modélisation des successions post-culturales. application à la
TRANSCRIPT

ORSAY N°D’ORDRE
UNIVERSITE DE PARIS-SUD U.F.R. SCIENTIFIQUE D’ORSAY
Thèse présentée pour obtenir le grade de
DOCTEUR EN SCIENCES DE L’UNIVERSITÉ PARIS 11, ORSAY
Discipline : Écologie
par
Yann MARTINEAU
Modélisation des successions post-culturales. Application à la gestion durable des agro-
écosystèmes des hautes Andes tropicales.
Soutenance prévue le 29 Septembre 2004 devant le jury constitué de : M. ABBADIE Luc, Examinateur M. GARNIER Eric, Rapporteur M. LEADLEY Paul, Examinateur M. SAUGIER Bernard, Directeur de thèse M. SOUSSANA Jean-François, Rapporteur

Département d’Ecophysiologie Végétale Laboratoire d’Ecologie, Systématique, Evolution Bâtiment 362 Université Paris-Sud XI 91405 Orsay Cedex [email protected]

I
Ce travail a bénéficié des financements Du Ministère de l’Education Nationale,
de l’Enseignement Supérieur et de la Recherche sous la forme d’une allocation moniteur polytechnicien (AMX),
De la Commission Européenne dans le
cadre du projet TROPANDES (INCO-DC N° ERBIC18-CT98-0263),
De la Fondation Européenne pour la
Science (ESF) dans le cadre du programme LINKECOL (Linking ecosystem and community ecology).

II
A mes racines, parents et grands-parents, qui m’ont transmis leurs semelles de vent, leur indépendance et une certaine idée de la liberté ;
Et aux pierres qui, au hasard des
rencontres, marquent les chemins : pierres tombales, cairns et pierres philosophales.

III
Remerciements

1
Sommaire
2
14 15 18 21
36 37 55 70
80 81 86 90
93
96
106
109
A1 A2
A40
A75
A96
3° 4° 4°
carte
Introduction 1 - Développement d’un modèle mécaniste de succession végétale (FAPROM) 11 - Pourquoi modéliser ? 12 - À partir de quoi modéliser ? 13 - Comment modéliser ? 2 - Applications du modèle, de la plante à l’écosystème 21 - Croissance d’un couvert végétal monospécifique 22 - Dynamique de la communauté végétale 23 - Rôle de la jachère dans l’agro-écosystème 3 - Bilan critique et perspectives 31 - Hypothèses du modèle 32 - Projet de thèse 33 - Réflexions personnelles et avenir du modèle FAPROM Conclusion Références Table des illustrations Table des matières Annexes 1 A process-based model of old-field succession linking ecosystem and community ecology. Martineau Y, Saugier B. Soumis. 2 Evidence of hidden fluxes in the estimation of Net Primary Production in herbaceous ecosystems. Sarmiento L, Martineau Y, Acevedo D, Coûteaux MM, Saugier B, Escalona A. En préparation. 3 Uses and abuses of logistic growth: considering turnover to avoid biological nonsense in ecological modelling. Martineau Y, Deredec A, Genton B, Saugier B. En projet. 4 Modèle mécaniste simplifié de succession secondaire. Piste de réflexion. Resumen en español Résumé en français Abstract in English Equations et paramètres du modèle

2
Introduction
Des systèmes de culture à jachère
Des successions post-culturales
Du projet TROPANDES
Des sites d’étude
Des rotations culturales andines
Des sols et de la fertilité
De la production végétale au cours de la succession post-culturale
Du modèle FAPROM et des questions étudiées
De l’articulation du travail et du plan de la synthèse
3
3
4
7
7
9
11
11
12

3
Ce travail a été développé dans le cadre du projet européen TROPANDES dont
l’objectif principal était de comprendre la dynamique de la fertilité du sol, base d’une gestion
durable du système de culture à jachère longue des hautes Andes tropicales. Il a pour but de
modéliser les successions post-culturales.
Des systèmes de culture à jachère
La jachère définit l’état d’une terre labourable qu’on abandonne un certain temps sans
produire de récolte (dictionnaire Larousse, 1979). Historiquement, la jachère était utilisée
pour laisser reposer la terre et la débarrasser des mauvaises herbes (Sebillotte et al. 1993). Au
cours du 20ème siècle, la jachère a été très largement abandonnée au profit de l’utilisation
d’engrais et de pesticides. Smil (2001) a souligné l’importance de la découverte du procédé
d’Haber-Bosch (synthèse chimique de fertilisants azotés) : l’alimentation de 40% de la
population mondiale en dépend aujourd’hui ! Toutefois, l’étude des jachères n’a pas perdu
son intérêt : (1) certains agronomes posent à l’échelle mondiale la question de la durabilité des
systèmes de culture intensifs (Crews & Peoples 2004) ; (2) la politique agricole commune
mise en place par l’Europe en 1992 encourage la mise en jachère de 15% des terres
cultivables pour limiter la production ce qui nécessite la recherche de nouveaux modes de
gestion des terres (Doré & Dalbiès 1994) ; (3) et enfin, objet de notre recherche, la jachère
reste dans de nombreuses régions du monde la pratique agricole incontournable pour pallier le
manque de fertilité du sol1 (Floret & Pontanier 2000). Dans le système traditionnel, les
paysans abandonnent les parcelles quand les rendements chutent. Au moment de rompre la
jachère, parfois plusieurs dizaines d’années plus tard, le travail de labour consiste à incorporer
la végétation naturelle à la terre. La végétation de la jachère joue ainsi le rôle d’engrais vert en
se décomposant lentement dans le sol pendant la culture.
Des successions post-culturales
La végétation au cours de la jachère suit un processus de succession. C’est ainsi qu’on
appelle en écologie le remplacement successif des espèces au cours du temps (Lepart &
Escarré 1983) : les espèces dites pionnières colonisent rapidement le sol laissé nu après la
culture ; des espèces dites intermédiaires s’installent progressivement tandis qu’évincées par
la compétition, les espèces pionnières disparaissent peu à peu ; les espèces de fin de
succession dominent plus tardivement. La dynamique post-culturale est qualifiée de
1 Nous entendons par fertilité l’aptitude du sol à produire.

4
succession secondaire : suite à la perturbation qu’il subit, l’écosystème tend à retourner à
l’état naturel (climax, Clements 1916). Dans les environnements aux conditions climatiques
extrêmes (écosystèmes arctiques et montagnards en particulier), on constate plus de
changements d’abondance que de remplacements stricto sensu d’espèces ; Muller (1952)
parle d’autosuccession et Urbanska (1997) de succession démographique. Les motifs de
succession sont déterminés par de nombreux facteurs parmi lesquels : (1) la composition
initiale de la végétation (Facelli & Pickett 1990) [conditions initiales] ; (2) les conditions
locales du milieu, en particulier la fertilité du sol (Inouye & Tilman 1995) [conditions
environnementales] ; et (3) la disponibilité des graines au sein du paysage (Frelich & Reich
1995) [conditions aux limites]. La structure et la dynamique des communautés végétales
résultent de compromis entre les capacités de dispersion et de compétition des espèces (Aerts
1999, Tilman 1997). De nombreux auteurs ont ainsi proposé de classer les espèces en
différentes stratégies : r-K (e.g. Hastings 1980, Tilman 1988, 1990, Ehrlen & Groenendael
1998) ; C-S-R, espèces compétitives, tolérantes au stress et rudérales (Grime 1974, 1988,
2001). Dans une succession, les espèces pionnières (stratégie r) sont suivies des espèces
compétitives puis d’espèces tolérantes (stratégie K).
Du projet TROPANDES
Le projet européen TROPANDES affiche comme objectif à long terme d’améliorer les
revenus des paysans et le niveau de vie des populations rurales qui pratiquent l’agriculture à
jachère longue dans les hautes Andes tropicales (Carballas et al. 2002a). Il s’est développé sur
une proposition de Lina Sarmiento dans la continuité des études menées à l’Universidad de
los Andes (Mérida, Venezuela) sur le paramo2 de Gavidia. Dans l’optique de définir les bases
agro-écologiques d’une agriculture à jachère durable, il a réuni, de 1998 à 2002, 7 équipes de
chercheurs européens et sud-américains (le Tableau 1 donne la liste des institutions
participantes) autour des trois axes de recherche suivants : (1) le système de culture au niveau
de l’exploitation et à l’échelle régionale ; (2) les interactions plantes/sol au cours de la
jachère ; et (3) les interactions plantes/sol au cours de la culture de pomme de terre. Le
Tableau 2 fournit le détail des études définies dans la proposition du projet. Il montre la
dimension pluridisciplinaire de la problématique et la démarche de développement durable
dans laquelle elle s’inscrit : la question de l’amélioration économique du système de culture
intègre différents aspects qui touchent à la fois l’économie, l’agronomie et l’écologie.
2 Paramo : lande caractéristique des régions humides des Andes du Nord.

5
Tableau 1. Les partenaires du projet européen TROPANDES (1998-2002). (1) Instituto de Investigaciones Agrobiologicas de Galicia (IIAG)
Consejo Superior de Investigaciones Científicas (CSIC), Santiago de Compostela, Espagne Tarsy Carballas, coordinateur
(2) Instituto de Ciencias Ambientales y Ecologicas (ICAE) Universidad de Los Andes (ULA), Mérida, Venezuela Maximina Monasterio
(3) Instituto de Ecologia (IE) Universidad Mayor de San Andres (UMSA), La Paz, Bolivie Stephan Beck
(4) Centre d’Ecologie Fonctionnelle et Evolutive (CEFE) Centre National de la Recherche scientifique (CNRS) UPR 90560, Montpellier, France Pierre Bottner
(5) Plant Research International Wageningen University and Research Centre, Pays-Bas Klaas Metselaar
(6) Laboratoire d’Ecologie, Systématique & Evolution (ESE) Université Paris Sud et CNRS UMR 8079, Orsay, France Bernard Saugier
(7) Laboratoire d’Etudes Rurales (LER) Institut de Recherche pour le Développement (IRD), Montpellier, France Dominique Hervé
Tableau 2. Les activités du projet européen TROPANDES (1998-2002). 1 – Bases agro-écologiques du système de culture à jachère longue des hautes Andes tropicales.
1.1 – Distribution spatiale de l’agriculture à jachère à l’échelle régionale. 1.2 – Dynamique du système culture/jachère à l’échelle de l’exploitation. 1.3 – Développement et application d’un modèle d’exploitation agricole. 1.4 – Application du modèle intégré à l’évaluation des stratégies de gestion d’exploitation (études de scénarios).
2 – Interactions plantes/sol au cours de la jachère. 2.1 – Diversité spécifique et production végétale.
2.1.1 – Typologie de la succession. 2.1.2 – Caractérisation qualitative et quantitative de la litière.
2.2 – Décomposition au cours de la jachère. 2.2.1 – Décomposition de la litière. 2.2.2 – Transferts de carbone et d’azote des plantes à la matière organique du sol.
2.3 – Effet de la jachère sur la MOS et sur la biomasse et l’activité microbiennes. 2.4 – Manipulation de la jachère (introduction de légumineuses). 2.5 – Développement et application de modèles.
2.5.1 – FAPROM : modèle de production de la jachère. 2.5.2 – MOMOS : modèle de décomposition de la matière organique du sol.
3 – Interactions plantes/sol pendant la culture. 3.1 – Développement de la pomme de terre à différents stades de succession. 3.2 – Transferts d’azote de la matière organique du sol à la pomme de terre. 3.3 – Décomposition des résidus de culture. 3.4 – Effet de la culture sur la MOS et sur la biomasse et l’activité microbiennes. 3.5 – Développement et application de modèles.
3.5.1 – LINTUL - SAHEL : modèles de culture et de bilan hydrique. 3.5.2 – MOMOS : modèle de décomposition de la matière organique du sol.

6
Trois objectifs concrets ont été définis au niveau de l’écosystème : (1) comprendre les
mécanismes de perte et de restauration de la fertilité du sol au cours d’un cycle
culture/jachère ; (2) tester des scénarios de gestion de la jachère qui permettent d’améliorer
les rendements de la culture de pomme de terre ; et (3) élaborer un modèle du fonctionnement
du système de culture à jachère longue des hautes Andes tropicales. Les études agro-
écologiques à l’échelle de la parcelle sont fondées sur l’hypothèse suivante : la perte rapide de
la fertilité au cours de la culture n’est pas due à un stock trop faible de nutriments mais à leur
faible disponibilité. La restauration de la fertilité résulterait d’une lente et progressive
mobilisation de l’azote par la biomasse microbienne, de la fraction stable à la fraction labile
de la matière organique. La jachère stimulerait l’activité de la biomasse microbienne.
Figure 1. Situation géographique et diagrammes climatiques des sites d’étude du projet TROPANDES.
A gauche : carte d’Amérique du Sud : Copyright © 2001 Yahoo! France & Hachette Multimédia / Hachette Livre. A droite : diagrammes climatiques. Les barres présentées sous les diagrammes climatiques symbolisent la durée de la saison sèche (en blanc) et la durée de la saison humide (en noir). Les espaces grisés représentent les périodes de l’année qui sont sèches ou humide en fonction du régime de précipitation de l’année considérée. a) Gavidia (Précipitations : 1300 mm par an ; Température moyenne annuelle : 8,4°C ; Amplitude diurne maximale : 17,3°C) b) Patarani (Précipitations : 400 mm par an ; Température moyenne annuelle : 10,0°C ; Amplitude diurne maximale : 29,4°C)

7
Des sites d’étude
L’étage supérieur des Andes tropicales s’étend entre 3200 et 4200 mètres d’altitude du
nord de l’Argentine et du Chili à la cordillère vénézuélienne. Le climat y subit l’influence
conjointe des tropiques et de la haute montagne : il se caractérise par deux saisons marquées,
l’une sèche, l’autre humide ; une température annuelle moyenne relativement basse ; et un fort
rayonnement. Les équipes du projet TROPANDES ont étudié en particulier deux sites aux
situations contrastées (Carballas et al. 2002a et b, Figure 1 p.6) : (1) le village de Gavidia
(8°35’ N, 70°52’ W, Sierra de Mérida, Venezuela) est situé entre 3200 et 3800
mètres d’altitude dans une ancienne vallée glaciaire ; le total annuel de précipitations entre
1990 et 1999 varie de 1100 à 1700 mm avec une moyenne de 1300 mm par an ; le
rayonnement global journalier moyen est de 15,8 MJ m-2 (Fontaine 2000) et la température
moyenne annuelle chute de 10°C à 3200 m à 6°C à 3800 m (8,4°C à la station climatique,
Llambi et al. 2003) ; (2) la communauté de Patarani (17°06’ S, 68°00’ W, Patacamaya,
Bolivie) est localisée sur l’Altiplano à 110 km au sud de La Paz, à 3800 m d’altitude ; le total
annuel de précipitation varie de 300 à 450 mm (en moyenne 409 mm), le rayonnement global
journalier moyen est de 21,6 MJ m-2 (Sarps 2001) et la température moyenne annuelle est de
10,0°C (moyennes des années 1981-1991, Ortuño et al. 2004) mais avec de très fortes
variations journalières en saison sèche (amplitude thermique maximale : 29,4°C) et en
moyenne, 201 jours de gel par an (Migueis et al. 1998). A ces conditions climatiques
extrêmes correspondent deux écosystèmes naturels distincts : (1) le paramo s’étend dans les
régions humides des Andes du Nord (Equateur, Colombie et Venezuela) et se caractérise par
des communautés dominées par des arbustes (tel que Hypericum laricifolium Juss.) et des
rosettes géantes endémiques (comme Espeletia schultzii Wedd., emblématique du paramo de
Gavidia ; Monasterio 1980) ; (2) la puna3, caractéristique des Andes centrales semi-arides
(Pérou et Altiplano bolivien) est une lande dominée par des touffes de graminées pérennes, en
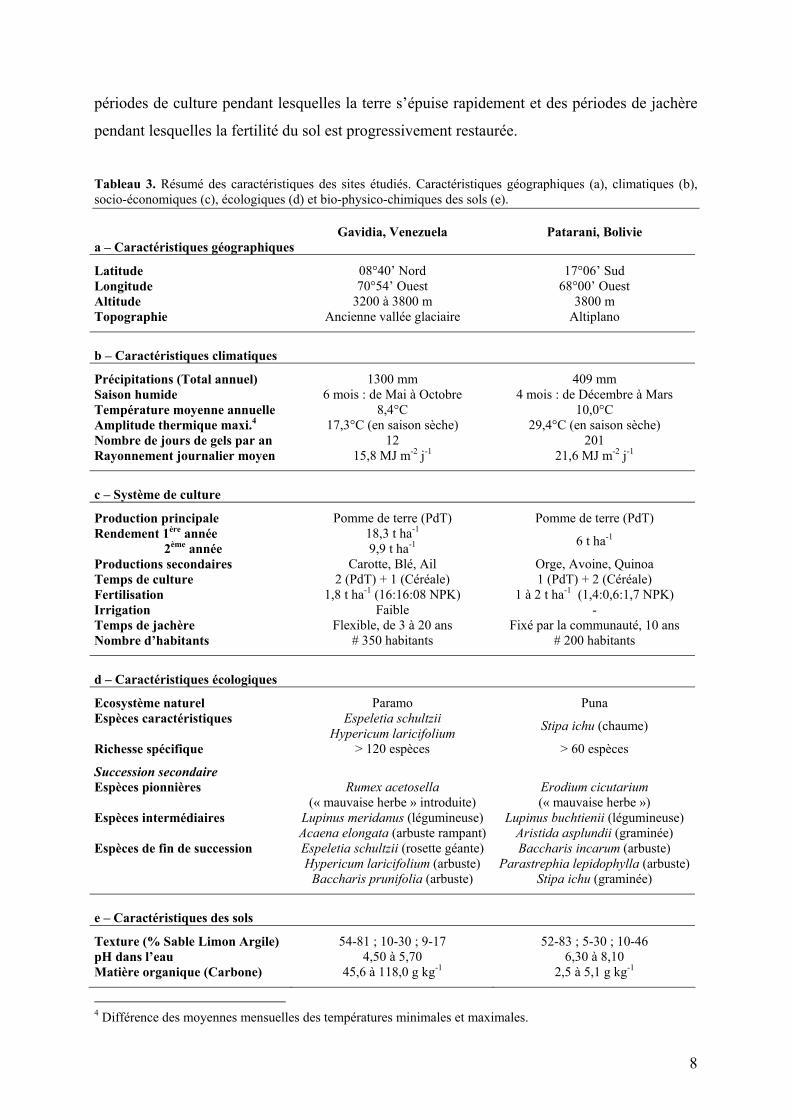
particulier, Stipa ichu. Le Tableau 3 (p.8) résume les caractéristiques géographiques,
climatiques, écologiques et socio-économiques des deux sites étudiés.
Des rotations culturales andines
Bien que les conditions naturelles soient peu favorables, les terres des hautes Andes
tropicales sont cultivées. La pomme de terre est la culture de subsistance des 3 millions de
sud-américains qui y vivent (Llambi et al. 2003). Le système agricole traditionnel alterne des
3 Puna : lande caractéristique des régions semi-arides des Andes centrales.
8
périodes de culture pendant lesquelles la terre s’épuise rapidement et des périodes de jachère
pendant lesquelles la fertilité du sol est progressivement restaurée.
Tableau 3. Résumé des caractéristiques des sites étudiés. Caractéristiques géographiques (a), climatiques (b), socio-économiques (c), écologiques (d) et bio-physico-chimiques des sols (e). Gavidia, Venezuela Patarani, Bolivie a – Caractéristiques géographiques
Latitude 08°40’ Nord 17°06’ Sud Longitude 70°54’ Ouest 68°00’ Ouest Altitude 3200 à 3800 m 3800 m Topographie Ancienne vallée glaciaire Altiplano
b – Caractéristiques climatiques
Précipitations (Total annuel) 1300 mm 409 mm Saison humide 6 mois : de Mai à Octobre 4 mois : de Décembre à Mars Température moyenne annuelle 8,4°C 10,0°C Amplitude thermique maxi.4 17,3°C (en saison sèche) 29,4°C (en saison sèche) Nombre de jours de gels par an 12 201 Rayonnement journalier moyen 15,8 MJ m-2 j-1 21,6 MJ m-2 j-1
c – Système de culture
Production principale Pomme de terre (PdT) Pomme de terre (PdT) Rendement 1ère année 18,3 t ha-1 2ème année 9,9 t ha-1 6 t ha-1
Productions secondaires Carotte, Blé, Ail Orge, Avoine, Quinoa Temps de culture 2 (PdT) + 1 (Céréale) 1 (PdT) + 2 (Céréale) Fertilisation 1,8 t ha-1 (16:16:08 NPK) 1 à 2 t ha-1 (1,4:0,6:1,7 NPK) Irrigation Faible - Temps de jachère Flexible, de 3 à 20 ans Fixé par la communauté, 10 ans Nombre d’habitants # 350 habitants # 200 habitants
d – Caractéristiques écologiques
Ecosystème naturel Paramo Puna Espèces caractéristiques Espeletia schultzii
Hypericum laricifolium Stipa ichu (chaume)
Richesse spécifique > 120 espèces > 60 espèces
Succession secondaire Espèces pionnières Rumex acetosella
(« mauvaise herbe » introduite) Erodium cicutarium (« mauvaise herbe »)
Espèces intermédiaires Lupinus meridanus (légumineuse) Acaena elongata (arbuste rampant)
Lupinus buchtienii (légumineuse) Aristida asplundii (graminée)
Espèces de fin de succession Espeletia schultzii (rosette géante) Hypericum laricifolium (arbuste)
Baccharis prunifolia (arbuste)
Baccharis incarum (arbuste) Parastrephia lepidophylla (arbuste)
Stipa ichu (graminée)
e – Caractéristiques des sols
Texture (% Sable Limon Argile) 54-81 ; 10-30 ; 9-17 52-83 ; 5-30 ; 10-46 pH dans l’eau 4,50 à 5,70 6,30 à 8,10 Matière organique (Carbone) 45,6 à 118,0 g kg-1 2,5 à 5,1 g kg-1
4 Différence des moyennes mensuelles des températures minimales et maximales.

9
La rotation culturale est plus ou moins figée : (1) à Gavidia, les parcelles sont
généralement abandonnées après deux années de pomme de terre, éventuellement suivies par
une troisième année de culture (blé, carotte, ail) ; la durée de la jachère est variable, de deux
ans pour les terres de fond de vallée, proches des maisons, à plusieurs dizaines d’années pour
les terres les plus éloignées ; (2) la gestion communautaire de Patarani fixe les règles
suivantes : un an de pomme de terre, deux ans de céréale (orge et quinoa) puis dix ans de
jachère (Hervé et al. 2003). L’accroissement des populations locales et l’augmentation
consécutive des besoins vivriers poussent les paysans à chercher de meilleurs rendements
(Gutierrez 1996). L’utilisation d’intrants (engrais organique et minéral) est assez répandue au
Venezuela mais son coût en limite l’usage en Bolivie5. Quoiqu’il en soit, les engrais ne
peuvent se substituer à la jachère (Sarmiento 1995). Dans certaines vallées vénézuéliennes
moins élevées (altitude inférieure à 3000 m), la culture intensive est cependant soutenue par la
fertilisation et l’irrigation des parcelles (Sarmiento et al. 1993, 2002). A Gavidia, le temps de
jachère tend aussi à se raccourcir mais la diminution des rendements aboutit finalement à
l’abandon de la terre après plusieurs années (Sarmiento & Bottner 2002). Sur l’Altiplano, on
constate également une tendance au raccourcissement du temps de jachère mais par le biais
d’une diminution des surfaces consacrées à ce système de culture, puisque les règles
communautaires fixent la durée de la jachère (Hervé & Rivière 1988).
En sus de son importance dans la récupération de la fertilité, la jachère joue d’autres
rôles : la période de repos et le travail du sol permettent de limiter les populations des
nématodes à kystes de la pomme de terre, de contrôler les parasites et de lutter contre les
plantes adventices ; les terres en jachère sont pâturées ; la végétation naturelle fournit des
matières premières, bois de feu, graminées pour le chaume des maisons (en Bolivie
notamment, Pestalozzi 2000, Camacho 2001), plantes aux vertus médicinales (les feuilles
d’Espeletia par exemple sont utilisées dans la préparation d’infusion) ; et à l’échelle du
paysage, le système de culture à jachère longue maintient une importante biodiversité (plus de
120 espèces végétales ont été répertoriées dans le paramo de Gavidia, Sarmiento et al. 2002).
Des sols et de la fertilité
Néanmoins, la jachère est surtout pratiquée à cause de la perte de fertilité du sol. Les
rendements décroissent en effet fortement après deux années consécutives de culture que ce
soit à Gavidia (Sarmiento 1995) ou sur l’Altiplano bolivien (Hervé 1994), et plus fortement
5 Sur l’Altiplano, le seul intrant est organique : déjections ovines (guano).
10
encore pour les rotations pomme de terre / pomme de terre que pour les rotations pomme de
terre / céréales (Hervé & Coûteaux, communication personnelle). Les courbes de dilution en
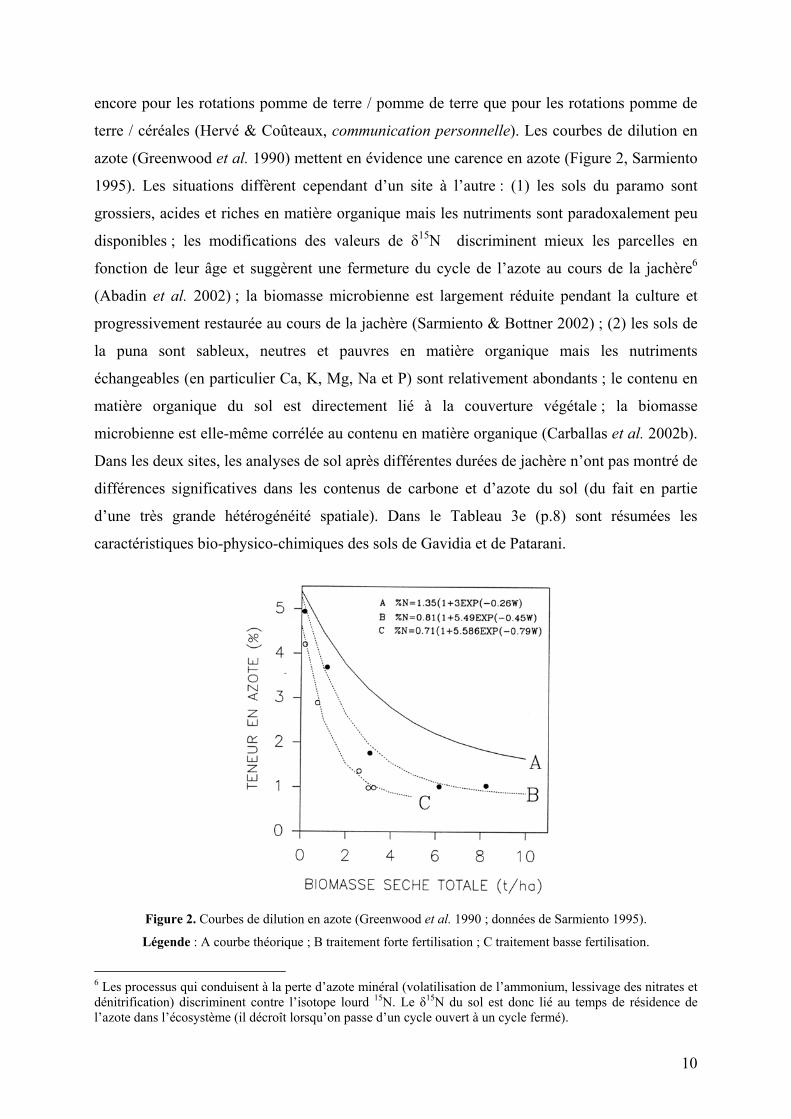
azote (Greenwood et al. 1990) mettent en évidence une carence en azote (Figure 2, Sarmiento
1995). Les situations diffèrent cependant d’un site à l’autre : (1) les sols du paramo sont
grossiers, acides et riches en matière organique mais les nutriments sont paradoxalement peu
disponibles ; les modifications des valeurs de δ15N discriminent mieux les parcelles en
fonction de leur âge et suggèrent une fermeture du cycle de l’azote au cours de la jachère6
(Abadin et al. 2002) ; la biomasse microbienne est largement réduite pendant la culture et
progressivement restaurée au cours de la jachère (Sarmiento & Bottner 2002) ; (2) les sols de
la puna sont sableux, neutres et pauvres en matière organique mais les nutriments
échangeables (en particulier Ca, K, Mg, Na et P) sont relativement abondants ; le contenu en
matière organique du sol est directement lié à la couverture végétale ; la biomasse
microbienne est elle-même corrélée au contenu en matière organique (Carballas et al. 2002b).
Dans les deux sites, les analyses de sol après différentes durées de jachère n’ont pas montré de
différences significatives dans les contenus de carbone et d’azote du sol (du fait en partie
d’une très grande hétérogénéité spatiale). Dans le Tableau 3e (p.8) sont résumées les
caractéristiques bio-physico-chimiques des sols de Gavidia et de Patarani.
Figure 2. Courbes de dilution en azote (Greenwood et al. 1990 ; données de Sarmiento 1995).
Légende : A courbe théorique ; B traitement forte fertilisation ; C traitement basse fertilisation.
6 Les processus qui conduisent à la perte d’azote minéral (volatilisation de l’ammonium, lessivage des nitrates et dénitrification) discriminent contre l’isotope lourd 15N. Le δ15N du sol est donc lié au temps de résidence de l’azote dans l’écosystème (il décroît lorsqu’on passe d’un cycle ouvert à un cycle fermé).

11
De la production végétale au cours de la succession post-culturale
Sous la responsabilité de Bernard Saugier, le partenaire numéro 6 du projet
TROPANDES (Université Paris-Sud, Laboratoire d’Ecologie Systématique & Evolution,
Département d’Ecophysiologie Végétale) était chargé de modéliser la production végétale de
la jachère au cours de la succession. La première esquisse de modèle (Saugier, rapport
intermédiaire 1998, communication personnelle) calculait la production primaire de
l’écosystème à partir de l’efficience d’utilisation de la lumière des différentes plantes. Dans ce
but, Magali Fontaine (2000) a mesuré pour les espèces les plus courantes du paramo (site
vénézuélien) la réponse de la photosynthèse à la lumière ainsi que plusieurs traits
écophysiologiques ou morphologiques (masse surfacique des feuilles, répartition de la
biomasse, angle foliaire moyen). Un travail similaire a été effectué en 2001 sur les espèces les
plus courantes de la puna (site bolivien) par Bernard Saugier et Jean-Yves Pontailler (données
de photosynthèse, 2003) et Audrey Sarps (masse surfacique des feuilles, répartition de la
biomasse et angle foliaire moyen, 2001). Mon travail de DEA s’est appuyé sur les travaux de
Magali Fontaine publiés par Llambi et al. (2003) pour construire la première version du
modèle FAPROM (Fallow Production Model, Martineau 2001).
Du modèle FAPROM et des questions étudiées
Nous avons construit un modèle mécaniste de succession secondaire fondé sur les
cycles biogéochimiques, en particulier, sur l’estimation de la photosynthèse pour quantifier la
dynamique de la production végétale de la jachère. La prise en compte des espèces principales
(caractéristiques des différents stades de succession) devait permettre à la fois d’intégrer les
différents taux de croissance des espèces et de quantifier la qualité de la litière produite. Le
modèle décrit ainsi le fonctionnement de l’écosystème du point de vue de ses constituants (les
espèces végétales assemblées) : il entre dans la catégorie des modèles multi-agents (Bonabeau
2002). Le modèle de communauté végétale fonctionne comme un système complexe auto-
organisé (Capra 1996) : il est caractérisé par un flux continu de matière et d’énergie, un état
éloigné de l’équilibre thermodynamique, l’émergence de motifs ordonnés au niveau supérieur
(patron de succession végétale), le rôle central des boucles de rétroaction (disponibilité des
ressources, eau, lumière et espace), et la description mathématique en terme d’équations non
linéaires. Le modèle représente un écosystème artificiel et les simulations des expériences in
silico. Ces simulations donnent des pistes de réflexion dans l’étude : (1) de la croissance
végétale (paramètres clefs, patrons et modélisation) ; (2) des interactions spécifiques directes
et indirectes (succession secondaire, compétition pour la lumière, compétition pour l’azote et

12
facilitation) ; (3) des services rendus par les écosystèmes, en particulier du rôle de la jachère
dans la restauration de la fertilité et dans la dynamique des cycles du carbone et de l’azote
(durée optimale de la jachère, scénario de gestion, jachère améliorée) ; (4) de la réponse des
écosystèmes aux changements globaux et plus spécifiquement aux changements d’utilisation
des terres (écosystème naturel, culture, jachère, pâturage) ; et (5) des liens entre diversité des
communautés et fonctionnement des écosystèmes (productivité, stabilité et risques liés à
l’extinction d’espèces).
De l’articulation du travail et du plan de la synthèse
La première partie de cette synthèse présentera la démarche que nous avons choisie
pour étudier la succession post-culturale. Nous expliciterons à la fois ce que la modélisation
peut apporter à la compréhension de la restauration de la fertilité et les étapes qui nous ont
conduit à l’élaboration du modèle FAPROM. Les principaux résultats obtenus au cours de la
thèse feront l’objet de la deuxième partie qui s’articulera autour des axes thématiques
suivants : (1) croissance végétale d’un couvert monospécifique, patrons et paramètres clefs ;
(2) succession secondaire, interactions entre espèces et indices de compétition ; (3) rôle de la
jachère dans le système de culture, cycle de l’azote et restauration de la fertilité. La troisième
partie exposera les qualités et les limites du modèle développé, les difficultés rencontrées au
cours du projet et les perspectives de prolongement du travail. Nous reproduisons en annexe
les articles rédigés au cours de la thèse : (1) A process-based model of old-field succession
linking ecosystem and community ecology. Martineau Y, Saugier B. 2004. Cet article
(soumis) présente le modèle FAPROM, décrit son fonctionnement et montre le comportement
général du modèle (succession végétale, sensibilité aux conditions environnementales et aux
conditions initiales, rôle de la jachère dans le cycle de l’azote) ; (2) Evidence of hidden fluxes
in the estimation of Net Primary Production in herbaceous ecosystems. Sarmiento L,
Martineau Y, Acevedo D, Couteaux MM, Escalona A, Saugier B. 2004. En préparation, cet
article fondé à la fois sur les données mensuelles de biomasse et sur un modèle de production
(variante de FAPROM), met en évidence qu’une part importante des flux de carbone n’est pas
prise en compte dans l’estimation de la production primaire nette obtenue à partir du seul
suivi des biomasses ; (3) Uses and abuses of logistic growth: considering turnover to avoid
biological nonsense in ecological modelling. Martineau Y, Deredec A, Genton B, Saugier B.
2004. Ce projet d’article met en garde contre l’utilisation abusive de l’équation logistique
dans la modélisation des dynamiques écologiques et propose une réécriture du modèle de
Smith (1963) obtenue par dégradation du modèle FAPROM. Il insiste sur l’importance du

13
turn-over de la matière et présente un modèle étendu qui peut être utilisé tant pour ajuster des
données que pour modéliser simplement les mécanismes de la croissance dans des systèmes
plus complexes.

14
1 - Développement d’un modèle mécaniste de succession végétale (FAPROM)
15
15
16
16
18
18
19
21
21
21
26
26
27
28
29
29
30
30
30
31
33
34
1.1 - POURQUOI MODELISER ?
1.1.1 - Qu’est-ce qu’un modèle ?
1.1.2 - Le modèle répond à des objectifs scientifiques généraux.
1.1.3 - Le modèle apporte son concours aux études de terrain.
12 - À PARTIR DE QUOI MODELISER ?
1.2.1 - Quel type de modèle est-il adapté à nos objectifs ?
1.2.2 - Le modèle est construit sur des hypothèses simplificatrices.
13 - COMMENT MODELISER ?
1.3.1 - Le modèle conceptuel repose sur les mécanismes écophysiologiques.
Architecture du couvert
Compétition pour la lumière et photosynthèse potentielle
Respiration des plantes
Absorption d’azote et assimilation réalisée
Allocation des assimilats
Sénescence des organes
Matière organique du sol
Cycle reproductif
Pâturage et cueillette
1.3.2 - Le modèle est paramétré à partir d’observations et de mesures de terrain.
1.3.3 - Le modèle informatique a été développé en FORTRAN puis VENSIM.
1.3.4 - Le modèle est utilisé pour estimer la dynamique de l’écosystème.
1.3.5 - Le modèle est couplé à des modèles de sol et de culture.

15
Cette première partie a pour objet de présenter le modèle FAPROM (Fallow
Production Model, Martineau & Saugier 2004, annexe 1) que nous avons développé à partir
de février 2001 (stage de DEA). Notre exposé s’articulera autour de trois questions : (1)
pourquoi modéliser ? (2) à partir de quoi modéliser ? et (3) comment modéliser ?
Le travail de modélisation que nous avons mené consistait à intégrer les mesures de
terrain (caractéristiques des sols, écophysiologie et morphologie des espèces) et prédire
quantitativement la production végétale au cours de la jachère, la dynamique de la litière et la
succession secondaire.
1.1 - POURQUOI MODELISER ? Où nous justifions l’approche de modélisation choisie.
Afin de justifier l’approche développée, nous allons faire le point sur les différents
objectifs de l’étude et montrer dans quelle mesure la modélisation est un moyen de répondre
aux questions scientifiques posées. Nous mettrons ici en exergue : (1) les buts de la
modélisation ; et (2) la contribution des modèles aux études de terrain.
1.1.1 - Qu’est-ce qu’un modèle ?
On entend généralement par modèle ce qui est donné comme exemple, ce qu’on
reproduit par imitation, ou la représentation elle-même de ce qu’on reproduit. En science, un
modèle est une abstraction qui simplifie la réalité en ignorant de nombreuses caractéristiques
du système réel étudié, pour se concentrer sur les aspects qui intéressent le modélisateur et qui
définissent la problématique du modèle (Coquillard & Hill 1997, Davi et al. 2003). Il existe
différents types de modèles scientifiques : les modèles verbaux, conceptuels ; les modèles
physiques (modèles réduits ou agrandis, par exemple, les modèles de molécules en chimie) ;
les modèles mathématiques, analytiques ou informatiques. Le modèle que nous proposons a
pour objectif d’estimer la production végétale au cours de la succession post-culturale ; il
repose sur des équations codées dans un programme informatique ; il entre dans la catégorie
des modèles de simulations.

