ribonuclease. These short dsRNA duplexesare then incorporated into a multicomponentnuclease complex referred to as the RNA-inducedsilencing complex (RISC). Based upon the specificsiRNA sequence, RISC targets cognate mRNAs fordegradation.14 siRNA has been shown to success-fully silence both endogenous and exogenousgenes in 293T cells.15,16
Cyclosporin A (CsA) is an effective immuno-suppressant used to treat patients who haveundergone organ transplantation as well as totreat certain autoimmune diseases such aspsoriasis and rheumatoid arthritis.17 However,CsA’s use has been limited by its adverse effectsincluding nephrotoxicity, hepatotoxicity, neuro-toxicity, hypertension, and dyslipidemia.18,19
CsA-induced renal toxicity is the most importantside effect which often results in discontinuationof CsA therapy.20,21 Renal toxicity is characterizedby a rise in serum creatinine levels and a decreasein the glomerular filtration rate22 and is oftenreversible following the discontinuation of lowCsA doses (<5 mg/kg/day). The majority ofpatients who have undergone kidney transplantsexperience a significant increase in total serumcholesterol levels.19,22,23 Yet the mechanisms stillremain unclear.
The objectives of our study were to modulate theLDLr expression using small interference RNAand overexpression in 293T cells and to investigatethe effect of decreased LDLr expression on CsAinternalization and CsA-induced cytotoxicity.We decreased LDLr expression using siRNAas shown by immunoblotting and RT-PCR andfunctionally by a decrease on the internalization offluorescent LDL. Cyclosporine-mediated cytotoxi-city in pSHAG and LDLr-792 transfected cellssuggest that a reduction in LDLr expression affectsCsA-mediated nephrotoxicity at physiologicaldoses. This effect was specific as transfectantstreated with gentamycin did not differ in theirviability. LDLr overexpression was associated with
OURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008

Table 1. Sequence of Inserts in pSHAG Constructsfor LDLr, AxV, and a Scrambled Sequence
LDLr-792:
2352 LEON ET AL.
an increased uptake of radiolabelled CsA. CsAinternalization and cytotoxicity may be partiallymediated by LDLr-dependent uptake.
50-TCACATTAACGCAGCCAACTTCATCGCT-GAAGCTTGAGCGGTGGAGTTGGCTGCGTTAGTGT-GATACTTTTTT-30
LDLr-973:50-ATCTTAAGGTCATTGCAGACGTGGGAAC-GAAGCTTGGTTCTCATGTCTGTAATGATCTTAA-GATCGG TTTTTT-30
AV2.1:50-TCCCCAGATGTATCTCCCTTAATCATGG-GAAGCTTGCTATGATTAAGGGGGATACGTCTGGG-GATTATTTTTT-30
pSHAG scrambled:50-TGTTCATATGTCTGCTCTTGTAGCAGTA-GAAGCTTGTACTGCTGCAGGAGCAGACGTATGAG-CACGTTTTTTT-30
The number (792 and 973) corresponds to the region in thecDNA sequence of the LDLr where the first oligonucleotidefrom the target sequence is located.
EXPERIMENTAL
Materials
Antibodies to annexin V (AxV) and actin werepurchased from Santa Cruz Biotechnology (SanFrancisco, CA). Antibodies to LDLr were boughtfrom Research Diagnostics (Flanders, NJ), whileantibodies to SR-BI and ABCA1 were purchasedfrom Novus Biologicals (Littleton, CO). Dulbeccomodified Eagle’s medium (DMEM), Hanks balan-ced salt solution (HBSS), L-glutamine, HEPES,penicillin and streptomycin, fetal bovine serum,Lipofectamine 2000, and minimal essential ami-noacids were bought from Invitrogen, Carlsbad,CA; PBS, polylysine, protease inhibitors mixtureand phenyl methyl sulfonyl fluoride (PMSF) wereobtained from Sigma Chemicals Co. (St Louis,MO). Secondary antimouse and antirabbit anti-bodies conjugates to HRPO were bought fromBIO-RAD (San Francisco, CA).
Cell Culture
Human embryonic kidney 293T cells wereobtained from the American Type Culture Collec-tion (Rockville, MD). 293T cells were grown inDMEM supplemented with 10% fetal calf serum,0.1 mM minimal nonessential amino acids, 10 mML-glutamine, 20 mM HEPES, and 100mg/mL eachof penicillin and streptomycin (Invitrogen).
RNA Interference (RNAi)
Modulation of LDL receptor expression in 293Tcells using siRNA: siRNA constructs were desig-ned using a web-based platform (http://katah-din.cshl.org). The regions chosen for targetingwere specific to either LDLr or AxV,24 the latestused as a specificity control, as determined by ahomology search using the BLAST engine. Thesequences used to create the pSHAG constructsare summarized in Table 1.
The inserts were cloned into the Bse RI/Bam HIcloning site of the pSHAG vector that directs thein vivo synthesis of short hairpin RNA (shRNA)molecules using a U6 promoter.15,25 Positiveclones were selected by the acquisition of bothkanamycin resistance and a new Hind III site.