16
1.1.2 - Le modèle répond à des objectifs scientifiques généraux.
La modélisation est un outil de recherche qui permet de synthétiser l’information
connue et d’en identifier les manques (Daalen & Shugart 1998). Le modèle FAPROM a un
but synthétique : la construction du modèle est l’occasion de rassembler l’information
existante, sur un plan général (mécanismes écophysiologiques, processus de succession,
cycles des nutriments) comme sur les sites d’études particuliers (connaissance du paramo et
de la puna, botanique, écophysiologie des espèces étudiées). Il a aussi un but exploratoire
dans le sens où il révèle l’information qui fait défaut dans notre compréhension du système : il
doit permettre en particulier d’identifier les paramètres clefs de la croissance végétale, les
facteurs qui déterminent la succession secondaire et le rôle de la diversité des communautés
dans le fonctionnement des écosystèmes. Enfin, le modèle comme outil de recherche a un but
explicatif : on cherche par exemple à déterminer les différentes stratégies végétales et à
comprendre la réponse de l’écosystème aux perturbations comme l’impact du pâturage sur la
dynamique de la communauté et la production de l’écosystème.
Calibré et validé, le modèle est également un outil d’ingénierie. Comme outil
diagnostique, il permet d’identifier les variables d’intérêt, ce qu’il importe de mesurer sur le
terrain et ce qui semble être ou ne pas être un bon indicateur de l’état de la parcelle. Dans un
cadre appliqué, il peut servir dans un but prédictif pour estimer les effets à court et long terme
d’une réduction du temps de jachère, pour étudier l’impact de la collecte des graminées et du
bois, et pour quantifier l’effet d’une jachère enrichie par semis de légumineuse. Il pourra
même dans le meilleur des cas se révéler être un outil de gestion qui permette de choisir entre
différentes pratiques culturales, d’évaluer la durée optimale de la jachère et de décider de la
mise en culture et de la mise en jachère des terres.
1.1.3 - Le modèle apporte son concours aux études de terrain.
Sur le terrain, il n’est pas toujours possible de mesurer ce dont on a besoin : (1) La
dynamique de certaines variables d’état est difficile à étudier in situ. L’utilisation de méthodes
destructives dans l’estimation de biomasse empêche d’en suivre la dynamique tandis que les
méthodes non destructives affectent les mesures d’une erreur souvent mal maîtrisée (Sarps
2001). (2) Certains flux ne sont pas directement mesurables ou sont difficilement accessibles.
Par exemple, les méthodes d’estimation de la production primaire nette des écosystèmes

17
uniquement à partir des relevés successifs de végétation dépendent très fortement de la
fréquence des données et de la variance des mesures, et ne sont pas robustes quand la variance
des mesures est trop élevée (cf. Sarmiento et al. 2004, annexe 2). Il est également difficile
d’estimer directement le turn-over des différents organes. La méthode d’analyse inverse
(Savenkoff et al. 2001, Leguerrier et al. 2003) pose un modèle a priori, établit des relations
entre les différents stocks et flux du modèle, et détermine les contraintes du système ; grâce
au principe de parcimonie, elle permet de compenser l’insuffisance des données : le modèle
permet de déduire les flux non mesurés à partir des informations disponibles (flux mesurés et
variables d’état). (3) Les incertitudes de mesure peuvent masquer la dynamique. Comment
déceler les variations temporelles quand la variabilité spatiale est importante ? Dans le cas
particulier de l’étude des variations de stocks de la matière organique du sol, aucune tendance
n’a pu être mise en évidence à cause de la forte hétérogénéité des sols (Sarmiento & Bottner
2002) et du fait que (4) les données de terrains sont coûteuses et partielles. Le suivi de
biomasse de 6 parcelles pendant 2 ans (cf. Sarmiento et al. 2004, annexe 2) aurait occupé une
personne à plein temps pendant 3 ans ! Il est évident qu’on ne peut pas multiplier les mesures
et en cela le modèle peut fournir une aide précieuse pour déterminer sur quelles variables faire
porter les efforts. Un modèle peut également permettre de générer des données utiles au
fonctionnement d’un autre modèle. Au cours des étapes de développement des modèles du
projet TROPANDES, nous avons fourni des données journalières de production de litière au
partenaire Montpelliérain qui développait le modèle de matière organique du sol (Pansu et al.
2004a et b). Réciproquement, nous avons utilisé des données d’évapotranspiration potentielle
fournies par le modèle de bilan hydrique (Metselaar, communication personnelle).
Le modèle constitue un laboratoire in silico. L’augmentation de la vitesse de calcul des
ordinateurs a ouvert un champ d’opportunités aux modélisateurs : (1) Nous sommes capables
d’effectuer un grand nombre de simulations en un temps raisonnable donc on peut tester
théoriquement l’impact de telle variable environnementale sur la production primaire de
l’écosystème, ou bien l’effet de tel paramètre sur la position de telle espèce dans la succession
secondaire. (2) Et nous pouvons évaluer les effets à long terme de nos scénarios (sous les
hypothèses du modèle bien évidemment) ce qui est également infaisable sur le terrain.
Comme système complexe, le modèle peut mettre en évidence des résultats contre intuitifs qui
résultent de combinaisons d’effets : il permet de dépasser les raisonnements conceptuels qui
se limitent à des résultats qualitatifs et principalement aux effets linéaires.

18
1.2 - À PARTIR DE QUOI MODELISER ?
Où nous situons le modèle dans la littérature et considérons le cahier des charges,
les contraintes de réalisation et les hypothèses du modèle.
Notre travail de modélisation a pour but d’estimer la dynamique de la production
végétale et la qualité de la litière produite au cours de la jachère. Aussi nous sommes-nous
intéressés au rôle des espèces tant pour tenir compte des différences de taux de croissance que
des différences de qualité du matériel végétal : le modèle doit quantifier la production
primaire de l’écosystème et reproduire la succession observée. Nous expliquerons dans cette
section quel type de modèle nous avons choisi et pourquoi, en regard des différents types de
modèles développés en écologie ; puis nous présenterons les hypothèses centrales du modèle,
les différents processus écophysiologiques pris en compte et les paramètres et variables qui
lient le modèle à la réalité.
Figure 3. Classification des modèles écologiques (cf. texte). a) types de modèle (Lavigne et al. 2004), b) modèles de succession végétale, c) modèles de production végétale.
1.2.1 - Quel type de modèle est-il adapté à nos objectifs ?
De nombreux types de modèles sont utilisés en écologie. Lavigne et al. (2004) propose
de classer les modèles sur un triangle dont les sommets représentent trois grandes classes
définies suivant l’objectif principal du modèle (Figure 3a) : (1) les modèles généraux et
théoriques qui donnent un cadre conceptuel au développement d’une question scientifique ;
(2) les modèles statistiques qui permettent de prédire avec précision dans le domaine étudié ;
(3) les modèles mécanistes utilisés pour comprendre les processus afin d’être en mesure
d’extrapoler les résultats au-delà du domaine d’étude et de pouvoir tester des scénarios et
Modèlesde trouées
Modèlesbiogéochimiques
Modèlesde compétition
type Lotka-Volterra
Modèlesde trouées
Modèlesbiogéochimiques
Modèlesde compétition
type Lotka-Volterra
Modèlesstatistiques
Modèlesmécanistes
Modèlesthéoriques
Modèlesstatistiques
Modèlesmécanistes
Modèlesthéoriques
ANOVA
Castanea
DSSAT
ANOVA
Castanea
DSSAT
a) b) c)

19
évaluer des effets à long terme. Un modèle peut être un compromis entre ces différents
aspects. Appliquée aux modèles de succession, cette classification place près des sommets du
triangle (Figure 3b) : (1) les modèles de compétition purement théoriques (par exemple Blatt
et al. 2001, basé sur les équations de Lotka-Volterra) ; (2) les modèles de trouées purement
statistiques (voir Bugmann 2001 pour une revue des “gap models” utilisés pour simuler les
dynamiques forestières) ; et (3) les modèles biogéochimiques purement mécanistes (dont
s’approchent les modèles de van Oene et al. 1999a et b, Bachelet et al. 2001, voir aussi
Caldwell 1995 pour une revue des modèles agronomiques à deux espèces). Si l’objectif est de
quantifier la production végétale, on trouve près des sommets (Figure 3c) : (1) les modèles
agronomiques génériques mais pas théoriques (Modèles DSSAT, CERES, Matthews &
Stephens 2002) ; (2) les modèles statistiques du type régression linéaire (efficience
photosynthétique) ou ANOVA (lorsque plusieurs variables explicatives sont prises en compte,
telles que l’âge de la jachère, l’espèce dominante, les caractéristiques du sol, etc.) ; et (3) les
modèles mécanistes écophysiologiques (Dufrêne et al. 2004) ou agronomiques (Parton et al.
1983, de Wit 1978). Dans le cadre du projet TROPANDES, on souhaitait (1) tester des
scénarios pour améliorer la gestion du système de culture (semis de lupin en début de jachère,
arrachage des arbustes, exclusion du pâturage) et (2) prédire les effets à long terme du
raccourcissement du temps de jachère. Or pour pouvoir étendre les résultats du modèle au-
delà des observations, le modèle doit être au moins partiellement mécaniste (Reynolds et al.
2001).
1.2.2 - Le modèle est construit sur des hypothèses simplificatrices.
La construction d’un modèle biogéochimique fondé sur la description des mécanismes
écophysiologiques permet de satisfaire simultanément nos deux objectifs : reproduire la
succession secondaire et quantifier la production végétale. Le modèle lie ainsi écologie des
communautés et écologie des écosystèmes : à chaque pas de temps, la composition spécifique
détermine les flux de l’écosystème (par l’estimation des processus écophysiologiques) ; et
inversement, les flux de l’écosystème structurent la communauté (par le biais des interactions
plantes/plantes et des interactions plantes/sol).
Nous avons volontairement insisté sur la description des mécanismes
écophysiologiques et avons réduit le système étudié à une « soupe végétale » dont la seule
dimension spatiale est la structure verticale de la communauté : les espèces (six ont été

20
paramétrées pour chacun des sites étudiés) sont réparties dans le couvert en fonction de leur
taille mais aléatoirement distribuées dans les strates qu’elles occupent. La hauteur des strates
a été fixée à 10 cm dans nos simulations. Les espèces sont en compétition pour la lumière et
l’azote. Chaque espèce est découpée en quatre organes : (1) les feuilles qui déterminent la
photosynthèse brute potentielle ; (2) les tiges qui déterminent la hauteur du couvert ; (3) les
racines qui déterminent l’absorption d’azote ; et (4) les graines qui participent au cycle
reproductif.
Le modèle décrit les cycles du carbone et de l’azote (Figure 4). Il simule au pas de
temps horaire, la photosynthèse ; au pas de temps journalier, les respirations de croissance et
d’entretien, l’allocation des assimilats, l’absorption, la fixation et la remobilisation de l’azote,
la sénescence des tissus et la chute de nécromasse ; et au pas de temps annuel, la dispersion et
la germination des graines. Ce modèle de végétation a été conçu pour être couplé à un modèle
de sol. Dans l’attente du couplage au modèle développé par Pansu et al. (2004a et b), nous
avons développé un petit modèle simplifié du fonctionnement de la matière organique du sol
pour simuler la rétroaction du cycle de l’azote sur la dynamique de la communauté.
B D LS
rg
F
Gs
Rm
C
Pd = (1-rg).Pg
Hs
a
B D LS
rg
F
Gs
Rm
C
Pd = (1-rg).Pg
Hs
a
B D LS
Uatm
FUsoil
R
N
Gs Hs
b
B D LS
Uatm
FUsoil
R
N
Gs Hs
b
aqd: rate of litter decomposition; asd: rate of soil organic matter decomposition; B: biomass; D: dead mass or standing necromass; F: fall of dead mass (litter accumulation); Gs: reproductive growth (seed germination); Hs: seed dispersion; L: litter; Nleached: mineral nitrogen lost by leaching; Nm: soil mineral nitrogen; Pd: daily net assimilation by photosynthesis; Pg: daily gross assimilation by photosynthesis; rg: fraction of Pg devoted to growth respiration; Rm: maintenance respiration; R: short recycling; S: senescence; SOM: soil organic matter; Uatm: atmospheric nitrogen uptake (leguminous only); Usoil: soil nitrogen uptake by roots; β: rate of litter burial; δ: daily nitrogen deposition.
Figure 4. Modélisation des cycles du carbone et de l’azote. a) cycle du carbone dans les plantes ; b) cycle d’azote dans les plantes ; c) cycle d’azote dans le sol.
SOM
NmL
β.L
Usoil
δ
N
aqd.L
Nleachedasd.SOM
F
c
SOM
NmL
β.L
Usoil
δ
N
aqd.L
Nleachedasd.SOM
F
c

21
1.3 - COMMENT MODELISER ? Où nous décrivons le modèle et les étapes de développement.
La description synthétique du modèle fait l’objet de l’article reproduit en annexe 1.
Nous donnerons ici une vue d’ensemble des voies explorées pour la modélisation des
différents processus écophysiologiques et quelques pistes d’amélioration du modèle. Dans la
suite de cette section, nous proposons un résumé des étapes du développement du modèle, de
son élaboration à son utilisation. On se réfèrera au Tableau 4 (paramètres utilisés et valeurs
estimées pour les six espèces vénézuéliennes) et au Tableau 5 (principales équations telles
qu’elles sont utilisées dans la dernière version du modèle). Le numéro des équations de
référence est cité dans le texte précédé de la lettre E.
Les Tableaux 4 et 5 sont reproduits pages suivantes et sur un volant cartonné recto-verso.
1.3.1 - Le modèle conceptuel repose sur les mécanismes écophysiologiques.
La Figure 4 (page précédente) schématise les cycles du carbone et de l’azote dans les
plantes et dans le sol tels qu’ils sont pris en compte dans le modèle FAPROM.
Architecture du couvert
Pour calculer la photosynthèse potentielle de chaque espèce, le modèle estime la
surface foliaire, la hauteur du couvert et la distribution de la surface foliaire dans le couvert.
La biomasse de feuilles (simulée par le modèle au pas de temps journalier) est convertie en
surface foliaire en supposant une masse surfacique de feuille constante au cours du temps
(E19). La surface foliaire est ensuite répartie de façon homogène dans les strates occupées par
l’espèce (E20). La hauteur du couvert est déterminée en fonction de la biomasse aérienne par
une régression linéaire (E18).
On peut substituer à ces hypothèses simples un calcul de la hauteur du couvert et des
fonctions de répartition des feuilles plus réalistes. La première version du modèle permettait
de choisir entre trois distributions ‘théoriques’ : en pyramide, homogène, en parapluie. Elle a
été abandonnée en attendant l’intégration complète des données de terrain. Ces données
pourront être directement utilisées pour simuler les distributions. (Suite du texte p.26)
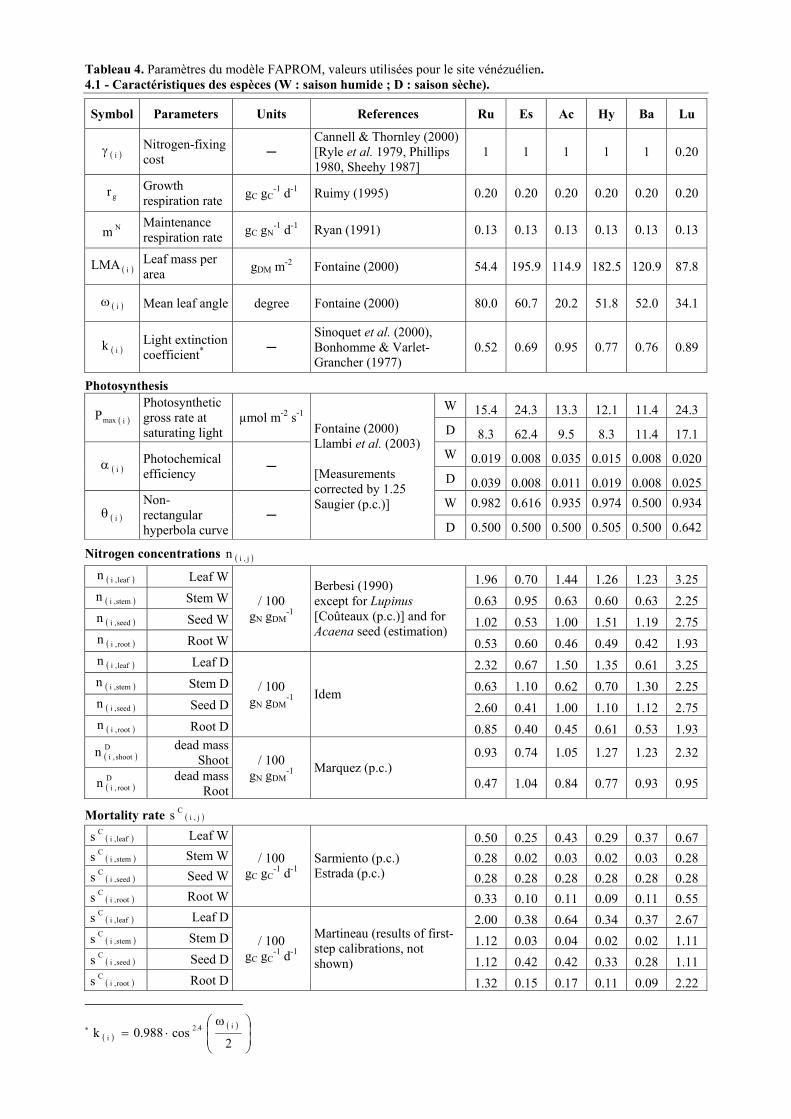
Tableau 4. Paramètres du modèle FAPROM, valeurs utilisées pour le site vénézuélien. 4.1 - Caractéristiques des espèces (W : saison humide ; D : saison sèche).
Symbol Parameters Units References Ru Es Ac Hy Ba Lu
( )iγ Nitrogen-fixing cost
Cannell & Thornley (2000) [Ryle et al. 1979, Phillips 1980, Sheehy 1987]
1 1 1 1 1 0.20
gr Growth respiration rate gC gC
-1 d-1 Ruimy (1995) 0.20 0.20 0.20 0.20 0.20 0.20
Nm Maintenance respiration rate gC gN
-1 d-1 Ryan (1991) 0.13 0.13 0.13 0.13 0.13 0.13
( )iLMA Leaf mass per area gDM m-2 Fontaine (2000) 54.4 195.9 114.9 182.5 120.9 87.8
( )iω Mean leaf angle degree Fontaine (2000) 80.0 60.7 20.2 51.8 52.0 34.1
( )ik Light extinction coefficient*
Sinoquet et al. (2000), Bonhomme & Varlet-Grancher (1977)
0.52 0.69 0.95 0.77 0.76 0.89
Photosynthesis W 15.4 24.3 13.3 12.1 11.4 24.3
( )imaxP Photosynthetic gross rate at saturating light
µmol m-2 s-1 D 8.3 62.4 9.5 8.3 11.4 17.1 W 0.019 0.008 0.035 0.015 0.008 0.020
( )iα Photochemical efficiency
D 0.039 0.008 0.011 0.019 0.008 0.025W 0.982 0.616 0.935 0.974 0.500 0.934
( )iθ Non-rectangular hyperbola curve
Fontaine (2000) Llambi et al. (2003) [Measurements corrected by 1.25 Saugier (p.c.)]
D 0.500 0.500 0.500 0.505 0.500 0.642
Nitrogen concentrations ( )j,in
( )leaf,in Leaf W 1.96 0.70 1.44 1.26 1.23 3.25 ( )stem,in Stem W 0.63 0.95 0.63 0.60 0.63 2.25 ( )seed,in Seed W 1.02 0.53 1.00 1.51 1.19 2.75 ( )root,in Root W
/ 100 gN gDM
-1
Berbesi (1990) except for Lupinus [Coûteaux (p.c.)] and for Acaena seed (estimation)
0.53 0.60 0.46 0.49 0.42 1.93 ( )leaf,in Leaf D 2.32 0.67 1.50 1.35 0.61 3.25 ( )stem,in Stem D 0.63 1.10 0.62 0.70 1.30 2.25 ( )seed,in Seed D 2.60 0.41 1.00 1.10 1.12 2.75 ( )root,in Root D
/ 100 gN gDM
-1 Idem
0.85 0.40 0.45 0.61 0.53 1.93
( )D
shoot,in dead mass Shoot 0.93 0.74 1.05 1.27 1.23 2.32
( )D
root,in dead mass Root
/ 100 gN gDM
-1 Marquez (p.c.) 0.47 1.04 0.84 0.77 0.93 0.95
Mortality rate ( )j,iCs
( )leaf,iCs Leaf W 0.50 0.25 0.43 0.29 0.37 0.67 ( )stem,i
Cs Stem W 0.28 0.02 0.03 0.02 0.03 0.28 ( )seed,i
Cs Seed W 0.28 0.28 0.28 0.28 0.28 0.28 ( )root,i
Cs Root W
/ 100 gC gC
-1 d-1 Sarmiento (p.c.) Estrada (p.c.)
0.33 0.10 0.11 0.09 0.11 0.55 ( )leaf,i
Cs Leaf D 2.00 0.38 0.64 0.34 0.37 2.67 ( )stem,i
Cs Stem D 1.12 0.03 0.04 0.02 0.02 1.11 ( )seed,i
Cs Seed D 1.12 0.42 0.42 0.33 0.28 1.11 ( )root,i
Cs Root D
/ 100 gC gC
-1 d-1
Martineau (results of first-step calibrations, not shown)
1.32 0.15 0.17 0.11 0.09 2.22
* ( )( )
ω⋅=
2cos988.0k
i4.2i
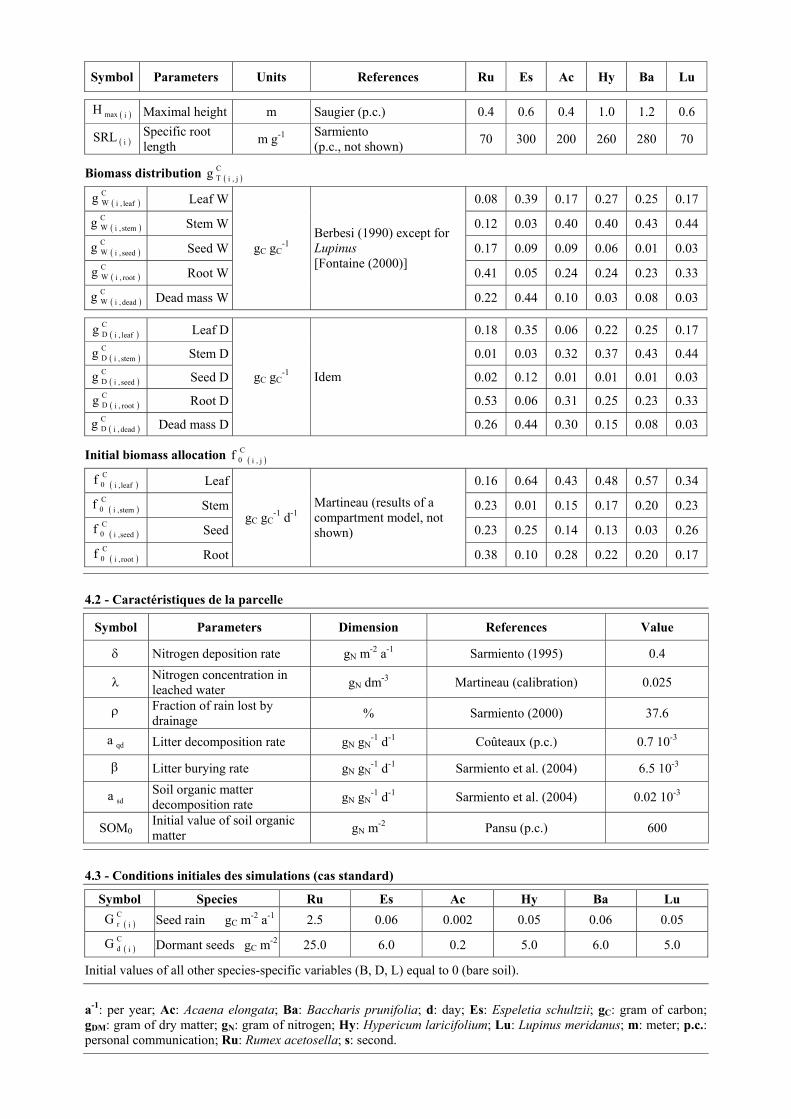
Symbol Parameters Units References Ru Es Ac Hy Ba Lu
( )imaxH Maximal height m Saugier (p.c.) 0.4 0.6 0.4 1.0 1.2 0.6
( )iSRL Specific root length m g-1 Sarmiento
(p.c., not shown) 70 300 200 260 280 70
Biomass distribution ( )j,iCTg
( )leaf,iCWg Leaf W 0.08 0.39 0.17 0.27 0.25 0.17
( )stem,iCWg Stem W 0.12 0.03 0.40 0.40 0.43 0.44
( )seed,iCWg Seed W 0.17 0.09 0.09 0.06 0.01 0.03
( )root,iCWg Root W 0.41 0.05 0.24 0.24 0.23 0.33
( )dead,iCWg Dead mass W
gC gC-1
Berbesi (1990) except for Lupinus [Fontaine (2000)]
0.22 0.44 0.10 0.03 0.08 0.03
( )leaf,iCDg Leaf D 0.18 0.35 0.06 0.22 0.25 0.17
( )stem,iCDg Stem D 0.01 0.03 0.32 0.37 0.43 0.44
( )seed,iCDg Seed D 0.02 0.12 0.01 0.01 0.01 0.03
( )root,iCDg Root D 0.53 0.06 0.31 0.25 0.23 0.33
( )dead,iCDg Dead mass D
gC gC-1 Idem
0.26 0.44 0.30 0.15 0.08 0.03
Initial biomass allocation ( )j,iC0f
( )leaf,iC0f Leaf 0.16 0.64 0.43 0.48 0.57 0.34
( )stem,iC0f Stem 0.23 0.01 0.15 0.17 0.20 0.23
( )seed,iC0f Seed 0.23 0.25 0.14 0.13 0.03 0.26
( )root,iC0f Root
gC gC-1 d-1
Martineau (results of a compartment model, not shown)
0.38 0.10 0.28 0.22 0.20 0.17
4.2 - Caractéristiques de la parcelle
Symbol Parameters Dimension References Value
δ Nitrogen deposition rate gN m-2 a-1 Sarmiento (1995) 0.4
λ Nitrogen concentration in leached water gN dm-3 Martineau (calibration) 0.025
ρ Fraction of rain lost by drainage % Sarmiento (2000) 37.6
qda Litter decomposition rate gN gN-1 d-1 Coûteaux (p.c.) 0.7 10-3
β Litter burying rate gN gN-1 d-1 Sarmiento et al. (2004) 6.5 10-3
sda Soil organic matter decomposition rate gN gN
-1 d-1 Sarmiento et al. (2004) 0.02 10-3
SOM0 Initial value of soil organic matter gN m-2 Pansu (p.c.) 600
4.3 - Conditions initiales des simulations (cas standard)
Symbol Species Ru Es Ac Hy Ba Lu
( )iCrG Seed rain gC m-2 a-1 2.5 0.06 0.002 0.05 0.06 0.05
( )iCdG Dormant seeds gC m-2 25.0 6.0 0.2 5.0 6.0 5.0
Initial values of all other species-specific variables (B, D, L) equal to 0 (bare soil).
a-1: per year; Ac: Acaena elongata; Ba: Baccharis prunifolia; d: day; Es: Espeletia schultzii; gC: gram of carbon; gDM: gram of dry matter; gN: gram of nitrogen; Hy: Hypericum laricifolium; Lu: Lupinus meridanus; m: meter; p.c.: personal communication; Ru: Rumex acetosella; s: second.

Tableau 5. Equations du modèle FAPROM (voir Tableau 4 pour la signification des paramètres et Figure 4 pour le schéma global du fonctionnement du modèle). E1, E2, … refer to equation number as quoted in model description. Description is made for species i, organ j; exponents C and N refer respectively to carbon and nitrogen contents.
t∆ represents the time step. 1t =∆ day for all our simulations.
5.1 - Carbon cycle: daily growth of biomass (B) and standing dead mass (D) [cf. Figure 4-a]. ( ) ( ) ( ) ( )( ) ( ) ( ) ( )j,i
Csj,i
Cj,i
Cmi
Csi
Cdj,i
Cj,i
C HSRGPft/B −−−+⋅=∆∆ E1
( ) ( ) ( )j,iC
j,iC
j,iC FSt/D −=∆∆ E2
( )j,iCf Fraction of carbon allocated to the compartment (for details see in Appendix I, annexe 1) E3
( )iCdP C-uptake: ( ) ( ) ( )( )i
Cmaxi
Ctheoi
Cd U;UminP = E4
( )iCsG Reproductive growth by seed germination
( )j,iCmR Maintenance respiration: ( ) ( )j,i
NNj,i
Cm BmR ⋅= E5
( )j,iCS Senescence: ( ) ( ) ( )j,i
Cj,i
Cj,i
C BsS ⋅= E6
( )j,iCsH Loss by seed dispersion
( )j,iCF Fall of standing dead mass, according to the standing dead/biomass ratio ( )j,i
Cν :
( ) ( ) ( )( )
( )
ν−
ν⋅−= 0;
1BDmaxF
j,iC
j,iC
j,iC
j,iC
j,iC E7
5.2 - Nitrogen cycle: daily growth of biomass (B) and standing dead mass (D) [cf. Figure 4-b].
( ) ( ) ( ) ( ) ( ) ( ) ( ) ( )j,iNsj,i
N
'j'j,i
Nai
Nsoili
Natmi
Nsj,i
Nj,i
N HSUUGft/B −−
ℜ+++⋅=∆∆ ∑ E8
( ) ( ) ( ) ( )j,iN
j,iNaj,i
Nj,i
N FSt/D −ℜ−=∆∆ E9
( )j,iNf Nitrogen allocation coefficient: ( )
( ) ( )
( ) ( )( )∑ ξ
ξ=
'j'j,i'j,i
C
j,ij,iC
j,iN
f
ff E10
( )j,iξ Carbon / nitrogen ratio: ( ) ( )j,ij,i nχ=ξ with carbon content 45.0=χ for all species and organs
( )j,in Nitrogen concentration in biomass
( )iNsG Nitrogen supply by seed rains: ( ) ( ) ( )seed,ii
Csi
Ns GG ξ= E11
( )iNatmU Symbiotic nitrogen fixation
( )iNsoilU Soil nitrogen uptake by plant roots
( )j,iNaℜ Actual nitrogen remobilisation:
( ) ( ) ( ) ( ) ( )( )
( )
ℜ⋅ℜℜ<ℜ=ℜ∑
∑'j 'j,i
Np
iNtheo
j,iNpj,i
Npi
Ntheo
'j 'j,iNpj,i
Na
uelse,then,uif E12
( )j,iNpℜ Potential nitrogen remobilisation: ( ) ( )
( )
( )
( )
( )j,i
j,iCm
necro,i
j,ij,i
Nj,i
Np
R1S
ξ+
ξ
ξ−⋅=ℜ E13
( )j,iNS Senescence: ( ) ( ) ( )j,i
Nj,i
Cj,i
N BsS ⋅= E14
( )j,iNsH Loss by seed dispersion
( )j,iNF Fall of standing dead mass: ( ) ( )
( )
( )j,iC
j,iN
j,iC
j,iN
D
DFF ⋅= E15
5.3 - Carbon and nitrogen uptake
( )iCtheoU Potential daily carbon uptake: ( ) ( ) ( )
⋅η⋅−= ∑∑
l hi
lhgi
Ctheo Pr1U (ηused to change units) E16
gr Growth respiration rate
( )ilhP Instantaneous photosynthesis of leaf layer l at solar time h: E17
( ) ( ) ( ) ( ) ( ) ( ) ( ) ( )( ) ( ) ( ) ( ) ( ) ( )iilhiimaxi
2
ilhiimaxi
lhiimaxili
lh 2AP4APAPLAIP θ⋅
⋅α⋅⋅θ⋅−⋅α+−⋅α+⋅=

Light competition
( )iH Height: ( ) ( ) ( )( )
( )
⋅=
shoot,iCmax
shoot,iC
imaximaxiB
BH;01.0max;HminH E18
with ( ) ( ) ( )stem,iC
leaf,iC
shoot,iC BBB += and ( )shoot,i
CmaxB the maximal value of ( )shoot,i
CB
( )iLAI Leaf area index: ( )( )
( )i
leaf,iC
i LMAB
LAI⋅χ
= with carbon content 45.0=χ E19
( )ilLAI Leaf area index in layer l, linearly related to height (for details see in Appendix II, annexe 1) E20
( )ilhA Light interception: ( )
( )
( ) ( )( ) ( )
⋅−−⋅⋅
⋅= ∑∑ 'i
'il'ilh
'i'il'i
ii
lh LAIkexp1I
LAIk
kA E21
0hI PAR above the canopy at solar time h:
π⋅
τ−⋅= 0;12hcosImaxI max
0h E22
maxI Daily maximal instantaneous radiation:τ⋅Γ
⋅Γ=
I
gUmax
RI E23
with data inputs ( gR : daily radiation and τ : day length) and constants ( UΓ : unit change and IΓ : integration)
1lhI + Instantaneous PAR above the layer l: ( ) ( )
⋅−⋅= ∑+
'i'il'i
lh
1lh LAIkexpII E24
Nitrogen competition
( )iNtheoU Potential nitrogen assimilation: ( ) ( ) ( ) ( )( )∑ ξ⋅=
jj,ij,i
Ci
Ctheoi
Ntheo fUU E25
( )iNtheoV Plant nitrogen demand: ( ) ( ) ( )
ℜ−= ∑ 0;UmaxV
jj,i
Nai
Ntheoi
Ntheo E26
( )iNsoilU Actual plant uptake: ( ) ( )
( ) ( )
( ) ( )
∆
∆⋅
⋅
⋅=
∑ tN
BSRL
BSRL;VminU m
'i
Croot,'i'i
Croot,ii
iNtheoi
Nsoil E27
mN Soil mineral nitrogen [ mm NN =∆ in limiting nitrogen]
( )iNatmU N-fixing: ( ) ( )( ) ( ) ( )( )i
Nsoili
Ntheoii
Natm UV1U −⋅γ−= E28
( )iCmaxU Actual carbon uptake: ( ) ( ) ( ) ( ) ( ) ( )( )∑∑ ⋅ξ⋅
++ℜ='j
'j,iN
'j,i'j
iNsoili
Natm'j,i
Nai
Cmax fUUU E29
5.4 - Seed cycle (cf. Table 4.3).
( )iCsG Seed germination: ( )
( ) ( )
( ) [ ] ( ) ( )( ) ( )[ ] ( )
Ω−⋅⋅δ−+=
==
1H1Gthen365ttif
GthenttifG
seed,iCsii
Crig
iCdi0
iCs E30
with ( )iδ fraction of seed dispersion; ( )igt time of seed germination and ( )i0t time of first germination
Ω Soil cover: ( ) ( )
⋅−−=Ω ∑
'i'i'i LAIkexp1 E31
( )j,iCsH Seed dispersion: ( ) ( ) [ ] ( ) 0else,Bthen,seedjand365ttifH seed,i
Cidj,i
Cs === E32
with ( )idt time of seed dispersion
5.5 - Nitrogen cycling through soil (cf. Table 4.2 and Figure 4-c).
Nsoilleached
Nsd
Nqdm UNSOMaLat/N −−++δ=∆∆ E33
NNqd
NN LLaFt/L β−−=∆∆ E34 N
sdNN SOMaLt/SOM −β=∆∆ E35
leachedN Nitrogen leached: ( )Nsoilmleached UtN,WminN −∆∆λρ= E36
NsoilU Plant uptake: ( )∑=
ii
Nsoil
Nsoil UU E37
NF Litter fall: ( )∑∑=i j
j,iNN FF E38
β, δ, λ, ρ, aqd and asd are defined in Table 4.2.

26
On fait l’hypothèse simplificatrice que dans chaque strate, les feuilles de chaque
espèce sont distribuées aléatoirement dans le plan horizontal et que les distributions des
feuillages de chaque espèce sont indépendantes.
Compétition pour la lumière et photosynthèse potentielle
Sous cette hypothèse, l’extinction de la lumière à travers le couvert végétal suit une loi
de Beer-Lambert (Monsi & Saeki 1953, E24). Dans chaque strate, la lumière absorbée est
répartie entre les espèces au prorata de leur indice local de surface foliaire modulé par leur
coefficient d’absorption (Sinoquet et al. 2000, E21). Pour calculer l’assimilation journalière
potentielle, on intègre sur les strates et sur les heures la réponse instantanée de la
photosynthèse à la lumière mesurée en champ (Thornley & Johnson 1990, E16 à 23). Pour
chaque espèce, deux courbes de réponse instantanée de la photosynthèse foliaire à la lumière
ont été obtenues (Fontaine 2000, Llambi et al. 2003, Saugier & Pontailler 2003) : l’une en
saison humide et l’autre en saison sèche (Gilmanov et al. 2003). Un modèle simplifié de bilan
hydrique détermine quels paramètres utiliser.
Une version antérieure du modèle distinguait lumière directe et lumière diffuse
(Spitters et al. 1986, Spitters 1986, Dufrêne et al. 2004). Malgré l’importance de cette
distinction (Gu et al. 2003 ont montré une augmentation de la photosynthèse consécutive à
l’éruption du Pinatubo), ce raffinement semblait disproportionné par rapport à la description
des autres processus, aussi l’avons-nous abandonné. Par ailleurs, l’application du modèle à
des sites aux saisons moins marquées justifierait d’inclure un véritable bilan hydrique dans le
modèle. Aucune dépendance de la photosynthèse à la température n’a été prise en compte
dans le modèle.
Respiration des plantes
Nous modélisons la respiration en suivant le découpage classique croissance /
entretien (Thornley 1970, Thornley & Cannell 2000, Figure 4a p.20). Nous prélevons d’abord
20% (rg) de l’assimilation photosynthétique pour assurer les besoins liés à la respiration de
croissance (E16). Puis nous prélevons une part proportionnelle au contenu en azote de chaque
organe pour assurer le renouvellement des substrats (Ryan 1991, E5) ; le coefficient de
proportionnalité (mN) est estimé d’après Ryan7. Nous réduisons en outre la respiration
7 ( )
⋅⋅= T
10Qln
exp36527m 10N avec Q10 = 2 et T la température moyenne annuelle du site.