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008
Clones containing the inserts of interest weregrown, plasmid was purified using an endotoxinfree Maxiprep kit (Qiagen, Mississauga, Canada)and used to transfect 293T cells. As a positivecontrol for the siRNA effect, we used an AxVconstruct (AV2.1) cloned in the pSHAG vector.24
It also served as a negative control for unspecificsiRNA effects.
siRNA transfection of 293T cells: a 6-well platewas coated with a 0.1% solution of polylysine(Sigma) to enhance the adherence of the cellswhich were seeded the day before of the transfec-tion. The cells were at 95% confluency the day ofthe transfection. The amount of DNA per well (6-well plate) was 4.0 mg diluted in 250 mLof OPTIMEM. The volume of Lipofectamine2000 per well was 10 mL in 250 mL. Briefly,4 mg of pSHAG, AV2.1, 792, or 973 plasmids wereindependently added to 500 mL of OPTIMEMmedium. One hundred microliters of Lipofecta-mine 2000 were incubated with 3 mL of OPTI-MEM. After vortexing and incubating for 5 min,600 mL of the Lipofectamine/OPTIMEM mixturewas added to the DNA/OPTIMEM solution. Thenew solution was incubated for 20 min at roomtemperature. After this period, the DNA/Lipofec-tamine/OPTIMEM mixture was added (500 mL) tothe cells. The cells were washed once with HBSSand then 1 mL of complete medium was added 3 hprior to adding the medium/DNA/Lipofectaminemix for an overnight incubation. The day after thetransfection, the cells were washed and newcomplete medium was added. The cells were left
DOI 10.1002/jps

LDL RECEPTOR LEVELS AFFECT CYCLOSPORIN A-MEDIATED CYTOTOXICITY 2353
in the plates for 3 days. After this period, the cellswere washed with HBSS, any extra remainingliquid was removed and the plates were frozenuntil the proteins were extracted, quantified andanalyzed by SDS–PAGE and immunoblotting.
Western Blot of LDL Receptor, Annexin V (AxV),ABCA1, and Actin
We used two different antibodies to detect the LDLreceptor: an antiLDLr mouse monoclonal antibody(Research Diagnostics RDI-PRO61087) and arabbit polyclonal antibody (Research DiagnosticsRDI-PRO61099). The rabbit antibody presented ahigher sensitivity (in agreement with the manu-facturer information) and was mostly used in thisstudy. Briefly, the cells were washed, lysed in RIPAbuffer, and solubilized protein was quantifiedusing the BIO-RAD protein determination assay.LDL receptor, AxV, ABCA1, and actin antibodieswere used at 1:400, 1:4000, 1:1000, and 1:5000dilutions, respectively. Amounts of 20–50 mg ofprotein lysates were resolved in SDS–PAGE in amini Protean or regular format gel electrophoresissystem (both from BIO-RAD). The protein samplesseparated by SDS–PAGE were transferred tonitrocellulose membranes (BIO-RAD) and blockedwith either 3% bovine serum albumin (SigmaChemical Co.) or 5% milk in 1X TBS with 0.1%Tween 20 for 1–2 h and then probed with theindicated primary antibody diluted in blockingsolution for 2 h or as otherwise specified. Mem-branes were washed three times, incubatedwith the appropriate secondary antibody coupledto peroxidase and proteins were detected byenhanced chemiluminescence (GE Healthcare,UK).
RT-PCR Analysis
The LDLr gene expression was assessed by RT-PCR. Briefly, the cells were seeded in 6-well platesat 50% confluency. After the different treatmentsthey were harvested and total RNA isolated withTRIzol1 Reagent (Invitrogen). cDNA was synthe-sized using 8 mg of RNA. The concentration ofcDNA reaction product was measured by usingPicogreen-Assay (molecular probes). The primerswere designed and purchased from Invitrogen.Parameters and conditions for the tested primerswere individually optimized. Primers (50–80 ng)were subjected to PCR for detection of LDLr aswell as the housekeeping gene small ribosomal
DOI 10.1002/jps J
subunit 18S. A sample (15 mL) from each PCRproduct was subjected to electrophoresis on a 1.5%agarose gel (containing ethidium bromide). A100 bp ladder was used to identify the size of PCRproducts. The fluorescent bands were imagedunder UV light and quantified.
DiI-LDL Studies
Procedures were adapted from5,26 and modified tosuit conditions for 293T cells. Briefly, 48 hpostseeding, cells were confluent with LDLreceptors being upregulated. Cells were washedand 500 mL of DMEM medium supplemented withdelipidated serum and BSA with or without 15 mLof DiI-LDL (1 mg/mL) were added to the cells whichwere then incubated for 15 min at 48C protectedfrom direct light. Fluorescence was then deter-mined by removing media and washing thor-oughly the cells with three ice-cold PBS washings,the cells were lysed using RIPA buffer supple-mented with PMSF and other protease inhibitors.Protein was quantified using the BIO-RADprotein determination kit with BSA as a standard.Fluorescence was determined at excitation andemission wavelengths, 522 and 578 nm, respec-tively, using the Cytofluor B. Multi-well plateReader Series 4000 from PerSeptive Biosystems.