27
d’entretien des feuilles de 25% pour tenir compte de l’utilisation directe de l’énergie
lumineuse par les feuilles pendant la journée (Penning de Vries et al. 1989, Cannell &
Thornley 2000).
Dans le modèle que nous avons construit pour estimer la production primaire nette
des parcelles vénézuéliennes dont la biomasse avait été suivie pendant deux ans (cf. annexe
2), nous prélevons d’abord ce qui correspond à la respiration d’entretien, puis 25% du reste
pour la respiration de croissance. Selon Cannell & Thornley 2000, cette méthode alternative
est équivalente à celle employée pour le modèle FAPROM. Une amélioration substantielle de
l’estimation de la respiration prendrait en compte la quantité de structures et de substrats
dans chaque organe et décrirait le détail des besoins énergétiques liés aux différents
processus écophysiologiques.
Absorption d’azote et assimilation réalisée
Les différentes espèces sont en compétition pour l’azote (Figure 4b p.20). Leur
demande d’azote minéral (dérivée de l’assimilation potentielle, du C/N des organes et des
coefficients d’allocation) est confrontée à l’offre d’azote minéral du sol (E25 à 27). Chaque
espèce absorbe une quantité d’azote minéral au prorata de la longueur de ses racines (modèle
de Berendse, van Oene et al. 1999a). La longueur des racines est calculée à partir de la
biomasse de racines simulée et du SRL (Specific root length : longueur de racine par unité de
masse). Au moment de la sénescence, une partie de l’azote peut-être remobilisée. Cette partie
est calculée à partir des teneurs en azote des différents organes vivants et morts (E12-13). Les
espèces légumineuses ont en outre la possibilité de compléter leur bilan d’azote par fixation
symbiotique (E28). Cette fixation présente un coût respiratoire additionnel : chaque gramme
d’azote fixé nécessite 0,2 g de carbone respiré (Cannell & Thornley 2000, Voisin et al. 2003).
Une version antérieure du modèle répartissait l’azote minéral entre les espèces au
prorata de leur demande d’azote (dérivée de l’assimilation potentielle). Le modèle de
Berendse semble plus adapté pour prendre en compte la compétition pour l’azote. On
pourrait poursuivre l’amélioration en distribuant les racines dans des strates de sol de façon
symétrique au modèle du couvert (mais nous manquons encore de données sur la distribution
des racines par espèce). Quant à la fixation symbiotique des légumineuses, elle était au
départ calculée comme une portion fixe de la demande d’azote. La prise en compte du coût de
la fixation permet une certaine plasticité de la réponse des légumineuses qui semble plus
réaliste : la plante ne fixe de l’azote qu’en conditions limitantes ; en absence d’azote minéral,
la plante a une fixation maximale d’azote.
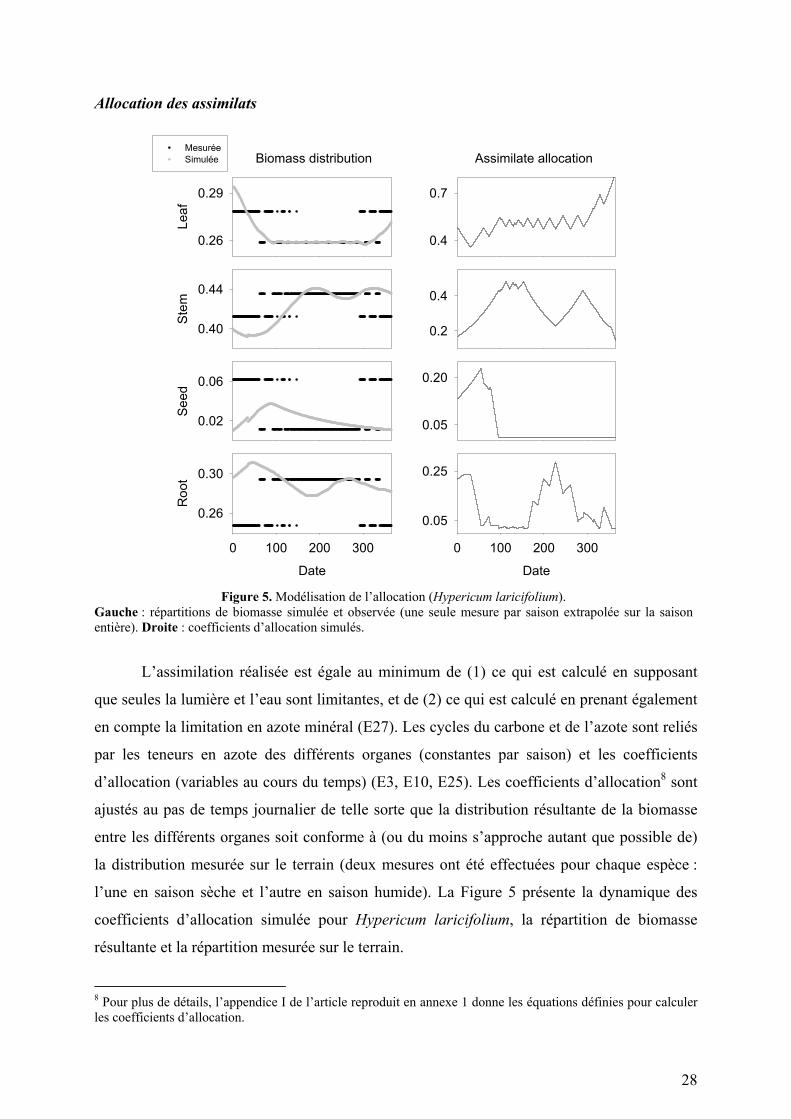
28
Allocation des assimilats
Leaf
0.26
0.29St
em
0.40
0.44
Seed
0.02
0.06
Date
0 100 200 300
Roo
t
0.26
0.30
0.4
0.7
0.2
0.4
0.05
0.20
Date
0 100 200 300
0.05
0.25
Biomass distribution Assimilate allocationMesuréeSimulée
Figure 5. Modélisation de l’allocation (Hypericum laricifolium).
Gauche : répartitions de biomasse simulée et observée (une seule mesure par saison extrapolée sur la saison entière). Droite : coefficients d’allocation simulés.
L’assimilation réalisée est égale au minimum de (1) ce qui est calculé en supposant
que seules la lumière et l’eau sont limitantes, et de (2) ce qui est calculé en prenant également
en compte la limitation en azote minéral (E27). Les cycles du carbone et de l’azote sont reliés
par les teneurs en azote des différents organes (constantes par saison) et les coefficients
d’allocation (variables au cours du temps) (E3, E10, E25). Les coefficients d’allocation8 sont
ajustés au pas de temps journalier de telle sorte que la distribution résultante de la biomasse
entre les différents organes soit conforme à (ou du moins s’approche autant que possible de)
la distribution mesurée sur le terrain (deux mesures ont été effectuées pour chaque espèce :
l’une en saison sèche et l’autre en saison humide). La Figure 5 présente la dynamique des
coefficients d’allocation simulée pour Hypericum laricifolium, la répartition de biomasse
résultante et la répartition mesurée sur le terrain.
8 Pour plus de détails, l’appendice I de l’article reproduit en annexe 1 donne les équations définies pour calculer les coefficients d’allocation.

29
La première version du modèle allouait les assimilats avec des coefficients fixes. Cette
hypothèse ne permettait pas de simuler une répartition de biomasse conforme aux mesures de
terrain. Améliorer la prise en compte des processus d’allocation nécessite vraisemblablement
d’étudier la plasticité de l’allocation (réponse de la plante à un déficit en azote minéral ;
croissance en hauteur stimulée par un déficit de lumière).
Sénescence des organes
La mortalité des tissus est modélisée par une équation du premier ordre : une fraction
constante de biomasse meurt à chaque pas de temps (E6, E14). Un taux de turn-over assure
ainsi le maintien des structures. Ce taux dépend de l’espèce, de l’organe et de la saison.
La prise en compte de la phénologie des espèces permettrait d’améliorer sensiblement
le modèle de sénescence. Noodén et al. (1997) fournissent des pistes pour intégrer l’action de
la sécheresse dans les processus de sénescence : la compensation de la photosynthèse brute
par la respiration d’entretien (due à la réduction de la photosynthèse consécutive à la
fermeture des stomates lors d’un stress hydrique) constituerait un signal des processus de
sénescence (le coût d’entretien des organes verts devient alors prohibitif).
Matière organique du sol
La chute de litière est simulée à partir du rapport de la nécromasse sur pied à la
biomasse, estimées sur les mesures de terrain (E7, E15). La litière au sol est partiellement
décomposée et partiellement enfouie sous l’action des macro-organismes du sol (Figure 4c
p.20). La matière organique du sol (dont une partie est récalcitrante) se décompose à un taux
beaucoup plus lent. Le bilan d’azote minéral (NO3- et NH4
+) tient compte de la minéralisation
de la litière et de la matière organique, de la déposition, du lessivage et de l’absorption par les
racines des plantes (E33 à 38).
Le couplage au modèle multicouche de Pansu & Bottner (2004 a et b) devrait
permettre (1) de prendre en compte la qualité de la litière produite et (2) d’ajuster les taux de
décomposition en fonction des conditions environnementales dans chaque couche (humidité
du sol, température). Il serait également intéressant de modéliser la diversité des micro- et
macro-organismes du sol afin de prendre en compte les boucles de rétroactions qui lient la
diversité des compartiments aériens et souterrains (De Deyn et al. 2004).

30
Cycle reproductif
Parallèlement à la croissance végétative, le modèle simule au pas de temps annuel la
reproduction. A chaque pas de temps les différentes espèces allouent de la biomasse à leurs
structures reproductives. A une date donnée (choisie en fonction de la phénologie de
l’espèce), les graines sont dispersées (E32). La banque de graines du sol est alimentée par une
partie de ces graines, par la pluie de graines qui arrive des parcelles voisines, et par les graines
en dormance (E30). Lorsque les conditions climatiques sont réunies (au début de la saison
humide suivante), une partie de ces graines germent (E31). On entend par germination la
germination proprement dite et la phase de recrutement : la biomasse des graines « qui
germent » est directement allouée aux organes de l’espèce.
La modélisation du cycle reproductif reste simpliste. Le modèle ne prend en compte
que la biomasse de graines ; il ne simule ni le nombre de graines ni le nombre de nouveaux
individus recrutés : il n’y a pas de modèle de dynamique de populations sous-jacent. Des
études complémentaires sur le terrain permettraient d’estimer les flux de graines intra- et
inter-parcelles, la banque de graines, les chances de recrutement, et le rapport de biomasses
graines dispersées / parties reproductives sur pied.
Pâturage et cueillette
Le modèle peut prendre en compte le prélèvement d’une partie de la biomasse d’une
ou de plusieurs espèces. Pour des études préparatoires sur l’impact du pâturage sur la
succession secondaire et sur la production primaire de l’écosystème, nous avons fait varier la
pression d’herbivorie (fréquence et intensité des prélèvements). Pour étudier l’impact de la
cueillette (prélèvement des arbustes), nous avons également fait varier la fréquence de la
perturbation.
Pour prendre correctement en compte le pâturage, il faudrait aussi modéliser l’apport
de matière organique par les fèces des animaux et étudier l’impact mécanique du piétinement
du bétail sur le sol et la végétation.
1.3.2 - Le modèle est construit à partir d’observations et de mesures de terrain.
Le modèle FAPROM a été développé pour prédire la dynamique de la végétation et la
production de litière. Les paramètres du modèle se devaient d’être estimables sinon
mesurables sur le terrain. Le modèle constitue un lien entre les mesures écophysiologiques et

31
la dynamique de la communauté. Bien que nous ayons recherché le meilleur compromis entre
réalisme et simplicité, le nombre de paramètres du modèle est élevé (une cinquantaine par
espèce, cf. Tableau 4). De nombreux auteurs ont publié leur point de vue quant à la
complexité des modèles (notamment Ginzburg & Jensen 2004, Lavigne et al. 2004) mais peu
de ces contributions s’intéressent précisément à ce type de modèle : ici, notre objectif est
clairement de faire le lien entre les processus instantanés (décrits à l’aide de traits spécifiques
mesurables sur le terrain) et la dynamique globale de la communauté (production primaire,
qualité de la litière, succession végétale). Les données interviennent donc à tous les niveaux
du modèle et sont au cœur de notre démarche : (1) les données écophysiologiques et
morphologiques déterminent les paramètres mécanistes de la croissance des différentes
espèces et du fonctionnement de l’écosystème (photosynthèse, distribution de la biomasse,
surface foliaire spécifique, angle foliaire moyen - cf. Tableau 4) ; (2) les données
météorologiques9 (rayonnement journalier, température et précipitations) et les éventuelles
variables exogènes (pluies de graines, flux d’azote minéral) constituent les variables de
forçage du système ; (3) les données écologiques permettent de déterminer les conditions
initiales de nos simulations et de valider le modèle : suivis de parcelle au cours de la jachère
(biovolume des espèces et analyses de sol), expérience de jachère améliorée (semis de
légumineuse), estimation de la production primaire nette de l’écosystème. Malheureusement,
nous n’avons pas encore accès à toutes les données écologiques.
1.3.3 - Le modèle informatique a été développé d’abord en FORTRAN puis sous
VENSIM10.
Conjointement au développement conceptuel du modèle, le développement
informatique constitue une part importante du travail. D’abord codé sous FORTRAN
(Martineau 2001), le modèle a été traduit en 2002 sous VENSIM pour être compatible avec
les autres modèles développés dans le cadre du projet TROPANDES en vue du couplage des
9 Les données climatiques du Venezuela ont été collectées par Lina Sarmiento à Gavidia. Lina Sarmiento et Julia Smith ont traité ces données et étendu le jeu de données disponible sur 34 ans en utilisant des corrélations entre plusieurs stations météorologiques voisines (pour plus de détails, se reporter à la partie Application de l’annexe 1 ou au Matériel & Méthodes de l’annexe 2). Les données climatiques de Bolivie ont été collectées par Jean Vacher à Patacamaya. Pierre Bottner a étendu le jeu de données disponible sur 20 ans en juxtaposant aléatoirement les années météorologiques complètes (1987-1991). 10 VENSIM est un logiciel de programmation graphique des systèmes dynamiques qui fonctionne en C (http://www.vensim.com).

32
modèles à l’échelle de l’écosystème. De nombreuses vérifications sont nécessaires pour
garantir l’adéquation entre le jeu d’équations du modèle conceptuel et le programme
informatique. L’utilisation du modèle sur deux sites différents a permis d’identifier certaines
erreurs et nous espérons qu’après trois années d’utilisation, le programme est complètement
épuré. De nombreuses discussions avec les différents partenaires du projet (en particulier Lina
Sarmiento) et d’autres chercheurs (notamment Frank Berendse, Eric Garnier et Paul Leadley)
ont contribué à l’évolution du modèle. La Figure 6 (ci-dessous) présente les différentes étapes
de développement du modèle : conception, estimation des paramètres, calibration sur les
données manquantes et validation sur des jeux de données indépendants.
Modèle
- Système d’équations qui décrit les mécanismes et les processus
- Traits écophysiologiques et morphologiques mesurés sur chaque espèce
Entrées
- Conditions initiales
- Conditions aux limites
- Conditions extérieures (données météo)
Sorties
- Cycle du carbone et de l’azote
- Dynamique de la communauté
Identification des paramètres
espèce par espèce, calibration des paramètres de
mortalité
Calibrationau niveau de la parcelle sur les
données manquantes (ex.: banque et pluies
de graines)
Validationsur les sorties réfutables (ex.: statut des espèces
dans la succession , biomasse totale,
production de litière)
ParcelleRégion
CommunautéJachère
EspèceEcosystème
Modèle
- Système d’équations qui décrit les mécanismes et les processus
- Traits écophysiologiques et morphologiques mesurés sur chaque espèce
Entrées
- Conditions initiales
- Conditions aux limites
- Conditions extérieures (données météo)
Sorties
- Cycle du carbone et de l’azote
- Dynamique de la communauté
Identification des paramètres
espèce par espèce, calibration des paramètres de
mortalité
Calibrationau niveau de la parcelle sur les
données manquantes (ex.: banque et pluies
de graines)
Validationsur les sorties réfutables (ex.: statut des espèces
dans la succession , biomasse totale,
production de litière)
ParcelleRégion
CommunautéJachère
EspèceEcosystème
Figure 6. Processus de modélisation.
Développer un modèle de ce type ne se limite pas à programmer un certain nombre de
fonctions : il faut également identifier l’ensemble des paramètres. Si la plupart des paramètres
utilisés dans le modèle étaient (ou devaient être) mesurées au cours du projet TROPANDES,
certains paramètres ont (au moins dans un premier temps) dû être calibrés : les paramètres de
mortalité, la banque de graines et les flux de graines. On a traité les espèces les unes après les
autres pour la procédure de calibration des paramètres de mortalité. On a considéré la banque
et les flux de graines comme les conditions initiales et les conditions aux limites des
simulations.
L’idée du projet TROPANDES était de développer un modèle générique qui puisse
être utilisé pour les deux sites d’études. Néanmoins en fonction des données disponibles sur

33
chaque site, il a fallu adapter le modèle au site particulier. Le modèle a été dans un premier
temps développé sur le site vénézuélien. Il a été par la suite adapté au site bolivien en fonction
des informations disponibles : par exemple, les concentrations en azote des différents organes
ont été supposées constantes sur l’année (pas de variations saisonnières).
Le travail sur la Bolivie date d’octobre 2002 et n’a pas été réactualisé depuis. La
version du modèle qui a été utilisée pour les simulations avec les données du site bolivien ne
tenait pas compte des derniers changements (juillet - août 2004) : les coefficients d’allocation
sont supposés constants ; l’azote minéral disponible est une variable exogène (considérée
comme constante) et est absorbé par les racines au prorata des demandes des plantes
(Martineau 2001).
1.3.4 - Le modèle est utilisé pour estimer la dynamique de l’écosystème.
Tableau 6. Variables d’intérêt et sorties réfutables du modèle FAPROM. Données collectées par les partenaires vénézuélien et bolivien.
Gavidia, Venezuela Patarani, Bolivie
Fonctionnement global de l’écosystème
Indice de surface foliaire (LAI) Couverture végétale Suivis de 123 parcelles Suivis de 19 parcelles
Biomasse aérienne Biomasse souterraine Production primaire
Expériences de production (suivi de 6 parcelles pendant 2 ans)
Expériences de production (12 parcelles)
Fixation de l’azote Manipulation de la jachère (semis de lupin)
Manipulation de la jachère (semis de lupin)
Succession végétale
Statut des espèces Dynamique de la biomasse Dynamique du LAI
Suivis de 123 parcelles Suivis de 19 parcelles
Répartition de la biomasse Production de litière
Expériences de production (suivi de 6 parcelles pendant 2 ans)
Expériences de production (12 parcelles)
Une fois la calibration du modèle effectuée se pose la question de la validation. Il
s’agit de vérifier l’adéquation des simulations aux observations de terrain sur un jeu de
données indépendant de ceux utilisés pour la calibration. Dans quelle mesure le modèle est-il
réfutable ? Pour que le modèle soit utile, il est nécessaire que les simulations aient un bon
degré de réalisme, c’est-à-dire que les prédictions soient « suffisamment » proches des
observations. Valider le modèle, c’est vérifier le réalisme d’un certain nombre de variables de

34
sortie du modèle. Le Tableau 6 (p.33) liste les variables d’intérêt et les sorties « réfutables »
du modèle. Pour garantir l’indépendance des jeux de données qui servent à la calibration et à
la validation du modèle, on calibre sur les paramètres écophysiologiques au niveau des
espèces et on valide sur les conditions initiales au niveau de la communauté. Des études
techniques complémentaires accompagnent l’étude du modèle : (1) test de la robustesse du
modèle ; (2) étude de sensibilité du modèle aux conditions initiales ; (3) étude de sensibilité
du modèle aux différents paramètres.
1.3.5 - Le modèle est couplé à des modèles de sol et de culture.
Région
Climat
Exploitation kExploitation 2
Exploitation 1Plot n
Plot 2Plot 1
SAHEL
MOMOS
LINTUL FAPROM
Région
Climat
Exploitation kExploitation 2
Exploitation 1Plot n
Plot 2Plot 1
SAHEL
MOMOS
LINTUL FAPROM
Figure 7. Intégration des modèles du projet TROPANDES (légende : cf. Tableau 7 page suivante).
Un des objectifs du projet TROPANDES était de réaliser un modèle complet du
système de culture à l’échelle de la parcelle (Figure 7). Il s’agissait de : (1) coupler les
modèles de végétation (FAPROM pour la jachère et LINTUL pour la culture, Metselaar,
communication personnelle) aux modèles de sol (MOMOS pour la matière organique du sol,
Pansu et al. 2004a et b, et SAHEL pour le bilan hydrique et l’eau dans le sol, Metselaar,
communication personnelle) ; puis de (2) coupler le modèle de jachère au modèle de culture.
Ces couplages ont nécessité de programmer une plate-forme d’échange des données entre les
différents modèles (les sorties d’un sous-modèle sont les entrées des autres, cf. Tableau 7) et
de modéliser la gestion agronomique de la parcelle (planification des activités agricoles,

35
quantité de semence, d’engrais, etc.). La Figure 8 présente le chronogramme des activités
agricoles.
Tableau 7. Variables échangées entre les différents sous-modèles du modèle agro-écologique de parcelle du projet TROPANDES. (pc : communisation personnelle)
SAHEL
Metselaar, pc
MOMOS Pansu et al. 2004a et b
LINTUL Metselaar, pc
FAPROM Martineau & Saugier 2004
SAHEL Bilan hydrique et eau dans le
sol Humidité du sol ETP
ETR ETP ETR
MOMOS Matière organique du sol Azote minéral Azote minéral
LINTUL Culture de pomme de terre
LAI Couverture
végétale
Fertilisation Résidus de
culture
FAPROM Végétation de la jachère
LAI Couverture
végétale
Production de litière
Biomasse
L’étape suivante serait d’inclure le modèle agro-écologique de la parcelle dans un
modèle de fonctionnement de l’exploitation agricole (PUMANI, Migueis et al. 1998, Modèle
multi-agent des décisions communautaires, Paz-Betancourt 1997) pour simuler la dynamique
du système à l’échelle de la vallée. En effet les décisions des paysans (mise en culture des
terres, mise en jachère, apport d’engrais, semis …) ne dépendent pas seulement des
contraintes agro-écologiques mais également fortement des contraintes socio-économiques
(coût du travail, des semences et des engrais, prix de la récolte, position géographique des
champs, décisions communautaires …).
Figure 8. Chronogramme des activités agricoles.
Légende : S : semis et fertilisation ; R : récolte ; L : labour ; Rompedura : rupture de la jachère, incorporation de la végétation naturelle à la terre.
Jachère Culture A1 Culture A2 Culture A3 Jachère
Rompedura Abandon de la parcelle
R R RPâturageCueillette
PâturageCueillette
S L S L S

36
2 - Applications du modèle, de la plante à l’écosystème
37
37
40
41
43
47
53
55
55
58
60
60
63
65
65
66
66
67
67
69
70
71
71
73
74
75
76
77
2.1 - CROISSANCE D’UN COUVERT VEGETAL MONOSPECIFIQUE
2.1.1 - Patron de croissance végétale, statut de succession et traits des espèces
Patron de croissance et statut de succession
Patron de croissance et traits des espèces (étude de sensibilité I)
Patron de croissance et mécanismes
2.1.2 - Application à l’estimation de la production primaire nette
Que nous apprend le suivi de biomasse des parcelles vénézuéliennes ?
2.2 - DYNAMIQUE DE LA COMMUNAUTE VEGETALE
2.2.1 - Fonctionnement du modèle avec six espèces en interaction
2.2.2 - Le modèle surestime les vitesses de croissance et de mortalité des espèces
2.2.3 - Etudes de sensibilité II
Sensibilité aux conditions initiales
Réponse de l’écosystème au niveau de ressources
2.2.4 - Compétition et coexistence
Sensibilité aux paramètres Hmax et SRL (étude de sensibilité III)
Rôle de la compétition dans le patron de succession
Conditions de coexistence
Modèle mécaniste simplifié
2.2.5 - Tentative de validation du modèle sur les parcelles boliviennes
2.2.6 - Combien d’espèces et quelles espèces faut-il prendre en compte ?
Comment gérer la sensibilité aux conditions initiales ?
2.3 - ROLE DE LA JACHERE DANS L’AGRO-ECOSYSTEME
2.3.1 - Cycle de l’azote au cours de la jachère, potentialité de la jachère améliorée
Quelles sont les fonctions agro-écologiques de la jachère ?
Propositions de gestion durable de l’agro-écosystème
A quelle distance sommes-nous d’un modèle d’aide à la décision ?
2.3.2 - Impact du pâturage
2.3.3 - Diversité des communautés et fonctionnement de l’écosystème

37
Cette deuxième partie présente une synthèse des résultats prédits par le modèle
FAPROM sur trois plans différents : (1) la croissance d’un couvert végétal monospécifique,
(2) la dynamique d’une communauté végétale et (3) le rôle de la jachère dans l’agro-
écosystème. Sauf mention contraire, les simulations ont été effectuées avec les valeurs de
paramètres définies dans le Tableau 4 (données vénézuéliennes).
2.1 - CROISSANCE D’UN COUVERT VEGETAL MONOSPECIFIQUE
Nous présentons dans un premier temps les résultats prédits par le modèle FAPROM
pour les différentes monocultures11. Nous analysons le patron de croissance des différentes
espèces étudiées au regard de la place que ces espèces occupent dans la succession et
interprétons leur patron de croissance en fonction de leurs traits. Afin de mieux comprendre
les mécanismes de la croissance végétale, nous dégradons ensuite le modèle FAPROM. Nous
obtenons un modèle mécaniste simplifié dont nous relions par une étude de sensibilité les
paramètres (qui déterminent le patron de croissance) aux traits mesurés (c’est-à-dire aux
paramètres qui servent à modéliser les espèces dans le modèle FAPROM). Nous exposons
enfin les résultats d’une étude préliminaire à la validation du modèle FAPROM : l’estimation
de la production primaire nette de l’écosystème par suivi de biomasse sur six parcelles
vénézuéliennes, assimilées à des monocultures de Rumex acetosella.
Les résultats concernant la dégradation du modèle FAPROM et l’étude du modèle
mécaniste simplifié font l’objet du projet d’article reproduit en annexe 3 ; l’estimation de la
production primaire nette des six parcelles vénézuéliennes fait l’objet de l’annexe 2.
2.1.1 - Patron de croissance végétale, statut de succession et traits des espèces
Avant de simuler la communauté végétale, nous avons simulé chaque espèce
séparément pour analyser le comportement du modèle pour chacune des monocultures et
calibrer les paramètres de mortalité manquants. La Figure 9 présente l’évolution au cours du
temps de la biomasse des différentes espèces (a - Venezuela, b - Bolivie). Le Tableau 8 (p.39)
donne les valeurs des paramètres utilisés pour la Bolivie. Aux variations saisonnières près, les
11 Nous utiliserons le terme monoculture pour désigner un couvert végétal monospécifique.

38
courbes de croissance suivent un motif sigmoïde caractéristique des croissances logistiques :
un début de croissance exponentielle et une saturation à la capacité de charge. Comment relier
les deux paramètres du modèle logistique12 aux 55 paramètres et 25 équations du modèle
FAPROM ?
Time
0 5 10 15 20
Stipa ichuLupinus buchtieniiBacharis incarum
b) Puna (Bolivia)
Time
0 2 4 6 8 10
Bio
mas
s (g
DM m
-2)
0
200
400
600
800
1000
1200
Rumex acetosellaAcaena elongataLupinus meridanus
Time
0 2 4 6 8 10
Espeletia schultziiHypericum laricifoliumBaccharis prunifolia
a) Paramo (Venezuela)
Time
0 5 10 15 20
Bio
mas
s (g
DM m
-2)
0
200
400
600
800
1000
1200
Aristida asplundiiErodium cicutariumParastrephia lepidophylla
Figure 9. Patron de croissance des 12 espèces paramétrées.
NB : Les échelles temporelles sont différentes (10 ans pour les espèces vénézuéliennes, en haut ; 20 ans pour les espèces boliviennes, en bas). Pour chaque espèce, les paramètres utilisés dans ces simulations correspondent à ceux des Tableaux 4 (Venezuela) et 8 (Bolivie) excepté pour les conditions initiales : 1 gC m-2 de graines en dormance (sauf pour Stipa ichu : 10 gC m-2) ; pas d’apport de graines en provenance de l’environnement au cours de la simulation.
12 L’équation logistique (
−⋅⋅=
Kx1xr
dtdx
, Verhulst 1824, Pearl & Reed 1920) est entièrement
déterminée par 2 paramètres : le taux de croissance intrinsèque de la population (r) et sa capacité de charge (K).
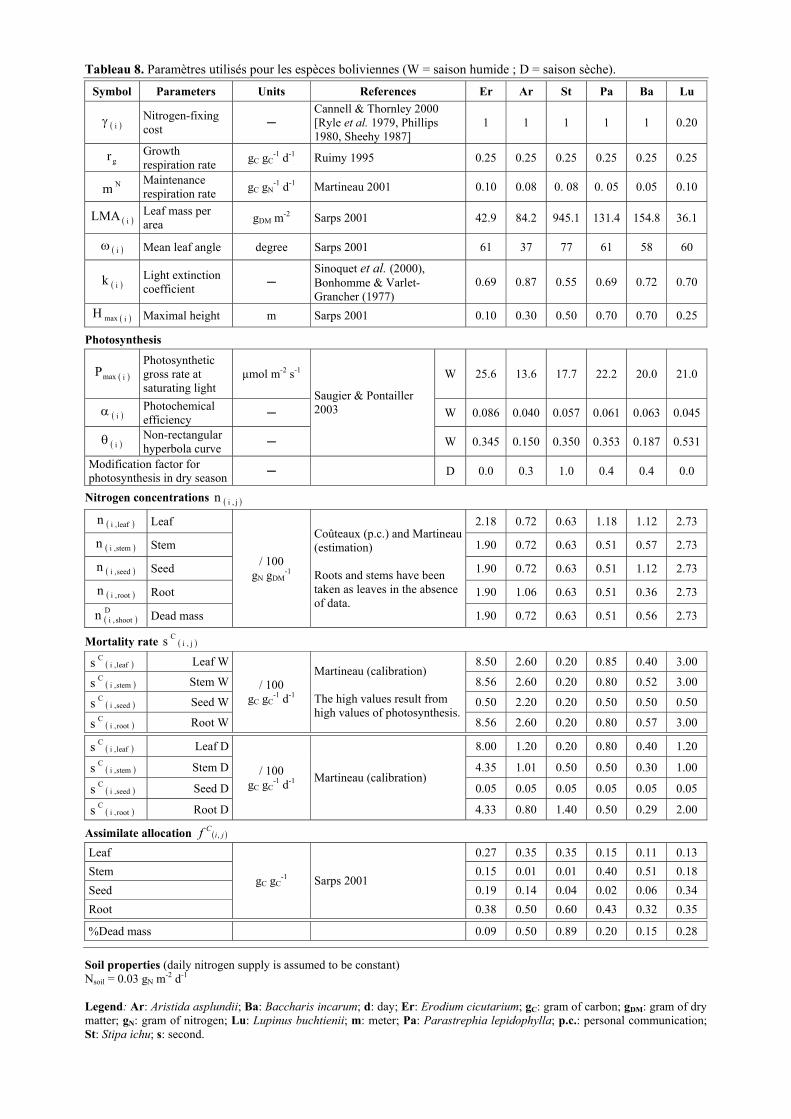
Tableau 8. Paramètres utilisés pour les espèces boliviennes (W = saison humide ; D = saison sèche).
Symbol Parameters Units References Er Ar St Pa Ba Lu
( )iγ Nitrogen-fixing cost
Cannell & Thornley 2000 [Ryle et al. 1979, Phillips 1980, Sheehy 1987]
1 1 1 1 1 0.20
gr Growth respiration rate gC gC
-1 d-1 Ruimy 1995 0.25 0.25 0.25 0.25 0.25 0.25
Nm Maintenance respiration rate gC gN
-1 d-1 Martineau 2001 0.10 0.08 0. 08 0. 05 0.05 0.10
( )iLMA Leaf mass per area gDM m-2 Sarps 2001 42.9 84.2 945.1 131.4 154.8 36.1
( )iω Mean leaf angle degree Sarps 2001 61 37 77 61 58 60
( )ik Light extinction coefficient
Sinoquet et al. (2000), Bonhomme & Varlet-Grancher (1977)
0.69 0.87 0.55 0.69 0.72 0.70
( )imaxH Maximal height m Sarps 2001 0.10 0.30 0.50 0.70 0.70 0.25
Photosynthesis
( )imaxP Photosynthetic gross rate at saturating light
µmol m-2 s-1 W 25.6 13.6 17.7 22.2 20.0 21.0
( )iα Photochemical efficiency W 0.086 0.040 0.057 0.061 0.063 0.045
( )iθ Non-rectangular hyperbola curve
Saugier & Pontailler 2003
W 0.345 0.150 0.350 0.353 0.187 0.531
Modification factor for photosynthesis in dry season D 0.0 0.3 1.0 0.4 0.4 0.0
Nitrogen concentrations ( )j,in
( )leaf,in Leaf 2.18 0.72 0.63 1.18 1.12 2.73
( )stem,in Stem 1.90 0.72 0.63 0.51 0.57 2.73
( )seed,in Seed 1.90 0.72 0.63 0.51 1.12 2.73
( )root,in Root 1.90 1.06 0.63 0.51 0.36 2.73
( )D
shoot,in Dead mass
/ 100 gN gDM
-1
Coûteaux (p.c.) and Martineau (estimation) Roots and stems have been taken as leaves in the absence of data.
1.90 0.72 0.63 0.51 0.56 2.73
Mortality rate ( )j,iCs
( )leaf,iCs Leaf W 8.50 2.60 0.20 0.85 0.40 3.00
( )stem,iCs Stem W 8.56 2.60 0.20 0.80 0.52 3.00
( )seed,iCs Seed W 0.50 2.20 0.20 0.50 0.50 0.50
( )root,iCs Root W
/ 100 gC gC
-1 d-1
Martineau (calibration) The high values result from high values of photosynthesis.
8.56 2.60 0.20 0.80 0.57 3.00
( )leaf,iCs Leaf D 8.00 1.20 0.20 0.80 0.40 1.20
( )stem,iCs Stem D 4.35 1.01 0.50 0.50 0.30 1.00
( )seed,iCs Seed D 0.05 0.05 0.05 0.05 0.05 0.05
( )root,iCs Root D
/ 100 gC gC
-1 d-1 Martineau (calibration)
4.33 0.80 1.40 0.50 0.29 2.00
Assimilate allocation ( )jiCf ,
Leaf 0.27 0.35 0.35 0.15 0.11 0.13 Stem 0.15 0.01 0.01 0.40 0.51 0.18 Seed 0.19 0.14 0.04 0.02 0.06 0.34 Root
gC gC-1 Sarps 2001
0.38 0.50 0.60 0.43 0.32 0.35
%Dead mass 0.09 0.50 0.89 0.20 0.15 0.28
Soil properties (daily nitrogen supply is assumed to be constant) Nsoil = 0.03 gN m-2 d-1 Legend: Ar: Aristida asplundii; Ba: Baccharis incarum; d: day; Er: Erodium cicutarium; gC: gram of carbon; gDM: gram of dry matter; gN: gram of nitrogen; Lu: Lupinus buchtienii; m: meter; Pa: Parastrephia lepidophylla; p.c.: personal communication; St: Stipa ichu; s: second.
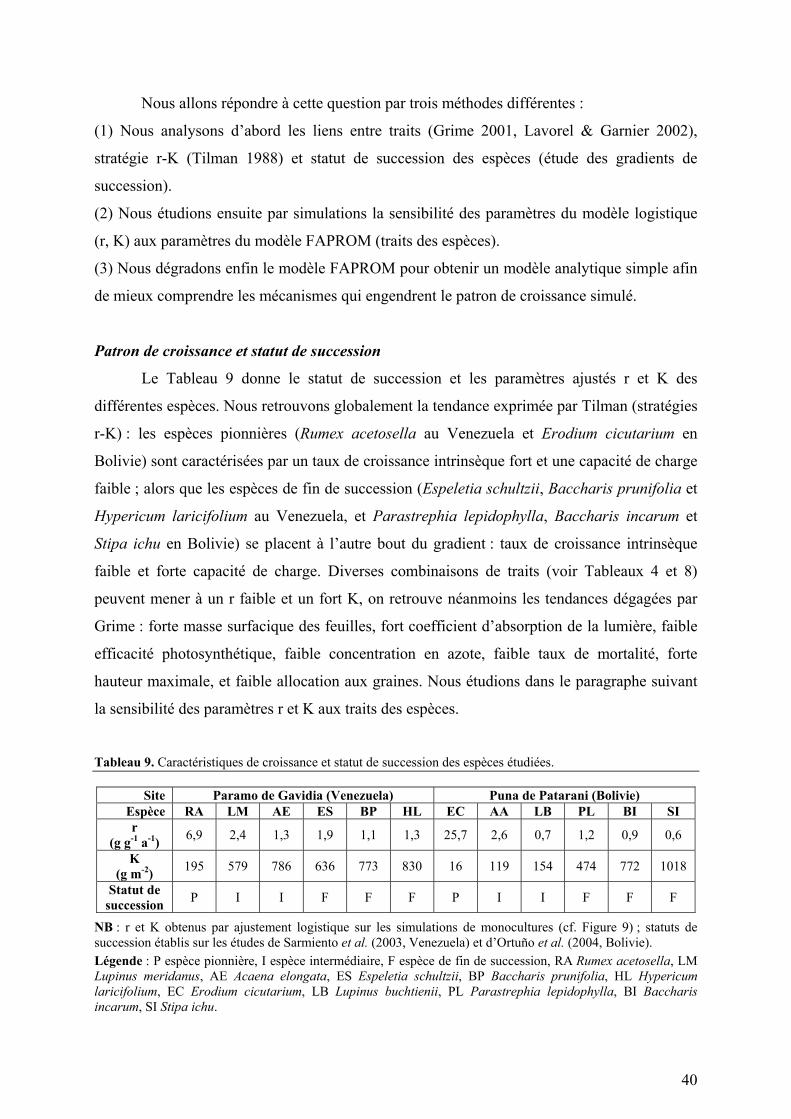
40
Nous allons répondre à cette question par trois méthodes différentes :
(1) Nous analysons d’abord les liens entre traits (Grime 2001, Lavorel & Garnier 2002),
stratégie r-K (Tilman 1988) et statut de succession des espèces (étude des gradients de
succession).
(2) Nous étudions ensuite par simulations la sensibilité des paramètres du modèle logistique
(r, K) aux paramètres du modèle FAPROM (traits des espèces).
(3) Nous dégradons enfin le modèle FAPROM pour obtenir un modèle analytique simple afin
de mieux comprendre les mécanismes qui engendrent le patron de croissance simulé.
Patron de croissance et statut de succession
Le Tableau 9 donne le statut de succession et les paramètres ajustés r et K des
différentes espèces. Nous retrouvons globalement la tendance exprimée par Tilman (stratégies
r-K) : les espèces pionnières (Rumex acetosella au Venezuela et Erodium cicutarium en
Bolivie) sont caractérisées par un taux de croissance intrinsèque fort et une capacité de charge
faible ; alors que les espèces de fin de succession (Espeletia schultzii, Baccharis prunifolia et
Hypericum laricifolium au Venezuela, et Parastrephia lepidophylla, Baccharis incarum et
Stipa ichu en Bolivie) se placent à l’autre bout du gradient : taux de croissance intrinsèque
faible et forte capacité de charge. Diverses combinaisons de traits (voir Tableaux 4 et 8)
peuvent mener à un r faible et un fort K, on retrouve néanmoins les tendances dégagées par
Grime : forte masse surfacique des feuilles, fort coefficient d’absorption de la lumière, faible
efficacité photosynthétique, faible concentration en azote, faible taux de mortalité, forte
hauteur maximale, et faible allocation aux graines. Nous étudions dans le paragraphe suivant
la sensibilité des paramètres r et K aux traits des espèces.
Tableau 9. Caractéristiques de croissance et statut de succession des espèces étudiées.
Site Paramo de Gavidia (Venezuela) Puna de Patarani (Bolivie) Espèce RA LM AE ES BP HL EC AA LB PL BI SI r
(g g-1 a-1) 6,9 2,4 1,3 1,9 1,1 1,3 25,7 2,6 0,7 1,2 0,9 0,6
K (g m-2) 195 579 786 636 773 830 16 119 154 474 772 1018
Statut de succession P I I F F F P I I F F F
NB : r et K obtenus par ajustement logistique sur les simulations de monocultures (cf. Figure 9) ; statuts de succession établis sur les études de Sarmiento et al. (2003, Venezuela) et d’Ortuño et al. (2004, Bolivie).
Légende : P espèce pionnière, I espèce intermédiaire, F espèce de fin de succession, RA Rumex acetosella, LM Lupinus meridanus, AE Acaena elongata, ES Espeletia schultzii, BP Baccharis prunifolia, HL Hypericum laricifolium, EC Erodium cicutarium, LB Lupinus buchtienii, PL Parastrephia lepidophylla, BI Baccharis incarum, SI Stipa ichu.
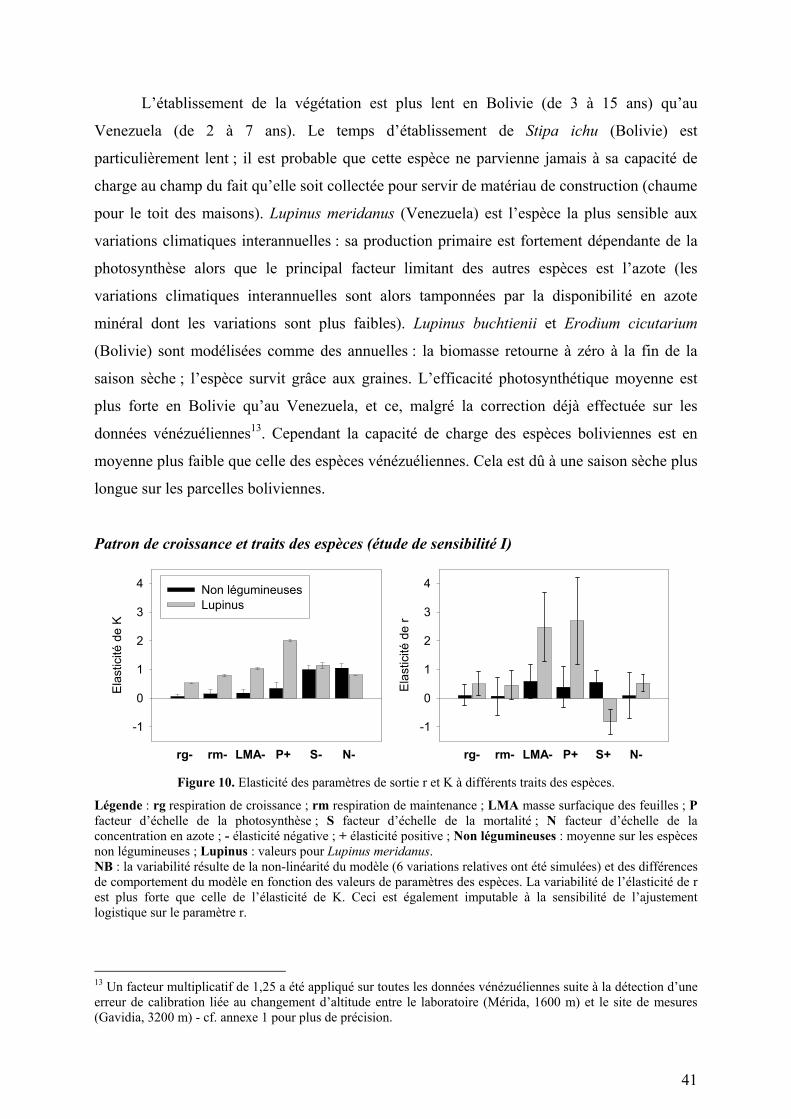
41
L’établissement de la végétation est plus lent en Bolivie (de 3 à 15 ans) qu’au
Venezuela (de 2 à 7 ans). Le temps d’établissement de Stipa ichu (Bolivie) est
particulièrement lent ; il est probable que cette espèce ne parvienne jamais à sa capacité de
charge au champ du fait qu’elle soit collectée pour servir de matériau de construction (chaume
pour le toit des maisons). Lupinus meridanus (Venezuela) est l’espèce la plus sensible aux
variations climatiques interannuelles : sa production primaire est fortement dépendante de la
photosynthèse alors que le principal facteur limitant des autres espèces est l’azote (les
variations climatiques interannuelles sont alors tamponnées par la disponibilité en azote
minéral dont les variations sont plus faibles). Lupinus buchtienii et Erodium cicutarium
(Bolivie) sont modélisées comme des annuelles : la biomasse retourne à zéro à la fin de la
saison sèche ; l’espèce survit grâce aux graines. L’efficacité photosynthétique moyenne est
plus forte en Bolivie qu’au Venezuela, et ce, malgré la correction déjà effectuée sur les
données vénézuéliennes13. Cependant la capacité de charge des espèces boliviennes est en
moyenne plus faible que celle des espèces vénézuéliennes. Cela est dû à une saison sèche plus
longue sur les parcelles boliviennes.
Patron de croissance et traits des espèces (étude de sensibilité I)
rg- rm- LMA- P+ S- N-
Ela
stic
ité d
e K
-1
0
1
2
3
4 Non légumineusesLupinus
rg- rm- LMA- P+ S+ N-
Ela
stic
ité d
e r
-1
0
1
2
3
4
Figure 10. Elasticité des paramètres de sortie r et K à différents traits des espèces.
Légende : rg respiration de croissance ; rm respiration de maintenance ; LMA masse surfacique des feuilles ; P facteur d’échelle de la photosynthèse ; S facteur d’échelle de la mortalité ; N facteur d’échelle de la concentration en azote ; - élasticité négative ; + élasticité positive ; Non légumineuses : moyenne sur les espèces non légumineuses ; Lupinus : valeurs pour Lupinus meridanus. NB : la variabilité résulte de la non-linéarité du modèle (6 variations relatives ont été simulées) et des différences de comportement du modèle en fonction des valeurs de paramètres des espèces. La variabilité de l’élasticité de r est plus forte que celle de l’élasticité de K. Ceci est également imputable à la sensibilité de l’ajustement logistique sur le paramètre r.
13 Un facteur multiplicatif de 1,25 a été appliqué sur toutes les données vénézuéliennes suite à la détection d’une erreur de calibration liée au changement d’altitude entre le laboratoire (Mérida, 1600 m) et le site de mesures (Gavidia, 3200 m) - cf. annexe 1 pour plus de précision.