Cyclosporin A and Gentamycin Cytotoxicity Studies
293T cells were seeded in 96-well plates precoat-ed with poly-L-lysine at a density of 20000–30000 cells/well. The next day they were washedwith HBSS and incubated with 200 mL of serialdilutions of Cyclosporin A, ranging from 1 to 10 mg/mL, or Gentamycin, ranging from 10 to 100 mg/mL.Both drugs were purchased from Sigma ChemicalCo. The cells were incubated with either drug for24 h at 378C and then the cytotoxicity was analyzedby MTS [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxy-methoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium]using a commercial kit (Promega, Madison, WI).Percent viability was determined as follows: readingof sample divided up the reading of the controltransfected cells (no Cyclosporin A added). Proteinconcentration was calculated using a proteindetermination assay.
Upregulation of the LDLr Expression andRadioactive Cyclosporin A Incorporation Studies
A human cDNA clone expressing the LDLr(OriGene Technologies, Rockville, MD) was used
OURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008

2354 LEON ET AL.
to transfect 293T cells. Briefly, 293T cells were setat 50% confluency on a six well plate. Thefollowing day the cells were washed and left torest for an hour in complete DMEM medium. TheDNA was incubated with the Lipofectamine 2000as found in the RNA Interference section. Theday, the cells were washed with HBSS and werelysed RIPA buffer to determine LDLr expressionby immunoblot. For the CsA incorporation stu-dies, on the day following the transfection, thecells were washed and the wells were replenishedwith incomplete DMEM (no FCS). After 2 h, thecells were incubated with 500 mL of completeDMEM with 0.1% BSA and 0.1 nM of radioactiveCsA (Amersham Pharmacia, stock of CsA 1 mCi/mL, 100 nM of drug). After 2 h of incubation, thesupernatants were recovered, the cells werewashed with HBSS twice and then lysed for30 min on ice using 500 mL of RIPA buffer plusprotease inhibitors. After this incubation,the lysates were cleared by centrifugation at9000 RPM for 3 min; the supernatants wereanalyzed for radioactive incorporation and also forLDLr protein expression by immunoblotting.
Statistics
Normalized protein expression were comparedbetween treatment groups by an unpaired t-test(INSTAT; GraphPad). Critical differences wereassessed by Tukey post hoc tests. A difference wasconsidered significant if the probability of chanceexplaining the results was reduced to less than 5%(p< 0.05). Data are expressed as mean� standarddeviation.
RESULTS
Construction and Validation of siRNATargeting the LDL Receptor
Constructs LDLr-792 and LDLr-973 were clonedinto the pSHAG plasmid and used to transformcompetent Escherichia coli cells. The isolatedclones were regrown and analyzed by plasmidminiprep and digestion with Hind III. Thelinearization of the clones that is, the acquisitionof the new Hind III site, suggested that theycontained an insert. The presence of the insert andthe lack of point mutations were further con-firmed by DNA sequencing (data not shown).These plasmids were used to transiently transfect293T cells as outlined before and gene expression
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008
was analyzed by RT-PCR. One of the LDLrconstructs, LDLr-973, showed only a minor effect(Fig. 1A; 20–30% reduction). The second constructLDLr-792 showed a 60% decrease in LDL receptorexpression.
The RT-PCR results were further confirmed byimmunoblot (Fig. 1B). In three experiments, theAxV construct downregulated AxV proteinexpression by 70%, as expected. The AxV down-regulation led to a significant increase in LDLrexpression (190%), both at the level of protein andmessenger (Fig. 1C). This confirms that thedownregulation of the LDLr expression is not afunction of an unspecific RNAi effect. As anadditional control of specificity, we tested theexpression of AxV and the ABC transporterABCA1 by immunoblot. AxV and ABCA1 expres-sion were not significantly different betweenthe pSHAG and LDLr-792-transfected cells(Fig. 2A and B).
We tested the downregulation of the LDLr at afunctional level by using a fluorescent LDL (DiI-LDL) to quantify the LDL internalization incontrol and LDLrLOW cells (Fig. 3). We found asignificant 30% decrease in DiI-LDL incorpora-tion in the LDLrLOW cells compared to the emptyvector-transfected cells. Our results suggest theinduction of compensatory mechanisms of LDLinternalization in the LDLrLOW cells.
Effect of LDLr Downregulation in CsA-MediatedCytotoxicity
We examined the dose-dependent cytotoxicityinduced by Cyclosporin A in 293T cells as assessedby an MTS assay. Between 1 and 10 mg/mL ofCsA there was a dose-dependent increase incytotoxicity (Fig. 4A). Interestingly, at doses 5and 10 mg/mL of CsA we did not find any furtherincrease in cytotoxicity.