42
Nous avons étudié la sensibilité des paramètres de sortie r et K au coefficient de
respiration de croissance (rg), au coefficient de respiration d’entretien (mN), à la masse
surfacique des feuilles (LMA), à la capacité photosynthétique de l’espèce (P, facteur
d’échelle14 du rendement quantique et de l’assimilation foliaire maximale), à la concentration
en azote de l’espèce (N, facteur d’échelle des concentrations en azote des différents organes)
et à la sénescence (S, facteur d’échelle des différents taux de mortalité des organes).
L’élasticité15 des sorties r et K aux paramètres rg, mN, LMA, P, N et S a été estimée avec les
données vénézuéliennes en moyennant l’élasticité obtenue pour 6 variations relatives de
chaque paramètre (-15%, -10%, -5%, +5%, +10% et +15%). Nous présentons d’une part les
résultats obtenus avec Lupinus meridanus, d’autre part la moyenne obtenue en utilisant les
paramètres des espèces non légumineuses (Figure 10 p.41). L’élasticité de r et K aux
paramètres liés à la limitation de la croissance par la ressource lumineuse (paramètres de la
photosynthèse, masse foliaire spécifique, coefficients de respiration de croissance et
d’entretien) est en moyenne 6,6 fois plus forte pour l’espèce légumineuse que pour les autres :
la croissance du lupin est fortement limitée par la lumière. Pour les espèces non légumineuses,
le paramètre qui donne la plus forte élasticité de K est la concentration en azote des organes :
leur croissance est fortement limitée par la disponibilité en azote minéral. Le taux de
croissance intrinsèque (r) est plus sensible que la capacité de charge (K) aux paramètres LMA
et P : la lumière est le principal facteur limitant de la phase de croissance exponentielle.
Les paramètres r et K définis sur les simulations de monocultures ne sont pas
sensibles à la hauteur maximale (absence de compétition interspécifique pour la lumière) ni à
la longueur de racine par unité de masse (absence de compétition interspécifique pour
l’azote). En effet, le modèle utilise la hauteur maximale uniquement pour répartir la surface
foliaire dans les strates de couvert. Pour une monoculture, il n’y a pas de compétition
interspécifique et la lumière absorbée dépend uniquement de la surface foliaire totale et non
de sa distribution dans les strates. Par contre, nous verrons que dans le cas d’un mélange
d’espèces, le modèle est sensible à la définition des hauteurs maximales et plus
14 On appelle facteur d’échelle d’un paramètre un coefficient multiplicatif qu’on applique à ce paramètre. Dans la simulation standard (effectuée avec les valeurs de référence), le facteur d’échelle est égal à 1. 15 L’élasticité de la sortie S au paramètre P est définie par analogie avec l’allongement relatif d’un élastique
rapporté à la variation relative de la force qui étire l’élastique, comme étant égale à : ( )PP/
SSPe S
∆∆= .
( ) 0Pe S > si une variation positive du paramètre P entraîne une variation positive de la sortie S.
( ) 1Pe S > si la variation du paramètre P est amplifiée dans la sortie S.

43
particulièrement des hauteurs relatives entre les différentes espèces. En ce qui concerne
l’absorption d’azote, le modèle ne simule pas la capacité d’absorption des différentes espèces
mais uniquement leur capacité relative. Une espèce seule peut (dans le modèle) absorber tout
l’azote minéral disponible quelle que soit sa biomasse de racines.
Patron de croissance et mécanismes
Pourquoi dégrader le modèle FAPROM ?
Nous avons entrepris cette étude théorique avec deux objectifs : mettre à jour les
mécanismes qui aboutissent au patron de croissance logistique et étudier les interactions entre
les différents paramètres. Il est difficile en effet de faire une étude exhaustive du
comportement d’un modèle de simulation au nombre élevé de paramètres. Il est plus simple
d’étudier un modèle analytique au nombre limité de paramètres. L’analyse mathématique
présente l’avantage de permettre d’étudier le comportement du modèle sur des ensembles plus
larges et continus de paramètres. La réduction du nombre de paramètres permet en outre de
visualiser les interactions entre ces différents paramètres et de mieux comprendre les patrons
qui en résultent. Le modèle réduit peut en outre être utilisé comme module d’un modèle plus
complexe.
Figure 11. Dégradation du modèle FAPROM : a) production primaire nette et production de litière en fonction de la biomasse ; b) taux de croissance résultant. Simulations effectuées avec les données d’Espeletia schultzii (Venezuela, Tableau 4), sans limitation d’azote (noir) et en conditions d’azote limitant (gris foncé). La production de litière (Figure 11a, gris clair) ne dépend pas du traitement. La Figure 11b présente en traits épais le taux de croissance annuelle en fonction de la biomasse moyenne annuelle pour les deux traitements, et en traits fins, l’ajustement avec la fonction de Smith (noir, R2 = 0.97) et l’ajustement avec la fonction logistique de Verhulst (gris, R2 = 0.91).
Flux
(gD
M m
-2 a
-1)
0
200
400
600
800
1000
1200
1400
NPP N-stressedNPP N-unlimitedLitter production
Yearly average biomass (gDM m-2)
0 200 400 600 800 1000 1200
Gro
wth
(gD
M m
-2 a
-1)
0
200
400
600
800N-stressedN-unlimitedSmithVerhulst
a
b

44
Dégradation du modèle FAPROM
Nous avons dégradé le modèle FAPROM par simplification des hypothèses (réduction
à un seul compartiment de biomasse, considération d’une unique saison moyenne) et
agrégation de paramètres (production primaire nette / mortalité). Nous discutons dans la suite
les mécanismes et le patron de croissance sur une espèce particulière (Espeletia schultzii). La
Figure 11 (page précédente) décompose le taux de croissance annuel d’Espeletia schultzii
tracé en fonction de sa biomasse moyenne annuelle (11b) en deux composantes : la
production primaire nette annuelle et la production annuelle de litière (11a).
La production primaire nette peut être ajustée par une fonction croissante saturante de
la biomasse (on a choisi une hyperbole, Tableau 10a). La réponse croissante saturante de la
production résulte de l’intégration de la réponse instantanée de la photosynthèse foliaire à la
lumière16. La première partie de la courbe (phase linéaire) résulte de la limitation de la
croissance par le rendement quantique, de la masse surfacique des feuilles et de la fraction de
biomasse consacrée à la surface foliaire. La seconde partie de la courbe (phase de saturation)
résulte de la limitation de la croissance par la ressource la plus limitante : lumière ou azote
minéral (dans l’application à nos sites d’étude, le facteur le plus limitant est l’azote minéral
pour les espèces non légumineuses).
La production de litière peut être ajustée par une fonction linéaire de la biomasse
(Tableau 10a). Le taux de mortalité agrégé au pas de temps annuel, à l’échelle de l’espèce,
résulte des paramètres de sénescence des différents organes et des coefficients d’allocation. Il
intègre les variations saisonnières.
La soustraction de la mortalité à la production donne le taux de croissance en fonction
de la biomasse. La courbe en cloche qui en résulte est responsable du profil logistique obtenu.
La Figure 11b montre également l’ajustement logistique (équation de Verhulst, cf. Tableau
10b, E1). Le caractère asymétrique de la courbe en cloche obtenue avec le modèle FAPROM
diffère de la parfaite symétrie obtenue avec le modèle logistique. Cette asymétrie se traduit
par le glissement du point d’inflexion à une biomasse inférieure à la demi-capacité de charge :
le taux de croissance absolu est maximal pour une biomasse P inférieure à la demi-capacité de
charge. P caractérise le point d’inflexion de la courbe qui représente la biomasse en fonction
du temps. La phase de croissance exponentielle est plus courte que la phase de saturation.
16 Une combinaison linéaire de fonctions hyperboliques saturantes peut être elle-même correctement ajustée par une fonction hyperbolique saturante (résultat non montré). On a choisi une fonction à deux paramètres (hyperbole équilatère) par souci de simplicité.

45
Tableau 10a. Dégradation du modèle FAPROM.
( ) ( )ttdtdx Β−Α=
( )tΑ : daily assimilation
( )tΒ : daily mortality
( )xaAxaAt⋅+⋅⋅≈Α
nN
A soil∝
( ) ( )nm
LMAIfP
r1a Ng ⋅−
⋅⋅−∝
( ) xqt ⋅≈Β Csq ∝
A : maximal assimilation (g d-1) a : marginal rate of assimilation (g g-1 d-1)
soilN : mineral nitrogen available in soil (gN d-1) n : average nitrogen concentration (gN g-1)
gr : rate of growth respiration (dmnl)
P : photosynthetic capacity (g m-2 d-1) ( )If : function of light intensity (dmnl)
LMA : leaf mass per area (g m-2) Nm : coefficient of maintenance respiration (g gN
-1 d-1) q : mortality rate (g g-1 d-1)
Cs : average senescence rate (g g-1 d-1)
NB : Cs représente la moyenne des taux de sénescence de l’espèce pondérée par la biomasse relative des différents organes. dmnl : sans dimension (dimensionless). Tableau 10b. Modèles de croissance et équivalence entre modèles.
E1
−⋅⋅=
Kx1xr
dtdx Verhulst 1838, Pearl & Reed 1924
E2
⋅⋅+
−
⋅⋅=
Kqxr1
Kx1
xrdtdx
Smith 1963
E3 xqxaAxaA
dtdx ⋅−
⋅+⋅⋅= derived from Martineau & Saugier 2004
E2 equivalent to E3 with the following change in variables:
E2→E3 qar −=
−⋅=
a1
q1AK E3→E2 qra +=
( )r
qrqKA
+⋅⋅=
Tableau 10c. Sensibilité du modèle de Smith
Output elasticity to parameters A: maximal assimilation a: marginal rate of
assimilation q: rate of mortality
r: relative growth rate 0
aq
1
1
−
1
qa
1
−−
K: carrying capacity 1 1qa
1
−
aq
1
1
−−
P: biomass for which the absolute growth rate is
maximal 1
aq
1
aq
1
21
−
+⋅
aq
1
121
−
⋅−

46
Réécriture du modèle de Smith (1963)
L’équation logistique a produit de nombreux résultats en écologie, tant dans le
domaine des croissances physiologiques qu’en dynamique des populations. Smith (1963)
revisite le modèle de Verhulst (Tableau 10b, E1) lorsqu’il constate des décalages temporels
dans la dynamique de population des Daphnia magna qu’il a étudié en microcosmes. Il
s’aperçoit que le taux de croissance relatif observé est plus faible que le taux de croissance
relatif attendu avec le modèle logistique. Pour expliquer ces différences, il introduit un
troisième paramètre : le taux de remplacement de la matière par unité de masse par unité de
temps dans la phase de saturation (le taux de turn-over à l’équilibre). Le Tableau 10b montre
l’équivalence entre l’équation obtenue par dégradation du modèle FAPROM (E3, écriture
mécaniste du taux de croissance égal à la différence entre les processus de production et les
processus de sénescence) et l’équation que Smith (E2, 1963) a obtenue en introduisant son
troisième paramètre. Le changement de variables effectué permet de passer des paramètres
mécanistes (taux d’assimilation marginal a, assimilation maximale A et taux de sénescence q)
aux paramètres intégrés (taux de croissance initial r, capacité de charge K et turn-over à
l’équilibre, égal à q). Le Tableau 10c donne les mesures d’élasticité des paramètres intégrés
aux paramètres mécanistes. On retrouve ici certains résultats énoncés précédemment dans
l’étude de sensibilité du modèle FAPROM : l’élasticité de la capacité de charge au taux de
sénescence est égal à 1
aq
1−
−− qui est proche de -1 lorsque q << a. L’intérêt du modèle
analytique tient au fait qu’il formule l’élasticité indépendamment des valeurs particulières
choisies pour l’estimer.
Le modèle de Smith ainsi réécrit peut être étendu (modèle pseudo-logistique). Cette
extension peut être utilisée en dynamique des populations. Comme cette utilisation déborde
du cadre de cette thèse, on trouvera le développement correspondant dans le projet d’article
reproduit en annexe 3.
Intérêt et limites du modèle de Smith et du modèle pseudo-logistique
De nombreux auteurs ont critiqué le modèle logistique pour son incapacité à
modéliser correctement les dynamiques de population (Smith 1963, Fagerström 1987,
Slobodkin 2001) parce qu’il ne prend notamment pas en compte les décalages temporels qui
se traduisent dans l’asymétrie des courbes. De nombreuses équations ont été proposées pour

47
modifier la position du point d’inflexion sans changer le nombre de paramètres (Gompertz17
1825) ou pour la laisser libre an ajoutant un ou plusieurs paramètres supplémentaires (von
Bertalanffy 1938, Zeide 1993, Birch 1999, Tsoularis 2001, Tsoularis & Wallace18 2002). Le
modèle de Smith propose une explication mécaniste aux décalages observés en introduisant le
taux de remplacement de la matière. La réécriture du modèle de Smith permet de séparer de
façon mécaniste les processus de synthèse des processus de destruction.
Nous discutons plus largement dans l’annexe 3 l’intérêt et les limites du modèle
pseudo-logistique, et sa place dans les modèles de dynamique des populations.
Les paramètres du modèle pseudo-logistique dépendent non seulement des
caractéristiques des différentes espèces mais également des conditions du milieu dans lequel
elles poussent. Tel quel, le modèle pseudo-logistique ne permet pas de modéliser la croissance
d’une plante en environnement changeant (notamment au sein d’une communauté végétale).
Pour établir un modèle mécaniste simple de succession végétale, il faut au préalable étudier la
réponse des paramètres pseudo-logistiques aux conditions du milieu (en particulier, la lumière
et l’azote disponible). Nous comptons poursuivre cette étude en discutant de la compétition
interspécifique (cf. deuxième partie et annexe 4).
2.1.2 - Application du modèle FAPROM à l’estimation de la production primaire nette
sur les parcelles vénézuéliennes
Dans le but de calibrer / valider le fonctionnement du modèle sur les monocultures,
nous utilisons les expériences de Sarmiento et al. (2004, annexe 2) pour estimer la production
primaire nette d’une monoculture de Rumex acetosella. La biomasse de racines et la
production souterraine de 2 parcelles (l’une de 0 à 2 ans de jachère, l’autre de 2 à 4 ans de
jachère), et, la biomasse aérienne, la nécromasse sur pied et la litière de 6 parcelles (3 de 0 à 2
17 Le modèle de Gompertz utilise le logarithme népérien pour estimer le taux de croissance de la population :
⋅⋅−=
Kxlnxr
dtdx
L’abscisse du point d’inflexion se déplace en K/e.
18 Le modèle logistique généralisé ajoute trois paramètres puissance au modèle logistique de Verhulst : γβ
α
−⋅⋅=
Kx1xr
dtdx
L’abscisse du point d’inflexion peut parcourir l’intervalle [0 ; K] suivant les
valeurs des paramètres α, β et γ.

48
ans de jachère, et 3 de 2 à 4 ans de jachère) ont été suivies régulièrement pendant deux ans
(respectivement, Figures 12 ci-dessous et 13 p.49).
Figure 12. Phytomasse de racine (a, b) et production souterraine (c, d) mesurée par la méthode des ‘ingrowth core’, Lauenroth 2000) de deux parcelles vénézuéliennes (gauche : parcelle de 0 à 2 ans de jachère ; droite : de 2 à 4 ans).
Nous avons estimé les productions primaires nettes aérienne et souterraine de
l’écosystème à partir de ces suivis de biomasse en utilisant trois méthodes :
- Méthode 1 : on somme uniquement les incréments positifs de matière aérienne entre deux
prélèvements (Sala & Austin 2000) ;
- Méthode 1bis : on ajoutant à cette somme la quantité de litière qui s’est décomposée entre
deux dates (estimée grâce aux expériences des sacs à litière (litterbags), Coûteaux et al.
2004b) ;
- Méthode 2 : on utilise les courbes de réponse instantanée de la photosynthèse foliaire à la
lumière et un modèle (dérivé du modèle FAPROM) pour simuler la photosynthèse
journalière, puis la respiration de croissance et d’entretien, et enfin la répartition des
assimilats entre les compartiments aérien et souterrain.
La méthode 1 (sommation des incréments positifs) présente deux inconvénients
majeurs : (1) elle ne tient pas compte des flux simultanés (production, sénescence,
décomposition) ; (2) elle n’est pas robuste, c’est-à-dire qu’elle est fortement sensible à
Young plot (0-2 years)
Bel
owgr
ound
phy
tom
ass
g m
-2
0
50
100
150
200
250 Plot 2
Bel
owgr
ound
pro
duct
ion
g m
-2 d
-1
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4Plot 2
Time
02/00 08/00 02/01 08/01 02/02
Wet season Time
02/00 08/00 02/01 08/01 02/02
Old plot (2-4 years)
g m
-2
0
50
100
150
200
250Plot 5
g m
-2 d-1
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4Plot 5
c d
ba
Young plot (0-2 years)
Bel
owgr
ound
phy
tom
ass
g m
-2
0
50
100
150
200
250 Plot 2
Bel
owgr
ound
pro
duct
ion
g m
-2 d
-1
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4Plot 2
Time
02/00 08/00 02/01 08/01 02/02
Wet season Time
02/00 08/00 02/01 08/01 02/02
Old plot (2-4 years)
g m
-2
0
50
100
150
200
250Plot 5
g m
-2 d-1
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4Plot 5
c d
ba
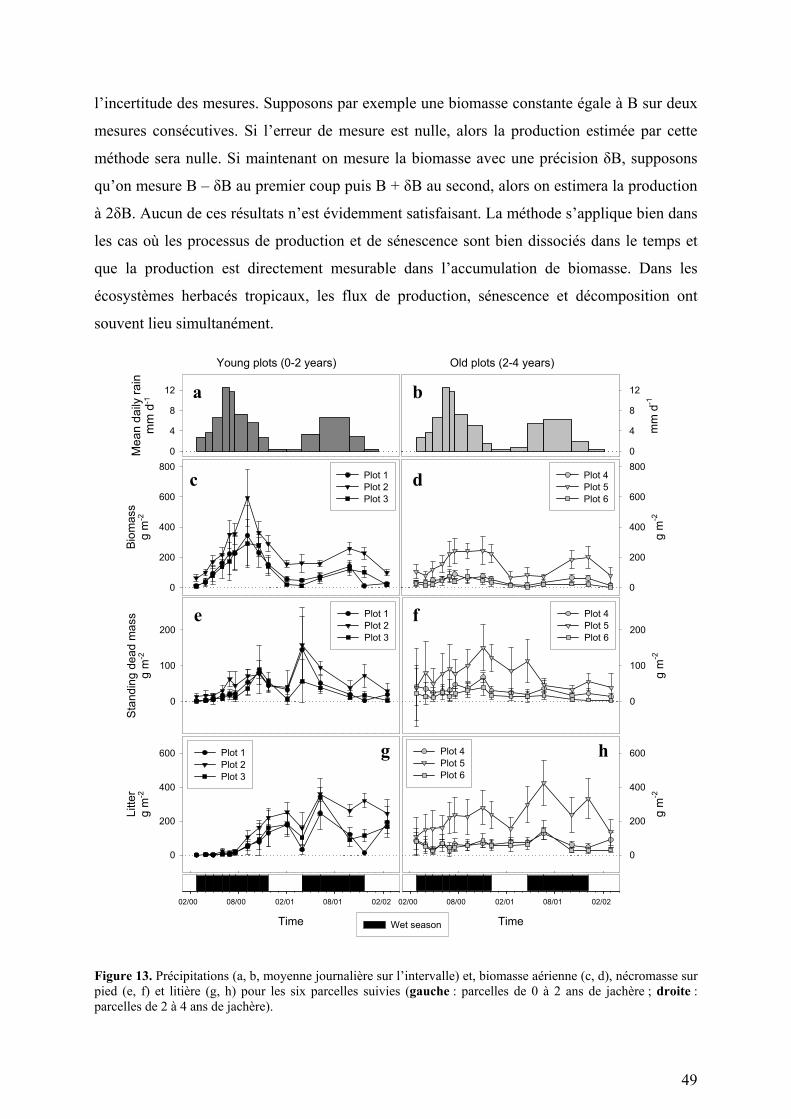
49
l’incertitude des mesures. Supposons par exemple une biomasse constante égale à B sur deux
mesures consécutives. Si l’erreur de mesure est nulle, alors la production estimée par cette
méthode sera nulle. Si maintenant on mesure la biomasse avec une précision δB, supposons
qu’on mesure B – δB au premier coup puis B + δB au second, alors on estimera la production
à 2δB. Aucun de ces résultats n’est évidemment satisfaisant. La méthode s’applique bien dans
les cas où les processus de production et de sénescence sont bien dissociés dans le temps et
que la production est directement mesurable dans l’accumulation de biomasse. Dans les
écosystèmes herbacés tropicaux, les flux de production, sénescence et décomposition ont
souvent lieu simultanément.
Figure 13. Précipitations (a, b, moyenne journalière sur l’intervalle) et, biomasse aérienne (c, d), nécromasse sur pied (e, f) et litière (g, h) pour les six parcelles suivies (gauche : parcelles de 0 à 2 ans de jachère ; droite : parcelles de 2 à 4 ans de jachère).
Young plots (0-2 years)
Mea
n da
ily ra
inm
m d
-1
0
4
8
12
Old plots (2-4 years)
mm
d-1
0
4
8
12
Biom
ass
g m
-2
0
200
400
600
800Plot 1Plot 2Plot 3
g m
-2
0
200
400
600
800Plot 4Plot 5Plot 6
Stan
ding
dea
d m
ass
g m
-2
0
100
200
Plot 1Plot 2Plot 3
g m
-2
0
100
200
Plot 4Plot 5Plot 6
Litte
rg
m-2
0
200
400
600 Plot 1Plot 2Plot 3
g m
-2
0
200
400
600Plot 4Plot 5Plot 6
Time
02/00 08/00 02/01 08/01 02/02
Wet season Time
02/00 08/00 02/01 08/01 02/02
a b
c d
e f
g h
Young plots (0-2 years)
Mea
n da
ily ra
inm
m d
-1
0
4
8
12
Old plots (2-4 years)
mm
d-1
0
4
8
12
Biom
ass
g m
-2
0
200
400
600
800Plot 1Plot 2Plot 3
g m
-2
0
200
400
600
800Plot 4Plot 5Plot 6
Stan
ding
dea
d m
ass
g m
-2
0
100
200
Plot 1Plot 2Plot 3
g m
-2
0
100
200
Plot 4Plot 5Plot 6
Litte
rg
m-2
0
200
400
600 Plot 1Plot 2Plot 3
g m
-2
0
200
400
600Plot 4Plot 5Plot 6
Time
02/00 08/00 02/01 08/01 02/02
Wet season Time
02/00 08/00 02/01 08/01 02/02
a b
c d
e f
g h

50
La méthode 1bis a été développée pour appliquer un correctif à la méthode 1, en
prenant également en compte ce qui se décompose au cours de chaque intervalle entre deux
mesures. Mais le taux de décomposition mesuré au champ ne permet pas d’expliquer la
disparition de litière observée sur le terrain. En effet, pour le compartiment aérien, la
décomposition estimée par le taux des sacs à litière (Tableau 11, colonne A) est environ 15
fois plus faible que les décréments observés (Tableau 11, colonne B). La méthode 1bis
n’apporte donc pas le correctif escompté sur la méthode 1. La différence entre la
décomposition estimée avec l’expérience des sacs à litière et la disparition de biomasse
observée au champ peut être expliquée par différents facteurs parmi lesquels : une sous-
estimation du taux de décomposition dans l’expérience des sacs à litière (modification des
conditions d’humidité, perturbation des interactions trophiques), une disparition de la
biomasse par enfouissement (action de la macrofaune du sol) ou par consommation par les
herbivores.
Pour le compartiment souterrain, le taux de décomposition des sacs à litière semble
compatible avec les décréments (les valeurs de la colonne A sont supérieures à celles de la
colonne B). Ceci suggère que la technique des sacs à litière modifie plus fortement les
conditions du milieu pour la litière aérienne que pour les racines mortes.
Tableau 11. Décomposition et/ou disparition de litière aérienne et souterraine.
Aboveground litter disappearance µ ± σ (g m-2 y-1) A B C
Plot 1 19.0 ± 1.6 450.2 ± 52.0 1342.8 ± 57.7 Plot 2 38.4 ± 1.4 502.2 ± 60.3 2171.4 ± 63.4
Young plots
Plot 3 24.2 ± 1.9 419.7 ± 47.1 1213.5 ± 67.5
Plot 4 14.3 ± 0.8 226.7 ± 25.8 654.0 ± 33.8 Plot 5 50.4 ± 2.9 583.6 ± 73.4 1682.2 ± 80.0
Old plots
Plot 6 12.2 ± 0.9 239.9 ± 32.8 460.4 ± 38.7
Belowground litter disappearance µ ± σ (g m-2 y-1) A B C
Young plot Plot 2 239.3 ± 11.4 94.6 ± 36.8 282.9 ± 34.2
Old plot Plot 5 265.5 ± 11.9 215.6 ± 35.1 561.8 ± 34.4
Légende : A : la décomposition est estimée à partir du taux mesuré avec l’expérience des Sacs à litière (Coûteaux et al. 2004b). B : la disparition de litière est calculée à partir des décréments de matière (Méthode 1, Sala & Austin 2002). C : la disparition de litière est calculée avec le modèle dynamique de production basé sur la photosynthèse, la respiration et l’allocation aux parties souterraines et aériennes (Méthode 2). Young plots : parcelles de 0 à 2 ans de jachère. Old plots : parcelles de 2 à 4 ans de jachère.
La méthode 2 repose sur les délicats changements d’échelle de la feuille au couvert
végétal, et des processus instantanés mesurés au champ, aux processus intégrés sur la journée
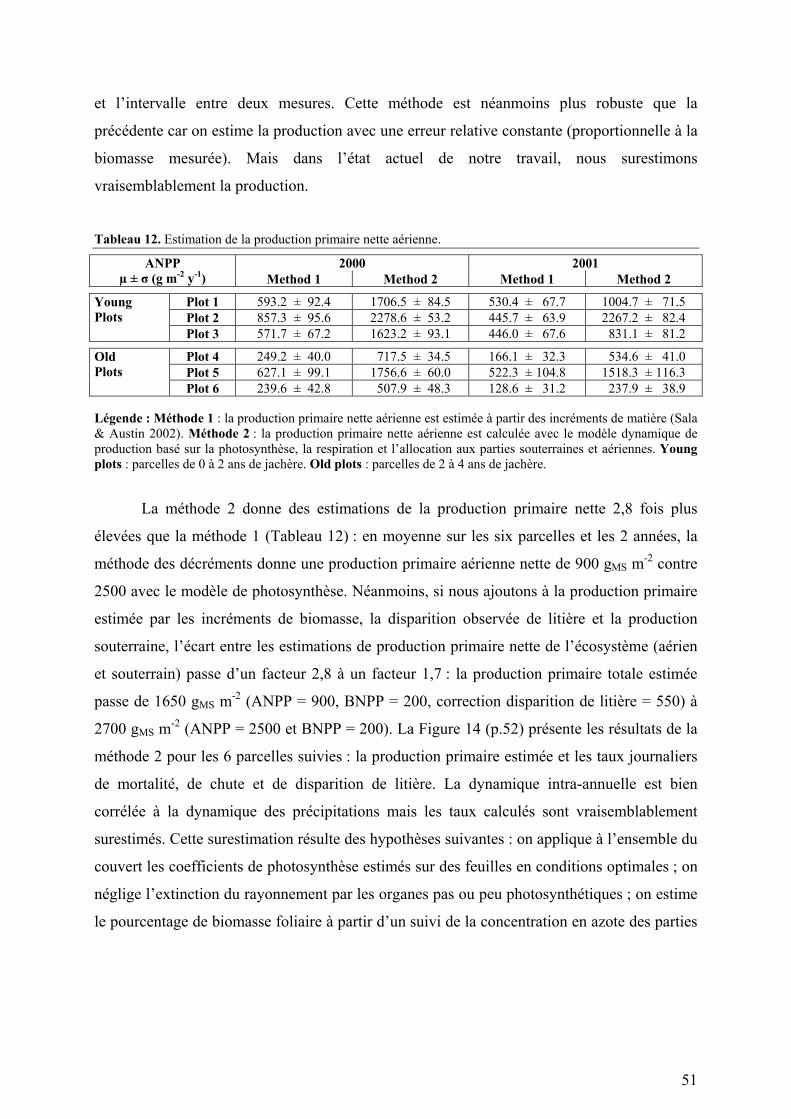
51
et l’intervalle entre deux mesures. Cette méthode est néanmoins plus robuste que la
précédente car on estime la production avec une erreur relative constante (proportionnelle à la
biomasse mesurée). Mais dans l’état actuel de notre travail, nous surestimons
vraisemblablement la production.
Tableau 12. Estimation de la production primaire nette aérienne.
2000 2001 ANPP µ ± σ (g m-2 y-1) Method 1 Method 2 Method 1 Method 2
Plot 1 593.2 ± 92.4 1706.5 ± 84.5 530.4 ± 67.7 1004.7 ± 71.5 Plot 2 857.3 ± 95.6 2278.6 ± 53.2 445.7 ± 63.9 2267.2 ± 82.4
Young Plots
Plot 3 571.7 ± 67.2 1623.2 ± 93.1 446.0 ± 67.6 831.1 ± 81.2
Plot 4 249.2 ± 40.0 717.5 ± 34.5 166.1 ± 32.3 534.6 ± 41.0 Plot 5 627.1 ± 99.1 1756.6 ± 60.0 522.3 ± 104.8 1518.3 ± 116.3
Old Plots
Plot 6 239.6 ± 42.8 507.9 ± 48.3 128.6 ± 31.2 237.9 ± 38.9
Légende : Méthode 1 : la production primaire nette aérienne est estimée à partir des incréments de matière (Sala & Austin 2002). Méthode 2 : la production primaire nette aérienne est calculée avec le modèle dynamique de production basé sur la photosynthèse, la respiration et l’allocation aux parties souterraines et aériennes. Young plots : parcelles de 0 à 2 ans de jachère. Old plots : parcelles de 2 à 4 ans de jachère.
La méthode 2 donne des estimations de la production primaire nette 2,8 fois plus
élevées que la méthode 1 (Tableau 12) : en moyenne sur les six parcelles et les 2 années, la
méthode des décréments donne une production primaire aérienne nette de 900 gMS m-2 contre
2500 avec le modèle de photosynthèse. Néanmoins, si nous ajoutons à la production primaire
estimée par les incréments de biomasse, la disparition observée de litière et la production
souterraine, l’écart entre les estimations de production primaire nette de l’écosystème (aérien
et souterrain) passe d’un facteur 2,8 à un facteur 1,7 : la production primaire totale estimée
passe de 1650 gMS m-2 (ANPP = 900, BNPP = 200, correction disparition de litière = 550) à
2700 gMS m-2 (ANPP = 2500 et BNPP = 200). La Figure 14 (p.52) présente les résultats de la
méthode 2 pour les 6 parcelles suivies : la production primaire estimée et les taux journaliers
de mortalité, de chute et de disparition de litière. La dynamique intra-annuelle est bien
corrélée à la dynamique des précipitations mais les taux calculés sont vraisemblablement
surestimés. Cette surestimation résulte des hypothèses suivantes : on applique à l’ensemble du
couvert les coefficients de photosynthèse estimés sur des feuilles en conditions optimales ; on
néglige l’extinction du rayonnement par les organes pas ou peu photosynthétiques ; on estime
le pourcentage de biomasse foliaire à partir d’un suivi de la concentration en azote des parties
52
aériennes19 (sans contrôle sur le terrain de la répartition de biomasse) ; on ne prend pas
complètement en compte les effets d’une carence en azote ; on utilise un bilan hydrique
simpliste (les paramètres de la saison sèche ont été estimés en début de saison sèche ; l’impact
moyen du stress hydrique sur l’ensemble de la saison sèche est sans doute plus fort). Nous
reviendrons sur ces problèmes dans l’évaluation de la capacité du modèle à reproduire le
patron de succession observé en champ.
Figure 14. Production primaire nette aérienne journalière (a, b), et taux de mortalité (c, d), de chute de litière (e, f) et de disparition (g, h) simulés par le modèle utilisé pour la méthode 2. Young plots : parcelles de 0 à 2 ans de jachère. Old plots : parcelles de 2 à 4 ans de jachère.
19 On suppose constante la teneur en azote des différents organes aériens (tiges et feuilles). On attribue les variations de la teneur en azote moyenne des parties aériennes à des modifications de la distribution de biomasse entre les deux compartiments (cf. Table 2, annexe 2).
Young plots (0-2 years)
Dai
ly A
NPP
g m
-2 d
-1
0
2
4
6
8
10
12 Plot 1Plot 2Plot 3
Mor
talit
y ra
ted-1
0.00
0.02
0.04
0.06
0.08
0.10 Plot 1Plot 2Plot 3
Det
achm
ent r
ate
d-1
0.0
0.2
0.4
0.6
0.8
1.0 Plot 1Plot 2Plot 3
Litte
r dis
appe
aran
ce ra
ted-1
0.0
0.2
0.4
0.6
0.8
1.0 Plot 1Plot 2Plot 3
Old plots (2-4 years)
g m
-2 d
-1
0
2
4
6
8
10
12Plot 4Plot 5Plot 6
d-1
0.00
0.02
0.04
0.06
0.08
0.10Plot 4Plot 5Plot 6
d-1
0.00
0.08
0.16
0.24
0.32
0.40Plot 4Plot 5Plot 6
d-1
0.00
0.03
0.06
0.09
0.12
0.15Plot 4Plot 5Plot 6
Time
02/00 08/00 02/01 08/01 02/02
Wet season Time
02/00 08/00 02/01 08/01 02/02
dc
ba
e f
g h
Young plots (0-2 years)
Dai
ly A
NPP
g m
-2 d
-1
0
2
4
6
8
10
12 Plot 1Plot 2Plot 3
Mor
talit
y ra
ted-1
0.00
0.02
0.04
0.06
0.08
0.10 Plot 1Plot 2Plot 3
Det
achm
ent r
ate
d-1
0.0
0.2
0.4
0.6
0.8
1.0 Plot 1Plot 2Plot 3
Litte
r dis
appe
aran
ce ra
ted-1
0.0
0.2
0.4
0.6
0.8
1.0 Plot 1Plot 2Plot 3
Old plots (2-4 years)
g m
-2 d
-1
0
2
4
6
8
10
12Plot 4Plot 5Plot 6
d-1
0.00
0.02
0.04
0.06
0.08
0.10Plot 4Plot 5Plot 6
d-1
0.00
0.08
0.16
0.24
0.32
0.40Plot 4Plot 5Plot 6
d-1
0.00
0.03
0.06
0.09
0.12
0.15Plot 4Plot 5Plot 6
Time
02/00 08/00 02/01 08/01 02/02
Wet season Time
02/00 08/00 02/01 08/01 02/02
dc
ba
e f
g h

53
Que nous apprend le suivi de biomasse des parcelles vénézuéliennes (annexe 2) ?
Cette étude met en évidence l’existence de flux cachés de l’écosystème (Tableau 13) :
ces flux correspondent à la part équivalente de production qui passe dans le même intervalle
de temps de la biomasse à la nécromasse sur pied par sénescence, de la nécromasse sur pied à
la litière au sol par chute de litière, et qui disparaît par décomposition et/ou disparition de
litière. La méthode traditionnelle des suivis de biomasse, nécromasse et litière est incapable
de détecter ces flux lorsqu’ils ont lieu simultanément. Il semble que ces flux cachés
représentent une part importante de la production primaire dans les écosystèmes herbacés
tropicaux au turn-over rapide et dont la période de production s’étend plus ou moins sur toute
l’année. Avec les hypothèses du modèle, on ne détecte par le suivi de biomasse qu’un tiers
des flux de carbone. Même si la part des flux cachés est surestimée, on peut considérer que
ces flux simultanés ne sont pas négligeables.
Tableau 13. Bilan de carbone de la parcelle.
Young plots (mean) Old plots (mean) Carbon flows g m-2 y-1 % g m-2 y-1 %
GPP 2506 100 1438 100Respiration 781 31 444 31NPP 1724 69 993 69
Aboveground NPP 1618 100 879 100Aboveground measured flows 574 35 322 37Aboveground hidden flows 1044 65 557 63
Légende : GPP production primaire brute (Gross Primary Production). NPP production primaire nette (Net Primary Production). Young plots : parcelles de 0 à 2 ans de jachère. Old plots : parcelles de 2 à 4 ans de jachère. NB : La production primaire nette aérienne a été estimée par la méthode 2 (modèle de photosynthèse). Les flux aériens mesurés sont estimés par la méthode 1 (incréments). Les flux cachés de l’écosystème sont définis comme la différence entre la production primaire nette aérienne et les flux mesurés.
Rumex acetosella représente 98, 93 et 63 % du biovolume20 des parcelles jeunes
étudiées (0-2 ans de jachère). On assimile dans notre étude ces parcelles à des monocultures
de Rumex. Le pic de biomasse mesuré (300-600 g m-2) est plus élevé que la capacité de charge
du Rumex simulée par le modèle FAPROM (100-200 g m-2). Dans FAPROM, nous
surestimons vraisemblablement les taux de mortalité du Rumex et sous-estimons donc la
biomasse. D’autre part, le modèle FAPROM simule mal la dynamique intra-annuelle du
Rumex. Nous surestimons les paramètres de mortalité en saison humide et nous les sous-
20 Le biovolume a été estimé à partir de la méthode des points quadrats (Fontaine 2000, Sarmiento et al. 2002). Il correspond au nombre de contacts entre une tige métallique et l’espèce considérée. Il peut être converti en biomasse moyennant un étalonnage préalable.

54
estimons en saison sèche. La dynamique qui en résulte (Figure 9a p.38) est beaucoup plus
lisse que la dynamique des données de terrain (Figure 13 p.49) Il faudra donc revoir les
paramètres de mortalité utilisés pour Rumex acetosella.
La procédure de calibration devrait ensuite se poursuivre avec le suivi d’autres
parcelles particulières : notamment les parcelles de paramo vierge ou les parcelles anciennes
qui peuvent être assimilées à des monocultures d’Espeletia schultzii ou d’Hypericum
laricifolium ; les parcelles des expériences de jachère améliorée (semées en lupin) peuvent
être considérées comme des monocultures de Lupinus meridanus. Une fois ces calibrations
faites pour les monocultures, la validation du modèle pourra être effectuée en confrontant la
dynamique de la communauté simulée aux suivis diachroniques du biovolume des parcelles
étudiées.

55
2.2 - DYNAMIQUE DE LA COMMUNAUTE VEGETALE
Après avoir identifié les paramètres caractéristiques des espèces à partir des
simulations de monocultures, nous avons étudié le comportement du modèle à l’échelle de la
communauté végétale. Nous présentons d’abord les résultats obtenus sur le site vénézuélien :
dynamique de la végétation, sensibilité aux conditions initiales (banque de graines) et aux
conditions environnementales (ressources disponibles), et étude de la compétition dans un
mélange de deux espèces. Nous présentons ensuite les résultats de validation du modèle pour
la Bolivie.
Les résultats concernant l’application au site vénézuélien font l’objet de l’article
reproduit en annexe 1.
2.2.1 - Fonctionnement du modèle avec six espèces en interaction
La calibration à l’échelle de la communauté consiste à définir les conditions initiales
de la simulation (banque de graines en dormance, teneur en azote organique du sol) et ses
conditions aux limites (pluies annuelles de graines, apports d’azote par déposition et départs
par lessivage). La Figure 15 (p.56) a été obtenue avec les conditions initiales fournies dans le
Tableau 4. Elle présente à la fois les biomasses simulées pour chaque espèce et les données de
biovolume relatif de ces espèces (Sarmiento et al. 2003). La simulation reproduit
correctement le motif de succession (ordre de dominance des différentes espèces) : Rumex
acetosella domine en début de jachère puis sa présence diminue, il disparaît en revanche plus
rapidement dans la simulation que dans les données de terrain (nous discutons ce point
ultérieurement) ; Lupinus meridanus présente un pic de biomasse après quelques années de
jachère ; la rosette géante Espeletia schultzii et les arbustes (Hypericum laricifolium,
Baccharis prunifolia et Acaena elongata) dominent plus tardivement. Des simulations à long
terme montrent que les espèces dominantes en fin de succession dépendent très largement du
niveau de ressource (disponibilité en azote minéral) et du climat (durée de la saison humide).
On détaille ce point dans les études de sensibilité aux ressources.
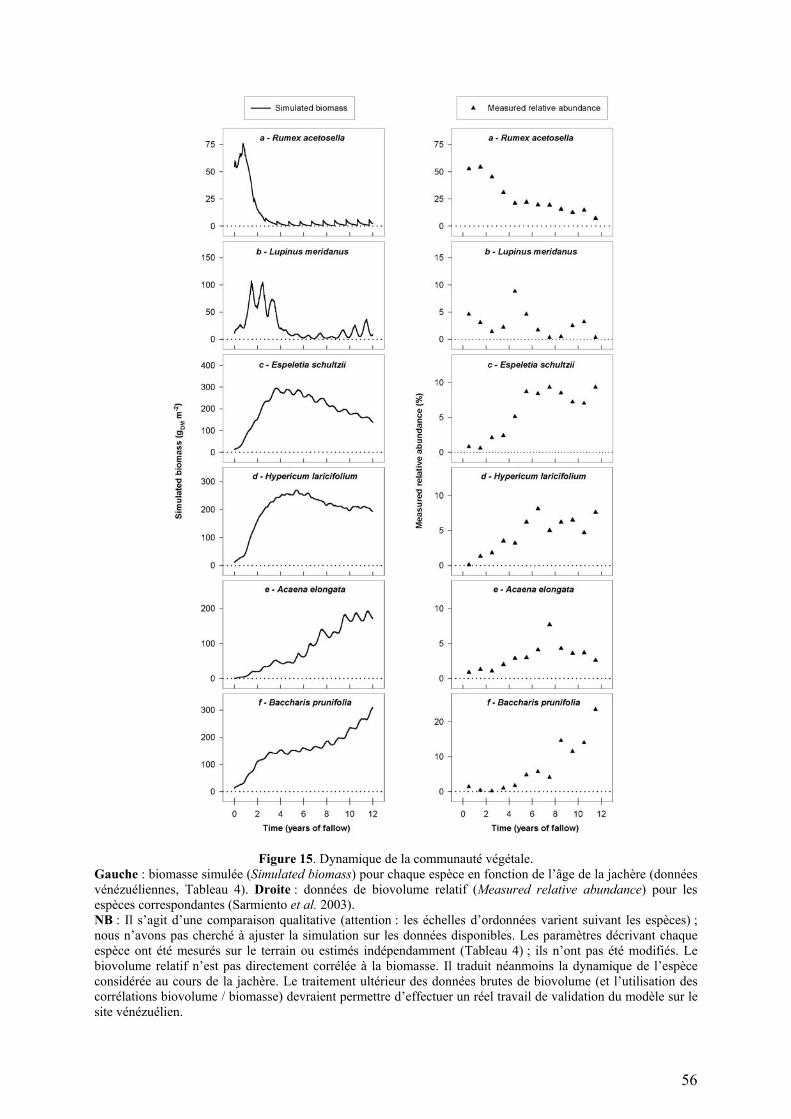
56
Figure 15. Dynamique de la communauté végétale.
Gauche : biomasse simulée (Simulated biomass) pour chaque espèce en fonction de l’âge de la jachère (données vénézuéliennes, Tableau 4). Droite : données de biovolume relatif (Measured relative abundance) pour les espèces correspondantes (Sarmiento et al. 2003). NB : Il s’agit d’une comparaison qualitative (attention : les échelles d’ordonnées varient suivant les espèces) ; nous n’avons pas cherché à ajuster la simulation sur les données disponibles. Les paramètres décrivant chaque espèce ont été mesurés sur le terrain ou estimés indépendamment (Tableau 4) ; ils n’ont pas été modifiés. Le biovolume relatif n’est pas directement corrélée à la biomasse. Il traduit néanmoins la dynamique de l’espèce considérée au cours de la jachère. Le traitement ultérieur des données brutes de biovolume (et l’utilisation des corrélations biovolume / biomasse) devraient permettre d’effectuer un réel travail de validation du modèle sur le site vénézuélien.

57
Figure 16. Variables du fonctionnement de l’écosystème. a) LAI, indice de surface foliaire ; b) productions journalières brutes potentielle et réelle, et nette réelle ; c) biomasse aérienne, nécromasse sur pied et biomasse souterraine. NB : les Figures 15 et 16 correspondent à la même simulation (données vénézuéliennes, paramètres du Tableau 4).
La Figure 16 présente l’indice de surface foliaire total de la communauté (LAI, Figure
16a), les productions brutes potentielle et réelle, et la production nette réelle (Figure 16b),
ainsi que la biomasse totale de la communauté (aérienne, souterraine et nécromasse sur pied,
Figure 16c). On appelle ici production brute potentielle la production limitée seulement par
les caractéristiques photosynthétiques des espèces et la ressource en eau (par l’intermédiaire
de la prise en compte de la saisonnalité de la réponse photosynthétique). La production brute
réelle est inférieure à la production potentielle parce que les divers organes ayant une

58
concentration en azote constante, la demande en azote nécessaire pour satisfaire la croissance
potentielle peut être supérieure à l’offre réelle en azote minéral du sol. Dans ce cas, la
croissance est réduite à celle permise par l’offre en azote. Il y a alors un surplus de carbone
qui n’est pas considéré dans le modèle. Ce surplus de carbone peut être interprété comme une
estimation potentielle de l’exsudation racinaire. La différence entre la production brute et la
production nette correspond à la respiration. En fin de succession, le LAI est environ de 2, la
biomasse totale de l’ordre de 1000 gDM m-2, la production nette journalière moyenne de 0,7 gC
m-2 d-1 et la respiration de 1,4 gC m-2 d-1. Le rapport entre production nette et production brute
est donc faible dans nos simulations (de l’ordre de 30%), mais augmente fortement avec la
disponibilité en azote minéral qui constitue le principal facteur limitant la production.
L’augmentation de la carence azotée conduit à diminuer la part de production brute allouée à
la synthèse de structures et à augmenter la part utilisée pour le turn-over des substrats
(respiration).
2.2.2 - Le modèle surestime les vitesses de croissance et de disparition des espèces.
La comparaison des simulations aux données de terrain met en évidence une
surestimation des vitesses de croissance et de mortalité. Cette surestimation peut résulter de
divers facteurs :
(1) Il peut y avoir une surestimation de la photosynthèse : les courbes de réponse instantanée
de la photosynthèse foliaire à la lumière ont été obtenues sur des feuilles pleinement
développées, en conditions optimales, et sont appliquées à l’ensemble des feuilles, quels que
soient leur statut dans la canopée ou leur âge. D’autre part, la photosynthèse en saison sèche a
été mesurée au début de la saison sèche ; elle diminue sans doute au cours de la saison. De
plus, le modèle ne prend pas en compte l’extinction de la lumière par les organes peu ou pas
photosynthétiques (tiges, nécromasse sur pied, parties reproductives) et donc surestime la
quantité de lumière absorbée par les feuilles. Par ailleurs, le rayonnement global journalier
(donnée climatique d’entrée du modèle) est réparti sur les heures à l’aide d’une fonction
sinusoïdale (cf. Tableau 5, équation E22). On remplace donc un rayonnement horaire variable
(entre le plein soleil et un couvert nuageux dense) par un rayonnement moyen qui tend à
surestimer la photosynthèse. Ainsi dans le modèle Castanea (Dufrêne et al. 2004), la

59
distribution sinusoïdale du rayonnement conduit à une surestimation de la photosynthèse de
l’ordre de 10% (Guerric Le Maire, communication personnelle).
(2) Les paramètres de mortalité ont pu être surestimés lors de la calibration. Les paramètres de
mortalité de la saison humide ont été estimés de façon indépendante à partir des durées de vie
des feuilles et de régressions utilisant la teneur en azote des différents organes (Sarmiento,
communication personnelle). Les paramètres de mortalité de la saison sèche ont été calibrés
de façon à obtenir des biomasses de monocultures conformes à celles attendues. Ils dépendent
donc à la fois de la biomasse totale et de la production ; diverses combinaisons de mortalité et
de production conduisent à la même biomasse. Ce problème de sur-paramétrisation se traduit
ici par une surestimation des vitesses de croissance et de mortalité.
(3) Le modèle ne prend pas en compte les positions respectives des plantes ni l’agrégation des
couverts végétaux. Les feuilles sont réparties au hasard ce qui accroît la compétition
interspécifique en mélangeant des espèces qui ne sont pas directement en contact dans la
réalité. Une solution permettant de limiter ce problème consisterait à utiliser un automate
cellulaire : chaque cellule correspondrait à une sous-parcelle dont on prédit l’évolution grâce
au modèle ; les seules informations transmises d’une cellule à l’autre seraient les flux de
graines. Cette solution pourrait d’abord être essayée sur le site vénézuélien où la densité de
pierres21 favorise la ségrégation spatiale et le développement de la végétation en taches.
(4) Le nombre limité d’espèces prises en compte peut aussi conduire à une surestimation de la
croissance. En effet, moins il y a d’espèces, plus chacune d’elles a d’espace et de ressources
pour se développer. De plus, les effets indirects sont souvent imprévisibles : une espèce rare
peut avoir un rôle d’arbitre dans la compétition entre deux autres espèces. En outre, la
validation pose la question du rôle des espèces considérées et de la représentativité des
groupes fonctionnels : pour le Venezuela (Sarmiento et al. 2003), les espèces prises en
compte représentent d’environ 95% du biovolume des espèces présentes en début de jachère
(principalement Rumex acetosella) à 50% après 12 ans (en particulier Espeletia schultzii,
Hypericum laricifolium et Baccharis prunifolia). Pour pouvoir valider le modèle, il faut
assimiler chaque espèce présente au champ à une espèce paramétrée dans le modèle en
fonction de ses caractéristiques fonctionnelles.
21 Les pierres couvrent jusqu’à 40% de la surface du sol dans certaines parcelles.
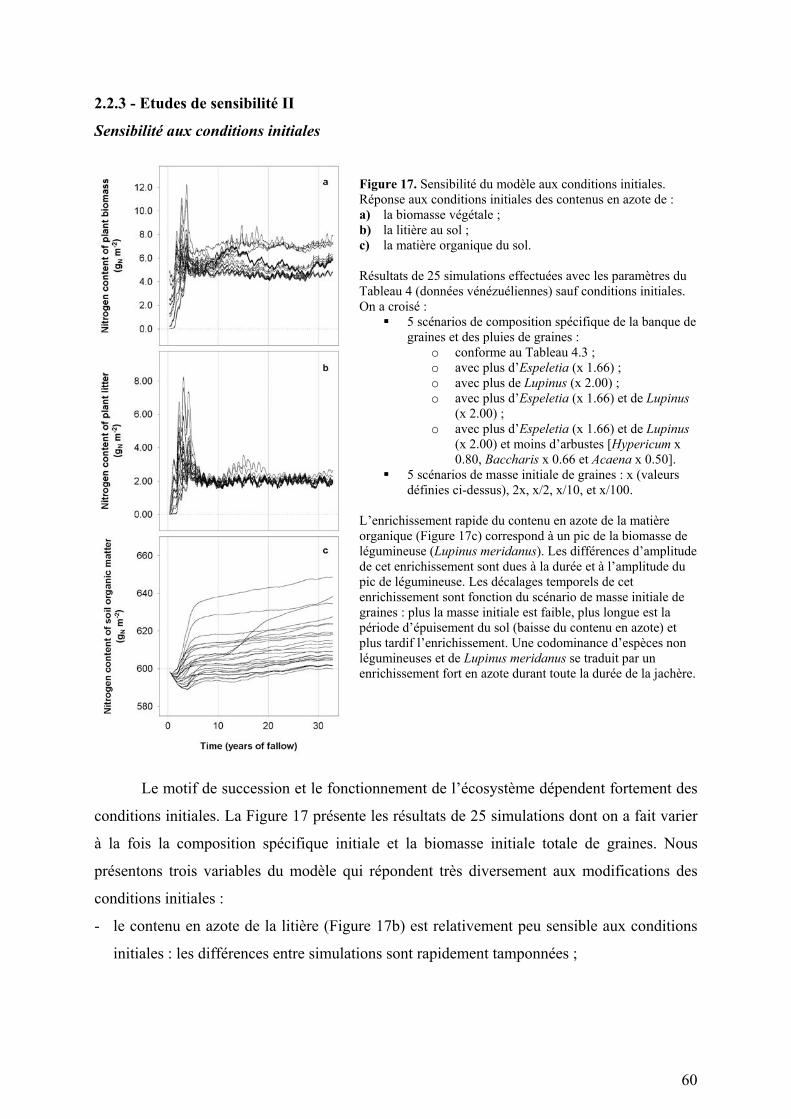
60
2.2.3 - Etudes de sensibilité II
Sensibilité aux conditions initiales
Figure 17. Sensibilité du modèle aux conditions initiales. Réponse aux conditions initiales des contenus en azote de : a) la biomasse végétale ; b) la litière au sol ; c) la matière organique du sol. Résultats de 25 simulations effectuées avec les paramètres du Tableau 4 (données vénézuéliennes) sauf conditions initiales. On a croisé :
5 scénarios de composition spécifique de la banque de graines et des pluies de graines :
o conforme au Tableau 4.3 ; o avec plus d’Espeletia (x 1.66) ; o avec plus de Lupinus (x 2.00) ; o avec plus d’Espeletia (x 1.66) et de Lupinus
(x 2.00) ; o avec plus d’Espeletia (x 1.66) et de Lupinus
(x 2.00) et moins d’arbustes [Hypericum x 0.80, Baccharis x 0.66 et Acaena x 0.50].
5 scénarios de masse initiale de graines : x (valeurs définies ci-dessus), 2x, x/2, x/10, et x/100.
L’enrichissement rapide du contenu en azote de la matière organique (Figure 17c) correspond à un pic de la biomasse de légumineuse (Lupinus meridanus). Les différences d’amplitude de cet enrichissement sont dues à la durée et à l’amplitude du pic de légumineuse. Les décalages temporels de cet enrichissement sont fonction du scénario de masse initiale de graines : plus la masse initiale est faible, plus longue est la période d’épuisement du sol (baisse du contenu en azote) et plus tardif l’enrichissement. Une codominance d’espèces non légumineuses et de Lupinus meridanus se traduit par un enrichissement fort en azote durant toute la durée de la jachère.
Le motif de succession et le fonctionnement de l’écosystème dépendent fortement des
conditions initiales. La Figure 17 présente les résultats de 25 simulations dont on a fait varier
à la fois la composition spécifique initiale et la biomasse initiale totale de graines. Nous
présentons trois variables du modèle qui répondent très diversement aux modifications des
conditions initiales :
- le contenu en azote de la litière (Figure 17b) est relativement peu sensible aux conditions
initiales : les différences entre simulations sont rapidement tamponnées ;

61
- le contenu en azote de la biomasse (Figure 17a) permet de classer les simulations en trois
groupes distincts qui correspondent à la composition en espèces dominantes en fin de
succession :
o le contenu en azote de la biomasse du premier groupe de simulations se
fixe aux alentours de 7,2 gN m-2 ; ces simulations aboutissent à un mélange
des trois espèces arbustives en coexistence (Hypericum laricifolium,
Baccharis prunifolia et Acaena elongata) ;
o le deuxième groupe (contenu en azote autour de 6,0 gN m-2) réunit les
simulations qui aboutissent à une dominance forte d’Acaena elongata en
coexistence avec Hypericum laricifolium ;
o le troisième groupe (contenu en azote autour de 4,9 gN m-2) correspond à
une dominance forte d’Espeletia schultzii en coexistence avec Acaena
elongata ;
- le contenu en azote de la matière organique du sol (Figure 17c) est très sensible aux
conditions initiales : la dynamique de ce compartiment est beaucoup plus lente ; il
accumule les différences successives et fonctionne comme une mémoire de l’histoire de la
parcelle ; les conditions initiales induisent des parcours de succession très différents et dont
les différences se répercutent fortement dans la teneur en azote de la matière organique du
sol. Toutes les simulations commencent par une diminution du contenu en azote de la
matière organique du sol ; cette diminution correspond à la phase d’établissement de la
végétation ; elle est d’autant plus forte que la banque de graines initiales est pauvre. Les
phases d’augmentation rapide du contenu en azote de la matière organique du sol
correspondent aux pics de biomasse de la légumineuse Lupinus meridanus.
On a également étudié dans un mélange d’Hypericum laricifolium et de Baccharis
prunifolia la sensibilité du recouvrement relatif d’Hypericum laricifolium aux paramètres
suivants : conditions initiales (biomasses de graines d’Hypericum laricifolium et de Baccharis
prunifolia), hauteur maximale et longueur de racine par unité de masse (SRL) d’Hypericum
laricifolium (Figure 18, p.62). Ce qui nous intéresse ici est de constater la discontinuité des
surfaces de dominance d’Hypericum laricifolium (en noir, recouvrement relatif au bout de 20
ans supérieur à 66%), de coexistence (en gris, recouvrement relatif au bout de 20 ans compris
entre 33 et 66%) et de dominance de Baccharis prunifolia (en blanc, recouvrement relatif
d’Hypericum laricifolium au bout de 20 ans inférieur à 33%). Les conditions initiales
parcourent l’intervalle [0 ; 50 gC m-2] de 2 en 2 pour la biomasse des graines en dormance,
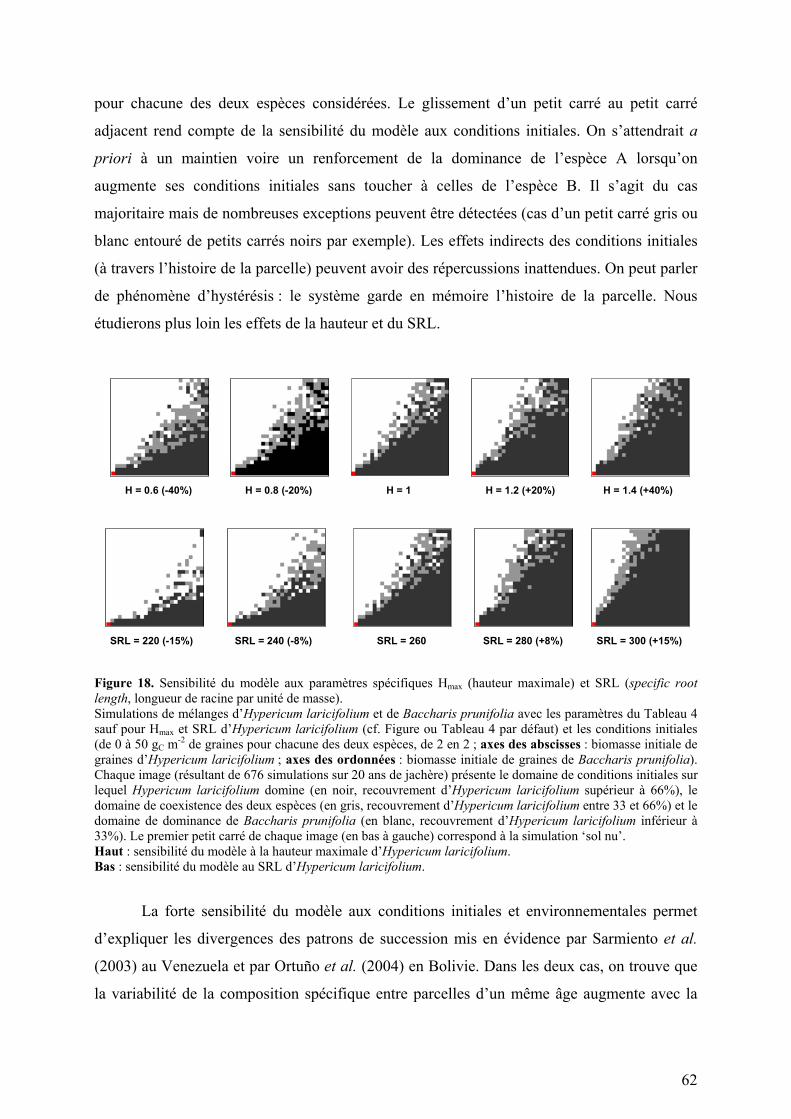
62
pour chacune des deux espèces considérées. Le glissement d’un petit carré au petit carré
adjacent rend compte de la sensibilité du modèle aux conditions initiales. On s’attendrait a
priori à un maintien voire un renforcement de la dominance de l’espèce A lorsqu’on
augmente ses conditions initiales sans toucher à celles de l’espèce B. Il s’agit du cas
majoritaire mais de nombreuses exceptions peuvent être détectées (cas d’un petit carré gris ou
blanc entouré de petits carrés noirs par exemple). Les effets indirects des conditions initiales
(à travers l’histoire de la parcelle) peuvent avoir des répercussions inattendues. On peut parler
de phénomène d’hystérésis : le système garde en mémoire l’histoire de la parcelle. Nous
étudierons plus loin les effets de la hauteur et du SRL.
Figure 18. Sensibilité du modèle aux paramètres spécifiques Hmax (hauteur maximale) et SRL (specific root length, longueur de racine par unité de masse). Simulations de mélanges d’Hypericum laricifolium et de Baccharis prunifolia avec les paramètres du Tableau 4 sauf pour Hmax et SRL d’Hypericum laricifolium (cf. Figure ou Tableau 4 par défaut) et les conditions initiales (de 0 à 50 gC m-2 de graines pour chacune des deux espèces, de 2 en 2 ; axes des abscisses : biomasse initiale de graines d’Hypericum laricifolium ; axes des ordonnées : biomasse initiale de graines de Baccharis prunifolia). Chaque image (résultant de 676 simulations sur 20 ans de jachère) présente le domaine de conditions initiales sur lequel Hypericum laricifolium domine (en noir, recouvrement d’Hypericum laricifolium supérieur à 66%), le domaine de coexistence des deux espèces (en gris, recouvrement d’Hypericum laricifolium entre 33 et 66%) et le domaine de dominance de Baccharis prunifolia (en blanc, recouvrement d’Hypericum laricifolium inférieur à 33%). Le premier petit carré de chaque image (en bas à gauche) correspond à la simulation ‘sol nu’. Haut : sensibilité du modèle à la hauteur maximale d’Hypericum laricifolium. Bas : sensibilité du modèle au SRL d’Hypericum laricifolium.
La forte sensibilité du modèle aux conditions initiales et environnementales permet
d’expliquer les divergences des patrons de succession mis en évidence par Sarmiento et al.
(2003) au Venezuela et par Ortuño et al. (2004) en Bolivie. Dans les deux cas, on trouve que
la variabilité de la composition spécifique entre parcelles d’un même âge augmente avec la
H = 0.6 (-40%) H = 0.8 (-20%)
SRL = 220 (-15%)
H = 1 H = 1.2 (+20%) H = 1.4 (+40%)
SRL = 240 (-8%) SRL = 260 SRL = 280 (+8%) SRL = 300 (+15%)
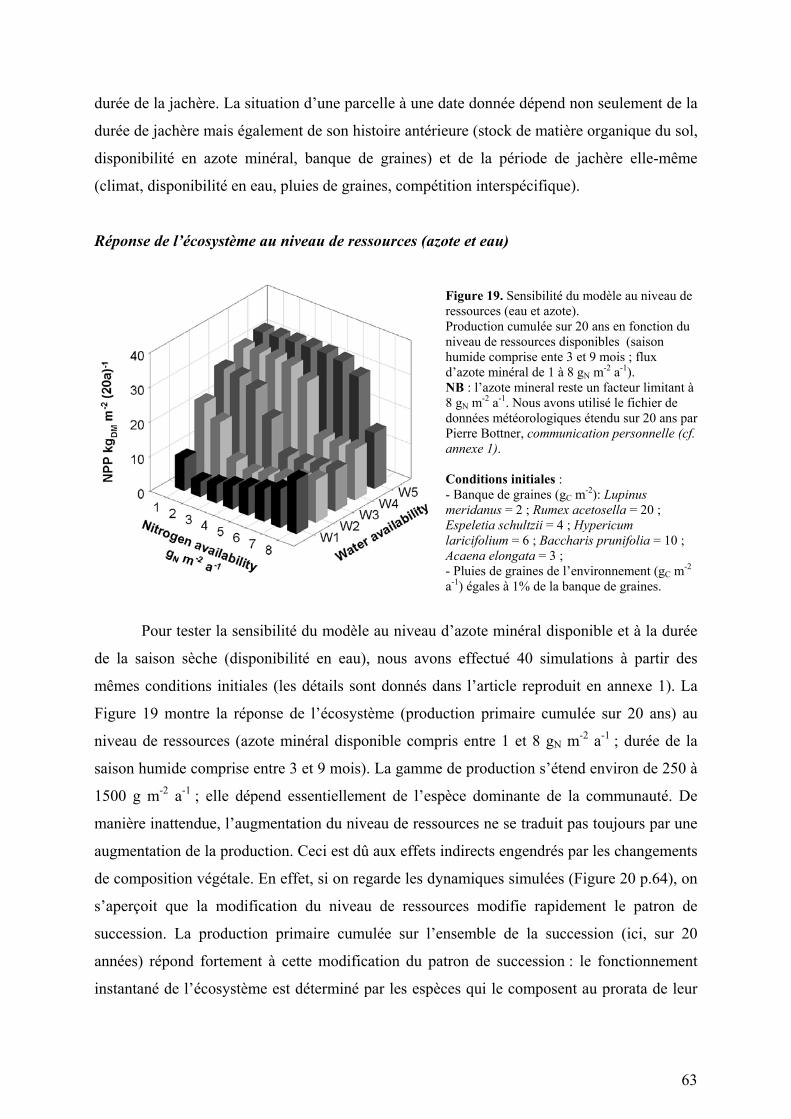
63
durée de la jachère. La situation d’une parcelle à une date donnée dépend non seulement de la
durée de jachère mais également de son histoire antérieure (stock de matière organique du sol,
disponibilité en azote minéral, banque de graines) et de la période de jachère elle-même
(climat, disponibilité en eau, pluies de graines, compétition interspécifique).
Réponse de l’écosystème au niveau de ressources (azote et eau)
Figure 19. Sensibilité du modèle au niveau de ressources (eau et azote). Production cumulée sur 20 ans en fonction du niveau de ressources disponibles (saison humide comprise ente 3 et 9 mois ; flux d’azote minéral de 1 à 8 gN m-2 a-1). NB : l’azote mineral reste un facteur limitant à 8 gN m-2 a-1. Nous avons utilisé le fichier de données météorologiques étendu sur 20 ans par Pierre Bottner, communication personnelle (cf. annexe 1). Conditions initiales : - Banque de graines (gC m-2): Lupinus meridanus = 2 ; Rumex acetosella = 20 ; Espeletia schultzii = 4 ; Hypericum laricifolium = 6 ; Baccharis prunifolia = 10 ; Acaena elongata = 3 ; - Pluies de graines de l’environnement (gC m-2 a-1) égales à 1% de la banque de graines.
Pour tester la sensibilité du modèle au niveau d’azote minéral disponible et à la durée
de la saison sèche (disponibilité en eau), nous avons effectué 40 simulations à partir des
mêmes conditions initiales (les détails sont donnés dans l’article reproduit en annexe 1). La
Figure 19 montre la réponse de l’écosystème (production primaire cumulée sur 20 ans) au
niveau de ressources (azote minéral disponible compris entre 1 et 8 gN m-2 a-1 ; durée de la
saison humide comprise entre 3 et 9 mois). La gamme de production s’étend environ de 250 à
1500 g m-2 a-1 ; elle dépend essentiellement de l’espèce dominante de la communauté. De
manière inattendue, l’augmentation du niveau de ressources ne se traduit pas toujours par une
augmentation de la production. Ceci est dû aux effets indirects engendrés par les changements
de composition végétale. En effet, si on regarde les dynamiques simulées (Figure 20 p.64), on
s’aperçoit que la modification du niveau de ressources modifie rapidement le patron de
succession. La production primaire cumulée sur l’ensemble de la succession (ici, sur 20
années) répond fortement à cette modification du patron de succession : le fonctionnement
instantané de l’écosystème est déterminé par les espèces qui le composent au prorata de leur
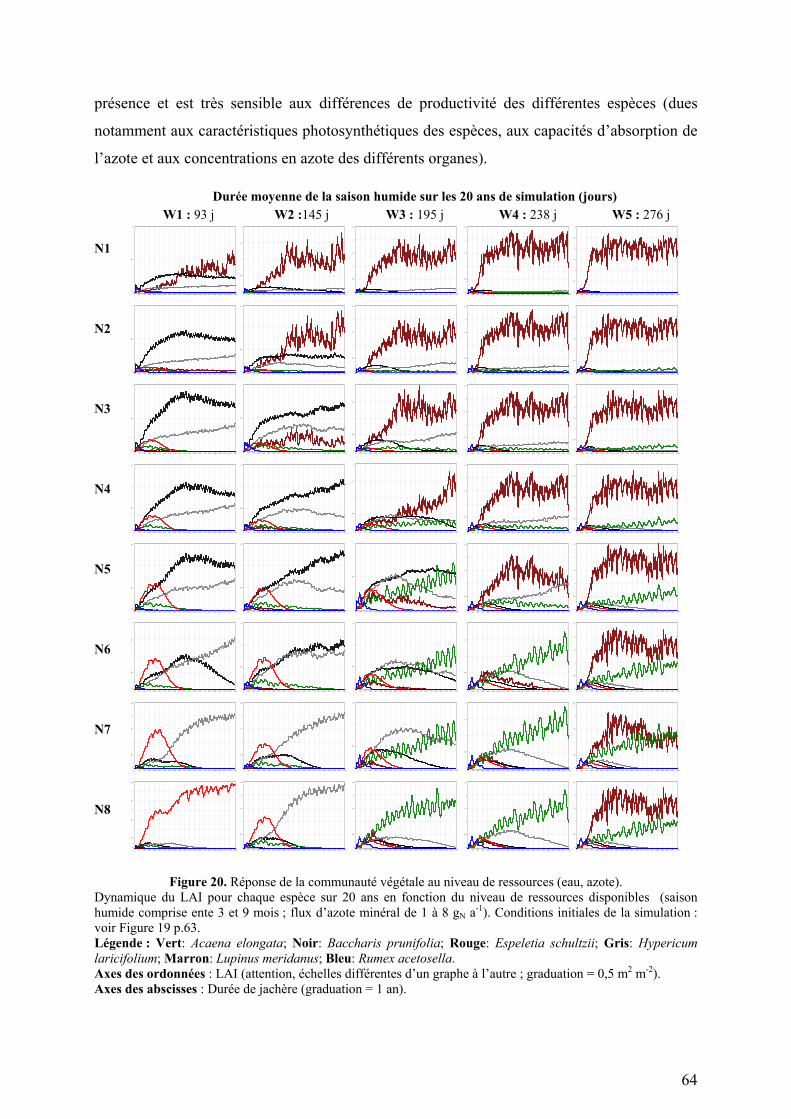
64
présence et est très sensible aux différences de productivité des différentes espèces (dues
notamment aux caractéristiques photosynthétiques des espèces, aux capacités d’absorption de
l’azote et aux concentrations en azote des différents organes). Durée moyenne de la saison humide sur les 20 ans de simulation (jours) W1 : 93 j W2 :145 j W3 : 195 j W4 : 238 j W5 : 276 j N1
N2
N3
N4
N5
N6
N7
N8
Figure 20. Réponse de la communauté végétale au niveau de ressources (eau, azote). Dynamique du LAI pour chaque espèce sur 20 ans en fonction du niveau de ressources disponibles (saison humide comprise ente 3 et 9 mois ; flux d’azote minéral de 1 à 8 gN a-1). Conditions initiales de la simulation : voir Figure 19 p.63. Légende : Vert: Acaena elongata; Noir: Baccharis prunifolia; Rouge: Espeletia schultzii; Gris: Hypericum laricifolium; Marron: Lupinus meridanus; Bleu: Rumex acetosella. Axes des ordonnées : LAI (attention, échelles différentes d’un graphe à l’autre ; graduation = 0,5 m2 m-2). Axes des abscisses : Durée de jachère (graduation = 1 an).
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
3
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
3
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
3
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
3.5
3
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
3
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300
2.5
2
1.5
1
0.5
00 365 730 1095 1460 1825 2190 2555 2920 3285 3650 4015 4380 4745 5110 5475 5840 6205 6570 6935 7300