Once we established that our 293T cell modelexpressed the LDLr, was responsive to siRNAdownregulation and that the cell viability wasaffected by CsA treatment, we addressed the effectof LDLr downregulation in CsA-mediated cyto-toxicity. Interestingly we found that CsA treat-ment decreased LDLr expression at doses of2.5 mg/mL and higher (Fig. 4B). In terms ofCsA-cytotoxicity there was a higher cell viabilityin LDLrLOW cells compared to pSHAG-transfectedcells (Fig. 4C) at a dose of 10 mg/mL of CsA. WhenpSHAG-transfected cells and untransfected 293Tcells viability in response to CsA was compared,
DOI 10.1002/jps

Figure 1. (A) Analysis of LDLr gene expression in 293T cells transfected with pSHAGconstructs. RT-PCR analysis of pSHAG, and pSHAG-LDL clones. Cells transfected withthe pSHAG constructs were washed and lysed. RNA was extracted and cDNA wasprepared. LDLr and 18 S ribosomal subunit were amplified using specific primers. Datapresented as mean�SD n¼ 3–4. �p< 0.05 with pSHAG. (B) Analysis of LDLr proteinexpression in 293T cells transfected with pSHAG constructs. Cells were transfected aspreviously described. Twenty-four hours posttransfection, the cells were washed and left48 h in complete medium. The cells were solubilized in modified RIPA buffer in thepresence of protease inhibitors. Lysates were separated by SDS–PAGE and proteinswere transferred to nitrocellulose for immunoblotting with a rabbit antiLDL receptor,goat antiactin, and rabbit antiAxV (not shown) antibodies (n¼ 4). (C) Densitometricanalysis of LDLr protein expression normalized using actin levels. �p< 0.05 withpSHAG.
LDL RECEPTOR LEVELS AFFECT CYCLOSPORIN A-MEDIATED CYTOTOXICITY 2355
we did not find a statistical difference betweenthese two groups (data not shown), indicating thatthe transfection with pSHAG itself is not respon-sible for the changes in cell viability. As anadditional control we examined the effect ofsiRNA production on CsA-mediated cytotoxicity.We used a scrambled siRNA sequence cloned inthe pSHAG vector by the same procedure that
DOI 10.1002/jps J
lead to the generation of the LDLr siRNA clones.We did not find any difference between theviability of cells transfected with pSHAG andpSHAG scrambled siRNA constructs treated withvarious doses of CsA (data not shown), suggestingthat the production of siRNA per se does not affectthe viability of the cells in response to differentdoses of CsA.
OURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008

Figure 2. (A) Analysis of the specificity of LDLr downregulation in 293T cells. AxVprotein expression is not affected by LDLr downregulation. Actin protein expression isshown as a control of protein loading. Data is representative of one of three differentexperiments. (B) Densitometric analysis of AxV expression in 293T transfected cells (A).(C) Analysis of the specificity of the LDLr downregulation in 293T cells. ABCA1 proteinexpression is not affected by LDLr downregulation. Actin protein expression is shown asa control of protein loading. Data is representative of one of three different experiments.(D) Densitometric analysis of ABCA1 protein expression normalized using actin levels.Mean and SD of three independent experiments.
2356 LEON ET AL.
Effect of LDLr Upregulation in CsA Uptake
The transfection with the LDLr overexpressionplasmid led to a marked increase in LDLr expres-sion as shown by immunoblotting (Fig. 5A). Theincrease in LDLr expression was also associatedwith a marked increase in DiI-LDL incorporation(data not shown). When the overexpressing cellswere incubated with radioactive CsA, these cells
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008
showed a 75% increase in CsA uptake compared tocontrols (Fig. 5B).
DISCUSSION
Currently there is no established method todownregulate the expression of the LDLr recep-tor. The studies on the LDL receptor include the
DOI 10.1002/jps

Figure 3. Analysis of the incorporation of fluorescent LDL (DiI-LDL) in pSHAG andLDLr-792 transfected 293T cells. The cells were incubated with DiI-LDL for 15 min at48C and then washed, lysed in RIPA buffer and fluorescence was measured using theCytofluor B fluorometer. The fluorescence was corrected by the protein concentrationand normalized in respect to the pSHAG transfected cells values. Mean and SD of threeexperiments. �p< 0.05 with pSHAG.
LDL RECEPTOR LEVELS AFFECT CYCLOSPORIN A-MEDIATED CYTOTOXICITY 2357
use of specific blocking antibodies which have aslimitation their short half life and the high costinvolved. Moreover, the incubation of humanmacrophages with antigen antibody complexesprepared with rabbit antiLDL and human LDL(LDL-IC) is followed by ingestion of those immunecomplexes (IC), massive cholesterol ester accu-mulation, cytokine release, and overexpression ofthe LDL receptor.27 There is a report on the use ofanother technique, a vector directed antisenseRNA28 that showed an 85% decrease in LDLrexpression in hepatocytes. However, this plasmidrequired the presence of a drug, G418, which mayinduce some other changes that could complicatethe interpretation of the results. Another limita-tion is the possibility of drug interactions betweenG418, the selective antibiotic, and Cyclosporin A.The lack of a methodology to downregulate theLDLr expression prompted us to design a siRNAstrategy to target this receptor.
Our laboratory has investigated renal cellcholesterol uptake and regulation and themechanisms of drug-induced cytotoxicity usingLLC-PK1 cells, a pig proximal tubule cell line.29 Inaddition, these cells have been established in vitroas a kidney model to investigate drug-inducedcytotoxicity.20,21,30,31 However, one of the disad-vantages of this model is its origin, so there is alimit on the inferences that we can make to ahuman model. Thus, we decided to use 293T cellsas model since we demonstrated that they expressthe LDL receptor and have been shown to be a
DOI 10.1002/jps J
useful model for siRNA downregulation. To ourknowledge this is the first report of the use ofRNAi targetting LDLr expression in any cellmodel.