65
En milieu sec et suffisamment riche en azote, la communauté est dominée par
Espeletia schultzii. Les productions les plus fortes sont atteintes en milieu pauvre en azote et
relativement humide : la légumineuse Lupinus meridanus domine alors la communauté
(Figure 20 p.64) ; elle fixe l’azote dont elle a besoin. Le modèle ne prend pas en compte les
facteurs limitants de la fixation symbiotique tel que le phosphore ; il est donc très
vraisemblable que la production des communautés dominées par le lupin soit surestimée.
2.2.4 - Compétition et coexistence
Sensibilité du modèle aux paramètres Hmax et SRL (étude de sensibilité III)
Utilisons maintenant la Figure 18 (p.62) pour tester la sensibilité du modèle à la
hauteur maximale (Hmax, Figure 18 haut) et à la longueur de racine par unité de masse (SRL,
Figure 18 bas). La modification du paramètre considéré induit une modification des surfaces
de dominance et de coexistence. La taille et la position de ces surfaces sont plus sensibles au
SRL qu’à la hauteur maximale. Ceci peut être interprété comme résultant du poids relatif de la
compétition pour l’azote par rapport à la compétition pour la lumière. Les conditions de
simulation mettent l’accent sur la carence en azote du système.
La Figure 21 présente l’élasticité de la biomasse à 5 ans d’Hypericum laricifolium et
de Baccharis prunifolia aux deux paramètres suivants : hauteur maximale et SRL
d’Hypericum laricifolium. On constate que c’est l’espèce dominée (Baccharis prunifolia dans
ce cas) qui réagit le plus fortement à une modification de ces paramètres. En outre, on
constate de nouveau que le modèle est plus fortement sensible au SRL qu’à la hauteur
maximale. L’extrême sensibilité du modèle au SRL devrait encourager à poursuivre des
études sur le terrain dans le but d’améliorer l’estimation de ce paramètre.
Figure 21. Sensibilité des biomasses d’Hypericum laricifolium et de Baccharis prunifolia aux paramètres Hmax et SRL d’Hypericum laricifolium. Paramètres du Tableau 4 excepté les conditions initiales : 1 gC de graines en dormance de chacune des deux espèces. On mesure la sensibilité du modèle sur la biomasse des deux espèces après 5 ans. Moyennes et écart-types de six variations de chaque paramètre (de -15% à +15% par pas de 5).
SRL Hmax
Ela
stic
ity
-2
-1
0
1
Hypericum laricifoliumBaccharis prunifolia

66
Rôle de la compétition dans le patron de succession
Les simulations effectuées montrent la capacité du modèle à reproduire le patron de
succession observé au champ. La compétition pour la lumière détermine très largement le
statut des espèces dans la succession. Les espèces de fin de succession sont caractérisées par
des traits22 liés à une grande taille et à une capacité d’ombrage importante (forte allocation
aux feuilles, longue durée de vie des feuilles).
La compétition pour l’azote détermine les espèces dominantes de chaque stade de
succession. En milieu très pauvre en azote, ce sont les espèces les plus ‘économes’ qui
dominent, c’est-à-dire les espèces qui sont caractérisées par des traits qui favorisent la
rétention des nutriments (turn-over lent des organes, allocation forte aux organes structuraux).
Dans les milieux riches (humides et peu carencés en azote), ce sont les espèces aux taux de
croissance les plus forts qui dominent.
Conditions de coexistence
Dans la version actuelle du modèle FAPROM, plusieurs espèces peuvent coexister
durablement, ce qui n’était pas le cas dans la première version. Cette modification du
comportement du modèle met en lumière l’importance des hypothèses de modélisation de la
compétition pour l’azote. La première version du modèle répartissait l’azote minéral entre les
espèces au prorata de leur demande en azote (Hypothèse 1). La version actuelle répartit
l’azote entre les espèces au prorata de la longueur (et de l’activité) de leurs racines
(Hypothèse 2). Cette différence est fondamentale : l’hypothèse 1 amplifiait la compétition
pour la lumière en favorisant l’absorption d’azote des espèces déjà favorisées par la lumière
(celles qui avaient la plus forte assimilation potentielle) ; l’hypothèse 2 dissocie plus
largement compétition pour la lumière et compétition pour l’azote. Cette dissociation favorise
la création de niches le long du gradient de ressources et l’existence d’équilibres
(dynamiques) de coexistence entre deux ou trois espèces. La prise en compte d’un plus grand
nombre de facteurs limitants favorisent la coexistence d’un plus grand nombre d’espèces.
D’autre part, les fluctuations climatiques favorisent également la coexistence par
dissociation de niches temporelles (saison sèche, saison humide). L’alternance d’années plutôt
humides et d’années plutôt sèches favorisent tour à tour des espèces différentes et entretient
donc durablement la coexistence. Les changements climatiques (diminution du nombre
d’années humides par décennie par exemple) peuvent entraîner des changements de
22 Traits de vie des espèces, traits de réponse et traits d’effet (Lavorel & Garnier 2002).

67
dominances. La diversité végétale peut jouer un rôle d’assurance (Yachi & Loreau 1999) en
limitant les fluctuations du fonctionnement de l’écosystème (nous rediscutons ultérieurement
cet aspect dans la partie 2.3).
Modèle mécaniste simplifié
L’annexe 4 présente des pistes de réflexion sur la façon de dégrader le modèle
FAPROM pour modéliser la dynamique d’une communauté végétale. Une étape importante
consiste à définir des indices mécanistes de compétition et à utiliser le modèle FAPROM pour
tester ces différents indices.
Les modèles mécanistes simplifiés que nous avons testés conservent une forte
sensibilité aux conditions initiales, contrairement aux modèles théoriques plus simples (tels
que les modèles du type Lotka-Volterra).
2.2.5 - Tentative de validation du modèle sur les parcelles boliviennes
a
Time (years after crop abandonment)
0 1 2 3 4
Bio
volu
me
0.0
1.0b
Time (years after crop abandonment)
10 11 12 13 14
Bio
volu
me
0.0
1.5Annual herbs (simulated)Perennial grass (simulated)Shrubs (simulated)Annual herbs (data)Perennial grass (data)Shrubs (data)
Figure 22. Biovolume par groupe fonctionnel (simulations et données). a) Parcelle de 0 à 4 ans de jachère ;
b) Parcelle de 10 à 14 ans de jachère. NB : Les données proviennent de Teresa Ortuño (communication personnelle). Les simulations ont été effectuées avec les conditions initiales optimisées définies dans le Tableau 14 (page suivante).
Nous présentons ici quelques résultats obtenus avec le modèle sur le site bolivien. La
Figure 22 présente la dynamique du biovolume simulée pour deux parcelles de Patacamaya
(l’une ‘jeune’ : de 0 à 4 ans de jachère ; l’autre ‘vieille’ : de 10 à 14 ans de jachère). Les
simulations sont confrontées aux données de Teresa Ortuño (communication personnelle) : 4
données en saison humide et deux en saison sèche (1998-2002). Nous avons groupé sur les
données de terrain et les simulations les différentes espèces en 3 groupes fonctionnels : les
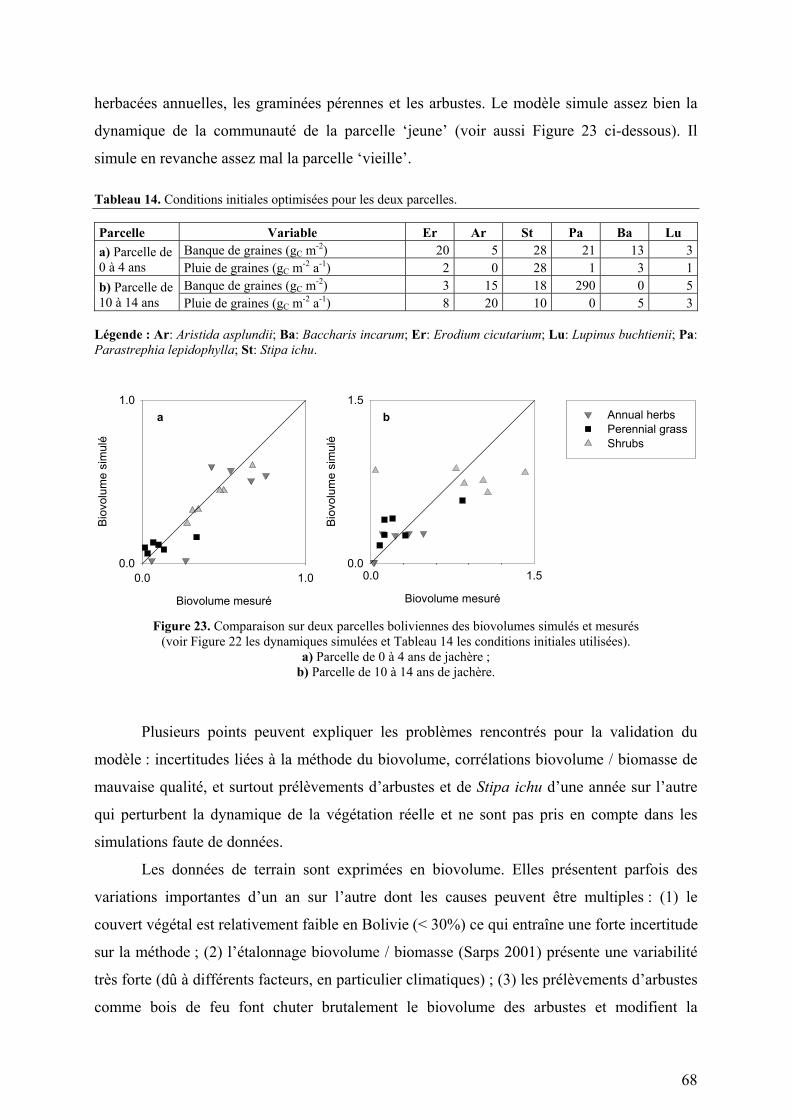
68
herbacées annuelles, les graminées pérennes et les arbustes. Le modèle simule assez bien la
dynamique de la communauté de la parcelle ‘jeune’ (voir aussi Figure 23 ci-dessous). Il
simule en revanche assez mal la parcelle ‘vieille’. Tableau 14. Conditions initiales optimisées pour les deux parcelles.
Parcelle Variable Er Ar St Pa Ba Lu Banque de graines (gC m-2) 20 5 28 21 13 3a) Parcelle de
0 à 4 ans Pluie de graines (gC m-2 a-1) 2 0 28 1 3 1Banque de graines (gC m-2) 3 15 18 290 0 5b) Parcelle de
10 à 14 ans Pluie de graines (gC m-2 a-1) 8 20 10 0 5 3 Légende : Ar: Aristida asplundii; Ba: Baccharis incarum; Er: Erodium cicutarium; Lu: Lupinus buchtienii; Pa: Parastrephia lepidophylla; St: Stipa ichu.
Biovolume mesuré
0.0 1.0
Biov
olum
e si
mul
é
0.0
1.0Annual herbsPerennial grassShrubs
Biovolume mesuré
0.0 1.5
Biov
olum
e si
mul
é
0.0
1.5a b
Figure 23. Comparaison sur deux parcelles boliviennes des biovolumes simulés et mesurés
(voir Figure 22 les dynamiques simulées et Tableau 14 les conditions initiales utilisées). a) Parcelle de 0 à 4 ans de jachère ;
b) Parcelle de 10 à 14 ans de jachère.
Plusieurs points peuvent expliquer les problèmes rencontrés pour la validation du
modèle : incertitudes liées à la méthode du biovolume, corrélations biovolume / biomasse de
mauvaise qualité, et surtout prélèvements d’arbustes et de Stipa ichu d’une année sur l’autre
qui perturbent la dynamique de la végétation réelle et ne sont pas pris en compte dans les
simulations faute de données.
Les données de terrain sont exprimées en biovolume. Elles présentent parfois des
variations importantes d’un an sur l’autre dont les causes peuvent être multiples : (1) le
couvert végétal est relativement faible en Bolivie (< 30%) ce qui entraîne une forte incertitude
sur la méthode ; (2) l’étalonnage biovolume / biomasse (Sarps 2001) présente une variabilité
très forte (dû à différents facteurs, en particulier climatiques) ; (3) les prélèvements d’arbustes
comme bois de feu font chuter brutalement le biovolume des arbustes et modifient la

69
dynamique globale de la communauté ; le manque d’information concernant ces ‘aléas’ rend
difficile l’optimisation des simulations.
Nous avons essayé de simuler les 19 parcelles boliviennes suivies pendant 4 ans par
Teresa Ortuño. Sur ces 19 parcelles, seules 14 parcelles sont exploitables. Le modèle
reproduit d’autant mieux la dynamique que la parcelle est jeune. Pour les parcelles en jachère
depuis plus de 10 ans, le modèle est incapable de simuler correctement les herbacées. Aucune
herbacée pérenne non graminée n’a été paramétrée et leur dynamique est assez différente de
celle d’Erodium cicutarium (espèce pionnière rudérale).
2.2.6 - Combien d’espèces et quelles espèces faut-il prendre en compte ?
Le fonctionnement instantané de l’écosystème est déterminé par les espèces au prorata
de leur présence (Biomass ratio hypothesis, Grime 1998, Garnier 2004). Aussi est-il important
de prendre en compte les espèces dominantes. Or on s’intéresse à la dynamique de la
communauté, il faut donc modéliser les espèces dominantes aux différents stades de
succession. La divergence des patrons de succession nécessite de s’intéresser à plusieurs
espèces par stade de succession. Si l’on souhaite modéliser des changements de conditions tel
que l’exclusion du pâturage, il faut également modéliser les espèces qui dominent dans ces
nouvelles conditions. La difficulté est de déterminer a priori les espèces qui joueront un rôle
déterminant a posteriori. C’est une problématique qui est largement débattue en biologie de la
conservation (Courchamp et al. 2003) : certaines espèces minoritaires sont amenées à jouer un
rôle clef dans l’écosystème après perturbation.
Les effets indirects via la compétition interspécifique peuvent avoir un rôle primordial
dans la dynamique de la communauté. Aussi certaines espèces rares peuvent-elles tenir un
rôle clef. Parmi ces espèces, on peut distinguer (1) les espèces opportunistes (telles
qu’Erodium cicutarium ou Lupinus buchtienii en Bolivie) qui peuvent exploser sur l’espace
nu laissé libre (après l’arrachage d’un arbuste par exemple) si les conditions météorologiques
et biogéochimiques sont favorables, et donc empêcher ou retarder le développement d’une
autre espèce, et (2) les espèces qui jouent un rôle fonctionnel particulier (les légumineuses par
exemple) et qui peuvent changer les conditions environnementales de façon décisive même si
elles ne dominent pas la communauté. Aussi est-il souhaitable de couvrir autant que possible
l’ensemble des groupes fonctionnels, et ce, pour les différents stades de succession.

70
En Bolivie, aucune herbacée pérenne (excepté les graminées) n’a été modélisée alors
que ce groupe participe pour un bon tiers au biovolume des parcelles de fin de succession. Les
premiers essais de validation du modèle sur les parcelles de Patarani (Bolivie) ont montré
qu’il était difficile de prédire la dynamique observée dans les parcelles âgées : non seulement,
on est incapable d’estimer la quantité d’herbacées, mais en plus, la non prise en compte de ce
groupe modifie les relations de compétition entre les autres groupes fonctionnels (graminées
et arbustes).
Au Venezuela, aucune graminée n’a été modélisée. Dans l’écosystème ouvert,
régulièrement pâturé, elles sont minoritaires et contribuent vraisemblablement assez peu au
fonctionnement de l’écosystème. Mais en champ clos, dans les expériences d’exclusion du
pâturage, elles dominent et modifient à la fois le fonctionnement de l’écosystème et la
dynamique de la communauté (en retardant le développement des espèces de fin de
succession).
La gestion matricielle des équations dans le modèle permet d’ajouter ou d’enlever
facilement des espèces. Il suffit de changer le nombre de modalités de l’indice espèce et de
modifier le fichier des paramètres d’entrée. Le temps de calcul de la version actuelle est de
l’ordre de 0,25 seconde par espèce et par an. Si on veut étudier la dynamique de 60 espèces
sur 100 ans, le temps de calcul devrait rester inférieur à la demi-heure. Le temps de calcul ne
semble donc pas limitant face aux efforts que nécessitent l’identification des paramètres et la
calibration.
Comment gérer la sensibilité aux conditions initiales ?
La forte sensibilité du modèle FAPROM aux conditions initiales peut sembler limiter
l’intérêt du modèle. Quelle crédibilité accorder à des simulations dont une petite variation des
conditions initiales produit des changements brutaux en sortie ? En apparence, ce
comportement peut mettre en évidence un manque de robustesse. Néanmoins, on peut aussi
supposer qu’on aurait le même manque de robustesse dans des expériences réelles de
compétition avec un nombre limité d’espèces. En outre, cette sensibilité aux conditions
initiales peut être limitée : (1) en prenant en compte suffisamment d’espèces (cf. ci-dessus) ;
(2) en étudiant le cycle reproductif des espèces pour définir les contraintes de variation des
conditions initiales ; (3) en étudiant non pas seulement la dynamique de la communauté mais
le fonctionnement global de l’écosystème.

71
2.3 - ROLE DE LA JACHERE DANS L’AGRO-ECOSYSTEME
Nous présentons maintenant les résultats prédits par le modèle FAPROM sur le
fonctionnement de l’écosystème. Nous avons étudié en particulier : (1) le rôle de la jachère
dans le système de culture et les possibilités de jachère améliorée, (2) l’impact du pâturage et
de la cueillette sur la dynamique de la communauté et le fonctionnement de la jachère, et (3)
le rôle de la biodiversité sur les services de l’écosystème.
2.3.1 - Cycle de l’azote au cours de la jachère et potentialités de la jachère améliorée
Figure 24. Dynamique du contenu en azote au cours de la jachère. a) Contenu en azote de la biomasse, de la nécromasse sur pied et de la litière (simulation de référence, paramètres vénézuéliens, Tableau 4 ; données de Sarmiento 1995 pour une parcelle de 12 ans) ; b) Contenu en azote de la matière organique du sol pour 3 simulations :
communauté végétale (simulation de référence, Tableau 4) ;
communauté végétale sans légumineuse ; sol nu.
La Figure 24a présente la dynamique de l’accumulation d’azote dans la biomasse, la
nécromasse sur pied et la litière. Le contenu en azote total de ces compartiments augmente en
début de jachère, atteint un pic au bout de 3 ans (11 gN m-2) et diminue légèrement pour se
fixer autour de 10 gN m-2. La continuation de la jachère au delà de 5 ans ne modifie pas le
contenu en azote total du compartiment végétal. La Figure 24b compare trois scénarios
différents : sol nu ; jachère sans légumineuse et jachère avec légumineuse. Le contenu en
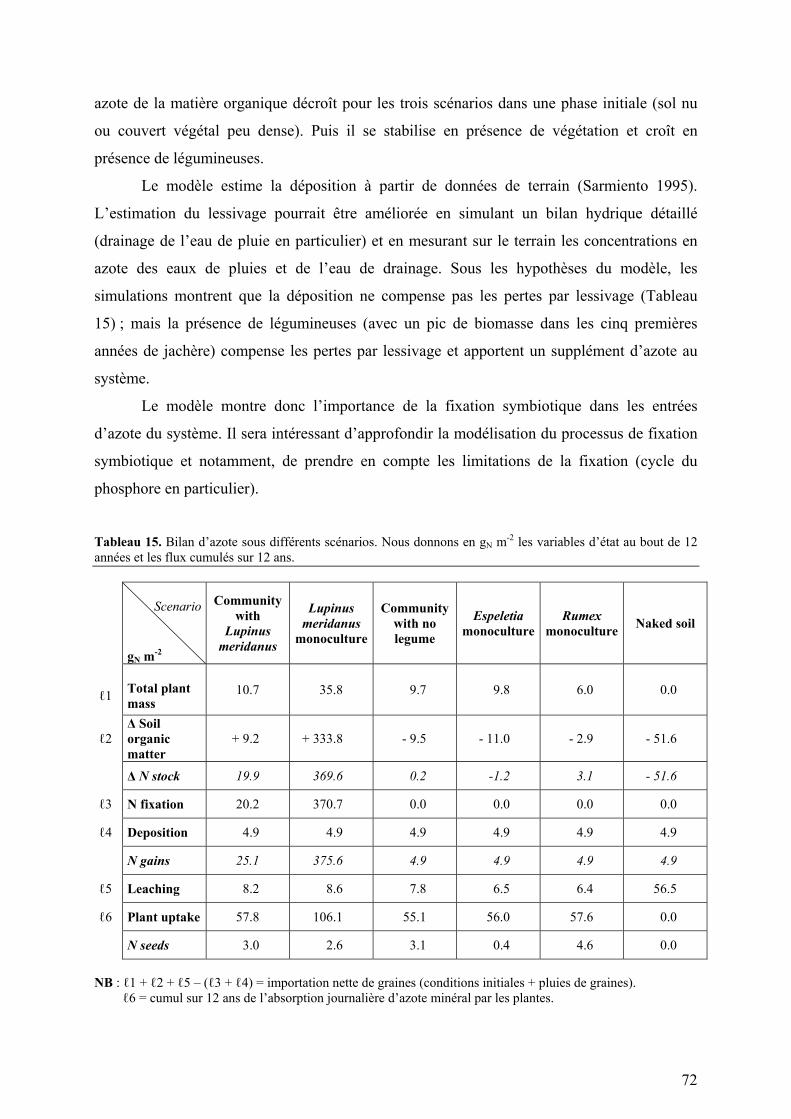
72
azote de la matière organique décroît pour les trois scénarios dans une phase initiale (sol nu
ou couvert végétal peu dense). Puis il se stabilise en présence de végétation et croît en
présence de légumineuses.
Le modèle estime la déposition à partir de données de terrain (Sarmiento 1995).
L’estimation du lessivage pourrait être améliorée en simulant un bilan hydrique détaillé
(drainage de l’eau de pluie en particulier) et en mesurant sur le terrain les concentrations en
azote des eaux de pluies et de l’eau de drainage. Sous les hypothèses du modèle, les
simulations montrent que la déposition ne compense pas les pertes par lessivage (Tableau
15) ; mais la présence de légumineuses (avec un pic de biomasse dans les cinq premières
années de jachère) compense les pertes par lessivage et apportent un supplément d’azote au
système.
Le modèle montre donc l’importance de la fixation symbiotique dans les entrées
d’azote du système. Il sera intéressant d’approfondir la modélisation du processus de fixation
symbiotique et notamment, de prendre en compte les limitations de la fixation (cycle du
phosphore en particulier).
Tableau 15. Bilan d’azote sous différents scénarios. Nous donnons en gN m-2 les variables d’état au bout de 12 années et les flux cumulés sur 12 ans.
Scenario gN m-2
Community with
Lupinus meridanus
Lupinus meridanus
monoculture
Community with no legume
Espeletia monoculture
Rumex monoculture Naked soil
ℓ1 Total plant mass
10.7 35.8 9.7 9.8 6.0 0.0
ℓ2 ∆ Soil organic matter
+ 9.2 + 333.8 - 9.5 - 11.0 - 2.9 - 51.6
∆ N stock 19.9 369.6 0.2 -1.2 3.1 - 51.6
ℓ3 N fixation 20.2 370.7 0.0 0.0 0.0 0.0
ℓ4 Deposition 4.9 4.9 4.9 4.9 4.9 4.9
N gains 25.1 375.6 4.9 4.9 4.9 4.9
ℓ5 Leaching 8.2 8.6 7.8 6.5 6.4 56.5
ℓ6 Plant uptake 57.8 106.1 55.1 56.0 57.6 0.0
N seeds 3.0 2.6 3.1 0.4 4.6 0.0
NB : ℓ1 + ℓ2 + ℓ5 – (ℓ3 + ℓ4) = importation nette de graines (conditions initiales + pluies de graines).
ℓ6 = cumul sur 12 ans de l’absorption journalière d’azote minéral par les plantes.

73
Quelles sont les fonctions agro-écologiques de la jachère ?
Le premier rôle de la jachère est de constituer un stock de biomasse qui est incorporée
à la terre à la fin de la jachère et qui a la fonction d’engrais vert en se décomposant au cours
de la culture. Ce stock atteint un plateau relativement rapidement dans les simulations (3 ou 4
ans23). Les prédictions du modèle sur la qualité de la litière produite (composition spécifique,
contenus en azote ou rapports C/N) peuvent être utilisées conjointement aux études menées
sur le terrain par Marie-Madeleine Coûteaux (Coûteaux et al. 2004a et b) sur les taux de
décomposition des différentes espèces (et type de litière) pour savoir à quel moment il est le
plus judicieux de rompre la jachère. A priori, les ligneux se décomposent moins vite que les
herbacées ou les graminées et un mélange approprié de ligneux et d’herbacées pourrait
permettre de gérer la fertilité (la mobilisation de l’azote minérale) sur les deux années de
culture.
Le second rôle de la jachère est d’accroître le stock de matière organique du sol, en
particulier, d’alimenter le pool d’azote organique : (1) la végétation naturelle limite les pertes
par lessivage (ce qui est cohérent avec la fermeture progressive du cycle de l’azote mise en
évidence par l’étude du δ15N de la matière organique du sol qui décroît au cours de la jachère
(Abadin et al. 2002, cf. p.10)) ; (2) les légumineuses apportent de l’azote au système par
fixation symbiotique. Le pic de biomasse des légumineuses (et de l’activité fixatrice) a
également lieu en début de jachère (dans les cinq premières années, ce qui est cohérent avec
les observations de terrain, à la fois au Venezuela (Sarmiento et al. 2003) et en Bolivie
(Ortuño et al. 2004)). L’amélioration de la jachère par semis de lupin permet d’augmenter
considérablement le stock d’azote (il s’agit d’une efficacité potentielle de la fixation
symbiotique puisque le modèle ne prend pas en compte pour l’instant la limitation de la
fixation par le phosphore).
Le troisième rôle agro-écologique de la jachère est d’éliminer les adventices. Les
simulations montrent une réduction importante des espèces rudérales au cours des 2 ou 3
premières années. Des études sur le terrain sur la banque de graines, les pluies de graines et la
dormance devraient permettre de déterminer la durée optimale de la jachère au regard de la
lutte contre les mauvaises herbes. Une jachère trop courte ne fait qu’accroître la banque de
graines des espèces rudérales et risque donc d’être néfaste pour la culture.
23 Mais il faut faire attention à la surestimation des vitesses dans le modèle et garder à l’esprit la sensibilité de la production et de la biomasse aux conditions initiales et environnementales.

74
D’autres rôles agro-écologiques doivent être pris en considération mais ne sont pas
intégrés au modèle : limiter les populations de nématodes à kystes de la pomme de terre et les
autres parasites de la culture ; stimuler l’activité de la biomasse microbienne et de la
microfaune du sol ; modifier la structure du sol.
Proposition de gestion durable de l’agro-écosystème
La durée optimale de la jachère, au regard de la gestion de la fertilité du sol, fait donc
l’objet de compromis entre ces différentes fonctions : reconstitution du stock de matière
organique, fixation symbiotique d’azote, qualité de la litière incorporée au sol, lutte contre les
adventices, limitation des parasites, stimulation de l’activité microbienne, modification de la
structure du sol. Elle n’est pas nécessairement très longue (environ 5 ans). Elle peut être
accélérée (pour certaines fonctions agro-écologiques en particulier ; il faudrait vérifier sur les
autres fonctions qu’on ne détériore pas l’efficacité de la jachère).
La jachère améliorée par semis de légumineuses offre de fortes potentialités (mais il
faudrait inclure le cycle du phosphore dans le modèle pour pouvoir prédire l’efficacité réelle
de la fixation). Il sera intéressant de confronter les prédictions du modèle aux expériences
menées sur le terrain. En Bolivie, la jachère semée en lupin a doublé les rendements de la
première année de culture (Dominique Hervé, communication personnelle). Au Venezuela,
des résultats similaires ont été obtenus (Lina Sarmiento, communication personnelle).
D’autres idées d’amélioration du système de jachère peuvent être testées avec le
modèle FAPROM : fertilisation de la jachère, modification de la qualité de litière par travail
de la jachère (arrachage de certaines espèces), alternance de périodes de culture et de jachère
plus courtes.
Il pourrait être intéressant par exemple de fertiliser la parcelle au cours de la jachère,
au moins un an avant la mise en culture. En effet, la fertilisation sur sol nu en début de culture
n’est pas optimale : une partie importante de l’azote minéral ajouté est lessivé avant le
développement de la culture. La fertilisation sur jachère serait sans doute un moyen
d’optimiser l’entrée d’azote dans le système. Par rapport à l’utilisation directe d’intrants
organiques, l’utilisation de la biomasse verte garantit une meilleure adéquation entre les
besoins de la culture et la disponibilité en azote minéral. Par rapport à la jachère
traditionnelle, la fertilisation sur jachère permet d’augmenter cette biomasse verte. Il faut
toutefois vérifier qu’elle ne favoriserait pas les adventices aux dépens des légumineuses et des
espèces de fin de succession.

75
L’utilisation conjointe du modèle FAPROM et des études sur la décomposition des
parties aériennes et souterraines des différentes espèces (Coûteaux et al. 2004a et b) devraient
permettre de définir la composition optimale de la communauté (au moment de rompre la
jachère) pour une régulation du flux d’azote minéral sur la période de culture. Le mélange
idéal de qualités de litière différentes permettrait de prolonger la période cultivable sur trois
années sans chute des rendements. Pour atteindre ce mélange idéal, on peut considérer un
travail de la jachère : arrachage de certaines espèces (ou modification de la pression de
pâturage, cf. paragraphes suivants) ; fertilisation à un certain moment ; irrigations des
parcelles. Ces solutions peuvent paraître surprenante mais il serait intéressant de les tester sur
le terrain.
Une autre modification du système traditionnel consisterait à raccourcir les temps de
culture et les périodes de jachère : l’alternance par exemple de cycles 1 / 4 (1 an de culture,
suivie de 4 ans de jachère). Il faut néanmoins veiller aux conséquences d’une telle pratique,
notamment sur la diversité de l’écosystème : on risque de privilégier à chaque cycle les
espèces rudérales. Peut-être faut-il combiner ces différentes idées (semis de lupin et
raccourcissement du temps de jachère, semis de lupin et fertilisation).
A quelle distance sommes-nous d’un modèle d’aide à la décision ?
Nous sommes relativement loin d’un modèle utilisable par un gestionnaire. En effet,
un certain nombre d’étapes mériterait au préalable d’être franchi : (1) la validation
approfondie du modèle FAPROM sur l’ensemble des données collectées sur le terrain afin de
tester la robustesse des prédictions sur l’ensemble le plus vaste de conditions différentes ; (2)
le développement éventuel d’autres modules (en particulier, le cycle du phosphore, le bilan
hydrique, la répartition spatiale dans un modèle d’automate cellulaire) ; (3) la validation des
modèles Sol et Culture ; (4) la validation du modèle intégré à l’échelle de la parcelle
(couplages Sol et Végétation, Culture et Jachère). Néanmoins le modèle permet dans sa
version actuelle de sélectionner des scénarios à tester sur le terrain.
Le modèle repose sur l’estimation instantanée des processus écophysiologiques à
l’échelle de la feuille (photosynthèse) ou de l’organe (mortalité, respiration). De nombreuses
hypothèses sont formulées pour permettre les changements d’échelle de l’organe à l’espèce,
de l’espèce à l’écosystème, de la seconde à la journée, et de la journée à l’année. Une
validation complète du modèle nécessiterait une validation des estimations des flux aux
échelles larges (écosystème, journée, année) en utilisant par exemple des mesures de tour à
flux (cependant difficile en pratique sur les terrains en pente du Venezuela) ou sous cloche.

76
D’autre part, les aspects socio-économiques ne doivent pas être négligés et il est
difficile de construire un modèle d’aide à la décision qui ne prenne pas en compte ces aspects.
En effet, les mesures d’amélioration de la jachère présentent un coût qu’il faudrait intégrer.
De plus, nous avons privilégié dans ce travail un aspect particulier de la fertilité (la
disponibilité de l’azote minéral), mais l’aptitude du sol à produire est fonction de quantité
d’autres facteurs (les autres nutriments minéraux, le climat, les intrants d’azote et d’eau, la
pratique culturale, la lutte contre les parasites et cætera).
2.3.2 - Impact du pâturage
a
Grazing pressure
0.00 0.05 0.10 0.15 0.20
g DM m
-2 a
-1
0
200
400
600
800NPPGrazed by animals
b
Grazing pressure
0.00 0.01 0.02 0.03 0.04
g DM m
-2 a
-1
0
200
400
600
800
Figure 25. Effet du pâturage sur la productivité de l’écosystème. a) Prélèvement d’Hypericum laricifolium dans un couvert végétal monospécifique.
b) Prélèvement d’Hypericum laricifolium dans la communauté végétale. La productivité de l’écosystème (NPP, moyenne sur 20 ans) est séparée en deux composantes : ce qui est prélevé par les animaux (Grazed by animals) et ce qui reste dans le cycle végétal (différence entre NPP et grazed by animals). La pression de pâturage correspond au pourcentage de biomasse de feuilles prélevée par les animaux, une fois par période de 30 jours. NB : Les échelles des abscisses sont différentes ! L’augmentation de la productivité de la communauté avec la pression de pâturage est essentiellement due à un changement de dominance : Hypericum laricifolium est progressivement exclu sous la pression directe du pâturage et les effets de la compétition entre espèces ; Espeletia schultzii bénéficie de l’exclusion d’Hypericum laricifolium et domine en fin de succession ; sa productivité est supérieure à celle d’Hypericum laricifolium.
Nous avons effectué des simulations pour comprendre l’impact du pâturage sur la
production végétale et sur le processus de succession. Les paramètres utilisés dans cette étude
correspondent à ceux du Venezuela (Tableau 4) mais cette étude est purement théorique :
aucune des espèces prises en compte dans le modèle n’est réellement appétante. La Figure 25a
présente l’effet du pâturage sur une monoculture sans prendre en compte l’impact des fèces
du bétail sur la matière organique ni l’impact mécanique du passage des animaux sur le
couvert végétal et la structure du sol. On constate que la productivité primaire est stimulée par

77
le pâturage dans une large gamme d’intensité de pâturage : la consommation d’une partie des
feuilles à intervalles réguliers augmente l’allocation aux feuilles. Les résultats diffèrent pour
les mélanges d’espèces. La réponse de l’écosystème au pâturage dépend très fortement des
effets indirects. On a étudié la réponse de la communauté au pâturage d’une seule espèce : une
espèce dominante en fin de succession en absence de pâturage. La consommation des
animaux est proportionnelle à la biomasse de l’espèce pâturée ; cette biomasse est beaucoup
plus sensible à l’intensité du pâturage dans une communauté plurispécifique (Figure 25b) que
lorsque l’espèce est en monoculture (Figure 25a). La productivité de l’écosystème dépend de
la communauté résultante. Cette étude montre principalement que les espèces pâturées seront
rares du fait que la compétition accélère l’effet d’exclusion du pâturage. Les expériences
menées sur le terrain (Sarmiento, communication personnelle) ont montré que les espèces
dominantes dans les champs clôturés (pâturage exclu) sont en général plus appétantes
(graminées notamment).
La cueillette des arbustes utilisés comme bois de feu et le prélèvement des touffes de
Stipa utilisées comme chaume modifient l’aspect des parcelles et la dynamique de la
communauté. L’espace libéré est recolonisé par les espèces pionnières et les espèces
opportunistes. L’absence de relevé précis de ces pratiques rend parfois difficile
l’interprétation des suivis de biovolumes.
2.3.3 - Relations entre la diversité des communautés et le fonctionnement de
l’écosystème24
Pour tester la relation entre la diversité de la communauté et le fonctionnement de
l’écosystème, nous avons tester l’ensemble des 26 = 64 assemblages possibles (il y a deux
modalités par espèce : présence/absence). Nous avons effectué les 64 simulations sur 20 ans
et comparons les productions primaires nettes. Si nous trions les simulations en fonction des
24 On s’intéresse à la biodiversité pour trois types de raisons au moins (Loreau 2002) : (1) elle procure un certain nombre de biens qui possèdent une valeur économique directe (nourriture, médicaments, gènes d’intérêt pour l’amélioration variétale, organismes utilisés en contrôle biologique) ; (2) elle est intrinsèquement liée au bien-être de l’homme pour des raisons éthiques, esthétiques, culturelles et scientifiques ; (3) elle peut contribuer à des "services" écologiques (production primaire et secondaire, pollinisation des plantes, régulation du climat, régulation du cycle de l’eau, maintien de la qualité de l’eau, maintien de la fertilité des sols). Au cours de la dernière décennie, les effets de la biodiversité sur les autres composantes du changement global ont reçu une attention croissante. En particulier, on a assisté à une croissance explosive des recherches sur les effets potentiels de la perte de biodiversité sur le fonctionnement des écosystèmes, et, par là, sur les "biens" et "services" écologiques qu’ils procurent aux sociétés humaines.

78
espèces présentes, on peut étudier la réponse de l’écosystème à la perte d’une espèce (Figure
26). On observe des patrons de réponse très différents suivant l’espèce à laquelle on
s’intéresse.
a
x espèces
0 1 2 3 4 5
g DM m
-2 a
-1
0
400
800
1200
1600 x espèces sans l'espèce ciblex espèces + l'espèce cible
b
x espèces
0 1 2 3 4 5
g DM m
-2 a
-1
0
400
800
1200
1600
Figure 26. Effet de la perte de diversité sur le fonctionnement de la communauté. a) Espèce cible : Lupinus meridanus ; b) Espèce cible: Baccharis prunifolia.
Axe des ordonnées : productivité primaire nette (moyenne annuelle sur 20 ans) ; axes des abscisses : nombre d’espèces dans la communauté (en noir), nombre d’espèces associées à l’espèce cible (en gris).
La disparition de Lupinus meridanus (Figure 26a, passage des barres grises aux barres
noires précédentes) provoque une diminution de la productivité de l’écosystème. Lupinus
meridanus est l’espèce la plus productive et la seule légumineuse prise en compte. La
communauté perd non seulement la contribution la plus forte à la productivité mais également
la fonction fixatrice d’azote. On peut également noter que l’augmentation de la diversité
réduit la variabilité (même lorsqu’on corrige par le nombre d’assemblages) : la diversité a un
effet ‘tampon’.
La perte de Baccharis prunifolia (Figure 26b, passage des barres grises aux barres
noires précédentes) provoque une augmentation de la productivité de l’écosystème. L’arbuste
Baccharis prunifolia a tendance à fermer la communauté. La perte de cette espèce laisse
l’opportunité à des espèces plus productives de se développer et de participer
significativement à la productivité de l’écosystème. D’autre part, si on ne considère que les
communautés végétales composées d’au moins Baccharis prunifolia (Figure 26b, barres
grises), il y a un net effet positif de la diversité sur la productivité de l’écosystème.
Ces simulations montrent qu’il y a un effet espèce majeur mais qu’on peut aussi mettre
en évidence sous certaines hypothèses sur les fonctions écologiques assurées par les
différentes espèces assemblées, un effet positif de la diversité sur le fonctionnement de
l’écosystème (diminution de la variabilité et augmentation de la productivité).

79
Certains assemblages révèlent des cas de complémentarité de niche par association de
groupes fonctionnels. En particulier, les mélanges Rumex acetosella et Lupinus meridanus
sont plus productifs sur 20 ans (en moyenne, 1705 g m-2 a-1) que les monocultures prises
séparément sur la même durée (respectivement 678 et 1697 g m-2 a-1). Ce type d’expérience
est très sensible à la durée de l’expérience. Dans les cas des assemblages Rumex acetosella et
Lupinus meridanus ils ne sont pas plus productifs que les monocultures sur des périodes de
deux ans. Si on regarde la production sur 2 ans et non plus sur 20 ans, le mélange Rumex
acetosella et Espeletia schultzii (635 g m-2 a-1) est plus productif que les monocultures
associées (respectivement 615 et 587 g m-2 a-1) ; de même, le mélange Acaena elongata et
Hypericum laricifolium (457 g m-2 a-1) produit plus que ses composantes seules
(respectivement 442 et 425 g m-2 a-1).
Pour tester l’hypothèse d’assurance, nous avons fait varier les conditions
environnementales et regardé si les mélanges résistaient mieux que les monocultures
associées aux changements de conditions. Certains mélanges (Espeletia schultzii et Rumex
acetosella par exemple) résistent mieux aux changements de conditions environnementales
(Espeletia schultzii résiste mieux à une carence en azote et au stress hydrique ; Rumex
acetosella est plus productif en milieu riche). Cependant, sur l’étendue des conditions
environnementales que nous avons testées, certaines espèces (telles que Lupinus meridanus)
garantissent la productivité la plus forte quel que soit le milieu.
Il serait intéressant de poursuivre ce type d’études en paramétrant un plus grand
nombre d’espèces.

80
3 - Bilan critique et perspectives
81
81
81
81
83
83
85
86
86
86
86
87
90
90
90
91
91
91
3.1 - HYPOTHESES DU MODELE
3.1.1 - Intérêt du modèle, synthèse des résultats obtenus
But du modèle
Thèses développées
3.1.2 - Critique des hypothèses et perspectives de prolongement
Limites du modèle
Quel modèle choisir (bis) ?
3.2 - PROJET DE THESE
3.2.1 - Dynamique du projet TROPANDES, travail en équipe
3.2.2 - Evaluation critique du travail
Dans quelle mesure les objectifs ont-ils été atteints ?
Analyse des difficultés rencontrées
3.3 - REFLEXIONS PERSONNELLES ET PERSPECTIVES
3.3.1 - De la multiplicité des métiers du chercheur
3.3.2 - Avenir du modèle FAPROM
Validation complète sur les données vénézuéliennes
Adaptation aux écosystèmes méditerranéens
Utilisation du modèle dans les études ‘diversité et fonctionnement’

81
Cette troisième partie a pour objectif d’évaluer le travail effectué : (1) nous revenons
d’abord sur le but et les résultats du modèle, et resituons notre étude dans les perspectives
actuelles de l’écologie ; (2) nous analysons ensuite dans quelle mesure les objectifs définis au
départ ont été atteints et quelles difficultés nous avons rencontrées au cours de la thèse ; (3)
nous discutons enfin l’expérience acquise en matière de recherche et l’avenir du modèle
FAPROM.
3.1 - RETOUR SUR LES HYPOTHESES DU MODELE
Nous commençons par résumer le but du modèle et les résultats obtenus puis nous
revenons sur les hypothèses du modèle, en montrons les limites et les possibles dépassements.
3.1.1 - Intérêt du modèle, synthèse des résultats obtenus
But du modèle
Nous avons développé un modèle biogéochimique de succession végétale qui lie
écophysiologie et écologie des communautés. Son originalité est d’assurer le couplage entre
les processus écophysiologiques instantanés et la dynamique de la végétation, et d’intégrer
tous les compartiments mesurés en identifiant des liens mécanistes entre eux. Il permet à la
fois d’établir la réponse des végétaux au cours de la succession (traits de réponse) et d’étudier
l’effet des végétaux sur le fonctionnement des écosystèmes (traits d’effet).
Thèses développées
Le modèle montre l’importance de prendre en compte le turn-over de la matière dans
la modélisation des dynamiques écologiques : (1) à l’échelle d’une espèce, le turn-over est
responsable des décalages temporels observés dans les dynamiques logistiques et se traduit
dans l’asymétrie des courbes de croissance ; (2) à l’échelle de la communauté, le turn-over
des espèces joue un rôle prépondérant dans les processus de compétition, en particulier, dans
les milieux pauvres en azote minéral, où les espèces qui dominent sont caractérisées par des
traits qui conduisent à une forte capacité de rétention des nutriments ; (3) à l’échelle de la
parcelle enfin, le turn-over est impliqué dans la dynamique de la fertilité du sol : la
disponibilité des nutriments est fonction de leur temps de résidence dans les différents

82
compartiments de la matière organique. Par conséquence, dans les écosystèmes herbacés
tropicaux au turn-over relativement rapide, la biomasse aérienne n’est pas une estimation
fiable de la productivité primaire nette de l’écosystème : d’importants flux cachés ne sont pas
détectés par les suivis de biomasse car les processus de production, sénescence, chute de
litière et décomposition ont lieu simultanément.
La durée optimale de la jachère fait l’objet de compromis entre les différentes
fonctions agro-écologiques de la jachère : constitution d’un stock de matière organique,
fixation symbiotique d’azote, qualité de la litière incorporée au sol, lutte contre les adventices,
limitation des parasites, stimulation de l’activité microbienne, modification de la structure du
sol. Elle n’est pas nécessairement très longue au regard des critères testés avec le modèle sur
les données vénézuéliennes : le stock de matière organique n’augmente plus au-delà de 5 ans
et c’est au cours de cette période qu’on a la fixation maximale d’azote avec le pic des
légumineuses. La jachère améliorée par semis de légumineuses offre de fortes potentialités
(qu’il faudrait modérer en incluant une éventuelle limitation pour le phosphore dans le
modèle). Le couplage aux études de décomposition de la matière organique devrait permettre
de définir la composition idéale de la litière et la durée optimale de la jachère pour ce critère.
D’autres idées pourront être testées, comme la fertilisation de la jachère ou la manipulation
par arrachage et prélèvements.
Le développement du modèle et l’analyse des résultats incitent à compléter les études
entreprises sur les sites d’applications, en particulier, ce qui concerne :
(1) le cycle reproductif des espèces considérées (banque et pluies de graines, dispersion,
dormance et recrutement), car la dynamique de la communauté et le fonctionnement de
l’écosystème sont fortement sensibles aux conditions initiales ;
(2) le bilan hydrique et le bilan d’azote pendant la jachère (apports par déposition et fixation,
et départs par lessivage), car la dynamique de la communauté et le fonctionnement de
l’écosystème sont fortement sensibles au niveau de ressources disponibles (eau et azote) ;
(3) le cycle du phosphore et les limitations de la fixation symbiotique, car celle-ci constitue la
principale entrée d’azote dans le système ;
(4) l’estimation de l’activité racinaire, représentée dans le modèle par SRL (Specific root
length, longueur de racines et de mycorhizes par unité de masse), car la compétition en azote
joue un rôle prépondérant dans la structuration de la communauté, notamment dans les
milieux pauvres en nutriments ;
(5) la structure du couvert, car elle détermine la compétition pour la lumière et est un facteur
déterminant dans l’exclusion des espèces pionnières ;

83
(6) la durée de vie des organes, car le taux de turn-over joue un rôle prépondérant dans la
dynamique du système ;
(7) le bilan de carbone à l’échelle de la parcelle (par mesures sous cloches ou par tour à flux),
car il permettra de valider le fonctionnement global du modèle.
3.1.2 - Critique des hypothèses et perspectives de prolongement
L’objectif du modèle était de prédire sur plusieurs décennies la dynamique de la
communauté et le fonctionnement de l’écosystème à partir des traits morphologiques et
écophysiologiques des espèces qui avaient été mesurés sur le terrain. Pour parvenir à cet
objectif, nous avons formulé de nombreuses hypothèses simplificatrices. Nous reprenons ici
ces hypothèses en montrant quelles en sont les limites et quel champ de l’écologie s’applique
à les étudier.
Limites du modèle
Dimension spatiale
Nous avons développé le modèle sur la base des mécanismes écophysiologiques et de
ce fait, volontairement réduit les aspects spatiaux du problème à la répartition verticale des
espèces dans le couvert végétal.
Nous négligeons ainsi la répartition des espèces dans le plan horizontal. Des modèles
3D ont été développés pour une plante unique ou pour des couverts réguliers monospécifiques
(vigne) ou bispécifique (herbe et trèfle, Sinoquet 1998). Ces modèles prédisent l’extinction du
rayonnement et l’assimilation photosynthétique du couvert. Mais ils nécessitent la description
complète de la scène (position de toutes les plantes, voire de tous les organes) et ils travaillent
généralement sur une scène fixe (qui n’est pas mise à jour à chaque pas de calcul). Le modèle
FAPROM assimile le couvert à une « soupe végétale » (hypothèse « champ moyen ») mais
prédit la dynamique de la communauté au pas de temps journalier sur plusieurs décennies.
Nous simplifions aussi excessivement la dimension spatiale à l’échelle du paysage en
supposant constant au cours du temps l’apport de graines en provenance des parcelles
voisines. Pour pouvoir simuler la diversité à l’échelle du paysage, il faudrait coupler le
modèle à une base de données géographiques et étudier les flux de graines entre parcelles.
Dans le but de simuler la dynamique de la diversité à l’échelle de l’écorégion, il peut s’avérer

84
intéressant d’utiliser au sein de chaque parcelle un modèle mécaniste simplifié de succession
secondaire, et un échiquier pour décrire les relations entre les différentes parcelles.
Paramètres constants, dimension temporelle
Nous avons supposé des paramètres constants par saison (parfois identiques sur les
deux saisons) et constants sur la durée complète de la simulation. Nous supposons une classe
d’âge unique par espèce, modélisons chaque espèce comme un individu moyen et négligeons
les éventuels stades de développement des plantes individuelles. Pour les concentrations en
azote par exemple, un modèle plus fin considèrerait plus précisément la phénologie et les
stades de développement et de croissance des différentes espèces. Nous avons axé notre
travail sur la compétition interspécifique et les mécanismes de la succession végétale.
Nous n’étudions pas non plus la plasticité phénotypique. Par exemple, nous supposons
toujours la même structure verticale du couvert pour une espèce donnée, quel que soit son
environnement. Certains modèles intègrent la morphologie des plantes et la plasticité des
caractères morphologiques ou écophysiologiques (Soussana et al. 2002, Dingkuhn et al.
2002) ; en revanche, ils ne simulent pas plus de deux espèces en interaction.
Nous n’avons pas intégré de dépendance de la photosynthèse à la température ni à la
teneur en CO2 de l’air. Nous ne pouvons donc pas simuler la réponse de l’écosystème aux
changements globaux. Pour aller plus loin dans cette direction, il faudrait en particulier
mesurer les courbes de réponse de la photosynthèse foliaire, de la respiration et de la
répartition des assimilats, à la température et au CO2. D’une manière générale, tout
raffinement demande d’introduire de nouveaux paramètres qui nécessitent donc de nouvelles
études sur le terrain pour les déterminer. Nous avons construit le modèle en essayant d’utiliser
toute l’information disponible sur les sites d’études, et dans la mesure du possible, en nous
limitant à cette information.
Nous focalisons notre étude sur une échelle de temps relativement courte au regard de
l’évolution des espèces et ne traitons donc pas des aspects évolutifs dans la dynamique de la
communauté.
Cycles du carbone et de l’azote
Le modèle décrit les cycles du carbone et de l’azote. Certains processus
écophysiologiques (comme la fixation symbiotique) sont vraisemblablement limités par
d’autres nutriments (P, K, Mg, Ca …). Il serait intéressant de tester (peut-être sur un modèle
simplifié dans un premier temps) quel est l’impact de la prise en compte du cycle d’un

85
troisième élément dans la dynamique de la communauté. Cet ajout au modèle nécessiterait
également un complément d’études sur le terrain.
Interactions entre le monde végétal et les animaux
Mis à part les simulations effectuées pour tester l’impact du pâturage sur la dynamique
de la communauté et le fonctionnement de l’écosystème, nous négligeons les interactions
entre le monde végétal et les animaux, en particulier : (1) les liens entre diversité aérienne et
diversité souterraine (nématodes, micro et macrofaune du sol, Wardle 2002, De Deyn et al.
2004) ; (2) les petits herbivores aériens (insectes, petits rongeurs…) ; les pollinisateurs et les
animaux qui interviennent dans le cycle reproductif des espèces (Molinillo & Farji Brener
1993 ont montré l’importance du bétail dans la dispersion des graines d’Acaena elongata).
Les étapes de validation du modèle, confrontation des simulations aux relevés de
terrain, peuvent mettre en évidence les limites des différentes hypothèses. Par exemple, le
suivi de biomasse des parcelles semées en lupin peut montrer l’importance de la limitation de
la fixation symbiotique et dans quelle mesure notre hypothèse de modélisation est capable de
reproduire la réalité.
Quel modèle choisir (bis) ?
A chaque question, son modèle. L’objet du modèle FAPROM est d’établir les liens
mécanistes entre les paramètres statiques (les traits des espèces mesurés sur le terrain), la
dynamique de la communauté et le fonctionnement de l’écosystème au cours de la jachère. Un
modèle mécaniste simplifié peut être utilisé pour aborder des questions qui nécessitent de
changer d’échelle spatiale (écologie du paysage), temporelle (évolution) ou thématique
(interactions trophiques, liens entre diversité aérienne et diversité souterraine). Les modèles
globaux plus théoriques, du type logistique de Verhulst ou Lotka-Volterra pour les
interactions entre espèces, permettent de dégager des tendances (stratégies r, K par exemple)
et d’étudier les interactions simples entre un nombre limité de paramètres. Les approches
globale (description du système dans son ensemble) et réductionniste (description du système
du point de vue de ses constituants) sont complémentaires. Suivant les questions qu’on met en
avant, il faudra pousser la description dans les détails ou au contraire la simplifier au
maximum.

86
3.2 - RETOUR SUR LE PROJET DE THESE
Nous revenons maintenant sur le déroulement du travail en analysant les aspects
positifs et les problèmes rencontrés dans le cadre du projet TROPANDES.
3.2.1 - Dynamique du projet TROPANDES, travail en équipe
Le projet TROPANDES fut une expérience humaine et polyglotte très enrichissante et
nous espérons que les collaborations entreprises seront durables. Nous avons particulièrement
apprécié le travail en équipe que nous avons développé avec les différents partenaires autour
des trois axes suivants : (1) l’application aux parcelles boliviennes ; (2) l’application aux
parcelles vénézuéliennes ; et (3) le couplage des modèles. Chacun des échanges que nous
avons eu a été fructueux et les questions des différents partenaires ont stimulé le travail. Le
travail en collaboration avec les équipes boliviennes et vénézuéliennes (en particulier Teresa
Ortuño en Bolivie, et Lina Sarmiento au Venezuela) a permis d’améliorer le modèle
(représentation de la respiration, de la fixation symbiotique, adaptation du modèle aux
données boliviennes). Le travail avec le groupe des modélisateurs (en particulier Marc Pansu
et Pierre Bottner à Montpellier et Klaas Metselaar à Wageningen) a permis d’avancer sur le
bilan d’azote à l’échelle de la parcelle.
Nous retiendrons l’importance des relations humaines, des contacts et de la confiance
dans un tel projet. A l’heure des nouvelles technologies, le courrier électronique simplifie
beaucoup les échanges matériels d’informations mais rien ne remplace les rencontres, les
discussions en face-à-face et les collaborations main dans la main pour faire avancer le travail
en équipe, d’autant plus quand les équipes sont multidisciplinaires et multiculturelles.
3.2.2 - Evaluation critique du travail
Dans quelle mesure les objectifs ont-ils été atteints ?
Dans le cadre du projet TROPANDES, nous étions chargé de modéliser, pour les sites
vénézuélien et bolivien, la dynamique de la production végétale au cours de la succession
post-culturale. La proposition du projet définissait les tâches suivantes : (1) le développement
d’un modèle générique de production de la jachère ; (2) la paramétrisation du modèle pour les

87
sites d’études particuliers ; (3) la validation du modèle ; (4) le couplage des modèles
biogéochimiques à l’échelle de la parcelle ; et (5) la compréhension des mécanismes de
restauration de la fertilité des sols dans le système de culture à jachères longues des hautes
Andes tropicales. Nous n’avons que partiellement rempli notre mission : si les objectifs 1 et 2
sont atteints, les objectifs 3, 4 et 5 restent inachevés.
Notre projet de thèse avait l’ambition d’inclure le modèle de parcelle dans le modèle
d’exploitation agricole développé par Bernardo Paz-Betancourt (1997) et de réunir les aspects
écologiques, agronomiques et socio-économiques du problème. Bien que nous ayons gardé ce
but en vue, nous avons rapidement revu nos objectifs à la baisse et seulement tenté de
modéliser le fonctionnement agro-écologique de la parcelle.
Analyse des difficultés rencontrées
Nous analysons dans la suite de cette partie les difficultés auxquelles nous avons été
confrontés et les disfonctionnements du projet.
Développement informatique
Nous avons initialement développé le modèle sous FORTRAN puisque tel était le
langage utilisé au laboratoire. Nous avons ensuite traduit le modèle (et poursuivi son
développement) sous VENSIM de façon à le rendre directement compatible aux autres
modèles développés dans le projet. Si c’était à refaire, quelle solution choisirions-nous ? Les
langages de programmation informatique (tels que FORTRAN) présentent l’avantage de ne
pas être limités dans leurs possibilités techniques (encore faut-il que le programmeur ait
suffisamment de dextérité). Mais ils ne sont pas vraiment ergonomiques, et sont difficilement
partageables, difficilement modifiables par un tiers non spécialiste, et nécessitent l’écriture
des fichiers de sorties dont on veut disposer. A contrario, les logiciels de programmation
graphique des systèmes dynamiques (tels que VENSIM) sont ergonomiques, intuitifs (c’est-à-
dire qu’ils fonctionnent comme les produits Microsoft) donc facilement utilisables et
modifiables par d’autres utilisateurs. D’autre part, toutes les sorties de simulation sont
disponibles par défaut, et le logiciel offre un certain nombre d’utilitaires comme une
plateforme de macro simulations (pour des études de sensibilité et des tirages de Monte-
Carlo) et de nombreuses fonctions mathématiques intégrées ( notamment les tirages aléatoires
suivant de multiples lois). Toutefois, il n’est pas possible de faire des boucles imbriquées et
d’appeler des sous-modèles en cours de calcul, ni d’augmenter le nombre de strates ou
d’espèces au cours d’une simulation. Aussi la solution à adopter dépend-elle de l’objectif

88
fixé : VENSIM est un bon compromis pour un modèle didactique permettant l’échange entre
chercheurs non modélisateurs. Pour un modélisateur également, l’interface de VENSIM
permet de visualiser rapidement le fonctionnement d’un modèle et d’en saisir d’un coup
d’oeil l’architecture globale. Mais pour développer des modèles plus complexes, comme le
modèle socio-économique du fonctionnement du système de culture à jachères longues à
l’échelle de la vallée, un langage de programmation sera nettement plus efficace. En effet,
sous VENSIM, pour modéliser l’alternance culture/jachère, nous avons été amenés à
additionner des zéros pour le modèle de jachère pendant la période de culture, et vice versa.
Au cours du développement du modèle, on a parfois l’impression de refaire sans arrêt
les mêmes simulations : à chaque fois que l’on modifie une équation ou qu’on affine
l’estimation d’un paramètre, on doit recommencer l’étude du comportement du modèle. Il
faudrait tout simplement pouvoir tout faire à la fin ! Mais on a besoin de résultats au cours du
développement et c’est à partir de ces résultats et de leur discussion qu’on améliore la
version courante du modèle.
Gestion de projet
Notre travail s’est intégré dans le cadre du projet TROPANDES mais il a commencé
au milieu du projet, alors que les principales orientations avaient été prises, avec leurs
conséquences sur le travail de terrain. Nous avons donc dû pour l’essentiel nous contenter des
mesures effectuées, sans pouvoir réellement interagir avec les expérimentateurs pour
compléter ces mesures dans un sens utile à notre modélisation. Ainsi le modèle FAPROM
nous a révélé l’importance des paramètres de mortalité et de reproduction (production et
dissémination des graines) mais nous avons dû déterminer ces paramètres par calibration du
modèle, faute d’en avoir des mesures sur le terrain. Pour valider notre modèle, il nous aurait
fallu un suivi aussi complet que possible de la biomasse par espèce tout au long de la
succession. Certaines de ces données existent mais n’ont pas encore été dépouillées. Elles
nous sont restées inaccessibles, faute d’avoir réussi, en dépit de nos efforts répétés, à
convaincre les expérimentateurs de l’intérêt de collecter ces mesures ou de les exploiter en
commun pour en tirer le meilleur parti.
Il faut reconnaître que le travail de terrain est lourd et contraignant. Ainsi, un suivi de
biomasse sur 6 parcelles pendant 2 années au Venezuela nécessite trois années complètes de
travail pour une personne (cf. annexe 2). On peut comprendre que les expérimentateurs aient
quelque réticence à ajouter des mesures à leur protocole et à laisser d’autres personnes

89
exploiter leurs données. Mais le projet dans son ensemble aurait gagné à ce que les
discussions entre modélisateurs et expérimentateurs aient lieu assez tôt dans le projet (ce fut
partiellement le cas), et surtout aient continué de façon régulière au fur et à mesure que les
données arrivaient et que les modèles progressaient. Peut-être ce dialogue permanent n’est-t-il
possible que lorsque le modélisateur est intégré dans une équipe d’expérimentateurs ; nos
courrier électroniques, complétés par plusieurs visites, ont permis d’avancer dans l’échange
mais de nombreuses données restent à exploiter.
C’est en couplant les modèles de jachère, de culture et de matière organique du sol que
nous avons réalisé l’importance du bilan d’azote, pour les modèles comme pour les mesures.
La première version du modèle de matière organique du sol donnait des flux d’azote minéral
beaucoup trop élevés qui modifiaient de façon irréaliste la dynamique de succession pendant
la jachère et qui n’étaient pas limitants pour la culture. Nous nous sommes alors tournés vers
les mesures pour valider ce bilan d’azote, ce qui a été possible au Venezuela mais pas en
Bolivie où il manquait les pertes d’azote par lessivage.
Couplage des modèles
Beaucoup d’énergie a été dépensée dans le couplage des modèles. Un premier
couplage opérationnel a été réalisé relativement rapidement mais la mise à jour séparée des
modèles a nécessité de refaire le couplage à plusieurs reprises. Le couplage a révélé des
différences d’approche de modélisation et des points d’intérêt non systématiquement
convergents. Par exemple, le modèle agronomique est basé sur le calcul des efficiences
d’utilisation de la lumière, de l’eau et de l’azote et calcule tout en kg ha-1 de matière sèche
puis définit les contenus de carbone et d’azote ; alors que le modèle écologique décrit les
cycles du carbone et de l’azote et calcule tous les stocks en g m-2 et les flux en g m-2 d-1. Il a
donc fallu ajouter une vue de synthèse qui permettent d’échanger les variables entre les
différents sous-modèles, vérifier l’homogénéité des unités et compléter le bilan d’azote
minéral à l’échelle de la parcelle.
A ce jour, le modèle couplé n’est toujours pas opérationnel : il nécessite encore la
validation complète de chacun des éléments et la validation globale du modèle de
fonctionnement de la parcelle sur un cycle culture / jachère.

90
3.3 - REFLEXIONS PERSONNELLES ET PERSPECTIVES
3.3.1 - De la multiplicité des métiers du chercheur
Je quitte quelques instants le nous conventionnel pour analyser la façon dont j’ai vécu
ce travail de thèse et la double dimension ingénierie / recherche du projet.
Ce travail de thèse aurait requis les compétences d’un chercheur, d’un ingénieur, d’un
programmeur, d’un consultant, d’un enseignant, d’un rédacteur, d’un graphiste et d’un
voyageur. La variété des activités du recherche fait sans doute la richesse de ce métier mais
l’aide de « professionnels » aurait souvent amélioré l’efficacité du travail. Une des
compétences acquise au cours de la thèse est de savoir repérer des amateurs plus éclairés
dans les domaines abordés et de tisser parmi eux son réseau de conseillers : un
mathématicien, un statisticien, un programmeur, un informaticien, un physiologiste, un
lecteur critique, un béotien, un assistant conception graphique et un traducteur spécialisé.
Le travail mené relève à la fois de l’ingénierie et de la recherche : le développement
du modèle nécessite de mettre les mains dans le cambouis, de se fixer des objectifs et d’être
pragmatique (tout paramètre se doit d’être identifié) ; la recherche scientifique avance en
posant sans cesse de nouvelles questions, dans un état de remise en cause permanent. Je
regrette de n’avoir pas su me détacher davantage de l’aspect technique du travail et de
n’avoir pas toujours su prendre du recul par rapport au développement du modèle, pour le
considérer réellement comme un outil et l’utiliser pour analyser les questions scientifiques. Il
faut savoir abandonner par moment le développement de l’outil pour pouvoir s’en servir
sereinement. Et il faut s’en servir pour pouvoir améliorer son développement. Quand le
concepteur et l’utilisateur sont bel et bien la même personne, il est difficile de mettre des
frontières entre ces deux activités.
3.3.2 - Avenir du modèle FAPROM
Nous terminons notre synthèse en évoquant l’avenir du modèle FAPROM : poursuite
de la validation sur les données vénézuéliennes, application à de nouveaux sites, les friches
méditerranéennes par exemple, et utilisation du modèle pour tester les relations entre diversité
des communautés et fonctionnement des écosystèmes.

91
Validation complète sur les données vénézuéliennes
Le modèle FAPROM devrait être utilisé par le partenaire vénézuélien qui pourra le
valider avec les séries de données diachroniques de biovolume. Dans ce but, nous animons en
juillet 2004 à l’Universidad de los Andes (Mérida, Venezuela) un atelier de modélisation qui a
pour double objectif de familiariser les participants à la modélisation des systèmes
dynamiques en écologie et de transmettre la dernière version du modèle FAPROM. Nous
espérons que le partenaire vénézuélien utilisera le modèle FAPROM, qu’il en poursuivra le
développement et que notre collaboration se poursuivra longtemps.
Adaptation aux écosystèmes méditerranéens
Eric Garnier (directeur de recherche CNRS au CEFE à Montpellier) a proposé en 2004
un sujet de thèse qui vise à étudier les conséquences écologiques de la déprise agricole en
région méditerranéenne, dans le cas de l’abandon progressif de la vigne au cours du 20ème
siècle. Cet abandon a créé une mosaïque d’habitats, correspondant à des parcelles
abandonnées à des moments divers représentant des états successionnels différents. L’objectif
est d’élaborer un outil de prédiction des patrons de succession végétale et des conséquences
sur le fonctionnement des écosystèmes en adaptant le modèle FAPROM au milieu
méditerranéen. Le modèle devrait être paramétré pour une vingtaine d’espèces végétales,
grâce aux données recueillies par l’équipe ECOPAR du CEFE dans un système de douze
friches proches de Montpellier et à des expérimentations complémentaires, notamment sur la
niche de régénération.
Utilisation du modèle dans l’étude des liens entre diversité et fonctionnement
Au cours de la dernière décennie, de nombreuses expérimentations ont eu pour
objectif de tester les relations entre diversité des communautés et fonctionnement des
écosystèmes. De nombreuses expériences ont utilisé des communautés assemblées au hasard
pour montrer que la richesse spécifique et la richesse en groupes fonctionnels des plantes ont
un effet positif sur la production primaire et la rétention des nutriments. Les expériences du
projet BIODEPTH (Hector et al. 1999) ont démontré des effets positifs de la diversité
végétale sur la production de biomasse aérienne sur un ensemble de 8 sites en Europe. Mais
l’interprétation de ces expériences a fait l’objet d’une controverse (Huston et al. 2000, Hector
et al. 2000) du fait que leurs résultats peuvent être générés par différents mécanismes.
Ce type d’expériences présente un jeu de données rêvé pour la validation du modèle
FAPROM : les suivis de biomasses de monocultures et de mélanges en conditions semi-

92
contrôlées. L’utilisation du modèle en contrepartie peut permettre de faire l’étude exhaustive
de toutes les combinaisons d’assemblages d’espèces pour contrôler l’effet d’échantillonnage.
Le modèle permet en outre d’estimer la production primaire nette et non pas seulement la
biomasse aérienne. Le modèle peut être intéressant pour généraliser les résultats de ces
expériences. Des contacts préliminaires ont été pris avec Nina Buchmann (expérimentations à
l’Institut Max Planck, Jena, Allemagne).

93
Conclusion

94
Nous souhaitons conclure en mettant en exergue les fruits de notre travail, les résultats
de notre recherche et les idées que nous souhaitons faire passer. Ces éléments de conclusion
rassemblent deux types de résultats : des résultats d’ordre méthodologique ou technique (1-4)
et des résultats centrés sur notre problématique, la gestion durable du système de culture à
jachère des hautes Andes tropicales (5-8).
(1) Nous avons développé un modèle biogéochimique de succession végétale qui lie
écophysiologie et écologie des communautés. Son originalité est d’assurer le couplage entre
processus instantanés et dynamique de la végétation et d’intégrer tous les compartiments
mesurés en identifiant des liens mécanistes entre eux. Il permet à la fois d’établir la réponse
des végétaux au cours de la succession et d’étudier l’effet des végétaux sur le fonctionnement
des écosystèmes. Disponible sur demande25, la dernière version est épurée et documentée.
(2) Nous avons mis en évidence l’existence de flux cachés de l’écosystème et montré que la
biomasse aérienne n’est pas une estimation fiable de la productivité primaire nette de
l’écosystème, en particulier, dans les écosystèmes herbacés tropicaux au turn-over rapide.
(3) Le modèle montre l’importance de prendre en compte le turn-over de la matière organique
dans la modélisation des dynamiques écologiques :
• A l’échelle d’une espèce, le turn-over est responsable des décalages temporels et se traduit
dans l’asymétrie des courbes de croissance.
• A l’échelle de la communauté, le turn-over des espèces joue un rôle prépondérant dans les
processus de compétition, en particulier dans les milieux pauvres en nutriments.
• A l’échelle de la parcelle, le turn-over est impliqué dans la dynamique de la fertilité du sol.
(4) Nous avons montré l’intérêt des modèles de simulations en complément des modèles
d’analyse mathématique (caractérisant les équilibres) pour comprendre la dynamique des
systèmes complexes. La prise en compte des effets indirects conduit parfois à des résultats
contre intuitifs (comme la baisse de production lorsque les ressources augmentent, cf. p.63).
La modélisation multi-agent des systèmes complexes offre des perspectives intéressantes dans
l’étude des dynamiques écologiques.