In our initial studies we used two constructstargeting the LDLr; only one of these (LDLr-792)proved successful. There is a great degree ofvariability on the efficiency of siRNA constructsand the tools to select effective target sequencesare still under development.32 One possible issuewould be the transfection efficiency. Undersimilar experimental conditions (pSHAG systemand 293T cells), we have found a 95% efficiency oftransfection using a b-galactosidase reporter gene(data not shown). Delivery of siRNAs into mam-malian cells by transfection of siRNA or DNAvectors expressing shRNA has been shown tomediate RNAi successfully.15,33,34 siRNA trans-fection is transient lasting only for a week or so,14
although DNA-based vectors may last longer withdrug selection. As an additional advantage ofthe system that we have used in our study is thepossibility of transferring the hairpin from thepSHAG construct to a lentiviral vector.16 Viralvectors have also been used to deliver siRNAsuccessfully, and these methods tend to providemore stable gene silencing.25,35
Our results show that the downregulation of theLDLr protein expression is associated withreduced transcript levels, indicating that thesiRNA effect is at the messenger level and notan unspecific protein degradation which can be
OURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008
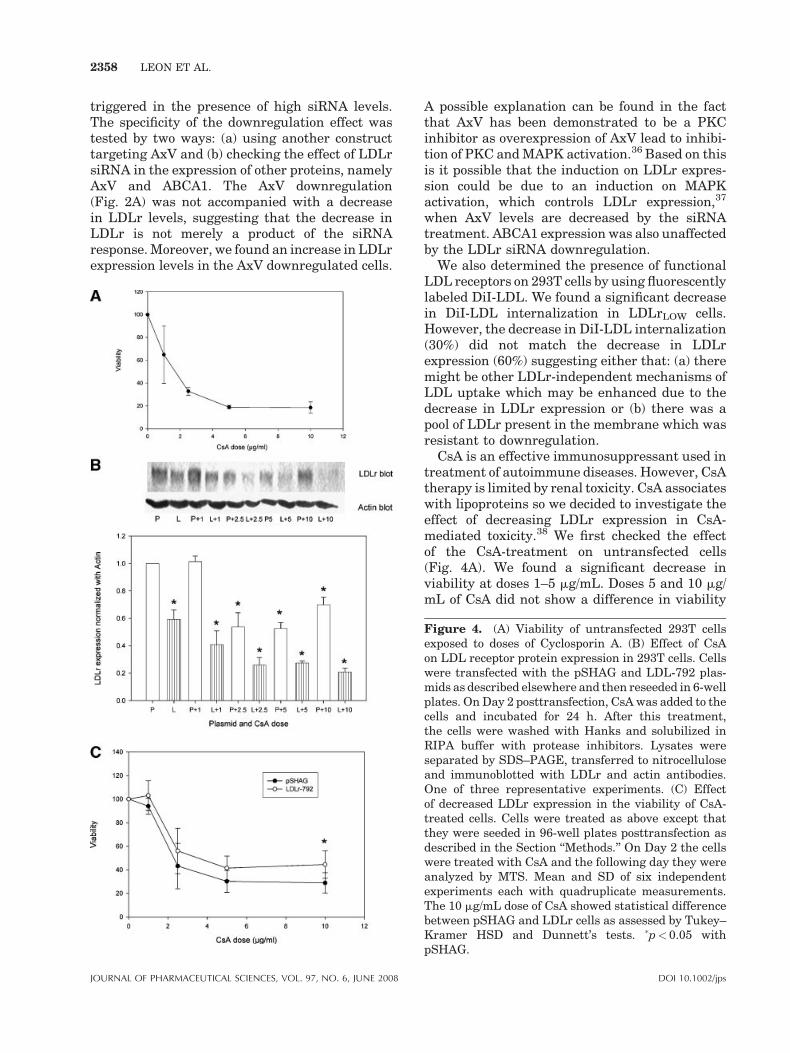
2358 LEON ET AL.
triggered in the presence of high siRNA levels.The specificity of the downregulation effect wastested by two ways: (a) using another constructtargeting AxV and (b) checking the effect of LDLrsiRNA in the expression of other proteins, namelyAxV and ABCA1. The AxV downregulation(Fig. 2A) was not accompanied with a decreasein LDLr levels, suggesting that the decrease inLDLr is not merely a product of the siRNAresponse. Moreover, we found an increase in LDLrexpression levels in the AxV downregulated cells.
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008
A possible explanation can be found in the factthat AxV has been demonstrated to be a PKCinhibitor as overexpression of AxV lead to inhibi-tion of PKC and MAPK activation.36 Based on thisis it possible that the induction on LDLr expres-sion could be due to an induction on MAPKactivation, which controls LDLr expression,37
when AxV levels are decreased by the siRNAtreatment. ABCA1 expression was also unaffectedby the LDLr siRNA downregulation.
We also determined the presence of functionalLDL receptors on 293T cells by using fluorescentlylabeled DiI-LDL. We found a significant decreasein DiI-LDL internalization in LDLrLOW cells.However, the decrease in DiI-LDL internalization(30%) did not match the decrease in LDLrexpression (60%) suggesting either that: (a) theremight be other LDLr-independent mechanisms ofLDL uptake which may be enhanced due to thedecrease in LDLr expression or (b) there was apool of LDLr present in the membrane which wasresistant to downregulation.
CsA is an effective immunosuppressant used intreatment of autoimmune diseases. However, CsAtherapy is limited by renal toxicity. CsA associateswith lipoproteins so we decided to investigate theeffect of decreasing LDLr expression in CsA-mediated toxicity.38 We first checked the effectof the CsA-treatment on untransfected cells(Fig. 4A). We found a significant decrease inviability at doses 1–5 mg/mL. Doses 5 and 10 mg/mL of CsA did not show a difference in viability
Figure 4. (A) Viability of untransfected 293T cellsexposed to doses of Cyclosporin A. (B) Effect of CsAon LDL receptor protein expression in 293T cells. Cellswere transfected with the pSHAG and LDL-792 plas-mids as described elsewhere and then reseeded in 6-wellplates. On Day 2 posttransfection, CsA was added to thecells and incubated for 24 h. After this treatment,the cells were washed with Hanks and solubilized inRIPA buffer with protease inhibitors. Lysates wereseparated by SDS–PAGE, transferred to nitrocelluloseand immunoblotted with LDLr and actin antibodies.One of three representative experiments. (C) Effectof decreased LDLr expression in the viability of CsA-treated cells. Cells were treated as above except thatthey were seeded in 96-well plates posttransfection asdescribed in the Section ‘‘Methods.’’ On Day 2 the cellswere treated with CsA and the following day they wereanalyzed by MTS. Mean and SD of six independentexperiments each with quadruplicate measurements.The 10 mg/mL dose of CsA showed statistical differencebetween pSHAG and LDLr cells as assessed by Tukey–Kramer HSD and Dunnett’s tests. �p< 0.05 withpSHAG.
DOI 10.1002/jps

Figure 5. (A) Overexpression of the LDLr in 293T cells. Cells were transfected with apCMV6-LDLr or b-galactosidase pCMV plasmids for 24 h. Cells were solubilized withRIPA buffer and lysates were cleared by centrifugation (11000 RPM for 20 min at 48C).Supernatants were used for protein determination, SDS–PAGE, and immunoblotting.Result shown is one representative of four independent experiments. (B) RadioactiveCsA uptake in cells that overexpress the LDLr. Cells were transfected with the LDLroverexpressing plasmid for 24 h. The cells were washed and rested in incomplete DMEMfor 2 h. Then they were incubated with radioactive CsA in complete medium as found inthe Experimental section. After the 2-h incubation, they were washed three times withPBS and then cells were lysed in RIPA buffer. Cleared lysates were subjected to proteindetermination and analyzed for radioactivity. Results shown are mean�SD of sixindependent transfections (�p¼ 0.0193). The DPM counts were normalized by proteincontent and then by the values obtained for the control cells in each experiment.
LDL RECEPTOR LEVELS AFFECT CYCLOSPORIN A-MEDIATED CYTOTOXICITY 2359
suggesting that either: (a) there is a population ofCsA-resistant 293T cells accounting for thisplateau in cytotoxicity or (b) that a reduction inLDLr expression at these doses of CsA precludeany further cell death.39 determined in a clinicalstudy that the maximum CsA plasma concentra-tions were around 2 mg/mL which are inline withthe CsA dose used in our present work.
We then evaluated the effects of CsA on LDLrexpression and cell viability in pSHAG-trans-fected cells. We found that at a dose of 1 mg/mL ofCsA there was no effect on LDLr protein expres-sion (Fig. 4B). However, higher doses of CsAdecreased LDLr protein expression. When wecompared the CsA-mediated cytotoxicity inpSHAG and LDLr-792 transfected 293T cells,we found a significant difference at a dose of 10mg/mL of CsA, where LDLr downregulation lead to an
DOI 10.1002/jps J
enhanced viability (reduced cytotoxicity, Fig. 4C).We also found that CsA decreased LDLr proteinexpression at doses above 1 mg/mL. The effect ofCsA on LDLr receptor is an interesting findingwhich might explain partially the increase inplasma cholesterol in patients treated with thisdrug. The downregulation of LDLr expression byCsA precludes the internalization of LDL particlesand leads to increased plasma cholesterol levels.We also tested the specificity of the LDLr down-regulation effect on CsA toxicity by using anothernephrotoxic drug, gentamycin. Schmitz et al.40
showed that gentamycin internalization wasmediated by megalin, another member of theLDL receptor family. We did not find anydifference in the viability of pSHAG or LDLrLOW
cells exposed to gentamycin suggesting that theeffect is specific for CsA.
OURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008

2360 LEON ET AL.
The overexpression of the LDLr led to anincrease in radioactive CsA uptake compared tocontrol cells (75%). Taken together, these resultssuggest that CsA internalization and nephrotoxi-city are partially mediated by LDLr-dependentuptake.