95
(5) La durée optimale de la jachère fait l’objet de compromis entre les différentes fonctions de
la jachère : constitution d’un stock de matière organique, fixation symbiotique d’azote, qualité
de la litière incorporée dans le sol, lutte contre les adventices. Elle n’est pas nécessairement
très longue pour son rôle agro-écologique26 mais d’autres usages (comme la production de
bois de feu en Bolivie) peuvent requérir plus de temps.
(6) La jachère améliorée par semis de légumineuses offre de fortes potentialités (mais il
faudrait inclure le cycle du phosphore dans le modèle pour pouvoir prédire l’efficacité réelle
de la fixation).
(7) Le modèle FAPROM est fortement sensible aux conditions initiales ce qui doit encourager
l’étude des cycles reproductifs des espèces considérées (banque de graines, dormance et
pluies de graines).
(8) Le développement du modèle et l’analyse des résultats incitent à compléter les études
entreprises sur les sites d’applications, en particulier, en ce qui concerne les graines, la
mortalité, l’estimation du SRL (longueur de racine par unité de masse), la fixation
symbiotique et le cycle du phosphore, la structure du couvert, le bilan hydrique et le bilan
d’azote pendant la jachère, et le bilan de carbone à l’échelle de l’écosystème.
26 Les simulations montrent une stabilisation de la biomasse totale après 5 ans de jachère. Le modèle requiert encore néanmoins une validation plus approfondie pour améliorer la confiance dans les prédictions, notamment sur la vitesse d’établissement de la végétation.

96
Références bibliographiques

97
Abadin J, Gonzalez-Prieto SJ, Sarmiento L, Villar MC, Carballas T. 2002. Successional dynamics in a long fallow agricultural system of the high tropical Andes. Soil Biology & Biochemistry 34: 1739-1748.
Aerts R. 1999. Interspecific competition in natural plant communities: mechanisms, trade-
offs, and plant-soil feedbacks. Journal of Experimental Botany 50: 29-37. Bachelet D, Lenihan JM, Daly C, Neilson RP, Ojima DS, Parton WJ. 2001. MC1, a dynamic
vegetation model for estimating the distribution of vegetation and associated carbon and nutrient fluxes. Technical Documentation Version 1.0. USDA F.S. General Technical Report PNW-GTR-508. Corvallis, OR. USDA Forest Service, Pacific Northwest Research Station.
Berbesi N. 1990. Estrategias de asignacion de biomasa y nutrientes en plantas del paramo
andino, en un gradiente sucesional y sus variaciones estacionales [Tesis de doctorado]. Merida (Venezuela): Facultad de Ciencias, Universidad de los Andes. 148 pp.
Bertalanffy L von. 1938. A quantitative theory of organic growth. Human Biology 10: 181-
213. Birch CPD. 1999. A new generalized logistic sigmoid growth equation compared with the
Richards growth equation. Annals of Botany 83: 713-723. Blatt SE, Janmaat JA, Harmsen R. 2001. Modelling succession to include an herbivore effect.
Ecological Modelling 139:123-136. Bonabeau E. 2002. Agent-based modeling: methods and techniques for simulating human
systems. Proceedings of the National Academy of Sciences 99: 7280-7287. Bonhomme R, Varlet-Grancher C. 1977. Application aux couverts végétaux des lois de
rayonnement en milieu diffusant I. Etablissement des lois et vérifications expérimentales. Annales agronomiques 28: 567-582.
Bugmann H. 2001. A review of forest gap models. Climatic Change 51: 259-305. Caldwell RM. 1995. Simulation models for intercropping systems. Sinoquet H, Cruz P,
editors. Ecophysiology of tropical intercropping. INRA Editions, Paris (France). 353-368.
Camacho, M. 2001. La gestión del espacio y las prácticas de manejo del suelo en la región
altiplánica de Bolivia: el caso de la comunidad de Patarani en la provincia Aroma [Tesis para optar al grado de Magister Sciencitae en Ecología y Conservación]. La Paz (Bolivia): Universidad Mayor de San Andres. 82 pp.
Cannell MGR, Thornley JHM. 2000. Modelling the components of plant respiration: some
guiding principles. Annals of Botany 85: 45-54. Capra F. 1996. The web of life: a new synthesis of mind and matter. Harper Collins
publishers, England. 320 pp.

98
Carballas T, Monasterio M, Beck S, Bottner P, Metselaar K, Saugier B, Hervé D. 2002a. Fertility management in the tropical Andean mountains: agroecological bases for a sustainable fallow agriculture (TROPANDES). Carballas T, coordinator. Final Report. CE, INCO-DC, Contract number: ERBIC18CT98-0263. 40 pp.
Carballas T, Monasterio M, Beck S, Bottner P, Metselaar K, Saugier B, Hervé D. 2002b.
Fertility management in the tropical Andean mountains: agroecological bases for a sustainable fallow agriculture (TROPANDES). Final Individual Report. CE, INCO-DC, Contract number: ERBIC18CT98-0263, 1998-2002. 99 pp.
Clements FE. 1916. Plant succession: an analysis of the development of vegetation.
Washington DC: Carnegie Institution of Washington DC. Publication number 242. 512 pp.
Coquillard P, Hill D. 1997. Modélisation et simulation d’écosystèmes. Masson: Paris. 273pp. Courchamp F, Woodroffe R, Roemer G. 2003. Removing protected populations to save
endangered species. Science 302: 1532. Coûteaux MM, Hervé D, Beck S. 2004. Decomposition of plant litter and roots in a long
fallow system (Bolivian Altiplano). Ecologia en Bolivia (sous presse). Coûteaux MM, Sarmiento L. 2004. Chemical composition control of above and belowground
plant litter decomposition in an old-field succession of the high Venezuelan Andes. En préparation.
Crews TE, Peoples MB. 2004. Legume versus fertilizer sources of nitrogen: ecological
tradeoffs and human needs. Agriculture, Ecosystems and Environment 102: 279-297. Daalen JC, Shugart HH. 1989. OUTENIQUA – A computer model to simulate succession in
the mixed evergreen forests of the southern Cape, South Africa. Landscape Ecology 2: 255-267.
Davi H, Deredec A, Martineau Y. 2003. Modélisation en écologie. Cours UVE, module
MMB, Université Paris-Sud. 77 pp. De Deyn GB, Raaijmakers CE, Zoomer HR, Berg MP, De Ruiter PC, Verhoef HA, Bezemer
TM, Van der Putten WH. 2003. Soil invertebrate fauna enhances grassland succession and diversity. Nature 422: 71-713.
Dingkuhn M, Baron C, Bonnal V, Combres JC, Mialet I. 2002. SARRAH v2.3 - Manuel
Utilisateur. CIRAD. 53 pp. Doré T, Dalbiès A. 1994. Retrait des terres et systèmes de culture. Le courrier de
l’environnement de l’INRA 21: 27-34. Dufrêne E, Davi H, François C, Le Maire G, Le Dantec V, Granier A. 2004. Modelling
carbon and water cycles in a Beech forest. Part I: Model description and uncertainty analysis on modelled NEE. Submitted in Ecological Modelling.

99
Ehrlen J, Groenendael JM van.1998. The trade-off between dispersibility and longevity: an important aspect of plant species diversity. Applied Vegetation Science 1: 29-36.
Facelli JM, Pickett STA. 1990. Markovian chains and the role of history in succession. Trends
in Ecology and Evolution 5: 27-30. Fagerström T. 1987. On theory, data and mathematics in ecology. Oikos 50: 258-261. Floret C, Pontanier R. 2000. La jachère en Afrique tropicale, rôles, aménagement,
alternatives. Volume 1. Actes du séminaire international, Dakar 13-16 avril 1999. John Libbey Eurotext (Paris). 777 pp.
Fontaine M. 2000. Caractéristiques écophysiologiques de quelques espèces d'une succession
végétale dans les Andes vénézuéliennes [Rapport de DEA]. Universités Paris VI, Paris XI, INAPG. Orsay (France). 42 pp.
Frelich LE, Reich PB. 1995. Spatial patterns and succession in a Minnesota southern-boreal
forest. Ecological Monographs 65: 325-346. Garnier E, Cortez J, Billès G, Navas ML, Roumet C, Debussche M, Laurent G, Blanchard A,
Aubry D, Bellmann A, Neill C, Toussaint JP. 2004. Plant functional markers capture ecosystem properties during secondary succession. Ecology: sous presse.
Gilmanov TG, Verma SB, Sims PL, Meyers TP, Bradford JA, Burba GG, Suyker AE. 2003.
Gross primary production and light response parameters of four Southern Plains ecosystems estimated using long-term CO2-flux tower measurements. Global Biogeochemical Cycles 17: 1071.
Ginzburg LR, Jensen CXJ. 2004. Rules of thumb for judging ecological theories. Trends in
Ecology and Evolution 19: 121-126. Gompertz B. 1825. On the nature of the function expressive of the law of human mortality,
and on a new mode of determining the value of life contingencies. Philosophical Transactions of The Royal Society (London) 123: 513-585.
Greenwood DJ, Lemaire G, Gosse G, Cruz P, Draycott A, Neeteson JJ. 1990. Decline in
percentage N of C3 and C4 crops with increasing plant mass. Annals of Botany 66: 425-436.
Grime JP. 1974. Vegetation classification by reference to strategies. Nature 250: 26-31. Grime JP. 1988. The C-S-R model of primary plant strategies-origins, implications and tests.
Gottlieb LD, Jain SK, editors. Plant evolutionary biology. New York (USA): Chapman and Hall. Ch. 14, p371-389.
Grime JP. 1998. Benefits of plant diversity to ecosystems: immediate, filter and founder
effects. Journal of Ecology 86: 902-910. Grime JP. 2001. Plant Strategies, Vegetation Processes, and Ecosystem Properties (2nd
edition). John Wiley & Sons LTD. 417 pp.

100
Gu L, Baldocchi DD, Wofsy SC, Munger JW, Michalsky JJ, Urbanski SP, Boden TA. 2003.
Response of a deciduous forest to the mount Pinatubo eruption: enhanced photosynthesis. Science 299: 2035-2038.
Gutiérrez A. 1996. Plan estratégico de desarollo agricola del Estado Mérida (PEDEM).
Documento sobre el sector agricola. Centro de Investigaciones Agroalimentarias, Universidad de los Andes, Mérida, Venezuela.
Hastings A. 1980. Disturbance, Coexistence, history and competition for space. Theoretical
Population Biology 18: 363-373. Hector A, Schmid B, Beierkuhnlein C, Caldeira MC, Diemer M, Dimitrakopoulos PG, Finn J,
Freitas H, Giller PS, Good J, Harris R, Högberg P, Huss-Danell K, Joshi J, Jumpponen, A., Körner C, Leadley PW, Loreau M, Minns A, Mulder CPH, O'Donovan G, Otway SJ, Pereira JS, Prinz A, Read DJ, Scherer-Lorenzen M, Schulze ED, Siamantziouras ASD, Spehn EM, Terry AC, Troumbis AY, Woodward FI, Yachi S, Lawton JH. 1999. Plant diversity and productivity experiments in European grasslands. Science 286: 1123-1127.
Hector A, Schmid B, Beierkuhnlein C, Caldeira MC, Diemer M, Dimitrakopoulos PG, Finn J,
Freitas H, Giller PS, Good J, Harris R, Högberg P, Huss-Danell K, Joshi J, Jumpponen, A., Körner C, Leadley PW, Loreau M, Minns A, Mulder CPH, O'Donovan G, Otway SJ, Pereira JS, Prinz A, Read DJ, Scherer-Lorenzen M, Schulze ED, Siamantziouras ASD, Spehn EM, Terry AC, Troumbis AY, Woodward FI, Yachi S, Lawton JH. 2000. No consistent effect of plant diversity on productivity? Response. Science 289: 1255.
Hervé D. 1994. Desarrollo sostenible en los Andes Altos: los sistemas de cultivos con
descanso largo pastoreado. Hervé D, Génin D, Rivière G, editors. Dinamicas del descanso de la tierra en los Andes. La Paz: IBTA-ORSTOM. 155-169.
Hervé D, Paz Betancourt B, Migueis J, Treuil JP. 2003. Introduction de la modélisation dans
une recherche interdisciplinaire : état et gestion des jachères dans les Andes. Nature Sciences Sociétés 11 : 243-254.
Hervé D, Rivière G. 1998. Les jachères longues pâturées dans les Andes. Acquis
interdisciplinaires. Nature Sciences Sociétés 6 : 5-19. Huston MA, Aarssen LW, Austin MP, Cade BS, Fridley JD, Garnier E, Grime JP, Hodgson J,
Lauenroth WK, Thompson K, Vandermeer JH, Wardle DA. No consistent effect of plant diversity on productivity. Science 289: 1255.
Inouye RS, Tilman D. 1995. Convergence and divergence of old-field vegetation after 11
years of nitrogen addition. Ecology 76: 1872-1887. Lauenroth WK. 2000. Methods of estimating belowground net primary production. In:
Methods in Ecosystem Science (eds Sala OE, Jackson RB, Mooney HA, Howarth RW). Springer. New York. 58-71.
Lavigne C, Devaux C, Deville A, Garnier A, Klein EK, Lecomte J, Pivard S, Gouyon PH.
2004. Potential and limits of modelling to predict the impact of transgenic crops in wild

101
species. In: den Nijs HCM, Bartsch D, Sweet J (eds). Introgression from genetically modified plants into wild relatives, CABI Publishing, Oxon, UK.
Lavorel S, Garnier E. 2002. Predicting changes in community composition and ecosystem
functioning from plant traits: revisiting the Holy Grail. Functional Ecology 16: 545-556. Leguerrier D, Niquil N, Boileau N, Rzeznik J, Sauriau PG, Le Moine O, Bacher C. 2003.
Food web numerical analysis of an intertidal mudflat ecosystem: Brouage, Marennes-Oléron Bay, France. Marine Ecology Progress Series 246 : 17-37.
Lepart J, Escarré J. 1983. La succession végétale, mécanismes et modèles : analyse
bibliographique. Bulletin d'Ecologie 14: 133-178. Llambi LD, Fontaine M, Rada F, Saugier B, Sarmiento L. 2003. Ecophysiology of dominant
plant species during old-field succession in a high tropical Andean system. Arctic, Antarctic, and Alpine Research 35: 36-42.
Loreau M. 2002. Conséquences de l’érosion de la biodiversité sur le maintien des
écosystèmes. CNRS. Lettre PIGB-PMRC Numéro 13. Martineau Y. 2001. Modélisation d’une succession secondaire dans le paramo des Andes
vénézuéliennes; croissance des plantes et compétition pour l’espace, la lumière et l’azote. Mémoire de DEA, Université Paris VI, France. 30 pp. & annexes.
Martineau Y, Saugier B. 2004. A process-based model of old-field succession linking
ecosystem and community ecology. Submitted. Matthews RB, Stephens W (eds). 2002. Crop-Soil Models: Applications in Developing
Countries. CAB International, Wallingford, UK. Migueis J, Hervé D, Génin D, Rivière G. 1998. PUMANI: un modèle de représentation de
l’activité agropastorale dans les Andes. Document ORSTOM Montpellier 9. 69 pp. & annexes.
Molinillo MF, Farji Brener AG. 1993. Cattle as a dispersal agent of Acaena elongata
(Rosaceae) in the cordillera of Merida, Venezuela. Journal of Range Management 46: 557-561.
Monasterio M. 1980. Las formaciones vegetales de los paramos de Venezuela. Monasterio
M, editor. Estudios ecologicos en los paramos Andinos. Mérida (Venezuela): Ediciones de la Universidad de los Andes. 45-91.
Monsi M, Saeki T. 1953. Über den Lichtfaktor in den Pflanzengesellschaften und seine
Bedeutung für die Stoffproduktion. Japanese Journal of Botany 14: 22-52. Muller CH. 1952. Plant succession in arctic heath and tundra in northern Scandinavia.
Bulletin of the Torrey Botanical Club 79: 296-309. Noodén LD, Guiamét JJ, John I. Senescence mechanisms. Physiologia Plantarum 101: 746-
753.

102
Ortuño T, Beck S, Sarmiento L. 2004. Dinamica sucesional de la vegetacion en un sistema
agricola con descansos largos en el Altiplano central boliviano. Ecologia en Bolivia (sous presse).
Pansu M, Bottner P, Sarmiento L, Metselaar K. 2004. Comparison of five soil organic matter
decomposition models using data from a 14C and 15N labelling field experiment. Soumis à Global Biogeochemical Cycles.
Pansu M, Metselaar K, Bottner P, Sarmiento L. 2004. Sensitivity analysis of two types of soil
organic matter decomposition models. Soumis à Global Biogeochemical Cycles. Parton WJ, Scurlock JMO, Ojima DS, Gilmanov TG, Scholes RJ, Schimel DS, Kirchner T,
Menaut JC, Seasetedt T, Garcia Moya E, Apinan Kamnalrut, Kinyamario JI. 1993. Observations and modelling of biomass and soil organic matter dynamics for the grassland biome worldwide. Global Biogeochemical Cycles 7: 785-809.
Paz-Betancourt B. 1997. Un modèle multi-agent pour simuler les accords de réciprocité dans
les Andes boliviennes [Thèse de Doctorat]. Université Claude Bernard Lyon 1 (France). 220 pp.
Pearl R, Reed LJ. 1920. On the rate of growth of the population of United States since 1790
and its mathematical representation. Proceedings of the National Academy of Sciences 6: 275-288.
Penning de Vries FWT, Jansen DM, ten Berge HFM, Bakema A. 1989. Simulation of
ecophysiological processes of growth in several annual crops. Pudoc: Wageningen (Netherlands). 269 pp.
Pestalozzi H. 2000. Sectoral fallow systems and the management of soil fertility: the
rationality of indigenous knowledge in the high Andes of Bolivia. Mountain Research and Development 20: 64-71.
Phillips DA. 1980. Efficiency of symbiotic nitrogen fixation in legumes. Annual Review of
Plant Physiology 31: 29-49. Reynolds JF, Bugmann H, Pitelka LF. 2001. How much physiology is needed in forest gap
models for simulating long-term vegetation response to global change? Challenges, limitations, and potentials. Climatic Change 51: 541-557.
Ruimy A. 1995. Modélisation de la productivité primaire nette continentale [Thèse de
doctorat]. Orsay (France): Université Paris-Sud. 36 pp. Ryan MG. 1991. A simple method for estimating gross carbon budgets for vegetation in forest
ecosystems. Tree Physiology 9: 255-266. Ryle GJA, Powell CE, Gordon AJ. 1979. The respiratory costs of nitrogen fixation in
soybean, cowpea and white clover. Journal of Experimental Botany 30: 135-144.

103
Sala OE, Austin AT. 2000. Methods of estimating aboveground net primary productivity. In: Methods in Ecosystem Science (eds Sala OE, Jackson RB, Mooney HA, Howarth RW), Springer. New York. 31-43.
Sarmiento L. 1995. Restauration de la fertilité dans un système agricole à jachère longue des
hautes Andes du Venezuela [Thèse de doctorat]. Orsay (France): Université Paris-Sud. 237 pp.
Sarmiento L. 2000. Water balance and soil loss under long fallow agriculture in the
Venezuelan Andes. Mountain Research and Development. 20: 246–253. Sarmiento L, Bottner P. 2002. Carbon and nitrogen dynamics in two soils with different
fallow times in the high tropical Andes: indications for fertility restoration. Applied Soil Ecology 19: 79-89.
Sarmiento L, Llambi LD, Escalona A, Marquez N. 2003. Vegetation patterns, regeneration
rates and divergence in an old-field succession in the high tropical Andes. Plant Ecology 166: 63-74.
Sarmiento L, Martineau Y, Acevedo D, Couteaux MM, Saugier B, Escalona A. 2004.
Evidence of hidden fluxes in the estimation of Net Primary Production in herbaceous ecosystem. In preparation.
Sarmiento L, Monasterio M, Montilla M. 1993. Ecological bases, sustainability and current
trends in traditional agriculture in the Venezuelan high Andes. Mountain Research and Development 13: 611-618.
Sarmiento L, Smith JK, Monasterio M. 2002. Balancing conservation of biodiversity and
economical profit in the agricultural of the high Venezuelan Andes: are fallow systems an alternative? Körner C, Spehn EM, editors. Mountain biodiversity – A global assessment. Parthenon publishing, London (UK). 283-293.
Sarps A. 2001. Caractérisation fonctionnelle des principales espèces d’une succession
secondaire. Mémoire de fin d’étude. ENITA Clermont-Ferrand. 30 pp. & annexes. Saugier B, Pontailler JY. 2004. Cycle global du carbone et photosynthèse des plantes de
l’Altiplano bolivien. Ecologia en Bolivia (sous presse). Savenkoff C, Vezina AF, Bundy A. 2001. Inverse analysis of the structure of the
Newfoundland-Labrador Shelf ecosystem. Canadian Technical Report of Fisheries and Aquatic Sciences 2354: 56 pp.
Sebillotte M, Allain S, Doré T, Meynard JM. 1993. La jachère et ses fonctions agronomiques,
économiques et environnementales. Le courrier de l’environnement de l’INRA 20: 11-22.
Sheehy JE. 1987. Photosynthesis and nitrogen fixation in legume plants. CRC Critical
Reviews in Plant Sciences. Boca Raton, Florida (USA): CRC Press Inc. 121-159.

104
Sinoquet H, Rakocevic M, Varlet-Grancher C. 2000. Comparison of models for daily light partitioning in multispecies canopies. Agricultural and Forest Meteorology 101: 251-263.
Sinoquet H, Thanisawanyangkura S, Mabrouk H, Kasemsap P. 1998. Characterization of the
light environment in canopies using 3D digitising and image processing. Annals of Botany 82: 203-212.
Slobodkin LB. 2001. The good, the bad and the reified. Evolutionary Ecology Research 3: 1-
13. Smil V. 2001. Enriching the Earth: Fritz Haber, Carl Bosch, and the Transformation of World
Food. MIT Press, Cambridge, MA. 339 pp. Smith FE. 1963. Population dynamics in Daphnia Magna and a new model for population
growth. Ecology 44: 651-663. Soussana JF, Minchin FR, MacDuff JH, Raistrick N, Abberton MT, Michaelson-Yeates TPT.
2002. A simple model of feed-back regulation for nitrate uptake and N2 fixation in contrasting phenotypes of white clover. Annals of Botany 90: 139-147.
Spitters CJT, Toussaint HAJ, Goudriaan J. 1986. Separating the diffuse and direct
components of global radiation and its implications for modelling canopy photosynthesis. Part I. Components of incoming solar radiation. Agricultural and Forest Meteorology 38: 217-229.
Spitters CJT. 1986. Separating the diffuse and direct components of global radiation and its
implications for modelling canopy photosynthesis. Part II. Calculation of canopy photosynthesis. Agricultural and Forest Meteorology 38: 231-242.
Thornley JHM. 1970. Respiration, growth and maintenance in plants. Nature 227: 304-305. Thornley JHM, Cannell MGR. 2000. Modelling the components of plant respiration:
representation and realism. Annals of Botany 85: 55-67. Thornley JHM, Johnson IR. 1990. Plant and Crop Modelling. Oxford Science Publications
670 pp. Tilman D. 1988. Plant strategies and the dynamics and structure of plant communities.
Monographs in Population Biology. Princeton, New Jersey (USA): Princeton University Press. 360 pp.
Tilman D. 1990. Constraints and tradeoffs: toward a predictive theory of competition and
succession. Oikos 58: 3-15. Tilman D. 1997. Community invasibility, recruitment limitation, and grassland biodiversity.
Ecology 78: 81-92. Tsoularis A. 2001. Analysis of logistic growth models. Research Letters in the Information
and Mathematical Sciences 2: 23-46.

105
Tsoularis A, Wallace J. 2002. Analysis of logistic growth models. Mathematical Biosciences
179: 21–55. Urbanska KM. 1997. Restoration ecology of alpine and arctic areas: are the classical concepts
of niche and succession directly applicable? Opera Botanica 132:189-200. van Oene H, Berendse F, De Kovel CGF. 1999a. Model analysis of the effects of historic CO2
levels and nitrogen inputs on vegetation succession. Ecological Applications 9: 920-935. van Oene H, van Deursen EJM, Berendse F. 1999b. Plant-herbivore interactions and its
consequence for succession in wetland ecosystems: a modelling approach. Ecosystems 2: 122-138.
Verhulst PF. 1838. Notice sur la loi que la population suit dans son accroissement.
Correspondance Mathématique et Physique 10 : 113-121. Voisin AS, Salon C, Jeudy C, Warembourg FR. 2003. Symbiotic N2 fixation activity in
relation to C economy of Pisum sativum L. as a function of plant phenology. Journal of Experimental Botany 54: 2733-2744.
Wardle DA. 2002. Communities and Ecosystems: linking the aboveground and belowground
components. Princeton (USA): Princeton University Press. 392 pp. Wit CT de. 1978. Simulation of assimilation, respiration and transpiration of crops,
Simulation Monographs, Pudoc, Wageningen. 141 pp. Yachi S, Loreau M. 1999. Biodiversity and ecosystem productivity in a fluctuating
environment: the insurance hypothesis. Proceedings of the National Academy of Sciences of the USA 96: 1463-1468.
Zeide B. 1993. Analysis of growth equations. Forest Science 39: 594-616.

106
Table des illustrations

107
TABLEAUX T 1 : Partenaires du projet TROPANDES.
T 2 : Activités du projet TROPANDES.
T 3 : Caractéristiques des sites étudiés.
T 4 : Paramètres utilisés dans le modèle FAPROM, données vénézuéliennes.
T 5 : Equations du modèle FAPROM.
T 6 : Variables d’intérêt et sorties « réfutables » du modèle FAPROM.
T 7 : Variables échangées entre les modèles.
T 8 : Paramètres utilisées dans le modèle FAPROM, données boliviennes.
T 9 : Caractéristiques de croissance et statut de succession des espèces étudiées.
T10 : Dégradation du modèle FAPROM (équations, paramètres).
T11 : Décomposition et/ou disparition de litière.
T12 : Estimation de la production primaire nette aérienne.
T13 : Bilan de carbone de la parcelle.
T14 : Conditions initiales optimisées pour deux parcelles boliviennes.
T15 : Bilan d’azote sous différents scénarios de jachère.
5
5
8
22
24
33
35
39
40
45
50
51
53
68
72
FIGURES F 1 : Situation géographique et diagrammes climatiques des sites d’étude.
F 2 : Courbes de dilution en azote (Greenwood et al. 1990 ; Sarmiento 1995).
F 3 : Classification des modèles écologiques.
F 4 : Modélisation des cycles du carbone et de l’azote.
F 5 : Modélisation de l’allocation (Hypericum laricifolium).
F 6 : Processus de modélisation.
F 7 : Intégration des modèles du projet TROPANDES.
F 8 : Chronogramme des activités agricoles.
F 9 : Patron de croissance des 12 espèces paramétrées.
F10 : Elasticité des paramètres de sortie r et K à différents traits des espèces.
F11 : Dégradation du modèle FAPROM (mécanismes de la croissance).
6
10
18
20
28
32
34
35
38
41
43

108
F12 : Phytomasse de racines et production souterraine (Sarmiento et al. 2004).
F13 : Suivis de biomasse aérienne (Sarmiento et al. 2004).
F14 : Production primaire nette journalière et flux simulés.
F15 : Dynamique de la communauté végétale.
F16 : Fonctionnement de l’écosystème.
F17 : Sensibilité du modèle aux conditions initiales.
F18 : Sensibilité du modèle aux paramètres Hmax et SRL.
F19 : Sensibilité du modèle au niveau de ressources (eau et azote).
F20 : Réponse de la communauté végétale au niveau de ressources (eau et azote).
F21 : Elasticité de la biomasse aux paramètres Hmax et SRL.
F22 : Biovolume par groupe fonctionnel sur deux parcelles boliviennes.
F23 : Biovolume simulé versus mesuré sur deux parcelles boliviennes.
F24 : Dynamique du contenu en azote au cours de la jachère.
F25 : Effet du pâturage sur la productivité de l’écosystème.
F26 : Effet de la perte de diversité sur le fonctionnement de la communauté.
48
49
52
56
57
60
62
63
64
65
67
68
71
76
78

109
Table des matières

110
I II III
1
2 3 3 4 7 7 9 11 11 12
14
15 15 16 16
18 18 19
21 21 21 26 26 27 28 29 29 30 30 30 31 33 34
Financement du travail Dédicace Remerciements Sommaire Introduction Des systèmes de culture à jachère Des successions post-culturales Du projet TROPANDES Des sites d’étude Des rotations culturales andines Des sols et de la fertilité De la production végétale au cours de la succession post-culturale Du modèle FAPROM et des questions étudiées De l’articulation du travail et du plan de la synthèse 1 - Développement d’un modèle mécaniste de succession
végétale (FAPROM) 1.1 - POURQUOI MODELISER ? 1.1.1 - Qu’est-ce qu’un modèle ? 1.1.2 - Le modèle répond à des objectifs scientifiques généraux. 1.1.3 - Le modèle apporte son concours aux études de terrain. 12 - À PARTIR DE QUOI MODELISER ? 1.2.1 - Quel type de modèle est-il adapté à nos objectifs ? 1.2.2 - Le modèle est construit sur des hypothèses simplificatrices. 13 - COMMENT MODELISER ? 1.3.1 - Le modèle conceptuel repose sur les mécanismes écophysiologiques. Architecture du couvert Compétition pour la lumière et photosynthèse potentielle Respiration des plantes Absorption d’azote et assimilation réalisée Allocation des assimilats Sénescence des organes Matière organique du sol Cycle reproductif Pâturage et cueillette 1.3.2 - Le modèle est paramétré à partir d’observations et de mesures de terrain. 1.3.3 - Le modèle informatique a été développé en FORTRAN puis VENSIM. 1.3.4 - Le modèle est utilisé pour estimer la dynamique de l’écosystème. 1.3.5 - Le modèle est couplé à des modèles de sol et de culture.

111
36
37 37 40 41 43 47 53
55 55 58 60 60 63 65 65 66 66 67 67 69 70
71 71 73 74 75 76 77
80
81 81 81 81 83 83 85
86 86 86 86 87
2 - Applications du modèle, de la plante à l’écosystème 2.1 - CROISSANCE D’UN COUVERT VEGETAL MONOSPECIFIQUE 2.1.1 - Patron de croissance végétale, statut de succession et traits des espèces Patron de croissance et statut de succession Patron de croissance et traits des espèces (étude de sensibilité I) Patron de croissance et mécanismes 2.1.2 - Application à l’estimation de la production primaire nette Que nous apprend le suivi de biomasse des parcelles vénézuéliennes ? 2.2 - DYNAMIQUE DE LA COMMUNAUTE VEGETALE 2.2.1 - Fonctionnement du modèle avec six espèces en interaction 2.2.2 - Le modèle surestime les vitesses de croissance et de disparition des espèces. 2.2.3 - Etudes de sensibilité II Sensibilité aux conditions initiales Réponse de l’écosystème au niveau de ressources (azote et eau) 2.2.4 - Compétition et coexistence Sensibilité aux paramètres Hmax et SRL (étude de sensibilité III) Rôle de la compétition dans le patron de succession Conditions de coexistence Modèle mécaniste simplifié 2.2.5 - Tentative de validation du modèle sur les parcelles boliviennes 2.2.6 - Combien d’espèces et quelles espèces faut-il prendre en compte ? Comment gérer la sensibilité aux conditions initiales ? 2.3 - ROLE DE LA JACHERE DANS L’AGRO-ECOSYSTEME 2.3.1 - Cycle de l’azote au cours de la jachère, potentialité de la jachère améliorée Quelles sont les fonctions agro-écologiques de la jachère ? Proposition de gestion durable de l’agro-écosystème A quelle distance sommes-nous d’un modèle d’aide à la décision ? 2.3.2 - Impact du pâturage 2.3.3 - Diversité des communautés et fonctionnement de l’écosystème 3 - Bilan critique et perspectives 3.1 - HYPOTHESES DU MODELE 3.1.1 - Intérêt du modèle, synthèse des résultats obtenus But du modèle Thèses développées 3.1.2 - Critique des hypothèses et perspectives de prolongement Limites du modèle Quel modèle choisir ? 3.2 - PROJET DE THESE 3.2.1 - Dynamique du projet TROPANDES, travail en équipe 3.2.2 - Evaluation critique du travail Dans quelle mesure les objectifs ont-ils été atteints ? Analyse des difficultés rencontrées

112
90 90 90 91 91 91
93
96
106
109
A1
A2
A40
A75
A96
3° 4° 4°
carte
3.3 - REFLEXIONS PERSONNELLES ET PERSPECTIVES 3.3.1 - De la multiplicité des métiers du chercheur 3.3.2 - Avenir du modèle FAPROM Validation complète sur les données vénézuéliennes Adaptation aux écosystèmes méditerranéens Utilisation du modèle dans les études ‘diversité et fonctionnement’ Conclusion Références Table des illustrations Table des matières Annexes 1 A process-based model of old-field succession linking ecosystem and community
ecology. Martineau Y, Saugier B. Article soumis. 2 Evidence of hidden fluxes in the estimation of Net Primary Production in
herbaceous ecosystems. Sarmiento L, Martineau Y, Acevedo D, Coûteaux MM, Saugier B, Escalona A. Article en préparation.
3 Uses and abuses of logistic growth: considering turnover to avoid biological
nonsense in ecological modelling. Martineau Y, Deredec A, Genton B, Saugier B. Projet d’article.
4 Modèle mécaniste simplifié de succession secondaire. Piste d’étude. Resumen en español Résumé en français Abstract in English Equations et paramètres du modèle


Resumen
El sistema agrícola tradicional de los altos Andes tropicales alterna cortos períodos de cultivo y barbechos largos. El crecimiento demográfico induce una intensificación de consecuencias arriesgadas (pérdida de fertilidad y de biodiversidad). El proyecto europeo TROPANDES estudió los procesos agro-ecológicos que controlan la fertilidad del suelo para proponer una alternativa duradera a la intensificación. La vegetación natural que coloniza las parcelas abandonadas se incorpora a la tierra en el momento de romper el barbecho. Desarrollamos un modelo biogeoquímico de sucesión vegetal que permite cuantificar la acumulación de biomasa durante el barbecho. FAPROM (Fallow Production Model) describe una mezcla con varias capas de seis especies en competencia por la luz y el nitrógeno mineral. Simula distintos procesos: fotosíntesis en intervalos de una hora; asignación del carbono, respiración, senecencia, absorción y fijación de nitrógeno en intervalos diarios; y reproducción en intervalos anuales. El modelo se aplicó a dos lugares particulares: el paramo húmedo (Venezuela) y la puna semiárida (Bolivia). Se obtuvieron distintos resultados: (1) Nuestras simulaciones muestran la importancia del turn-over de la materia (respiración, senescencia) para considerar el crecimiento, la dinámica de la vegetación y la producción primaria del ecosistema. (2) La dinámica de la vegetación y el funcionamiento del ecosistema dependen estrechamente de las condiciones iniciales (semillas) y de los recursos (agua, nitrógeno). (3) El modelo permite cuantificar la importancia relativa de los procesos que controlan el balance nitrogenado bajo distintas situaciones (con o sin leguminosas).
Palabras clave: Bolivia; Carbono; Dinámica de la vegetación; Fotosíntesis; Funcionamiento del ecosistema; Nitrógeno; Páramo; Producción primaria; Puna; Venezuela.

Résumé Le système agricole traditionnel des hautes Andes tropicales alterne courtes périodes
de culture et jachères longues. La croissance démographique des populations locales induit une intensification aux conséquences risquées (perte de fertilité et de biodiversité). Le projet européen TROPANDES a étudié les processus agro-écologiques qui contrôlent la fertilité du sol pour proposer une alternative durable à l’intensification. La végétation naturelle qui colonise les parcelles abandonnées est incorporée à la terre au moment de rompre la jachère. Aussi avons-nous développé un modèle biogéochimique de succession végétale qui permet de quantifier l’accumulation de biomasse pendant la jachère. FAPROM (Fallow Production Model) décrit un mélange multicouche de six espèces en compétition pour la lumière et l’azote minéral. Il simule divers processus : photosynthèse au pas de temps horaire ; allocation du carbone, respiration, sénescence, absorption et fixation d'azote au pas de temps journalier ; et reproduction au pas de temps annuel. Le modèle a été appliqué à deux sites particuliers : le paramo humide (Venezuela) et la puna semi-aride (Bolivie). Nos simulations montrent l’importance du turn-over de la matière (respiration, sénescence) pour estimer la croissance des plantes, la dynamique de la végétation et la production primaire de l’écosystème. La dynamique de la végétation et le fonctionnement de l’écosystème dépendent étroitement des conditions initiales (graines) et des ressources (eau et azote). Le modèle permet en outre de quantifier l’importance relative des processus qui contrôlent le bilan azoté sous différents scénarios (avec ou sans légumineuses). Mots clefs : Azote ; Bolivie ; Carbone ; Dynamique de la végétation ; Fonctionnement de l’écosystème ; Paramo ; Photosynthèse ; Production primaire ; Puna ; Venezuela.
Abstract The traditional cropping system of the high tropical Andes alternates short cropping
periods and long fallows. Demographic growth induced an intensification with hazardous consequences (loss of fertility and biodiversity). The European project TROPANDES studied the agro-ecological processes that control soil fertility to give a basis for a sustainable alternative. The natural vegetation colonizing fallow plots is incorporated to the ground at crop start. Therefore we built a biogeochemical model of plant succession to account for the accumulation of biomass during fallow. FAPROM (Fallow Production Model) describes a multi-layer mixture of six species in competition for light and mineral nitrogen. It simulates various processes: photosynthesis at a hourly time step; assimilates allocation, respiration, tissue mortality, nitrogen absorption and nitrogen fixation at a daily time-step; and reproduction at a yearly time step. The model was applied to two particular sites: the wet paramo (Venezuela) and the semi-arid puna (Bolivia). Simulations show the importance of matter turnover (respiration, senescence) to estimate plant growth, vegetation dynamics and ecosystem primary production. Vegetation dynamics and ecosystem functioning depend on initial conditions (seed bank) and on resources (water and nitrogen). The model helps quantifying the relative importance of the processes that control the nitrogen balance under several scenarios (with or without legumes). Keywords: Bolivia; Carbon; Ecosystem functioning; Nitrogen; Paramo; Photosynthesis; Primary production; Puna; Vegetation dynamics ; Venezuela.