It is well established that certain drugsassociate with lipoproteins; as such, lipoproteinscan act as a natural drug delivery system forhydrophobic drugs or lipid-based formulations.Nevertheless, it is the association of these drugswith lipoproteins that may be able to explain theirpharmacological activity, pharmacokinetic prop-erties as well as its toxicities. Furthermore, byunderstanding the uptake mechanisms of specificdrug delivery systems can provide better ther-apeutic treatments and improve administration topatients who experience side effects or lowefficacy. Hence, the elucidation of whether theLDL receptor super-family can play a relevantrole in uptake of specific drugs will be able toimprove drug targeting and hence formulationconsiderations. Our results support the appro-priateness of this cell model in studies involvingrenal cell cholesterol uptake and drug uptake andmetabolism.
ACKNOWLEDGMENTS
We thank Dr. G. Hannon and Dr. N.E. Reiner forproviding us with the pSHAG vector and thepSHAG-Annexin V plasmid, respectively. Theauthors wish to thank Mr. Stephen Lee and Ms.Wendy Li for excellent technical assistance. Thiswork was supported with grants from the Cana-dian Institutes of Health Research to KMW andJSH.
REFERENCES
1. Anderson RG, Goldstein JL, Brown MS. 1976.Localization of low density lipoprotein receptorson plasma membrane of normal human fibroblastsand their absence in cells from a familial hyperch-olesterolemia homozygote. Proc Natl Acad Sci USA73:2434–2438.
2. Goldstein JL, Basu SK, Brown MS. 1983. Receptor-mediated endocytosis of low-density lipoprotein incultured cells. Methods Enzymol 98:241–260.
3. Jeon H, Blacklow SC. 2005. Structure and physio-logic function of the low-density lipoprotein recep-tor. Annu Rev Biochem 74:535–562.
JOURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008
4. Beglova N, Blacklow SC. 2005. The LDL receptor:How acid pulls the trigger. Trends Biochem Sci 30:309–317.
5. Goldstein JL, Brown MS. 1974. Binding and degra-dation of low density lipoproteins by culturedhuman fibroblasts. Comparison of cells from a nor-mal subject and from a patient with homozygousfamilial hypercholesterolemia. J Biol Chem 249:5153–5162.
6. Chung NS, Wasan KM. 2004. Potential role of thelow-density lipoprotein receptor family as media-tors of cellular drug uptake. Adv Drug Deliv Rev56:1315–1334.
7. Schmitz G, Bruning T, Kovacs E, Barlage S. 1993.Fluorescence flow cytometry of human leukocytesin the detection of LDL receptor defects in thedifferential diagnosis of hypercholesterolemia.Arterioscler Thromb 13:1053–1065.
8. Hobbs HH, Russell DW, Brown MS, Goldstein JL.1990. The LDL receptor locus in familial hyperch-olesterolemia: Mutational analysis of a membraneprotein. Annu Rev Genet 24:133–170.
9. Brocks DR, Wasan KM. 2002. The influence of lipidson stereoselective pharmacokinetics of halofan-trine: Important implications in food-effect studiesinvolving drugs that bind to lipoproteins. J PharmSci 91:1817–1826.
10. McIntosh MP, Charman WN, Campbell M, PorterCJ. 2004. Influence of physicochemical propertieson the patterns of association of a series of aliphaticesters of halofantrine with plasma lipoproteins.J Control Release 95:275–289.
11. Peteherych KD, Wasan KM. 2001. Effects of lipo-proteins on Cyclosporine A toxicity and uptake inLLC-PK1 pig kidney cells. J Pharm Sci 90:1395–1406.
12. Kim T, Lu SK, Brunner LJ. 2003. The effect oflipoprotein-associated Cyclosporine on drug meta-bolism and toxicity in rats. PDA J Pharm Sci Tech-nol 57:410–424.
13. Sanghvi A, Warty VS, Diven W, Starzl FT. 1989.Receptor-mediated cellular uptake of Cyclosporine.Transplant Proc 21:858–859.
14. Hannon GJ. 2002. RNA interference. Nature 418:244–251.
15. Paddison PJ, Caudy AA, Hannon GJ. 2002b. Stablesuppression of gene expression by RNAi in mamma-lian cells. Proc Natl Acad Sci USA 99:1443–1448.
16. Lee JS, Hmama Z, Mui A, Reiner NE. 2004. Stablegene silencing in human monocytic cell lines usinglentiviral-delivered small interference RNA. Silen-cing of the P110alpha isoform of phosphoinositide 3-kinase reveals differential regulation of adherenceinduced by 1alpha,25-dihydroxycholecalciferol andbacterial lipopolysaccharide. J Biol Chem 279:9379–9388.
17. Hirano T, Kawamura T, Fukuda S, Kohsaka S,Yoshikawa N, Yoshida M, Oka K. 2003. Implication
DOI 10.1002/jps

LDL RECEPTOR LEVELS AFFECT CYCLOSPORIN A-MEDIATED CYTOTOXICITY 2361
of cholesterol in Cyclosporine pharmacodynamicsin minimal change nephrotic syndrome. Clin Phar-macol Ther 74:581–590.
18. Serkova NJ, Christians U, Benet LZ. 2004. Bio-chemical mechanisms of cyclosporine neurotoxicity.Mol Interv 4:97–107.
19. Midtvedt K. 2004. Therapeutic drug monitoring ofcyclosporine. Transplant Proc 36:430S–433S.
20. Cole E, Cheung F, Wong PY, Fung LS, Skorecki K,Levy GA. 1989. Toxic effects on renal cells in cul-ture—a comparison of Cyclosporin A and its meta-bolites. Transplant Proc 21:943–945.
21. Healy E, Dempsey M, Lally C, Ryan MP. 1998.Apoptosis and necrosis: Mechanisms of cell deathinduced by Cyclosporine A in a renal proximaltubular cell line. Kidney Int 54:1955–1966.
22. Bren A, Pajek J, Grego K, Buturovic J, Ponikvar R,Lindic J, Knap B, Vizjak A, Ferluga D, Kandus A.2005. Follow-up of kidney graft recipients withCyclosporine-associated hemolytic-uremic syn-drome and thrombotic microangiopathy. Trans-plant Proc 37:1889–1891.
23. Raine AE, Carter R, Mann JI, Morris PJ. 1988.Adverse effect of Cyclosporin on plasma cholesterolin renal transplant recipients. Nephrol Dial Trans-plant 3:458–463.
24. Leon C, Nandan D, Lopez M, Moeenrezakhanlou A,Reiner NE. 2006. Annexin V associates with theIFN gamma receptor and regulates IFN gammasignaling. J Immunol
25. Paddison PJ, Caudy AA, Bernstein E, Hannon GJ,Conklin DS. 2002a. Short hairpin RNAs (ShRNAs)induce sequence-specific silencing in mammaliancells. Genes Dev 16:948–958.
26. Stephan ZF, Yurachek EC. 1993. Rapid fluoro-metric assay of LDL receptor activity by DiI-labeledLDL. J Lipid Res 34:325–330.
27. Lopes-Virella MF, Binzafar N, Rackley S, Takei A,La VM, Virella G. 1997. The Uptake of LDL-IC byhuman macrophages: Predominant involvement ofthe Fc gamma RI receptor. Atherosclerosis135:161–170.
28. Truong TQ, Falstrault L, Tremblay C, Brissette L.1999. Low density lipoprotein-receptor plays amajor role in the binding of very low density lipo-proteins and their remnants on HepG2 Cells. Int JBiochem Cell Biol 31:695–705.
29. Chung NS, Sachs-Barrable K, Lee SD, Wasan KM.2005. Suitability of LLC-PK1 pig kidney cells forthe study of drug action on renal cell cholesterol
DOI 10.1002/jps J
uptake: Identification and characterization of low-density lipoprotein receptors. J Pharmacol ToxicolMethods 51:139–145.
30. Becker GM, Gandolfi AJ, Nagle RB. 1987. Effects ofCyclosporin A on a kidney epithelial cell line (LLC-PK1). Res Commun Chem Pathol Pharmacol56:277–280.
31. Massicot F, Lamouri A, Martin C, Pham-Huy C,Heymans F, Warnet JM, Godfroid JJ, Claude JR.1997. Preventive effects of two PAF-antagonists,PMS 536 and PMS 549,on Cyclosporin-inducedLLC-PK1 oxidative injury. J Lipid Mediat CellSignal 15:203–214.
32. Aronin N. 2006. Target selectivity in mRNA silen-cing. Gene Ther 13:509–516.
33. Elbashir SM, Harborth J, Lendeckel W, Yalcin A,Weber K, Tuschl T. 2001. Duplexes of 21-nucleotideRNAs mediate RNA interference in cultured mam-malian cells. Nature 411:494–498.
34. Brummelkamp TR, Bernards R, Agami R. 2002.A System for stable expression of short interferingRNAs in mammalian cells. Science 296:550–553.
35. Tiscornia G, Singer O, Ikawa M, Verma IM. 2003.A general method for gene knockdown in mice byusing lentiviral vectors expressing small interfer-ing RNA. Proc Natl Acad Sci USA 100:1844–1848.
36. Sato H, Ogata H, De Luca LM. 2000. Annexin Vinhibits the 12-O-tetradecanoylphorbol-13-acetate-induced activation of Ras/extracellular signal-regu-lated kinase (ERK) signaling pathway upstream ofShc in MCF-7 cells. Oncogene 19:2904–2912.
37. Metah KD. 2002. Role of mitogen-activated proteinkinases and protein kinase C in regulating low-density lipoprotein receptor expression. Gene Expr10:153–164.
38. Luke DR. 1992. Immunosuppressive effect of cyclo-sporine in the hyperlipidemic rat model. BiopharmDrug Dispos 13:635–645.
39. Barbari AG, Masri MA, Stephan AG, El Ghoul B,Rizk S, Mourad N, Kamel GS, Kilani HE, KaramAS. 2006. Cyclosporine lymphocyte maximumlevel monitoring in de novo kidney transplantpatients: A prospective study. Exp Clin Transplant4:400–405.
40. Schmitz C, Hilpert J, Jacobsen C, Boensch C,Christensen EI, Luft FC, Willnow TE. 2002.Megalin deficiency offers protection from renalaminoglycoside accumulation. J Biol Chem 277:618–622.
OURNAL OF PHARMACEUTICAL SCIENCES, VOL. 97, NO. 6, JUNE 2008